Embed Size (px)
Citation preview

Lecture 15:Regulation of Proteins 2:
Allosteric Control of Hemoglobin
Hemoglobin and Myoglobin
Allosteric Transition in Hemoglobin
Physiological Role of Hemoglobin
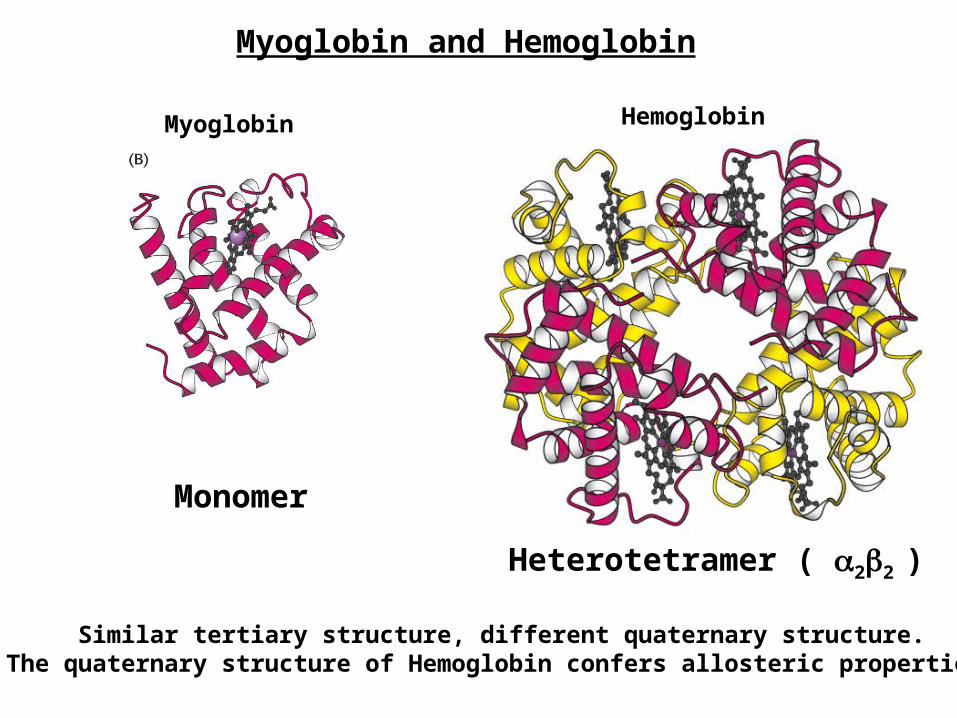
Myoglobin and Hemoglobin
Monomer
Heterotetramer ( 22 )
Myoglobin Hemoglobin
Similar tertiary structure, different quaternary structure.The quaternary structure of Hemoglobin confers allosteric properties.

Myoglobin:
Biological role: oxygen storage protein (1 binding site)
Binds oxygen in muscle cells, keeping it until needed.
Hemoglobin:
Biological role: oxygen transport protein. (4 binding sites.)
Circulates in red blood cells.
Binds oxygen when in lungs, and releases it whenin oxygen-requiring tissues.
Affinity for oxygen is modulated so as to bind tightlyin lungs, but release easily when in tissues. Themodulation relies on the allosteric properties ofhemoglobin.

Binding of Oxygen by Hemoglobin
(In solution, dissolved gases such as oxygen are described in terms ofpartial pressure, not concentration. )
In lungs, the partial pressure of oxygen is high, ~100 torr.In tissues, the partial pressure of oxygen is lower, ~20 torr.
Hemoglobin must bind oxygen tightly enough that it can fill up its siteswhen in the lungs, but not so tightly that it cannot release oxygenwhen it reaches tissues where oxygen is needed.
In other words, it would be most efficient if, over the range ofoxygen levels between lungs and tissues, the molecule couldgo from completely saturated (4 sites occupied) to completelyunloaded (zero sites occupied.)
Hemoglobin comes close to achieving this through cooperativity.

Cooperative Binding
The fractional saturation is the percentage of the total binding sites occupied.
Hemoglobin has a sigmoidal binding response to oxygen- oxygen bindingby hemoglobin is cooperative. (the binding of oxygen at one siteinfluences binding at other sites)
Hemoglobin:
100 torr: 98% saturated20 torr: 32% saturated
66% change possible due to cooperativity
(a non-cooperativetransport proteincould achieve 38%at best)

Basis of Hemoglobin Cooperativity
The ability of the oxygen-binding sites to influence one another results from a conformational change in hemoglobin.
The T state has a low affinity for oxygen, while the R state hasa high affinity for oxygen. The abrupt change in oxygen affinityresults from a switch between the low affinity state and thehigh affinity state.

Oxygen binding sites in Hemoglobin
Oxygen is bound on an iron ion on prosthetic groups called heme.There are 4 hemes in hemoglobin between 24 and 40 Angstromsapart. Each iron has six coordination sites, one of whichcan be occupied by oxygen.
Heme = Fe2+ ion and protoporphyrin
N N
NN
Histidine
Oxygen
Fe

In the absence of oxygen, the iron ion lies 0.4 Angstromsoutside the plane of the porphyrin ring.

When oxygen binds, the iron ion moves into the plane of theporphyrin ring, pulling the histidine upwards and closer to theporphyrin.

In turn, force on the histidine causes upward displacement of a helix which leads directly to the interface region of the tetramer.
The changes at the heme due to oxygen binding are “communicated” through the protein to the interface, causing the rearrangement of the quaternary structure, which in turn affects the affinity for oxygen at the other sites.
(Two distinct stablequaternary arrangements)

Free Energy
All sites boundNo sites bound
Fractional Occupancy
0 1
R state
T state
In the absence of oxygen, the T state is more stable, and soa larger percentage of the tetramers are in the T state.
But when oxygen levels are high, most of the sites are bound,and the R state predominates.

pO2
Sites Occupied
The sigmoidal shape of the binding curve results from theswitch between the T and R states.
Effect of the Allosteric Switch on Binding Affinity

All T All RMixed T and R subunits
Sequential Model
Models for Cooperativity
All RAll T
Concerted Model
Most allosteric proteins fall somewhere in between the two extremes.

Allosteric Effectors of Hemoglobin
The affinity of hemoglobin for oxygen can be controlled by externaleffectors, which affect the equilibrium between the T and R states.Favoring the T state assists hemoglobin in unloading its boundoxygen.
2,3-Bis-phosphoglycerate: BPG decreases the affinity of hemoglobin foroxygen, assisting it to unload the oxygen molecules where theyare needed.
The Bohr effect: Carbon dioxide and hydrogen ions also decreasethe affinity of hemoglobin for oxygen, so that a larger fraction of itsoxygen can be delivered in the tissues.

BPG Binding Stabilizes the T state
( - 5 charge)
Purified hemoglobin has much higher affinityfor oxygen than when in red blood cells. A substance in red blood cells, 2,3-bisphosphoglycerate (BPG), binds the T state of hemoglobin, effectively decreasing the affinity for oxygen. The binding of BPG is stabilized by ionic interactions.

Fetal hemoglobin is less sensitive to BPG
During early development, the human fetus expresses different and hemoglobin genes. These are similar but not identical to the hemoglobingenes expressed in adults. The fetal chain differs in that residue 143(a histidine in adult hemoglobin) is a serine, which is uncharged. Anionic interaction at the BPG binding site is removed, decreasing the abilityof BPG to bind. As a result, fetal hemoglobin has a higher affinityfor oxygen, allowing transfer from the maternal adult hemoglobin.

Increased BPG Contributes to Altitude AdaptationThe low levels of oxygen found at high altitude make it more difficult totransport sufficient oxygen from lungs to tissues. To overcome thisdifficult, a complicated adaptation process takes place in which morered blood cells and more hemoglobin are produced, which requires severalweeks. (Mountain climbers often have a series of camps where theypause to allow time for this process.)
There is also a faster physiological response, which takes place over thecourse of only a few hours. Red blood cells can increase their levels ofBPG, decreasing the affinity of hemoglobin for oxygen, with the result thatmore oxygen can be unloaded in the tissues.
1
2
3
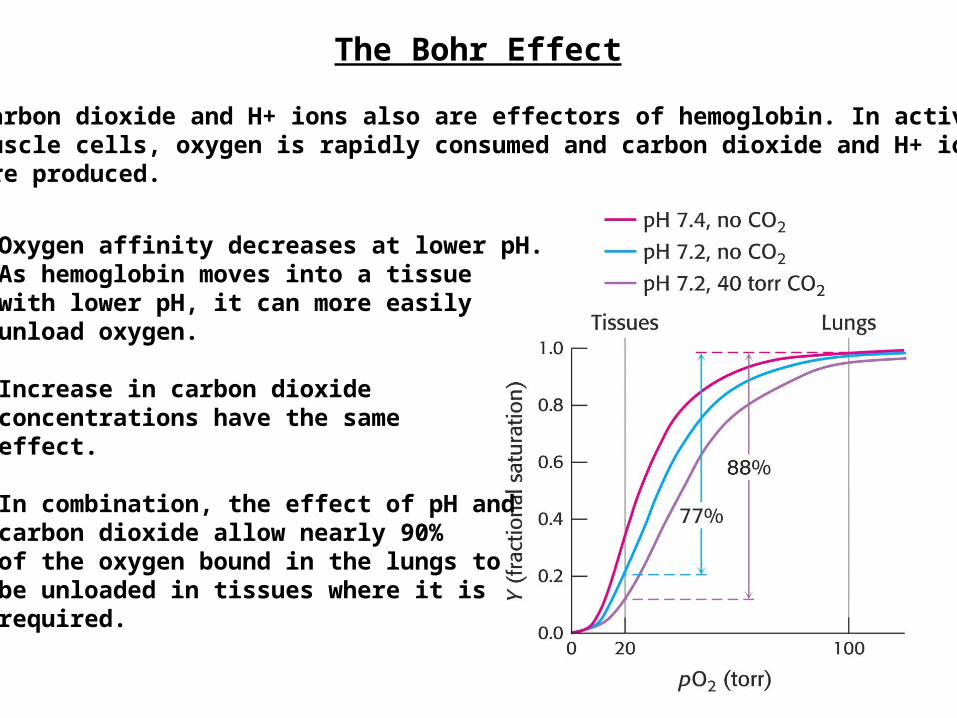
The Bohr Effect
Carbon dioxide and H+ ions also are effectors of hemoglobin. In activemuscle cells, oxygen is rapidly consumed and carbon dioxide and H+ ionsare produced.
Oxygen affinity decreases at lower pH.As hemoglobin moves into a tissuewith lower pH, it can more easilyunload oxygen.
Increase in carbon dioxideconcentrations have the sameeffect.
In combination, the effect of pH andcarbon dioxide allow nearly 90%of the oxygen bound in the lungs tobe unloaded in tissues where it isrequired.

The Bohr Effect can be explained in terms of ionic interactions.
Structural Basis of the Bohr Effect
When protonated, His 146 on the chain is positively charged, and can participate in an ionic interaction with Asp 94, which stabilizes the T state. At low pH, the T state is favored, decreasing the oxygen affinity.
The amino termini of the hemoglobin chains lie at an interface between and subunits.
Carbon dioxide can react with thepositively charged amino termini toform negatively charged carbamategroups that form ionic interactionswhich also stabilize the T state.

Summary:
Hemoglobin is an oxygen transport protein that carries oxygen from thelungs, where oxygen levels are high, to tissues where it is needed.
Hemoglobin exhibits cooperative binding of oxygen which is the resultof a conformational switch between a low-affinity state and a high-affinitystate.
The affinity of hemoglobin for oxygen can be regulated by allostericeffectors to improve its physiological performance.
Key Concepts:Function of hemoglobinConformational changes due to oxygen bindingCooperativityRole of BPGRole of Bohr Effect





























