Embed Size (px)
Citation preview

Leaf Energy Balance
Leaf temperature has already been implicated in photorespiration. What determines leaf temperature?
Leaves, unlike most animal bodies, do not store a significant amount of energy. Leaf temperature is determined by dynamic balance between energy input and energy loss…
If a leaf is holding a constant temperature, then:
Rn = H + E
Rn is radiant energy input (solar energy absorbed).
H is sensible heat loss due to convection and conduction,
and E is latent heat loss (energy loss from transpiration and evaporation of water from the leaf).

We need to examine each of these terms and how they affect the energy balance of leaves. First, radiant energy…
We begin with light absorption by leaves. Leaves absorb radiation in the visible and shorter wavelength ranges, i.e. <700 nm. Leaves also reflect a portion of this spectral range.
However, leaves also absorb and radiate energy in the long wavelength region of the spectrum (>700 nm, infrared).
The energy of a photon is given by h, where is frequency, or the inverse of wavelength. Shorter wavelength = higher frequency and greater energy.

The amount of energy emitted by a surface is given by the “blackbody” or Stephan-Boltzmann equation:
= T4
is the flux density of energy emitted in W/m2. is the emissivity of the surface. is the Stephan-Boltzmann constant, 5.67x10-8 W/m2/K4.
The emissivity of real objects, including leaves, is in the range 0.9 < < 0.98.
So the key to radiation from an object is its temperature, measured in degrees Kelvin.
The sun, at 6000K, radiates ~73,000 W/m2.The earth, global average temperature ~290K, radiates ~400 W/m2.A leaf during daylight at ~300K radiates ~500 W/m2.

The radiant energy balance during the day is normally positive; more energy is absorbed than radiated. During the night that balance is usually reversed; more energy is radiated than absorbed.
However, the whole picture is more complicated than that sounds…

The second term in the equation for energy balance is sensible heat loss, comprising energy loss due to convection and conduction.
First conduction. This is direct transfer of heat energy from one body to another (bodies diffusely defined).
In this case one body is the leaf itself, and the other ‘body’ is surrounding atmosphere. Due to surface resistance, there is a boundary layer of air around a leaf that is the main portion with which conductive exchange occurs.
Even if the leaf is smooth surfaced, there is a boundary layer. However, many leaves have various forms of projections (trichomes, hairs, etc.) that tend to retain a more isolating boundary layer.

The leaves of tomato plants are covered by trichomes…
The leaves of stinging nettles have two kinds of ‘hairs’, the hollow stinging, needle-like large hairs and smaller hairs. The smaller ones are important in creating a boundary layer.

In some species the hairs are reflective, and reduce the amount of radiant heat energy absorbed. Such leaves (or other plant structures) may appear white or pale due to light reflection.

Boundary layers can insulate the leaf from being heated in hot air, as well. They influence both conduction and…
Convection is transport of energy by a volume of fluid (here air). It is far more important in leaf energy dynamics. Convection is limited by the occurrence and existence of the boundary layer, since that layer ‘disconnects’ the leaf from the air movements around it.
Here’s the basic diagram:

There is something called a characteristic dimension of a leaf that is determined by its size and shape (and, of course, surface texture due to trichomes, hairs, etc.), and is directly proportional to the boundary layer resistance of the leaf.
Leaves with a small characteristic dimension tend to remain at temperatures close to actual air temperature. Leaves with large characteristic dimensions can diverge in temperature from ambient temperature, either above or below surrounding air depending on radiant energy input and stomatal resistance that affects latent heat exchange (evaporation and transpiration).

Finally, there is latent heat exchange. This is the energy lost by the leaf as the result of evaporation (including transpiration) from the leaf. The energy required to change state from liquid to vapor (the heat of vaporization) is 2450 joules/gm at 20°C. The number of grams of water vaporizing in transpiration is affected by stomatal resistance (rs in the figure that follows), air temperature, boundary layer resistance (indicated by leaf characteristic dimension), wind speed (affecting convection), and relative humidity.
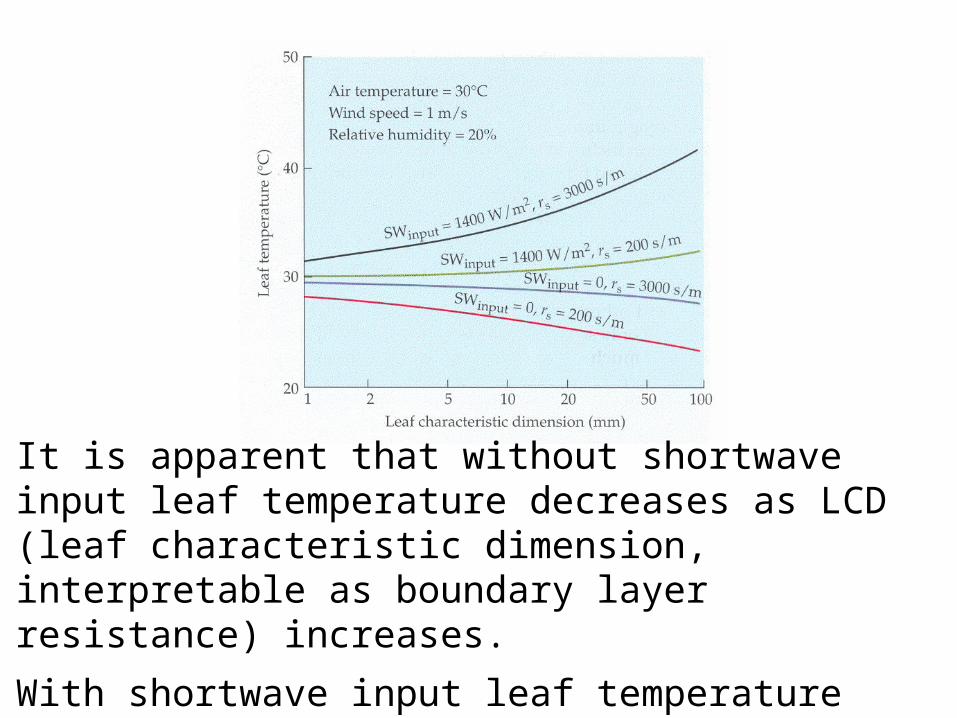
It is apparent that without shortwave input leaf temperature decreases as LCD (leaf characteristic dimension, interpretable as boundary layer resistance) increases.
With shortwave input leaf temperature increases as LCD increases; the increase is larger with greater boundary layer resistance.

The final segment of the chapter is about adaptation to extreme climate. There are really two temperatures of interest, whether the extreme is high temperature or low – one is called an activity limit, which means the temperature at which a plant ceases to function effectively, and the other more extreme, is the lethal limit, beyond which a plant dies.
Exposure to high temperaturesBeyond a temperature of ~40C (consult table 3.2 for more detailed data), plants shift from normal protein synthesis to rapid synthesis of a characteristic set of heat shock proteins. Many are homologous across a very wide taxonomic spectrum of plants, e.g. HSP70. Higher plants synthesize a group of small HSPs , all in the range of 15-18 kDa. These proteins protect against short exposures to high temperatures.

One, appropriately named, ubiqitin, apparently identifies denatured proteins for proteolysis.
When high temperature exceeds tolerance limits, the usual cause of cell death is loss of membrane integrity, with resultant ion leakage.
Nobel studied acclimation in agaves and cacti. Plants can adapt. He raised the growth temperature from 30/20 (day/night) to 50/40, and found that the temperature causing cell death increased by 1.6 to 15.8 among species. For these hot desert species, the lethal temperatures were generally >60 C. We don’t really know how this acclimation is achieved.

Low temperature injury
Low temperature injuries occur in two ways: tropical plants typically suffer what is called chilling injury (wilting, growth inhibition, germination failure, reproductive failure) below temperatures of ~10C. Damage from short exposure is reversible.
The more serious form of damage is freezing injury. Some of the tissue water freezes. Without acclimation freezing injury can occur at -1 to -3C. With acclimation tissues can survive to -40C, and seeds can survive to much lower temperatures.
The mechanism of chilling injury is also membrane damage, increasing permeability. The transition is usually rapid/abrupt with temperature decrease in sensitive plants. It has been suggested that membranes change from ‘fluid’ to ‘gel’.

A different suggestion is that microtubule function is associated with chilling injury.
Freezing injury results (both directly and indirectly) from ice crystal formation. Typically, ice crystals form initially extracellularly and in cell walls. Difference in the vapor pressure of ice and water cause cells to lose water. Some damage may result from dehydration and solute concentration within cells. Other damage can result from intracellular ice crystal formation.
Plants, as well as insects, are able to acclimate and achieve tolerance in part chemically. In plants, it is typically soluble carbohydrates and abscisic acid. Another protection is supercooling. Ice formation requires nucleation sites; inside cells such sites can be absent, so that water cools below even the freezing point set by solute concentration.

Some trees can prevent ice formation down to a temperature called the homogenous nucleation temperature (a temperature where ice crystals form spontaneously even without nucleation sites). Oaks, elms and maples can avoid ice formation down to -41 to -47C.
Chilling can even be advantageous. The text mentions cold air drainage. Cold air is denser than warm air and sinks. In northern Scandinavia the arctic birch Betula pendula is attacked by a moth that can totally defoliate plants. However, the moth has limited temperature tolerance (at least for an arctic species), so that birches at lower elevations in ‘bowls’ avoid fatal damage, while those higher up are killed.
What follows (if I can find the slide) is a picture of this situation from northern Finland.





























