Embed Size (px)
Citation preview

BioOne sees sustainable scholarly publishing as an inherently collaborative enterprise connecting authors nonprofitpublishers academic institutions research libraries and research funders in the common goal of maximizing access tocritical research
LARVAL DEVELOPMENT OF THE GHOSTSHRIMP CALLICHIRUS ISLAGRANDE (DECAPODATHALASSINIDEA CALLIANASSIDAE) UNDERLABORATORY CONDITIONSAuthor(s) Karen M Strasser and Darryl L FelderSource Journal of Crustacean Biology 20(1)100-117 2000Published By The Crustacean SocietyDOI httpdxdoiorg1016510278-0372(2000)020[0100LDOTGS]20CO2URL httpwwwbiooneorgdoifull1016510278-0372282000290205B01003ALDOTGS5D20CO3B2
BioOne (wwwbiooneorg) is a nonprofit online aggregation of core research in thebiological ecological and environmental sciences BioOne provides a sustainable onlineplatform for over 170 journals and books published by nonprofit societies associationsmuseums institutions and presses
Your use of this PDF the BioOne Web site and all posted and associated contentindicates your acceptance of BioOnersquos Terms of Use available at wwwbiooneorgpageterms_of_use
Usage of BioOne content is strictly limited to personal educational and non-commercialuse Commercial inquiries or rights and permissions requests should be directed to theindividual publisher as copyright holder
In a recent revision of the Callianassidae(Manning and Felder 1991) the genus Cal-lianassa was restricted from its former world-wide status to certain eastern Atlantic popu-lations based on morphological charactersAmerican species that were previously in-cluded in the genus Callianassa were placedin a number of genera based on life historyand morphology as in the case of the genusCallichirus As redefined by Manning andFelder (1986) the genus Callichirus now in-cludes at minimum C major (Say 1818) Cislagrande (Schmitt 1935) C seilacheri(Bott 1955) C garthi (Retamal 1975) andC adamas (Kensley 1974) and at least oneundescribed species from the Atlantic coastsof Brazil and Colombia which has been of-ten referred to as C major (see Staton andFelder 1995 D L Felder personal obser-vation) For a number of other callianassidspecies generic status remains unresolvedand they will herein be referred to as Cal-lianassa sensu lato (sl)
Detailed descriptions of larvae are avail-able for relatively few species in the familyCallianassidae all of which currently fall intothe subfamilies Callianassinae (Callianassasl kewalramanii Sankolli Sankolli andShenoy 1975 Neotrypaea uncinata (HMilne Edwards) Aste and Retamal 1984 Csl petalura Stimpson Konishi et al 1990)
and Callichirinae (Callichirus sp (formerlyC major) Rodrigues 1976 C garthi (Reta-mal) Aste and Retamal 1983 Sergio mirim(Rodrigues) Rodrigues 1984 Lepidophthal-mus louisianensis (Schmitt) L sinuensisLemaitre and Rodrigues Nates et al 1997Callichirus major Strasser and Felder 1999a)Larval development of Callichirus islagrandehas not been described even though this spe-cies is well studied (Felder and Griffis 1994Staton and Felder 1995 Strasser and Felder1998 1999b)
Callichirus islagrande is a Gulf of Mexicoendemic found typically on barrier islands inlow to high intertidal habitats This speciesinhabits the northern and western Gulf ofMexico ranging from northwestern Florida toParaiso Tabasco Mexico (Staton and Felder1995) A somewhat discontinuous distribu-tion of C islagrande occurs between south-eastern Louisiana and the northeastern coastof Texas where shorelines tend to lack thecleaner quartzite sand and high energybeaches that C islagrande prefers (Staton andFelder 1995)
The present study describes the zoeal anddecapodid stages provides illustrations foridentification and reports the number and du-ration of larval stages of Callichirus isla-grande Comparisons of these characteristicsin C islagrande are made with other con-
JOURNAL OF CRUSTACEAN BIOLOGY 20(1) 100ndash117 2000
LARVAL DEVELOPMENT OF THE GHOST SHRIMP CALLICHIRUSISLAGRANDE (DECAPODA THALASSINIDEA CALLIANASSIDAE)
UNDER LABORATORY CONDITIONS
Karen M Strasser and Darryl L Felder
Department of Biology University of Louisiana Lafayette Louisiana 70504 USA(DLF e-mail dfelderlouisianaedu) (KMS current address) Box 108F University of Tampa 401 W
Kennedy Boulevard Tampa Florida 33606 USA (e-mail kstrasseralphautampaedu)
A B S T R A C T
Descriptions of the zoeal and decapodid stages of the ghost shrimp Callichirus islagrande werebased on laboratory cultures from a population inhabiting the coast of Louisiana Whereas mor-phological studies were based upon animals from mass cultures 40ndash120 larvae from each of 8parental females were reared individually to obtain stage-duration data Larvae passed through 4 or5 zoeal stages before molting to the decapodid stage The proportion of individuals that molted toa fifth zoeal stage varied widely (9ndash89) between clutches of different parental females The firstjuvenile stage was usually reached 16ndash20 days after hatching and durations of larval stages weresimilar in larvae from different females Morphological comparisons with congeneric species re-vealed larvae of C islagrande to be more similar to larvae of C garthi a species from the coastof Chile than to larvae of a Gulf of Mexico population of Callichirus major
100
STRASSER AND FELDER LARVAL DEVELOPMENT IN CALLICHIRUS ISLAGRANDE 101
generic species and members of the familyCallianassidae
MATERIALS AND METHODS
Ovigerous females were collected in March May andJuly 1996 from the bayward side of a barrier island (IslesDernieres Louisiana) with yabby pumps (as in Felder1978) Adult females were maintained in 20-cm diame-ter finger bowls (at 27degC 25 ppt salinity) with daily wa-ter changes until eggs hatched Sea water was taken fromwell offshore Louisiana filtered through a 30-microm screenaerated and diluted to 25 ppt salinity with deionized wa-ter before use in the culture of animals To determinestage durations 40ndash120 larvae from each of 8 parentalfemales were reared individually through the first juve-nile stage (J1) ZI (first zoeal stage) larvae were movedto individual compartments upon hatching and maintainedat 27degC in filtered sea water on a 1212 light dark cy-cle Each day larvae were moved to containers with newsea water fed freshly hatched nauplii of Artemia (GreatSalt Lake) and examined to assess their stage of devel-opment Observations were terminated when animalsreached the first juvenile stage (J1)
Animals used for morphological comparisons were ob-tained from mass cultures Larvae were maintained in 20-cm diameter finger bowls (100ndash200 individuals per bowl)with daily water changes under conditions mentionedabove Animals were fixed in 70 ethanol stained withchlorozol black and transferred to glycerine prior to dis-section At least 10 animals were dissected for each stageand both right and left appendages were used for setalcounts Line illustrations were made on a Nikon invertedmicroscope fitted with a camera lucida Measurementswere made with a calibrated ocular micrometer Carapacelength (CL) was measured from the tip of the rostrum tothe posterior midpoint of the carapace and total length(TL) was measured from the rostral tip to the posteriormidpoint of the telson For each appendage the arrange-ment of setae was listed sequentially from proximal todistal margins as in Konishi (1989) and Nates et al(1997) Setal groups on successive segments were sepa-rated with a comma () Groups of setae on the same seg-ment or on different lobes of the same segment articleor endite were separated with a plus (+) A question mark() was used to designate questionable distinctions be-tween setae and aesthetascs Roman numerals were usedto describe the pattern of processes on the posterior mar-gin of the telson
RESULTS
Although there was no attempt to quantifythe duration of embryonic developmentovigerous females were held for up to 28 daysbefore the eggs hatched (usually at night)Prior to hatching eggs changed gradually incolor from bright orange-red to a translucent-brown Late-stage embryos that were droppedby the female before hatching and larvae thathatched as prezoeae were not viable evenwith aeration Prezoeae did not appear to bedeveloped fully and could not swim All zoealstages fed readily on nauplii of Artemia aswell as on each other The decapodid stage(D) appeared to consume much less than didthe preceding zoeal stage Strong positivephototaxis was exhibited by the first threezoeal (ZI to ZIII) stages The fourth zoealstage (ZIV) in general appeared to be at-tracted to light although some larvae reactedindifferently Decapodids did not exhibit ob-vious phototaxis
Larvae of Callichirus islagrande passedthrough four or five zoeal stages before molt-ing to D (Fig 1) these were usually com-pleted 11ndash14 days after hatching Mean du-rations of each stage were relatively consis-tent between larvae from different females(Table 1) However duration of ZIV was ap-proximately a day shorter when animalsmolted from ZIV to ZV than when animalsmolted directly to D (Table 2) Among theeight cultures observed in this study the per-centage of animals molting to ZV ranged from9ndash89 of the larvae that survived past ZIV
Description of Zoeal and Decapodid Stages
Near the mandibles the larval stages anddecapodid had two red chromatophores whichusually appeared fused when expanded Sev-
Table 1 Mean duration in days (plusmn95 CI) of the first to fifth zoeal stages (ZI to ZV) and the decapodid stage (D)in larval cultures from 8 ovigerous females of Callichirus islagrande Each culture consists of larvae from a differ-ent parental female Only animals that reached J1 (N) were used to calculate mean durations N0 is the starting num-ber of ZI larvae
Culture N N0 ZI ZII ZIII ZIV ZV D
1 100 120 226 (plusmn009) 204 (plusmn005) 222 (plusmn009) 388 (plusmn009) 389 (plusmn012) 570 (plusmn022)2 14 60 207 (plusmn016) 200 (plusmn000) 250 (plusmn031) 336 (plusmn060) 430 (plusmn028) 664 (plusmn117)3 34 55 206 (plusmn008) 206 (plusmn008) 244 (plusmn017) 421 (plusmn024) 420 (plusmn016) 568 (plusmn039)4 102 120 205 (plusmn004) 202 (plusmn005) 232 (plusmn010) 359 (plusmn010) 320 (plusmn009) 518 (plusmn028)5 31 40 205 (plusmn011) 200 (plusmn000) 226 (plusmn016) 323 (plusmn028) 400 (plusmn037) 494 (plusmn023)6 49 60 206 (plusmn007) 204 (plusmn008) 243 (plusmn014) 339 (plusmn020) 364 (plusmn018) 547 (plusmn046)7 36 54 217 (plusmn013) 211 (plusmn011) 214 (plusmn012) 294 (plusmn021) 327 (plusmn015) 439 (plusmn019)8 44 60 216 (plusmn013) 225 (plusmn015) 236 (plusmn017) 316 (plusmn026) 342 (plusmn017) 430 (plusmn030)
102 JOURNAL OF CRUSTACEAN BIOLOGY VOL 20 NO 1 2000
eral other red chromatophores were presenton the lateral edges of the carapace forminga red line when expanded
Morphological descriptions of Callichirusislagrande are as detailed below for stages ZIto ZIV and D ZV resembled ZIV in appear-ance However appendages were like thoseof ZIV D or some gradation between thetwo Setal patterns are given from proximal todistal segments for each appendage Carapacelength (CL) and total length (TL) are given asthe mean plusmn 95 CI in mm followed by therange of measurements from 10 specimens
Zoea IFigs 2a 3andashj
SizemdashCL = 123 plusmn 006 mm range 110ndash141mm TL = 326 plusmn 007 mm range 311ndash336mm
Carapace (Fig 2a)mdashShorter than abdomenrostrum elongate rounded in cross sectionand smooth eyes fused to carapace
Abdomen (Fig 2a)mdashSomites 2ndash5 with pos-terolateral projections and posterodorsalspines dorsal spine elongate on somite 2sometimes extending beyond posterior mar-gin of somite 3 short dorsal spines on somites3ndash5
Antennule (Fig 3a)mdashElongate exopodal andendopodal lobes not distinct 4ndash6 terminalaesthetascs 1 long plumose seta on future en-dopodal lobe
Antenna (Fig 3b)mdashProtopod with 1 distalspine between rami endopod with 2 longplumose setae scaphocerite (exopod) armedwith 1 strong distolateral spine 8 plumose se-tae on inner margin and 1 plumose seta onouter margin
Mandible (Fig 3c d)mdashIncisor process with6 or 7 teeth on right mandible 5ndash7 teeth onleft mandible molar process denticulate onboth mandibles
Maxillule (Fig 3e)mdashCoxal endite with 8ndash10marginal setae basal endite with 3 plumosesetae 3 or 4 large dentate spines and 4 or 5smaller dentate spines inner margin with 0or 1 plumose seta endopod 3-segmented dis-tinction between segments sometimes ob-scure 2 2 (3 or 4) setae protopod withoutsetae
Maxilla (Fig 3f )mdashCoxal endite bilobed 11or 12 setae on proximal lobe and 4 or 5 se-
Table 2 Percentage of larval Callichirus islangrande that molted from the fourth zoeal stage (ZIV) to the fifthzoeal stage (ZV) or directly to the decapodid stage (D) in larval cultures from 8 ovigerous females (Culture num-bers 1ndash8) Each batch consisted of larvae from a different parental female N0 = number surviving beyond ZIV N1
= number molting ZIV to ZV N1J = percentage of N1 surviving to J1 N2 = number molting ZIV to D N2J = per-centage of N2 surviving to J1 The mean duration in days (plusmn95 CI) is given for animals that molted from ZIV toZV and from ZIV to D
From ZIV To ZV To D
Survived to J1 Duration Survived to J1 DurationCulture N0 N1 N1J of ZIV N2 N2J of ZIV
1 105 11 82 30 (plusmn030) 94 97 40 (plusmn008)2 47 42 24 32 (plusmn023) 5 80 44 (plusmn111)3 45 8 62 30 (plusmn00) 37 78 44 (plusmn018)4 109 10 50 31 (plusmn078) 99 98 36 (plusmn010)5 38 19 79 28 (plusmn026) 19 84 37 (plusmn027)6 58 35 80 31 (plusmn017) 23 91 39 (plusmn030)7 48 30 73 26 (plusmn023) 18 78 34 (plusmn025)8 51 29 83 26 (plusmn019) 22 91 38 (plusmn019)
Fig 1 Mean proportion of larvae at each stage duringdevelopment of Callichirus islagrande Mean proportionsare derived from the individual rearings of 8 parental fe-males The first through fifth zoeal stages (ZI to ZV) thedecapodid stage (D) and all first-stage juveniles thatpassed D (J) are labeled under the peak of each line
STRASSER AND FELDER LARVAL DEVELOPMENT IN CALLICHIRUS ISLAGRANDE 103
tae on distal lobe basal edite bilobed with6ndash9 setae on each lobe endopod with 5 lobes3+2+2+2+(2 or 3) setae scaphognathite with16ndash19 setae all setae plumose
Maxilliped 1 (Fig 3g)mdashCoxa with 4ndash8 se-tae basis with 12ndash17 setae endopod 4-seg-mented (3 or 4) (1 or 2) (1ndash3) (3 or 4) se-tae exopod 2-segmented proximal segment
without setae distal segment with 5 or 6 se-tae on distal margin bilobed epipod withoutsetae all setae plumose
Maxilliped 2 (Fig 3h)mdashCoxa with 1 or 2 se-tae basis with 3ndash6 setae endopod 4-seg-mented (4 or 5) (0 or 1) (1 or 2) (3ndash6) se-tae exopod unsegmented 6ndash8 setae on dis-tal margin all setae plumose
Fig 2 Callichirus islagrande dorsal view of larvae and decapodid a ZI stage b ZII stage c ZIII stage d ZIVstage e decapodid stage Scale = 1 mm
104 JOURNAL OF CRUSTACEAN BIOLOGY VOL 20 NO 1 2000
Maxilliped 3 (Fig 3i)mdashCoxa with 0 or 1seta basis with 1 or 2 setae endopod 4-seg-mented (0 or 1) (0ndash2) (2 or 3) (3ndash5) se-tae exopod unsegmented 6ndash8 setae on dis-tal margin all setae plumose
PereiopodsmdashNot developed
PleopodsmdashNot developed
Telson (Fig 3j)mdashNarrow anterior not dif-ferentiated from abdominal segment 6 trian-gular posterior anal spine present with(12ndash14)+1+(13 or 14) processes on posterior
margin arranged as (I II III IV III II I)(I) outermost process a plumodenticulate seta(II) second process ldquoanomuran hairrdquo (III)10ndash12 plumodenticulate setae and (IV) cen-ter process a naked spine longer than otherprocesses
Zoea IIFigs 2b 4andashl
SizemdashCL = 144 plusmn 003 mm range 138ndash153mm TL = 364 plusmn 006 mm range 355ndash380mm
Fig 3 Callichirus islagrande ZI stage a antennule b antenna c right mandible d left mandible e maxillulef maxilla (plumose condition not shown) g maxilliped 1 h maxilliped 2 i maxilliped 3 j telson Scales = 02mm
STRASSER AND FELDER LARVAL DEVELOPMENT IN CALLICHIRUS ISLAGRANDE 105
Carapace (Fig 2b)mdashWith antennal spinesanterolateral margins serrated rostrum broadand dorsoventrally flattened with thin distalprojection and distolateral serrations eyesseparated from carapace
Abdomen (Fig 2b)mdashNo marked change fromstage ZI
Antennule (Fig 4a)mdashElongate weaklybilobed peduncle weakly 2-segmented 0ndash3short setae on distal margin of proximal seg-ment 2 long plumose setae and 0ndash3 short se-tae on distal end of distal segment endopodwith 1 terminal plumose seta exopodite with6ndash8 terminal aesthetascs
Antenna (Fig 4b)mdashProtopod with 2 distalspines 1 on outer margin and 1 between ramiendopod without setae scaphocerite armedwith 1 strong distolateral spine 10 or 11plumose setae on inner margin and 1plumose seta on outer margin
Mandible (Fig 4c d)mdashNo marked changefrom ZI
Maxillule (Fig 4e)mdashCoxal endite with 8ndash10marginal plumose setae basal endite with 3plumose setae 4 or 5 large dentate spines and4 or 5 smaller dentate spines inner marginwith 0 or 1 plumose seta endopod 3-seg-mented distinction between segments some-times obscure 2 2 (4 or 5) setae protopodwithout setae
Maxilla (Fig 4f )mdashCoxal endite bilobed12ndash15 setae on proximal lobe 4 or 5 setaeon distal lobe basal endite bilobed with 6ndash8setae on proximal lobe 5ndash9 setae on distallobe endopod with 5 lobes (2 or 3)+2+2+2+3setae scaphognathite with 19ndash24 setae allsetae plumose
Maxilliped 1 (Fig 4g)mdashCoxa with 4ndash8 se-tae basis with 13ndash17 marginal setae endo-pod 4-segmented (3 or 4) (2 or 3) (1 or 2)4 setae exopod 2-segmented proximal seg-ment without setae distal segment with 6 or7 setae on distal margin bilobed epipod with-out setae all setae plumose
Maxilliped 2 (Fig 4h)mdashCoxa with 0ndash2 se-tae basis with 4ndash6 setae endopod 4-seg-mented (4ndash7) 1 (2 or 3) (4ndash6) setae exo-pod unsegmented 7 or 8 setae on distal mar-gin all setae plumose
Maxilliped 3 (Fig 4i)mdashCoxa without setaebasis with 0ndash2 setae endopod 4-segmented
(l or 2) (1ndash3) (2ndash4) (3ndash6) setae exopod un-segmented 6ndash8 setae on distal margin all se-tae plumose
Pereiopods (Fig 4j k)mdashFirst and second pe-reiopod present both without setae
PleopodsmdashNot developed
Telson (Fig 4l)mdashNo change from ZI
Zoea IIIFigs 2c 5andashh 6andashh
SizemdashCL = 165 plusmn 003 mm range 160ndash172mm TL = 413 plusmn 009 mm range 399ndash436mm
Carapace (Fig 2c)mdashNo marked changefrom ZII
Abdomen (Fig 2c)mdashSomites 2ndash5 with pos-terolateral projections dorsal spine elongateon somite 2 sometimes extending beyondposterior margin of somite 3 short dorsalspines on somites 3ndash6
Antennule (Fig 5a)mdashBiramous peduncle2-segmented proximal segment with (0 or1)+(2 or 3) short setae on outer margin and1 long plumose seta on inner margin distalsegment with 2 or 3 setae on inner margin 1seta between rami and 0ndash3 short seta onouter margin endopod with 0 or 1 terminalplumose seta exopod with 6 or 7 terminaland 0ndash2 subterminal aesthetascs
Antenna (Fig 5b)mdashProtopod with 1 spineat outer distal margin endopod with 0 or 1short seta scaphocerite armed with 1 strongdistolateral spine 11ndash13 plumose setae on in-ner margin and 1 plumose seta on outer mar-gin
Mandible (Fig 5c d)mdashIncisor process with7 or 8 teeth on right mandible 6ndash8 teeth onleft mandible molar process denticulate onboth mandibles buds of palps present
Maxillule (Fig 5e)mdashCoxal endite with10ndash12 marginal plumose setae basal enditewith 3 plumose setae 4 or 5 large dentatespines and 4 or 5 smaller dentate spinesinner margin with 1 plumose seta endopod3-segmented distinction between segmentssometimes obscure 2 2 4 setae protopodwithout setation
Maxilla (Fig 5f )mdashCoxal endite bilobed14ndash17 setae on proximal lobe 4ndash6 setae ondistal lobe basal endite bilobed with 7ndash10 se-
106 JOURNAL OF CRUSTACEAN BIOLOGY VOL 20 NO 1 2000
tae on proximal lobe 6ndash9 setae on distal lobeendopod with 5 lobes 3+2+2+2+(2 or 3) se-tae scaphognathite with 21ndash26 setae all se-tae plumose
Maxilliped 1 (Fig 5g)mdashCoxa with 4ndash9 se-tae basis with 14ndash18 marginal setae endo-pod 4-segmented (3 or 4) 2 2 (3 or 4) se-tae exopod 2-segmented proximal segment
without setae distal segment with 5ndash7 setaeon distal margin bilobed epipod without se-tae all setae plumose
Maxilliped 2 (Fig 5h)mdashCoxa with 0ndash2 se-tae basis with 5 or 6 setae endopod 4-seg-mented (4ndash6) 1 3 (4ndash6) setae exopod un-segmented 7 or 8 setae on distal margin allsetae plumose
Fig 4 Callichirus islagrande ZII stage a antennule b antenna c right mandible d left mandible e maxillulef maxilla (plumose condition not shown) g maxilliped 1 h maxilliped 2 i maxilliped 3 j pereiopod 1 k pe-reiopod 2 1 telson Scales = 02 mm
STRASSER AND FELDER LARVAL DEVELOPMENT IN CALLICHIRUS ISLAGRANDE 107
Maxilliped 3 (Fig 6a)mdashCoxa with 0 or 1seta basis with 1ndash3 setae endopod 4-seg-mented (1 or 2) 2 (2ndash4) (4 or 5) setae exo-pod unsegmented 7 or 8 setae on distal mar-gin all setae plumose
Pereiopod 1 (Fig 6b)mdashChelate and bira-mous coxa without setae basis with 0 or 1seta endopod 4-segmented 0 (1 or 2) (2ndash4)2 setae exopod with 6 or 7 setae on distalmargin
Fig 5 Callichirus islagrande ZIII stage a antennule b antenna c right mandible d left mandible e maxil-lule f maxilla (plumose condition not shown) g maxilliped 1 h maxilliped 2 All setae on the maxilla were plumoseScales = 02 mm
108 JOURNAL OF CRUSTACEAN BIOLOGY VOL 20 NO 1 2000
Pereiopod 2 (Fig 6c)mdashChelate and biramouscoxa without setae basis with 0 or 1 seta en-dopod 4-segmented 0 (0ndash2) (0ndash4) 2 setaeexopod with 5ndash7 setae on distal margin
Pereiopods 3ndash5 (Fig 6d e f )mdashWithout setae
Pleopods (Figs 1c 6g)mdashBilobed buds onabdominal segments 2ndash5 first pair smallerthan others
Telson and Uropods (Fig 6h)mdashNo changein arrangement of posterior telsonal processesuropods biramous endopod without setae
Fig 6 Callichirus islagrande ZIII stage a maxilliped 3 b pereiopod 1 c pereiopod 2 d pereiopod 3 e pe-reiopod 4 f pereiopod 5 g pleopods from fourth abdominal somite h telson Scales = 02 mm
STRASSER AND FELDER LARVAL DEVELOPMENT IN CALLICHIRUS ISLAGRANDE 109
exopod with 10 or 11 marginal plumose se-tae ventral spine near base of protopod
Zoea IVFigs 2d 7andashh 8andashi
SizemdashCL = 192 plusmn 009 mm range 169ndash215mm TL = 482 plusmn 025 mm range 424ndash536mm
Carapace (Fig 2d)mdashNo marked changefrom ZIII
Abdomen (Fig 2d)mdashNo marked change fromZIII
Antennule (Fig 7a)mdashBiramous peduncle2-segmented with 1+(1ndash4) short setae onouter margin and 0 or 1 plumose seta on in-ner margin of proximal segment distal seg-ment with 3ndash5 plumose setae on inner mar-gin 1 plumose seta between rami and 0ndash3short setae on outer margin endopod with 1or 2 plumose setae exopod with 6 or 7 ter-minal and 2 subterminal aesthetascs
Antenna (Fig 7b)mdashProtopod with 1 distalspine on outer margin endopod with 0ndash2 se-tae scaphocerite armed with 1 strong disto-lateral spine 12ndash14 plumose setae on innermargin and 1 plumose seta on outer margin
Mandible (Fig 7c d)mdashIncisor process with8 teeth on right mandible 6ndash8 teeth on leftmandible molar process denticulate on bothmandibles palps present
Maxillule (Fig 7e)mdashCoxal endite with12ndash14 marginal plumose setae basal enditewith 3 plumose setae 5 or 6 large dentatespines and 5 or 6 smaller dentate spinesinner margin with 1 plumose seta endopod3-segmented distinction between segmentssometimes obscure 2 2 (3 or 4) setae pro-topod without setation
Maxilla (Fig 7f )mdashCoxal endite bilobed14ndash19 setae on proximal lobe 5 setae on dis-tal lobe basal endite bilobed 6ndash10 setae onproximal lobe 8 or 9 setae on distal lobeendopod with 5 lobes 3+(2 or 3)+2+2+3 se-tae scaphognathite with 27ndash32 setae all se-tae plumose
Maxilliped 1 (Fig 7g)mdashCoxa with 2ndash8 se-tae basis with 16ndash19 marginal setae endo-pod 4-segmented (3 or 4) 2 2 4 setae exo-
pod 2-segmented proximal segment withoutsetae distal segment with 6 or 7 setae on dis-tal margin bilobed epipod without setae allsetae plumose
Maxilliped 2 (Fig 7h)mdashCoxa with 0ndash3 se-tae basis with 4ndash6 setae endopod 4-seg-mented (5ndash8) 1 (3ndash5) (4 or 5) setae exo-pod unsegmented 7 or 8 setae on distal mar-gin all setae plumose
Maxilliped 3 (Fig 8a)mdashCoxa with 0 or 1seta basis with 2ndash4 setae endopod 4-seg-mented (l or 2) 2 (3ndash5) (3 or 4) setae exo-pod unsegmented 7 or 8 setae on distal mar-gin all setae plumose
Pereiopod 1 (Fig 8b)mdashBiramous cheliformcoxa without setae basis with 0ndash2 setae en-dopod 4-segmented (0 or 1) (1 or 2) (2ndash4)(1 or 2) setae exopod with 5ndash7 setae on dis-tal margin
Pereiopod 2 (Fig 8c)mdashBiramous cheliformcoxa without setae basis with 0 or 1 setaendopod 4-segmented 0 (1 or 2) (3ndash5) (2or 3) setae exopod with 5ndash7 setae on distalmargin
Pereiopod 3 (Fig 8d)mdashBiramous coxa andbasis without setae endopod 4-segmented0 (0ndash2) (3 or 4) 2 setae propodus blade-like exopod with 4 or 5 terminal setae
Pereiopod 4 (Fig 8e)mdashBiramous coxa andbasis without setae endopod 4-segmented (0or 1) (0ndash2) (2ndash4) 2 setae exopod with 0ndash2setae
Pereiopod 5 (Fig 8f )mdashUniramous 2 setaeon terminal segment 4 or 5 setae on adja-cent segment
Pleopods (Fig 8g h)mdashLarger than at ZIIIstill without setae
Telson and Uropods (Fig 8i)mdashTelson trian-gular narrow anteriorly no anal spine pos-terior margin with (12ndash15)+1+(13 or 14)processes arranged as (I II III IV III II I)(I) outermost process a plumodenticulate seta(II) second process ldquoanomuran hairrdquo (III) 10ndash13plumodenticulate setae and (IV) naked spineuropods biramous endopod with 7ndash11 marginalplumose setae exopod with 1 naked distolat-eral spine 12ndash14 marginal plumose setae
110 JOURNAL OF CRUSTACEAN BIOLOGY VOL 20 NO 1 2000
Decapodid (first postlarva)Figs 2e 9cndashj l0andashi
SizemdashCL = 133 plusmn 002 mm range129ndash138 mm TL = 378 plusmn 011 mm range361ndash405 mm
Carapace (Fig 2e)mdashShorter than abdomenno anterolateral serrations with pterygostomial
projections rostrum reduced without serra-tions sometimes bending down between eyes
Abdomen (Fig 2e)mdashSomites broad and flat-tened posterior projections rounded with 0ndash7setae somite 2 rectangular somites 3ndash5 oval
Antennule (Fig 9c)mdashPeduncle 3-segmentedproximal segment with 17ndash24 setae 13ndash15 of
Fig 7 Callichirus islagrande ZIV stage a antennule b antenna c right mandible d left mandible e maxil-lule f maxilla (plumose condition not shown) g maxilliped 1 h maxilliped 2 All setae on the maxilla were plumoseScales = 02 mm
STRASSER AND FELDER LARVAL DEVELOPMENT IN CALLICHIRUS ISLAGRANDE 111
these associated with statocyst intermediatesegment with 6ndash10 setae distal segment of pe-duncle with 2 vertical rows of setae one with4 or 5 setae other with 2ndash4 setae 1ndash3 addi-tional setae on outer margin and 3ndash5 short se-tae on distal margin endopod 4ndash6 segmented(1ndash3) (0 or 1) (1ndash4) (1ndash4) (3ndash6) (3ndash6) se-tae exopod 3ndash5-segmented 0 (0ndash4) (0ndash4)(1ndash4) setae +2 asthetascs (5ndash8) asthetascs ()
Antenna (Fig 9d)mdashPeduncle 2-segmentedproximal segment with 0 or 1 seta distal seg-ment with 1ndash3 setae on inner margin spinepresent in some flagellum ranging from 14ndash18
segments (0 or 1) (2ndash4) variable region(1ndash3 articles with 0ndash2 setae) (0ndash4) (0ndash2)(2ndash5) (1ndash4) (3ndash6) (0ndash3) (2ndash5) (3ndash6)(0ndash3) (3ndash6) (0 or 1) (1ndash7) (4ndash9) setae exo-pod reduced to small lobe with 0ndash5 setae
Mandible (Fig 9e f )mdashBoth mandibles sim-ilar incisor process with 5ndash8 teeth molarprocess smooth or with small ridges palp3-segmented 0 (0 or 1) (8ndash12) setae
Maxillule (Fig 9g)mdashCoxal endite with15ndash22 marginal setae and 0 or 1 submarginal
Fig 8 Callichirus islagrande ZIV stage a maxilliped 3 b pereiopod 1 c pereiopod 2 d pereiopod 3 e pe-reiopod 4 f pereiopod 5 g pleopods from first abdominal somite h pleopods from fourth abdominal somite i tel-son Scales = 02 mm
112 JOURNAL OF CRUSTACEAN BIOLOGY VOL 20 NO 1 2000
seta basal endite with 3ndash5 plumose setae14ndash19 dentate spines 0ndash2 plumose setae oninner margin endopod reduced unsegmentedwith 0ndash8 setae
Maxilla (Fig 9h)mdashCoxal endite trilobed(17ndash21)+(7ndash10)+(6ndash9) setae basal enditebilobed 8ndash10 setae on proximal lobe 8ndash15setae on distal lobe endopod reduced with
Fig 9 Callichirus islagrande ZV stage a one example of the antenna b one example of pleopods Decapodidstage c antennule d antenna e right mandible f left mandible g maxillule h maxilla (plumose condition notshown) i maxilliped 1 j maxilliped 2 Scales = 02 mm
STRASSER AND FELDER LARVAL DEVELOPMENT IN CALLICHIRUS ISLAGRANDE 113
5ndash11 setae scaphognathite with 37ndash45 se-tae all setae plumose
Maxilliped 1 (Fig 9i)mdashCoxa with 8ndash12 se-tae basis highly setose with 27ndash39 setae en-dopod reduced with 0ndash11 setae exopod un-segmented 6ndash14 setae bilobed epipod with-out setae
Maxilliped 2 (Fig 9j)mdashCoxa partially fusedto basis with 1ndash4 setae basis with 3ndash5 se-
tae endopod 4-segmented (16ndash23) (0 or 1)(6ndash11) (4ndash8) setae exopod with 0ndash8 setaeepipod present but without setae
Maxilliped 3 (Fig 10a)mdashCoxa partiallyfused to basis with 0ndash3 setae basis with 1ndash4setae endopod 5-segmented (11ndash19) (9ndash15)(7ndash11) (8ndash21) (5ndash11) setae exopod usuallyreduced and without setae occasionally with2ndash9 setae epipod present
Fig 10 Callichirus islagrande decapodid stage a maxilliped 3 b pereiopod 1 c pereiopod 2 d pereiopod 3e pereiopod 4 f pereiopod 5 g pleopods from first abdominal somite h pleopods from fourth abdominal somitei telson Scales = 02 mm
114 JOURNAL OF CRUSTACEAN BIOLOGY VOL 20 NO 1 2000
Pereiopod 1 (Fig 10b)mdashChelate biramous7-segmented coxa with 0ndash6 setae basis with2ndash5 setae ischium with 4ndash7 setae and 0ndash3spines merus with 5ndash8 setae carpus with10ndash14 setae propodus with 27ndash37 setae and3ndash4 spines on inner margin of immovable fin-ger dactylus with 21ndash33 setae exopod usu-ally reduced and without setae occasionallywith 4ndash7 setae
Pereiopod 2 (Fig 10c)mdashChelate biramous7-segmented coxa with 2ndash5 setae basis with1ndash5 setae ischium with 4ndash10 setae meruswith 15ndash20 setae carpus with 8ndash12 setaepropodus with 30ndash40 setae and 1ndash4 spineson inner margin of immovable finger dacty-lus with 21ndash30 setae exopod usually reduced0ndash7 setae
Pereiopod 3 (Fig 10d)mdashBiramous 7-seg-mented coxa with 2ndash5 setae basis with 1ndash3setae ischium with 5ndash8 setae merus with6ndash9 setae carpus with 7ndash11 setae propodusbladelike with 25ndash34 setae 1 spine on innerdistal margin dactylus with 15ndash21 setae exo-pod usually reduced 0ndash5 setae
Pereiopod 4 (Fig 10e)mdashBiramous 7-seg-mented coxa with 2ndash7 setae basis with 1ndash4setae ischium with 3ndash8 setae merus with5ndash11 setae carpus with 4ndash9 setae propoduswith 27ndash38 setae 1 spine on inner distal mar-gin dactylus with 15ndash19 setae exopod usu-ally reduced with 0ndash2 setae
Pereiopod 5 (Fig 10f )mdashCheliform unira-mous 7-segmented coxa with 2ndash6 setae ba-sis with l or 2 setae ischium with 2ndash5 setaemerus with 5ndash9 setae carpus with 4ndash7 se-tae propodus with 22ndash30 setae 1 spine onimmovable finger dactylus with 6ndash10 setaeexopod absent
Pleopods (somite 2) (Fig 10g)mdashBiramouscoxa partially fused to basis without setaebasis with 0ndash3 setae endopod with 3ndash5hooked setae on appendix interna 10ndash17 mar-ginal plumose setae 0ndash2 submarginal nakedsetae exopod with 21ndash27 marginal plumosesetae and 0ndash1 submarginal naked seta
Pleopods (somites 3ndash5) (Fig 10h)mdashBiramouscoxa without setae basis with 3ndash6 marginalplumose setae endopod with 3ndash6 hooked se-tae on appendix interna 10ndash18 marginalplumose setae and 0ndash3 naked submarginal se-tae exopod with 20ndash29 marginal plumose se-tae and 0 or 1 submarginal naked seta
Telson and Uropods (Fig 10i)mdashWith 24ndash36processes on posterior margin median pro-cess usually absent or reduced uropods bi-ramous endopod with 19ndash25 plumose setaeexopod with 35ndash58 plumose setae protopodwith 3ndash5 setae
DISCUSSION
As in other species of Callichirus (see Ro-drigues 1976 Aste and Retamal 1983Strasser and Felder in press b) larval devel-opment of C islagrande included four or fivezoeal stages before metamorphosis to the de-capodid ZI larvae may be identified by thepresence of an elongate rounded rostrumsessile eyes and a lack of pereiopodspleopods or uropods (Fig 1a) At ZII lar-vae acquired biramous buds of pereiopods 1and 2 eyes were separated from the carapaceand the rostrum became flattened and serratedlaterally (Fig 1b) Buds of pleopods on ab-dominal segments 2ndash5 pereiopods 3ndash5 anduropods were developed at ZIII (Fig 1c) Inaddition to further development of structuresmentioned thus far ZIV larvae could be dis-tinguished by presence of a distal spine on theexopod and setae on the endopod of each uro-pod (Fig 1d) When it occurred ZV typicallyresembled ZIV but varied widely in mor-phology Appendages at ZV resembled thoseof ZIV the decapodid stage or some grada-tion between the two (see Figs 9a b for ex-amples) The decapodid stage was similar toother species of Callichirus in the reductionof larval characters such as the elongate ros-trum dorsal spines on the abdomen and ex-opods of the pereiopods (Fig le) Acquisitionof fan-like pleopods marked the transition toa primarily abdominal mode of swimming atthis stage
In the Florida Atlantic population of C ma-jor both salinity and presence of putative set-tlement cues at ZIV impacted the observednumber of zoeal stages and the duration ofZIV (Strasser and Felder in preparation1999c) In C islagrande the percentage oflarvae that passed through five versus fourzoeal stages varied (9ndash89) between parentalfemales but was not shown to depend onsalinity or presence of various putative set-tlement cues (Strasser and Felder 1998 inpreparation) Larvae that molted from ZIVto ZV were usually normal morphologicallyand had mortality similar to those that molteddirectly from ZIV to D In contrast larvae of
STRASSER AND FELDER LARVAL DEVELOPMENT IN CALLICHIRUS ISLAGRANDE 115
the Florida Atlantic C major that molted toZV had a higher incidence of mortality anddeformity at D than did those that had by-passed ZV (Strasser and Felder 1999a c)
The number of zoeal stages appears to beless variable in Callichirus garthi and theGulf of Mexico population of C major thanin congeneric populations studied to dateCallichirus garthi was reported to have fivezoeal stages and a long larval duration reach-ing the decapodid stage 48 days after hatch-ing (Aste and Retamal 1983) Developmentof the Gulf population of C major wasshorter in duration (decapodid reached about10 days after hatching) and typically includedfour zoeal stages (Strasser and Felder 1999a)animals rarely passed through three or fivezoeal stages before the molt to the decapodidAlthough the number of stages is reducedfrom that observed in C garthi developmentin the Gulf population of C major cannot beclassified as abbreviated There are species inboth the Callianassinae (Callianassa slkraussi Stebbing C sl kewalramanii and Csl tyrrhena (Petagna)) and Callichirinae(Sergio mirim Lepidophthalmus sinuensisand L louisianensis) that have only two zoealstages before the molt to the decapodid(Forbes 1973 Sankolli and Shenoy 1975Rodrigues 1984 Thessalou-Legaki 1990Nates et al 1997)
Compared to other species of the genusCallichirus C islagrande is morphologicallycloser to C garthi (from Chile) than to theGulf of Mexico population of C major whichoverlaps in distribution with C islagrande Inthe zoeal phase of C islagrande and Cgarthi the telson is concave and the outer-most process on the ventral margin is articu-lated In both C major and Callichirus sp theposterior margin of the telson is straight andthe outermost process is unarticulated duringthe zoeal stages (Rodrigues 1976 Strasserand Felder 1999a) There are also differencesin spination of the abdomen In zoeae of Cislagrande and C garthi the dorsal spine ofabdominal segment 3 is not longer than thosefound on segments 4 and 5 (Aste and Reta-mal 1983) In contrast the dorsal spine of ab-dominal segment 3 is approximately twice aslong as the dorsal spines on segments 4 and5 in early zoeal stages of C major and Cal-lichirus sp (Rodrigues 1976 Aste and Re-tamal 1983 Strasser and Felder 1999a) Thegreatest difference between these groups is
the number of pleopod pairs at zoeal and de-capodid stages Callichirus islagrande and Cgarthi have pleopods on abdominal segments2ndash5 while pleopods are present only on ab-dominal segments 3ndash5 in C major and Cal-lichirus sp (Rodrigues 1976 Aste and Re-tamal 1983 Strasser and Felder 1999a) Al-though Callichirus islagrande and C garthiare similar morphologically there are differ-ences between these two species Callichirusislagrande as well as C major and Cal-lichirus sp develops pleopod buds andchelate pereiopods at ZIII whereas C garthidoes not acquire these characters until ZIV
There are few callianassid species forwhich the larval development has been de-scribed (Sankolli and Shenoy 1975 Ro-drigues 1976 1984 Aste and Retamal 19831984 Konishi et al 1990 Nates et al 1997Strasser and Felder 1999a) all of which aremembers of the Callichirinae and Cal-lianassinae Among species there is a greatdeal of overlap in larval characters makingit difficult to draw distinctions between thesesubfamilies At present information on lar-val development of species in the Callichiri-nae is limited to larval descriptions of Cal-lichirus major C garthi Callichirus sp Cislagrande Sergio mirim Lepidophthalmuslouisianensis and L sinuensis (see Ro-drigues 1976 1984 Aste and Retamal 1983Nates et al 1997 Strasser and Felder1999a) As mentioned above species of Cal-lichirus have been shown to have four to fivezoeal stages while the other members exhibitabbreviated development that consists of onlytwo zoeal stages The most obvious differencebetween larvae of the species of Callichirinaewith abbreviated development is the numberof pleopod pairs Larvae of S mirim havepleopods on abdominal segments 3ndash5whereas L louisianensis and L sinuensishave pleopods on segments 2ndash5 (Rodrigues1984 Nates et al 1997)
Ghost shrimps of the genus Callichirus maybe distinguished from other genera of the Cal-lianassidae by the cuticular pattern on the dor-sal surface of the abdomen and by the strap-like shape of the uropodal endopods (Manningand Felder 1991) Whereas these charactersmay be used to distinguish adults of the genusCallichirus they are not present at zoeal anddecapodid stages Larvae may be distin-guished from all other callianassid genera byhaving four to five zoeal stages and numer-
116 JOURNAL OF CRUSTACEAN BIOLOGY VOL 20 NO 1 2000
ous processes (12ndash25+1+12ndash25) on the pos-terior margin of the telson during the earlyzoeal stages Larvae of Callichirus may alsodiffer from other callianassids with extendeddevelopment by the retention of the anomu-ran hair as the second process on the poste-rior margin of the telson in the late zoealstages Whereas this process is converted toa spine in Neotrypaea uncinata at ZV and Cal-lianassa sl petalura at ZIV the anomuranhair is retained at ZIV in Callichirus majorand C islagrande (see Rodrigues 1976Strasser and Felder 1999a) Although notmentioned specifically in the descriptions theanomuran hair appears to be present in illus-trations of Callichirus sp at ZIII and Cgarthii at ZV (Rodrigues 1976 fig 35 Asteand Retamal 1983 fig 8Q) A bilobed epi-pod on the first maxilliped of ZI is consistentlyfound in Callichirus but is also present at ZIin Lepidophthalmus sinuensis (see Nates etal 1997) Other callianassids acquire thischaracter at ZII (L louisianensis Nates et al1997 Callianassa sl kewlramanii Sankolliand Shenoy 1975) at ZV (Neotrypaea unci-nata Aste and Retamal 1984) or at the de-capodid stage (Sergio mirim Rodrigues 1984C sl petalura Konishi et al 1990) Thusthe number of zoeal stages and setation of thetelson may serve as the best characters to dis-tinguish larvae of Callichirus from other gen-era However since larval descriptions arelacking for many callianassid species it is un-known if these characters (in combination)will prove to be diagnostic until descriptionsfor other species are published
ACKNOWLEDGEMENTS
We thank Robert Bourgeois Barbara Hasek JimMaKinster Sergio Nates and Kurt Strasser who pro-vided assistance in the field and laboratory and MaryE Rice Director of the Smithsonian Marine StationLink Port Florida who facilitated access to station fa-cilities We also thank R Bauer R Jaeger R Lemaitreand G Watson for their early comments on this manu-script and Raymond B Manning who facilitated fund-ing and gave advice on collecting sites Direct supportfor this study was provided to Karen M Strasser by adoctoral fellowship from the University of Louisianaa graduate student research grant from the LUMCON(Louisiana Universities Marine Consortium) Founda-tion and a summer research award from the Smith-sonian Marine Station Additional support for field andlab activities was provided to D L Felder under Loui-siana Sea Grant College Program Grant RCFB-21 USFish and Wildlife Service Cooperative Agreement 14-16-0009-89-96 (Task Order No 6) US Department of En-ergy grant DE-FG02-97ER12220 and several small proj-ect grants from the Smithsonian Marine Station Views
expressed herein are those of the authors and do not nec-essarily reflect views of supporting government agenciesor subagencies mentioned above This is contributionNumber 466 of the Smithsonian Marine Station and con-tribution Number 67 of the ULL Laboratory for Crus-tacean Research All experiments conducted in this studycomply with current applicable state and federal laws
LITERATURE CITED
Aste A and M Retamal 1983 Desarrollo larval deCallianassa garthi Retamal 1975 bajo condiciones delaboratoriomdashCiencia y Tecnologiacutea del Mar CONA 75ndash26
mdashmdashmdash and mdashmdashmdash 1984 Desarrollo larval de Cal-lianassa uncinata H Milne Edwards1837 (DecapodaCallianassidae) bajo condiciones de laboratoriomdashGayana Zooloacutegica 48 41ndash56
Felder D L 1978 Osmotic and ionic regulation in sev-eral western Atlantic Callianassidae (Crustacea De-capoda Thalassinidea)mdashBiological Bulletin 154409ndash429
mdashmdashmdash and R B Griffis 1994 Dominant infaunalcommunities at risk in shoreline habitats burrowingthalassinid CrustaceamdashOCS Study number MMS94ndash0007 United States Department of the InteriorMinerals Management Service Gulf of Mexico OCSRegional Office New Orleans Louisiana Pp 1ndash87
Forbes A T 1973 An unusual abbreviated larval lifehistory in the estuarine burrowing prawn Callianassakraussi (Crustacea Decapoda Thalassinidea)mdashMa-rine Biology 22 361ndash365
Konishi K 1989 Larval development of the mudshrimp Upogebia (Upogebia) major (de Haan) (Crus-tacea Thalassinidea Upogebiidae) under laboratoryconditions with comments on larval characters of tha-lassinid familiesmdashBulletin of the National ResearchInstitute of Aquaculture 15 1ndash17
mdashmdashmdash R Quintana and Y Fukuda 1990 A completedescription of larval stages of the ghost shrimp Cal-lianassa petalura Stimpson (Crustacea ThalassinideaCallianassidae) under laboratory conditionsmdashBulletinof the National Research Institute of Aquaculture 1727ndash49
Manning R B and D L Felder 1986 The status ofthe callianassid genus Callichirus Stimpson 1866(Crustacea Decapoda Thalassinidea)mdashProceedingsof the Biological Society of Washington 99 437ndash443
mdashmdashmdash and mdashmdashmdash 1991 Revision of the AmericanCallianassidae (Crustacea Decapoda Thalassinidea)mdashProceedings of the Biological Society of Washington104 764ndash794
Nates S F D L Felder and R Lemaitre 1997 Com-parative larval development in two species of the bur-rowing ghost shrimp genus Lepidophthalmus (Decap-oda Callianassidae)mdashJournal of Crustacean Biology17 497ndash519
Rodrigues S de A 1976 Sobre a reproduccedilatildeo embri-ologia e desenvolvimento larval de Callichirus majorSay 1818 (Crustacea Decapoda Thalassinidea)mdashBo-letim de Zoologia Universidade de Satildeo Paulo 185ndash104
mdashmdashmdash 1984 Desenvolvimento poacutes-embrionaacuterio deCallichirus mirim (Rodrigues 1971) obtido emcondiccedilotildees artificiais (Crustacea Decapoda Tha-lassinidea)mdashBoletim de Zoologia Universidade deSatildeo Paulo 8 239ndash256
STRASSER AND FELDER LARVAL DEVELOPMENT IN CALLICHIRUS ISLAGRANDE 117
Sankolli K N and S Shenoy 1975 Larval develop-ment of mud shrimp Callianassa (Callichirus) kewal-ramanii Sankolli in the laboratory (Crustacea Decap-oda)mdashBulletin of the Department of Marine ScienceUniversity of Cochin 4 704ndash720
Staton J L and D L Felder 1995 Genetic variationin populations of the ghost shrimp genus Callichirus(Crustacea Decapoda Thalassinoidea) in the westernAtlantic and Gulf of MexicomdashBulletin of Marine Sci-ence 56 523ndash536
Strasser K M and D L Felder 1998 Settlement cuesin successive developmental stages of the ghost shrimpCallichirus major and C islagrande (Crustacea De-capoda Thalassinidea)mdashMarine Biology
mdashmdashmdash and mdashmdashmdash 1999a Larval development of twopopulations of the ghost shrimp Callichirus major (De-capoda Thalassinidea)mdashJournal of Crustacean Biology
mdashmdashmdash and mdashmdashmdash 1999b Sand as a stimulus for set-tlement in the ghost shrimp Callichirus major (Say)and C islagrande (Schmitt) (Crustacea DecapodaThalassinidea)mdashJournal of Experimental Marine Bi-ology and Ecology 239 211ndash222
mdashmdashmdash and mdashmdashmdash 1999c Settlement cues in theFlorida Atlantic population of the ghost shrimp Cal-lichirus major (Crustacea Decapoda Thalassinidea)mdashMarine Ecology Progress Series
Thessalou-Legaki M 1990 Advanced larval develop-ment of Callianassa tyrrhena (Decapoda Tha-lassinidea) and the effect of environmental factorsmdashJournal of Crustacean Biology 10 659ndash666
RECEIVED 19 September 1998ACCEPTED 3 December 1998

In a recent revision of the Callianassidae(Manning and Felder 1991) the genus Cal-lianassa was restricted from its former world-wide status to certain eastern Atlantic popu-lations based on morphological charactersAmerican species that were previously in-cluded in the genus Callianassa were placedin a number of genera based on life historyand morphology as in the case of the genusCallichirus As redefined by Manning andFelder (1986) the genus Callichirus now in-cludes at minimum C major (Say 1818) Cislagrande (Schmitt 1935) C seilacheri(Bott 1955) C garthi (Retamal 1975) andC adamas (Kensley 1974) and at least oneundescribed species from the Atlantic coastsof Brazil and Colombia which has been of-ten referred to as C major (see Staton andFelder 1995 D L Felder personal obser-vation) For a number of other callianassidspecies generic status remains unresolvedand they will herein be referred to as Cal-lianassa sensu lato (sl)
Detailed descriptions of larvae are avail-able for relatively few species in the familyCallianassidae all of which currently fall intothe subfamilies Callianassinae (Callianassasl kewalramanii Sankolli Sankolli andShenoy 1975 Neotrypaea uncinata (HMilne Edwards) Aste and Retamal 1984 Csl petalura Stimpson Konishi et al 1990)
and Callichirinae (Callichirus sp (formerlyC major) Rodrigues 1976 C garthi (Reta-mal) Aste and Retamal 1983 Sergio mirim(Rodrigues) Rodrigues 1984 Lepidophthal-mus louisianensis (Schmitt) L sinuensisLemaitre and Rodrigues Nates et al 1997Callichirus major Strasser and Felder 1999a)Larval development of Callichirus islagrandehas not been described even though this spe-cies is well studied (Felder and Griffis 1994Staton and Felder 1995 Strasser and Felder1998 1999b)
Callichirus islagrande is a Gulf of Mexicoendemic found typically on barrier islands inlow to high intertidal habitats This speciesinhabits the northern and western Gulf ofMexico ranging from northwestern Florida toParaiso Tabasco Mexico (Staton and Felder1995) A somewhat discontinuous distribu-tion of C islagrande occurs between south-eastern Louisiana and the northeastern coastof Texas where shorelines tend to lack thecleaner quartzite sand and high energybeaches that C islagrande prefers (Staton andFelder 1995)
The present study describes the zoeal anddecapodid stages provides illustrations foridentification and reports the number and du-ration of larval stages of Callichirus isla-grande Comparisons of these characteristicsin C islagrande are made with other con-
JOURNAL OF CRUSTACEAN BIOLOGY 20(1) 100ndash117 2000
LARVAL DEVELOPMENT OF THE GHOST SHRIMP CALLICHIRUSISLAGRANDE (DECAPODA THALASSINIDEA CALLIANASSIDAE)
UNDER LABORATORY CONDITIONS
Karen M Strasser and Darryl L Felder
Department of Biology University of Louisiana Lafayette Louisiana 70504 USA(DLF e-mail dfelderlouisianaedu) (KMS current address) Box 108F University of Tampa 401 W
Kennedy Boulevard Tampa Florida 33606 USA (e-mail kstrasseralphautampaedu)
A B S T R A C T
Descriptions of the zoeal and decapodid stages of the ghost shrimp Callichirus islagrande werebased on laboratory cultures from a population inhabiting the coast of Louisiana Whereas mor-phological studies were based upon animals from mass cultures 40ndash120 larvae from each of 8parental females were reared individually to obtain stage-duration data Larvae passed through 4 or5 zoeal stages before molting to the decapodid stage The proportion of individuals that molted toa fifth zoeal stage varied widely (9ndash89) between clutches of different parental females The firstjuvenile stage was usually reached 16ndash20 days after hatching and durations of larval stages weresimilar in larvae from different females Morphological comparisons with congeneric species re-vealed larvae of C islagrande to be more similar to larvae of C garthi a species from the coastof Chile than to larvae of a Gulf of Mexico population of Callichirus major
100
STRASSER AND FELDER LARVAL DEVELOPMENT IN CALLICHIRUS ISLAGRANDE 101
generic species and members of the familyCallianassidae
MATERIALS AND METHODS
Ovigerous females were collected in March May andJuly 1996 from the bayward side of a barrier island (IslesDernieres Louisiana) with yabby pumps (as in Felder1978) Adult females were maintained in 20-cm diame-ter finger bowls (at 27degC 25 ppt salinity) with daily wa-ter changes until eggs hatched Sea water was taken fromwell offshore Louisiana filtered through a 30-microm screenaerated and diluted to 25 ppt salinity with deionized wa-ter before use in the culture of animals To determinestage durations 40ndash120 larvae from each of 8 parentalfemales were reared individually through the first juve-nile stage (J1) ZI (first zoeal stage) larvae were movedto individual compartments upon hatching and maintainedat 27degC in filtered sea water on a 1212 light dark cy-cle Each day larvae were moved to containers with newsea water fed freshly hatched nauplii of Artemia (GreatSalt Lake) and examined to assess their stage of devel-opment Observations were terminated when animalsreached the first juvenile stage (J1)
Animals used for morphological comparisons were ob-tained from mass cultures Larvae were maintained in 20-cm diameter finger bowls (100ndash200 individuals per bowl)with daily water changes under conditions mentionedabove Animals were fixed in 70 ethanol stained withchlorozol black and transferred to glycerine prior to dis-section At least 10 animals were dissected for each stageand both right and left appendages were used for setalcounts Line illustrations were made on a Nikon invertedmicroscope fitted with a camera lucida Measurementswere made with a calibrated ocular micrometer Carapacelength (CL) was measured from the tip of the rostrum tothe posterior midpoint of the carapace and total length(TL) was measured from the rostral tip to the posteriormidpoint of the telson For each appendage the arrange-ment of setae was listed sequentially from proximal todistal margins as in Konishi (1989) and Nates et al(1997) Setal groups on successive segments were sepa-rated with a comma () Groups of setae on the same seg-ment or on different lobes of the same segment articleor endite were separated with a plus (+) A question mark() was used to designate questionable distinctions be-tween setae and aesthetascs Roman numerals were usedto describe the pattern of processes on the posterior mar-gin of the telson
RESULTS
Although there was no attempt to quantifythe duration of embryonic developmentovigerous females were held for up to 28 daysbefore the eggs hatched (usually at night)Prior to hatching eggs changed gradually incolor from bright orange-red to a translucent-brown Late-stage embryos that were droppedby the female before hatching and larvae thathatched as prezoeae were not viable evenwith aeration Prezoeae did not appear to bedeveloped fully and could not swim All zoealstages fed readily on nauplii of Artemia aswell as on each other The decapodid stage(D) appeared to consume much less than didthe preceding zoeal stage Strong positivephototaxis was exhibited by the first threezoeal (ZI to ZIII) stages The fourth zoealstage (ZIV) in general appeared to be at-tracted to light although some larvae reactedindifferently Decapodids did not exhibit ob-vious phototaxis
Larvae of Callichirus islagrande passedthrough four or five zoeal stages before molt-ing to D (Fig 1) these were usually com-pleted 11ndash14 days after hatching Mean du-rations of each stage were relatively consis-tent between larvae from different females(Table 1) However duration of ZIV was ap-proximately a day shorter when animalsmolted from ZIV to ZV than when animalsmolted directly to D (Table 2) Among theeight cultures observed in this study the per-centage of animals molting to ZV ranged from9ndash89 of the larvae that survived past ZIV
Description of Zoeal and Decapodid Stages
Near the mandibles the larval stages anddecapodid had two red chromatophores whichusually appeared fused when expanded Sev-
Table 1 Mean duration in days (plusmn95 CI) of the first to fifth zoeal stages (ZI to ZV) and the decapodid stage (D)in larval cultures from 8 ovigerous females of Callichirus islagrande Each culture consists of larvae from a differ-ent parental female Only animals that reached J1 (N) were used to calculate mean durations N0 is the starting num-ber of ZI larvae
Culture N N0 ZI ZII ZIII ZIV ZV D
1 100 120 226 (plusmn009) 204 (plusmn005) 222 (plusmn009) 388 (plusmn009) 389 (plusmn012) 570 (plusmn022)2 14 60 207 (plusmn016) 200 (plusmn000) 250 (plusmn031) 336 (plusmn060) 430 (plusmn028) 664 (plusmn117)3 34 55 206 (plusmn008) 206 (plusmn008) 244 (plusmn017) 421 (plusmn024) 420 (plusmn016) 568 (plusmn039)4 102 120 205 (plusmn004) 202 (plusmn005) 232 (plusmn010) 359 (plusmn010) 320 (plusmn009) 518 (plusmn028)5 31 40 205 (plusmn011) 200 (plusmn000) 226 (plusmn016) 323 (plusmn028) 400 (plusmn037) 494 (plusmn023)6 49 60 206 (plusmn007) 204 (plusmn008) 243 (plusmn014) 339 (plusmn020) 364 (plusmn018) 547 (plusmn046)7 36 54 217 (plusmn013) 211 (plusmn011) 214 (plusmn012) 294 (plusmn021) 327 (plusmn015) 439 (plusmn019)8 44 60 216 (plusmn013) 225 (plusmn015) 236 (plusmn017) 316 (plusmn026) 342 (plusmn017) 430 (plusmn030)
102 JOURNAL OF CRUSTACEAN BIOLOGY VOL 20 NO 1 2000
eral other red chromatophores were presenton the lateral edges of the carapace forminga red line when expanded
Morphological descriptions of Callichirusislagrande are as detailed below for stages ZIto ZIV and D ZV resembled ZIV in appear-ance However appendages were like thoseof ZIV D or some gradation between thetwo Setal patterns are given from proximal todistal segments for each appendage Carapacelength (CL) and total length (TL) are given asthe mean plusmn 95 CI in mm followed by therange of measurements from 10 specimens
Zoea IFigs 2a 3andashj
SizemdashCL = 123 plusmn 006 mm range 110ndash141mm TL = 326 plusmn 007 mm range 311ndash336mm
Carapace (Fig 2a)mdashShorter than abdomenrostrum elongate rounded in cross sectionand smooth eyes fused to carapace
Abdomen (Fig 2a)mdashSomites 2ndash5 with pos-terolateral projections and posterodorsalspines dorsal spine elongate on somite 2sometimes extending beyond posterior mar-gin of somite 3 short dorsal spines on somites3ndash5
Antennule (Fig 3a)mdashElongate exopodal andendopodal lobes not distinct 4ndash6 terminalaesthetascs 1 long plumose seta on future en-dopodal lobe
Antenna (Fig 3b)mdashProtopod with 1 distalspine between rami endopod with 2 longplumose setae scaphocerite (exopod) armedwith 1 strong distolateral spine 8 plumose se-tae on inner margin and 1 plumose seta onouter margin
Mandible (Fig 3c d)mdashIncisor process with6 or 7 teeth on right mandible 5ndash7 teeth onleft mandible molar process denticulate onboth mandibles
Maxillule (Fig 3e)mdashCoxal endite with 8ndash10marginal setae basal endite with 3 plumosesetae 3 or 4 large dentate spines and 4 or 5smaller dentate spines inner margin with 0or 1 plumose seta endopod 3-segmented dis-tinction between segments sometimes ob-scure 2 2 (3 or 4) setae protopod withoutsetae
Maxilla (Fig 3f )mdashCoxal endite bilobed 11or 12 setae on proximal lobe and 4 or 5 se-
Table 2 Percentage of larval Callichirus islangrande that molted from the fourth zoeal stage (ZIV) to the fifthzoeal stage (ZV) or directly to the decapodid stage (D) in larval cultures from 8 ovigerous females (Culture num-bers 1ndash8) Each batch consisted of larvae from a different parental female N0 = number surviving beyond ZIV N1
= number molting ZIV to ZV N1J = percentage of N1 surviving to J1 N2 = number molting ZIV to D N2J = per-centage of N2 surviving to J1 The mean duration in days (plusmn95 CI) is given for animals that molted from ZIV toZV and from ZIV to D
From ZIV To ZV To D
Survived to J1 Duration Survived to J1 DurationCulture N0 N1 N1J of ZIV N2 N2J of ZIV
1 105 11 82 30 (plusmn030) 94 97 40 (plusmn008)2 47 42 24 32 (plusmn023) 5 80 44 (plusmn111)3 45 8 62 30 (plusmn00) 37 78 44 (plusmn018)4 109 10 50 31 (plusmn078) 99 98 36 (plusmn010)5 38 19 79 28 (plusmn026) 19 84 37 (plusmn027)6 58 35 80 31 (plusmn017) 23 91 39 (plusmn030)7 48 30 73 26 (plusmn023) 18 78 34 (plusmn025)8 51 29 83 26 (plusmn019) 22 91 38 (plusmn019)
Fig 1 Mean proportion of larvae at each stage duringdevelopment of Callichirus islagrande Mean proportionsare derived from the individual rearings of 8 parental fe-males The first through fifth zoeal stages (ZI to ZV) thedecapodid stage (D) and all first-stage juveniles thatpassed D (J) are labeled under the peak of each line
STRASSER AND FELDER LARVAL DEVELOPMENT IN CALLICHIRUS ISLAGRANDE 103
tae on distal lobe basal edite bilobed with6ndash9 setae on each lobe endopod with 5 lobes3+2+2+2+(2 or 3) setae scaphognathite with16ndash19 setae all setae plumose
Maxilliped 1 (Fig 3g)mdashCoxa with 4ndash8 se-tae basis with 12ndash17 setae endopod 4-seg-mented (3 or 4) (1 or 2) (1ndash3) (3 or 4) se-tae exopod 2-segmented proximal segment
without setae distal segment with 5 or 6 se-tae on distal margin bilobed epipod withoutsetae all setae plumose
Maxilliped 2 (Fig 3h)mdashCoxa with 1 or 2 se-tae basis with 3ndash6 setae endopod 4-seg-mented (4 or 5) (0 or 1) (1 or 2) (3ndash6) se-tae exopod unsegmented 6ndash8 setae on dis-tal margin all setae plumose
Fig 2 Callichirus islagrande dorsal view of larvae and decapodid a ZI stage b ZII stage c ZIII stage d ZIVstage e decapodid stage Scale = 1 mm
104 JOURNAL OF CRUSTACEAN BIOLOGY VOL 20 NO 1 2000
Maxilliped 3 (Fig 3i)mdashCoxa with 0 or 1seta basis with 1 or 2 setae endopod 4-seg-mented (0 or 1) (0ndash2) (2 or 3) (3ndash5) se-tae exopod unsegmented 6ndash8 setae on dis-tal margin all setae plumose
PereiopodsmdashNot developed
PleopodsmdashNot developed
Telson (Fig 3j)mdashNarrow anterior not dif-ferentiated from abdominal segment 6 trian-gular posterior anal spine present with(12ndash14)+1+(13 or 14) processes on posterior
margin arranged as (I II III IV III II I)(I) outermost process a plumodenticulate seta(II) second process ldquoanomuran hairrdquo (III)10ndash12 plumodenticulate setae and (IV) cen-ter process a naked spine longer than otherprocesses
Zoea IIFigs 2b 4andashl
SizemdashCL = 144 plusmn 003 mm range 138ndash153mm TL = 364 plusmn 006 mm range 355ndash380mm
Fig 3 Callichirus islagrande ZI stage a antennule b antenna c right mandible d left mandible e maxillulef maxilla (plumose condition not shown) g maxilliped 1 h maxilliped 2 i maxilliped 3 j telson Scales = 02mm
STRASSER AND FELDER LARVAL DEVELOPMENT IN CALLICHIRUS ISLAGRANDE 105
Carapace (Fig 2b)mdashWith antennal spinesanterolateral margins serrated rostrum broadand dorsoventrally flattened with thin distalprojection and distolateral serrations eyesseparated from carapace
Abdomen (Fig 2b)mdashNo marked change fromstage ZI
Antennule (Fig 4a)mdashElongate weaklybilobed peduncle weakly 2-segmented 0ndash3short setae on distal margin of proximal seg-ment 2 long plumose setae and 0ndash3 short se-tae on distal end of distal segment endopodwith 1 terminal plumose seta exopodite with6ndash8 terminal aesthetascs
Antenna (Fig 4b)mdashProtopod with 2 distalspines 1 on outer margin and 1 between ramiendopod without setae scaphocerite armedwith 1 strong distolateral spine 10 or 11plumose setae on inner margin and 1plumose seta on outer margin
Mandible (Fig 4c d)mdashNo marked changefrom ZI
Maxillule (Fig 4e)mdashCoxal endite with 8ndash10marginal plumose setae basal endite with 3plumose setae 4 or 5 large dentate spines and4 or 5 smaller dentate spines inner marginwith 0 or 1 plumose seta endopod 3-seg-mented distinction between segments some-times obscure 2 2 (4 or 5) setae protopodwithout setae
Maxilla (Fig 4f )mdashCoxal endite bilobed12ndash15 setae on proximal lobe 4 or 5 setaeon distal lobe basal endite bilobed with 6ndash8setae on proximal lobe 5ndash9 setae on distallobe endopod with 5 lobes (2 or 3)+2+2+2+3setae scaphognathite with 19ndash24 setae allsetae plumose
Maxilliped 1 (Fig 4g)mdashCoxa with 4ndash8 se-tae basis with 13ndash17 marginal setae endo-pod 4-segmented (3 or 4) (2 or 3) (1 or 2)4 setae exopod 2-segmented proximal seg-ment without setae distal segment with 6 or7 setae on distal margin bilobed epipod with-out setae all setae plumose
Maxilliped 2 (Fig 4h)mdashCoxa with 0ndash2 se-tae basis with 4ndash6 setae endopod 4-seg-mented (4ndash7) 1 (2 or 3) (4ndash6) setae exo-pod unsegmented 7 or 8 setae on distal mar-gin all setae plumose
Maxilliped 3 (Fig 4i)mdashCoxa without setaebasis with 0ndash2 setae endopod 4-segmented
(l or 2) (1ndash3) (2ndash4) (3ndash6) setae exopod un-segmented 6ndash8 setae on distal margin all se-tae plumose
Pereiopods (Fig 4j k)mdashFirst and second pe-reiopod present both without setae
PleopodsmdashNot developed
Telson (Fig 4l)mdashNo change from ZI
Zoea IIIFigs 2c 5andashh 6andashh
SizemdashCL = 165 plusmn 003 mm range 160ndash172mm TL = 413 plusmn 009 mm range 399ndash436mm
Carapace (Fig 2c)mdashNo marked changefrom ZII
Abdomen (Fig 2c)mdashSomites 2ndash5 with pos-terolateral projections dorsal spine elongateon somite 2 sometimes extending beyondposterior margin of somite 3 short dorsalspines on somites 3ndash6
Antennule (Fig 5a)mdashBiramous peduncle2-segmented proximal segment with (0 or1)+(2 or 3) short setae on outer margin and1 long plumose seta on inner margin distalsegment with 2 or 3 setae on inner margin 1seta between rami and 0ndash3 short seta onouter margin endopod with 0 or 1 terminalplumose seta exopod with 6 or 7 terminaland 0ndash2 subterminal aesthetascs
Antenna (Fig 5b)mdashProtopod with 1 spineat outer distal margin endopod with 0 or 1short seta scaphocerite armed with 1 strongdistolateral spine 11ndash13 plumose setae on in-ner margin and 1 plumose seta on outer mar-gin
Mandible (Fig 5c d)mdashIncisor process with7 or 8 teeth on right mandible 6ndash8 teeth onleft mandible molar process denticulate onboth mandibles buds of palps present
Maxillule (Fig 5e)mdashCoxal endite with10ndash12 marginal plumose setae basal enditewith 3 plumose setae 4 or 5 large dentatespines and 4 or 5 smaller dentate spinesinner margin with 1 plumose seta endopod3-segmented distinction between segmentssometimes obscure 2 2 4 setae protopodwithout setation
Maxilla (Fig 5f )mdashCoxal endite bilobed14ndash17 setae on proximal lobe 4ndash6 setae ondistal lobe basal endite bilobed with 7ndash10 se-
106 JOURNAL OF CRUSTACEAN BIOLOGY VOL 20 NO 1 2000
tae on proximal lobe 6ndash9 setae on distal lobeendopod with 5 lobes 3+2+2+2+(2 or 3) se-tae scaphognathite with 21ndash26 setae all se-tae plumose
Maxilliped 1 (Fig 5g)mdashCoxa with 4ndash9 se-tae basis with 14ndash18 marginal setae endo-pod 4-segmented (3 or 4) 2 2 (3 or 4) se-tae exopod 2-segmented proximal segment
without setae distal segment with 5ndash7 setaeon distal margin bilobed epipod without se-tae all setae plumose
Maxilliped 2 (Fig 5h)mdashCoxa with 0ndash2 se-tae basis with 5 or 6 setae endopod 4-seg-mented (4ndash6) 1 3 (4ndash6) setae exopod un-segmented 7 or 8 setae on distal margin allsetae plumose
Fig 4 Callichirus islagrande ZII stage a antennule b antenna c right mandible d left mandible e maxillulef maxilla (plumose condition not shown) g maxilliped 1 h maxilliped 2 i maxilliped 3 j pereiopod 1 k pe-reiopod 2 1 telson Scales = 02 mm
STRASSER AND FELDER LARVAL DEVELOPMENT IN CALLICHIRUS ISLAGRANDE 107
Maxilliped 3 (Fig 6a)mdashCoxa with 0 or 1seta basis with 1ndash3 setae endopod 4-seg-mented (1 or 2) 2 (2ndash4) (4 or 5) setae exo-pod unsegmented 7 or 8 setae on distal mar-gin all setae plumose
Pereiopod 1 (Fig 6b)mdashChelate and bira-mous coxa without setae basis with 0 or 1seta endopod 4-segmented 0 (1 or 2) (2ndash4)2 setae exopod with 6 or 7 setae on distalmargin
Fig 5 Callichirus islagrande ZIII stage a antennule b antenna c right mandible d left mandible e maxil-lule f maxilla (plumose condition not shown) g maxilliped 1 h maxilliped 2 All setae on the maxilla were plumoseScales = 02 mm
108 JOURNAL OF CRUSTACEAN BIOLOGY VOL 20 NO 1 2000
Pereiopod 2 (Fig 6c)mdashChelate and biramouscoxa without setae basis with 0 or 1 seta en-dopod 4-segmented 0 (0ndash2) (0ndash4) 2 setaeexopod with 5ndash7 setae on distal margin
Pereiopods 3ndash5 (Fig 6d e f )mdashWithout setae
Pleopods (Figs 1c 6g)mdashBilobed buds onabdominal segments 2ndash5 first pair smallerthan others
Telson and Uropods (Fig 6h)mdashNo changein arrangement of posterior telsonal processesuropods biramous endopod without setae
Fig 6 Callichirus islagrande ZIII stage a maxilliped 3 b pereiopod 1 c pereiopod 2 d pereiopod 3 e pe-reiopod 4 f pereiopod 5 g pleopods from fourth abdominal somite h telson Scales = 02 mm
STRASSER AND FELDER LARVAL DEVELOPMENT IN CALLICHIRUS ISLAGRANDE 109
exopod with 10 or 11 marginal plumose se-tae ventral spine near base of protopod
Zoea IVFigs 2d 7andashh 8andashi
SizemdashCL = 192 plusmn 009 mm range 169ndash215mm TL = 482 plusmn 025 mm range 424ndash536mm
Carapace (Fig 2d)mdashNo marked changefrom ZIII
Abdomen (Fig 2d)mdashNo marked change fromZIII
Antennule (Fig 7a)mdashBiramous peduncle2-segmented with 1+(1ndash4) short setae onouter margin and 0 or 1 plumose seta on in-ner margin of proximal segment distal seg-ment with 3ndash5 plumose setae on inner mar-gin 1 plumose seta between rami and 0ndash3short setae on outer margin endopod with 1or 2 plumose setae exopod with 6 or 7 ter-minal and 2 subterminal aesthetascs
Antenna (Fig 7b)mdashProtopod with 1 distalspine on outer margin endopod with 0ndash2 se-tae scaphocerite armed with 1 strong disto-lateral spine 12ndash14 plumose setae on innermargin and 1 plumose seta on outer margin
Mandible (Fig 7c d)mdashIncisor process with8 teeth on right mandible 6ndash8 teeth on leftmandible molar process denticulate on bothmandibles palps present
Maxillule (Fig 7e)mdashCoxal endite with12ndash14 marginal plumose setae basal enditewith 3 plumose setae 5 or 6 large dentatespines and 5 or 6 smaller dentate spinesinner margin with 1 plumose seta endopod3-segmented distinction between segmentssometimes obscure 2 2 (3 or 4) setae pro-topod without setation
Maxilla (Fig 7f )mdashCoxal endite bilobed14ndash19 setae on proximal lobe 5 setae on dis-tal lobe basal endite bilobed 6ndash10 setae onproximal lobe 8 or 9 setae on distal lobeendopod with 5 lobes 3+(2 or 3)+2+2+3 se-tae scaphognathite with 27ndash32 setae all se-tae plumose
Maxilliped 1 (Fig 7g)mdashCoxa with 2ndash8 se-tae basis with 16ndash19 marginal setae endo-pod 4-segmented (3 or 4) 2 2 4 setae exo-
pod 2-segmented proximal segment withoutsetae distal segment with 6 or 7 setae on dis-tal margin bilobed epipod without setae allsetae plumose
Maxilliped 2 (Fig 7h)mdashCoxa with 0ndash3 se-tae basis with 4ndash6 setae endopod 4-seg-mented (5ndash8) 1 (3ndash5) (4 or 5) setae exo-pod unsegmented 7 or 8 setae on distal mar-gin all setae plumose
Maxilliped 3 (Fig 8a)mdashCoxa with 0 or 1seta basis with 2ndash4 setae endopod 4-seg-mented (l or 2) 2 (3ndash5) (3 or 4) setae exo-pod unsegmented 7 or 8 setae on distal mar-gin all setae plumose
Pereiopod 1 (Fig 8b)mdashBiramous cheliformcoxa without setae basis with 0ndash2 setae en-dopod 4-segmented (0 or 1) (1 or 2) (2ndash4)(1 or 2) setae exopod with 5ndash7 setae on dis-tal margin
Pereiopod 2 (Fig 8c)mdashBiramous cheliformcoxa without setae basis with 0 or 1 setaendopod 4-segmented 0 (1 or 2) (3ndash5) (2or 3) setae exopod with 5ndash7 setae on distalmargin
Pereiopod 3 (Fig 8d)mdashBiramous coxa andbasis without setae endopod 4-segmented0 (0ndash2) (3 or 4) 2 setae propodus blade-like exopod with 4 or 5 terminal setae
Pereiopod 4 (Fig 8e)mdashBiramous coxa andbasis without setae endopod 4-segmented (0or 1) (0ndash2) (2ndash4) 2 setae exopod with 0ndash2setae
Pereiopod 5 (Fig 8f )mdashUniramous 2 setaeon terminal segment 4 or 5 setae on adja-cent segment
Pleopods (Fig 8g h)mdashLarger than at ZIIIstill without setae
Telson and Uropods (Fig 8i)mdashTelson trian-gular narrow anteriorly no anal spine pos-terior margin with (12ndash15)+1+(13 or 14)processes arranged as (I II III IV III II I)(I) outermost process a plumodenticulate seta(II) second process ldquoanomuran hairrdquo (III) 10ndash13plumodenticulate setae and (IV) naked spineuropods biramous endopod with 7ndash11 marginalplumose setae exopod with 1 naked distolat-eral spine 12ndash14 marginal plumose setae
110 JOURNAL OF CRUSTACEAN BIOLOGY VOL 20 NO 1 2000
Decapodid (first postlarva)Figs 2e 9cndashj l0andashi
SizemdashCL = 133 plusmn 002 mm range129ndash138 mm TL = 378 plusmn 011 mm range361ndash405 mm
Carapace (Fig 2e)mdashShorter than abdomenno anterolateral serrations with pterygostomial
projections rostrum reduced without serra-tions sometimes bending down between eyes
Abdomen (Fig 2e)mdashSomites broad and flat-tened posterior projections rounded with 0ndash7setae somite 2 rectangular somites 3ndash5 oval
Antennule (Fig 9c)mdashPeduncle 3-segmentedproximal segment with 17ndash24 setae 13ndash15 of
Fig 7 Callichirus islagrande ZIV stage a antennule b antenna c right mandible d left mandible e maxil-lule f maxilla (plumose condition not shown) g maxilliped 1 h maxilliped 2 All setae on the maxilla were plumoseScales = 02 mm
STRASSER AND FELDER LARVAL DEVELOPMENT IN CALLICHIRUS ISLAGRANDE 111
these associated with statocyst intermediatesegment with 6ndash10 setae distal segment of pe-duncle with 2 vertical rows of setae one with4 or 5 setae other with 2ndash4 setae 1ndash3 addi-tional setae on outer margin and 3ndash5 short se-tae on distal margin endopod 4ndash6 segmented(1ndash3) (0 or 1) (1ndash4) (1ndash4) (3ndash6) (3ndash6) se-tae exopod 3ndash5-segmented 0 (0ndash4) (0ndash4)(1ndash4) setae +2 asthetascs (5ndash8) asthetascs ()
Antenna (Fig 9d)mdashPeduncle 2-segmentedproximal segment with 0 or 1 seta distal seg-ment with 1ndash3 setae on inner margin spinepresent in some flagellum ranging from 14ndash18
segments (0 or 1) (2ndash4) variable region(1ndash3 articles with 0ndash2 setae) (0ndash4) (0ndash2)(2ndash5) (1ndash4) (3ndash6) (0ndash3) (2ndash5) (3ndash6)(0ndash3) (3ndash6) (0 or 1) (1ndash7) (4ndash9) setae exo-pod reduced to small lobe with 0ndash5 setae
Mandible (Fig 9e f )mdashBoth mandibles sim-ilar incisor process with 5ndash8 teeth molarprocess smooth or with small ridges palp3-segmented 0 (0 or 1) (8ndash12) setae
Maxillule (Fig 9g)mdashCoxal endite with15ndash22 marginal setae and 0 or 1 submarginal
Fig 8 Callichirus islagrande ZIV stage a maxilliped 3 b pereiopod 1 c pereiopod 2 d pereiopod 3 e pe-reiopod 4 f pereiopod 5 g pleopods from first abdominal somite h pleopods from fourth abdominal somite i tel-son Scales = 02 mm
112 JOURNAL OF CRUSTACEAN BIOLOGY VOL 20 NO 1 2000
seta basal endite with 3ndash5 plumose setae14ndash19 dentate spines 0ndash2 plumose setae oninner margin endopod reduced unsegmentedwith 0ndash8 setae
Maxilla (Fig 9h)mdashCoxal endite trilobed(17ndash21)+(7ndash10)+(6ndash9) setae basal enditebilobed 8ndash10 setae on proximal lobe 8ndash15setae on distal lobe endopod reduced with
Fig 9 Callichirus islagrande ZV stage a one example of the antenna b one example of pleopods Decapodidstage c antennule d antenna e right mandible f left mandible g maxillule h maxilla (plumose condition notshown) i maxilliped 1 j maxilliped 2 Scales = 02 mm
STRASSER AND FELDER LARVAL DEVELOPMENT IN CALLICHIRUS ISLAGRANDE 113
5ndash11 setae scaphognathite with 37ndash45 se-tae all setae plumose
Maxilliped 1 (Fig 9i)mdashCoxa with 8ndash12 se-tae basis highly setose with 27ndash39 setae en-dopod reduced with 0ndash11 setae exopod un-segmented 6ndash14 setae bilobed epipod with-out setae
Maxilliped 2 (Fig 9j)mdashCoxa partially fusedto basis with 1ndash4 setae basis with 3ndash5 se-
tae endopod 4-segmented (16ndash23) (0 or 1)(6ndash11) (4ndash8) setae exopod with 0ndash8 setaeepipod present but without setae
Maxilliped 3 (Fig 10a)mdashCoxa partiallyfused to basis with 0ndash3 setae basis with 1ndash4setae endopod 5-segmented (11ndash19) (9ndash15)(7ndash11) (8ndash21) (5ndash11) setae exopod usuallyreduced and without setae occasionally with2ndash9 setae epipod present
Fig 10 Callichirus islagrande decapodid stage a maxilliped 3 b pereiopod 1 c pereiopod 2 d pereiopod 3e pereiopod 4 f pereiopod 5 g pleopods from first abdominal somite h pleopods from fourth abdominal somitei telson Scales = 02 mm
114 JOURNAL OF CRUSTACEAN BIOLOGY VOL 20 NO 1 2000
Pereiopod 1 (Fig 10b)mdashChelate biramous7-segmented coxa with 0ndash6 setae basis with2ndash5 setae ischium with 4ndash7 setae and 0ndash3spines merus with 5ndash8 setae carpus with10ndash14 setae propodus with 27ndash37 setae and3ndash4 spines on inner margin of immovable fin-ger dactylus with 21ndash33 setae exopod usu-ally reduced and without setae occasionallywith 4ndash7 setae
Pereiopod 2 (Fig 10c)mdashChelate biramous7-segmented coxa with 2ndash5 setae basis with1ndash5 setae ischium with 4ndash10 setae meruswith 15ndash20 setae carpus with 8ndash12 setaepropodus with 30ndash40 setae and 1ndash4 spineson inner margin of immovable finger dacty-lus with 21ndash30 setae exopod usually reduced0ndash7 setae
Pereiopod 3 (Fig 10d)mdashBiramous 7-seg-mented coxa with 2ndash5 setae basis with 1ndash3setae ischium with 5ndash8 setae merus with6ndash9 setae carpus with 7ndash11 setae propodusbladelike with 25ndash34 setae 1 spine on innerdistal margin dactylus with 15ndash21 setae exo-pod usually reduced 0ndash5 setae
Pereiopod 4 (Fig 10e)mdashBiramous 7-seg-mented coxa with 2ndash7 setae basis with 1ndash4setae ischium with 3ndash8 setae merus with5ndash11 setae carpus with 4ndash9 setae propoduswith 27ndash38 setae 1 spine on inner distal mar-gin dactylus with 15ndash19 setae exopod usu-ally reduced with 0ndash2 setae
Pereiopod 5 (Fig 10f )mdashCheliform unira-mous 7-segmented coxa with 2ndash6 setae ba-sis with l or 2 setae ischium with 2ndash5 setaemerus with 5ndash9 setae carpus with 4ndash7 se-tae propodus with 22ndash30 setae 1 spine onimmovable finger dactylus with 6ndash10 setaeexopod absent
Pleopods (somite 2) (Fig 10g)mdashBiramouscoxa partially fused to basis without setaebasis with 0ndash3 setae endopod with 3ndash5hooked setae on appendix interna 10ndash17 mar-ginal plumose setae 0ndash2 submarginal nakedsetae exopod with 21ndash27 marginal plumosesetae and 0ndash1 submarginal naked seta
Pleopods (somites 3ndash5) (Fig 10h)mdashBiramouscoxa without setae basis with 3ndash6 marginalplumose setae endopod with 3ndash6 hooked se-tae on appendix interna 10ndash18 marginalplumose setae and 0ndash3 naked submarginal se-tae exopod with 20ndash29 marginal plumose se-tae and 0 or 1 submarginal naked seta
Telson and Uropods (Fig 10i)mdashWith 24ndash36processes on posterior margin median pro-cess usually absent or reduced uropods bi-ramous endopod with 19ndash25 plumose setaeexopod with 35ndash58 plumose setae protopodwith 3ndash5 setae
DISCUSSION
As in other species of Callichirus (see Ro-drigues 1976 Aste and Retamal 1983Strasser and Felder in press b) larval devel-opment of C islagrande included four or fivezoeal stages before metamorphosis to the de-capodid ZI larvae may be identified by thepresence of an elongate rounded rostrumsessile eyes and a lack of pereiopodspleopods or uropods (Fig 1a) At ZII lar-vae acquired biramous buds of pereiopods 1and 2 eyes were separated from the carapaceand the rostrum became flattened and serratedlaterally (Fig 1b) Buds of pleopods on ab-dominal segments 2ndash5 pereiopods 3ndash5 anduropods were developed at ZIII (Fig 1c) Inaddition to further development of structuresmentioned thus far ZIV larvae could be dis-tinguished by presence of a distal spine on theexopod and setae on the endopod of each uro-pod (Fig 1d) When it occurred ZV typicallyresembled ZIV but varied widely in mor-phology Appendages at ZV resembled thoseof ZIV the decapodid stage or some grada-tion between the two (see Figs 9a b for ex-amples) The decapodid stage was similar toother species of Callichirus in the reductionof larval characters such as the elongate ros-trum dorsal spines on the abdomen and ex-opods of the pereiopods (Fig le) Acquisitionof fan-like pleopods marked the transition toa primarily abdominal mode of swimming atthis stage
In the Florida Atlantic population of C ma-jor both salinity and presence of putative set-tlement cues at ZIV impacted the observednumber of zoeal stages and the duration ofZIV (Strasser and Felder in preparation1999c) In C islagrande the percentage oflarvae that passed through five versus fourzoeal stages varied (9ndash89) between parentalfemales but was not shown to depend onsalinity or presence of various putative set-tlement cues (Strasser and Felder 1998 inpreparation) Larvae that molted from ZIVto ZV were usually normal morphologicallyand had mortality similar to those that molteddirectly from ZIV to D In contrast larvae of
STRASSER AND FELDER LARVAL DEVELOPMENT IN CALLICHIRUS ISLAGRANDE 115
the Florida Atlantic C major that molted toZV had a higher incidence of mortality anddeformity at D than did those that had by-passed ZV (Strasser and Felder 1999a c)
The number of zoeal stages appears to beless variable in Callichirus garthi and theGulf of Mexico population of C major thanin congeneric populations studied to dateCallichirus garthi was reported to have fivezoeal stages and a long larval duration reach-ing the decapodid stage 48 days after hatch-ing (Aste and Retamal 1983) Developmentof the Gulf population of C major wasshorter in duration (decapodid reached about10 days after hatching) and typically includedfour zoeal stages (Strasser and Felder 1999a)animals rarely passed through three or fivezoeal stages before the molt to the decapodidAlthough the number of stages is reducedfrom that observed in C garthi developmentin the Gulf population of C major cannot beclassified as abbreviated There are species inboth the Callianassinae (Callianassa slkraussi Stebbing C sl kewalramanii and Csl tyrrhena (Petagna)) and Callichirinae(Sergio mirim Lepidophthalmus sinuensisand L louisianensis) that have only two zoealstages before the molt to the decapodid(Forbes 1973 Sankolli and Shenoy 1975Rodrigues 1984 Thessalou-Legaki 1990Nates et al 1997)
Compared to other species of the genusCallichirus C islagrande is morphologicallycloser to C garthi (from Chile) than to theGulf of Mexico population of C major whichoverlaps in distribution with C islagrande Inthe zoeal phase of C islagrande and Cgarthi the telson is concave and the outer-most process on the ventral margin is articu-lated In both C major and Callichirus sp theposterior margin of the telson is straight andthe outermost process is unarticulated duringthe zoeal stages (Rodrigues 1976 Strasserand Felder 1999a) There are also differencesin spination of the abdomen In zoeae of Cislagrande and C garthi the dorsal spine ofabdominal segment 3 is not longer than thosefound on segments 4 and 5 (Aste and Reta-mal 1983) In contrast the dorsal spine of ab-dominal segment 3 is approximately twice aslong as the dorsal spines on segments 4 and5 in early zoeal stages of C major and Cal-lichirus sp (Rodrigues 1976 Aste and Re-tamal 1983 Strasser and Felder 1999a) Thegreatest difference between these groups is
the number of pleopod pairs at zoeal and de-capodid stages Callichirus islagrande and Cgarthi have pleopods on abdominal segments2ndash5 while pleopods are present only on ab-dominal segments 3ndash5 in C major and Cal-lichirus sp (Rodrigues 1976 Aste and Re-tamal 1983 Strasser and Felder 1999a) Al-though Callichirus islagrande and C garthiare similar morphologically there are differ-ences between these two species Callichirusislagrande as well as C major and Cal-lichirus sp develops pleopod buds andchelate pereiopods at ZIII whereas C garthidoes not acquire these characters until ZIV
There are few callianassid species forwhich the larval development has been de-scribed (Sankolli and Shenoy 1975 Ro-drigues 1976 1984 Aste and Retamal 19831984 Konishi et al 1990 Nates et al 1997Strasser and Felder 1999a) all of which aremembers of the Callichirinae and Cal-lianassinae Among species there is a greatdeal of overlap in larval characters makingit difficult to draw distinctions between thesesubfamilies At present information on lar-val development of species in the Callichiri-nae is limited to larval descriptions of Cal-lichirus major C garthi Callichirus sp Cislagrande Sergio mirim Lepidophthalmuslouisianensis and L sinuensis (see Ro-drigues 1976 1984 Aste and Retamal 1983Nates et al 1997 Strasser and Felder1999a) As mentioned above species of Cal-lichirus have been shown to have four to fivezoeal stages while the other members exhibitabbreviated development that consists of onlytwo zoeal stages The most obvious differencebetween larvae of the species of Callichirinaewith abbreviated development is the numberof pleopod pairs Larvae of S mirim havepleopods on abdominal segments 3ndash5whereas L louisianensis and L sinuensishave pleopods on segments 2ndash5 (Rodrigues1984 Nates et al 1997)
Ghost shrimps of the genus Callichirus maybe distinguished from other genera of the Cal-lianassidae by the cuticular pattern on the dor-sal surface of the abdomen and by the strap-like shape of the uropodal endopods (Manningand Felder 1991) Whereas these charactersmay be used to distinguish adults of the genusCallichirus they are not present at zoeal anddecapodid stages Larvae may be distin-guished from all other callianassid genera byhaving four to five zoeal stages and numer-
116 JOURNAL OF CRUSTACEAN BIOLOGY VOL 20 NO 1 2000
ous processes (12ndash25+1+12ndash25) on the pos-terior margin of the telson during the earlyzoeal stages Larvae of Callichirus may alsodiffer from other callianassids with extendeddevelopment by the retention of the anomu-ran hair as the second process on the poste-rior margin of the telson in the late zoealstages Whereas this process is converted toa spine in Neotrypaea uncinata at ZV and Cal-lianassa sl petalura at ZIV the anomuranhair is retained at ZIV in Callichirus majorand C islagrande (see Rodrigues 1976Strasser and Felder 1999a) Although notmentioned specifically in the descriptions theanomuran hair appears to be present in illus-trations of Callichirus sp at ZIII and Cgarthii at ZV (Rodrigues 1976 fig 35 Asteand Retamal 1983 fig 8Q) A bilobed epi-pod on the first maxilliped of ZI is consistentlyfound in Callichirus but is also present at ZIin Lepidophthalmus sinuensis (see Nates etal 1997) Other callianassids acquire thischaracter at ZII (L louisianensis Nates et al1997 Callianassa sl kewlramanii Sankolliand Shenoy 1975) at ZV (Neotrypaea unci-nata Aste and Retamal 1984) or at the de-capodid stage (Sergio mirim Rodrigues 1984C sl petalura Konishi et al 1990) Thusthe number of zoeal stages and setation of thetelson may serve as the best characters to dis-tinguish larvae of Callichirus from other gen-era However since larval descriptions arelacking for many callianassid species it is un-known if these characters (in combination)will prove to be diagnostic until descriptionsfor other species are published
ACKNOWLEDGEMENTS
We thank Robert Bourgeois Barbara Hasek JimMaKinster Sergio Nates and Kurt Strasser who pro-vided assistance in the field and laboratory and MaryE Rice Director of the Smithsonian Marine StationLink Port Florida who facilitated access to station fa-cilities We also thank R Bauer R Jaeger R Lemaitreand G Watson for their early comments on this manu-script and Raymond B Manning who facilitated fund-ing and gave advice on collecting sites Direct supportfor this study was provided to Karen M Strasser by adoctoral fellowship from the University of Louisianaa graduate student research grant from the LUMCON(Louisiana Universities Marine Consortium) Founda-tion and a summer research award from the Smith-sonian Marine Station Additional support for field andlab activities was provided to D L Felder under Loui-siana Sea Grant College Program Grant RCFB-21 USFish and Wildlife Service Cooperative Agreement 14-16-0009-89-96 (Task Order No 6) US Department of En-ergy grant DE-FG02-97ER12220 and several small proj-ect grants from the Smithsonian Marine Station Views
expressed herein are those of the authors and do not nec-essarily reflect views of supporting government agenciesor subagencies mentioned above This is contributionNumber 466 of the Smithsonian Marine Station and con-tribution Number 67 of the ULL Laboratory for Crus-tacean Research All experiments conducted in this studycomply with current applicable state and federal laws
LITERATURE CITED
Aste A and M Retamal 1983 Desarrollo larval deCallianassa garthi Retamal 1975 bajo condiciones delaboratoriomdashCiencia y Tecnologiacutea del Mar CONA 75ndash26
mdashmdashmdash and mdashmdashmdash 1984 Desarrollo larval de Cal-lianassa uncinata H Milne Edwards1837 (DecapodaCallianassidae) bajo condiciones de laboratoriomdashGayana Zooloacutegica 48 41ndash56
Felder D L 1978 Osmotic and ionic regulation in sev-eral western Atlantic Callianassidae (Crustacea De-capoda Thalassinidea)mdashBiological Bulletin 154409ndash429
mdashmdashmdash and R B Griffis 1994 Dominant infaunalcommunities at risk in shoreline habitats burrowingthalassinid CrustaceamdashOCS Study number MMS94ndash0007 United States Department of the InteriorMinerals Management Service Gulf of Mexico OCSRegional Office New Orleans Louisiana Pp 1ndash87
Forbes A T 1973 An unusual abbreviated larval lifehistory in the estuarine burrowing prawn Callianassakraussi (Crustacea Decapoda Thalassinidea)mdashMa-rine Biology 22 361ndash365
Konishi K 1989 Larval development of the mudshrimp Upogebia (Upogebia) major (de Haan) (Crus-tacea Thalassinidea Upogebiidae) under laboratoryconditions with comments on larval characters of tha-lassinid familiesmdashBulletin of the National ResearchInstitute of Aquaculture 15 1ndash17
mdashmdashmdash R Quintana and Y Fukuda 1990 A completedescription of larval stages of the ghost shrimp Cal-lianassa petalura Stimpson (Crustacea ThalassinideaCallianassidae) under laboratory conditionsmdashBulletinof the National Research Institute of Aquaculture 1727ndash49
Manning R B and D L Felder 1986 The status ofthe callianassid genus Callichirus Stimpson 1866(Crustacea Decapoda Thalassinidea)mdashProceedingsof the Biological Society of Washington 99 437ndash443
mdashmdashmdash and mdashmdashmdash 1991 Revision of the AmericanCallianassidae (Crustacea Decapoda Thalassinidea)mdashProceedings of the Biological Society of Washington104 764ndash794
Nates S F D L Felder and R Lemaitre 1997 Com-parative larval development in two species of the bur-rowing ghost shrimp genus Lepidophthalmus (Decap-oda Callianassidae)mdashJournal of Crustacean Biology17 497ndash519
Rodrigues S de A 1976 Sobre a reproduccedilatildeo embri-ologia e desenvolvimento larval de Callichirus majorSay 1818 (Crustacea Decapoda Thalassinidea)mdashBo-letim de Zoologia Universidade de Satildeo Paulo 185ndash104
mdashmdashmdash 1984 Desenvolvimento poacutes-embrionaacuterio deCallichirus mirim (Rodrigues 1971) obtido emcondiccedilotildees artificiais (Crustacea Decapoda Tha-lassinidea)mdashBoletim de Zoologia Universidade deSatildeo Paulo 8 239ndash256
STRASSER AND FELDER LARVAL DEVELOPMENT IN CALLICHIRUS ISLAGRANDE 117
Sankolli K N and S Shenoy 1975 Larval develop-ment of mud shrimp Callianassa (Callichirus) kewal-ramanii Sankolli in the laboratory (Crustacea Decap-oda)mdashBulletin of the Department of Marine ScienceUniversity of Cochin 4 704ndash720
Staton J L and D L Felder 1995 Genetic variationin populations of the ghost shrimp genus Callichirus(Crustacea Decapoda Thalassinoidea) in the westernAtlantic and Gulf of MexicomdashBulletin of Marine Sci-ence 56 523ndash536
Strasser K M and D L Felder 1998 Settlement cuesin successive developmental stages of the ghost shrimpCallichirus major and C islagrande (Crustacea De-capoda Thalassinidea)mdashMarine Biology
mdashmdashmdash and mdashmdashmdash 1999a Larval development of twopopulations of the ghost shrimp Callichirus major (De-capoda Thalassinidea)mdashJournal of Crustacean Biology
mdashmdashmdash and mdashmdashmdash 1999b Sand as a stimulus for set-tlement in the ghost shrimp Callichirus major (Say)and C islagrande (Schmitt) (Crustacea DecapodaThalassinidea)mdashJournal of Experimental Marine Bi-ology and Ecology 239 211ndash222
mdashmdashmdash and mdashmdashmdash 1999c Settlement cues in theFlorida Atlantic population of the ghost shrimp Cal-lichirus major (Crustacea Decapoda Thalassinidea)mdashMarine Ecology Progress Series
Thessalou-Legaki M 1990 Advanced larval develop-ment of Callianassa tyrrhena (Decapoda Tha-lassinidea) and the effect of environmental factorsmdashJournal of Crustacean Biology 10 659ndash666
RECEIVED 19 September 1998ACCEPTED 3 December 1998

STRASSER AND FELDER LARVAL DEVELOPMENT IN CALLICHIRUS ISLAGRANDE 101
generic species and members of the familyCallianassidae
MATERIALS AND METHODS
Ovigerous females were collected in March May andJuly 1996 from the bayward side of a barrier island (IslesDernieres Louisiana) with yabby pumps (as in Felder1978) Adult females were maintained in 20-cm diame-ter finger bowls (at 27degC 25 ppt salinity) with daily wa-ter changes until eggs hatched Sea water was taken fromwell offshore Louisiana filtered through a 30-microm screenaerated and diluted to 25 ppt salinity with deionized wa-ter before use in the culture of animals To determinestage durations 40ndash120 larvae from each of 8 parentalfemales were reared individually through the first juve-nile stage (J1) ZI (first zoeal stage) larvae were movedto individual compartments upon hatching and maintainedat 27degC in filtered sea water on a 1212 light dark cy-cle Each day larvae were moved to containers with newsea water fed freshly hatched nauplii of Artemia (GreatSalt Lake) and examined to assess their stage of devel-opment Observations were terminated when animalsreached the first juvenile stage (J1)
Animals used for morphological comparisons were ob-tained from mass cultures Larvae were maintained in 20-cm diameter finger bowls (100ndash200 individuals per bowl)with daily water changes under conditions mentionedabove Animals were fixed in 70 ethanol stained withchlorozol black and transferred to glycerine prior to dis-section At least 10 animals were dissected for each stageand both right and left appendages were used for setalcounts Line illustrations were made on a Nikon invertedmicroscope fitted with a camera lucida Measurementswere made with a calibrated ocular micrometer Carapacelength (CL) was measured from the tip of the rostrum tothe posterior midpoint of the carapace and total length(TL) was measured from the rostral tip to the posteriormidpoint of the telson For each appendage the arrange-ment of setae was listed sequentially from proximal todistal margins as in Konishi (1989) and Nates et al(1997) Setal groups on successive segments were sepa-rated with a comma () Groups of setae on the same seg-ment or on different lobes of the same segment articleor endite were separated with a plus (+) A question mark() was used to designate questionable distinctions be-tween setae and aesthetascs Roman numerals were usedto describe the pattern of processes on the posterior mar-gin of the telson
RESULTS
Although there was no attempt to quantifythe duration of embryonic developmentovigerous females were held for up to 28 daysbefore the eggs hatched (usually at night)Prior to hatching eggs changed gradually incolor from bright orange-red to a translucent-brown Late-stage embryos that were droppedby the female before hatching and larvae thathatched as prezoeae were not viable evenwith aeration Prezoeae did not appear to bedeveloped fully and could not swim All zoealstages fed readily on nauplii of Artemia aswell as on each other The decapodid stage(D) appeared to consume much less than didthe preceding zoeal stage Strong positivephototaxis was exhibited by the first threezoeal (ZI to ZIII) stages The fourth zoealstage (ZIV) in general appeared to be at-tracted to light although some larvae reactedindifferently Decapodids did not exhibit ob-vious phototaxis
Larvae of Callichirus islagrande passedthrough four or five zoeal stages before molt-ing to D (Fig 1) these were usually com-pleted 11ndash14 days after hatching Mean du-rations of each stage were relatively consis-tent between larvae from different females(Table 1) However duration of ZIV was ap-proximately a day shorter when animalsmolted from ZIV to ZV than when animalsmolted directly to D (Table 2) Among theeight cultures observed in this study the per-centage of animals molting to ZV ranged from9ndash89 of the larvae that survived past ZIV
Description of Zoeal and Decapodid Stages
Near the mandibles the larval stages anddecapodid had two red chromatophores whichusually appeared fused when expanded Sev-
Table 1 Mean duration in days (plusmn95 CI) of the first to fifth zoeal stages (ZI to ZV) and the decapodid stage (D)in larval cultures from 8 ovigerous females of Callichirus islagrande Each culture consists of larvae from a differ-ent parental female Only animals that reached J1 (N) were used to calculate mean durations N0 is the starting num-ber of ZI larvae
Culture N N0 ZI ZII ZIII ZIV ZV D
1 100 120 226 (plusmn009) 204 (plusmn005) 222 (plusmn009) 388 (plusmn009) 389 (plusmn012) 570 (plusmn022)2 14 60 207 (plusmn016) 200 (plusmn000) 250 (plusmn031) 336 (plusmn060) 430 (plusmn028) 664 (plusmn117)3 34 55 206 (plusmn008) 206 (plusmn008) 244 (plusmn017) 421 (plusmn024) 420 (plusmn016) 568 (plusmn039)4 102 120 205 (plusmn004) 202 (plusmn005) 232 (plusmn010) 359 (plusmn010) 320 (plusmn009) 518 (plusmn028)5 31 40 205 (plusmn011) 200 (plusmn000) 226 (plusmn016) 323 (plusmn028) 400 (plusmn037) 494 (plusmn023)6 49 60 206 (plusmn007) 204 (plusmn008) 243 (plusmn014) 339 (plusmn020) 364 (plusmn018) 547 (plusmn046)7 36 54 217 (plusmn013) 211 (plusmn011) 214 (plusmn012) 294 (plusmn021) 327 (plusmn015) 439 (plusmn019)8 44 60 216 (plusmn013) 225 (plusmn015) 236 (plusmn017) 316 (plusmn026) 342 (plusmn017) 430 (plusmn030)
102 JOURNAL OF CRUSTACEAN BIOLOGY VOL 20 NO 1 2000
eral other red chromatophores were presenton the lateral edges of the carapace forminga red line when expanded
Morphological descriptions of Callichirusislagrande are as detailed below for stages ZIto ZIV and D ZV resembled ZIV in appear-ance However appendages were like thoseof ZIV D or some gradation between thetwo Setal patterns are given from proximal todistal segments for each appendage Carapacelength (CL) and total length (TL) are given asthe mean plusmn 95 CI in mm followed by therange of measurements from 10 specimens
Zoea IFigs 2a 3andashj
SizemdashCL = 123 plusmn 006 mm range 110ndash141mm TL = 326 plusmn 007 mm range 311ndash336mm
Carapace (Fig 2a)mdashShorter than abdomenrostrum elongate rounded in cross sectionand smooth eyes fused to carapace
Abdomen (Fig 2a)mdashSomites 2ndash5 with pos-terolateral projections and posterodorsalspines dorsal spine elongate on somite 2sometimes extending beyond posterior mar-gin of somite 3 short dorsal spines on somites3ndash5
Antennule (Fig 3a)mdashElongate exopodal andendopodal lobes not distinct 4ndash6 terminalaesthetascs 1 long plumose seta on future en-dopodal lobe
Antenna (Fig 3b)mdashProtopod with 1 distalspine between rami endopod with 2 longplumose setae scaphocerite (exopod) armedwith 1 strong distolateral spine 8 plumose se-tae on inner margin and 1 plumose seta onouter margin
Mandible (Fig 3c d)mdashIncisor process with6 or 7 teeth on right mandible 5ndash7 teeth onleft mandible molar process denticulate onboth mandibles
Maxillule (Fig 3e)mdashCoxal endite with 8ndash10marginal setae basal endite with 3 plumosesetae 3 or 4 large dentate spines and 4 or 5smaller dentate spines inner margin with 0or 1 plumose seta endopod 3-segmented dis-tinction between segments sometimes ob-scure 2 2 (3 or 4) setae protopod withoutsetae
Maxilla (Fig 3f )mdashCoxal endite bilobed 11or 12 setae on proximal lobe and 4 or 5 se-
Table 2 Percentage of larval Callichirus islangrande that molted from the fourth zoeal stage (ZIV) to the fifthzoeal stage (ZV) or directly to the decapodid stage (D) in larval cultures from 8 ovigerous females (Culture num-bers 1ndash8) Each batch consisted of larvae from a different parental female N0 = number surviving beyond ZIV N1
= number molting ZIV to ZV N1J = percentage of N1 surviving to J1 N2 = number molting ZIV to D N2J = per-centage of N2 surviving to J1 The mean duration in days (plusmn95 CI) is given for animals that molted from ZIV toZV and from ZIV to D
From ZIV To ZV To D
Survived to J1 Duration Survived to J1 DurationCulture N0 N1 N1J of ZIV N2 N2J of ZIV
1 105 11 82 30 (plusmn030) 94 97 40 (plusmn008)2 47 42 24 32 (plusmn023) 5 80 44 (plusmn111)3 45 8 62 30 (plusmn00) 37 78 44 (plusmn018)4 109 10 50 31 (plusmn078) 99 98 36 (plusmn010)5 38 19 79 28 (plusmn026) 19 84 37 (plusmn027)6 58 35 80 31 (plusmn017) 23 91 39 (plusmn030)7 48 30 73 26 (plusmn023) 18 78 34 (plusmn025)8 51 29 83 26 (plusmn019) 22 91 38 (plusmn019)
Fig 1 Mean proportion of larvae at each stage duringdevelopment of Callichirus islagrande Mean proportionsare derived from the individual rearings of 8 parental fe-males The first through fifth zoeal stages (ZI to ZV) thedecapodid stage (D) and all first-stage juveniles thatpassed D (J) are labeled under the peak of each line
STRASSER AND FELDER LARVAL DEVELOPMENT IN CALLICHIRUS ISLAGRANDE 103
tae on distal lobe basal edite bilobed with6ndash9 setae on each lobe endopod with 5 lobes3+2+2+2+(2 or 3) setae scaphognathite with16ndash19 setae all setae plumose
Maxilliped 1 (Fig 3g)mdashCoxa with 4ndash8 se-tae basis with 12ndash17 setae endopod 4-seg-mented (3 or 4) (1 or 2) (1ndash3) (3 or 4) se-tae exopod 2-segmented proximal segment
without setae distal segment with 5 or 6 se-tae on distal margin bilobed epipod withoutsetae all setae plumose
Maxilliped 2 (Fig 3h)mdashCoxa with 1 or 2 se-tae basis with 3ndash6 setae endopod 4-seg-mented (4 or 5) (0 or 1) (1 or 2) (3ndash6) se-tae exopod unsegmented 6ndash8 setae on dis-tal margin all setae plumose
Fig 2 Callichirus islagrande dorsal view of larvae and decapodid a ZI stage b ZII stage c ZIII stage d ZIVstage e decapodid stage Scale = 1 mm
104 JOURNAL OF CRUSTACEAN BIOLOGY VOL 20 NO 1 2000
Maxilliped 3 (Fig 3i)mdashCoxa with 0 or 1seta basis with 1 or 2 setae endopod 4-seg-mented (0 or 1) (0ndash2) (2 or 3) (3ndash5) se-tae exopod unsegmented 6ndash8 setae on dis-tal margin all setae plumose
PereiopodsmdashNot developed
PleopodsmdashNot developed
Telson (Fig 3j)mdashNarrow anterior not dif-ferentiated from abdominal segment 6 trian-gular posterior anal spine present with(12ndash14)+1+(13 or 14) processes on posterior
margin arranged as (I II III IV III II I)(I) outermost process a plumodenticulate seta(II) second process ldquoanomuran hairrdquo (III)10ndash12 plumodenticulate setae and (IV) cen-ter process a naked spine longer than otherprocesses
Zoea IIFigs 2b 4andashl
SizemdashCL = 144 plusmn 003 mm range 138ndash153mm TL = 364 plusmn 006 mm range 355ndash380mm
Fig 3 Callichirus islagrande ZI stage a antennule b antenna c right mandible d left mandible e maxillulef maxilla (plumose condition not shown) g maxilliped 1 h maxilliped 2 i maxilliped 3 j telson Scales = 02mm
STRASSER AND FELDER LARVAL DEVELOPMENT IN CALLICHIRUS ISLAGRANDE 105
Carapace (Fig 2b)mdashWith antennal spinesanterolateral margins serrated rostrum broadand dorsoventrally flattened with thin distalprojection and distolateral serrations eyesseparated from carapace
Abdomen (Fig 2b)mdashNo marked change fromstage ZI
Antennule (Fig 4a)mdashElongate weaklybilobed peduncle weakly 2-segmented 0ndash3short setae on distal margin of proximal seg-ment 2 long plumose setae and 0ndash3 short se-tae on distal end of distal segment endopodwith 1 terminal plumose seta exopodite with6ndash8 terminal aesthetascs
Antenna (Fig 4b)mdashProtopod with 2 distalspines 1 on outer margin and 1 between ramiendopod without setae scaphocerite armedwith 1 strong distolateral spine 10 or 11plumose setae on inner margin and 1plumose seta on outer margin
Mandible (Fig 4c d)mdashNo marked changefrom ZI
Maxillule (Fig 4e)mdashCoxal endite with 8ndash10marginal plumose setae basal endite with 3plumose setae 4 or 5 large dentate spines and4 or 5 smaller dentate spines inner marginwith 0 or 1 plumose seta endopod 3-seg-mented distinction between segments some-times obscure 2 2 (4 or 5) setae protopodwithout setae
Maxilla (Fig 4f )mdashCoxal endite bilobed12ndash15 setae on proximal lobe 4 or 5 setaeon distal lobe basal endite bilobed with 6ndash8setae on proximal lobe 5ndash9 setae on distallobe endopod with 5 lobes (2 or 3)+2+2+2+3setae scaphognathite with 19ndash24 setae allsetae plumose
Maxilliped 1 (Fig 4g)mdashCoxa with 4ndash8 se-tae basis with 13ndash17 marginal setae endo-pod 4-segmented (3 or 4) (2 or 3) (1 or 2)4 setae exopod 2-segmented proximal seg-ment without setae distal segment with 6 or7 setae on distal margin bilobed epipod with-out setae all setae plumose
Maxilliped 2 (Fig 4h)mdashCoxa with 0ndash2 se-tae basis with 4ndash6 setae endopod 4-seg-mented (4ndash7) 1 (2 or 3) (4ndash6) setae exo-pod unsegmented 7 or 8 setae on distal mar-gin all setae plumose
Maxilliped 3 (Fig 4i)mdashCoxa without setaebasis with 0ndash2 setae endopod 4-segmented
(l or 2) (1ndash3) (2ndash4) (3ndash6) setae exopod un-segmented 6ndash8 setae on distal margin all se-tae plumose
Pereiopods (Fig 4j k)mdashFirst and second pe-reiopod present both without setae
PleopodsmdashNot developed
Telson (Fig 4l)mdashNo change from ZI
Zoea IIIFigs 2c 5andashh 6andashh
SizemdashCL = 165 plusmn 003 mm range 160ndash172mm TL = 413 plusmn 009 mm range 399ndash436mm
Carapace (Fig 2c)mdashNo marked changefrom ZII
Abdomen (Fig 2c)mdashSomites 2ndash5 with pos-terolateral projections dorsal spine elongateon somite 2 sometimes extending beyondposterior margin of somite 3 short dorsalspines on somites 3ndash6
Antennule (Fig 5a)mdashBiramous peduncle2-segmented proximal segment with (0 or1)+(2 or 3) short setae on outer margin and1 long plumose seta on inner margin distalsegment with 2 or 3 setae on inner margin 1seta between rami and 0ndash3 short seta onouter margin endopod with 0 or 1 terminalplumose seta exopod with 6 or 7 terminaland 0ndash2 subterminal aesthetascs
Antenna (Fig 5b)mdashProtopod with 1 spineat outer distal margin endopod with 0 or 1short seta scaphocerite armed with 1 strongdistolateral spine 11ndash13 plumose setae on in-ner margin and 1 plumose seta on outer mar-gin
Mandible (Fig 5c d)mdashIncisor process with7 or 8 teeth on right mandible 6ndash8 teeth onleft mandible molar process denticulate onboth mandibles buds of palps present
Maxillule (Fig 5e)mdashCoxal endite with10ndash12 marginal plumose setae basal enditewith 3 plumose setae 4 or 5 large dentatespines and 4 or 5 smaller dentate spinesinner margin with 1 plumose seta endopod3-segmented distinction between segmentssometimes obscure 2 2 4 setae protopodwithout setation
Maxilla (Fig 5f )mdashCoxal endite bilobed14ndash17 setae on proximal lobe 4ndash6 setae ondistal lobe basal endite bilobed with 7ndash10 se-
106 JOURNAL OF CRUSTACEAN BIOLOGY VOL 20 NO 1 2000
tae on proximal lobe 6ndash9 setae on distal lobeendopod with 5 lobes 3+2+2+2+(2 or 3) se-tae scaphognathite with 21ndash26 setae all se-tae plumose
Maxilliped 1 (Fig 5g)mdashCoxa with 4ndash9 se-tae basis with 14ndash18 marginal setae endo-pod 4-segmented (3 or 4) 2 2 (3 or 4) se-tae exopod 2-segmented proximal segment
without setae distal segment with 5ndash7 setaeon distal margin bilobed epipod without se-tae all setae plumose
Maxilliped 2 (Fig 5h)mdashCoxa with 0ndash2 se-tae basis with 5 or 6 setae endopod 4-seg-mented (4ndash6) 1 3 (4ndash6) setae exopod un-segmented 7 or 8 setae on distal margin allsetae plumose
Fig 4 Callichirus islagrande ZII stage a antennule b antenna c right mandible d left mandible e maxillulef maxilla (plumose condition not shown) g maxilliped 1 h maxilliped 2 i maxilliped 3 j pereiopod 1 k pe-reiopod 2 1 telson Scales = 02 mm
STRASSER AND FELDER LARVAL DEVELOPMENT IN CALLICHIRUS ISLAGRANDE 107
Maxilliped 3 (Fig 6a)mdashCoxa with 0 or 1seta basis with 1ndash3 setae endopod 4-seg-mented (1 or 2) 2 (2ndash4) (4 or 5) setae exo-pod unsegmented 7 or 8 setae on distal mar-gin all setae plumose
Pereiopod 1 (Fig 6b)mdashChelate and bira-mous coxa without setae basis with 0 or 1seta endopod 4-segmented 0 (1 or 2) (2ndash4)2 setae exopod with 6 or 7 setae on distalmargin
Fig 5 Callichirus islagrande ZIII stage a antennule b antenna c right mandible d left mandible e maxil-lule f maxilla (plumose condition not shown) g maxilliped 1 h maxilliped 2 All setae on the maxilla were plumoseScales = 02 mm
108 JOURNAL OF CRUSTACEAN BIOLOGY VOL 20 NO 1 2000
Pereiopod 2 (Fig 6c)mdashChelate and biramouscoxa without setae basis with 0 or 1 seta en-dopod 4-segmented 0 (0ndash2) (0ndash4) 2 setaeexopod with 5ndash7 setae on distal margin
Pereiopods 3ndash5 (Fig 6d e f )mdashWithout setae
Pleopods (Figs 1c 6g)mdashBilobed buds onabdominal segments 2ndash5 first pair smallerthan others
Telson and Uropods (Fig 6h)mdashNo changein arrangement of posterior telsonal processesuropods biramous endopod without setae
Fig 6 Callichirus islagrande ZIII stage a maxilliped 3 b pereiopod 1 c pereiopod 2 d pereiopod 3 e pe-reiopod 4 f pereiopod 5 g pleopods from fourth abdominal somite h telson Scales = 02 mm
STRASSER AND FELDER LARVAL DEVELOPMENT IN CALLICHIRUS ISLAGRANDE 109
exopod with 10 or 11 marginal plumose se-tae ventral spine near base of protopod
Zoea IVFigs 2d 7andashh 8andashi
SizemdashCL = 192 plusmn 009 mm range 169ndash215mm TL = 482 plusmn 025 mm range 424ndash536mm
Carapace (Fig 2d)mdashNo marked changefrom ZIII
Abdomen (Fig 2d)mdashNo marked change fromZIII
Antennule (Fig 7a)mdashBiramous peduncle2-segmented with 1+(1ndash4) short setae onouter margin and 0 or 1 plumose seta on in-ner margin of proximal segment distal seg-ment with 3ndash5 plumose setae on inner mar-gin 1 plumose seta between rami and 0ndash3short setae on outer margin endopod with 1or 2 plumose setae exopod with 6 or 7 ter-minal and 2 subterminal aesthetascs
Antenna (Fig 7b)mdashProtopod with 1 distalspine on outer margin endopod with 0ndash2 se-tae scaphocerite armed with 1 strong disto-lateral spine 12ndash14 plumose setae on innermargin and 1 plumose seta on outer margin
Mandible (Fig 7c d)mdashIncisor process with8 teeth on right mandible 6ndash8 teeth on leftmandible molar process denticulate on bothmandibles palps present
Maxillule (Fig 7e)mdashCoxal endite with12ndash14 marginal plumose setae basal enditewith 3 plumose setae 5 or 6 large dentatespines and 5 or 6 smaller dentate spinesinner margin with 1 plumose seta endopod3-segmented distinction between segmentssometimes obscure 2 2 (3 or 4) setae pro-topod without setation
Maxilla (Fig 7f )mdashCoxal endite bilobed14ndash19 setae on proximal lobe 5 setae on dis-tal lobe basal endite bilobed 6ndash10 setae onproximal lobe 8 or 9 setae on distal lobeendopod with 5 lobes 3+(2 or 3)+2+2+3 se-tae scaphognathite with 27ndash32 setae all se-tae plumose
Maxilliped 1 (Fig 7g)mdashCoxa with 2ndash8 se-tae basis with 16ndash19 marginal setae endo-pod 4-segmented (3 or 4) 2 2 4 setae exo-
pod 2-segmented proximal segment withoutsetae distal segment with 6 or 7 setae on dis-tal margin bilobed epipod without setae allsetae plumose
Maxilliped 2 (Fig 7h)mdashCoxa with 0ndash3 se-tae basis with 4ndash6 setae endopod 4-seg-mented (5ndash8) 1 (3ndash5) (4 or 5) setae exo-pod unsegmented 7 or 8 setae on distal mar-gin all setae plumose
Maxilliped 3 (Fig 8a)mdashCoxa with 0 or 1seta basis with 2ndash4 setae endopod 4-seg-mented (l or 2) 2 (3ndash5) (3 or 4) setae exo-pod unsegmented 7 or 8 setae on distal mar-gin all setae plumose
Pereiopod 1 (Fig 8b)mdashBiramous cheliformcoxa without setae basis with 0ndash2 setae en-dopod 4-segmented (0 or 1) (1 or 2) (2ndash4)(1 or 2) setae exopod with 5ndash7 setae on dis-tal margin
Pereiopod 2 (Fig 8c)mdashBiramous cheliformcoxa without setae basis with 0 or 1 setaendopod 4-segmented 0 (1 or 2) (3ndash5) (2or 3) setae exopod with 5ndash7 setae on distalmargin
Pereiopod 3 (Fig 8d)mdashBiramous coxa andbasis without setae endopod 4-segmented0 (0ndash2) (3 or 4) 2 setae propodus blade-like exopod with 4 or 5 terminal setae
Pereiopod 4 (Fig 8e)mdashBiramous coxa andbasis without setae endopod 4-segmented (0or 1) (0ndash2) (2ndash4) 2 setae exopod with 0ndash2setae
Pereiopod 5 (Fig 8f )mdashUniramous 2 setaeon terminal segment 4 or 5 setae on adja-cent segment
Pleopods (Fig 8g h)mdashLarger than at ZIIIstill without setae
Telson and Uropods (Fig 8i)mdashTelson trian-gular narrow anteriorly no anal spine pos-terior margin with (12ndash15)+1+(13 or 14)processes arranged as (I II III IV III II I)(I) outermost process a plumodenticulate seta(II) second process ldquoanomuran hairrdquo (III) 10ndash13plumodenticulate setae and (IV) naked spineuropods biramous endopod with 7ndash11 marginalplumose setae exopod with 1 naked distolat-eral spine 12ndash14 marginal plumose setae
110 JOURNAL OF CRUSTACEAN BIOLOGY VOL 20 NO 1 2000
Decapodid (first postlarva)Figs 2e 9cndashj l0andashi
SizemdashCL = 133 plusmn 002 mm range129ndash138 mm TL = 378 plusmn 011 mm range361ndash405 mm
Carapace (Fig 2e)mdashShorter than abdomenno anterolateral serrations with pterygostomial
projections rostrum reduced without serra-tions sometimes bending down between eyes
Abdomen (Fig 2e)mdashSomites broad and flat-tened posterior projections rounded with 0ndash7setae somite 2 rectangular somites 3ndash5 oval
Antennule (Fig 9c)mdashPeduncle 3-segmentedproximal segment with 17ndash24 setae 13ndash15 of
Fig 7 Callichirus islagrande ZIV stage a antennule b antenna c right mandible d left mandible e maxil-lule f maxilla (plumose condition not shown) g maxilliped 1 h maxilliped 2 All setae on the maxilla were plumoseScales = 02 mm
STRASSER AND FELDER LARVAL DEVELOPMENT IN CALLICHIRUS ISLAGRANDE 111
these associated with statocyst intermediatesegment with 6ndash10 setae distal segment of pe-duncle with 2 vertical rows of setae one with4 or 5 setae other with 2ndash4 setae 1ndash3 addi-tional setae on outer margin and 3ndash5 short se-tae on distal margin endopod 4ndash6 segmented(1ndash3) (0 or 1) (1ndash4) (1ndash4) (3ndash6) (3ndash6) se-tae exopod 3ndash5-segmented 0 (0ndash4) (0ndash4)(1ndash4) setae +2 asthetascs (5ndash8) asthetascs ()
Antenna (Fig 9d)mdashPeduncle 2-segmentedproximal segment with 0 or 1 seta distal seg-ment with 1ndash3 setae on inner margin spinepresent in some flagellum ranging from 14ndash18
segments (0 or 1) (2ndash4) variable region(1ndash3 articles with 0ndash2 setae) (0ndash4) (0ndash2)(2ndash5) (1ndash4) (3ndash6) (0ndash3) (2ndash5) (3ndash6)(0ndash3) (3ndash6) (0 or 1) (1ndash7) (4ndash9) setae exo-pod reduced to small lobe with 0ndash5 setae
Mandible (Fig 9e f )mdashBoth mandibles sim-ilar incisor process with 5ndash8 teeth molarprocess smooth or with small ridges palp3-segmented 0 (0 or 1) (8ndash12) setae
Maxillule (Fig 9g)mdashCoxal endite with15ndash22 marginal setae and 0 or 1 submarginal
Fig 8 Callichirus islagrande ZIV stage a maxilliped 3 b pereiopod 1 c pereiopod 2 d pereiopod 3 e pe-reiopod 4 f pereiopod 5 g pleopods from first abdominal somite h pleopods from fourth abdominal somite i tel-son Scales = 02 mm
112 JOURNAL OF CRUSTACEAN BIOLOGY VOL 20 NO 1 2000
seta basal endite with 3ndash5 plumose setae14ndash19 dentate spines 0ndash2 plumose setae oninner margin endopod reduced unsegmentedwith 0ndash8 setae
Maxilla (Fig 9h)mdashCoxal endite trilobed(17ndash21)+(7ndash10)+(6ndash9) setae basal enditebilobed 8ndash10 setae on proximal lobe 8ndash15setae on distal lobe endopod reduced with
Fig 9 Callichirus islagrande ZV stage a one example of the antenna b one example of pleopods Decapodidstage c antennule d antenna e right mandible f left mandible g maxillule h maxilla (plumose condition notshown) i maxilliped 1 j maxilliped 2 Scales = 02 mm
STRASSER AND FELDER LARVAL DEVELOPMENT IN CALLICHIRUS ISLAGRANDE 113
5ndash11 setae scaphognathite with 37ndash45 se-tae all setae plumose
Maxilliped 1 (Fig 9i)mdashCoxa with 8ndash12 se-tae basis highly setose with 27ndash39 setae en-dopod reduced with 0ndash11 setae exopod un-segmented 6ndash14 setae bilobed epipod with-out setae
Maxilliped 2 (Fig 9j)mdashCoxa partially fusedto basis with 1ndash4 setae basis with 3ndash5 se-
tae endopod 4-segmented (16ndash23) (0 or 1)(6ndash11) (4ndash8) setae exopod with 0ndash8 setaeepipod present but without setae
Maxilliped 3 (Fig 10a)mdashCoxa partiallyfused to basis with 0ndash3 setae basis with 1ndash4setae endopod 5-segmented (11ndash19) (9ndash15)(7ndash11) (8ndash21) (5ndash11) setae exopod usuallyreduced and without setae occasionally with2ndash9 setae epipod present
Fig 10 Callichirus islagrande decapodid stage a maxilliped 3 b pereiopod 1 c pereiopod 2 d pereiopod 3e pereiopod 4 f pereiopod 5 g pleopods from first abdominal somite h pleopods from fourth abdominal somitei telson Scales = 02 mm
114 JOURNAL OF CRUSTACEAN BIOLOGY VOL 20 NO 1 2000
Pereiopod 1 (Fig 10b)mdashChelate biramous7-segmented coxa with 0ndash6 setae basis with2ndash5 setae ischium with 4ndash7 setae and 0ndash3spines merus with 5ndash8 setae carpus with10ndash14 setae propodus with 27ndash37 setae and3ndash4 spines on inner margin of immovable fin-ger dactylus with 21ndash33 setae exopod usu-ally reduced and without setae occasionallywith 4ndash7 setae
Pereiopod 2 (Fig 10c)mdashChelate biramous7-segmented coxa with 2ndash5 setae basis with1ndash5 setae ischium with 4ndash10 setae meruswith 15ndash20 setae carpus with 8ndash12 setaepropodus with 30ndash40 setae and 1ndash4 spineson inner margin of immovable finger dacty-lus with 21ndash30 setae exopod usually reduced0ndash7 setae
Pereiopod 3 (Fig 10d)mdashBiramous 7-seg-mented coxa with 2ndash5 setae basis with 1ndash3setae ischium with 5ndash8 setae merus with6ndash9 setae carpus with 7ndash11 setae propodusbladelike with 25ndash34 setae 1 spine on innerdistal margin dactylus with 15ndash21 setae exo-pod usually reduced 0ndash5 setae
Pereiopod 4 (Fig 10e)mdashBiramous 7-seg-mented coxa with 2ndash7 setae basis with 1ndash4setae ischium with 3ndash8 setae merus with5ndash11 setae carpus with 4ndash9 setae propoduswith 27ndash38 setae 1 spine on inner distal mar-gin dactylus with 15ndash19 setae exopod usu-ally reduced with 0ndash2 setae
Pereiopod 5 (Fig 10f )mdashCheliform unira-mous 7-segmented coxa with 2ndash6 setae ba-sis with l or 2 setae ischium with 2ndash5 setaemerus with 5ndash9 setae carpus with 4ndash7 se-tae propodus with 22ndash30 setae 1 spine onimmovable finger dactylus with 6ndash10 setaeexopod absent
Pleopods (somite 2) (Fig 10g)mdashBiramouscoxa partially fused to basis without setaebasis with 0ndash3 setae endopod with 3ndash5hooked setae on appendix interna 10ndash17 mar-ginal plumose setae 0ndash2 submarginal nakedsetae exopod with 21ndash27 marginal plumosesetae and 0ndash1 submarginal naked seta
Pleopods (somites 3ndash5) (Fig 10h)mdashBiramouscoxa without setae basis with 3ndash6 marginalplumose setae endopod with 3ndash6 hooked se-tae on appendix interna 10ndash18 marginalplumose setae and 0ndash3 naked submarginal se-tae exopod with 20ndash29 marginal plumose se-tae and 0 or 1 submarginal naked seta
Telson and Uropods (Fig 10i)mdashWith 24ndash36processes on posterior margin median pro-cess usually absent or reduced uropods bi-ramous endopod with 19ndash25 plumose setaeexopod with 35ndash58 plumose setae protopodwith 3ndash5 setae
DISCUSSION
As in other species of Callichirus (see Ro-drigues 1976 Aste and Retamal 1983Strasser and Felder in press b) larval devel-opment of C islagrande included four or fivezoeal stages before metamorphosis to the de-capodid ZI larvae may be identified by thepresence of an elongate rounded rostrumsessile eyes and a lack of pereiopodspleopods or uropods (Fig 1a) At ZII lar-vae acquired biramous buds of pereiopods 1and 2 eyes were separated from the carapaceand the rostrum became flattened and serratedlaterally (Fig 1b) Buds of pleopods on ab-dominal segments 2ndash5 pereiopods 3ndash5 anduropods were developed at ZIII (Fig 1c) Inaddition to further development of structuresmentioned thus far ZIV larvae could be dis-tinguished by presence of a distal spine on theexopod and setae on the endopod of each uro-pod (Fig 1d) When it occurred ZV typicallyresembled ZIV but varied widely in mor-phology Appendages at ZV resembled thoseof ZIV the decapodid stage or some grada-tion between the two (see Figs 9a b for ex-amples) The decapodid stage was similar toother species of Callichirus in the reductionof larval characters such as the elongate ros-trum dorsal spines on the abdomen and ex-opods of the pereiopods (Fig le) Acquisitionof fan-like pleopods marked the transition toa primarily abdominal mode of swimming atthis stage
In the Florida Atlantic population of C ma-jor both salinity and presence of putative set-tlement cues at ZIV impacted the observednumber of zoeal stages and the duration ofZIV (Strasser and Felder in preparation1999c) In C islagrande the percentage oflarvae that passed through five versus fourzoeal stages varied (9ndash89) between parentalfemales but was not shown to depend onsalinity or presence of various putative set-tlement cues (Strasser and Felder 1998 inpreparation) Larvae that molted from ZIVto ZV were usually normal morphologicallyand had mortality similar to those that molteddirectly from ZIV to D In contrast larvae of
STRASSER AND FELDER LARVAL DEVELOPMENT IN CALLICHIRUS ISLAGRANDE 115
the Florida Atlantic C major that molted toZV had a higher incidence of mortality anddeformity at D than did those that had by-passed ZV (Strasser and Felder 1999a c)
The number of zoeal stages appears to beless variable in Callichirus garthi and theGulf of Mexico population of C major thanin congeneric populations studied to dateCallichirus garthi was reported to have fivezoeal stages and a long larval duration reach-ing the decapodid stage 48 days after hatch-ing (Aste and Retamal 1983) Developmentof the Gulf population of C major wasshorter in duration (decapodid reached about10 days after hatching) and typically includedfour zoeal stages (Strasser and Felder 1999a)animals rarely passed through three or fivezoeal stages before the molt to the decapodidAlthough the number of stages is reducedfrom that observed in C garthi developmentin the Gulf population of C major cannot beclassified as abbreviated There are species inboth the Callianassinae (Callianassa slkraussi Stebbing C sl kewalramanii and Csl tyrrhena (Petagna)) and Callichirinae(Sergio mirim Lepidophthalmus sinuensisand L louisianensis) that have only two zoealstages before the molt to the decapodid(Forbes 1973 Sankolli and Shenoy 1975Rodrigues 1984 Thessalou-Legaki 1990Nates et al 1997)
Compared to other species of the genusCallichirus C islagrande is morphologicallycloser to C garthi (from Chile) than to theGulf of Mexico population of C major whichoverlaps in distribution with C islagrande Inthe zoeal phase of C islagrande and Cgarthi the telson is concave and the outer-most process on the ventral margin is articu-lated In both C major and Callichirus sp theposterior margin of the telson is straight andthe outermost process is unarticulated duringthe zoeal stages (Rodrigues 1976 Strasserand Felder 1999a) There are also differencesin spination of the abdomen In zoeae of Cislagrande and C garthi the dorsal spine ofabdominal segment 3 is not longer than thosefound on segments 4 and 5 (Aste and Reta-mal 1983) In contrast the dorsal spine of ab-dominal segment 3 is approximately twice aslong as the dorsal spines on segments 4 and5 in early zoeal stages of C major and Cal-lichirus sp (Rodrigues 1976 Aste and Re-tamal 1983 Strasser and Felder 1999a) Thegreatest difference between these groups is
the number of pleopod pairs at zoeal and de-capodid stages Callichirus islagrande and Cgarthi have pleopods on abdominal segments2ndash5 while pleopods are present only on ab-dominal segments 3ndash5 in C major and Cal-lichirus sp (Rodrigues 1976 Aste and Re-tamal 1983 Strasser and Felder 1999a) Al-though Callichirus islagrande and C garthiare similar morphologically there are differ-ences between these two species Callichirusislagrande as well as C major and Cal-lichirus sp develops pleopod buds andchelate pereiopods at ZIII whereas C garthidoes not acquire these characters until ZIV
There are few callianassid species forwhich the larval development has been de-scribed (Sankolli and Shenoy 1975 Ro-drigues 1976 1984 Aste and Retamal 19831984 Konishi et al 1990 Nates et al 1997Strasser and Felder 1999a) all of which aremembers of the Callichirinae and Cal-lianassinae Among species there is a greatdeal of overlap in larval characters makingit difficult to draw distinctions between thesesubfamilies At present information on lar-val development of species in the Callichiri-nae is limited to larval descriptions of Cal-lichirus major C garthi Callichirus sp Cislagrande Sergio mirim Lepidophthalmuslouisianensis and L sinuensis (see Ro-drigues 1976 1984 Aste and Retamal 1983Nates et al 1997 Strasser and Felder1999a) As mentioned above species of Cal-lichirus have been shown to have four to fivezoeal stages while the other members exhibitabbreviated development that consists of onlytwo zoeal stages The most obvious differencebetween larvae of the species of Callichirinaewith abbreviated development is the numberof pleopod pairs Larvae of S mirim havepleopods on abdominal segments 3ndash5whereas L louisianensis and L sinuensishave pleopods on segments 2ndash5 (Rodrigues1984 Nates et al 1997)
Ghost shrimps of the genus Callichirus maybe distinguished from other genera of the Cal-lianassidae by the cuticular pattern on the dor-sal surface of the abdomen and by the strap-like shape of the uropodal endopods (Manningand Felder 1991) Whereas these charactersmay be used to distinguish adults of the genusCallichirus they are not present at zoeal anddecapodid stages Larvae may be distin-guished from all other callianassid genera byhaving four to five zoeal stages and numer-
116 JOURNAL OF CRUSTACEAN BIOLOGY VOL 20 NO 1 2000
ous processes (12ndash25+1+12ndash25) on the pos-terior margin of the telson during the earlyzoeal stages Larvae of Callichirus may alsodiffer from other callianassids with extendeddevelopment by the retention of the anomu-ran hair as the second process on the poste-rior margin of the telson in the late zoealstages Whereas this process is converted toa spine in Neotrypaea uncinata at ZV and Cal-lianassa sl petalura at ZIV the anomuranhair is retained at ZIV in Callichirus majorand C islagrande (see Rodrigues 1976Strasser and Felder 1999a) Although notmentioned specifically in the descriptions theanomuran hair appears to be present in illus-trations of Callichirus sp at ZIII and Cgarthii at ZV (Rodrigues 1976 fig 35 Asteand Retamal 1983 fig 8Q) A bilobed epi-pod on the first maxilliped of ZI is consistentlyfound in Callichirus but is also present at ZIin Lepidophthalmus sinuensis (see Nates etal 1997) Other callianassids acquire thischaracter at ZII (L louisianensis Nates et al1997 Callianassa sl kewlramanii Sankolliand Shenoy 1975) at ZV (Neotrypaea unci-nata Aste and Retamal 1984) or at the de-capodid stage (Sergio mirim Rodrigues 1984C sl petalura Konishi et al 1990) Thusthe number of zoeal stages and setation of thetelson may serve as the best characters to dis-tinguish larvae of Callichirus from other gen-era However since larval descriptions arelacking for many callianassid species it is un-known if these characters (in combination)will prove to be diagnostic until descriptionsfor other species are published
ACKNOWLEDGEMENTS
We thank Robert Bourgeois Barbara Hasek JimMaKinster Sergio Nates and Kurt Strasser who pro-vided assistance in the field and laboratory and MaryE Rice Director of the Smithsonian Marine StationLink Port Florida who facilitated access to station fa-cilities We also thank R Bauer R Jaeger R Lemaitreand G Watson for their early comments on this manu-script and Raymond B Manning who facilitated fund-ing and gave advice on collecting sites Direct supportfor this study was provided to Karen M Strasser by adoctoral fellowship from the University of Louisianaa graduate student research grant from the LUMCON(Louisiana Universities Marine Consortium) Founda-tion and a summer research award from the Smith-sonian Marine Station Additional support for field andlab activities was provided to D L Felder under Loui-siana Sea Grant College Program Grant RCFB-21 USFish and Wildlife Service Cooperative Agreement 14-16-0009-89-96 (Task Order No 6) US Department of En-ergy grant DE-FG02-97ER12220 and several small proj-ect grants from the Smithsonian Marine Station Views
expressed herein are those of the authors and do not nec-essarily reflect views of supporting government agenciesor subagencies mentioned above This is contributionNumber 466 of the Smithsonian Marine Station and con-tribution Number 67 of the ULL Laboratory for Crus-tacean Research All experiments conducted in this studycomply with current applicable state and federal laws
LITERATURE CITED
Aste A and M Retamal 1983 Desarrollo larval deCallianassa garthi Retamal 1975 bajo condiciones delaboratoriomdashCiencia y Tecnologiacutea del Mar CONA 75ndash26
mdashmdashmdash and mdashmdashmdash 1984 Desarrollo larval de Cal-lianassa uncinata H Milne Edwards1837 (DecapodaCallianassidae) bajo condiciones de laboratoriomdashGayana Zooloacutegica 48 41ndash56
Felder D L 1978 Osmotic and ionic regulation in sev-eral western Atlantic Callianassidae (Crustacea De-capoda Thalassinidea)mdashBiological Bulletin 154409ndash429
mdashmdashmdash and R B Griffis 1994 Dominant infaunalcommunities at risk in shoreline habitats burrowingthalassinid CrustaceamdashOCS Study number MMS94ndash0007 United States Department of the InteriorMinerals Management Service Gulf of Mexico OCSRegional Office New Orleans Louisiana Pp 1ndash87
Forbes A T 1973 An unusual abbreviated larval lifehistory in the estuarine burrowing prawn Callianassakraussi (Crustacea Decapoda Thalassinidea)mdashMa-rine Biology 22 361ndash365
Konishi K 1989 Larval development of the mudshrimp Upogebia (Upogebia) major (de Haan) (Crus-tacea Thalassinidea Upogebiidae) under laboratoryconditions with comments on larval characters of tha-lassinid familiesmdashBulletin of the National ResearchInstitute of Aquaculture 15 1ndash17
mdashmdashmdash R Quintana and Y Fukuda 1990 A completedescription of larval stages of the ghost shrimp Cal-lianassa petalura Stimpson (Crustacea ThalassinideaCallianassidae) under laboratory conditionsmdashBulletinof the National Research Institute of Aquaculture 1727ndash49
Manning R B and D L Felder 1986 The status ofthe callianassid genus Callichirus Stimpson 1866(Crustacea Decapoda Thalassinidea)mdashProceedingsof the Biological Society of Washington 99 437ndash443
mdashmdashmdash and mdashmdashmdash 1991 Revision of the AmericanCallianassidae (Crustacea Decapoda Thalassinidea)mdashProceedings of the Biological Society of Washington104 764ndash794
Nates S F D L Felder and R Lemaitre 1997 Com-parative larval development in two species of the bur-rowing ghost shrimp genus Lepidophthalmus (Decap-oda Callianassidae)mdashJournal of Crustacean Biology17 497ndash519
Rodrigues S de A 1976 Sobre a reproduccedilatildeo embri-ologia e desenvolvimento larval de Callichirus majorSay 1818 (Crustacea Decapoda Thalassinidea)mdashBo-letim de Zoologia Universidade de Satildeo Paulo 185ndash104
mdashmdashmdash 1984 Desenvolvimento poacutes-embrionaacuterio deCallichirus mirim (Rodrigues 1971) obtido emcondiccedilotildees artificiais (Crustacea Decapoda Tha-lassinidea)mdashBoletim de Zoologia Universidade deSatildeo Paulo 8 239ndash256
STRASSER AND FELDER LARVAL DEVELOPMENT IN CALLICHIRUS ISLAGRANDE 117
Sankolli K N and S Shenoy 1975 Larval develop-ment of mud shrimp Callianassa (Callichirus) kewal-ramanii Sankolli in the laboratory (Crustacea Decap-oda)mdashBulletin of the Department of Marine ScienceUniversity of Cochin 4 704ndash720
Staton J L and D L Felder 1995 Genetic variationin populations of the ghost shrimp genus Callichirus(Crustacea Decapoda Thalassinoidea) in the westernAtlantic and Gulf of MexicomdashBulletin of Marine Sci-ence 56 523ndash536
Strasser K M and D L Felder 1998 Settlement cuesin successive developmental stages of the ghost shrimpCallichirus major and C islagrande (Crustacea De-capoda Thalassinidea)mdashMarine Biology
mdashmdashmdash and mdashmdashmdash 1999a Larval development of twopopulations of the ghost shrimp Callichirus major (De-capoda Thalassinidea)mdashJournal of Crustacean Biology
mdashmdashmdash and mdashmdashmdash 1999b Sand as a stimulus for set-tlement in the ghost shrimp Callichirus major (Say)and C islagrande (Schmitt) (Crustacea DecapodaThalassinidea)mdashJournal of Experimental Marine Bi-ology and Ecology 239 211ndash222
mdashmdashmdash and mdashmdashmdash 1999c Settlement cues in theFlorida Atlantic population of the ghost shrimp Cal-lichirus major (Crustacea Decapoda Thalassinidea)mdashMarine Ecology Progress Series
Thessalou-Legaki M 1990 Advanced larval develop-ment of Callianassa tyrrhena (Decapoda Tha-lassinidea) and the effect of environmental factorsmdashJournal of Crustacean Biology 10 659ndash666
RECEIVED 19 September 1998ACCEPTED 3 December 1998

102 JOURNAL OF CRUSTACEAN BIOLOGY VOL 20 NO 1 2000
eral other red chromatophores were presenton the lateral edges of the carapace forminga red line when expanded
Morphological descriptions of Callichirusislagrande are as detailed below for stages ZIto ZIV and D ZV resembled ZIV in appear-ance However appendages were like thoseof ZIV D or some gradation between thetwo Setal patterns are given from proximal todistal segments for each appendage Carapacelength (CL) and total length (TL) are given asthe mean plusmn 95 CI in mm followed by therange of measurements from 10 specimens
Zoea IFigs 2a 3andashj
SizemdashCL = 123 plusmn 006 mm range 110ndash141mm TL = 326 plusmn 007 mm range 311ndash336mm
Carapace (Fig 2a)mdashShorter than abdomenrostrum elongate rounded in cross sectionand smooth eyes fused to carapace
Abdomen (Fig 2a)mdashSomites 2ndash5 with pos-terolateral projections and posterodorsalspines dorsal spine elongate on somite 2sometimes extending beyond posterior mar-gin of somite 3 short dorsal spines on somites3ndash5
Antennule (Fig 3a)mdashElongate exopodal andendopodal lobes not distinct 4ndash6 terminalaesthetascs 1 long plumose seta on future en-dopodal lobe
Antenna (Fig 3b)mdashProtopod with 1 distalspine between rami endopod with 2 longplumose setae scaphocerite (exopod) armedwith 1 strong distolateral spine 8 plumose se-tae on inner margin and 1 plumose seta onouter margin
Mandible (Fig 3c d)mdashIncisor process with6 or 7 teeth on right mandible 5ndash7 teeth onleft mandible molar process denticulate onboth mandibles
Maxillule (Fig 3e)mdashCoxal endite with 8ndash10marginal setae basal endite with 3 plumosesetae 3 or 4 large dentate spines and 4 or 5smaller dentate spines inner margin with 0or 1 plumose seta endopod 3-segmented dis-tinction between segments sometimes ob-scure 2 2 (3 or 4) setae protopod withoutsetae
Maxilla (Fig 3f )mdashCoxal endite bilobed 11or 12 setae on proximal lobe and 4 or 5 se-
Table 2 Percentage of larval Callichirus islangrande that molted from the fourth zoeal stage (ZIV) to the fifthzoeal stage (ZV) or directly to the decapodid stage (D) in larval cultures from 8 ovigerous females (Culture num-bers 1ndash8) Each batch consisted of larvae from a different parental female N0 = number surviving beyond ZIV N1
= number molting ZIV to ZV N1J = percentage of N1 surviving to J1 N2 = number molting ZIV to D N2J = per-centage of N2 surviving to J1 The mean duration in days (plusmn95 CI) is given for animals that molted from ZIV toZV and from ZIV to D
From ZIV To ZV To D
Survived to J1 Duration Survived to J1 DurationCulture N0 N1 N1J of ZIV N2 N2J of ZIV
1 105 11 82 30 (plusmn030) 94 97 40 (plusmn008)2 47 42 24 32 (plusmn023) 5 80 44 (plusmn111)3 45 8 62 30 (plusmn00) 37 78 44 (plusmn018)4 109 10 50 31 (plusmn078) 99 98 36 (plusmn010)5 38 19 79 28 (plusmn026) 19 84 37 (plusmn027)6 58 35 80 31 (plusmn017) 23 91 39 (plusmn030)7 48 30 73 26 (plusmn023) 18 78 34 (plusmn025)8 51 29 83 26 (plusmn019) 22 91 38 (plusmn019)
Fig 1 Mean proportion of larvae at each stage duringdevelopment of Callichirus islagrande Mean proportionsare derived from the individual rearings of 8 parental fe-males The first through fifth zoeal stages (ZI to ZV) thedecapodid stage (D) and all first-stage juveniles thatpassed D (J) are labeled under the peak of each line
STRASSER AND FELDER LARVAL DEVELOPMENT IN CALLICHIRUS ISLAGRANDE 103
tae on distal lobe basal edite bilobed with6ndash9 setae on each lobe endopod with 5 lobes3+2+2+2+(2 or 3) setae scaphognathite with16ndash19 setae all setae plumose
Maxilliped 1 (Fig 3g)mdashCoxa with 4ndash8 se-tae basis with 12ndash17 setae endopod 4-seg-mented (3 or 4) (1 or 2) (1ndash3) (3 or 4) se-tae exopod 2-segmented proximal segment
without setae distal segment with 5 or 6 se-tae on distal margin bilobed epipod withoutsetae all setae plumose
Maxilliped 2 (Fig 3h)mdashCoxa with 1 or 2 se-tae basis with 3ndash6 setae endopod 4-seg-mented (4 or 5) (0 or 1) (1 or 2) (3ndash6) se-tae exopod unsegmented 6ndash8 setae on dis-tal margin all setae plumose
Fig 2 Callichirus islagrande dorsal view of larvae and decapodid a ZI stage b ZII stage c ZIII stage d ZIVstage e decapodid stage Scale = 1 mm
104 JOURNAL OF CRUSTACEAN BIOLOGY VOL 20 NO 1 2000
Maxilliped 3 (Fig 3i)mdashCoxa with 0 or 1seta basis with 1 or 2 setae endopod 4-seg-mented (0 or 1) (0ndash2) (2 or 3) (3ndash5) se-tae exopod unsegmented 6ndash8 setae on dis-tal margin all setae plumose
PereiopodsmdashNot developed
PleopodsmdashNot developed
Telson (Fig 3j)mdashNarrow anterior not dif-ferentiated from abdominal segment 6 trian-gular posterior anal spine present with(12ndash14)+1+(13 or 14) processes on posterior
margin arranged as (I II III IV III II I)(I) outermost process a plumodenticulate seta(II) second process ldquoanomuran hairrdquo (III)10ndash12 plumodenticulate setae and (IV) cen-ter process a naked spine longer than otherprocesses
Zoea IIFigs 2b 4andashl
SizemdashCL = 144 plusmn 003 mm range 138ndash153mm TL = 364 plusmn 006 mm range 355ndash380mm
Fig 3 Callichirus islagrande ZI stage a antennule b antenna c right mandible d left mandible e maxillulef maxilla (plumose condition not shown) g maxilliped 1 h maxilliped 2 i maxilliped 3 j telson Scales = 02mm
STRASSER AND FELDER LARVAL DEVELOPMENT IN CALLICHIRUS ISLAGRANDE 105
Carapace (Fig 2b)mdashWith antennal spinesanterolateral margins serrated rostrum broadand dorsoventrally flattened with thin distalprojection and distolateral serrations eyesseparated from carapace
Abdomen (Fig 2b)mdashNo marked change fromstage ZI
Antennule (Fig 4a)mdashElongate weaklybilobed peduncle weakly 2-segmented 0ndash3short setae on distal margin of proximal seg-ment 2 long plumose setae and 0ndash3 short se-tae on distal end of distal segment endopodwith 1 terminal plumose seta exopodite with6ndash8 terminal aesthetascs
Antenna (Fig 4b)mdashProtopod with 2 distalspines 1 on outer margin and 1 between ramiendopod without setae scaphocerite armedwith 1 strong distolateral spine 10 or 11plumose setae on inner margin and 1plumose seta on outer margin
Mandible (Fig 4c d)mdashNo marked changefrom ZI
Maxillule (Fig 4e)mdashCoxal endite with 8ndash10marginal plumose setae basal endite with 3plumose setae 4 or 5 large dentate spines and4 or 5 smaller dentate spines inner marginwith 0 or 1 plumose seta endopod 3-seg-mented distinction between segments some-times obscure 2 2 (4 or 5) setae protopodwithout setae
Maxilla (Fig 4f )mdashCoxal endite bilobed12ndash15 setae on proximal lobe 4 or 5 setaeon distal lobe basal endite bilobed with 6ndash8setae on proximal lobe 5ndash9 setae on distallobe endopod with 5 lobes (2 or 3)+2+2+2+3setae scaphognathite with 19ndash24 setae allsetae plumose
Maxilliped 1 (Fig 4g)mdashCoxa with 4ndash8 se-tae basis with 13ndash17 marginal setae endo-pod 4-segmented (3 or 4) (2 or 3) (1 or 2)4 setae exopod 2-segmented proximal seg-ment without setae distal segment with 6 or7 setae on distal margin bilobed epipod with-out setae all setae plumose
Maxilliped 2 (Fig 4h)mdashCoxa with 0ndash2 se-tae basis with 4ndash6 setae endopod 4-seg-mented (4ndash7) 1 (2 or 3) (4ndash6) setae exo-pod unsegmented 7 or 8 setae on distal mar-gin all setae plumose
Maxilliped 3 (Fig 4i)mdashCoxa without setaebasis with 0ndash2 setae endopod 4-segmented
(l or 2) (1ndash3) (2ndash4) (3ndash6) setae exopod un-segmented 6ndash8 setae on distal margin all se-tae plumose
Pereiopods (Fig 4j k)mdashFirst and second pe-reiopod present both without setae
PleopodsmdashNot developed
Telson (Fig 4l)mdashNo change from ZI
Zoea IIIFigs 2c 5andashh 6andashh
SizemdashCL = 165 plusmn 003 mm range 160ndash172mm TL = 413 plusmn 009 mm range 399ndash436mm
Carapace (Fig 2c)mdashNo marked changefrom ZII
Abdomen (Fig 2c)mdashSomites 2ndash5 with pos-terolateral projections dorsal spine elongateon somite 2 sometimes extending beyondposterior margin of somite 3 short dorsalspines on somites 3ndash6
Antennule (Fig 5a)mdashBiramous peduncle2-segmented proximal segment with (0 or1)+(2 or 3) short setae on outer margin and1 long plumose seta on inner margin distalsegment with 2 or 3 setae on inner margin 1seta between rami and 0ndash3 short seta onouter margin endopod with 0 or 1 terminalplumose seta exopod with 6 or 7 terminaland 0ndash2 subterminal aesthetascs
Antenna (Fig 5b)mdashProtopod with 1 spineat outer distal margin endopod with 0 or 1short seta scaphocerite armed with 1 strongdistolateral spine 11ndash13 plumose setae on in-ner margin and 1 plumose seta on outer mar-gin
Mandible (Fig 5c d)mdashIncisor process with7 or 8 teeth on right mandible 6ndash8 teeth onleft mandible molar process denticulate onboth mandibles buds of palps present
Maxillule (Fig 5e)mdashCoxal endite with10ndash12 marginal plumose setae basal enditewith 3 plumose setae 4 or 5 large dentatespines and 4 or 5 smaller dentate spinesinner margin with 1 plumose seta endopod3-segmented distinction between segmentssometimes obscure 2 2 4 setae protopodwithout setation
Maxilla (Fig 5f )mdashCoxal endite bilobed14ndash17 setae on proximal lobe 4ndash6 setae ondistal lobe basal endite bilobed with 7ndash10 se-
106 JOURNAL OF CRUSTACEAN BIOLOGY VOL 20 NO 1 2000
tae on proximal lobe 6ndash9 setae on distal lobeendopod with 5 lobes 3+2+2+2+(2 or 3) se-tae scaphognathite with 21ndash26 setae all se-tae plumose
Maxilliped 1 (Fig 5g)mdashCoxa with 4ndash9 se-tae basis with 14ndash18 marginal setae endo-pod 4-segmented (3 or 4) 2 2 (3 or 4) se-tae exopod 2-segmented proximal segment
without setae distal segment with 5ndash7 setaeon distal margin bilobed epipod without se-tae all setae plumose
Maxilliped 2 (Fig 5h)mdashCoxa with 0ndash2 se-tae basis with 5 or 6 setae endopod 4-seg-mented (4ndash6) 1 3 (4ndash6) setae exopod un-segmented 7 or 8 setae on distal margin allsetae plumose
Fig 4 Callichirus islagrande ZII stage a antennule b antenna c right mandible d left mandible e maxillulef maxilla (plumose condition not shown) g maxilliped 1 h maxilliped 2 i maxilliped 3 j pereiopod 1 k pe-reiopod 2 1 telson Scales = 02 mm
STRASSER AND FELDER LARVAL DEVELOPMENT IN CALLICHIRUS ISLAGRANDE 107
Maxilliped 3 (Fig 6a)mdashCoxa with 0 or 1seta basis with 1ndash3 setae endopod 4-seg-mented (1 or 2) 2 (2ndash4) (4 or 5) setae exo-pod unsegmented 7 or 8 setae on distal mar-gin all setae plumose
Pereiopod 1 (Fig 6b)mdashChelate and bira-mous coxa without setae basis with 0 or 1seta endopod 4-segmented 0 (1 or 2) (2ndash4)2 setae exopod with 6 or 7 setae on distalmargin
Fig 5 Callichirus islagrande ZIII stage a antennule b antenna c right mandible d left mandible e maxil-lule f maxilla (plumose condition not shown) g maxilliped 1 h maxilliped 2 All setae on the maxilla were plumoseScales = 02 mm
108 JOURNAL OF CRUSTACEAN BIOLOGY VOL 20 NO 1 2000
Pereiopod 2 (Fig 6c)mdashChelate and biramouscoxa without setae basis with 0 or 1 seta en-dopod 4-segmented 0 (0ndash2) (0ndash4) 2 setaeexopod with 5ndash7 setae on distal margin
Pereiopods 3ndash5 (Fig 6d e f )mdashWithout setae
Pleopods (Figs 1c 6g)mdashBilobed buds onabdominal segments 2ndash5 first pair smallerthan others
Telson and Uropods (Fig 6h)mdashNo changein arrangement of posterior telsonal processesuropods biramous endopod without setae
Fig 6 Callichirus islagrande ZIII stage a maxilliped 3 b pereiopod 1 c pereiopod 2 d pereiopod 3 e pe-reiopod 4 f pereiopod 5 g pleopods from fourth abdominal somite h telson Scales = 02 mm
STRASSER AND FELDER LARVAL DEVELOPMENT IN CALLICHIRUS ISLAGRANDE 109
exopod with 10 or 11 marginal plumose se-tae ventral spine near base of protopod
Zoea IVFigs 2d 7andashh 8andashi
SizemdashCL = 192 plusmn 009 mm range 169ndash215mm TL = 482 plusmn 025 mm range 424ndash536mm
Carapace (Fig 2d)mdashNo marked changefrom ZIII
Abdomen (Fig 2d)mdashNo marked change fromZIII
Antennule (Fig 7a)mdashBiramous peduncle2-segmented with 1+(1ndash4) short setae onouter margin and 0 or 1 plumose seta on in-ner margin of proximal segment distal seg-ment with 3ndash5 plumose setae on inner mar-gin 1 plumose seta between rami and 0ndash3short setae on outer margin endopod with 1or 2 plumose setae exopod with 6 or 7 ter-minal and 2 subterminal aesthetascs
Antenna (Fig 7b)mdashProtopod with 1 distalspine on outer margin endopod with 0ndash2 se-tae scaphocerite armed with 1 strong disto-lateral spine 12ndash14 plumose setae on innermargin and 1 plumose seta on outer margin
Mandible (Fig 7c d)mdashIncisor process with8 teeth on right mandible 6ndash8 teeth on leftmandible molar process denticulate on bothmandibles palps present
Maxillule (Fig 7e)mdashCoxal endite with12ndash14 marginal plumose setae basal enditewith 3 plumose setae 5 or 6 large dentatespines and 5 or 6 smaller dentate spinesinner margin with 1 plumose seta endopod3-segmented distinction between segmentssometimes obscure 2 2 (3 or 4) setae pro-topod without setation
Maxilla (Fig 7f )mdashCoxal endite bilobed14ndash19 setae on proximal lobe 5 setae on dis-tal lobe basal endite bilobed 6ndash10 setae onproximal lobe 8 or 9 setae on distal lobeendopod with 5 lobes 3+(2 or 3)+2+2+3 se-tae scaphognathite with 27ndash32 setae all se-tae plumose
Maxilliped 1 (Fig 7g)mdashCoxa with 2ndash8 se-tae basis with 16ndash19 marginal setae endo-pod 4-segmented (3 or 4) 2 2 4 setae exo-
pod 2-segmented proximal segment withoutsetae distal segment with 6 or 7 setae on dis-tal margin bilobed epipod without setae allsetae plumose
Maxilliped 2 (Fig 7h)mdashCoxa with 0ndash3 se-tae basis with 4ndash6 setae endopod 4-seg-mented (5ndash8) 1 (3ndash5) (4 or 5) setae exo-pod unsegmented 7 or 8 setae on distal mar-gin all setae plumose
Maxilliped 3 (Fig 8a)mdashCoxa with 0 or 1seta basis with 2ndash4 setae endopod 4-seg-mented (l or 2) 2 (3ndash5) (3 or 4) setae exo-pod unsegmented 7 or 8 setae on distal mar-gin all setae plumose
Pereiopod 1 (Fig 8b)mdashBiramous cheliformcoxa without setae basis with 0ndash2 setae en-dopod 4-segmented (0 or 1) (1 or 2) (2ndash4)(1 or 2) setae exopod with 5ndash7 setae on dis-tal margin
Pereiopod 2 (Fig 8c)mdashBiramous cheliformcoxa without setae basis with 0 or 1 setaendopod 4-segmented 0 (1 or 2) (3ndash5) (2or 3) setae exopod with 5ndash7 setae on distalmargin
Pereiopod 3 (Fig 8d)mdashBiramous coxa andbasis without setae endopod 4-segmented0 (0ndash2) (3 or 4) 2 setae propodus blade-like exopod with 4 or 5 terminal setae
Pereiopod 4 (Fig 8e)mdashBiramous coxa andbasis without setae endopod 4-segmented (0or 1) (0ndash2) (2ndash4) 2 setae exopod with 0ndash2setae
Pereiopod 5 (Fig 8f )mdashUniramous 2 setaeon terminal segment 4 or 5 setae on adja-cent segment
Pleopods (Fig 8g h)mdashLarger than at ZIIIstill without setae
Telson and Uropods (Fig 8i)mdashTelson trian-gular narrow anteriorly no anal spine pos-terior margin with (12ndash15)+1+(13 or 14)processes arranged as (I II III IV III II I)(I) outermost process a plumodenticulate seta(II) second process ldquoanomuran hairrdquo (III) 10ndash13plumodenticulate setae and (IV) naked spineuropods biramous endopod with 7ndash11 marginalplumose setae exopod with 1 naked distolat-eral spine 12ndash14 marginal plumose setae
110 JOURNAL OF CRUSTACEAN BIOLOGY VOL 20 NO 1 2000
Decapodid (first postlarva)Figs 2e 9cndashj l0andashi
SizemdashCL = 133 plusmn 002 mm range129ndash138 mm TL = 378 plusmn 011 mm range361ndash405 mm
Carapace (Fig 2e)mdashShorter than abdomenno anterolateral serrations with pterygostomial
projections rostrum reduced without serra-tions sometimes bending down between eyes
Abdomen (Fig 2e)mdashSomites broad and flat-tened posterior projections rounded with 0ndash7setae somite 2 rectangular somites 3ndash5 oval
Antennule (Fig 9c)mdashPeduncle 3-segmentedproximal segment with 17ndash24 setae 13ndash15 of
Fig 7 Callichirus islagrande ZIV stage a antennule b antenna c right mandible d left mandible e maxil-lule f maxilla (plumose condition not shown) g maxilliped 1 h maxilliped 2 All setae on the maxilla were plumoseScales = 02 mm
STRASSER AND FELDER LARVAL DEVELOPMENT IN CALLICHIRUS ISLAGRANDE 111
these associated with statocyst intermediatesegment with 6ndash10 setae distal segment of pe-duncle with 2 vertical rows of setae one with4 or 5 setae other with 2ndash4 setae 1ndash3 addi-tional setae on outer margin and 3ndash5 short se-tae on distal margin endopod 4ndash6 segmented(1ndash3) (0 or 1) (1ndash4) (1ndash4) (3ndash6) (3ndash6) se-tae exopod 3ndash5-segmented 0 (0ndash4) (0ndash4)(1ndash4) setae +2 asthetascs (5ndash8) asthetascs ()
Antenna (Fig 9d)mdashPeduncle 2-segmentedproximal segment with 0 or 1 seta distal seg-ment with 1ndash3 setae on inner margin spinepresent in some flagellum ranging from 14ndash18
segments (0 or 1) (2ndash4) variable region(1ndash3 articles with 0ndash2 setae) (0ndash4) (0ndash2)(2ndash5) (1ndash4) (3ndash6) (0ndash3) (2ndash5) (3ndash6)(0ndash3) (3ndash6) (0 or 1) (1ndash7) (4ndash9) setae exo-pod reduced to small lobe with 0ndash5 setae
Mandible (Fig 9e f )mdashBoth mandibles sim-ilar incisor process with 5ndash8 teeth molarprocess smooth or with small ridges palp3-segmented 0 (0 or 1) (8ndash12) setae
Maxillule (Fig 9g)mdashCoxal endite with15ndash22 marginal setae and 0 or 1 submarginal
Fig 8 Callichirus islagrande ZIV stage a maxilliped 3 b pereiopod 1 c pereiopod 2 d pereiopod 3 e pe-reiopod 4 f pereiopod 5 g pleopods from first abdominal somite h pleopods from fourth abdominal somite i tel-son Scales = 02 mm
112 JOURNAL OF CRUSTACEAN BIOLOGY VOL 20 NO 1 2000
seta basal endite with 3ndash5 plumose setae14ndash19 dentate spines 0ndash2 plumose setae oninner margin endopod reduced unsegmentedwith 0ndash8 setae
Maxilla (Fig 9h)mdashCoxal endite trilobed(17ndash21)+(7ndash10)+(6ndash9) setae basal enditebilobed 8ndash10 setae on proximal lobe 8ndash15setae on distal lobe endopod reduced with
Fig 9 Callichirus islagrande ZV stage a one example of the antenna b one example of pleopods Decapodidstage c antennule d antenna e right mandible f left mandible g maxillule h maxilla (plumose condition notshown) i maxilliped 1 j maxilliped 2 Scales = 02 mm
STRASSER AND FELDER LARVAL DEVELOPMENT IN CALLICHIRUS ISLAGRANDE 113
5ndash11 setae scaphognathite with 37ndash45 se-tae all setae plumose
Maxilliped 1 (Fig 9i)mdashCoxa with 8ndash12 se-tae basis highly setose with 27ndash39 setae en-dopod reduced with 0ndash11 setae exopod un-segmented 6ndash14 setae bilobed epipod with-out setae
Maxilliped 2 (Fig 9j)mdashCoxa partially fusedto basis with 1ndash4 setae basis with 3ndash5 se-
tae endopod 4-segmented (16ndash23) (0 or 1)(6ndash11) (4ndash8) setae exopod with 0ndash8 setaeepipod present but without setae
Maxilliped 3 (Fig 10a)mdashCoxa partiallyfused to basis with 0ndash3 setae basis with 1ndash4setae endopod 5-segmented (11ndash19) (9ndash15)(7ndash11) (8ndash21) (5ndash11) setae exopod usuallyreduced and without setae occasionally with2ndash9 setae epipod present
Fig 10 Callichirus islagrande decapodid stage a maxilliped 3 b pereiopod 1 c pereiopod 2 d pereiopod 3e pereiopod 4 f pereiopod 5 g pleopods from first abdominal somite h pleopods from fourth abdominal somitei telson Scales = 02 mm
114 JOURNAL OF CRUSTACEAN BIOLOGY VOL 20 NO 1 2000
Pereiopod 1 (Fig 10b)mdashChelate biramous7-segmented coxa with 0ndash6 setae basis with2ndash5 setae ischium with 4ndash7 setae and 0ndash3spines merus with 5ndash8 setae carpus with10ndash14 setae propodus with 27ndash37 setae and3ndash4 spines on inner margin of immovable fin-ger dactylus with 21ndash33 setae exopod usu-ally reduced and without setae occasionallywith 4ndash7 setae
Pereiopod 2 (Fig 10c)mdashChelate biramous7-segmented coxa with 2ndash5 setae basis with1ndash5 setae ischium with 4ndash10 setae meruswith 15ndash20 setae carpus with 8ndash12 setaepropodus with 30ndash40 setae and 1ndash4 spineson inner margin of immovable finger dacty-lus with 21ndash30 setae exopod usually reduced0ndash7 setae
Pereiopod 3 (Fig 10d)mdashBiramous 7-seg-mented coxa with 2ndash5 setae basis with 1ndash3setae ischium with 5ndash8 setae merus with6ndash9 setae carpus with 7ndash11 setae propodusbladelike with 25ndash34 setae 1 spine on innerdistal margin dactylus with 15ndash21 setae exo-pod usually reduced 0ndash5 setae
Pereiopod 4 (Fig 10e)mdashBiramous 7-seg-mented coxa with 2ndash7 setae basis with 1ndash4setae ischium with 3ndash8 setae merus with5ndash11 setae carpus with 4ndash9 setae propoduswith 27ndash38 setae 1 spine on inner distal mar-gin dactylus with 15ndash19 setae exopod usu-ally reduced with 0ndash2 setae
Pereiopod 5 (Fig 10f )mdashCheliform unira-mous 7-segmented coxa with 2ndash6 setae ba-sis with l or 2 setae ischium with 2ndash5 setaemerus with 5ndash9 setae carpus with 4ndash7 se-tae propodus with 22ndash30 setae 1 spine onimmovable finger dactylus with 6ndash10 setaeexopod absent
Pleopods (somite 2) (Fig 10g)mdashBiramouscoxa partially fused to basis without setaebasis with 0ndash3 setae endopod with 3ndash5hooked setae on appendix interna 10ndash17 mar-ginal plumose setae 0ndash2 submarginal nakedsetae exopod with 21ndash27 marginal plumosesetae and 0ndash1 submarginal naked seta
Pleopods (somites 3ndash5) (Fig 10h)mdashBiramouscoxa without setae basis with 3ndash6 marginalplumose setae endopod with 3ndash6 hooked se-tae on appendix interna 10ndash18 marginalplumose setae and 0ndash3 naked submarginal se-tae exopod with 20ndash29 marginal plumose se-tae and 0 or 1 submarginal naked seta
Telson and Uropods (Fig 10i)mdashWith 24ndash36processes on posterior margin median pro-cess usually absent or reduced uropods bi-ramous endopod with 19ndash25 plumose setaeexopod with 35ndash58 plumose setae protopodwith 3ndash5 setae
DISCUSSION
As in other species of Callichirus (see Ro-drigues 1976 Aste and Retamal 1983Strasser and Felder in press b) larval devel-opment of C islagrande included four or fivezoeal stages before metamorphosis to the de-capodid ZI larvae may be identified by thepresence of an elongate rounded rostrumsessile eyes and a lack of pereiopodspleopods or uropods (Fig 1a) At ZII lar-vae acquired biramous buds of pereiopods 1and 2 eyes were separated from the carapaceand the rostrum became flattened and serratedlaterally (Fig 1b) Buds of pleopods on ab-dominal segments 2ndash5 pereiopods 3ndash5 anduropods were developed at ZIII (Fig 1c) Inaddition to further development of structuresmentioned thus far ZIV larvae could be dis-tinguished by presence of a distal spine on theexopod and setae on the endopod of each uro-pod (Fig 1d) When it occurred ZV typicallyresembled ZIV but varied widely in mor-phology Appendages at ZV resembled thoseof ZIV the decapodid stage or some grada-tion between the two (see Figs 9a b for ex-amples) The decapodid stage was similar toother species of Callichirus in the reductionof larval characters such as the elongate ros-trum dorsal spines on the abdomen and ex-opods of the pereiopods (Fig le) Acquisitionof fan-like pleopods marked the transition toa primarily abdominal mode of swimming atthis stage
In the Florida Atlantic population of C ma-jor both salinity and presence of putative set-tlement cues at ZIV impacted the observednumber of zoeal stages and the duration ofZIV (Strasser and Felder in preparation1999c) In C islagrande the percentage oflarvae that passed through five versus fourzoeal stages varied (9ndash89) between parentalfemales but was not shown to depend onsalinity or presence of various putative set-tlement cues (Strasser and Felder 1998 inpreparation) Larvae that molted from ZIVto ZV were usually normal morphologicallyand had mortality similar to those that molteddirectly from ZIV to D In contrast larvae of
STRASSER AND FELDER LARVAL DEVELOPMENT IN CALLICHIRUS ISLAGRANDE 115
the Florida Atlantic C major that molted toZV had a higher incidence of mortality anddeformity at D than did those that had by-passed ZV (Strasser and Felder 1999a c)
The number of zoeal stages appears to beless variable in Callichirus garthi and theGulf of Mexico population of C major thanin congeneric populations studied to dateCallichirus garthi was reported to have fivezoeal stages and a long larval duration reach-ing the decapodid stage 48 days after hatch-ing (Aste and Retamal 1983) Developmentof the Gulf population of C major wasshorter in duration (decapodid reached about10 days after hatching) and typically includedfour zoeal stages (Strasser and Felder 1999a)animals rarely passed through three or fivezoeal stages before the molt to the decapodidAlthough the number of stages is reducedfrom that observed in C garthi developmentin the Gulf population of C major cannot beclassified as abbreviated There are species inboth the Callianassinae (Callianassa slkraussi Stebbing C sl kewalramanii and Csl tyrrhena (Petagna)) and Callichirinae(Sergio mirim Lepidophthalmus sinuensisand L louisianensis) that have only two zoealstages before the molt to the decapodid(Forbes 1973 Sankolli and Shenoy 1975Rodrigues 1984 Thessalou-Legaki 1990Nates et al 1997)
Compared to other species of the genusCallichirus C islagrande is morphologicallycloser to C garthi (from Chile) than to theGulf of Mexico population of C major whichoverlaps in distribution with C islagrande Inthe zoeal phase of C islagrande and Cgarthi the telson is concave and the outer-most process on the ventral margin is articu-lated In both C major and Callichirus sp theposterior margin of the telson is straight andthe outermost process is unarticulated duringthe zoeal stages (Rodrigues 1976 Strasserand Felder 1999a) There are also differencesin spination of the abdomen In zoeae of Cislagrande and C garthi the dorsal spine ofabdominal segment 3 is not longer than thosefound on segments 4 and 5 (Aste and Reta-mal 1983) In contrast the dorsal spine of ab-dominal segment 3 is approximately twice aslong as the dorsal spines on segments 4 and5 in early zoeal stages of C major and Cal-lichirus sp (Rodrigues 1976 Aste and Re-tamal 1983 Strasser and Felder 1999a) Thegreatest difference between these groups is
the number of pleopod pairs at zoeal and de-capodid stages Callichirus islagrande and Cgarthi have pleopods on abdominal segments2ndash5 while pleopods are present only on ab-dominal segments 3ndash5 in C major and Cal-lichirus sp (Rodrigues 1976 Aste and Re-tamal 1983 Strasser and Felder 1999a) Al-though Callichirus islagrande and C garthiare similar morphologically there are differ-ences between these two species Callichirusislagrande as well as C major and Cal-lichirus sp develops pleopod buds andchelate pereiopods at ZIII whereas C garthidoes not acquire these characters until ZIV
There are few callianassid species forwhich the larval development has been de-scribed (Sankolli and Shenoy 1975 Ro-drigues 1976 1984 Aste and Retamal 19831984 Konishi et al 1990 Nates et al 1997Strasser and Felder 1999a) all of which aremembers of the Callichirinae and Cal-lianassinae Among species there is a greatdeal of overlap in larval characters makingit difficult to draw distinctions between thesesubfamilies At present information on lar-val development of species in the Callichiri-nae is limited to larval descriptions of Cal-lichirus major C garthi Callichirus sp Cislagrande Sergio mirim Lepidophthalmuslouisianensis and L sinuensis (see Ro-drigues 1976 1984 Aste and Retamal 1983Nates et al 1997 Strasser and Felder1999a) As mentioned above species of Cal-lichirus have been shown to have four to fivezoeal stages while the other members exhibitabbreviated development that consists of onlytwo zoeal stages The most obvious differencebetween larvae of the species of Callichirinaewith abbreviated development is the numberof pleopod pairs Larvae of S mirim havepleopods on abdominal segments 3ndash5whereas L louisianensis and L sinuensishave pleopods on segments 2ndash5 (Rodrigues1984 Nates et al 1997)
Ghost shrimps of the genus Callichirus maybe distinguished from other genera of the Cal-lianassidae by the cuticular pattern on the dor-sal surface of the abdomen and by the strap-like shape of the uropodal endopods (Manningand Felder 1991) Whereas these charactersmay be used to distinguish adults of the genusCallichirus they are not present at zoeal anddecapodid stages Larvae may be distin-guished from all other callianassid genera byhaving four to five zoeal stages and numer-
116 JOURNAL OF CRUSTACEAN BIOLOGY VOL 20 NO 1 2000
ous processes (12ndash25+1+12ndash25) on the pos-terior margin of the telson during the earlyzoeal stages Larvae of Callichirus may alsodiffer from other callianassids with extendeddevelopment by the retention of the anomu-ran hair as the second process on the poste-rior margin of the telson in the late zoealstages Whereas this process is converted toa spine in Neotrypaea uncinata at ZV and Cal-lianassa sl petalura at ZIV the anomuranhair is retained at ZIV in Callichirus majorand C islagrande (see Rodrigues 1976Strasser and Felder 1999a) Although notmentioned specifically in the descriptions theanomuran hair appears to be present in illus-trations of Callichirus sp at ZIII and Cgarthii at ZV (Rodrigues 1976 fig 35 Asteand Retamal 1983 fig 8Q) A bilobed epi-pod on the first maxilliped of ZI is consistentlyfound in Callichirus but is also present at ZIin Lepidophthalmus sinuensis (see Nates etal 1997) Other callianassids acquire thischaracter at ZII (L louisianensis Nates et al1997 Callianassa sl kewlramanii Sankolliand Shenoy 1975) at ZV (Neotrypaea unci-nata Aste and Retamal 1984) or at the de-capodid stage (Sergio mirim Rodrigues 1984C sl petalura Konishi et al 1990) Thusthe number of zoeal stages and setation of thetelson may serve as the best characters to dis-tinguish larvae of Callichirus from other gen-era However since larval descriptions arelacking for many callianassid species it is un-known if these characters (in combination)will prove to be diagnostic until descriptionsfor other species are published
ACKNOWLEDGEMENTS
We thank Robert Bourgeois Barbara Hasek JimMaKinster Sergio Nates and Kurt Strasser who pro-vided assistance in the field and laboratory and MaryE Rice Director of the Smithsonian Marine StationLink Port Florida who facilitated access to station fa-cilities We also thank R Bauer R Jaeger R Lemaitreand G Watson for their early comments on this manu-script and Raymond B Manning who facilitated fund-ing and gave advice on collecting sites Direct supportfor this study was provided to Karen M Strasser by adoctoral fellowship from the University of Louisianaa graduate student research grant from the LUMCON(Louisiana Universities Marine Consortium) Founda-tion and a summer research award from the Smith-sonian Marine Station Additional support for field andlab activities was provided to D L Felder under Loui-siana Sea Grant College Program Grant RCFB-21 USFish and Wildlife Service Cooperative Agreement 14-16-0009-89-96 (Task Order No 6) US Department of En-ergy grant DE-FG02-97ER12220 and several small proj-ect grants from the Smithsonian Marine Station Views
expressed herein are those of the authors and do not nec-essarily reflect views of supporting government agenciesor subagencies mentioned above This is contributionNumber 466 of the Smithsonian Marine Station and con-tribution Number 67 of the ULL Laboratory for Crus-tacean Research All experiments conducted in this studycomply with current applicable state and federal laws
LITERATURE CITED
Aste A and M Retamal 1983 Desarrollo larval deCallianassa garthi Retamal 1975 bajo condiciones delaboratoriomdashCiencia y Tecnologiacutea del Mar CONA 75ndash26
mdashmdashmdash and mdashmdashmdash 1984 Desarrollo larval de Cal-lianassa uncinata H Milne Edwards1837 (DecapodaCallianassidae) bajo condiciones de laboratoriomdashGayana Zooloacutegica 48 41ndash56
Felder D L 1978 Osmotic and ionic regulation in sev-eral western Atlantic Callianassidae (Crustacea De-capoda Thalassinidea)mdashBiological Bulletin 154409ndash429
mdashmdashmdash and R B Griffis 1994 Dominant infaunalcommunities at risk in shoreline habitats burrowingthalassinid CrustaceamdashOCS Study number MMS94ndash0007 United States Department of the InteriorMinerals Management Service Gulf of Mexico OCSRegional Office New Orleans Louisiana Pp 1ndash87
Forbes A T 1973 An unusual abbreviated larval lifehistory in the estuarine burrowing prawn Callianassakraussi (Crustacea Decapoda Thalassinidea)mdashMa-rine Biology 22 361ndash365
Konishi K 1989 Larval development of the mudshrimp Upogebia (Upogebia) major (de Haan) (Crus-tacea Thalassinidea Upogebiidae) under laboratoryconditions with comments on larval characters of tha-lassinid familiesmdashBulletin of the National ResearchInstitute of Aquaculture 15 1ndash17
mdashmdashmdash R Quintana and Y Fukuda 1990 A completedescription of larval stages of the ghost shrimp Cal-lianassa petalura Stimpson (Crustacea ThalassinideaCallianassidae) under laboratory conditionsmdashBulletinof the National Research Institute of Aquaculture 1727ndash49
Manning R B and D L Felder 1986 The status ofthe callianassid genus Callichirus Stimpson 1866(Crustacea Decapoda Thalassinidea)mdashProceedingsof the Biological Society of Washington 99 437ndash443
mdashmdashmdash and mdashmdashmdash 1991 Revision of the AmericanCallianassidae (Crustacea Decapoda Thalassinidea)mdashProceedings of the Biological Society of Washington104 764ndash794
Nates S F D L Felder and R Lemaitre 1997 Com-parative larval development in two species of the bur-rowing ghost shrimp genus Lepidophthalmus (Decap-oda Callianassidae)mdashJournal of Crustacean Biology17 497ndash519
Rodrigues S de A 1976 Sobre a reproduccedilatildeo embri-ologia e desenvolvimento larval de Callichirus majorSay 1818 (Crustacea Decapoda Thalassinidea)mdashBo-letim de Zoologia Universidade de Satildeo Paulo 185ndash104
mdashmdashmdash 1984 Desenvolvimento poacutes-embrionaacuterio deCallichirus mirim (Rodrigues 1971) obtido emcondiccedilotildees artificiais (Crustacea Decapoda Tha-lassinidea)mdashBoletim de Zoologia Universidade deSatildeo Paulo 8 239ndash256
STRASSER AND FELDER LARVAL DEVELOPMENT IN CALLICHIRUS ISLAGRANDE 117
Sankolli K N and S Shenoy 1975 Larval develop-ment of mud shrimp Callianassa (Callichirus) kewal-ramanii Sankolli in the laboratory (Crustacea Decap-oda)mdashBulletin of the Department of Marine ScienceUniversity of Cochin 4 704ndash720
Staton J L and D L Felder 1995 Genetic variationin populations of the ghost shrimp genus Callichirus(Crustacea Decapoda Thalassinoidea) in the westernAtlantic and Gulf of MexicomdashBulletin of Marine Sci-ence 56 523ndash536
Strasser K M and D L Felder 1998 Settlement cuesin successive developmental stages of the ghost shrimpCallichirus major and C islagrande (Crustacea De-capoda Thalassinidea)mdashMarine Biology
mdashmdashmdash and mdashmdashmdash 1999a Larval development of twopopulations of the ghost shrimp Callichirus major (De-capoda Thalassinidea)mdashJournal of Crustacean Biology
mdashmdashmdash and mdashmdashmdash 1999b Sand as a stimulus for set-tlement in the ghost shrimp Callichirus major (Say)and C islagrande (Schmitt) (Crustacea DecapodaThalassinidea)mdashJournal of Experimental Marine Bi-ology and Ecology 239 211ndash222
mdashmdashmdash and mdashmdashmdash 1999c Settlement cues in theFlorida Atlantic population of the ghost shrimp Cal-lichirus major (Crustacea Decapoda Thalassinidea)mdashMarine Ecology Progress Series
Thessalou-Legaki M 1990 Advanced larval develop-ment of Callianassa tyrrhena (Decapoda Tha-lassinidea) and the effect of environmental factorsmdashJournal of Crustacean Biology 10 659ndash666
RECEIVED 19 September 1998ACCEPTED 3 December 1998

STRASSER AND FELDER LARVAL DEVELOPMENT IN CALLICHIRUS ISLAGRANDE 103
tae on distal lobe basal edite bilobed with6ndash9 setae on each lobe endopod with 5 lobes3+2+2+2+(2 or 3) setae scaphognathite with16ndash19 setae all setae plumose
Maxilliped 1 (Fig 3g)mdashCoxa with 4ndash8 se-tae basis with 12ndash17 setae endopod 4-seg-mented (3 or 4) (1 or 2) (1ndash3) (3 or 4) se-tae exopod 2-segmented proximal segment
without setae distal segment with 5 or 6 se-tae on distal margin bilobed epipod withoutsetae all setae plumose
Maxilliped 2 (Fig 3h)mdashCoxa with 1 or 2 se-tae basis with 3ndash6 setae endopod 4-seg-mented (4 or 5) (0 or 1) (1 or 2) (3ndash6) se-tae exopod unsegmented 6ndash8 setae on dis-tal margin all setae plumose
Fig 2 Callichirus islagrande dorsal view of larvae and decapodid a ZI stage b ZII stage c ZIII stage d ZIVstage e decapodid stage Scale = 1 mm
104 JOURNAL OF CRUSTACEAN BIOLOGY VOL 20 NO 1 2000
Maxilliped 3 (Fig 3i)mdashCoxa with 0 or 1seta basis with 1 or 2 setae endopod 4-seg-mented (0 or 1) (0ndash2) (2 or 3) (3ndash5) se-tae exopod unsegmented 6ndash8 setae on dis-tal margin all setae plumose
PereiopodsmdashNot developed
PleopodsmdashNot developed
Telson (Fig 3j)mdashNarrow anterior not dif-ferentiated from abdominal segment 6 trian-gular posterior anal spine present with(12ndash14)+1+(13 or 14) processes on posterior
margin arranged as (I II III IV III II I)(I) outermost process a plumodenticulate seta(II) second process ldquoanomuran hairrdquo (III)10ndash12 plumodenticulate setae and (IV) cen-ter process a naked spine longer than otherprocesses
Zoea IIFigs 2b 4andashl
SizemdashCL = 144 plusmn 003 mm range 138ndash153mm TL = 364 plusmn 006 mm range 355ndash380mm
Fig 3 Callichirus islagrande ZI stage a antennule b antenna c right mandible d left mandible e maxillulef maxilla (plumose condition not shown) g maxilliped 1 h maxilliped 2 i maxilliped 3 j telson Scales = 02mm
STRASSER AND FELDER LARVAL DEVELOPMENT IN CALLICHIRUS ISLAGRANDE 105
Carapace (Fig 2b)mdashWith antennal spinesanterolateral margins serrated rostrum broadand dorsoventrally flattened with thin distalprojection and distolateral serrations eyesseparated from carapace
Abdomen (Fig 2b)mdashNo marked change fromstage ZI
Antennule (Fig 4a)mdashElongate weaklybilobed peduncle weakly 2-segmented 0ndash3short setae on distal margin of proximal seg-ment 2 long plumose setae and 0ndash3 short se-tae on distal end of distal segment endopodwith 1 terminal plumose seta exopodite with6ndash8 terminal aesthetascs
Antenna (Fig 4b)mdashProtopod with 2 distalspines 1 on outer margin and 1 between ramiendopod without setae scaphocerite armedwith 1 strong distolateral spine 10 or 11plumose setae on inner margin and 1plumose seta on outer margin
Mandible (Fig 4c d)mdashNo marked changefrom ZI
Maxillule (Fig 4e)mdashCoxal endite with 8ndash10marginal plumose setae basal endite with 3plumose setae 4 or 5 large dentate spines and4 or 5 smaller dentate spines inner marginwith 0 or 1 plumose seta endopod 3-seg-mented distinction between segments some-times obscure 2 2 (4 or 5) setae protopodwithout setae
Maxilla (Fig 4f )mdashCoxal endite bilobed12ndash15 setae on proximal lobe 4 or 5 setaeon distal lobe basal endite bilobed with 6ndash8setae on proximal lobe 5ndash9 setae on distallobe endopod with 5 lobes (2 or 3)+2+2+2+3setae scaphognathite with 19ndash24 setae allsetae plumose
Maxilliped 1 (Fig 4g)mdashCoxa with 4ndash8 se-tae basis with 13ndash17 marginal setae endo-pod 4-segmented (3 or 4) (2 or 3) (1 or 2)4 setae exopod 2-segmented proximal seg-ment without setae distal segment with 6 or7 setae on distal margin bilobed epipod with-out setae all setae plumose
Maxilliped 2 (Fig 4h)mdashCoxa with 0ndash2 se-tae basis with 4ndash6 setae endopod 4-seg-mented (4ndash7) 1 (2 or 3) (4ndash6) setae exo-pod unsegmented 7 or 8 setae on distal mar-gin all setae plumose
Maxilliped 3 (Fig 4i)mdashCoxa without setaebasis with 0ndash2 setae endopod 4-segmented
(l or 2) (1ndash3) (2ndash4) (3ndash6) setae exopod un-segmented 6ndash8 setae on distal margin all se-tae plumose
Pereiopods (Fig 4j k)mdashFirst and second pe-reiopod present both without setae
PleopodsmdashNot developed
Telson (Fig 4l)mdashNo change from ZI
Zoea IIIFigs 2c 5andashh 6andashh
SizemdashCL = 165 plusmn 003 mm range 160ndash172mm TL = 413 plusmn 009 mm range 399ndash436mm
Carapace (Fig 2c)mdashNo marked changefrom ZII
Abdomen (Fig 2c)mdashSomites 2ndash5 with pos-terolateral projections dorsal spine elongateon somite 2 sometimes extending beyondposterior margin of somite 3 short dorsalspines on somites 3ndash6
Antennule (Fig 5a)mdashBiramous peduncle2-segmented proximal segment with (0 or1)+(2 or 3) short setae on outer margin and1 long plumose seta on inner margin distalsegment with 2 or 3 setae on inner margin 1seta between rami and 0ndash3 short seta onouter margin endopod with 0 or 1 terminalplumose seta exopod with 6 or 7 terminaland 0ndash2 subterminal aesthetascs
Antenna (Fig 5b)mdashProtopod with 1 spineat outer distal margin endopod with 0 or 1short seta scaphocerite armed with 1 strongdistolateral spine 11ndash13 plumose setae on in-ner margin and 1 plumose seta on outer mar-gin
Mandible (Fig 5c d)mdashIncisor process with7 or 8 teeth on right mandible 6ndash8 teeth onleft mandible molar process denticulate onboth mandibles buds of palps present
Maxillule (Fig 5e)mdashCoxal endite with10ndash12 marginal plumose setae basal enditewith 3 plumose setae 4 or 5 large dentatespines and 4 or 5 smaller dentate spinesinner margin with 1 plumose seta endopod3-segmented distinction between segmentssometimes obscure 2 2 4 setae protopodwithout setation
Maxilla (Fig 5f )mdashCoxal endite bilobed14ndash17 setae on proximal lobe 4ndash6 setae ondistal lobe basal endite bilobed with 7ndash10 se-
106 JOURNAL OF CRUSTACEAN BIOLOGY VOL 20 NO 1 2000
tae on proximal lobe 6ndash9 setae on distal lobeendopod with 5 lobes 3+2+2+2+(2 or 3) se-tae scaphognathite with 21ndash26 setae all se-tae plumose
Maxilliped 1 (Fig 5g)mdashCoxa with 4ndash9 se-tae basis with 14ndash18 marginal setae endo-pod 4-segmented (3 or 4) 2 2 (3 or 4) se-tae exopod 2-segmented proximal segment
without setae distal segment with 5ndash7 setaeon distal margin bilobed epipod without se-tae all setae plumose
Maxilliped 2 (Fig 5h)mdashCoxa with 0ndash2 se-tae basis with 5 or 6 setae endopod 4-seg-mented (4ndash6) 1 3 (4ndash6) setae exopod un-segmented 7 or 8 setae on distal margin allsetae plumose
Fig 4 Callichirus islagrande ZII stage a antennule b antenna c right mandible d left mandible e maxillulef maxilla (plumose condition not shown) g maxilliped 1 h maxilliped 2 i maxilliped 3 j pereiopod 1 k pe-reiopod 2 1 telson Scales = 02 mm
STRASSER AND FELDER LARVAL DEVELOPMENT IN CALLICHIRUS ISLAGRANDE 107
Maxilliped 3 (Fig 6a)mdashCoxa with 0 or 1seta basis with 1ndash3 setae endopod 4-seg-mented (1 or 2) 2 (2ndash4) (4 or 5) setae exo-pod unsegmented 7 or 8 setae on distal mar-gin all setae plumose
Pereiopod 1 (Fig 6b)mdashChelate and bira-mous coxa without setae basis with 0 or 1seta endopod 4-segmented 0 (1 or 2) (2ndash4)2 setae exopod with 6 or 7 setae on distalmargin
Fig 5 Callichirus islagrande ZIII stage a antennule b antenna c right mandible d left mandible e maxil-lule f maxilla (plumose condition not shown) g maxilliped 1 h maxilliped 2 All setae on the maxilla were plumoseScales = 02 mm
108 JOURNAL OF CRUSTACEAN BIOLOGY VOL 20 NO 1 2000
Pereiopod 2 (Fig 6c)mdashChelate and biramouscoxa without setae basis with 0 or 1 seta en-dopod 4-segmented 0 (0ndash2) (0ndash4) 2 setaeexopod with 5ndash7 setae on distal margin
Pereiopods 3ndash5 (Fig 6d e f )mdashWithout setae
Pleopods (Figs 1c 6g)mdashBilobed buds onabdominal segments 2ndash5 first pair smallerthan others
Telson and Uropods (Fig 6h)mdashNo changein arrangement of posterior telsonal processesuropods biramous endopod without setae
Fig 6 Callichirus islagrande ZIII stage a maxilliped 3 b pereiopod 1 c pereiopod 2 d pereiopod 3 e pe-reiopod 4 f pereiopod 5 g pleopods from fourth abdominal somite h telson Scales = 02 mm
STRASSER AND FELDER LARVAL DEVELOPMENT IN CALLICHIRUS ISLAGRANDE 109
exopod with 10 or 11 marginal plumose se-tae ventral spine near base of protopod
Zoea IVFigs 2d 7andashh 8andashi
SizemdashCL = 192 plusmn 009 mm range 169ndash215mm TL = 482 plusmn 025 mm range 424ndash536mm
Carapace (Fig 2d)mdashNo marked changefrom ZIII
Abdomen (Fig 2d)mdashNo marked change fromZIII
Antennule (Fig 7a)mdashBiramous peduncle2-segmented with 1+(1ndash4) short setae onouter margin and 0 or 1 plumose seta on in-ner margin of proximal segment distal seg-ment with 3ndash5 plumose setae on inner mar-gin 1 plumose seta between rami and 0ndash3short setae on outer margin endopod with 1or 2 plumose setae exopod with 6 or 7 ter-minal and 2 subterminal aesthetascs
Antenna (Fig 7b)mdashProtopod with 1 distalspine on outer margin endopod with 0ndash2 se-tae scaphocerite armed with 1 strong disto-lateral spine 12ndash14 plumose setae on innermargin and 1 plumose seta on outer margin
Mandible (Fig 7c d)mdashIncisor process with8 teeth on right mandible 6ndash8 teeth on leftmandible molar process denticulate on bothmandibles palps present
Maxillule (Fig 7e)mdashCoxal endite with12ndash14 marginal plumose setae basal enditewith 3 plumose setae 5 or 6 large dentatespines and 5 or 6 smaller dentate spinesinner margin with 1 plumose seta endopod3-segmented distinction between segmentssometimes obscure 2 2 (3 or 4) setae pro-topod without setation
Maxilla (Fig 7f )mdashCoxal endite bilobed14ndash19 setae on proximal lobe 5 setae on dis-tal lobe basal endite bilobed 6ndash10 setae onproximal lobe 8 or 9 setae on distal lobeendopod with 5 lobes 3+(2 or 3)+2+2+3 se-tae scaphognathite with 27ndash32 setae all se-tae plumose
Maxilliped 1 (Fig 7g)mdashCoxa with 2ndash8 se-tae basis with 16ndash19 marginal setae endo-pod 4-segmented (3 or 4) 2 2 4 setae exo-
pod 2-segmented proximal segment withoutsetae distal segment with 6 or 7 setae on dis-tal margin bilobed epipod without setae allsetae plumose
Maxilliped 2 (Fig 7h)mdashCoxa with 0ndash3 se-tae basis with 4ndash6 setae endopod 4-seg-mented (5ndash8) 1 (3ndash5) (4 or 5) setae exo-pod unsegmented 7 or 8 setae on distal mar-gin all setae plumose
Maxilliped 3 (Fig 8a)mdashCoxa with 0 or 1seta basis with 2ndash4 setae endopod 4-seg-mented (l or 2) 2 (3ndash5) (3 or 4) setae exo-pod unsegmented 7 or 8 setae on distal mar-gin all setae plumose
Pereiopod 1 (Fig 8b)mdashBiramous cheliformcoxa without setae basis with 0ndash2 setae en-dopod 4-segmented (0 or 1) (1 or 2) (2ndash4)(1 or 2) setae exopod with 5ndash7 setae on dis-tal margin
Pereiopod 2 (Fig 8c)mdashBiramous cheliformcoxa without setae basis with 0 or 1 setaendopod 4-segmented 0 (1 or 2) (3ndash5) (2or 3) setae exopod with 5ndash7 setae on distalmargin
Pereiopod 3 (Fig 8d)mdashBiramous coxa andbasis without setae endopod 4-segmented0 (0ndash2) (3 or 4) 2 setae propodus blade-like exopod with 4 or 5 terminal setae
Pereiopod 4 (Fig 8e)mdashBiramous coxa andbasis without setae endopod 4-segmented (0or 1) (0ndash2) (2ndash4) 2 setae exopod with 0ndash2setae
Pereiopod 5 (Fig 8f )mdashUniramous 2 setaeon terminal segment 4 or 5 setae on adja-cent segment
Pleopods (Fig 8g h)mdashLarger than at ZIIIstill without setae
Telson and Uropods (Fig 8i)mdashTelson trian-gular narrow anteriorly no anal spine pos-terior margin with (12ndash15)+1+(13 or 14)processes arranged as (I II III IV III II I)(I) outermost process a plumodenticulate seta(II) second process ldquoanomuran hairrdquo (III) 10ndash13plumodenticulate setae and (IV) naked spineuropods biramous endopod with 7ndash11 marginalplumose setae exopod with 1 naked distolat-eral spine 12ndash14 marginal plumose setae
110 JOURNAL OF CRUSTACEAN BIOLOGY VOL 20 NO 1 2000
Decapodid (first postlarva)Figs 2e 9cndashj l0andashi
SizemdashCL = 133 plusmn 002 mm range129ndash138 mm TL = 378 plusmn 011 mm range361ndash405 mm
Carapace (Fig 2e)mdashShorter than abdomenno anterolateral serrations with pterygostomial
projections rostrum reduced without serra-tions sometimes bending down between eyes
Abdomen (Fig 2e)mdashSomites broad and flat-tened posterior projections rounded with 0ndash7setae somite 2 rectangular somites 3ndash5 oval
Antennule (Fig 9c)mdashPeduncle 3-segmentedproximal segment with 17ndash24 setae 13ndash15 of
Fig 7 Callichirus islagrande ZIV stage a antennule b antenna c right mandible d left mandible e maxil-lule f maxilla (plumose condition not shown) g maxilliped 1 h maxilliped 2 All setae on the maxilla were plumoseScales = 02 mm
STRASSER AND FELDER LARVAL DEVELOPMENT IN CALLICHIRUS ISLAGRANDE 111
these associated with statocyst intermediatesegment with 6ndash10 setae distal segment of pe-duncle with 2 vertical rows of setae one with4 or 5 setae other with 2ndash4 setae 1ndash3 addi-tional setae on outer margin and 3ndash5 short se-tae on distal margin endopod 4ndash6 segmented(1ndash3) (0 or 1) (1ndash4) (1ndash4) (3ndash6) (3ndash6) se-tae exopod 3ndash5-segmented 0 (0ndash4) (0ndash4)(1ndash4) setae +2 asthetascs (5ndash8) asthetascs ()
Antenna (Fig 9d)mdashPeduncle 2-segmentedproximal segment with 0 or 1 seta distal seg-ment with 1ndash3 setae on inner margin spinepresent in some flagellum ranging from 14ndash18
segments (0 or 1) (2ndash4) variable region(1ndash3 articles with 0ndash2 setae) (0ndash4) (0ndash2)(2ndash5) (1ndash4) (3ndash6) (0ndash3) (2ndash5) (3ndash6)(0ndash3) (3ndash6) (0 or 1) (1ndash7) (4ndash9) setae exo-pod reduced to small lobe with 0ndash5 setae
Mandible (Fig 9e f )mdashBoth mandibles sim-ilar incisor process with 5ndash8 teeth molarprocess smooth or with small ridges palp3-segmented 0 (0 or 1) (8ndash12) setae
Maxillule (Fig 9g)mdashCoxal endite with15ndash22 marginal setae and 0 or 1 submarginal
Fig 8 Callichirus islagrande ZIV stage a maxilliped 3 b pereiopod 1 c pereiopod 2 d pereiopod 3 e pe-reiopod 4 f pereiopod 5 g pleopods from first abdominal somite h pleopods from fourth abdominal somite i tel-son Scales = 02 mm
112 JOURNAL OF CRUSTACEAN BIOLOGY VOL 20 NO 1 2000
seta basal endite with 3ndash5 plumose setae14ndash19 dentate spines 0ndash2 plumose setae oninner margin endopod reduced unsegmentedwith 0ndash8 setae
Maxilla (Fig 9h)mdashCoxal endite trilobed(17ndash21)+(7ndash10)+(6ndash9) setae basal enditebilobed 8ndash10 setae on proximal lobe 8ndash15setae on distal lobe endopod reduced with
Fig 9 Callichirus islagrande ZV stage a one example of the antenna b one example of pleopods Decapodidstage c antennule d antenna e right mandible f left mandible g maxillule h maxilla (plumose condition notshown) i maxilliped 1 j maxilliped 2 Scales = 02 mm
STRASSER AND FELDER LARVAL DEVELOPMENT IN CALLICHIRUS ISLAGRANDE 113
5ndash11 setae scaphognathite with 37ndash45 se-tae all setae plumose
Maxilliped 1 (Fig 9i)mdashCoxa with 8ndash12 se-tae basis highly setose with 27ndash39 setae en-dopod reduced with 0ndash11 setae exopod un-segmented 6ndash14 setae bilobed epipod with-out setae
Maxilliped 2 (Fig 9j)mdashCoxa partially fusedto basis with 1ndash4 setae basis with 3ndash5 se-
tae endopod 4-segmented (16ndash23) (0 or 1)(6ndash11) (4ndash8) setae exopod with 0ndash8 setaeepipod present but without setae
Maxilliped 3 (Fig 10a)mdashCoxa partiallyfused to basis with 0ndash3 setae basis with 1ndash4setae endopod 5-segmented (11ndash19) (9ndash15)(7ndash11) (8ndash21) (5ndash11) setae exopod usuallyreduced and without setae occasionally with2ndash9 setae epipod present
Fig 10 Callichirus islagrande decapodid stage a maxilliped 3 b pereiopod 1 c pereiopod 2 d pereiopod 3e pereiopod 4 f pereiopod 5 g pleopods from first abdominal somite h pleopods from fourth abdominal somitei telson Scales = 02 mm
114 JOURNAL OF CRUSTACEAN BIOLOGY VOL 20 NO 1 2000
Pereiopod 1 (Fig 10b)mdashChelate biramous7-segmented coxa with 0ndash6 setae basis with2ndash5 setae ischium with 4ndash7 setae and 0ndash3spines merus with 5ndash8 setae carpus with10ndash14 setae propodus with 27ndash37 setae and3ndash4 spines on inner margin of immovable fin-ger dactylus with 21ndash33 setae exopod usu-ally reduced and without setae occasionallywith 4ndash7 setae
Pereiopod 2 (Fig 10c)mdashChelate biramous7-segmented coxa with 2ndash5 setae basis with1ndash5 setae ischium with 4ndash10 setae meruswith 15ndash20 setae carpus with 8ndash12 setaepropodus with 30ndash40 setae and 1ndash4 spineson inner margin of immovable finger dacty-lus with 21ndash30 setae exopod usually reduced0ndash7 setae
Pereiopod 3 (Fig 10d)mdashBiramous 7-seg-mented coxa with 2ndash5 setae basis with 1ndash3setae ischium with 5ndash8 setae merus with6ndash9 setae carpus with 7ndash11 setae propodusbladelike with 25ndash34 setae 1 spine on innerdistal margin dactylus with 15ndash21 setae exo-pod usually reduced 0ndash5 setae
Pereiopod 4 (Fig 10e)mdashBiramous 7-seg-mented coxa with 2ndash7 setae basis with 1ndash4setae ischium with 3ndash8 setae merus with5ndash11 setae carpus with 4ndash9 setae propoduswith 27ndash38 setae 1 spine on inner distal mar-gin dactylus with 15ndash19 setae exopod usu-ally reduced with 0ndash2 setae
Pereiopod 5 (Fig 10f )mdashCheliform unira-mous 7-segmented coxa with 2ndash6 setae ba-sis with l or 2 setae ischium with 2ndash5 setaemerus with 5ndash9 setae carpus with 4ndash7 se-tae propodus with 22ndash30 setae 1 spine onimmovable finger dactylus with 6ndash10 setaeexopod absent
Pleopods (somite 2) (Fig 10g)mdashBiramouscoxa partially fused to basis without setaebasis with 0ndash3 setae endopod with 3ndash5hooked setae on appendix interna 10ndash17 mar-ginal plumose setae 0ndash2 submarginal nakedsetae exopod with 21ndash27 marginal plumosesetae and 0ndash1 submarginal naked seta
Pleopods (somites 3ndash5) (Fig 10h)mdashBiramouscoxa without setae basis with 3ndash6 marginalplumose setae endopod with 3ndash6 hooked se-tae on appendix interna 10ndash18 marginalplumose setae and 0ndash3 naked submarginal se-tae exopod with 20ndash29 marginal plumose se-tae and 0 or 1 submarginal naked seta
Telson and Uropods (Fig 10i)mdashWith 24ndash36processes on posterior margin median pro-cess usually absent or reduced uropods bi-ramous endopod with 19ndash25 plumose setaeexopod with 35ndash58 plumose setae protopodwith 3ndash5 setae
DISCUSSION
As in other species of Callichirus (see Ro-drigues 1976 Aste and Retamal 1983Strasser and Felder in press b) larval devel-opment of C islagrande included four or fivezoeal stages before metamorphosis to the de-capodid ZI larvae may be identified by thepresence of an elongate rounded rostrumsessile eyes and a lack of pereiopodspleopods or uropods (Fig 1a) At ZII lar-vae acquired biramous buds of pereiopods 1and 2 eyes were separated from the carapaceand the rostrum became flattened and serratedlaterally (Fig 1b) Buds of pleopods on ab-dominal segments 2ndash5 pereiopods 3ndash5 anduropods were developed at ZIII (Fig 1c) Inaddition to further development of structuresmentioned thus far ZIV larvae could be dis-tinguished by presence of a distal spine on theexopod and setae on the endopod of each uro-pod (Fig 1d) When it occurred ZV typicallyresembled ZIV but varied widely in mor-phology Appendages at ZV resembled thoseof ZIV the decapodid stage or some grada-tion between the two (see Figs 9a b for ex-amples) The decapodid stage was similar toother species of Callichirus in the reductionof larval characters such as the elongate ros-trum dorsal spines on the abdomen and ex-opods of the pereiopods (Fig le) Acquisitionof fan-like pleopods marked the transition toa primarily abdominal mode of swimming atthis stage
In the Florida Atlantic population of C ma-jor both salinity and presence of putative set-tlement cues at ZIV impacted the observednumber of zoeal stages and the duration ofZIV (Strasser and Felder in preparation1999c) In C islagrande the percentage oflarvae that passed through five versus fourzoeal stages varied (9ndash89) between parentalfemales but was not shown to depend onsalinity or presence of various putative set-tlement cues (Strasser and Felder 1998 inpreparation) Larvae that molted from ZIVto ZV were usually normal morphologicallyand had mortality similar to those that molteddirectly from ZIV to D In contrast larvae of
STRASSER AND FELDER LARVAL DEVELOPMENT IN CALLICHIRUS ISLAGRANDE 115
the Florida Atlantic C major that molted toZV had a higher incidence of mortality anddeformity at D than did those that had by-passed ZV (Strasser and Felder 1999a c)
The number of zoeal stages appears to beless variable in Callichirus garthi and theGulf of Mexico population of C major thanin congeneric populations studied to dateCallichirus garthi was reported to have fivezoeal stages and a long larval duration reach-ing the decapodid stage 48 days after hatch-ing (Aste and Retamal 1983) Developmentof the Gulf population of C major wasshorter in duration (decapodid reached about10 days after hatching) and typically includedfour zoeal stages (Strasser and Felder 1999a)animals rarely passed through three or fivezoeal stages before the molt to the decapodidAlthough the number of stages is reducedfrom that observed in C garthi developmentin the Gulf population of C major cannot beclassified as abbreviated There are species inboth the Callianassinae (Callianassa slkraussi Stebbing C sl kewalramanii and Csl tyrrhena (Petagna)) and Callichirinae(Sergio mirim Lepidophthalmus sinuensisand L louisianensis) that have only two zoealstages before the molt to the decapodid(Forbes 1973 Sankolli and Shenoy 1975Rodrigues 1984 Thessalou-Legaki 1990Nates et al 1997)
Compared to other species of the genusCallichirus C islagrande is morphologicallycloser to C garthi (from Chile) than to theGulf of Mexico population of C major whichoverlaps in distribution with C islagrande Inthe zoeal phase of C islagrande and Cgarthi the telson is concave and the outer-most process on the ventral margin is articu-lated In both C major and Callichirus sp theposterior margin of the telson is straight andthe outermost process is unarticulated duringthe zoeal stages (Rodrigues 1976 Strasserand Felder 1999a) There are also differencesin spination of the abdomen In zoeae of Cislagrande and C garthi the dorsal spine ofabdominal segment 3 is not longer than thosefound on segments 4 and 5 (Aste and Reta-mal 1983) In contrast the dorsal spine of ab-dominal segment 3 is approximately twice aslong as the dorsal spines on segments 4 and5 in early zoeal stages of C major and Cal-lichirus sp (Rodrigues 1976 Aste and Re-tamal 1983 Strasser and Felder 1999a) Thegreatest difference between these groups is
the number of pleopod pairs at zoeal and de-capodid stages Callichirus islagrande and Cgarthi have pleopods on abdominal segments2ndash5 while pleopods are present only on ab-dominal segments 3ndash5 in C major and Cal-lichirus sp (Rodrigues 1976 Aste and Re-tamal 1983 Strasser and Felder 1999a) Al-though Callichirus islagrande and C garthiare similar morphologically there are differ-ences between these two species Callichirusislagrande as well as C major and Cal-lichirus sp develops pleopod buds andchelate pereiopods at ZIII whereas C garthidoes not acquire these characters until ZIV
There are few callianassid species forwhich the larval development has been de-scribed (Sankolli and Shenoy 1975 Ro-drigues 1976 1984 Aste and Retamal 19831984 Konishi et al 1990 Nates et al 1997Strasser and Felder 1999a) all of which aremembers of the Callichirinae and Cal-lianassinae Among species there is a greatdeal of overlap in larval characters makingit difficult to draw distinctions between thesesubfamilies At present information on lar-val development of species in the Callichiri-nae is limited to larval descriptions of Cal-lichirus major C garthi Callichirus sp Cislagrande Sergio mirim Lepidophthalmuslouisianensis and L sinuensis (see Ro-drigues 1976 1984 Aste and Retamal 1983Nates et al 1997 Strasser and Felder1999a) As mentioned above species of Cal-lichirus have been shown to have four to fivezoeal stages while the other members exhibitabbreviated development that consists of onlytwo zoeal stages The most obvious differencebetween larvae of the species of Callichirinaewith abbreviated development is the numberof pleopod pairs Larvae of S mirim havepleopods on abdominal segments 3ndash5whereas L louisianensis and L sinuensishave pleopods on segments 2ndash5 (Rodrigues1984 Nates et al 1997)
Ghost shrimps of the genus Callichirus maybe distinguished from other genera of the Cal-lianassidae by the cuticular pattern on the dor-sal surface of the abdomen and by the strap-like shape of the uropodal endopods (Manningand Felder 1991) Whereas these charactersmay be used to distinguish adults of the genusCallichirus they are not present at zoeal anddecapodid stages Larvae may be distin-guished from all other callianassid genera byhaving four to five zoeal stages and numer-
116 JOURNAL OF CRUSTACEAN BIOLOGY VOL 20 NO 1 2000
ous processes (12ndash25+1+12ndash25) on the pos-terior margin of the telson during the earlyzoeal stages Larvae of Callichirus may alsodiffer from other callianassids with extendeddevelopment by the retention of the anomu-ran hair as the second process on the poste-rior margin of the telson in the late zoealstages Whereas this process is converted toa spine in Neotrypaea uncinata at ZV and Cal-lianassa sl petalura at ZIV the anomuranhair is retained at ZIV in Callichirus majorand C islagrande (see Rodrigues 1976Strasser and Felder 1999a) Although notmentioned specifically in the descriptions theanomuran hair appears to be present in illus-trations of Callichirus sp at ZIII and Cgarthii at ZV (Rodrigues 1976 fig 35 Asteand Retamal 1983 fig 8Q) A bilobed epi-pod on the first maxilliped of ZI is consistentlyfound in Callichirus but is also present at ZIin Lepidophthalmus sinuensis (see Nates etal 1997) Other callianassids acquire thischaracter at ZII (L louisianensis Nates et al1997 Callianassa sl kewlramanii Sankolliand Shenoy 1975) at ZV (Neotrypaea unci-nata Aste and Retamal 1984) or at the de-capodid stage (Sergio mirim Rodrigues 1984C sl petalura Konishi et al 1990) Thusthe number of zoeal stages and setation of thetelson may serve as the best characters to dis-tinguish larvae of Callichirus from other gen-era However since larval descriptions arelacking for many callianassid species it is un-known if these characters (in combination)will prove to be diagnostic until descriptionsfor other species are published
ACKNOWLEDGEMENTS
We thank Robert Bourgeois Barbara Hasek JimMaKinster Sergio Nates and Kurt Strasser who pro-vided assistance in the field and laboratory and MaryE Rice Director of the Smithsonian Marine StationLink Port Florida who facilitated access to station fa-cilities We also thank R Bauer R Jaeger R Lemaitreand G Watson for their early comments on this manu-script and Raymond B Manning who facilitated fund-ing and gave advice on collecting sites Direct supportfor this study was provided to Karen M Strasser by adoctoral fellowship from the University of Louisianaa graduate student research grant from the LUMCON(Louisiana Universities Marine Consortium) Founda-tion and a summer research award from the Smith-sonian Marine Station Additional support for field andlab activities was provided to D L Felder under Loui-siana Sea Grant College Program Grant RCFB-21 USFish and Wildlife Service Cooperative Agreement 14-16-0009-89-96 (Task Order No 6) US Department of En-ergy grant DE-FG02-97ER12220 and several small proj-ect grants from the Smithsonian Marine Station Views
expressed herein are those of the authors and do not nec-essarily reflect views of supporting government agenciesor subagencies mentioned above This is contributionNumber 466 of the Smithsonian Marine Station and con-tribution Number 67 of the ULL Laboratory for Crus-tacean Research All experiments conducted in this studycomply with current applicable state and federal laws
LITERATURE CITED
Aste A and M Retamal 1983 Desarrollo larval deCallianassa garthi Retamal 1975 bajo condiciones delaboratoriomdashCiencia y Tecnologiacutea del Mar CONA 75ndash26
mdashmdashmdash and mdashmdashmdash 1984 Desarrollo larval de Cal-lianassa uncinata H Milne Edwards1837 (DecapodaCallianassidae) bajo condiciones de laboratoriomdashGayana Zooloacutegica 48 41ndash56
Felder D L 1978 Osmotic and ionic regulation in sev-eral western Atlantic Callianassidae (Crustacea De-capoda Thalassinidea)mdashBiological Bulletin 154409ndash429
mdashmdashmdash and R B Griffis 1994 Dominant infaunalcommunities at risk in shoreline habitats burrowingthalassinid CrustaceamdashOCS Study number MMS94ndash0007 United States Department of the InteriorMinerals Management Service Gulf of Mexico OCSRegional Office New Orleans Louisiana Pp 1ndash87
Forbes A T 1973 An unusual abbreviated larval lifehistory in the estuarine burrowing prawn Callianassakraussi (Crustacea Decapoda Thalassinidea)mdashMa-rine Biology 22 361ndash365
Konishi K 1989 Larval development of the mudshrimp Upogebia (Upogebia) major (de Haan) (Crus-tacea Thalassinidea Upogebiidae) under laboratoryconditions with comments on larval characters of tha-lassinid familiesmdashBulletin of the National ResearchInstitute of Aquaculture 15 1ndash17
mdashmdashmdash R Quintana and Y Fukuda 1990 A completedescription of larval stages of the ghost shrimp Cal-lianassa petalura Stimpson (Crustacea ThalassinideaCallianassidae) under laboratory conditionsmdashBulletinof the National Research Institute of Aquaculture 1727ndash49
Manning R B and D L Felder 1986 The status ofthe callianassid genus Callichirus Stimpson 1866(Crustacea Decapoda Thalassinidea)mdashProceedingsof the Biological Society of Washington 99 437ndash443
mdashmdashmdash and mdashmdashmdash 1991 Revision of the AmericanCallianassidae (Crustacea Decapoda Thalassinidea)mdashProceedings of the Biological Society of Washington104 764ndash794
Nates S F D L Felder and R Lemaitre 1997 Com-parative larval development in two species of the bur-rowing ghost shrimp genus Lepidophthalmus (Decap-oda Callianassidae)mdashJournal of Crustacean Biology17 497ndash519
Rodrigues S de A 1976 Sobre a reproduccedilatildeo embri-ologia e desenvolvimento larval de Callichirus majorSay 1818 (Crustacea Decapoda Thalassinidea)mdashBo-letim de Zoologia Universidade de Satildeo Paulo 185ndash104
mdashmdashmdash 1984 Desenvolvimento poacutes-embrionaacuterio deCallichirus mirim (Rodrigues 1971) obtido emcondiccedilotildees artificiais (Crustacea Decapoda Tha-lassinidea)mdashBoletim de Zoologia Universidade deSatildeo Paulo 8 239ndash256
STRASSER AND FELDER LARVAL DEVELOPMENT IN CALLICHIRUS ISLAGRANDE 117
Sankolli K N and S Shenoy 1975 Larval develop-ment of mud shrimp Callianassa (Callichirus) kewal-ramanii Sankolli in the laboratory (Crustacea Decap-oda)mdashBulletin of the Department of Marine ScienceUniversity of Cochin 4 704ndash720
Staton J L and D L Felder 1995 Genetic variationin populations of the ghost shrimp genus Callichirus(Crustacea Decapoda Thalassinoidea) in the westernAtlantic and Gulf of MexicomdashBulletin of Marine Sci-ence 56 523ndash536
Strasser K M and D L Felder 1998 Settlement cuesin successive developmental stages of the ghost shrimpCallichirus major and C islagrande (Crustacea De-capoda Thalassinidea)mdashMarine Biology
mdashmdashmdash and mdashmdashmdash 1999a Larval development of twopopulations of the ghost shrimp Callichirus major (De-capoda Thalassinidea)mdashJournal of Crustacean Biology
mdashmdashmdash and mdashmdashmdash 1999b Sand as a stimulus for set-tlement in the ghost shrimp Callichirus major (Say)and C islagrande (Schmitt) (Crustacea DecapodaThalassinidea)mdashJournal of Experimental Marine Bi-ology and Ecology 239 211ndash222
mdashmdashmdash and mdashmdashmdash 1999c Settlement cues in theFlorida Atlantic population of the ghost shrimp Cal-lichirus major (Crustacea Decapoda Thalassinidea)mdashMarine Ecology Progress Series
Thessalou-Legaki M 1990 Advanced larval develop-ment of Callianassa tyrrhena (Decapoda Tha-lassinidea) and the effect of environmental factorsmdashJournal of Crustacean Biology 10 659ndash666
RECEIVED 19 September 1998ACCEPTED 3 December 1998

104 JOURNAL OF CRUSTACEAN BIOLOGY VOL 20 NO 1 2000
Maxilliped 3 (Fig 3i)mdashCoxa with 0 or 1seta basis with 1 or 2 setae endopod 4-seg-mented (0 or 1) (0ndash2) (2 or 3) (3ndash5) se-tae exopod unsegmented 6ndash8 setae on dis-tal margin all setae plumose
PereiopodsmdashNot developed
PleopodsmdashNot developed
Telson (Fig 3j)mdashNarrow anterior not dif-ferentiated from abdominal segment 6 trian-gular posterior anal spine present with(12ndash14)+1+(13 or 14) processes on posterior
margin arranged as (I II III IV III II I)(I) outermost process a plumodenticulate seta(II) second process ldquoanomuran hairrdquo (III)10ndash12 plumodenticulate setae and (IV) cen-ter process a naked spine longer than otherprocesses
Zoea IIFigs 2b 4andashl
SizemdashCL = 144 plusmn 003 mm range 138ndash153mm TL = 364 plusmn 006 mm range 355ndash380mm
Fig 3 Callichirus islagrande ZI stage a antennule b antenna c right mandible d left mandible e maxillulef maxilla (plumose condition not shown) g maxilliped 1 h maxilliped 2 i maxilliped 3 j telson Scales = 02mm
STRASSER AND FELDER LARVAL DEVELOPMENT IN CALLICHIRUS ISLAGRANDE 105
Carapace (Fig 2b)mdashWith antennal spinesanterolateral margins serrated rostrum broadand dorsoventrally flattened with thin distalprojection and distolateral serrations eyesseparated from carapace
Abdomen (Fig 2b)mdashNo marked change fromstage ZI
Antennule (Fig 4a)mdashElongate weaklybilobed peduncle weakly 2-segmented 0ndash3short setae on distal margin of proximal seg-ment 2 long plumose setae and 0ndash3 short se-tae on distal end of distal segment endopodwith 1 terminal plumose seta exopodite with6ndash8 terminal aesthetascs
Antenna (Fig 4b)mdashProtopod with 2 distalspines 1 on outer margin and 1 between ramiendopod without setae scaphocerite armedwith 1 strong distolateral spine 10 or 11plumose setae on inner margin and 1plumose seta on outer margin
Mandible (Fig 4c d)mdashNo marked changefrom ZI
Maxillule (Fig 4e)mdashCoxal endite with 8ndash10marginal plumose setae basal endite with 3plumose setae 4 or 5 large dentate spines and4 or 5 smaller dentate spines inner marginwith 0 or 1 plumose seta endopod 3-seg-mented distinction between segments some-times obscure 2 2 (4 or 5) setae protopodwithout setae
Maxilla (Fig 4f )mdashCoxal endite bilobed12ndash15 setae on proximal lobe 4 or 5 setaeon distal lobe basal endite bilobed with 6ndash8setae on proximal lobe 5ndash9 setae on distallobe endopod with 5 lobes (2 or 3)+2+2+2+3setae scaphognathite with 19ndash24 setae allsetae plumose
Maxilliped 1 (Fig 4g)mdashCoxa with 4ndash8 se-tae basis with 13ndash17 marginal setae endo-pod 4-segmented (3 or 4) (2 or 3) (1 or 2)4 setae exopod 2-segmented proximal seg-ment without setae distal segment with 6 or7 setae on distal margin bilobed epipod with-out setae all setae plumose
Maxilliped 2 (Fig 4h)mdashCoxa with 0ndash2 se-tae basis with 4ndash6 setae endopod 4-seg-mented (4ndash7) 1 (2 or 3) (4ndash6) setae exo-pod unsegmented 7 or 8 setae on distal mar-gin all setae plumose
Maxilliped 3 (Fig 4i)mdashCoxa without setaebasis with 0ndash2 setae endopod 4-segmented
(l or 2) (1ndash3) (2ndash4) (3ndash6) setae exopod un-segmented 6ndash8 setae on distal margin all se-tae plumose
Pereiopods (Fig 4j k)mdashFirst and second pe-reiopod present both without setae
PleopodsmdashNot developed
Telson (Fig 4l)mdashNo change from ZI
Zoea IIIFigs 2c 5andashh 6andashh
SizemdashCL = 165 plusmn 003 mm range 160ndash172mm TL = 413 plusmn 009 mm range 399ndash436mm
Carapace (Fig 2c)mdashNo marked changefrom ZII
Abdomen (Fig 2c)mdashSomites 2ndash5 with pos-terolateral projections dorsal spine elongateon somite 2 sometimes extending beyondposterior margin of somite 3 short dorsalspines on somites 3ndash6
Antennule (Fig 5a)mdashBiramous peduncle2-segmented proximal segment with (0 or1)+(2 or 3) short setae on outer margin and1 long plumose seta on inner margin distalsegment with 2 or 3 setae on inner margin 1seta between rami and 0ndash3 short seta onouter margin endopod with 0 or 1 terminalplumose seta exopod with 6 or 7 terminaland 0ndash2 subterminal aesthetascs
Antenna (Fig 5b)mdashProtopod with 1 spineat outer distal margin endopod with 0 or 1short seta scaphocerite armed with 1 strongdistolateral spine 11ndash13 plumose setae on in-ner margin and 1 plumose seta on outer mar-gin
Mandible (Fig 5c d)mdashIncisor process with7 or 8 teeth on right mandible 6ndash8 teeth onleft mandible molar process denticulate onboth mandibles buds of palps present
Maxillule (Fig 5e)mdashCoxal endite with10ndash12 marginal plumose setae basal enditewith 3 plumose setae 4 or 5 large dentatespines and 4 or 5 smaller dentate spinesinner margin with 1 plumose seta endopod3-segmented distinction between segmentssometimes obscure 2 2 4 setae protopodwithout setation
Maxilla (Fig 5f )mdashCoxal endite bilobed14ndash17 setae on proximal lobe 4ndash6 setae ondistal lobe basal endite bilobed with 7ndash10 se-
106 JOURNAL OF CRUSTACEAN BIOLOGY VOL 20 NO 1 2000
tae on proximal lobe 6ndash9 setae on distal lobeendopod with 5 lobes 3+2+2+2+(2 or 3) se-tae scaphognathite with 21ndash26 setae all se-tae plumose
Maxilliped 1 (Fig 5g)mdashCoxa with 4ndash9 se-tae basis with 14ndash18 marginal setae endo-pod 4-segmented (3 or 4) 2 2 (3 or 4) se-tae exopod 2-segmented proximal segment
without setae distal segment with 5ndash7 setaeon distal margin bilobed epipod without se-tae all setae plumose
Maxilliped 2 (Fig 5h)mdashCoxa with 0ndash2 se-tae basis with 5 or 6 setae endopod 4-seg-mented (4ndash6) 1 3 (4ndash6) setae exopod un-segmented 7 or 8 setae on distal margin allsetae plumose
Fig 4 Callichirus islagrande ZII stage a antennule b antenna c right mandible d left mandible e maxillulef maxilla (plumose condition not shown) g maxilliped 1 h maxilliped 2 i maxilliped 3 j pereiopod 1 k pe-reiopod 2 1 telson Scales = 02 mm
STRASSER AND FELDER LARVAL DEVELOPMENT IN CALLICHIRUS ISLAGRANDE 107
Maxilliped 3 (Fig 6a)mdashCoxa with 0 or 1seta basis with 1ndash3 setae endopod 4-seg-mented (1 or 2) 2 (2ndash4) (4 or 5) setae exo-pod unsegmented 7 or 8 setae on distal mar-gin all setae plumose
Pereiopod 1 (Fig 6b)mdashChelate and bira-mous coxa without setae basis with 0 or 1seta endopod 4-segmented 0 (1 or 2) (2ndash4)2 setae exopod with 6 or 7 setae on distalmargin
Fig 5 Callichirus islagrande ZIII stage a antennule b antenna c right mandible d left mandible e maxil-lule f maxilla (plumose condition not shown) g maxilliped 1 h maxilliped 2 All setae on the maxilla were plumoseScales = 02 mm
108 JOURNAL OF CRUSTACEAN BIOLOGY VOL 20 NO 1 2000
Pereiopod 2 (Fig 6c)mdashChelate and biramouscoxa without setae basis with 0 or 1 seta en-dopod 4-segmented 0 (0ndash2) (0ndash4) 2 setaeexopod with 5ndash7 setae on distal margin
Pereiopods 3ndash5 (Fig 6d e f )mdashWithout setae
Pleopods (Figs 1c 6g)mdashBilobed buds onabdominal segments 2ndash5 first pair smallerthan others
Telson and Uropods (Fig 6h)mdashNo changein arrangement of posterior telsonal processesuropods biramous endopod without setae
Fig 6 Callichirus islagrande ZIII stage a maxilliped 3 b pereiopod 1 c pereiopod 2 d pereiopod 3 e pe-reiopod 4 f pereiopod 5 g pleopods from fourth abdominal somite h telson Scales = 02 mm
STRASSER AND FELDER LARVAL DEVELOPMENT IN CALLICHIRUS ISLAGRANDE 109
exopod with 10 or 11 marginal plumose se-tae ventral spine near base of protopod
Zoea IVFigs 2d 7andashh 8andashi
SizemdashCL = 192 plusmn 009 mm range 169ndash215mm TL = 482 plusmn 025 mm range 424ndash536mm
Carapace (Fig 2d)mdashNo marked changefrom ZIII
Abdomen (Fig 2d)mdashNo marked change fromZIII
Antennule (Fig 7a)mdashBiramous peduncle2-segmented with 1+(1ndash4) short setae onouter margin and 0 or 1 plumose seta on in-ner margin of proximal segment distal seg-ment with 3ndash5 plumose setae on inner mar-gin 1 plumose seta between rami and 0ndash3short setae on outer margin endopod with 1or 2 plumose setae exopod with 6 or 7 ter-minal and 2 subterminal aesthetascs
Antenna (Fig 7b)mdashProtopod with 1 distalspine on outer margin endopod with 0ndash2 se-tae scaphocerite armed with 1 strong disto-lateral spine 12ndash14 plumose setae on innermargin and 1 plumose seta on outer margin
Mandible (Fig 7c d)mdashIncisor process with8 teeth on right mandible 6ndash8 teeth on leftmandible molar process denticulate on bothmandibles palps present
Maxillule (Fig 7e)mdashCoxal endite with12ndash14 marginal plumose setae basal enditewith 3 plumose setae 5 or 6 large dentatespines and 5 or 6 smaller dentate spinesinner margin with 1 plumose seta endopod3-segmented distinction between segmentssometimes obscure 2 2 (3 or 4) setae pro-topod without setation
Maxilla (Fig 7f )mdashCoxal endite bilobed14ndash19 setae on proximal lobe 5 setae on dis-tal lobe basal endite bilobed 6ndash10 setae onproximal lobe 8 or 9 setae on distal lobeendopod with 5 lobes 3+(2 or 3)+2+2+3 se-tae scaphognathite with 27ndash32 setae all se-tae plumose
Maxilliped 1 (Fig 7g)mdashCoxa with 2ndash8 se-tae basis with 16ndash19 marginal setae endo-pod 4-segmented (3 or 4) 2 2 4 setae exo-
pod 2-segmented proximal segment withoutsetae distal segment with 6 or 7 setae on dis-tal margin bilobed epipod without setae allsetae plumose
Maxilliped 2 (Fig 7h)mdashCoxa with 0ndash3 se-tae basis with 4ndash6 setae endopod 4-seg-mented (5ndash8) 1 (3ndash5) (4 or 5) setae exo-pod unsegmented 7 or 8 setae on distal mar-gin all setae plumose
Maxilliped 3 (Fig 8a)mdashCoxa with 0 or 1seta basis with 2ndash4 setae endopod 4-seg-mented (l or 2) 2 (3ndash5) (3 or 4) setae exo-pod unsegmented 7 or 8 setae on distal mar-gin all setae plumose
Pereiopod 1 (Fig 8b)mdashBiramous cheliformcoxa without setae basis with 0ndash2 setae en-dopod 4-segmented (0 or 1) (1 or 2) (2ndash4)(1 or 2) setae exopod with 5ndash7 setae on dis-tal margin
Pereiopod 2 (Fig 8c)mdashBiramous cheliformcoxa without setae basis with 0 or 1 setaendopod 4-segmented 0 (1 or 2) (3ndash5) (2or 3) setae exopod with 5ndash7 setae on distalmargin
Pereiopod 3 (Fig 8d)mdashBiramous coxa andbasis without setae endopod 4-segmented0 (0ndash2) (3 or 4) 2 setae propodus blade-like exopod with 4 or 5 terminal setae
Pereiopod 4 (Fig 8e)mdashBiramous coxa andbasis without setae endopod 4-segmented (0or 1) (0ndash2) (2ndash4) 2 setae exopod with 0ndash2setae
Pereiopod 5 (Fig 8f )mdashUniramous 2 setaeon terminal segment 4 or 5 setae on adja-cent segment
Pleopods (Fig 8g h)mdashLarger than at ZIIIstill without setae
Telson and Uropods (Fig 8i)mdashTelson trian-gular narrow anteriorly no anal spine pos-terior margin with (12ndash15)+1+(13 or 14)processes arranged as (I II III IV III II I)(I) outermost process a plumodenticulate seta(II) second process ldquoanomuran hairrdquo (III) 10ndash13plumodenticulate setae and (IV) naked spineuropods biramous endopod with 7ndash11 marginalplumose setae exopod with 1 naked distolat-eral spine 12ndash14 marginal plumose setae
110 JOURNAL OF CRUSTACEAN BIOLOGY VOL 20 NO 1 2000
Decapodid (first postlarva)Figs 2e 9cndashj l0andashi
SizemdashCL = 133 plusmn 002 mm range129ndash138 mm TL = 378 plusmn 011 mm range361ndash405 mm
Carapace (Fig 2e)mdashShorter than abdomenno anterolateral serrations with pterygostomial
projections rostrum reduced without serra-tions sometimes bending down between eyes
Abdomen (Fig 2e)mdashSomites broad and flat-tened posterior projections rounded with 0ndash7setae somite 2 rectangular somites 3ndash5 oval
Antennule (Fig 9c)mdashPeduncle 3-segmentedproximal segment with 17ndash24 setae 13ndash15 of
Fig 7 Callichirus islagrande ZIV stage a antennule b antenna c right mandible d left mandible e maxil-lule f maxilla (plumose condition not shown) g maxilliped 1 h maxilliped 2 All setae on the maxilla were plumoseScales = 02 mm
STRASSER AND FELDER LARVAL DEVELOPMENT IN CALLICHIRUS ISLAGRANDE 111
these associated with statocyst intermediatesegment with 6ndash10 setae distal segment of pe-duncle with 2 vertical rows of setae one with4 or 5 setae other with 2ndash4 setae 1ndash3 addi-tional setae on outer margin and 3ndash5 short se-tae on distal margin endopod 4ndash6 segmented(1ndash3) (0 or 1) (1ndash4) (1ndash4) (3ndash6) (3ndash6) se-tae exopod 3ndash5-segmented 0 (0ndash4) (0ndash4)(1ndash4) setae +2 asthetascs (5ndash8) asthetascs ()
Antenna (Fig 9d)mdashPeduncle 2-segmentedproximal segment with 0 or 1 seta distal seg-ment with 1ndash3 setae on inner margin spinepresent in some flagellum ranging from 14ndash18
segments (0 or 1) (2ndash4) variable region(1ndash3 articles with 0ndash2 setae) (0ndash4) (0ndash2)(2ndash5) (1ndash4) (3ndash6) (0ndash3) (2ndash5) (3ndash6)(0ndash3) (3ndash6) (0 or 1) (1ndash7) (4ndash9) setae exo-pod reduced to small lobe with 0ndash5 setae
Mandible (Fig 9e f )mdashBoth mandibles sim-ilar incisor process with 5ndash8 teeth molarprocess smooth or with small ridges palp3-segmented 0 (0 or 1) (8ndash12) setae
Maxillule (Fig 9g)mdashCoxal endite with15ndash22 marginal setae and 0 or 1 submarginal
Fig 8 Callichirus islagrande ZIV stage a maxilliped 3 b pereiopod 1 c pereiopod 2 d pereiopod 3 e pe-reiopod 4 f pereiopod 5 g pleopods from first abdominal somite h pleopods from fourth abdominal somite i tel-son Scales = 02 mm
112 JOURNAL OF CRUSTACEAN BIOLOGY VOL 20 NO 1 2000
seta basal endite with 3ndash5 plumose setae14ndash19 dentate spines 0ndash2 plumose setae oninner margin endopod reduced unsegmentedwith 0ndash8 setae
Maxilla (Fig 9h)mdashCoxal endite trilobed(17ndash21)+(7ndash10)+(6ndash9) setae basal enditebilobed 8ndash10 setae on proximal lobe 8ndash15setae on distal lobe endopod reduced with
Fig 9 Callichirus islagrande ZV stage a one example of the antenna b one example of pleopods Decapodidstage c antennule d antenna e right mandible f left mandible g maxillule h maxilla (plumose condition notshown) i maxilliped 1 j maxilliped 2 Scales = 02 mm
STRASSER AND FELDER LARVAL DEVELOPMENT IN CALLICHIRUS ISLAGRANDE 113
5ndash11 setae scaphognathite with 37ndash45 se-tae all setae plumose
Maxilliped 1 (Fig 9i)mdashCoxa with 8ndash12 se-tae basis highly setose with 27ndash39 setae en-dopod reduced with 0ndash11 setae exopod un-segmented 6ndash14 setae bilobed epipod with-out setae
Maxilliped 2 (Fig 9j)mdashCoxa partially fusedto basis with 1ndash4 setae basis with 3ndash5 se-
tae endopod 4-segmented (16ndash23) (0 or 1)(6ndash11) (4ndash8) setae exopod with 0ndash8 setaeepipod present but without setae
Maxilliped 3 (Fig 10a)mdashCoxa partiallyfused to basis with 0ndash3 setae basis with 1ndash4setae endopod 5-segmented (11ndash19) (9ndash15)(7ndash11) (8ndash21) (5ndash11) setae exopod usuallyreduced and without setae occasionally with2ndash9 setae epipod present
Fig 10 Callichirus islagrande decapodid stage a maxilliped 3 b pereiopod 1 c pereiopod 2 d pereiopod 3e pereiopod 4 f pereiopod 5 g pleopods from first abdominal somite h pleopods from fourth abdominal somitei telson Scales = 02 mm
114 JOURNAL OF CRUSTACEAN BIOLOGY VOL 20 NO 1 2000
Pereiopod 1 (Fig 10b)mdashChelate biramous7-segmented coxa with 0ndash6 setae basis with2ndash5 setae ischium with 4ndash7 setae and 0ndash3spines merus with 5ndash8 setae carpus with10ndash14 setae propodus with 27ndash37 setae and3ndash4 spines on inner margin of immovable fin-ger dactylus with 21ndash33 setae exopod usu-ally reduced and without setae occasionallywith 4ndash7 setae
Pereiopod 2 (Fig 10c)mdashChelate biramous7-segmented coxa with 2ndash5 setae basis with1ndash5 setae ischium with 4ndash10 setae meruswith 15ndash20 setae carpus with 8ndash12 setaepropodus with 30ndash40 setae and 1ndash4 spineson inner margin of immovable finger dacty-lus with 21ndash30 setae exopod usually reduced0ndash7 setae
Pereiopod 3 (Fig 10d)mdashBiramous 7-seg-mented coxa with 2ndash5 setae basis with 1ndash3setae ischium with 5ndash8 setae merus with6ndash9 setae carpus with 7ndash11 setae propodusbladelike with 25ndash34 setae 1 spine on innerdistal margin dactylus with 15ndash21 setae exo-pod usually reduced 0ndash5 setae
Pereiopod 4 (Fig 10e)mdashBiramous 7-seg-mented coxa with 2ndash7 setae basis with 1ndash4setae ischium with 3ndash8 setae merus with5ndash11 setae carpus with 4ndash9 setae propoduswith 27ndash38 setae 1 spine on inner distal mar-gin dactylus with 15ndash19 setae exopod usu-ally reduced with 0ndash2 setae
Pereiopod 5 (Fig 10f )mdashCheliform unira-mous 7-segmented coxa with 2ndash6 setae ba-sis with l or 2 setae ischium with 2ndash5 setaemerus with 5ndash9 setae carpus with 4ndash7 se-tae propodus with 22ndash30 setae 1 spine onimmovable finger dactylus with 6ndash10 setaeexopod absent
Pleopods (somite 2) (Fig 10g)mdashBiramouscoxa partially fused to basis without setaebasis with 0ndash3 setae endopod with 3ndash5hooked setae on appendix interna 10ndash17 mar-ginal plumose setae 0ndash2 submarginal nakedsetae exopod with 21ndash27 marginal plumosesetae and 0ndash1 submarginal naked seta
Pleopods (somites 3ndash5) (Fig 10h)mdashBiramouscoxa without setae basis with 3ndash6 marginalplumose setae endopod with 3ndash6 hooked se-tae on appendix interna 10ndash18 marginalplumose setae and 0ndash3 naked submarginal se-tae exopod with 20ndash29 marginal plumose se-tae and 0 or 1 submarginal naked seta
Telson and Uropods (Fig 10i)mdashWith 24ndash36processes on posterior margin median pro-cess usually absent or reduced uropods bi-ramous endopod with 19ndash25 plumose setaeexopod with 35ndash58 plumose setae protopodwith 3ndash5 setae
DISCUSSION
As in other species of Callichirus (see Ro-drigues 1976 Aste and Retamal 1983Strasser and Felder in press b) larval devel-opment of C islagrande included four or fivezoeal stages before metamorphosis to the de-capodid ZI larvae may be identified by thepresence of an elongate rounded rostrumsessile eyes and a lack of pereiopodspleopods or uropods (Fig 1a) At ZII lar-vae acquired biramous buds of pereiopods 1and 2 eyes were separated from the carapaceand the rostrum became flattened and serratedlaterally (Fig 1b) Buds of pleopods on ab-dominal segments 2ndash5 pereiopods 3ndash5 anduropods were developed at ZIII (Fig 1c) Inaddition to further development of structuresmentioned thus far ZIV larvae could be dis-tinguished by presence of a distal spine on theexopod and setae on the endopod of each uro-pod (Fig 1d) When it occurred ZV typicallyresembled ZIV but varied widely in mor-phology Appendages at ZV resembled thoseof ZIV the decapodid stage or some grada-tion between the two (see Figs 9a b for ex-amples) The decapodid stage was similar toother species of Callichirus in the reductionof larval characters such as the elongate ros-trum dorsal spines on the abdomen and ex-opods of the pereiopods (Fig le) Acquisitionof fan-like pleopods marked the transition toa primarily abdominal mode of swimming atthis stage
In the Florida Atlantic population of C ma-jor both salinity and presence of putative set-tlement cues at ZIV impacted the observednumber of zoeal stages and the duration ofZIV (Strasser and Felder in preparation1999c) In C islagrande the percentage oflarvae that passed through five versus fourzoeal stages varied (9ndash89) between parentalfemales but was not shown to depend onsalinity or presence of various putative set-tlement cues (Strasser and Felder 1998 inpreparation) Larvae that molted from ZIVto ZV were usually normal morphologicallyand had mortality similar to those that molteddirectly from ZIV to D In contrast larvae of
STRASSER AND FELDER LARVAL DEVELOPMENT IN CALLICHIRUS ISLAGRANDE 115
the Florida Atlantic C major that molted toZV had a higher incidence of mortality anddeformity at D than did those that had by-passed ZV (Strasser and Felder 1999a c)
The number of zoeal stages appears to beless variable in Callichirus garthi and theGulf of Mexico population of C major thanin congeneric populations studied to dateCallichirus garthi was reported to have fivezoeal stages and a long larval duration reach-ing the decapodid stage 48 days after hatch-ing (Aste and Retamal 1983) Developmentof the Gulf population of C major wasshorter in duration (decapodid reached about10 days after hatching) and typically includedfour zoeal stages (Strasser and Felder 1999a)animals rarely passed through three or fivezoeal stages before the molt to the decapodidAlthough the number of stages is reducedfrom that observed in C garthi developmentin the Gulf population of C major cannot beclassified as abbreviated There are species inboth the Callianassinae (Callianassa slkraussi Stebbing C sl kewalramanii and Csl tyrrhena (Petagna)) and Callichirinae(Sergio mirim Lepidophthalmus sinuensisand L louisianensis) that have only two zoealstages before the molt to the decapodid(Forbes 1973 Sankolli and Shenoy 1975Rodrigues 1984 Thessalou-Legaki 1990Nates et al 1997)
Compared to other species of the genusCallichirus C islagrande is morphologicallycloser to C garthi (from Chile) than to theGulf of Mexico population of C major whichoverlaps in distribution with C islagrande Inthe zoeal phase of C islagrande and Cgarthi the telson is concave and the outer-most process on the ventral margin is articu-lated In both C major and Callichirus sp theposterior margin of the telson is straight andthe outermost process is unarticulated duringthe zoeal stages (Rodrigues 1976 Strasserand Felder 1999a) There are also differencesin spination of the abdomen In zoeae of Cislagrande and C garthi the dorsal spine ofabdominal segment 3 is not longer than thosefound on segments 4 and 5 (Aste and Reta-mal 1983) In contrast the dorsal spine of ab-dominal segment 3 is approximately twice aslong as the dorsal spines on segments 4 and5 in early zoeal stages of C major and Cal-lichirus sp (Rodrigues 1976 Aste and Re-tamal 1983 Strasser and Felder 1999a) Thegreatest difference between these groups is
the number of pleopod pairs at zoeal and de-capodid stages Callichirus islagrande and Cgarthi have pleopods on abdominal segments2ndash5 while pleopods are present only on ab-dominal segments 3ndash5 in C major and Cal-lichirus sp (Rodrigues 1976 Aste and Re-tamal 1983 Strasser and Felder 1999a) Al-though Callichirus islagrande and C garthiare similar morphologically there are differ-ences between these two species Callichirusislagrande as well as C major and Cal-lichirus sp develops pleopod buds andchelate pereiopods at ZIII whereas C garthidoes not acquire these characters until ZIV
There are few callianassid species forwhich the larval development has been de-scribed (Sankolli and Shenoy 1975 Ro-drigues 1976 1984 Aste and Retamal 19831984 Konishi et al 1990 Nates et al 1997Strasser and Felder 1999a) all of which aremembers of the Callichirinae and Cal-lianassinae Among species there is a greatdeal of overlap in larval characters makingit difficult to draw distinctions between thesesubfamilies At present information on lar-val development of species in the Callichiri-nae is limited to larval descriptions of Cal-lichirus major C garthi Callichirus sp Cislagrande Sergio mirim Lepidophthalmuslouisianensis and L sinuensis (see Ro-drigues 1976 1984 Aste and Retamal 1983Nates et al 1997 Strasser and Felder1999a) As mentioned above species of Cal-lichirus have been shown to have four to fivezoeal stages while the other members exhibitabbreviated development that consists of onlytwo zoeal stages The most obvious differencebetween larvae of the species of Callichirinaewith abbreviated development is the numberof pleopod pairs Larvae of S mirim havepleopods on abdominal segments 3ndash5whereas L louisianensis and L sinuensishave pleopods on segments 2ndash5 (Rodrigues1984 Nates et al 1997)
Ghost shrimps of the genus Callichirus maybe distinguished from other genera of the Cal-lianassidae by the cuticular pattern on the dor-sal surface of the abdomen and by the strap-like shape of the uropodal endopods (Manningand Felder 1991) Whereas these charactersmay be used to distinguish adults of the genusCallichirus they are not present at zoeal anddecapodid stages Larvae may be distin-guished from all other callianassid genera byhaving four to five zoeal stages and numer-
116 JOURNAL OF CRUSTACEAN BIOLOGY VOL 20 NO 1 2000
ous processes (12ndash25+1+12ndash25) on the pos-terior margin of the telson during the earlyzoeal stages Larvae of Callichirus may alsodiffer from other callianassids with extendeddevelopment by the retention of the anomu-ran hair as the second process on the poste-rior margin of the telson in the late zoealstages Whereas this process is converted toa spine in Neotrypaea uncinata at ZV and Cal-lianassa sl petalura at ZIV the anomuranhair is retained at ZIV in Callichirus majorand C islagrande (see Rodrigues 1976Strasser and Felder 1999a) Although notmentioned specifically in the descriptions theanomuran hair appears to be present in illus-trations of Callichirus sp at ZIII and Cgarthii at ZV (Rodrigues 1976 fig 35 Asteand Retamal 1983 fig 8Q) A bilobed epi-pod on the first maxilliped of ZI is consistentlyfound in Callichirus but is also present at ZIin Lepidophthalmus sinuensis (see Nates etal 1997) Other callianassids acquire thischaracter at ZII (L louisianensis Nates et al1997 Callianassa sl kewlramanii Sankolliand Shenoy 1975) at ZV (Neotrypaea unci-nata Aste and Retamal 1984) or at the de-capodid stage (Sergio mirim Rodrigues 1984C sl petalura Konishi et al 1990) Thusthe number of zoeal stages and setation of thetelson may serve as the best characters to dis-tinguish larvae of Callichirus from other gen-era However since larval descriptions arelacking for many callianassid species it is un-known if these characters (in combination)will prove to be diagnostic until descriptionsfor other species are published
ACKNOWLEDGEMENTS
We thank Robert Bourgeois Barbara Hasek JimMaKinster Sergio Nates and Kurt Strasser who pro-vided assistance in the field and laboratory and MaryE Rice Director of the Smithsonian Marine StationLink Port Florida who facilitated access to station fa-cilities We also thank R Bauer R Jaeger R Lemaitreand G Watson for their early comments on this manu-script and Raymond B Manning who facilitated fund-ing and gave advice on collecting sites Direct supportfor this study was provided to Karen M Strasser by adoctoral fellowship from the University of Louisianaa graduate student research grant from the LUMCON(Louisiana Universities Marine Consortium) Founda-tion and a summer research award from the Smith-sonian Marine Station Additional support for field andlab activities was provided to D L Felder under Loui-siana Sea Grant College Program Grant RCFB-21 USFish and Wildlife Service Cooperative Agreement 14-16-0009-89-96 (Task Order No 6) US Department of En-ergy grant DE-FG02-97ER12220 and several small proj-ect grants from the Smithsonian Marine Station Views
expressed herein are those of the authors and do not nec-essarily reflect views of supporting government agenciesor subagencies mentioned above This is contributionNumber 466 of the Smithsonian Marine Station and con-tribution Number 67 of the ULL Laboratory for Crus-tacean Research All experiments conducted in this studycomply with current applicable state and federal laws
LITERATURE CITED
Aste A and M Retamal 1983 Desarrollo larval deCallianassa garthi Retamal 1975 bajo condiciones delaboratoriomdashCiencia y Tecnologiacutea del Mar CONA 75ndash26
mdashmdashmdash and mdashmdashmdash 1984 Desarrollo larval de Cal-lianassa uncinata H Milne Edwards1837 (DecapodaCallianassidae) bajo condiciones de laboratoriomdashGayana Zooloacutegica 48 41ndash56
Felder D L 1978 Osmotic and ionic regulation in sev-eral western Atlantic Callianassidae (Crustacea De-capoda Thalassinidea)mdashBiological Bulletin 154409ndash429
mdashmdashmdash and R B Griffis 1994 Dominant infaunalcommunities at risk in shoreline habitats burrowingthalassinid CrustaceamdashOCS Study number MMS94ndash0007 United States Department of the InteriorMinerals Management Service Gulf of Mexico OCSRegional Office New Orleans Louisiana Pp 1ndash87
Forbes A T 1973 An unusual abbreviated larval lifehistory in the estuarine burrowing prawn Callianassakraussi (Crustacea Decapoda Thalassinidea)mdashMa-rine Biology 22 361ndash365
Konishi K 1989 Larval development of the mudshrimp Upogebia (Upogebia) major (de Haan) (Crus-tacea Thalassinidea Upogebiidae) under laboratoryconditions with comments on larval characters of tha-lassinid familiesmdashBulletin of the National ResearchInstitute of Aquaculture 15 1ndash17
mdashmdashmdash R Quintana and Y Fukuda 1990 A completedescription of larval stages of the ghost shrimp Cal-lianassa petalura Stimpson (Crustacea ThalassinideaCallianassidae) under laboratory conditionsmdashBulletinof the National Research Institute of Aquaculture 1727ndash49
Manning R B and D L Felder 1986 The status ofthe callianassid genus Callichirus Stimpson 1866(Crustacea Decapoda Thalassinidea)mdashProceedingsof the Biological Society of Washington 99 437ndash443
mdashmdashmdash and mdashmdashmdash 1991 Revision of the AmericanCallianassidae (Crustacea Decapoda Thalassinidea)mdashProceedings of the Biological Society of Washington104 764ndash794
Nates S F D L Felder and R Lemaitre 1997 Com-parative larval development in two species of the bur-rowing ghost shrimp genus Lepidophthalmus (Decap-oda Callianassidae)mdashJournal of Crustacean Biology17 497ndash519
Rodrigues S de A 1976 Sobre a reproduccedilatildeo embri-ologia e desenvolvimento larval de Callichirus majorSay 1818 (Crustacea Decapoda Thalassinidea)mdashBo-letim de Zoologia Universidade de Satildeo Paulo 185ndash104
mdashmdashmdash 1984 Desenvolvimento poacutes-embrionaacuterio deCallichirus mirim (Rodrigues 1971) obtido emcondiccedilotildees artificiais (Crustacea Decapoda Tha-lassinidea)mdashBoletim de Zoologia Universidade deSatildeo Paulo 8 239ndash256
STRASSER AND FELDER LARVAL DEVELOPMENT IN CALLICHIRUS ISLAGRANDE 117
Sankolli K N and S Shenoy 1975 Larval develop-ment of mud shrimp Callianassa (Callichirus) kewal-ramanii Sankolli in the laboratory (Crustacea Decap-oda)mdashBulletin of the Department of Marine ScienceUniversity of Cochin 4 704ndash720
Staton J L and D L Felder 1995 Genetic variationin populations of the ghost shrimp genus Callichirus(Crustacea Decapoda Thalassinoidea) in the westernAtlantic and Gulf of MexicomdashBulletin of Marine Sci-ence 56 523ndash536
Strasser K M and D L Felder 1998 Settlement cuesin successive developmental stages of the ghost shrimpCallichirus major and C islagrande (Crustacea De-capoda Thalassinidea)mdashMarine Biology
mdashmdashmdash and mdashmdashmdash 1999a Larval development of twopopulations of the ghost shrimp Callichirus major (De-capoda Thalassinidea)mdashJournal of Crustacean Biology
mdashmdashmdash and mdashmdashmdash 1999b Sand as a stimulus for set-tlement in the ghost shrimp Callichirus major (Say)and C islagrande (Schmitt) (Crustacea DecapodaThalassinidea)mdashJournal of Experimental Marine Bi-ology and Ecology 239 211ndash222
mdashmdashmdash and mdashmdashmdash 1999c Settlement cues in theFlorida Atlantic population of the ghost shrimp Cal-lichirus major (Crustacea Decapoda Thalassinidea)mdashMarine Ecology Progress Series
Thessalou-Legaki M 1990 Advanced larval develop-ment of Callianassa tyrrhena (Decapoda Tha-lassinidea) and the effect of environmental factorsmdashJournal of Crustacean Biology 10 659ndash666
RECEIVED 19 September 1998ACCEPTED 3 December 1998

STRASSER AND FELDER LARVAL DEVELOPMENT IN CALLICHIRUS ISLAGRANDE 105
Carapace (Fig 2b)mdashWith antennal spinesanterolateral margins serrated rostrum broadand dorsoventrally flattened with thin distalprojection and distolateral serrations eyesseparated from carapace
Abdomen (Fig 2b)mdashNo marked change fromstage ZI
Antennule (Fig 4a)mdashElongate weaklybilobed peduncle weakly 2-segmented 0ndash3short setae on distal margin of proximal seg-ment 2 long plumose setae and 0ndash3 short se-tae on distal end of distal segment endopodwith 1 terminal plumose seta exopodite with6ndash8 terminal aesthetascs
Antenna (Fig 4b)mdashProtopod with 2 distalspines 1 on outer margin and 1 between ramiendopod without setae scaphocerite armedwith 1 strong distolateral spine 10 or 11plumose setae on inner margin and 1plumose seta on outer margin
Mandible (Fig 4c d)mdashNo marked changefrom ZI
Maxillule (Fig 4e)mdashCoxal endite with 8ndash10marginal plumose setae basal endite with 3plumose setae 4 or 5 large dentate spines and4 or 5 smaller dentate spines inner marginwith 0 or 1 plumose seta endopod 3-seg-mented distinction between segments some-times obscure 2 2 (4 or 5) setae protopodwithout setae
Maxilla (Fig 4f )mdashCoxal endite bilobed12ndash15 setae on proximal lobe 4 or 5 setaeon distal lobe basal endite bilobed with 6ndash8setae on proximal lobe 5ndash9 setae on distallobe endopod with 5 lobes (2 or 3)+2+2+2+3setae scaphognathite with 19ndash24 setae allsetae plumose
Maxilliped 1 (Fig 4g)mdashCoxa with 4ndash8 se-tae basis with 13ndash17 marginal setae endo-pod 4-segmented (3 or 4) (2 or 3) (1 or 2)4 setae exopod 2-segmented proximal seg-ment without setae distal segment with 6 or7 setae on distal margin bilobed epipod with-out setae all setae plumose
Maxilliped 2 (Fig 4h)mdashCoxa with 0ndash2 se-tae basis with 4ndash6 setae endopod 4-seg-mented (4ndash7) 1 (2 or 3) (4ndash6) setae exo-pod unsegmented 7 or 8 setae on distal mar-gin all setae plumose
Maxilliped 3 (Fig 4i)mdashCoxa without setaebasis with 0ndash2 setae endopod 4-segmented
(l or 2) (1ndash3) (2ndash4) (3ndash6) setae exopod un-segmented 6ndash8 setae on distal margin all se-tae plumose
Pereiopods (Fig 4j k)mdashFirst and second pe-reiopod present both without setae
PleopodsmdashNot developed
Telson (Fig 4l)mdashNo change from ZI
Zoea IIIFigs 2c 5andashh 6andashh
SizemdashCL = 165 plusmn 003 mm range 160ndash172mm TL = 413 plusmn 009 mm range 399ndash436mm
Carapace (Fig 2c)mdashNo marked changefrom ZII
Abdomen (Fig 2c)mdashSomites 2ndash5 with pos-terolateral projections dorsal spine elongateon somite 2 sometimes extending beyondposterior margin of somite 3 short dorsalspines on somites 3ndash6
Antennule (Fig 5a)mdashBiramous peduncle2-segmented proximal segment with (0 or1)+(2 or 3) short setae on outer margin and1 long plumose seta on inner margin distalsegment with 2 or 3 setae on inner margin 1seta between rami and 0ndash3 short seta onouter margin endopod with 0 or 1 terminalplumose seta exopod with 6 or 7 terminaland 0ndash2 subterminal aesthetascs
Antenna (Fig 5b)mdashProtopod with 1 spineat outer distal margin endopod with 0 or 1short seta scaphocerite armed with 1 strongdistolateral spine 11ndash13 plumose setae on in-ner margin and 1 plumose seta on outer mar-gin
Mandible (Fig 5c d)mdashIncisor process with7 or 8 teeth on right mandible 6ndash8 teeth onleft mandible molar process denticulate onboth mandibles buds of palps present
Maxillule (Fig 5e)mdashCoxal endite with10ndash12 marginal plumose setae basal enditewith 3 plumose setae 4 or 5 large dentatespines and 4 or 5 smaller dentate spinesinner margin with 1 plumose seta endopod3-segmented distinction between segmentssometimes obscure 2 2 4 setae protopodwithout setation
Maxilla (Fig 5f )mdashCoxal endite bilobed14ndash17 setae on proximal lobe 4ndash6 setae ondistal lobe basal endite bilobed with 7ndash10 se-
106 JOURNAL OF CRUSTACEAN BIOLOGY VOL 20 NO 1 2000
tae on proximal lobe 6ndash9 setae on distal lobeendopod with 5 lobes 3+2+2+2+(2 or 3) se-tae scaphognathite with 21ndash26 setae all se-tae plumose
Maxilliped 1 (Fig 5g)mdashCoxa with 4ndash9 se-tae basis with 14ndash18 marginal setae endo-pod 4-segmented (3 or 4) 2 2 (3 or 4) se-tae exopod 2-segmented proximal segment
without setae distal segment with 5ndash7 setaeon distal margin bilobed epipod without se-tae all setae plumose
Maxilliped 2 (Fig 5h)mdashCoxa with 0ndash2 se-tae basis with 5 or 6 setae endopod 4-seg-mented (4ndash6) 1 3 (4ndash6) setae exopod un-segmented 7 or 8 setae on distal margin allsetae plumose
Fig 4 Callichirus islagrande ZII stage a antennule b antenna c right mandible d left mandible e maxillulef maxilla (plumose condition not shown) g maxilliped 1 h maxilliped 2 i maxilliped 3 j pereiopod 1 k pe-reiopod 2 1 telson Scales = 02 mm
STRASSER AND FELDER LARVAL DEVELOPMENT IN CALLICHIRUS ISLAGRANDE 107
Maxilliped 3 (Fig 6a)mdashCoxa with 0 or 1seta basis with 1ndash3 setae endopod 4-seg-mented (1 or 2) 2 (2ndash4) (4 or 5) setae exo-pod unsegmented 7 or 8 setae on distal mar-gin all setae plumose
Pereiopod 1 (Fig 6b)mdashChelate and bira-mous coxa without setae basis with 0 or 1seta endopod 4-segmented 0 (1 or 2) (2ndash4)2 setae exopod with 6 or 7 setae on distalmargin
Fig 5 Callichirus islagrande ZIII stage a antennule b antenna c right mandible d left mandible e maxil-lule f maxilla (plumose condition not shown) g maxilliped 1 h maxilliped 2 All setae on the maxilla were plumoseScales = 02 mm
108 JOURNAL OF CRUSTACEAN BIOLOGY VOL 20 NO 1 2000
Pereiopod 2 (Fig 6c)mdashChelate and biramouscoxa without setae basis with 0 or 1 seta en-dopod 4-segmented 0 (0ndash2) (0ndash4) 2 setaeexopod with 5ndash7 setae on distal margin
Pereiopods 3ndash5 (Fig 6d e f )mdashWithout setae
Pleopods (Figs 1c 6g)mdashBilobed buds onabdominal segments 2ndash5 first pair smallerthan others
Telson and Uropods (Fig 6h)mdashNo changein arrangement of posterior telsonal processesuropods biramous endopod without setae
Fig 6 Callichirus islagrande ZIII stage a maxilliped 3 b pereiopod 1 c pereiopod 2 d pereiopod 3 e pe-reiopod 4 f pereiopod 5 g pleopods from fourth abdominal somite h telson Scales = 02 mm
STRASSER AND FELDER LARVAL DEVELOPMENT IN CALLICHIRUS ISLAGRANDE 109
exopod with 10 or 11 marginal plumose se-tae ventral spine near base of protopod
Zoea IVFigs 2d 7andashh 8andashi
SizemdashCL = 192 plusmn 009 mm range 169ndash215mm TL = 482 plusmn 025 mm range 424ndash536mm
Carapace (Fig 2d)mdashNo marked changefrom ZIII
Abdomen (Fig 2d)mdashNo marked change fromZIII
Antennule (Fig 7a)mdashBiramous peduncle2-segmented with 1+(1ndash4) short setae onouter margin and 0 or 1 plumose seta on in-ner margin of proximal segment distal seg-ment with 3ndash5 plumose setae on inner mar-gin 1 plumose seta between rami and 0ndash3short setae on outer margin endopod with 1or 2 plumose setae exopod with 6 or 7 ter-minal and 2 subterminal aesthetascs
Antenna (Fig 7b)mdashProtopod with 1 distalspine on outer margin endopod with 0ndash2 se-tae scaphocerite armed with 1 strong disto-lateral spine 12ndash14 plumose setae on innermargin and 1 plumose seta on outer margin
Mandible (Fig 7c d)mdashIncisor process with8 teeth on right mandible 6ndash8 teeth on leftmandible molar process denticulate on bothmandibles palps present
Maxillule (Fig 7e)mdashCoxal endite with12ndash14 marginal plumose setae basal enditewith 3 plumose setae 5 or 6 large dentatespines and 5 or 6 smaller dentate spinesinner margin with 1 plumose seta endopod3-segmented distinction between segmentssometimes obscure 2 2 (3 or 4) setae pro-topod without setation
Maxilla (Fig 7f )mdashCoxal endite bilobed14ndash19 setae on proximal lobe 5 setae on dis-tal lobe basal endite bilobed 6ndash10 setae onproximal lobe 8 or 9 setae on distal lobeendopod with 5 lobes 3+(2 or 3)+2+2+3 se-tae scaphognathite with 27ndash32 setae all se-tae plumose
Maxilliped 1 (Fig 7g)mdashCoxa with 2ndash8 se-tae basis with 16ndash19 marginal setae endo-pod 4-segmented (3 or 4) 2 2 4 setae exo-
pod 2-segmented proximal segment withoutsetae distal segment with 6 or 7 setae on dis-tal margin bilobed epipod without setae allsetae plumose
Maxilliped 2 (Fig 7h)mdashCoxa with 0ndash3 se-tae basis with 4ndash6 setae endopod 4-seg-mented (5ndash8) 1 (3ndash5) (4 or 5) setae exo-pod unsegmented 7 or 8 setae on distal mar-gin all setae plumose
Maxilliped 3 (Fig 8a)mdashCoxa with 0 or 1seta basis with 2ndash4 setae endopod 4-seg-mented (l or 2) 2 (3ndash5) (3 or 4) setae exo-pod unsegmented 7 or 8 setae on distal mar-gin all setae plumose
Pereiopod 1 (Fig 8b)mdashBiramous cheliformcoxa without setae basis with 0ndash2 setae en-dopod 4-segmented (0 or 1) (1 or 2) (2ndash4)(1 or 2) setae exopod with 5ndash7 setae on dis-tal margin
Pereiopod 2 (Fig 8c)mdashBiramous cheliformcoxa without setae basis with 0 or 1 setaendopod 4-segmented 0 (1 or 2) (3ndash5) (2or 3) setae exopod with 5ndash7 setae on distalmargin
Pereiopod 3 (Fig 8d)mdashBiramous coxa andbasis without setae endopod 4-segmented0 (0ndash2) (3 or 4) 2 setae propodus blade-like exopod with 4 or 5 terminal setae
Pereiopod 4 (Fig 8e)mdashBiramous coxa andbasis without setae endopod 4-segmented (0or 1) (0ndash2) (2ndash4) 2 setae exopod with 0ndash2setae
Pereiopod 5 (Fig 8f )mdashUniramous 2 setaeon terminal segment 4 or 5 setae on adja-cent segment
Pleopods (Fig 8g h)mdashLarger than at ZIIIstill without setae
Telson and Uropods (Fig 8i)mdashTelson trian-gular narrow anteriorly no anal spine pos-terior margin with (12ndash15)+1+(13 or 14)processes arranged as (I II III IV III II I)(I) outermost process a plumodenticulate seta(II) second process ldquoanomuran hairrdquo (III) 10ndash13plumodenticulate setae and (IV) naked spineuropods biramous endopod with 7ndash11 marginalplumose setae exopod with 1 naked distolat-eral spine 12ndash14 marginal plumose setae
110 JOURNAL OF CRUSTACEAN BIOLOGY VOL 20 NO 1 2000
Decapodid (first postlarva)Figs 2e 9cndashj l0andashi
SizemdashCL = 133 plusmn 002 mm range129ndash138 mm TL = 378 plusmn 011 mm range361ndash405 mm
Carapace (Fig 2e)mdashShorter than abdomenno anterolateral serrations with pterygostomial
projections rostrum reduced without serra-tions sometimes bending down between eyes
Abdomen (Fig 2e)mdashSomites broad and flat-tened posterior projections rounded with 0ndash7setae somite 2 rectangular somites 3ndash5 oval
Antennule (Fig 9c)mdashPeduncle 3-segmentedproximal segment with 17ndash24 setae 13ndash15 of
Fig 7 Callichirus islagrande ZIV stage a antennule b antenna c right mandible d left mandible e maxil-lule f maxilla (plumose condition not shown) g maxilliped 1 h maxilliped 2 All setae on the maxilla were plumoseScales = 02 mm
STRASSER AND FELDER LARVAL DEVELOPMENT IN CALLICHIRUS ISLAGRANDE 111
these associated with statocyst intermediatesegment with 6ndash10 setae distal segment of pe-duncle with 2 vertical rows of setae one with4 or 5 setae other with 2ndash4 setae 1ndash3 addi-tional setae on outer margin and 3ndash5 short se-tae on distal margin endopod 4ndash6 segmented(1ndash3) (0 or 1) (1ndash4) (1ndash4) (3ndash6) (3ndash6) se-tae exopod 3ndash5-segmented 0 (0ndash4) (0ndash4)(1ndash4) setae +2 asthetascs (5ndash8) asthetascs ()
Antenna (Fig 9d)mdashPeduncle 2-segmentedproximal segment with 0 or 1 seta distal seg-ment with 1ndash3 setae on inner margin spinepresent in some flagellum ranging from 14ndash18
segments (0 or 1) (2ndash4) variable region(1ndash3 articles with 0ndash2 setae) (0ndash4) (0ndash2)(2ndash5) (1ndash4) (3ndash6) (0ndash3) (2ndash5) (3ndash6)(0ndash3) (3ndash6) (0 or 1) (1ndash7) (4ndash9) setae exo-pod reduced to small lobe with 0ndash5 setae
Mandible (Fig 9e f )mdashBoth mandibles sim-ilar incisor process with 5ndash8 teeth molarprocess smooth or with small ridges palp3-segmented 0 (0 or 1) (8ndash12) setae
Maxillule (Fig 9g)mdashCoxal endite with15ndash22 marginal setae and 0 or 1 submarginal
Fig 8 Callichirus islagrande ZIV stage a maxilliped 3 b pereiopod 1 c pereiopod 2 d pereiopod 3 e pe-reiopod 4 f pereiopod 5 g pleopods from first abdominal somite h pleopods from fourth abdominal somite i tel-son Scales = 02 mm
112 JOURNAL OF CRUSTACEAN BIOLOGY VOL 20 NO 1 2000
seta basal endite with 3ndash5 plumose setae14ndash19 dentate spines 0ndash2 plumose setae oninner margin endopod reduced unsegmentedwith 0ndash8 setae
Maxilla (Fig 9h)mdashCoxal endite trilobed(17ndash21)+(7ndash10)+(6ndash9) setae basal enditebilobed 8ndash10 setae on proximal lobe 8ndash15setae on distal lobe endopod reduced with
Fig 9 Callichirus islagrande ZV stage a one example of the antenna b one example of pleopods Decapodidstage c antennule d antenna e right mandible f left mandible g maxillule h maxilla (plumose condition notshown) i maxilliped 1 j maxilliped 2 Scales = 02 mm
STRASSER AND FELDER LARVAL DEVELOPMENT IN CALLICHIRUS ISLAGRANDE 113
5ndash11 setae scaphognathite with 37ndash45 se-tae all setae plumose
Maxilliped 1 (Fig 9i)mdashCoxa with 8ndash12 se-tae basis highly setose with 27ndash39 setae en-dopod reduced with 0ndash11 setae exopod un-segmented 6ndash14 setae bilobed epipod with-out setae
Maxilliped 2 (Fig 9j)mdashCoxa partially fusedto basis with 1ndash4 setae basis with 3ndash5 se-
tae endopod 4-segmented (16ndash23) (0 or 1)(6ndash11) (4ndash8) setae exopod with 0ndash8 setaeepipod present but without setae
Maxilliped 3 (Fig 10a)mdashCoxa partiallyfused to basis with 0ndash3 setae basis with 1ndash4setae endopod 5-segmented (11ndash19) (9ndash15)(7ndash11) (8ndash21) (5ndash11) setae exopod usuallyreduced and without setae occasionally with2ndash9 setae epipod present
Fig 10 Callichirus islagrande decapodid stage a maxilliped 3 b pereiopod 1 c pereiopod 2 d pereiopod 3e pereiopod 4 f pereiopod 5 g pleopods from first abdominal somite h pleopods from fourth abdominal somitei telson Scales = 02 mm
114 JOURNAL OF CRUSTACEAN BIOLOGY VOL 20 NO 1 2000
Pereiopod 1 (Fig 10b)mdashChelate biramous7-segmented coxa with 0ndash6 setae basis with2ndash5 setae ischium with 4ndash7 setae and 0ndash3spines merus with 5ndash8 setae carpus with10ndash14 setae propodus with 27ndash37 setae and3ndash4 spines on inner margin of immovable fin-ger dactylus with 21ndash33 setae exopod usu-ally reduced and without setae occasionallywith 4ndash7 setae
Pereiopod 2 (Fig 10c)mdashChelate biramous7-segmented coxa with 2ndash5 setae basis with1ndash5 setae ischium with 4ndash10 setae meruswith 15ndash20 setae carpus with 8ndash12 setaepropodus with 30ndash40 setae and 1ndash4 spineson inner margin of immovable finger dacty-lus with 21ndash30 setae exopod usually reduced0ndash7 setae
Pereiopod 3 (Fig 10d)mdashBiramous 7-seg-mented coxa with 2ndash5 setae basis with 1ndash3setae ischium with 5ndash8 setae merus with6ndash9 setae carpus with 7ndash11 setae propodusbladelike with 25ndash34 setae 1 spine on innerdistal margin dactylus with 15ndash21 setae exo-pod usually reduced 0ndash5 setae
Pereiopod 4 (Fig 10e)mdashBiramous 7-seg-mented coxa with 2ndash7 setae basis with 1ndash4setae ischium with 3ndash8 setae merus with5ndash11 setae carpus with 4ndash9 setae propoduswith 27ndash38 setae 1 spine on inner distal mar-gin dactylus with 15ndash19 setae exopod usu-ally reduced with 0ndash2 setae
Pereiopod 5 (Fig 10f )mdashCheliform unira-mous 7-segmented coxa with 2ndash6 setae ba-sis with l or 2 setae ischium with 2ndash5 setaemerus with 5ndash9 setae carpus with 4ndash7 se-tae propodus with 22ndash30 setae 1 spine onimmovable finger dactylus with 6ndash10 setaeexopod absent
Pleopods (somite 2) (Fig 10g)mdashBiramouscoxa partially fused to basis without setaebasis with 0ndash3 setae endopod with 3ndash5hooked setae on appendix interna 10ndash17 mar-ginal plumose setae 0ndash2 submarginal nakedsetae exopod with 21ndash27 marginal plumosesetae and 0ndash1 submarginal naked seta
Pleopods (somites 3ndash5) (Fig 10h)mdashBiramouscoxa without setae basis with 3ndash6 marginalplumose setae endopod with 3ndash6 hooked se-tae on appendix interna 10ndash18 marginalplumose setae and 0ndash3 naked submarginal se-tae exopod with 20ndash29 marginal plumose se-tae and 0 or 1 submarginal naked seta
Telson and Uropods (Fig 10i)mdashWith 24ndash36processes on posterior margin median pro-cess usually absent or reduced uropods bi-ramous endopod with 19ndash25 plumose setaeexopod with 35ndash58 plumose setae protopodwith 3ndash5 setae
DISCUSSION
As in other species of Callichirus (see Ro-drigues 1976 Aste and Retamal 1983Strasser and Felder in press b) larval devel-opment of C islagrande included four or fivezoeal stages before metamorphosis to the de-capodid ZI larvae may be identified by thepresence of an elongate rounded rostrumsessile eyes and a lack of pereiopodspleopods or uropods (Fig 1a) At ZII lar-vae acquired biramous buds of pereiopods 1and 2 eyes were separated from the carapaceand the rostrum became flattened and serratedlaterally (Fig 1b) Buds of pleopods on ab-dominal segments 2ndash5 pereiopods 3ndash5 anduropods were developed at ZIII (Fig 1c) Inaddition to further development of structuresmentioned thus far ZIV larvae could be dis-tinguished by presence of a distal spine on theexopod and setae on the endopod of each uro-pod (Fig 1d) When it occurred ZV typicallyresembled ZIV but varied widely in mor-phology Appendages at ZV resembled thoseof ZIV the decapodid stage or some grada-tion between the two (see Figs 9a b for ex-amples) The decapodid stage was similar toother species of Callichirus in the reductionof larval characters such as the elongate ros-trum dorsal spines on the abdomen and ex-opods of the pereiopods (Fig le) Acquisitionof fan-like pleopods marked the transition toa primarily abdominal mode of swimming atthis stage
In the Florida Atlantic population of C ma-jor both salinity and presence of putative set-tlement cues at ZIV impacted the observednumber of zoeal stages and the duration ofZIV (Strasser and Felder in preparation1999c) In C islagrande the percentage oflarvae that passed through five versus fourzoeal stages varied (9ndash89) between parentalfemales but was not shown to depend onsalinity or presence of various putative set-tlement cues (Strasser and Felder 1998 inpreparation) Larvae that molted from ZIVto ZV were usually normal morphologicallyand had mortality similar to those that molteddirectly from ZIV to D In contrast larvae of
STRASSER AND FELDER LARVAL DEVELOPMENT IN CALLICHIRUS ISLAGRANDE 115
the Florida Atlantic C major that molted toZV had a higher incidence of mortality anddeformity at D than did those that had by-passed ZV (Strasser and Felder 1999a c)
The number of zoeal stages appears to beless variable in Callichirus garthi and theGulf of Mexico population of C major thanin congeneric populations studied to dateCallichirus garthi was reported to have fivezoeal stages and a long larval duration reach-ing the decapodid stage 48 days after hatch-ing (Aste and Retamal 1983) Developmentof the Gulf population of C major wasshorter in duration (decapodid reached about10 days after hatching) and typically includedfour zoeal stages (Strasser and Felder 1999a)animals rarely passed through three or fivezoeal stages before the molt to the decapodidAlthough the number of stages is reducedfrom that observed in C garthi developmentin the Gulf population of C major cannot beclassified as abbreviated There are species inboth the Callianassinae (Callianassa slkraussi Stebbing C sl kewalramanii and Csl tyrrhena (Petagna)) and Callichirinae(Sergio mirim Lepidophthalmus sinuensisand L louisianensis) that have only two zoealstages before the molt to the decapodid(Forbes 1973 Sankolli and Shenoy 1975Rodrigues 1984 Thessalou-Legaki 1990Nates et al 1997)
Compared to other species of the genusCallichirus C islagrande is morphologicallycloser to C garthi (from Chile) than to theGulf of Mexico population of C major whichoverlaps in distribution with C islagrande Inthe zoeal phase of C islagrande and Cgarthi the telson is concave and the outer-most process on the ventral margin is articu-lated In both C major and Callichirus sp theposterior margin of the telson is straight andthe outermost process is unarticulated duringthe zoeal stages (Rodrigues 1976 Strasserand Felder 1999a) There are also differencesin spination of the abdomen In zoeae of Cislagrande and C garthi the dorsal spine ofabdominal segment 3 is not longer than thosefound on segments 4 and 5 (Aste and Reta-mal 1983) In contrast the dorsal spine of ab-dominal segment 3 is approximately twice aslong as the dorsal spines on segments 4 and5 in early zoeal stages of C major and Cal-lichirus sp (Rodrigues 1976 Aste and Re-tamal 1983 Strasser and Felder 1999a) Thegreatest difference between these groups is
the number of pleopod pairs at zoeal and de-capodid stages Callichirus islagrande and Cgarthi have pleopods on abdominal segments2ndash5 while pleopods are present only on ab-dominal segments 3ndash5 in C major and Cal-lichirus sp (Rodrigues 1976 Aste and Re-tamal 1983 Strasser and Felder 1999a) Al-though Callichirus islagrande and C garthiare similar morphologically there are differ-ences between these two species Callichirusislagrande as well as C major and Cal-lichirus sp develops pleopod buds andchelate pereiopods at ZIII whereas C garthidoes not acquire these characters until ZIV
There are few callianassid species forwhich the larval development has been de-scribed (Sankolli and Shenoy 1975 Ro-drigues 1976 1984 Aste and Retamal 19831984 Konishi et al 1990 Nates et al 1997Strasser and Felder 1999a) all of which aremembers of the Callichirinae and Cal-lianassinae Among species there is a greatdeal of overlap in larval characters makingit difficult to draw distinctions between thesesubfamilies At present information on lar-val development of species in the Callichiri-nae is limited to larval descriptions of Cal-lichirus major C garthi Callichirus sp Cislagrande Sergio mirim Lepidophthalmuslouisianensis and L sinuensis (see Ro-drigues 1976 1984 Aste and Retamal 1983Nates et al 1997 Strasser and Felder1999a) As mentioned above species of Cal-lichirus have been shown to have four to fivezoeal stages while the other members exhibitabbreviated development that consists of onlytwo zoeal stages The most obvious differencebetween larvae of the species of Callichirinaewith abbreviated development is the numberof pleopod pairs Larvae of S mirim havepleopods on abdominal segments 3ndash5whereas L louisianensis and L sinuensishave pleopods on segments 2ndash5 (Rodrigues1984 Nates et al 1997)
Ghost shrimps of the genus Callichirus maybe distinguished from other genera of the Cal-lianassidae by the cuticular pattern on the dor-sal surface of the abdomen and by the strap-like shape of the uropodal endopods (Manningand Felder 1991) Whereas these charactersmay be used to distinguish adults of the genusCallichirus they are not present at zoeal anddecapodid stages Larvae may be distin-guished from all other callianassid genera byhaving four to five zoeal stages and numer-
116 JOURNAL OF CRUSTACEAN BIOLOGY VOL 20 NO 1 2000
ous processes (12ndash25+1+12ndash25) on the pos-terior margin of the telson during the earlyzoeal stages Larvae of Callichirus may alsodiffer from other callianassids with extendeddevelopment by the retention of the anomu-ran hair as the second process on the poste-rior margin of the telson in the late zoealstages Whereas this process is converted toa spine in Neotrypaea uncinata at ZV and Cal-lianassa sl petalura at ZIV the anomuranhair is retained at ZIV in Callichirus majorand C islagrande (see Rodrigues 1976Strasser and Felder 1999a) Although notmentioned specifically in the descriptions theanomuran hair appears to be present in illus-trations of Callichirus sp at ZIII and Cgarthii at ZV (Rodrigues 1976 fig 35 Asteand Retamal 1983 fig 8Q) A bilobed epi-pod on the first maxilliped of ZI is consistentlyfound in Callichirus but is also present at ZIin Lepidophthalmus sinuensis (see Nates etal 1997) Other callianassids acquire thischaracter at ZII (L louisianensis Nates et al1997 Callianassa sl kewlramanii Sankolliand Shenoy 1975) at ZV (Neotrypaea unci-nata Aste and Retamal 1984) or at the de-capodid stage (Sergio mirim Rodrigues 1984C sl petalura Konishi et al 1990) Thusthe number of zoeal stages and setation of thetelson may serve as the best characters to dis-tinguish larvae of Callichirus from other gen-era However since larval descriptions arelacking for many callianassid species it is un-known if these characters (in combination)will prove to be diagnostic until descriptionsfor other species are published
ACKNOWLEDGEMENTS
We thank Robert Bourgeois Barbara Hasek JimMaKinster Sergio Nates and Kurt Strasser who pro-vided assistance in the field and laboratory and MaryE Rice Director of the Smithsonian Marine StationLink Port Florida who facilitated access to station fa-cilities We also thank R Bauer R Jaeger R Lemaitreand G Watson for their early comments on this manu-script and Raymond B Manning who facilitated fund-ing and gave advice on collecting sites Direct supportfor this study was provided to Karen M Strasser by adoctoral fellowship from the University of Louisianaa graduate student research grant from the LUMCON(Louisiana Universities Marine Consortium) Founda-tion and a summer research award from the Smith-sonian Marine Station Additional support for field andlab activities was provided to D L Felder under Loui-siana Sea Grant College Program Grant RCFB-21 USFish and Wildlife Service Cooperative Agreement 14-16-0009-89-96 (Task Order No 6) US Department of En-ergy grant DE-FG02-97ER12220 and several small proj-ect grants from the Smithsonian Marine Station Views
expressed herein are those of the authors and do not nec-essarily reflect views of supporting government agenciesor subagencies mentioned above This is contributionNumber 466 of the Smithsonian Marine Station and con-tribution Number 67 of the ULL Laboratory for Crus-tacean Research All experiments conducted in this studycomply with current applicable state and federal laws
LITERATURE CITED
Aste A and M Retamal 1983 Desarrollo larval deCallianassa garthi Retamal 1975 bajo condiciones delaboratoriomdashCiencia y Tecnologiacutea del Mar CONA 75ndash26
mdashmdashmdash and mdashmdashmdash 1984 Desarrollo larval de Cal-lianassa uncinata H Milne Edwards1837 (DecapodaCallianassidae) bajo condiciones de laboratoriomdashGayana Zooloacutegica 48 41ndash56
Felder D L 1978 Osmotic and ionic regulation in sev-eral western Atlantic Callianassidae (Crustacea De-capoda Thalassinidea)mdashBiological Bulletin 154409ndash429
mdashmdashmdash and R B Griffis 1994 Dominant infaunalcommunities at risk in shoreline habitats burrowingthalassinid CrustaceamdashOCS Study number MMS94ndash0007 United States Department of the InteriorMinerals Management Service Gulf of Mexico OCSRegional Office New Orleans Louisiana Pp 1ndash87
Forbes A T 1973 An unusual abbreviated larval lifehistory in the estuarine burrowing prawn Callianassakraussi (Crustacea Decapoda Thalassinidea)mdashMa-rine Biology 22 361ndash365
Konishi K 1989 Larval development of the mudshrimp Upogebia (Upogebia) major (de Haan) (Crus-tacea Thalassinidea Upogebiidae) under laboratoryconditions with comments on larval characters of tha-lassinid familiesmdashBulletin of the National ResearchInstitute of Aquaculture 15 1ndash17
mdashmdashmdash R Quintana and Y Fukuda 1990 A completedescription of larval stages of the ghost shrimp Cal-lianassa petalura Stimpson (Crustacea ThalassinideaCallianassidae) under laboratory conditionsmdashBulletinof the National Research Institute of Aquaculture 1727ndash49
Manning R B and D L Felder 1986 The status ofthe callianassid genus Callichirus Stimpson 1866(Crustacea Decapoda Thalassinidea)mdashProceedingsof the Biological Society of Washington 99 437ndash443
mdashmdashmdash and mdashmdashmdash 1991 Revision of the AmericanCallianassidae (Crustacea Decapoda Thalassinidea)mdashProceedings of the Biological Society of Washington104 764ndash794
Nates S F D L Felder and R Lemaitre 1997 Com-parative larval development in two species of the bur-rowing ghost shrimp genus Lepidophthalmus (Decap-oda Callianassidae)mdashJournal of Crustacean Biology17 497ndash519
Rodrigues S de A 1976 Sobre a reproduccedilatildeo embri-ologia e desenvolvimento larval de Callichirus majorSay 1818 (Crustacea Decapoda Thalassinidea)mdashBo-letim de Zoologia Universidade de Satildeo Paulo 185ndash104
mdashmdashmdash 1984 Desenvolvimento poacutes-embrionaacuterio deCallichirus mirim (Rodrigues 1971) obtido emcondiccedilotildees artificiais (Crustacea Decapoda Tha-lassinidea)mdashBoletim de Zoologia Universidade deSatildeo Paulo 8 239ndash256
STRASSER AND FELDER LARVAL DEVELOPMENT IN CALLICHIRUS ISLAGRANDE 117
Sankolli K N and S Shenoy 1975 Larval develop-ment of mud shrimp Callianassa (Callichirus) kewal-ramanii Sankolli in the laboratory (Crustacea Decap-oda)mdashBulletin of the Department of Marine ScienceUniversity of Cochin 4 704ndash720
Staton J L and D L Felder 1995 Genetic variationin populations of the ghost shrimp genus Callichirus(Crustacea Decapoda Thalassinoidea) in the westernAtlantic and Gulf of MexicomdashBulletin of Marine Sci-ence 56 523ndash536
Strasser K M and D L Felder 1998 Settlement cuesin successive developmental stages of the ghost shrimpCallichirus major and C islagrande (Crustacea De-capoda Thalassinidea)mdashMarine Biology
mdashmdashmdash and mdashmdashmdash 1999a Larval development of twopopulations of the ghost shrimp Callichirus major (De-capoda Thalassinidea)mdashJournal of Crustacean Biology
mdashmdashmdash and mdashmdashmdash 1999b Sand as a stimulus for set-tlement in the ghost shrimp Callichirus major (Say)and C islagrande (Schmitt) (Crustacea DecapodaThalassinidea)mdashJournal of Experimental Marine Bi-ology and Ecology 239 211ndash222
mdashmdashmdash and mdashmdashmdash 1999c Settlement cues in theFlorida Atlantic population of the ghost shrimp Cal-lichirus major (Crustacea Decapoda Thalassinidea)mdashMarine Ecology Progress Series
Thessalou-Legaki M 1990 Advanced larval develop-ment of Callianassa tyrrhena (Decapoda Tha-lassinidea) and the effect of environmental factorsmdashJournal of Crustacean Biology 10 659ndash666
RECEIVED 19 September 1998ACCEPTED 3 December 1998

106 JOURNAL OF CRUSTACEAN BIOLOGY VOL 20 NO 1 2000
tae on proximal lobe 6ndash9 setae on distal lobeendopod with 5 lobes 3+2+2+2+(2 or 3) se-tae scaphognathite with 21ndash26 setae all se-tae plumose
Maxilliped 1 (Fig 5g)mdashCoxa with 4ndash9 se-tae basis with 14ndash18 marginal setae endo-pod 4-segmented (3 or 4) 2 2 (3 or 4) se-tae exopod 2-segmented proximal segment
without setae distal segment with 5ndash7 setaeon distal margin bilobed epipod without se-tae all setae plumose
Maxilliped 2 (Fig 5h)mdashCoxa with 0ndash2 se-tae basis with 5 or 6 setae endopod 4-seg-mented (4ndash6) 1 3 (4ndash6) setae exopod un-segmented 7 or 8 setae on distal margin allsetae plumose
Fig 4 Callichirus islagrande ZII stage a antennule b antenna c right mandible d left mandible e maxillulef maxilla (plumose condition not shown) g maxilliped 1 h maxilliped 2 i maxilliped 3 j pereiopod 1 k pe-reiopod 2 1 telson Scales = 02 mm
STRASSER AND FELDER LARVAL DEVELOPMENT IN CALLICHIRUS ISLAGRANDE 107
Maxilliped 3 (Fig 6a)mdashCoxa with 0 or 1seta basis with 1ndash3 setae endopod 4-seg-mented (1 or 2) 2 (2ndash4) (4 or 5) setae exo-pod unsegmented 7 or 8 setae on distal mar-gin all setae plumose
Pereiopod 1 (Fig 6b)mdashChelate and bira-mous coxa without setae basis with 0 or 1seta endopod 4-segmented 0 (1 or 2) (2ndash4)2 setae exopod with 6 or 7 setae on distalmargin
Fig 5 Callichirus islagrande ZIII stage a antennule b antenna c right mandible d left mandible e maxil-lule f maxilla (plumose condition not shown) g maxilliped 1 h maxilliped 2 All setae on the maxilla were plumoseScales = 02 mm
108 JOURNAL OF CRUSTACEAN BIOLOGY VOL 20 NO 1 2000
Pereiopod 2 (Fig 6c)mdashChelate and biramouscoxa without setae basis with 0 or 1 seta en-dopod 4-segmented 0 (0ndash2) (0ndash4) 2 setaeexopod with 5ndash7 setae on distal margin
Pereiopods 3ndash5 (Fig 6d e f )mdashWithout setae
Pleopods (Figs 1c 6g)mdashBilobed buds onabdominal segments 2ndash5 first pair smallerthan others
Telson and Uropods (Fig 6h)mdashNo changein arrangement of posterior telsonal processesuropods biramous endopod without setae
Fig 6 Callichirus islagrande ZIII stage a maxilliped 3 b pereiopod 1 c pereiopod 2 d pereiopod 3 e pe-reiopod 4 f pereiopod 5 g pleopods from fourth abdominal somite h telson Scales = 02 mm
STRASSER AND FELDER LARVAL DEVELOPMENT IN CALLICHIRUS ISLAGRANDE 109
exopod with 10 or 11 marginal plumose se-tae ventral spine near base of protopod
Zoea IVFigs 2d 7andashh 8andashi
SizemdashCL = 192 plusmn 009 mm range 169ndash215mm TL = 482 plusmn 025 mm range 424ndash536mm
Carapace (Fig 2d)mdashNo marked changefrom ZIII
Abdomen (Fig 2d)mdashNo marked change fromZIII
Antennule (Fig 7a)mdashBiramous peduncle2-segmented with 1+(1ndash4) short setae onouter margin and 0 or 1 plumose seta on in-ner margin of proximal segment distal seg-ment with 3ndash5 plumose setae on inner mar-gin 1 plumose seta between rami and 0ndash3short setae on outer margin endopod with 1or 2 plumose setae exopod with 6 or 7 ter-minal and 2 subterminal aesthetascs
Antenna (Fig 7b)mdashProtopod with 1 distalspine on outer margin endopod with 0ndash2 se-tae scaphocerite armed with 1 strong disto-lateral spine 12ndash14 plumose setae on innermargin and 1 plumose seta on outer margin
Mandible (Fig 7c d)mdashIncisor process with8 teeth on right mandible 6ndash8 teeth on leftmandible molar process denticulate on bothmandibles palps present
Maxillule (Fig 7e)mdashCoxal endite with12ndash14 marginal plumose setae basal enditewith 3 plumose setae 5 or 6 large dentatespines and 5 or 6 smaller dentate spinesinner margin with 1 plumose seta endopod3-segmented distinction between segmentssometimes obscure 2 2 (3 or 4) setae pro-topod without setation
Maxilla (Fig 7f )mdashCoxal endite bilobed14ndash19 setae on proximal lobe 5 setae on dis-tal lobe basal endite bilobed 6ndash10 setae onproximal lobe 8 or 9 setae on distal lobeendopod with 5 lobes 3+(2 or 3)+2+2+3 se-tae scaphognathite with 27ndash32 setae all se-tae plumose
Maxilliped 1 (Fig 7g)mdashCoxa with 2ndash8 se-tae basis with 16ndash19 marginal setae endo-pod 4-segmented (3 or 4) 2 2 4 setae exo-
pod 2-segmented proximal segment withoutsetae distal segment with 6 or 7 setae on dis-tal margin bilobed epipod without setae allsetae plumose
Maxilliped 2 (Fig 7h)mdashCoxa with 0ndash3 se-tae basis with 4ndash6 setae endopod 4-seg-mented (5ndash8) 1 (3ndash5) (4 or 5) setae exo-pod unsegmented 7 or 8 setae on distal mar-gin all setae plumose
Maxilliped 3 (Fig 8a)mdashCoxa with 0 or 1seta basis with 2ndash4 setae endopod 4-seg-mented (l or 2) 2 (3ndash5) (3 or 4) setae exo-pod unsegmented 7 or 8 setae on distal mar-gin all setae plumose
Pereiopod 1 (Fig 8b)mdashBiramous cheliformcoxa without setae basis with 0ndash2 setae en-dopod 4-segmented (0 or 1) (1 or 2) (2ndash4)(1 or 2) setae exopod with 5ndash7 setae on dis-tal margin
Pereiopod 2 (Fig 8c)mdashBiramous cheliformcoxa without setae basis with 0 or 1 setaendopod 4-segmented 0 (1 or 2) (3ndash5) (2or 3) setae exopod with 5ndash7 setae on distalmargin
Pereiopod 3 (Fig 8d)mdashBiramous coxa andbasis without setae endopod 4-segmented0 (0ndash2) (3 or 4) 2 setae propodus blade-like exopod with 4 or 5 terminal setae
Pereiopod 4 (Fig 8e)mdashBiramous coxa andbasis without setae endopod 4-segmented (0or 1) (0ndash2) (2ndash4) 2 setae exopod with 0ndash2setae
Pereiopod 5 (Fig 8f )mdashUniramous 2 setaeon terminal segment 4 or 5 setae on adja-cent segment
Pleopods (Fig 8g h)mdashLarger than at ZIIIstill without setae
Telson and Uropods (Fig 8i)mdashTelson trian-gular narrow anteriorly no anal spine pos-terior margin with (12ndash15)+1+(13 or 14)processes arranged as (I II III IV III II I)(I) outermost process a plumodenticulate seta(II) second process ldquoanomuran hairrdquo (III) 10ndash13plumodenticulate setae and (IV) naked spineuropods biramous endopod with 7ndash11 marginalplumose setae exopod with 1 naked distolat-eral spine 12ndash14 marginal plumose setae
110 JOURNAL OF CRUSTACEAN BIOLOGY VOL 20 NO 1 2000
Decapodid (first postlarva)Figs 2e 9cndashj l0andashi
SizemdashCL = 133 plusmn 002 mm range129ndash138 mm TL = 378 plusmn 011 mm range361ndash405 mm
Carapace (Fig 2e)mdashShorter than abdomenno anterolateral serrations with pterygostomial
projections rostrum reduced without serra-tions sometimes bending down between eyes
Abdomen (Fig 2e)mdashSomites broad and flat-tened posterior projections rounded with 0ndash7setae somite 2 rectangular somites 3ndash5 oval
Antennule (Fig 9c)mdashPeduncle 3-segmentedproximal segment with 17ndash24 setae 13ndash15 of
Fig 7 Callichirus islagrande ZIV stage a antennule b antenna c right mandible d left mandible e maxil-lule f maxilla (plumose condition not shown) g maxilliped 1 h maxilliped 2 All setae on the maxilla were plumoseScales = 02 mm
STRASSER AND FELDER LARVAL DEVELOPMENT IN CALLICHIRUS ISLAGRANDE 111
these associated with statocyst intermediatesegment with 6ndash10 setae distal segment of pe-duncle with 2 vertical rows of setae one with4 or 5 setae other with 2ndash4 setae 1ndash3 addi-tional setae on outer margin and 3ndash5 short se-tae on distal margin endopod 4ndash6 segmented(1ndash3) (0 or 1) (1ndash4) (1ndash4) (3ndash6) (3ndash6) se-tae exopod 3ndash5-segmented 0 (0ndash4) (0ndash4)(1ndash4) setae +2 asthetascs (5ndash8) asthetascs ()
Antenna (Fig 9d)mdashPeduncle 2-segmentedproximal segment with 0 or 1 seta distal seg-ment with 1ndash3 setae on inner margin spinepresent in some flagellum ranging from 14ndash18
segments (0 or 1) (2ndash4) variable region(1ndash3 articles with 0ndash2 setae) (0ndash4) (0ndash2)(2ndash5) (1ndash4) (3ndash6) (0ndash3) (2ndash5) (3ndash6)(0ndash3) (3ndash6) (0 or 1) (1ndash7) (4ndash9) setae exo-pod reduced to small lobe with 0ndash5 setae
Mandible (Fig 9e f )mdashBoth mandibles sim-ilar incisor process with 5ndash8 teeth molarprocess smooth or with small ridges palp3-segmented 0 (0 or 1) (8ndash12) setae
Maxillule (Fig 9g)mdashCoxal endite with15ndash22 marginal setae and 0 or 1 submarginal
Fig 8 Callichirus islagrande ZIV stage a maxilliped 3 b pereiopod 1 c pereiopod 2 d pereiopod 3 e pe-reiopod 4 f pereiopod 5 g pleopods from first abdominal somite h pleopods from fourth abdominal somite i tel-son Scales = 02 mm
112 JOURNAL OF CRUSTACEAN BIOLOGY VOL 20 NO 1 2000
seta basal endite with 3ndash5 plumose setae14ndash19 dentate spines 0ndash2 plumose setae oninner margin endopod reduced unsegmentedwith 0ndash8 setae
Maxilla (Fig 9h)mdashCoxal endite trilobed(17ndash21)+(7ndash10)+(6ndash9) setae basal enditebilobed 8ndash10 setae on proximal lobe 8ndash15setae on distal lobe endopod reduced with
Fig 9 Callichirus islagrande ZV stage a one example of the antenna b one example of pleopods Decapodidstage c antennule d antenna e right mandible f left mandible g maxillule h maxilla (plumose condition notshown) i maxilliped 1 j maxilliped 2 Scales = 02 mm
STRASSER AND FELDER LARVAL DEVELOPMENT IN CALLICHIRUS ISLAGRANDE 113
5ndash11 setae scaphognathite with 37ndash45 se-tae all setae plumose
Maxilliped 1 (Fig 9i)mdashCoxa with 8ndash12 se-tae basis highly setose with 27ndash39 setae en-dopod reduced with 0ndash11 setae exopod un-segmented 6ndash14 setae bilobed epipod with-out setae
Maxilliped 2 (Fig 9j)mdashCoxa partially fusedto basis with 1ndash4 setae basis with 3ndash5 se-
tae endopod 4-segmented (16ndash23) (0 or 1)(6ndash11) (4ndash8) setae exopod with 0ndash8 setaeepipod present but without setae
Maxilliped 3 (Fig 10a)mdashCoxa partiallyfused to basis with 0ndash3 setae basis with 1ndash4setae endopod 5-segmented (11ndash19) (9ndash15)(7ndash11) (8ndash21) (5ndash11) setae exopod usuallyreduced and without setae occasionally with2ndash9 setae epipod present
Fig 10 Callichirus islagrande decapodid stage a maxilliped 3 b pereiopod 1 c pereiopod 2 d pereiopod 3e pereiopod 4 f pereiopod 5 g pleopods from first abdominal somite h pleopods from fourth abdominal somitei telson Scales = 02 mm
114 JOURNAL OF CRUSTACEAN BIOLOGY VOL 20 NO 1 2000
Pereiopod 1 (Fig 10b)mdashChelate biramous7-segmented coxa with 0ndash6 setae basis with2ndash5 setae ischium with 4ndash7 setae and 0ndash3spines merus with 5ndash8 setae carpus with10ndash14 setae propodus with 27ndash37 setae and3ndash4 spines on inner margin of immovable fin-ger dactylus with 21ndash33 setae exopod usu-ally reduced and without setae occasionallywith 4ndash7 setae
Pereiopod 2 (Fig 10c)mdashChelate biramous7-segmented coxa with 2ndash5 setae basis with1ndash5 setae ischium with 4ndash10 setae meruswith 15ndash20 setae carpus with 8ndash12 setaepropodus with 30ndash40 setae and 1ndash4 spineson inner margin of immovable finger dacty-lus with 21ndash30 setae exopod usually reduced0ndash7 setae
Pereiopod 3 (Fig 10d)mdashBiramous 7-seg-mented coxa with 2ndash5 setae basis with 1ndash3setae ischium with 5ndash8 setae merus with6ndash9 setae carpus with 7ndash11 setae propodusbladelike with 25ndash34 setae 1 spine on innerdistal margin dactylus with 15ndash21 setae exo-pod usually reduced 0ndash5 setae
Pereiopod 4 (Fig 10e)mdashBiramous 7-seg-mented coxa with 2ndash7 setae basis with 1ndash4setae ischium with 3ndash8 setae merus with5ndash11 setae carpus with 4ndash9 setae propoduswith 27ndash38 setae 1 spine on inner distal mar-gin dactylus with 15ndash19 setae exopod usu-ally reduced with 0ndash2 setae
Pereiopod 5 (Fig 10f )mdashCheliform unira-mous 7-segmented coxa with 2ndash6 setae ba-sis with l or 2 setae ischium with 2ndash5 setaemerus with 5ndash9 setae carpus with 4ndash7 se-tae propodus with 22ndash30 setae 1 spine onimmovable finger dactylus with 6ndash10 setaeexopod absent
Pleopods (somite 2) (Fig 10g)mdashBiramouscoxa partially fused to basis without setaebasis with 0ndash3 setae endopod with 3ndash5hooked setae on appendix interna 10ndash17 mar-ginal plumose setae 0ndash2 submarginal nakedsetae exopod with 21ndash27 marginal plumosesetae and 0ndash1 submarginal naked seta
Pleopods (somites 3ndash5) (Fig 10h)mdashBiramouscoxa without setae basis with 3ndash6 marginalplumose setae endopod with 3ndash6 hooked se-tae on appendix interna 10ndash18 marginalplumose setae and 0ndash3 naked submarginal se-tae exopod with 20ndash29 marginal plumose se-tae and 0 or 1 submarginal naked seta
Telson and Uropods (Fig 10i)mdashWith 24ndash36processes on posterior margin median pro-cess usually absent or reduced uropods bi-ramous endopod with 19ndash25 plumose setaeexopod with 35ndash58 plumose setae protopodwith 3ndash5 setae
DISCUSSION
As in other species of Callichirus (see Ro-drigues 1976 Aste and Retamal 1983Strasser and Felder in press b) larval devel-opment of C islagrande included four or fivezoeal stages before metamorphosis to the de-capodid ZI larvae may be identified by thepresence of an elongate rounded rostrumsessile eyes and a lack of pereiopodspleopods or uropods (Fig 1a) At ZII lar-vae acquired biramous buds of pereiopods 1and 2 eyes were separated from the carapaceand the rostrum became flattened and serratedlaterally (Fig 1b) Buds of pleopods on ab-dominal segments 2ndash5 pereiopods 3ndash5 anduropods were developed at ZIII (Fig 1c) Inaddition to further development of structuresmentioned thus far ZIV larvae could be dis-tinguished by presence of a distal spine on theexopod and setae on the endopod of each uro-pod (Fig 1d) When it occurred ZV typicallyresembled ZIV but varied widely in mor-phology Appendages at ZV resembled thoseof ZIV the decapodid stage or some grada-tion between the two (see Figs 9a b for ex-amples) The decapodid stage was similar toother species of Callichirus in the reductionof larval characters such as the elongate ros-trum dorsal spines on the abdomen and ex-opods of the pereiopods (Fig le) Acquisitionof fan-like pleopods marked the transition toa primarily abdominal mode of swimming atthis stage
In the Florida Atlantic population of C ma-jor both salinity and presence of putative set-tlement cues at ZIV impacted the observednumber of zoeal stages and the duration ofZIV (Strasser and Felder in preparation1999c) In C islagrande the percentage oflarvae that passed through five versus fourzoeal stages varied (9ndash89) between parentalfemales but was not shown to depend onsalinity or presence of various putative set-tlement cues (Strasser and Felder 1998 inpreparation) Larvae that molted from ZIVto ZV were usually normal morphologicallyand had mortality similar to those that molteddirectly from ZIV to D In contrast larvae of
STRASSER AND FELDER LARVAL DEVELOPMENT IN CALLICHIRUS ISLAGRANDE 115
the Florida Atlantic C major that molted toZV had a higher incidence of mortality anddeformity at D than did those that had by-passed ZV (Strasser and Felder 1999a c)
The number of zoeal stages appears to beless variable in Callichirus garthi and theGulf of Mexico population of C major thanin congeneric populations studied to dateCallichirus garthi was reported to have fivezoeal stages and a long larval duration reach-ing the decapodid stage 48 days after hatch-ing (Aste and Retamal 1983) Developmentof the Gulf population of C major wasshorter in duration (decapodid reached about10 days after hatching) and typically includedfour zoeal stages (Strasser and Felder 1999a)animals rarely passed through three or fivezoeal stages before the molt to the decapodidAlthough the number of stages is reducedfrom that observed in C garthi developmentin the Gulf population of C major cannot beclassified as abbreviated There are species inboth the Callianassinae (Callianassa slkraussi Stebbing C sl kewalramanii and Csl tyrrhena (Petagna)) and Callichirinae(Sergio mirim Lepidophthalmus sinuensisand L louisianensis) that have only two zoealstages before the molt to the decapodid(Forbes 1973 Sankolli and Shenoy 1975Rodrigues 1984 Thessalou-Legaki 1990Nates et al 1997)
Compared to other species of the genusCallichirus C islagrande is morphologicallycloser to C garthi (from Chile) than to theGulf of Mexico population of C major whichoverlaps in distribution with C islagrande Inthe zoeal phase of C islagrande and Cgarthi the telson is concave and the outer-most process on the ventral margin is articu-lated In both C major and Callichirus sp theposterior margin of the telson is straight andthe outermost process is unarticulated duringthe zoeal stages (Rodrigues 1976 Strasserand Felder 1999a) There are also differencesin spination of the abdomen In zoeae of Cislagrande and C garthi the dorsal spine ofabdominal segment 3 is not longer than thosefound on segments 4 and 5 (Aste and Reta-mal 1983) In contrast the dorsal spine of ab-dominal segment 3 is approximately twice aslong as the dorsal spines on segments 4 and5 in early zoeal stages of C major and Cal-lichirus sp (Rodrigues 1976 Aste and Re-tamal 1983 Strasser and Felder 1999a) Thegreatest difference between these groups is
the number of pleopod pairs at zoeal and de-capodid stages Callichirus islagrande and Cgarthi have pleopods on abdominal segments2ndash5 while pleopods are present only on ab-dominal segments 3ndash5 in C major and Cal-lichirus sp (Rodrigues 1976 Aste and Re-tamal 1983 Strasser and Felder 1999a) Al-though Callichirus islagrande and C garthiare similar morphologically there are differ-ences between these two species Callichirusislagrande as well as C major and Cal-lichirus sp develops pleopod buds andchelate pereiopods at ZIII whereas C garthidoes not acquire these characters until ZIV
There are few callianassid species forwhich the larval development has been de-scribed (Sankolli and Shenoy 1975 Ro-drigues 1976 1984 Aste and Retamal 19831984 Konishi et al 1990 Nates et al 1997Strasser and Felder 1999a) all of which aremembers of the Callichirinae and Cal-lianassinae Among species there is a greatdeal of overlap in larval characters makingit difficult to draw distinctions between thesesubfamilies At present information on lar-val development of species in the Callichiri-nae is limited to larval descriptions of Cal-lichirus major C garthi Callichirus sp Cislagrande Sergio mirim Lepidophthalmuslouisianensis and L sinuensis (see Ro-drigues 1976 1984 Aste and Retamal 1983Nates et al 1997 Strasser and Felder1999a) As mentioned above species of Cal-lichirus have been shown to have four to fivezoeal stages while the other members exhibitabbreviated development that consists of onlytwo zoeal stages The most obvious differencebetween larvae of the species of Callichirinaewith abbreviated development is the numberof pleopod pairs Larvae of S mirim havepleopods on abdominal segments 3ndash5whereas L louisianensis and L sinuensishave pleopods on segments 2ndash5 (Rodrigues1984 Nates et al 1997)
Ghost shrimps of the genus Callichirus maybe distinguished from other genera of the Cal-lianassidae by the cuticular pattern on the dor-sal surface of the abdomen and by the strap-like shape of the uropodal endopods (Manningand Felder 1991) Whereas these charactersmay be used to distinguish adults of the genusCallichirus they are not present at zoeal anddecapodid stages Larvae may be distin-guished from all other callianassid genera byhaving four to five zoeal stages and numer-
116 JOURNAL OF CRUSTACEAN BIOLOGY VOL 20 NO 1 2000
ous processes (12ndash25+1+12ndash25) on the pos-terior margin of the telson during the earlyzoeal stages Larvae of Callichirus may alsodiffer from other callianassids with extendeddevelopment by the retention of the anomu-ran hair as the second process on the poste-rior margin of the telson in the late zoealstages Whereas this process is converted toa spine in Neotrypaea uncinata at ZV and Cal-lianassa sl petalura at ZIV the anomuranhair is retained at ZIV in Callichirus majorand C islagrande (see Rodrigues 1976Strasser and Felder 1999a) Although notmentioned specifically in the descriptions theanomuran hair appears to be present in illus-trations of Callichirus sp at ZIII and Cgarthii at ZV (Rodrigues 1976 fig 35 Asteand Retamal 1983 fig 8Q) A bilobed epi-pod on the first maxilliped of ZI is consistentlyfound in Callichirus but is also present at ZIin Lepidophthalmus sinuensis (see Nates etal 1997) Other callianassids acquire thischaracter at ZII (L louisianensis Nates et al1997 Callianassa sl kewlramanii Sankolliand Shenoy 1975) at ZV (Neotrypaea unci-nata Aste and Retamal 1984) or at the de-capodid stage (Sergio mirim Rodrigues 1984C sl petalura Konishi et al 1990) Thusthe number of zoeal stages and setation of thetelson may serve as the best characters to dis-tinguish larvae of Callichirus from other gen-era However since larval descriptions arelacking for many callianassid species it is un-known if these characters (in combination)will prove to be diagnostic until descriptionsfor other species are published
ACKNOWLEDGEMENTS
We thank Robert Bourgeois Barbara Hasek JimMaKinster Sergio Nates and Kurt Strasser who pro-vided assistance in the field and laboratory and MaryE Rice Director of the Smithsonian Marine StationLink Port Florida who facilitated access to station fa-cilities We also thank R Bauer R Jaeger R Lemaitreand G Watson for their early comments on this manu-script and Raymond B Manning who facilitated fund-ing and gave advice on collecting sites Direct supportfor this study was provided to Karen M Strasser by adoctoral fellowship from the University of Louisianaa graduate student research grant from the LUMCON(Louisiana Universities Marine Consortium) Founda-tion and a summer research award from the Smith-sonian Marine Station Additional support for field andlab activities was provided to D L Felder under Loui-siana Sea Grant College Program Grant RCFB-21 USFish and Wildlife Service Cooperative Agreement 14-16-0009-89-96 (Task Order No 6) US Department of En-ergy grant DE-FG02-97ER12220 and several small proj-ect grants from the Smithsonian Marine Station Views
expressed herein are those of the authors and do not nec-essarily reflect views of supporting government agenciesor subagencies mentioned above This is contributionNumber 466 of the Smithsonian Marine Station and con-tribution Number 67 of the ULL Laboratory for Crus-tacean Research All experiments conducted in this studycomply with current applicable state and federal laws
LITERATURE CITED
Aste A and M Retamal 1983 Desarrollo larval deCallianassa garthi Retamal 1975 bajo condiciones delaboratoriomdashCiencia y Tecnologiacutea del Mar CONA 75ndash26
mdashmdashmdash and mdashmdashmdash 1984 Desarrollo larval de Cal-lianassa uncinata H Milne Edwards1837 (DecapodaCallianassidae) bajo condiciones de laboratoriomdashGayana Zooloacutegica 48 41ndash56
Felder D L 1978 Osmotic and ionic regulation in sev-eral western Atlantic Callianassidae (Crustacea De-capoda Thalassinidea)mdashBiological Bulletin 154409ndash429
mdashmdashmdash and R B Griffis 1994 Dominant infaunalcommunities at risk in shoreline habitats burrowingthalassinid CrustaceamdashOCS Study number MMS94ndash0007 United States Department of the InteriorMinerals Management Service Gulf of Mexico OCSRegional Office New Orleans Louisiana Pp 1ndash87
Forbes A T 1973 An unusual abbreviated larval lifehistory in the estuarine burrowing prawn Callianassakraussi (Crustacea Decapoda Thalassinidea)mdashMa-rine Biology 22 361ndash365
Konishi K 1989 Larval development of the mudshrimp Upogebia (Upogebia) major (de Haan) (Crus-tacea Thalassinidea Upogebiidae) under laboratoryconditions with comments on larval characters of tha-lassinid familiesmdashBulletin of the National ResearchInstitute of Aquaculture 15 1ndash17
mdashmdashmdash R Quintana and Y Fukuda 1990 A completedescription of larval stages of the ghost shrimp Cal-lianassa petalura Stimpson (Crustacea ThalassinideaCallianassidae) under laboratory conditionsmdashBulletinof the National Research Institute of Aquaculture 1727ndash49
Manning R B and D L Felder 1986 The status ofthe callianassid genus Callichirus Stimpson 1866(Crustacea Decapoda Thalassinidea)mdashProceedingsof the Biological Society of Washington 99 437ndash443
mdashmdashmdash and mdashmdashmdash 1991 Revision of the AmericanCallianassidae (Crustacea Decapoda Thalassinidea)mdashProceedings of the Biological Society of Washington104 764ndash794
Nates S F D L Felder and R Lemaitre 1997 Com-parative larval development in two species of the bur-rowing ghost shrimp genus Lepidophthalmus (Decap-oda Callianassidae)mdashJournal of Crustacean Biology17 497ndash519
Rodrigues S de A 1976 Sobre a reproduccedilatildeo embri-ologia e desenvolvimento larval de Callichirus majorSay 1818 (Crustacea Decapoda Thalassinidea)mdashBo-letim de Zoologia Universidade de Satildeo Paulo 185ndash104
mdashmdashmdash 1984 Desenvolvimento poacutes-embrionaacuterio deCallichirus mirim (Rodrigues 1971) obtido emcondiccedilotildees artificiais (Crustacea Decapoda Tha-lassinidea)mdashBoletim de Zoologia Universidade deSatildeo Paulo 8 239ndash256
STRASSER AND FELDER LARVAL DEVELOPMENT IN CALLICHIRUS ISLAGRANDE 117
Sankolli K N and S Shenoy 1975 Larval develop-ment of mud shrimp Callianassa (Callichirus) kewal-ramanii Sankolli in the laboratory (Crustacea Decap-oda)mdashBulletin of the Department of Marine ScienceUniversity of Cochin 4 704ndash720
Staton J L and D L Felder 1995 Genetic variationin populations of the ghost shrimp genus Callichirus(Crustacea Decapoda Thalassinoidea) in the westernAtlantic and Gulf of MexicomdashBulletin of Marine Sci-ence 56 523ndash536
Strasser K M and D L Felder 1998 Settlement cuesin successive developmental stages of the ghost shrimpCallichirus major and C islagrande (Crustacea De-capoda Thalassinidea)mdashMarine Biology
mdashmdashmdash and mdashmdashmdash 1999a Larval development of twopopulations of the ghost shrimp Callichirus major (De-capoda Thalassinidea)mdashJournal of Crustacean Biology
mdashmdashmdash and mdashmdashmdash 1999b Sand as a stimulus for set-tlement in the ghost shrimp Callichirus major (Say)and C islagrande (Schmitt) (Crustacea DecapodaThalassinidea)mdashJournal of Experimental Marine Bi-ology and Ecology 239 211ndash222
mdashmdashmdash and mdashmdashmdash 1999c Settlement cues in theFlorida Atlantic population of the ghost shrimp Cal-lichirus major (Crustacea Decapoda Thalassinidea)mdashMarine Ecology Progress Series
Thessalou-Legaki M 1990 Advanced larval develop-ment of Callianassa tyrrhena (Decapoda Tha-lassinidea) and the effect of environmental factorsmdashJournal of Crustacean Biology 10 659ndash666
RECEIVED 19 September 1998ACCEPTED 3 December 1998

STRASSER AND FELDER LARVAL DEVELOPMENT IN CALLICHIRUS ISLAGRANDE 107
Maxilliped 3 (Fig 6a)mdashCoxa with 0 or 1seta basis with 1ndash3 setae endopod 4-seg-mented (1 or 2) 2 (2ndash4) (4 or 5) setae exo-pod unsegmented 7 or 8 setae on distal mar-gin all setae plumose
Pereiopod 1 (Fig 6b)mdashChelate and bira-mous coxa without setae basis with 0 or 1seta endopod 4-segmented 0 (1 or 2) (2ndash4)2 setae exopod with 6 or 7 setae on distalmargin
Fig 5 Callichirus islagrande ZIII stage a antennule b antenna c right mandible d left mandible e maxil-lule f maxilla (plumose condition not shown) g maxilliped 1 h maxilliped 2 All setae on the maxilla were plumoseScales = 02 mm
108 JOURNAL OF CRUSTACEAN BIOLOGY VOL 20 NO 1 2000
Pereiopod 2 (Fig 6c)mdashChelate and biramouscoxa without setae basis with 0 or 1 seta en-dopod 4-segmented 0 (0ndash2) (0ndash4) 2 setaeexopod with 5ndash7 setae on distal margin
Pereiopods 3ndash5 (Fig 6d e f )mdashWithout setae
Pleopods (Figs 1c 6g)mdashBilobed buds onabdominal segments 2ndash5 first pair smallerthan others
Telson and Uropods (Fig 6h)mdashNo changein arrangement of posterior telsonal processesuropods biramous endopod without setae
Fig 6 Callichirus islagrande ZIII stage a maxilliped 3 b pereiopod 1 c pereiopod 2 d pereiopod 3 e pe-reiopod 4 f pereiopod 5 g pleopods from fourth abdominal somite h telson Scales = 02 mm
STRASSER AND FELDER LARVAL DEVELOPMENT IN CALLICHIRUS ISLAGRANDE 109
exopod with 10 or 11 marginal plumose se-tae ventral spine near base of protopod
Zoea IVFigs 2d 7andashh 8andashi
SizemdashCL = 192 plusmn 009 mm range 169ndash215mm TL = 482 plusmn 025 mm range 424ndash536mm
Carapace (Fig 2d)mdashNo marked changefrom ZIII
Abdomen (Fig 2d)mdashNo marked change fromZIII
Antennule (Fig 7a)mdashBiramous peduncle2-segmented with 1+(1ndash4) short setae onouter margin and 0 or 1 plumose seta on in-ner margin of proximal segment distal seg-ment with 3ndash5 plumose setae on inner mar-gin 1 plumose seta between rami and 0ndash3short setae on outer margin endopod with 1or 2 plumose setae exopod with 6 or 7 ter-minal and 2 subterminal aesthetascs
Antenna (Fig 7b)mdashProtopod with 1 distalspine on outer margin endopod with 0ndash2 se-tae scaphocerite armed with 1 strong disto-lateral spine 12ndash14 plumose setae on innermargin and 1 plumose seta on outer margin
Mandible (Fig 7c d)mdashIncisor process with8 teeth on right mandible 6ndash8 teeth on leftmandible molar process denticulate on bothmandibles palps present
Maxillule (Fig 7e)mdashCoxal endite with12ndash14 marginal plumose setae basal enditewith 3 plumose setae 5 or 6 large dentatespines and 5 or 6 smaller dentate spinesinner margin with 1 plumose seta endopod3-segmented distinction between segmentssometimes obscure 2 2 (3 or 4) setae pro-topod without setation
Maxilla (Fig 7f )mdashCoxal endite bilobed14ndash19 setae on proximal lobe 5 setae on dis-tal lobe basal endite bilobed 6ndash10 setae onproximal lobe 8 or 9 setae on distal lobeendopod with 5 lobes 3+(2 or 3)+2+2+3 se-tae scaphognathite with 27ndash32 setae all se-tae plumose
Maxilliped 1 (Fig 7g)mdashCoxa with 2ndash8 se-tae basis with 16ndash19 marginal setae endo-pod 4-segmented (3 or 4) 2 2 4 setae exo-
pod 2-segmented proximal segment withoutsetae distal segment with 6 or 7 setae on dis-tal margin bilobed epipod without setae allsetae plumose
Maxilliped 2 (Fig 7h)mdashCoxa with 0ndash3 se-tae basis with 4ndash6 setae endopod 4-seg-mented (5ndash8) 1 (3ndash5) (4 or 5) setae exo-pod unsegmented 7 or 8 setae on distal mar-gin all setae plumose
Maxilliped 3 (Fig 8a)mdashCoxa with 0 or 1seta basis with 2ndash4 setae endopod 4-seg-mented (l or 2) 2 (3ndash5) (3 or 4) setae exo-pod unsegmented 7 or 8 setae on distal mar-gin all setae plumose
Pereiopod 1 (Fig 8b)mdashBiramous cheliformcoxa without setae basis with 0ndash2 setae en-dopod 4-segmented (0 or 1) (1 or 2) (2ndash4)(1 or 2) setae exopod with 5ndash7 setae on dis-tal margin
Pereiopod 2 (Fig 8c)mdashBiramous cheliformcoxa without setae basis with 0 or 1 setaendopod 4-segmented 0 (1 or 2) (3ndash5) (2or 3) setae exopod with 5ndash7 setae on distalmargin
Pereiopod 3 (Fig 8d)mdashBiramous coxa andbasis without setae endopod 4-segmented0 (0ndash2) (3 or 4) 2 setae propodus blade-like exopod with 4 or 5 terminal setae
Pereiopod 4 (Fig 8e)mdashBiramous coxa andbasis without setae endopod 4-segmented (0or 1) (0ndash2) (2ndash4) 2 setae exopod with 0ndash2setae
Pereiopod 5 (Fig 8f )mdashUniramous 2 setaeon terminal segment 4 or 5 setae on adja-cent segment
Pleopods (Fig 8g h)mdashLarger than at ZIIIstill without setae
Telson and Uropods (Fig 8i)mdashTelson trian-gular narrow anteriorly no anal spine pos-terior margin with (12ndash15)+1+(13 or 14)processes arranged as (I II III IV III II I)(I) outermost process a plumodenticulate seta(II) second process ldquoanomuran hairrdquo (III) 10ndash13plumodenticulate setae and (IV) naked spineuropods biramous endopod with 7ndash11 marginalplumose setae exopod with 1 naked distolat-eral spine 12ndash14 marginal plumose setae
110 JOURNAL OF CRUSTACEAN BIOLOGY VOL 20 NO 1 2000
Decapodid (first postlarva)Figs 2e 9cndashj l0andashi
SizemdashCL = 133 plusmn 002 mm range129ndash138 mm TL = 378 plusmn 011 mm range361ndash405 mm
Carapace (Fig 2e)mdashShorter than abdomenno anterolateral serrations with pterygostomial
projections rostrum reduced without serra-tions sometimes bending down between eyes
Abdomen (Fig 2e)mdashSomites broad and flat-tened posterior projections rounded with 0ndash7setae somite 2 rectangular somites 3ndash5 oval
Antennule (Fig 9c)mdashPeduncle 3-segmentedproximal segment with 17ndash24 setae 13ndash15 of
Fig 7 Callichirus islagrande ZIV stage a antennule b antenna c right mandible d left mandible e maxil-lule f maxilla (plumose condition not shown) g maxilliped 1 h maxilliped 2 All setae on the maxilla were plumoseScales = 02 mm
STRASSER AND FELDER LARVAL DEVELOPMENT IN CALLICHIRUS ISLAGRANDE 111
these associated with statocyst intermediatesegment with 6ndash10 setae distal segment of pe-duncle with 2 vertical rows of setae one with4 or 5 setae other with 2ndash4 setae 1ndash3 addi-tional setae on outer margin and 3ndash5 short se-tae on distal margin endopod 4ndash6 segmented(1ndash3) (0 or 1) (1ndash4) (1ndash4) (3ndash6) (3ndash6) se-tae exopod 3ndash5-segmented 0 (0ndash4) (0ndash4)(1ndash4) setae +2 asthetascs (5ndash8) asthetascs ()
Antenna (Fig 9d)mdashPeduncle 2-segmentedproximal segment with 0 or 1 seta distal seg-ment with 1ndash3 setae on inner margin spinepresent in some flagellum ranging from 14ndash18
segments (0 or 1) (2ndash4) variable region(1ndash3 articles with 0ndash2 setae) (0ndash4) (0ndash2)(2ndash5) (1ndash4) (3ndash6) (0ndash3) (2ndash5) (3ndash6)(0ndash3) (3ndash6) (0 or 1) (1ndash7) (4ndash9) setae exo-pod reduced to small lobe with 0ndash5 setae
Mandible (Fig 9e f )mdashBoth mandibles sim-ilar incisor process with 5ndash8 teeth molarprocess smooth or with small ridges palp3-segmented 0 (0 or 1) (8ndash12) setae
Maxillule (Fig 9g)mdashCoxal endite with15ndash22 marginal setae and 0 or 1 submarginal
Fig 8 Callichirus islagrande ZIV stage a maxilliped 3 b pereiopod 1 c pereiopod 2 d pereiopod 3 e pe-reiopod 4 f pereiopod 5 g pleopods from first abdominal somite h pleopods from fourth abdominal somite i tel-son Scales = 02 mm
112 JOURNAL OF CRUSTACEAN BIOLOGY VOL 20 NO 1 2000
seta basal endite with 3ndash5 plumose setae14ndash19 dentate spines 0ndash2 plumose setae oninner margin endopod reduced unsegmentedwith 0ndash8 setae
Maxilla (Fig 9h)mdashCoxal endite trilobed(17ndash21)+(7ndash10)+(6ndash9) setae basal enditebilobed 8ndash10 setae on proximal lobe 8ndash15setae on distal lobe endopod reduced with
Fig 9 Callichirus islagrande ZV stage a one example of the antenna b one example of pleopods Decapodidstage c antennule d antenna e right mandible f left mandible g maxillule h maxilla (plumose condition notshown) i maxilliped 1 j maxilliped 2 Scales = 02 mm
STRASSER AND FELDER LARVAL DEVELOPMENT IN CALLICHIRUS ISLAGRANDE 113
5ndash11 setae scaphognathite with 37ndash45 se-tae all setae plumose
Maxilliped 1 (Fig 9i)mdashCoxa with 8ndash12 se-tae basis highly setose with 27ndash39 setae en-dopod reduced with 0ndash11 setae exopod un-segmented 6ndash14 setae bilobed epipod with-out setae
Maxilliped 2 (Fig 9j)mdashCoxa partially fusedto basis with 1ndash4 setae basis with 3ndash5 se-
tae endopod 4-segmented (16ndash23) (0 or 1)(6ndash11) (4ndash8) setae exopod with 0ndash8 setaeepipod present but without setae
Maxilliped 3 (Fig 10a)mdashCoxa partiallyfused to basis with 0ndash3 setae basis with 1ndash4setae endopod 5-segmented (11ndash19) (9ndash15)(7ndash11) (8ndash21) (5ndash11) setae exopod usuallyreduced and without setae occasionally with2ndash9 setae epipod present
Fig 10 Callichirus islagrande decapodid stage a maxilliped 3 b pereiopod 1 c pereiopod 2 d pereiopod 3e pereiopod 4 f pereiopod 5 g pleopods from first abdominal somite h pleopods from fourth abdominal somitei telson Scales = 02 mm
114 JOURNAL OF CRUSTACEAN BIOLOGY VOL 20 NO 1 2000
Pereiopod 1 (Fig 10b)mdashChelate biramous7-segmented coxa with 0ndash6 setae basis with2ndash5 setae ischium with 4ndash7 setae and 0ndash3spines merus with 5ndash8 setae carpus with10ndash14 setae propodus with 27ndash37 setae and3ndash4 spines on inner margin of immovable fin-ger dactylus with 21ndash33 setae exopod usu-ally reduced and without setae occasionallywith 4ndash7 setae
Pereiopod 2 (Fig 10c)mdashChelate biramous7-segmented coxa with 2ndash5 setae basis with1ndash5 setae ischium with 4ndash10 setae meruswith 15ndash20 setae carpus with 8ndash12 setaepropodus with 30ndash40 setae and 1ndash4 spineson inner margin of immovable finger dacty-lus with 21ndash30 setae exopod usually reduced0ndash7 setae
Pereiopod 3 (Fig 10d)mdashBiramous 7-seg-mented coxa with 2ndash5 setae basis with 1ndash3setae ischium with 5ndash8 setae merus with6ndash9 setae carpus with 7ndash11 setae propodusbladelike with 25ndash34 setae 1 spine on innerdistal margin dactylus with 15ndash21 setae exo-pod usually reduced 0ndash5 setae
Pereiopod 4 (Fig 10e)mdashBiramous 7-seg-mented coxa with 2ndash7 setae basis with 1ndash4setae ischium with 3ndash8 setae merus with5ndash11 setae carpus with 4ndash9 setae propoduswith 27ndash38 setae 1 spine on inner distal mar-gin dactylus with 15ndash19 setae exopod usu-ally reduced with 0ndash2 setae
Pereiopod 5 (Fig 10f )mdashCheliform unira-mous 7-segmented coxa with 2ndash6 setae ba-sis with l or 2 setae ischium with 2ndash5 setaemerus with 5ndash9 setae carpus with 4ndash7 se-tae propodus with 22ndash30 setae 1 spine onimmovable finger dactylus with 6ndash10 setaeexopod absent
Pleopods (somite 2) (Fig 10g)mdashBiramouscoxa partially fused to basis without setaebasis with 0ndash3 setae endopod with 3ndash5hooked setae on appendix interna 10ndash17 mar-ginal plumose setae 0ndash2 submarginal nakedsetae exopod with 21ndash27 marginal plumosesetae and 0ndash1 submarginal naked seta
Pleopods (somites 3ndash5) (Fig 10h)mdashBiramouscoxa without setae basis with 3ndash6 marginalplumose setae endopod with 3ndash6 hooked se-tae on appendix interna 10ndash18 marginalplumose setae and 0ndash3 naked submarginal se-tae exopod with 20ndash29 marginal plumose se-tae and 0 or 1 submarginal naked seta
Telson and Uropods (Fig 10i)mdashWith 24ndash36processes on posterior margin median pro-cess usually absent or reduced uropods bi-ramous endopod with 19ndash25 plumose setaeexopod with 35ndash58 plumose setae protopodwith 3ndash5 setae
DISCUSSION
As in other species of Callichirus (see Ro-drigues 1976 Aste and Retamal 1983Strasser and Felder in press b) larval devel-opment of C islagrande included four or fivezoeal stages before metamorphosis to the de-capodid ZI larvae may be identified by thepresence of an elongate rounded rostrumsessile eyes and a lack of pereiopodspleopods or uropods (Fig 1a) At ZII lar-vae acquired biramous buds of pereiopods 1and 2 eyes were separated from the carapaceand the rostrum became flattened and serratedlaterally (Fig 1b) Buds of pleopods on ab-dominal segments 2ndash5 pereiopods 3ndash5 anduropods were developed at ZIII (Fig 1c) Inaddition to further development of structuresmentioned thus far ZIV larvae could be dis-tinguished by presence of a distal spine on theexopod and setae on the endopod of each uro-pod (Fig 1d) When it occurred ZV typicallyresembled ZIV but varied widely in mor-phology Appendages at ZV resembled thoseof ZIV the decapodid stage or some grada-tion between the two (see Figs 9a b for ex-amples) The decapodid stage was similar toother species of Callichirus in the reductionof larval characters such as the elongate ros-trum dorsal spines on the abdomen and ex-opods of the pereiopods (Fig le) Acquisitionof fan-like pleopods marked the transition toa primarily abdominal mode of swimming atthis stage
In the Florida Atlantic population of C ma-jor both salinity and presence of putative set-tlement cues at ZIV impacted the observednumber of zoeal stages and the duration ofZIV (Strasser and Felder in preparation1999c) In C islagrande the percentage oflarvae that passed through five versus fourzoeal stages varied (9ndash89) between parentalfemales but was not shown to depend onsalinity or presence of various putative set-tlement cues (Strasser and Felder 1998 inpreparation) Larvae that molted from ZIVto ZV were usually normal morphologicallyand had mortality similar to those that molteddirectly from ZIV to D In contrast larvae of
STRASSER AND FELDER LARVAL DEVELOPMENT IN CALLICHIRUS ISLAGRANDE 115
the Florida Atlantic C major that molted toZV had a higher incidence of mortality anddeformity at D than did those that had by-passed ZV (Strasser and Felder 1999a c)
The number of zoeal stages appears to beless variable in Callichirus garthi and theGulf of Mexico population of C major thanin congeneric populations studied to dateCallichirus garthi was reported to have fivezoeal stages and a long larval duration reach-ing the decapodid stage 48 days after hatch-ing (Aste and Retamal 1983) Developmentof the Gulf population of C major wasshorter in duration (decapodid reached about10 days after hatching) and typically includedfour zoeal stages (Strasser and Felder 1999a)animals rarely passed through three or fivezoeal stages before the molt to the decapodidAlthough the number of stages is reducedfrom that observed in C garthi developmentin the Gulf population of C major cannot beclassified as abbreviated There are species inboth the Callianassinae (Callianassa slkraussi Stebbing C sl kewalramanii and Csl tyrrhena (Petagna)) and Callichirinae(Sergio mirim Lepidophthalmus sinuensisand L louisianensis) that have only two zoealstages before the molt to the decapodid(Forbes 1973 Sankolli and Shenoy 1975Rodrigues 1984 Thessalou-Legaki 1990Nates et al 1997)
Compared to other species of the genusCallichirus C islagrande is morphologicallycloser to C garthi (from Chile) than to theGulf of Mexico population of C major whichoverlaps in distribution with C islagrande Inthe zoeal phase of C islagrande and Cgarthi the telson is concave and the outer-most process on the ventral margin is articu-lated In both C major and Callichirus sp theposterior margin of the telson is straight andthe outermost process is unarticulated duringthe zoeal stages (Rodrigues 1976 Strasserand Felder 1999a) There are also differencesin spination of the abdomen In zoeae of Cislagrande and C garthi the dorsal spine ofabdominal segment 3 is not longer than thosefound on segments 4 and 5 (Aste and Reta-mal 1983) In contrast the dorsal spine of ab-dominal segment 3 is approximately twice aslong as the dorsal spines on segments 4 and5 in early zoeal stages of C major and Cal-lichirus sp (Rodrigues 1976 Aste and Re-tamal 1983 Strasser and Felder 1999a) Thegreatest difference between these groups is
the number of pleopod pairs at zoeal and de-capodid stages Callichirus islagrande and Cgarthi have pleopods on abdominal segments2ndash5 while pleopods are present only on ab-dominal segments 3ndash5 in C major and Cal-lichirus sp (Rodrigues 1976 Aste and Re-tamal 1983 Strasser and Felder 1999a) Al-though Callichirus islagrande and C garthiare similar morphologically there are differ-ences between these two species Callichirusislagrande as well as C major and Cal-lichirus sp develops pleopod buds andchelate pereiopods at ZIII whereas C garthidoes not acquire these characters until ZIV
There are few callianassid species forwhich the larval development has been de-scribed (Sankolli and Shenoy 1975 Ro-drigues 1976 1984 Aste and Retamal 19831984 Konishi et al 1990 Nates et al 1997Strasser and Felder 1999a) all of which aremembers of the Callichirinae and Cal-lianassinae Among species there is a greatdeal of overlap in larval characters makingit difficult to draw distinctions between thesesubfamilies At present information on lar-val development of species in the Callichiri-nae is limited to larval descriptions of Cal-lichirus major C garthi Callichirus sp Cislagrande Sergio mirim Lepidophthalmuslouisianensis and L sinuensis (see Ro-drigues 1976 1984 Aste and Retamal 1983Nates et al 1997 Strasser and Felder1999a) As mentioned above species of Cal-lichirus have been shown to have four to fivezoeal stages while the other members exhibitabbreviated development that consists of onlytwo zoeal stages The most obvious differencebetween larvae of the species of Callichirinaewith abbreviated development is the numberof pleopod pairs Larvae of S mirim havepleopods on abdominal segments 3ndash5whereas L louisianensis and L sinuensishave pleopods on segments 2ndash5 (Rodrigues1984 Nates et al 1997)
Ghost shrimps of the genus Callichirus maybe distinguished from other genera of the Cal-lianassidae by the cuticular pattern on the dor-sal surface of the abdomen and by the strap-like shape of the uropodal endopods (Manningand Felder 1991) Whereas these charactersmay be used to distinguish adults of the genusCallichirus they are not present at zoeal anddecapodid stages Larvae may be distin-guished from all other callianassid genera byhaving four to five zoeal stages and numer-
116 JOURNAL OF CRUSTACEAN BIOLOGY VOL 20 NO 1 2000
ous processes (12ndash25+1+12ndash25) on the pos-terior margin of the telson during the earlyzoeal stages Larvae of Callichirus may alsodiffer from other callianassids with extendeddevelopment by the retention of the anomu-ran hair as the second process on the poste-rior margin of the telson in the late zoealstages Whereas this process is converted toa spine in Neotrypaea uncinata at ZV and Cal-lianassa sl petalura at ZIV the anomuranhair is retained at ZIV in Callichirus majorand C islagrande (see Rodrigues 1976Strasser and Felder 1999a) Although notmentioned specifically in the descriptions theanomuran hair appears to be present in illus-trations of Callichirus sp at ZIII and Cgarthii at ZV (Rodrigues 1976 fig 35 Asteand Retamal 1983 fig 8Q) A bilobed epi-pod on the first maxilliped of ZI is consistentlyfound in Callichirus but is also present at ZIin Lepidophthalmus sinuensis (see Nates etal 1997) Other callianassids acquire thischaracter at ZII (L louisianensis Nates et al1997 Callianassa sl kewlramanii Sankolliand Shenoy 1975) at ZV (Neotrypaea unci-nata Aste and Retamal 1984) or at the de-capodid stage (Sergio mirim Rodrigues 1984C sl petalura Konishi et al 1990) Thusthe number of zoeal stages and setation of thetelson may serve as the best characters to dis-tinguish larvae of Callichirus from other gen-era However since larval descriptions arelacking for many callianassid species it is un-known if these characters (in combination)will prove to be diagnostic until descriptionsfor other species are published
ACKNOWLEDGEMENTS
We thank Robert Bourgeois Barbara Hasek JimMaKinster Sergio Nates and Kurt Strasser who pro-vided assistance in the field and laboratory and MaryE Rice Director of the Smithsonian Marine StationLink Port Florida who facilitated access to station fa-cilities We also thank R Bauer R Jaeger R Lemaitreand G Watson for their early comments on this manu-script and Raymond B Manning who facilitated fund-ing and gave advice on collecting sites Direct supportfor this study was provided to Karen M Strasser by adoctoral fellowship from the University of Louisianaa graduate student research grant from the LUMCON(Louisiana Universities Marine Consortium) Founda-tion and a summer research award from the Smith-sonian Marine Station Additional support for field andlab activities was provided to D L Felder under Loui-siana Sea Grant College Program Grant RCFB-21 USFish and Wildlife Service Cooperative Agreement 14-16-0009-89-96 (Task Order No 6) US Department of En-ergy grant DE-FG02-97ER12220 and several small proj-ect grants from the Smithsonian Marine Station Views
expressed herein are those of the authors and do not nec-essarily reflect views of supporting government agenciesor subagencies mentioned above This is contributionNumber 466 of the Smithsonian Marine Station and con-tribution Number 67 of the ULL Laboratory for Crus-tacean Research All experiments conducted in this studycomply with current applicable state and federal laws
LITERATURE CITED
Aste A and M Retamal 1983 Desarrollo larval deCallianassa garthi Retamal 1975 bajo condiciones delaboratoriomdashCiencia y Tecnologiacutea del Mar CONA 75ndash26
mdashmdashmdash and mdashmdashmdash 1984 Desarrollo larval de Cal-lianassa uncinata H Milne Edwards1837 (DecapodaCallianassidae) bajo condiciones de laboratoriomdashGayana Zooloacutegica 48 41ndash56
Felder D L 1978 Osmotic and ionic regulation in sev-eral western Atlantic Callianassidae (Crustacea De-capoda Thalassinidea)mdashBiological Bulletin 154409ndash429
mdashmdashmdash and R B Griffis 1994 Dominant infaunalcommunities at risk in shoreline habitats burrowingthalassinid CrustaceamdashOCS Study number MMS94ndash0007 United States Department of the InteriorMinerals Management Service Gulf of Mexico OCSRegional Office New Orleans Louisiana Pp 1ndash87
Forbes A T 1973 An unusual abbreviated larval lifehistory in the estuarine burrowing prawn Callianassakraussi (Crustacea Decapoda Thalassinidea)mdashMa-rine Biology 22 361ndash365
Konishi K 1989 Larval development of the mudshrimp Upogebia (Upogebia) major (de Haan) (Crus-tacea Thalassinidea Upogebiidae) under laboratoryconditions with comments on larval characters of tha-lassinid familiesmdashBulletin of the National ResearchInstitute of Aquaculture 15 1ndash17
mdashmdashmdash R Quintana and Y Fukuda 1990 A completedescription of larval stages of the ghost shrimp Cal-lianassa petalura Stimpson (Crustacea ThalassinideaCallianassidae) under laboratory conditionsmdashBulletinof the National Research Institute of Aquaculture 1727ndash49
Manning R B and D L Felder 1986 The status ofthe callianassid genus Callichirus Stimpson 1866(Crustacea Decapoda Thalassinidea)mdashProceedingsof the Biological Society of Washington 99 437ndash443
mdashmdashmdash and mdashmdashmdash 1991 Revision of the AmericanCallianassidae (Crustacea Decapoda Thalassinidea)mdashProceedings of the Biological Society of Washington104 764ndash794
Nates S F D L Felder and R Lemaitre 1997 Com-parative larval development in two species of the bur-rowing ghost shrimp genus Lepidophthalmus (Decap-oda Callianassidae)mdashJournal of Crustacean Biology17 497ndash519
Rodrigues S de A 1976 Sobre a reproduccedilatildeo embri-ologia e desenvolvimento larval de Callichirus majorSay 1818 (Crustacea Decapoda Thalassinidea)mdashBo-letim de Zoologia Universidade de Satildeo Paulo 185ndash104
mdashmdashmdash 1984 Desenvolvimento poacutes-embrionaacuterio deCallichirus mirim (Rodrigues 1971) obtido emcondiccedilotildees artificiais (Crustacea Decapoda Tha-lassinidea)mdashBoletim de Zoologia Universidade deSatildeo Paulo 8 239ndash256
STRASSER AND FELDER LARVAL DEVELOPMENT IN CALLICHIRUS ISLAGRANDE 117
Sankolli K N and S Shenoy 1975 Larval develop-ment of mud shrimp Callianassa (Callichirus) kewal-ramanii Sankolli in the laboratory (Crustacea Decap-oda)mdashBulletin of the Department of Marine ScienceUniversity of Cochin 4 704ndash720
Staton J L and D L Felder 1995 Genetic variationin populations of the ghost shrimp genus Callichirus(Crustacea Decapoda Thalassinoidea) in the westernAtlantic and Gulf of MexicomdashBulletin of Marine Sci-ence 56 523ndash536
Strasser K M and D L Felder 1998 Settlement cuesin successive developmental stages of the ghost shrimpCallichirus major and C islagrande (Crustacea De-capoda Thalassinidea)mdashMarine Biology
mdashmdashmdash and mdashmdashmdash 1999a Larval development of twopopulations of the ghost shrimp Callichirus major (De-capoda Thalassinidea)mdashJournal of Crustacean Biology
mdashmdashmdash and mdashmdashmdash 1999b Sand as a stimulus for set-tlement in the ghost shrimp Callichirus major (Say)and C islagrande (Schmitt) (Crustacea DecapodaThalassinidea)mdashJournal of Experimental Marine Bi-ology and Ecology 239 211ndash222
mdashmdashmdash and mdashmdashmdash 1999c Settlement cues in theFlorida Atlantic population of the ghost shrimp Cal-lichirus major (Crustacea Decapoda Thalassinidea)mdashMarine Ecology Progress Series
Thessalou-Legaki M 1990 Advanced larval develop-ment of Callianassa tyrrhena (Decapoda Tha-lassinidea) and the effect of environmental factorsmdashJournal of Crustacean Biology 10 659ndash666
RECEIVED 19 September 1998ACCEPTED 3 December 1998

108 JOURNAL OF CRUSTACEAN BIOLOGY VOL 20 NO 1 2000
Pereiopod 2 (Fig 6c)mdashChelate and biramouscoxa without setae basis with 0 or 1 seta en-dopod 4-segmented 0 (0ndash2) (0ndash4) 2 setaeexopod with 5ndash7 setae on distal margin
Pereiopods 3ndash5 (Fig 6d e f )mdashWithout setae
Pleopods (Figs 1c 6g)mdashBilobed buds onabdominal segments 2ndash5 first pair smallerthan others
Telson and Uropods (Fig 6h)mdashNo changein arrangement of posterior telsonal processesuropods biramous endopod without setae
Fig 6 Callichirus islagrande ZIII stage a maxilliped 3 b pereiopod 1 c pereiopod 2 d pereiopod 3 e pe-reiopod 4 f pereiopod 5 g pleopods from fourth abdominal somite h telson Scales = 02 mm
STRASSER AND FELDER LARVAL DEVELOPMENT IN CALLICHIRUS ISLAGRANDE 109
exopod with 10 or 11 marginal plumose se-tae ventral spine near base of protopod
Zoea IVFigs 2d 7andashh 8andashi
SizemdashCL = 192 plusmn 009 mm range 169ndash215mm TL = 482 plusmn 025 mm range 424ndash536mm
Carapace (Fig 2d)mdashNo marked changefrom ZIII
Abdomen (Fig 2d)mdashNo marked change fromZIII
Antennule (Fig 7a)mdashBiramous peduncle2-segmented with 1+(1ndash4) short setae onouter margin and 0 or 1 plumose seta on in-ner margin of proximal segment distal seg-ment with 3ndash5 plumose setae on inner mar-gin 1 plumose seta between rami and 0ndash3short setae on outer margin endopod with 1or 2 plumose setae exopod with 6 or 7 ter-minal and 2 subterminal aesthetascs
Antenna (Fig 7b)mdashProtopod with 1 distalspine on outer margin endopod with 0ndash2 se-tae scaphocerite armed with 1 strong disto-lateral spine 12ndash14 plumose setae on innermargin and 1 plumose seta on outer margin
Mandible (Fig 7c d)mdashIncisor process with8 teeth on right mandible 6ndash8 teeth on leftmandible molar process denticulate on bothmandibles palps present
Maxillule (Fig 7e)mdashCoxal endite with12ndash14 marginal plumose setae basal enditewith 3 plumose setae 5 or 6 large dentatespines and 5 or 6 smaller dentate spinesinner margin with 1 plumose seta endopod3-segmented distinction between segmentssometimes obscure 2 2 (3 or 4) setae pro-topod without setation
Maxilla (Fig 7f )mdashCoxal endite bilobed14ndash19 setae on proximal lobe 5 setae on dis-tal lobe basal endite bilobed 6ndash10 setae onproximal lobe 8 or 9 setae on distal lobeendopod with 5 lobes 3+(2 or 3)+2+2+3 se-tae scaphognathite with 27ndash32 setae all se-tae plumose
Maxilliped 1 (Fig 7g)mdashCoxa with 2ndash8 se-tae basis with 16ndash19 marginal setae endo-pod 4-segmented (3 or 4) 2 2 4 setae exo-
pod 2-segmented proximal segment withoutsetae distal segment with 6 or 7 setae on dis-tal margin bilobed epipod without setae allsetae plumose
Maxilliped 2 (Fig 7h)mdashCoxa with 0ndash3 se-tae basis with 4ndash6 setae endopod 4-seg-mented (5ndash8) 1 (3ndash5) (4 or 5) setae exo-pod unsegmented 7 or 8 setae on distal mar-gin all setae plumose
Maxilliped 3 (Fig 8a)mdashCoxa with 0 or 1seta basis with 2ndash4 setae endopod 4-seg-mented (l or 2) 2 (3ndash5) (3 or 4) setae exo-pod unsegmented 7 or 8 setae on distal mar-gin all setae plumose
Pereiopod 1 (Fig 8b)mdashBiramous cheliformcoxa without setae basis with 0ndash2 setae en-dopod 4-segmented (0 or 1) (1 or 2) (2ndash4)(1 or 2) setae exopod with 5ndash7 setae on dis-tal margin
Pereiopod 2 (Fig 8c)mdashBiramous cheliformcoxa without setae basis with 0 or 1 setaendopod 4-segmented 0 (1 or 2) (3ndash5) (2or 3) setae exopod with 5ndash7 setae on distalmargin
Pereiopod 3 (Fig 8d)mdashBiramous coxa andbasis without setae endopod 4-segmented0 (0ndash2) (3 or 4) 2 setae propodus blade-like exopod with 4 or 5 terminal setae
Pereiopod 4 (Fig 8e)mdashBiramous coxa andbasis without setae endopod 4-segmented (0or 1) (0ndash2) (2ndash4) 2 setae exopod with 0ndash2setae
Pereiopod 5 (Fig 8f )mdashUniramous 2 setaeon terminal segment 4 or 5 setae on adja-cent segment
Pleopods (Fig 8g h)mdashLarger than at ZIIIstill without setae
Telson and Uropods (Fig 8i)mdashTelson trian-gular narrow anteriorly no anal spine pos-terior margin with (12ndash15)+1+(13 or 14)processes arranged as (I II III IV III II I)(I) outermost process a plumodenticulate seta(II) second process ldquoanomuran hairrdquo (III) 10ndash13plumodenticulate setae and (IV) naked spineuropods biramous endopod with 7ndash11 marginalplumose setae exopod with 1 naked distolat-eral spine 12ndash14 marginal plumose setae
110 JOURNAL OF CRUSTACEAN BIOLOGY VOL 20 NO 1 2000
Decapodid (first postlarva)Figs 2e 9cndashj l0andashi
SizemdashCL = 133 plusmn 002 mm range129ndash138 mm TL = 378 plusmn 011 mm range361ndash405 mm
Carapace (Fig 2e)mdashShorter than abdomenno anterolateral serrations with pterygostomial
projections rostrum reduced without serra-tions sometimes bending down between eyes
Abdomen (Fig 2e)mdashSomites broad and flat-tened posterior projections rounded with 0ndash7setae somite 2 rectangular somites 3ndash5 oval
Antennule (Fig 9c)mdashPeduncle 3-segmentedproximal segment with 17ndash24 setae 13ndash15 of
Fig 7 Callichirus islagrande ZIV stage a antennule b antenna c right mandible d left mandible e maxil-lule f maxilla (plumose condition not shown) g maxilliped 1 h maxilliped 2 All setae on the maxilla were plumoseScales = 02 mm
STRASSER AND FELDER LARVAL DEVELOPMENT IN CALLICHIRUS ISLAGRANDE 111
these associated with statocyst intermediatesegment with 6ndash10 setae distal segment of pe-duncle with 2 vertical rows of setae one with4 or 5 setae other with 2ndash4 setae 1ndash3 addi-tional setae on outer margin and 3ndash5 short se-tae on distal margin endopod 4ndash6 segmented(1ndash3) (0 or 1) (1ndash4) (1ndash4) (3ndash6) (3ndash6) se-tae exopod 3ndash5-segmented 0 (0ndash4) (0ndash4)(1ndash4) setae +2 asthetascs (5ndash8) asthetascs ()
Antenna (Fig 9d)mdashPeduncle 2-segmentedproximal segment with 0 or 1 seta distal seg-ment with 1ndash3 setae on inner margin spinepresent in some flagellum ranging from 14ndash18
segments (0 or 1) (2ndash4) variable region(1ndash3 articles with 0ndash2 setae) (0ndash4) (0ndash2)(2ndash5) (1ndash4) (3ndash6) (0ndash3) (2ndash5) (3ndash6)(0ndash3) (3ndash6) (0 or 1) (1ndash7) (4ndash9) setae exo-pod reduced to small lobe with 0ndash5 setae
Mandible (Fig 9e f )mdashBoth mandibles sim-ilar incisor process with 5ndash8 teeth molarprocess smooth or with small ridges palp3-segmented 0 (0 or 1) (8ndash12) setae
Maxillule (Fig 9g)mdashCoxal endite with15ndash22 marginal setae and 0 or 1 submarginal
Fig 8 Callichirus islagrande ZIV stage a maxilliped 3 b pereiopod 1 c pereiopod 2 d pereiopod 3 e pe-reiopod 4 f pereiopod 5 g pleopods from first abdominal somite h pleopods from fourth abdominal somite i tel-son Scales = 02 mm
112 JOURNAL OF CRUSTACEAN BIOLOGY VOL 20 NO 1 2000
seta basal endite with 3ndash5 plumose setae14ndash19 dentate spines 0ndash2 plumose setae oninner margin endopod reduced unsegmentedwith 0ndash8 setae
Maxilla (Fig 9h)mdashCoxal endite trilobed(17ndash21)+(7ndash10)+(6ndash9) setae basal enditebilobed 8ndash10 setae on proximal lobe 8ndash15setae on distal lobe endopod reduced with
Fig 9 Callichirus islagrande ZV stage a one example of the antenna b one example of pleopods Decapodidstage c antennule d antenna e right mandible f left mandible g maxillule h maxilla (plumose condition notshown) i maxilliped 1 j maxilliped 2 Scales = 02 mm
STRASSER AND FELDER LARVAL DEVELOPMENT IN CALLICHIRUS ISLAGRANDE 113
5ndash11 setae scaphognathite with 37ndash45 se-tae all setae plumose
Maxilliped 1 (Fig 9i)mdashCoxa with 8ndash12 se-tae basis highly setose with 27ndash39 setae en-dopod reduced with 0ndash11 setae exopod un-segmented 6ndash14 setae bilobed epipod with-out setae
Maxilliped 2 (Fig 9j)mdashCoxa partially fusedto basis with 1ndash4 setae basis with 3ndash5 se-
tae endopod 4-segmented (16ndash23) (0 or 1)(6ndash11) (4ndash8) setae exopod with 0ndash8 setaeepipod present but without setae
Maxilliped 3 (Fig 10a)mdashCoxa partiallyfused to basis with 0ndash3 setae basis with 1ndash4setae endopod 5-segmented (11ndash19) (9ndash15)(7ndash11) (8ndash21) (5ndash11) setae exopod usuallyreduced and without setae occasionally with2ndash9 setae epipod present
Fig 10 Callichirus islagrande decapodid stage a maxilliped 3 b pereiopod 1 c pereiopod 2 d pereiopod 3e pereiopod 4 f pereiopod 5 g pleopods from first abdominal somite h pleopods from fourth abdominal somitei telson Scales = 02 mm
114 JOURNAL OF CRUSTACEAN BIOLOGY VOL 20 NO 1 2000
Pereiopod 1 (Fig 10b)mdashChelate biramous7-segmented coxa with 0ndash6 setae basis with2ndash5 setae ischium with 4ndash7 setae and 0ndash3spines merus with 5ndash8 setae carpus with10ndash14 setae propodus with 27ndash37 setae and3ndash4 spines on inner margin of immovable fin-ger dactylus with 21ndash33 setae exopod usu-ally reduced and without setae occasionallywith 4ndash7 setae
Pereiopod 2 (Fig 10c)mdashChelate biramous7-segmented coxa with 2ndash5 setae basis with1ndash5 setae ischium with 4ndash10 setae meruswith 15ndash20 setae carpus with 8ndash12 setaepropodus with 30ndash40 setae and 1ndash4 spineson inner margin of immovable finger dacty-lus with 21ndash30 setae exopod usually reduced0ndash7 setae
Pereiopod 3 (Fig 10d)mdashBiramous 7-seg-mented coxa with 2ndash5 setae basis with 1ndash3setae ischium with 5ndash8 setae merus with6ndash9 setae carpus with 7ndash11 setae propodusbladelike with 25ndash34 setae 1 spine on innerdistal margin dactylus with 15ndash21 setae exo-pod usually reduced 0ndash5 setae
Pereiopod 4 (Fig 10e)mdashBiramous 7-seg-mented coxa with 2ndash7 setae basis with 1ndash4setae ischium with 3ndash8 setae merus with5ndash11 setae carpus with 4ndash9 setae propoduswith 27ndash38 setae 1 spine on inner distal mar-gin dactylus with 15ndash19 setae exopod usu-ally reduced with 0ndash2 setae
Pereiopod 5 (Fig 10f )mdashCheliform unira-mous 7-segmented coxa with 2ndash6 setae ba-sis with l or 2 setae ischium with 2ndash5 setaemerus with 5ndash9 setae carpus with 4ndash7 se-tae propodus with 22ndash30 setae 1 spine onimmovable finger dactylus with 6ndash10 setaeexopod absent
Pleopods (somite 2) (Fig 10g)mdashBiramouscoxa partially fused to basis without setaebasis with 0ndash3 setae endopod with 3ndash5hooked setae on appendix interna 10ndash17 mar-ginal plumose setae 0ndash2 submarginal nakedsetae exopod with 21ndash27 marginal plumosesetae and 0ndash1 submarginal naked seta
Pleopods (somites 3ndash5) (Fig 10h)mdashBiramouscoxa without setae basis with 3ndash6 marginalplumose setae endopod with 3ndash6 hooked se-tae on appendix interna 10ndash18 marginalplumose setae and 0ndash3 naked submarginal se-tae exopod with 20ndash29 marginal plumose se-tae and 0 or 1 submarginal naked seta
Telson and Uropods (Fig 10i)mdashWith 24ndash36processes on posterior margin median pro-cess usually absent or reduced uropods bi-ramous endopod with 19ndash25 plumose setaeexopod with 35ndash58 plumose setae protopodwith 3ndash5 setae
DISCUSSION
As in other species of Callichirus (see Ro-drigues 1976 Aste and Retamal 1983Strasser and Felder in press b) larval devel-opment of C islagrande included four or fivezoeal stages before metamorphosis to the de-capodid ZI larvae may be identified by thepresence of an elongate rounded rostrumsessile eyes and a lack of pereiopodspleopods or uropods (Fig 1a) At ZII lar-vae acquired biramous buds of pereiopods 1and 2 eyes were separated from the carapaceand the rostrum became flattened and serratedlaterally (Fig 1b) Buds of pleopods on ab-dominal segments 2ndash5 pereiopods 3ndash5 anduropods were developed at ZIII (Fig 1c) Inaddition to further development of structuresmentioned thus far ZIV larvae could be dis-tinguished by presence of a distal spine on theexopod and setae on the endopod of each uro-pod (Fig 1d) When it occurred ZV typicallyresembled ZIV but varied widely in mor-phology Appendages at ZV resembled thoseof ZIV the decapodid stage or some grada-tion between the two (see Figs 9a b for ex-amples) The decapodid stage was similar toother species of Callichirus in the reductionof larval characters such as the elongate ros-trum dorsal spines on the abdomen and ex-opods of the pereiopods (Fig le) Acquisitionof fan-like pleopods marked the transition toa primarily abdominal mode of swimming atthis stage
In the Florida Atlantic population of C ma-jor both salinity and presence of putative set-tlement cues at ZIV impacted the observednumber of zoeal stages and the duration ofZIV (Strasser and Felder in preparation1999c) In C islagrande the percentage oflarvae that passed through five versus fourzoeal stages varied (9ndash89) between parentalfemales but was not shown to depend onsalinity or presence of various putative set-tlement cues (Strasser and Felder 1998 inpreparation) Larvae that molted from ZIVto ZV were usually normal morphologicallyand had mortality similar to those that molteddirectly from ZIV to D In contrast larvae of
STRASSER AND FELDER LARVAL DEVELOPMENT IN CALLICHIRUS ISLAGRANDE 115
the Florida Atlantic C major that molted toZV had a higher incidence of mortality anddeformity at D than did those that had by-passed ZV (Strasser and Felder 1999a c)
The number of zoeal stages appears to beless variable in Callichirus garthi and theGulf of Mexico population of C major thanin congeneric populations studied to dateCallichirus garthi was reported to have fivezoeal stages and a long larval duration reach-ing the decapodid stage 48 days after hatch-ing (Aste and Retamal 1983) Developmentof the Gulf population of C major wasshorter in duration (decapodid reached about10 days after hatching) and typically includedfour zoeal stages (Strasser and Felder 1999a)animals rarely passed through three or fivezoeal stages before the molt to the decapodidAlthough the number of stages is reducedfrom that observed in C garthi developmentin the Gulf population of C major cannot beclassified as abbreviated There are species inboth the Callianassinae (Callianassa slkraussi Stebbing C sl kewalramanii and Csl tyrrhena (Petagna)) and Callichirinae(Sergio mirim Lepidophthalmus sinuensisand L louisianensis) that have only two zoealstages before the molt to the decapodid(Forbes 1973 Sankolli and Shenoy 1975Rodrigues 1984 Thessalou-Legaki 1990Nates et al 1997)
Compared to other species of the genusCallichirus C islagrande is morphologicallycloser to C garthi (from Chile) than to theGulf of Mexico population of C major whichoverlaps in distribution with C islagrande Inthe zoeal phase of C islagrande and Cgarthi the telson is concave and the outer-most process on the ventral margin is articu-lated In both C major and Callichirus sp theposterior margin of the telson is straight andthe outermost process is unarticulated duringthe zoeal stages (Rodrigues 1976 Strasserand Felder 1999a) There are also differencesin spination of the abdomen In zoeae of Cislagrande and C garthi the dorsal spine ofabdominal segment 3 is not longer than thosefound on segments 4 and 5 (Aste and Reta-mal 1983) In contrast the dorsal spine of ab-dominal segment 3 is approximately twice aslong as the dorsal spines on segments 4 and5 in early zoeal stages of C major and Cal-lichirus sp (Rodrigues 1976 Aste and Re-tamal 1983 Strasser and Felder 1999a) Thegreatest difference between these groups is
the number of pleopod pairs at zoeal and de-capodid stages Callichirus islagrande and Cgarthi have pleopods on abdominal segments2ndash5 while pleopods are present only on ab-dominal segments 3ndash5 in C major and Cal-lichirus sp (Rodrigues 1976 Aste and Re-tamal 1983 Strasser and Felder 1999a) Al-though Callichirus islagrande and C garthiare similar morphologically there are differ-ences between these two species Callichirusislagrande as well as C major and Cal-lichirus sp develops pleopod buds andchelate pereiopods at ZIII whereas C garthidoes not acquire these characters until ZIV
There are few callianassid species forwhich the larval development has been de-scribed (Sankolli and Shenoy 1975 Ro-drigues 1976 1984 Aste and Retamal 19831984 Konishi et al 1990 Nates et al 1997Strasser and Felder 1999a) all of which aremembers of the Callichirinae and Cal-lianassinae Among species there is a greatdeal of overlap in larval characters makingit difficult to draw distinctions between thesesubfamilies At present information on lar-val development of species in the Callichiri-nae is limited to larval descriptions of Cal-lichirus major C garthi Callichirus sp Cislagrande Sergio mirim Lepidophthalmuslouisianensis and L sinuensis (see Ro-drigues 1976 1984 Aste and Retamal 1983Nates et al 1997 Strasser and Felder1999a) As mentioned above species of Cal-lichirus have been shown to have four to fivezoeal stages while the other members exhibitabbreviated development that consists of onlytwo zoeal stages The most obvious differencebetween larvae of the species of Callichirinaewith abbreviated development is the numberof pleopod pairs Larvae of S mirim havepleopods on abdominal segments 3ndash5whereas L louisianensis and L sinuensishave pleopods on segments 2ndash5 (Rodrigues1984 Nates et al 1997)
Ghost shrimps of the genus Callichirus maybe distinguished from other genera of the Cal-lianassidae by the cuticular pattern on the dor-sal surface of the abdomen and by the strap-like shape of the uropodal endopods (Manningand Felder 1991) Whereas these charactersmay be used to distinguish adults of the genusCallichirus they are not present at zoeal anddecapodid stages Larvae may be distin-guished from all other callianassid genera byhaving four to five zoeal stages and numer-
116 JOURNAL OF CRUSTACEAN BIOLOGY VOL 20 NO 1 2000
ous processes (12ndash25+1+12ndash25) on the pos-terior margin of the telson during the earlyzoeal stages Larvae of Callichirus may alsodiffer from other callianassids with extendeddevelopment by the retention of the anomu-ran hair as the second process on the poste-rior margin of the telson in the late zoealstages Whereas this process is converted toa spine in Neotrypaea uncinata at ZV and Cal-lianassa sl petalura at ZIV the anomuranhair is retained at ZIV in Callichirus majorand C islagrande (see Rodrigues 1976Strasser and Felder 1999a) Although notmentioned specifically in the descriptions theanomuran hair appears to be present in illus-trations of Callichirus sp at ZIII and Cgarthii at ZV (Rodrigues 1976 fig 35 Asteand Retamal 1983 fig 8Q) A bilobed epi-pod on the first maxilliped of ZI is consistentlyfound in Callichirus but is also present at ZIin Lepidophthalmus sinuensis (see Nates etal 1997) Other callianassids acquire thischaracter at ZII (L louisianensis Nates et al1997 Callianassa sl kewlramanii Sankolliand Shenoy 1975) at ZV (Neotrypaea unci-nata Aste and Retamal 1984) or at the de-capodid stage (Sergio mirim Rodrigues 1984C sl petalura Konishi et al 1990) Thusthe number of zoeal stages and setation of thetelson may serve as the best characters to dis-tinguish larvae of Callichirus from other gen-era However since larval descriptions arelacking for many callianassid species it is un-known if these characters (in combination)will prove to be diagnostic until descriptionsfor other species are published
ACKNOWLEDGEMENTS
We thank Robert Bourgeois Barbara Hasek JimMaKinster Sergio Nates and Kurt Strasser who pro-vided assistance in the field and laboratory and MaryE Rice Director of the Smithsonian Marine StationLink Port Florida who facilitated access to station fa-cilities We also thank R Bauer R Jaeger R Lemaitreand G Watson for their early comments on this manu-script and Raymond B Manning who facilitated fund-ing and gave advice on collecting sites Direct supportfor this study was provided to Karen M Strasser by adoctoral fellowship from the University of Louisianaa graduate student research grant from the LUMCON(Louisiana Universities Marine Consortium) Founda-tion and a summer research award from the Smith-sonian Marine Station Additional support for field andlab activities was provided to D L Felder under Loui-siana Sea Grant College Program Grant RCFB-21 USFish and Wildlife Service Cooperative Agreement 14-16-0009-89-96 (Task Order No 6) US Department of En-ergy grant DE-FG02-97ER12220 and several small proj-ect grants from the Smithsonian Marine Station Views
expressed herein are those of the authors and do not nec-essarily reflect views of supporting government agenciesor subagencies mentioned above This is contributionNumber 466 of the Smithsonian Marine Station and con-tribution Number 67 of the ULL Laboratory for Crus-tacean Research All experiments conducted in this studycomply with current applicable state and federal laws
LITERATURE CITED
Aste A and M Retamal 1983 Desarrollo larval deCallianassa garthi Retamal 1975 bajo condiciones delaboratoriomdashCiencia y Tecnologiacutea del Mar CONA 75ndash26
mdashmdashmdash and mdashmdashmdash 1984 Desarrollo larval de Cal-lianassa uncinata H Milne Edwards1837 (DecapodaCallianassidae) bajo condiciones de laboratoriomdashGayana Zooloacutegica 48 41ndash56
Felder D L 1978 Osmotic and ionic regulation in sev-eral western Atlantic Callianassidae (Crustacea De-capoda Thalassinidea)mdashBiological Bulletin 154409ndash429
mdashmdashmdash and R B Griffis 1994 Dominant infaunalcommunities at risk in shoreline habitats burrowingthalassinid CrustaceamdashOCS Study number MMS94ndash0007 United States Department of the InteriorMinerals Management Service Gulf of Mexico OCSRegional Office New Orleans Louisiana Pp 1ndash87
Forbes A T 1973 An unusual abbreviated larval lifehistory in the estuarine burrowing prawn Callianassakraussi (Crustacea Decapoda Thalassinidea)mdashMa-rine Biology 22 361ndash365
Konishi K 1989 Larval development of the mudshrimp Upogebia (Upogebia) major (de Haan) (Crus-tacea Thalassinidea Upogebiidae) under laboratoryconditions with comments on larval characters of tha-lassinid familiesmdashBulletin of the National ResearchInstitute of Aquaculture 15 1ndash17
mdashmdashmdash R Quintana and Y Fukuda 1990 A completedescription of larval stages of the ghost shrimp Cal-lianassa petalura Stimpson (Crustacea ThalassinideaCallianassidae) under laboratory conditionsmdashBulletinof the National Research Institute of Aquaculture 1727ndash49
Manning R B and D L Felder 1986 The status ofthe callianassid genus Callichirus Stimpson 1866(Crustacea Decapoda Thalassinidea)mdashProceedingsof the Biological Society of Washington 99 437ndash443
mdashmdashmdash and mdashmdashmdash 1991 Revision of the AmericanCallianassidae (Crustacea Decapoda Thalassinidea)mdashProceedings of the Biological Society of Washington104 764ndash794
Nates S F D L Felder and R Lemaitre 1997 Com-parative larval development in two species of the bur-rowing ghost shrimp genus Lepidophthalmus (Decap-oda Callianassidae)mdashJournal of Crustacean Biology17 497ndash519
Rodrigues S de A 1976 Sobre a reproduccedilatildeo embri-ologia e desenvolvimento larval de Callichirus majorSay 1818 (Crustacea Decapoda Thalassinidea)mdashBo-letim de Zoologia Universidade de Satildeo Paulo 185ndash104
mdashmdashmdash 1984 Desenvolvimento poacutes-embrionaacuterio deCallichirus mirim (Rodrigues 1971) obtido emcondiccedilotildees artificiais (Crustacea Decapoda Tha-lassinidea)mdashBoletim de Zoologia Universidade deSatildeo Paulo 8 239ndash256
STRASSER AND FELDER LARVAL DEVELOPMENT IN CALLICHIRUS ISLAGRANDE 117
Sankolli K N and S Shenoy 1975 Larval develop-ment of mud shrimp Callianassa (Callichirus) kewal-ramanii Sankolli in the laboratory (Crustacea Decap-oda)mdashBulletin of the Department of Marine ScienceUniversity of Cochin 4 704ndash720
Staton J L and D L Felder 1995 Genetic variationin populations of the ghost shrimp genus Callichirus(Crustacea Decapoda Thalassinoidea) in the westernAtlantic and Gulf of MexicomdashBulletin of Marine Sci-ence 56 523ndash536
Strasser K M and D L Felder 1998 Settlement cuesin successive developmental stages of the ghost shrimpCallichirus major and C islagrande (Crustacea De-capoda Thalassinidea)mdashMarine Biology
mdashmdashmdash and mdashmdashmdash 1999a Larval development of twopopulations of the ghost shrimp Callichirus major (De-capoda Thalassinidea)mdashJournal of Crustacean Biology
mdashmdashmdash and mdashmdashmdash 1999b Sand as a stimulus for set-tlement in the ghost shrimp Callichirus major (Say)and C islagrande (Schmitt) (Crustacea DecapodaThalassinidea)mdashJournal of Experimental Marine Bi-ology and Ecology 239 211ndash222
mdashmdashmdash and mdashmdashmdash 1999c Settlement cues in theFlorida Atlantic population of the ghost shrimp Cal-lichirus major (Crustacea Decapoda Thalassinidea)mdashMarine Ecology Progress Series
Thessalou-Legaki M 1990 Advanced larval develop-ment of Callianassa tyrrhena (Decapoda Tha-lassinidea) and the effect of environmental factorsmdashJournal of Crustacean Biology 10 659ndash666
RECEIVED 19 September 1998ACCEPTED 3 December 1998

STRASSER AND FELDER LARVAL DEVELOPMENT IN CALLICHIRUS ISLAGRANDE 109
exopod with 10 or 11 marginal plumose se-tae ventral spine near base of protopod
Zoea IVFigs 2d 7andashh 8andashi
SizemdashCL = 192 plusmn 009 mm range 169ndash215mm TL = 482 plusmn 025 mm range 424ndash536mm
Carapace (Fig 2d)mdashNo marked changefrom ZIII
Abdomen (Fig 2d)mdashNo marked change fromZIII
Antennule (Fig 7a)mdashBiramous peduncle2-segmented with 1+(1ndash4) short setae onouter margin and 0 or 1 plumose seta on in-ner margin of proximal segment distal seg-ment with 3ndash5 plumose setae on inner mar-gin 1 plumose seta between rami and 0ndash3short setae on outer margin endopod with 1or 2 plumose setae exopod with 6 or 7 ter-minal and 2 subterminal aesthetascs
Antenna (Fig 7b)mdashProtopod with 1 distalspine on outer margin endopod with 0ndash2 se-tae scaphocerite armed with 1 strong disto-lateral spine 12ndash14 plumose setae on innermargin and 1 plumose seta on outer margin
Mandible (Fig 7c d)mdashIncisor process with8 teeth on right mandible 6ndash8 teeth on leftmandible molar process denticulate on bothmandibles palps present
Maxillule (Fig 7e)mdashCoxal endite with12ndash14 marginal plumose setae basal enditewith 3 plumose setae 5 or 6 large dentatespines and 5 or 6 smaller dentate spinesinner margin with 1 plumose seta endopod3-segmented distinction between segmentssometimes obscure 2 2 (3 or 4) setae pro-topod without setation
Maxilla (Fig 7f )mdashCoxal endite bilobed14ndash19 setae on proximal lobe 5 setae on dis-tal lobe basal endite bilobed 6ndash10 setae onproximal lobe 8 or 9 setae on distal lobeendopod with 5 lobes 3+(2 or 3)+2+2+3 se-tae scaphognathite with 27ndash32 setae all se-tae plumose
Maxilliped 1 (Fig 7g)mdashCoxa with 2ndash8 se-tae basis with 16ndash19 marginal setae endo-pod 4-segmented (3 or 4) 2 2 4 setae exo-
pod 2-segmented proximal segment withoutsetae distal segment with 6 or 7 setae on dis-tal margin bilobed epipod without setae allsetae plumose
Maxilliped 2 (Fig 7h)mdashCoxa with 0ndash3 se-tae basis with 4ndash6 setae endopod 4-seg-mented (5ndash8) 1 (3ndash5) (4 or 5) setae exo-pod unsegmented 7 or 8 setae on distal mar-gin all setae plumose
Maxilliped 3 (Fig 8a)mdashCoxa with 0 or 1seta basis with 2ndash4 setae endopod 4-seg-mented (l or 2) 2 (3ndash5) (3 or 4) setae exo-pod unsegmented 7 or 8 setae on distal mar-gin all setae plumose
Pereiopod 1 (Fig 8b)mdashBiramous cheliformcoxa without setae basis with 0ndash2 setae en-dopod 4-segmented (0 or 1) (1 or 2) (2ndash4)(1 or 2) setae exopod with 5ndash7 setae on dis-tal margin
Pereiopod 2 (Fig 8c)mdashBiramous cheliformcoxa without setae basis with 0 or 1 setaendopod 4-segmented 0 (1 or 2) (3ndash5) (2or 3) setae exopod with 5ndash7 setae on distalmargin
Pereiopod 3 (Fig 8d)mdashBiramous coxa andbasis without setae endopod 4-segmented0 (0ndash2) (3 or 4) 2 setae propodus blade-like exopod with 4 or 5 terminal setae
Pereiopod 4 (Fig 8e)mdashBiramous coxa andbasis without setae endopod 4-segmented (0or 1) (0ndash2) (2ndash4) 2 setae exopod with 0ndash2setae
Pereiopod 5 (Fig 8f )mdashUniramous 2 setaeon terminal segment 4 or 5 setae on adja-cent segment
Pleopods (Fig 8g h)mdashLarger than at ZIIIstill without setae
Telson and Uropods (Fig 8i)mdashTelson trian-gular narrow anteriorly no anal spine pos-terior margin with (12ndash15)+1+(13 or 14)processes arranged as (I II III IV III II I)(I) outermost process a plumodenticulate seta(II) second process ldquoanomuran hairrdquo (III) 10ndash13plumodenticulate setae and (IV) naked spineuropods biramous endopod with 7ndash11 marginalplumose setae exopod with 1 naked distolat-eral spine 12ndash14 marginal plumose setae
110 JOURNAL OF CRUSTACEAN BIOLOGY VOL 20 NO 1 2000
Decapodid (first postlarva)Figs 2e 9cndashj l0andashi
SizemdashCL = 133 plusmn 002 mm range129ndash138 mm TL = 378 plusmn 011 mm range361ndash405 mm
Carapace (Fig 2e)mdashShorter than abdomenno anterolateral serrations with pterygostomial
projections rostrum reduced without serra-tions sometimes bending down between eyes
Abdomen (Fig 2e)mdashSomites broad and flat-tened posterior projections rounded with 0ndash7setae somite 2 rectangular somites 3ndash5 oval
Antennule (Fig 9c)mdashPeduncle 3-segmentedproximal segment with 17ndash24 setae 13ndash15 of
Fig 7 Callichirus islagrande ZIV stage a antennule b antenna c right mandible d left mandible e maxil-lule f maxilla (plumose condition not shown) g maxilliped 1 h maxilliped 2 All setae on the maxilla were plumoseScales = 02 mm
STRASSER AND FELDER LARVAL DEVELOPMENT IN CALLICHIRUS ISLAGRANDE 111
these associated with statocyst intermediatesegment with 6ndash10 setae distal segment of pe-duncle with 2 vertical rows of setae one with4 or 5 setae other with 2ndash4 setae 1ndash3 addi-tional setae on outer margin and 3ndash5 short se-tae on distal margin endopod 4ndash6 segmented(1ndash3) (0 or 1) (1ndash4) (1ndash4) (3ndash6) (3ndash6) se-tae exopod 3ndash5-segmented 0 (0ndash4) (0ndash4)(1ndash4) setae +2 asthetascs (5ndash8) asthetascs ()
Antenna (Fig 9d)mdashPeduncle 2-segmentedproximal segment with 0 or 1 seta distal seg-ment with 1ndash3 setae on inner margin spinepresent in some flagellum ranging from 14ndash18
segments (0 or 1) (2ndash4) variable region(1ndash3 articles with 0ndash2 setae) (0ndash4) (0ndash2)(2ndash5) (1ndash4) (3ndash6) (0ndash3) (2ndash5) (3ndash6)(0ndash3) (3ndash6) (0 or 1) (1ndash7) (4ndash9) setae exo-pod reduced to small lobe with 0ndash5 setae
Mandible (Fig 9e f )mdashBoth mandibles sim-ilar incisor process with 5ndash8 teeth molarprocess smooth or with small ridges palp3-segmented 0 (0 or 1) (8ndash12) setae
Maxillule (Fig 9g)mdashCoxal endite with15ndash22 marginal setae and 0 or 1 submarginal
Fig 8 Callichirus islagrande ZIV stage a maxilliped 3 b pereiopod 1 c pereiopod 2 d pereiopod 3 e pe-reiopod 4 f pereiopod 5 g pleopods from first abdominal somite h pleopods from fourth abdominal somite i tel-son Scales = 02 mm
112 JOURNAL OF CRUSTACEAN BIOLOGY VOL 20 NO 1 2000
seta basal endite with 3ndash5 plumose setae14ndash19 dentate spines 0ndash2 plumose setae oninner margin endopod reduced unsegmentedwith 0ndash8 setae
Maxilla (Fig 9h)mdashCoxal endite trilobed(17ndash21)+(7ndash10)+(6ndash9) setae basal enditebilobed 8ndash10 setae on proximal lobe 8ndash15setae on distal lobe endopod reduced with
Fig 9 Callichirus islagrande ZV stage a one example of the antenna b one example of pleopods Decapodidstage c antennule d antenna e right mandible f left mandible g maxillule h maxilla (plumose condition notshown) i maxilliped 1 j maxilliped 2 Scales = 02 mm
STRASSER AND FELDER LARVAL DEVELOPMENT IN CALLICHIRUS ISLAGRANDE 113
5ndash11 setae scaphognathite with 37ndash45 se-tae all setae plumose
Maxilliped 1 (Fig 9i)mdashCoxa with 8ndash12 se-tae basis highly setose with 27ndash39 setae en-dopod reduced with 0ndash11 setae exopod un-segmented 6ndash14 setae bilobed epipod with-out setae
Maxilliped 2 (Fig 9j)mdashCoxa partially fusedto basis with 1ndash4 setae basis with 3ndash5 se-
tae endopod 4-segmented (16ndash23) (0 or 1)(6ndash11) (4ndash8) setae exopod with 0ndash8 setaeepipod present but without setae
Maxilliped 3 (Fig 10a)mdashCoxa partiallyfused to basis with 0ndash3 setae basis with 1ndash4setae endopod 5-segmented (11ndash19) (9ndash15)(7ndash11) (8ndash21) (5ndash11) setae exopod usuallyreduced and without setae occasionally with2ndash9 setae epipod present
Fig 10 Callichirus islagrande decapodid stage a maxilliped 3 b pereiopod 1 c pereiopod 2 d pereiopod 3e pereiopod 4 f pereiopod 5 g pleopods from first abdominal somite h pleopods from fourth abdominal somitei telson Scales = 02 mm
114 JOURNAL OF CRUSTACEAN BIOLOGY VOL 20 NO 1 2000
Pereiopod 1 (Fig 10b)mdashChelate biramous7-segmented coxa with 0ndash6 setae basis with2ndash5 setae ischium with 4ndash7 setae and 0ndash3spines merus with 5ndash8 setae carpus with10ndash14 setae propodus with 27ndash37 setae and3ndash4 spines on inner margin of immovable fin-ger dactylus with 21ndash33 setae exopod usu-ally reduced and without setae occasionallywith 4ndash7 setae
Pereiopod 2 (Fig 10c)mdashChelate biramous7-segmented coxa with 2ndash5 setae basis with1ndash5 setae ischium with 4ndash10 setae meruswith 15ndash20 setae carpus with 8ndash12 setaepropodus with 30ndash40 setae and 1ndash4 spineson inner margin of immovable finger dacty-lus with 21ndash30 setae exopod usually reduced0ndash7 setae
Pereiopod 3 (Fig 10d)mdashBiramous 7-seg-mented coxa with 2ndash5 setae basis with 1ndash3setae ischium with 5ndash8 setae merus with6ndash9 setae carpus with 7ndash11 setae propodusbladelike with 25ndash34 setae 1 spine on innerdistal margin dactylus with 15ndash21 setae exo-pod usually reduced 0ndash5 setae
Pereiopod 4 (Fig 10e)mdashBiramous 7-seg-mented coxa with 2ndash7 setae basis with 1ndash4setae ischium with 3ndash8 setae merus with5ndash11 setae carpus with 4ndash9 setae propoduswith 27ndash38 setae 1 spine on inner distal mar-gin dactylus with 15ndash19 setae exopod usu-ally reduced with 0ndash2 setae
Pereiopod 5 (Fig 10f )mdashCheliform unira-mous 7-segmented coxa with 2ndash6 setae ba-sis with l or 2 setae ischium with 2ndash5 setaemerus with 5ndash9 setae carpus with 4ndash7 se-tae propodus with 22ndash30 setae 1 spine onimmovable finger dactylus with 6ndash10 setaeexopod absent
Pleopods (somite 2) (Fig 10g)mdashBiramouscoxa partially fused to basis without setaebasis with 0ndash3 setae endopod with 3ndash5hooked setae on appendix interna 10ndash17 mar-ginal plumose setae 0ndash2 submarginal nakedsetae exopod with 21ndash27 marginal plumosesetae and 0ndash1 submarginal naked seta
Pleopods (somites 3ndash5) (Fig 10h)mdashBiramouscoxa without setae basis with 3ndash6 marginalplumose setae endopod with 3ndash6 hooked se-tae on appendix interna 10ndash18 marginalplumose setae and 0ndash3 naked submarginal se-tae exopod with 20ndash29 marginal plumose se-tae and 0 or 1 submarginal naked seta
Telson and Uropods (Fig 10i)mdashWith 24ndash36processes on posterior margin median pro-cess usually absent or reduced uropods bi-ramous endopod with 19ndash25 plumose setaeexopod with 35ndash58 plumose setae protopodwith 3ndash5 setae
DISCUSSION
As in other species of Callichirus (see Ro-drigues 1976 Aste and Retamal 1983Strasser and Felder in press b) larval devel-opment of C islagrande included four or fivezoeal stages before metamorphosis to the de-capodid ZI larvae may be identified by thepresence of an elongate rounded rostrumsessile eyes and a lack of pereiopodspleopods or uropods (Fig 1a) At ZII lar-vae acquired biramous buds of pereiopods 1and 2 eyes were separated from the carapaceand the rostrum became flattened and serratedlaterally (Fig 1b) Buds of pleopods on ab-dominal segments 2ndash5 pereiopods 3ndash5 anduropods were developed at ZIII (Fig 1c) Inaddition to further development of structuresmentioned thus far ZIV larvae could be dis-tinguished by presence of a distal spine on theexopod and setae on the endopod of each uro-pod (Fig 1d) When it occurred ZV typicallyresembled ZIV but varied widely in mor-phology Appendages at ZV resembled thoseof ZIV the decapodid stage or some grada-tion between the two (see Figs 9a b for ex-amples) The decapodid stage was similar toother species of Callichirus in the reductionof larval characters such as the elongate ros-trum dorsal spines on the abdomen and ex-opods of the pereiopods (Fig le) Acquisitionof fan-like pleopods marked the transition toa primarily abdominal mode of swimming atthis stage
In the Florida Atlantic population of C ma-jor both salinity and presence of putative set-tlement cues at ZIV impacted the observednumber of zoeal stages and the duration ofZIV (Strasser and Felder in preparation1999c) In C islagrande the percentage oflarvae that passed through five versus fourzoeal stages varied (9ndash89) between parentalfemales but was not shown to depend onsalinity or presence of various putative set-tlement cues (Strasser and Felder 1998 inpreparation) Larvae that molted from ZIVto ZV were usually normal morphologicallyand had mortality similar to those that molteddirectly from ZIV to D In contrast larvae of
STRASSER AND FELDER LARVAL DEVELOPMENT IN CALLICHIRUS ISLAGRANDE 115
the Florida Atlantic C major that molted toZV had a higher incidence of mortality anddeformity at D than did those that had by-passed ZV (Strasser and Felder 1999a c)
The number of zoeal stages appears to beless variable in Callichirus garthi and theGulf of Mexico population of C major thanin congeneric populations studied to dateCallichirus garthi was reported to have fivezoeal stages and a long larval duration reach-ing the decapodid stage 48 days after hatch-ing (Aste and Retamal 1983) Developmentof the Gulf population of C major wasshorter in duration (decapodid reached about10 days after hatching) and typically includedfour zoeal stages (Strasser and Felder 1999a)animals rarely passed through three or fivezoeal stages before the molt to the decapodidAlthough the number of stages is reducedfrom that observed in C garthi developmentin the Gulf population of C major cannot beclassified as abbreviated There are species inboth the Callianassinae (Callianassa slkraussi Stebbing C sl kewalramanii and Csl tyrrhena (Petagna)) and Callichirinae(Sergio mirim Lepidophthalmus sinuensisand L louisianensis) that have only two zoealstages before the molt to the decapodid(Forbes 1973 Sankolli and Shenoy 1975Rodrigues 1984 Thessalou-Legaki 1990Nates et al 1997)
Compared to other species of the genusCallichirus C islagrande is morphologicallycloser to C garthi (from Chile) than to theGulf of Mexico population of C major whichoverlaps in distribution with C islagrande Inthe zoeal phase of C islagrande and Cgarthi the telson is concave and the outer-most process on the ventral margin is articu-lated In both C major and Callichirus sp theposterior margin of the telson is straight andthe outermost process is unarticulated duringthe zoeal stages (Rodrigues 1976 Strasserand Felder 1999a) There are also differencesin spination of the abdomen In zoeae of Cislagrande and C garthi the dorsal spine ofabdominal segment 3 is not longer than thosefound on segments 4 and 5 (Aste and Reta-mal 1983) In contrast the dorsal spine of ab-dominal segment 3 is approximately twice aslong as the dorsal spines on segments 4 and5 in early zoeal stages of C major and Cal-lichirus sp (Rodrigues 1976 Aste and Re-tamal 1983 Strasser and Felder 1999a) Thegreatest difference between these groups is
the number of pleopod pairs at zoeal and de-capodid stages Callichirus islagrande and Cgarthi have pleopods on abdominal segments2ndash5 while pleopods are present only on ab-dominal segments 3ndash5 in C major and Cal-lichirus sp (Rodrigues 1976 Aste and Re-tamal 1983 Strasser and Felder 1999a) Al-though Callichirus islagrande and C garthiare similar morphologically there are differ-ences between these two species Callichirusislagrande as well as C major and Cal-lichirus sp develops pleopod buds andchelate pereiopods at ZIII whereas C garthidoes not acquire these characters until ZIV
There are few callianassid species forwhich the larval development has been de-scribed (Sankolli and Shenoy 1975 Ro-drigues 1976 1984 Aste and Retamal 19831984 Konishi et al 1990 Nates et al 1997Strasser and Felder 1999a) all of which aremembers of the Callichirinae and Cal-lianassinae Among species there is a greatdeal of overlap in larval characters makingit difficult to draw distinctions between thesesubfamilies At present information on lar-val development of species in the Callichiri-nae is limited to larval descriptions of Cal-lichirus major C garthi Callichirus sp Cislagrande Sergio mirim Lepidophthalmuslouisianensis and L sinuensis (see Ro-drigues 1976 1984 Aste and Retamal 1983Nates et al 1997 Strasser and Felder1999a) As mentioned above species of Cal-lichirus have been shown to have four to fivezoeal stages while the other members exhibitabbreviated development that consists of onlytwo zoeal stages The most obvious differencebetween larvae of the species of Callichirinaewith abbreviated development is the numberof pleopod pairs Larvae of S mirim havepleopods on abdominal segments 3ndash5whereas L louisianensis and L sinuensishave pleopods on segments 2ndash5 (Rodrigues1984 Nates et al 1997)
Ghost shrimps of the genus Callichirus maybe distinguished from other genera of the Cal-lianassidae by the cuticular pattern on the dor-sal surface of the abdomen and by the strap-like shape of the uropodal endopods (Manningand Felder 1991) Whereas these charactersmay be used to distinguish adults of the genusCallichirus they are not present at zoeal anddecapodid stages Larvae may be distin-guished from all other callianassid genera byhaving four to five zoeal stages and numer-
116 JOURNAL OF CRUSTACEAN BIOLOGY VOL 20 NO 1 2000
ous processes (12ndash25+1+12ndash25) on the pos-terior margin of the telson during the earlyzoeal stages Larvae of Callichirus may alsodiffer from other callianassids with extendeddevelopment by the retention of the anomu-ran hair as the second process on the poste-rior margin of the telson in the late zoealstages Whereas this process is converted toa spine in Neotrypaea uncinata at ZV and Cal-lianassa sl petalura at ZIV the anomuranhair is retained at ZIV in Callichirus majorand C islagrande (see Rodrigues 1976Strasser and Felder 1999a) Although notmentioned specifically in the descriptions theanomuran hair appears to be present in illus-trations of Callichirus sp at ZIII and Cgarthii at ZV (Rodrigues 1976 fig 35 Asteand Retamal 1983 fig 8Q) A bilobed epi-pod on the first maxilliped of ZI is consistentlyfound in Callichirus but is also present at ZIin Lepidophthalmus sinuensis (see Nates etal 1997) Other callianassids acquire thischaracter at ZII (L louisianensis Nates et al1997 Callianassa sl kewlramanii Sankolliand Shenoy 1975) at ZV (Neotrypaea unci-nata Aste and Retamal 1984) or at the de-capodid stage (Sergio mirim Rodrigues 1984C sl petalura Konishi et al 1990) Thusthe number of zoeal stages and setation of thetelson may serve as the best characters to dis-tinguish larvae of Callichirus from other gen-era However since larval descriptions arelacking for many callianassid species it is un-known if these characters (in combination)will prove to be diagnostic until descriptionsfor other species are published
ACKNOWLEDGEMENTS
We thank Robert Bourgeois Barbara Hasek JimMaKinster Sergio Nates and Kurt Strasser who pro-vided assistance in the field and laboratory and MaryE Rice Director of the Smithsonian Marine StationLink Port Florida who facilitated access to station fa-cilities We also thank R Bauer R Jaeger R Lemaitreand G Watson for their early comments on this manu-script and Raymond B Manning who facilitated fund-ing and gave advice on collecting sites Direct supportfor this study was provided to Karen M Strasser by adoctoral fellowship from the University of Louisianaa graduate student research grant from the LUMCON(Louisiana Universities Marine Consortium) Founda-tion and a summer research award from the Smith-sonian Marine Station Additional support for field andlab activities was provided to D L Felder under Loui-siana Sea Grant College Program Grant RCFB-21 USFish and Wildlife Service Cooperative Agreement 14-16-0009-89-96 (Task Order No 6) US Department of En-ergy grant DE-FG02-97ER12220 and several small proj-ect grants from the Smithsonian Marine Station Views
expressed herein are those of the authors and do not nec-essarily reflect views of supporting government agenciesor subagencies mentioned above This is contributionNumber 466 of the Smithsonian Marine Station and con-tribution Number 67 of the ULL Laboratory for Crus-tacean Research All experiments conducted in this studycomply with current applicable state and federal laws
LITERATURE CITED
Aste A and M Retamal 1983 Desarrollo larval deCallianassa garthi Retamal 1975 bajo condiciones delaboratoriomdashCiencia y Tecnologiacutea del Mar CONA 75ndash26
mdashmdashmdash and mdashmdashmdash 1984 Desarrollo larval de Cal-lianassa uncinata H Milne Edwards1837 (DecapodaCallianassidae) bajo condiciones de laboratoriomdashGayana Zooloacutegica 48 41ndash56
Felder D L 1978 Osmotic and ionic regulation in sev-eral western Atlantic Callianassidae (Crustacea De-capoda Thalassinidea)mdashBiological Bulletin 154409ndash429
mdashmdashmdash and R B Griffis 1994 Dominant infaunalcommunities at risk in shoreline habitats burrowingthalassinid CrustaceamdashOCS Study number MMS94ndash0007 United States Department of the InteriorMinerals Management Service Gulf of Mexico OCSRegional Office New Orleans Louisiana Pp 1ndash87
Forbes A T 1973 An unusual abbreviated larval lifehistory in the estuarine burrowing prawn Callianassakraussi (Crustacea Decapoda Thalassinidea)mdashMa-rine Biology 22 361ndash365
Konishi K 1989 Larval development of the mudshrimp Upogebia (Upogebia) major (de Haan) (Crus-tacea Thalassinidea Upogebiidae) under laboratoryconditions with comments on larval characters of tha-lassinid familiesmdashBulletin of the National ResearchInstitute of Aquaculture 15 1ndash17
mdashmdashmdash R Quintana and Y Fukuda 1990 A completedescription of larval stages of the ghost shrimp Cal-lianassa petalura Stimpson (Crustacea ThalassinideaCallianassidae) under laboratory conditionsmdashBulletinof the National Research Institute of Aquaculture 1727ndash49
Manning R B and D L Felder 1986 The status ofthe callianassid genus Callichirus Stimpson 1866(Crustacea Decapoda Thalassinidea)mdashProceedingsof the Biological Society of Washington 99 437ndash443
mdashmdashmdash and mdashmdashmdash 1991 Revision of the AmericanCallianassidae (Crustacea Decapoda Thalassinidea)mdashProceedings of the Biological Society of Washington104 764ndash794
Nates S F D L Felder and R Lemaitre 1997 Com-parative larval development in two species of the bur-rowing ghost shrimp genus Lepidophthalmus (Decap-oda Callianassidae)mdashJournal of Crustacean Biology17 497ndash519
Rodrigues S de A 1976 Sobre a reproduccedilatildeo embri-ologia e desenvolvimento larval de Callichirus majorSay 1818 (Crustacea Decapoda Thalassinidea)mdashBo-letim de Zoologia Universidade de Satildeo Paulo 185ndash104
mdashmdashmdash 1984 Desenvolvimento poacutes-embrionaacuterio deCallichirus mirim (Rodrigues 1971) obtido emcondiccedilotildees artificiais (Crustacea Decapoda Tha-lassinidea)mdashBoletim de Zoologia Universidade deSatildeo Paulo 8 239ndash256
STRASSER AND FELDER LARVAL DEVELOPMENT IN CALLICHIRUS ISLAGRANDE 117
Sankolli K N and S Shenoy 1975 Larval develop-ment of mud shrimp Callianassa (Callichirus) kewal-ramanii Sankolli in the laboratory (Crustacea Decap-oda)mdashBulletin of the Department of Marine ScienceUniversity of Cochin 4 704ndash720
Staton J L and D L Felder 1995 Genetic variationin populations of the ghost shrimp genus Callichirus(Crustacea Decapoda Thalassinoidea) in the westernAtlantic and Gulf of MexicomdashBulletin of Marine Sci-ence 56 523ndash536
Strasser K M and D L Felder 1998 Settlement cuesin successive developmental stages of the ghost shrimpCallichirus major and C islagrande (Crustacea De-capoda Thalassinidea)mdashMarine Biology
mdashmdashmdash and mdashmdashmdash 1999a Larval development of twopopulations of the ghost shrimp Callichirus major (De-capoda Thalassinidea)mdashJournal of Crustacean Biology
mdashmdashmdash and mdashmdashmdash 1999b Sand as a stimulus for set-tlement in the ghost shrimp Callichirus major (Say)and C islagrande (Schmitt) (Crustacea DecapodaThalassinidea)mdashJournal of Experimental Marine Bi-ology and Ecology 239 211ndash222
mdashmdashmdash and mdashmdashmdash 1999c Settlement cues in theFlorida Atlantic population of the ghost shrimp Cal-lichirus major (Crustacea Decapoda Thalassinidea)mdashMarine Ecology Progress Series
Thessalou-Legaki M 1990 Advanced larval develop-ment of Callianassa tyrrhena (Decapoda Tha-lassinidea) and the effect of environmental factorsmdashJournal of Crustacean Biology 10 659ndash666
RECEIVED 19 September 1998ACCEPTED 3 December 1998

110 JOURNAL OF CRUSTACEAN BIOLOGY VOL 20 NO 1 2000
Decapodid (first postlarva)Figs 2e 9cndashj l0andashi
SizemdashCL = 133 plusmn 002 mm range129ndash138 mm TL = 378 plusmn 011 mm range361ndash405 mm
Carapace (Fig 2e)mdashShorter than abdomenno anterolateral serrations with pterygostomial
projections rostrum reduced without serra-tions sometimes bending down between eyes
Abdomen (Fig 2e)mdashSomites broad and flat-tened posterior projections rounded with 0ndash7setae somite 2 rectangular somites 3ndash5 oval
Antennule (Fig 9c)mdashPeduncle 3-segmentedproximal segment with 17ndash24 setae 13ndash15 of
Fig 7 Callichirus islagrande ZIV stage a antennule b antenna c right mandible d left mandible e maxil-lule f maxilla (plumose condition not shown) g maxilliped 1 h maxilliped 2 All setae on the maxilla were plumoseScales = 02 mm
STRASSER AND FELDER LARVAL DEVELOPMENT IN CALLICHIRUS ISLAGRANDE 111
these associated with statocyst intermediatesegment with 6ndash10 setae distal segment of pe-duncle with 2 vertical rows of setae one with4 or 5 setae other with 2ndash4 setae 1ndash3 addi-tional setae on outer margin and 3ndash5 short se-tae on distal margin endopod 4ndash6 segmented(1ndash3) (0 or 1) (1ndash4) (1ndash4) (3ndash6) (3ndash6) se-tae exopod 3ndash5-segmented 0 (0ndash4) (0ndash4)(1ndash4) setae +2 asthetascs (5ndash8) asthetascs ()
Antenna (Fig 9d)mdashPeduncle 2-segmentedproximal segment with 0 or 1 seta distal seg-ment with 1ndash3 setae on inner margin spinepresent in some flagellum ranging from 14ndash18
segments (0 or 1) (2ndash4) variable region(1ndash3 articles with 0ndash2 setae) (0ndash4) (0ndash2)(2ndash5) (1ndash4) (3ndash6) (0ndash3) (2ndash5) (3ndash6)(0ndash3) (3ndash6) (0 or 1) (1ndash7) (4ndash9) setae exo-pod reduced to small lobe with 0ndash5 setae
Mandible (Fig 9e f )mdashBoth mandibles sim-ilar incisor process with 5ndash8 teeth molarprocess smooth or with small ridges palp3-segmented 0 (0 or 1) (8ndash12) setae
Maxillule (Fig 9g)mdashCoxal endite with15ndash22 marginal setae and 0 or 1 submarginal
Fig 8 Callichirus islagrande ZIV stage a maxilliped 3 b pereiopod 1 c pereiopod 2 d pereiopod 3 e pe-reiopod 4 f pereiopod 5 g pleopods from first abdominal somite h pleopods from fourth abdominal somite i tel-son Scales = 02 mm
112 JOURNAL OF CRUSTACEAN BIOLOGY VOL 20 NO 1 2000
seta basal endite with 3ndash5 plumose setae14ndash19 dentate spines 0ndash2 plumose setae oninner margin endopod reduced unsegmentedwith 0ndash8 setae
Maxilla (Fig 9h)mdashCoxal endite trilobed(17ndash21)+(7ndash10)+(6ndash9) setae basal enditebilobed 8ndash10 setae on proximal lobe 8ndash15setae on distal lobe endopod reduced with
Fig 9 Callichirus islagrande ZV stage a one example of the antenna b one example of pleopods Decapodidstage c antennule d antenna e right mandible f left mandible g maxillule h maxilla (plumose condition notshown) i maxilliped 1 j maxilliped 2 Scales = 02 mm
STRASSER AND FELDER LARVAL DEVELOPMENT IN CALLICHIRUS ISLAGRANDE 113
5ndash11 setae scaphognathite with 37ndash45 se-tae all setae plumose
Maxilliped 1 (Fig 9i)mdashCoxa with 8ndash12 se-tae basis highly setose with 27ndash39 setae en-dopod reduced with 0ndash11 setae exopod un-segmented 6ndash14 setae bilobed epipod with-out setae
Maxilliped 2 (Fig 9j)mdashCoxa partially fusedto basis with 1ndash4 setae basis with 3ndash5 se-
tae endopod 4-segmented (16ndash23) (0 or 1)(6ndash11) (4ndash8) setae exopod with 0ndash8 setaeepipod present but without setae
Maxilliped 3 (Fig 10a)mdashCoxa partiallyfused to basis with 0ndash3 setae basis with 1ndash4setae endopod 5-segmented (11ndash19) (9ndash15)(7ndash11) (8ndash21) (5ndash11) setae exopod usuallyreduced and without setae occasionally with2ndash9 setae epipod present
Fig 10 Callichirus islagrande decapodid stage a maxilliped 3 b pereiopod 1 c pereiopod 2 d pereiopod 3e pereiopod 4 f pereiopod 5 g pleopods from first abdominal somite h pleopods from fourth abdominal somitei telson Scales = 02 mm
114 JOURNAL OF CRUSTACEAN BIOLOGY VOL 20 NO 1 2000
Pereiopod 1 (Fig 10b)mdashChelate biramous7-segmented coxa with 0ndash6 setae basis with2ndash5 setae ischium with 4ndash7 setae and 0ndash3spines merus with 5ndash8 setae carpus with10ndash14 setae propodus with 27ndash37 setae and3ndash4 spines on inner margin of immovable fin-ger dactylus with 21ndash33 setae exopod usu-ally reduced and without setae occasionallywith 4ndash7 setae
Pereiopod 2 (Fig 10c)mdashChelate biramous7-segmented coxa with 2ndash5 setae basis with1ndash5 setae ischium with 4ndash10 setae meruswith 15ndash20 setae carpus with 8ndash12 setaepropodus with 30ndash40 setae and 1ndash4 spineson inner margin of immovable finger dacty-lus with 21ndash30 setae exopod usually reduced0ndash7 setae
Pereiopod 3 (Fig 10d)mdashBiramous 7-seg-mented coxa with 2ndash5 setae basis with 1ndash3setae ischium with 5ndash8 setae merus with6ndash9 setae carpus with 7ndash11 setae propodusbladelike with 25ndash34 setae 1 spine on innerdistal margin dactylus with 15ndash21 setae exo-pod usually reduced 0ndash5 setae
Pereiopod 4 (Fig 10e)mdashBiramous 7-seg-mented coxa with 2ndash7 setae basis with 1ndash4setae ischium with 3ndash8 setae merus with5ndash11 setae carpus with 4ndash9 setae propoduswith 27ndash38 setae 1 spine on inner distal mar-gin dactylus with 15ndash19 setae exopod usu-ally reduced with 0ndash2 setae
Pereiopod 5 (Fig 10f )mdashCheliform unira-mous 7-segmented coxa with 2ndash6 setae ba-sis with l or 2 setae ischium with 2ndash5 setaemerus with 5ndash9 setae carpus with 4ndash7 se-tae propodus with 22ndash30 setae 1 spine onimmovable finger dactylus with 6ndash10 setaeexopod absent
Pleopods (somite 2) (Fig 10g)mdashBiramouscoxa partially fused to basis without setaebasis with 0ndash3 setae endopod with 3ndash5hooked setae on appendix interna 10ndash17 mar-ginal plumose setae 0ndash2 submarginal nakedsetae exopod with 21ndash27 marginal plumosesetae and 0ndash1 submarginal naked seta
Pleopods (somites 3ndash5) (Fig 10h)mdashBiramouscoxa without setae basis with 3ndash6 marginalplumose setae endopod with 3ndash6 hooked se-tae on appendix interna 10ndash18 marginalplumose setae and 0ndash3 naked submarginal se-tae exopod with 20ndash29 marginal plumose se-tae and 0 or 1 submarginal naked seta
Telson and Uropods (Fig 10i)mdashWith 24ndash36processes on posterior margin median pro-cess usually absent or reduced uropods bi-ramous endopod with 19ndash25 plumose setaeexopod with 35ndash58 plumose setae protopodwith 3ndash5 setae
DISCUSSION
As in other species of Callichirus (see Ro-drigues 1976 Aste and Retamal 1983Strasser and Felder in press b) larval devel-opment of C islagrande included four or fivezoeal stages before metamorphosis to the de-capodid ZI larvae may be identified by thepresence of an elongate rounded rostrumsessile eyes and a lack of pereiopodspleopods or uropods (Fig 1a) At ZII lar-vae acquired biramous buds of pereiopods 1and 2 eyes were separated from the carapaceand the rostrum became flattened and serratedlaterally (Fig 1b) Buds of pleopods on ab-dominal segments 2ndash5 pereiopods 3ndash5 anduropods were developed at ZIII (Fig 1c) Inaddition to further development of structuresmentioned thus far ZIV larvae could be dis-tinguished by presence of a distal spine on theexopod and setae on the endopod of each uro-pod (Fig 1d) When it occurred ZV typicallyresembled ZIV but varied widely in mor-phology Appendages at ZV resembled thoseof ZIV the decapodid stage or some grada-tion between the two (see Figs 9a b for ex-amples) The decapodid stage was similar toother species of Callichirus in the reductionof larval characters such as the elongate ros-trum dorsal spines on the abdomen and ex-opods of the pereiopods (Fig le) Acquisitionof fan-like pleopods marked the transition toa primarily abdominal mode of swimming atthis stage
In the Florida Atlantic population of C ma-jor both salinity and presence of putative set-tlement cues at ZIV impacted the observednumber of zoeal stages and the duration ofZIV (Strasser and Felder in preparation1999c) In C islagrande the percentage oflarvae that passed through five versus fourzoeal stages varied (9ndash89) between parentalfemales but was not shown to depend onsalinity or presence of various putative set-tlement cues (Strasser and Felder 1998 inpreparation) Larvae that molted from ZIVto ZV were usually normal morphologicallyand had mortality similar to those that molteddirectly from ZIV to D In contrast larvae of
STRASSER AND FELDER LARVAL DEVELOPMENT IN CALLICHIRUS ISLAGRANDE 115
the Florida Atlantic C major that molted toZV had a higher incidence of mortality anddeformity at D than did those that had by-passed ZV (Strasser and Felder 1999a c)
The number of zoeal stages appears to beless variable in Callichirus garthi and theGulf of Mexico population of C major thanin congeneric populations studied to dateCallichirus garthi was reported to have fivezoeal stages and a long larval duration reach-ing the decapodid stage 48 days after hatch-ing (Aste and Retamal 1983) Developmentof the Gulf population of C major wasshorter in duration (decapodid reached about10 days after hatching) and typically includedfour zoeal stages (Strasser and Felder 1999a)animals rarely passed through three or fivezoeal stages before the molt to the decapodidAlthough the number of stages is reducedfrom that observed in C garthi developmentin the Gulf population of C major cannot beclassified as abbreviated There are species inboth the Callianassinae (Callianassa slkraussi Stebbing C sl kewalramanii and Csl tyrrhena (Petagna)) and Callichirinae(Sergio mirim Lepidophthalmus sinuensisand L louisianensis) that have only two zoealstages before the molt to the decapodid(Forbes 1973 Sankolli and Shenoy 1975Rodrigues 1984 Thessalou-Legaki 1990Nates et al 1997)
Compared to other species of the genusCallichirus C islagrande is morphologicallycloser to C garthi (from Chile) than to theGulf of Mexico population of C major whichoverlaps in distribution with C islagrande Inthe zoeal phase of C islagrande and Cgarthi the telson is concave and the outer-most process on the ventral margin is articu-lated In both C major and Callichirus sp theposterior margin of the telson is straight andthe outermost process is unarticulated duringthe zoeal stages (Rodrigues 1976 Strasserand Felder 1999a) There are also differencesin spination of the abdomen In zoeae of Cislagrande and C garthi the dorsal spine ofabdominal segment 3 is not longer than thosefound on segments 4 and 5 (Aste and Reta-mal 1983) In contrast the dorsal spine of ab-dominal segment 3 is approximately twice aslong as the dorsal spines on segments 4 and5 in early zoeal stages of C major and Cal-lichirus sp (Rodrigues 1976 Aste and Re-tamal 1983 Strasser and Felder 1999a) Thegreatest difference between these groups is
the number of pleopod pairs at zoeal and de-capodid stages Callichirus islagrande and Cgarthi have pleopods on abdominal segments2ndash5 while pleopods are present only on ab-dominal segments 3ndash5 in C major and Cal-lichirus sp (Rodrigues 1976 Aste and Re-tamal 1983 Strasser and Felder 1999a) Al-though Callichirus islagrande and C garthiare similar morphologically there are differ-ences between these two species Callichirusislagrande as well as C major and Cal-lichirus sp develops pleopod buds andchelate pereiopods at ZIII whereas C garthidoes not acquire these characters until ZIV
There are few callianassid species forwhich the larval development has been de-scribed (Sankolli and Shenoy 1975 Ro-drigues 1976 1984 Aste and Retamal 19831984 Konishi et al 1990 Nates et al 1997Strasser and Felder 1999a) all of which aremembers of the Callichirinae and Cal-lianassinae Among species there is a greatdeal of overlap in larval characters makingit difficult to draw distinctions between thesesubfamilies At present information on lar-val development of species in the Callichiri-nae is limited to larval descriptions of Cal-lichirus major C garthi Callichirus sp Cislagrande Sergio mirim Lepidophthalmuslouisianensis and L sinuensis (see Ro-drigues 1976 1984 Aste and Retamal 1983Nates et al 1997 Strasser and Felder1999a) As mentioned above species of Cal-lichirus have been shown to have four to fivezoeal stages while the other members exhibitabbreviated development that consists of onlytwo zoeal stages The most obvious differencebetween larvae of the species of Callichirinaewith abbreviated development is the numberof pleopod pairs Larvae of S mirim havepleopods on abdominal segments 3ndash5whereas L louisianensis and L sinuensishave pleopods on segments 2ndash5 (Rodrigues1984 Nates et al 1997)
Ghost shrimps of the genus Callichirus maybe distinguished from other genera of the Cal-lianassidae by the cuticular pattern on the dor-sal surface of the abdomen and by the strap-like shape of the uropodal endopods (Manningand Felder 1991) Whereas these charactersmay be used to distinguish adults of the genusCallichirus they are not present at zoeal anddecapodid stages Larvae may be distin-guished from all other callianassid genera byhaving four to five zoeal stages and numer-
116 JOURNAL OF CRUSTACEAN BIOLOGY VOL 20 NO 1 2000
ous processes (12ndash25+1+12ndash25) on the pos-terior margin of the telson during the earlyzoeal stages Larvae of Callichirus may alsodiffer from other callianassids with extendeddevelopment by the retention of the anomu-ran hair as the second process on the poste-rior margin of the telson in the late zoealstages Whereas this process is converted toa spine in Neotrypaea uncinata at ZV and Cal-lianassa sl petalura at ZIV the anomuranhair is retained at ZIV in Callichirus majorand C islagrande (see Rodrigues 1976Strasser and Felder 1999a) Although notmentioned specifically in the descriptions theanomuran hair appears to be present in illus-trations of Callichirus sp at ZIII and Cgarthii at ZV (Rodrigues 1976 fig 35 Asteand Retamal 1983 fig 8Q) A bilobed epi-pod on the first maxilliped of ZI is consistentlyfound in Callichirus but is also present at ZIin Lepidophthalmus sinuensis (see Nates etal 1997) Other callianassids acquire thischaracter at ZII (L louisianensis Nates et al1997 Callianassa sl kewlramanii Sankolliand Shenoy 1975) at ZV (Neotrypaea unci-nata Aste and Retamal 1984) or at the de-capodid stage (Sergio mirim Rodrigues 1984C sl petalura Konishi et al 1990) Thusthe number of zoeal stages and setation of thetelson may serve as the best characters to dis-tinguish larvae of Callichirus from other gen-era However since larval descriptions arelacking for many callianassid species it is un-known if these characters (in combination)will prove to be diagnostic until descriptionsfor other species are published
ACKNOWLEDGEMENTS
We thank Robert Bourgeois Barbara Hasek JimMaKinster Sergio Nates and Kurt Strasser who pro-vided assistance in the field and laboratory and MaryE Rice Director of the Smithsonian Marine StationLink Port Florida who facilitated access to station fa-cilities We also thank R Bauer R Jaeger R Lemaitreand G Watson for their early comments on this manu-script and Raymond B Manning who facilitated fund-ing and gave advice on collecting sites Direct supportfor this study was provided to Karen M Strasser by adoctoral fellowship from the University of Louisianaa graduate student research grant from the LUMCON(Louisiana Universities Marine Consortium) Founda-tion and a summer research award from the Smith-sonian Marine Station Additional support for field andlab activities was provided to D L Felder under Loui-siana Sea Grant College Program Grant RCFB-21 USFish and Wildlife Service Cooperative Agreement 14-16-0009-89-96 (Task Order No 6) US Department of En-ergy grant DE-FG02-97ER12220 and several small proj-ect grants from the Smithsonian Marine Station Views
expressed herein are those of the authors and do not nec-essarily reflect views of supporting government agenciesor subagencies mentioned above This is contributionNumber 466 of the Smithsonian Marine Station and con-tribution Number 67 of the ULL Laboratory for Crus-tacean Research All experiments conducted in this studycomply with current applicable state and federal laws
LITERATURE CITED
Aste A and M Retamal 1983 Desarrollo larval deCallianassa garthi Retamal 1975 bajo condiciones delaboratoriomdashCiencia y Tecnologiacutea del Mar CONA 75ndash26
mdashmdashmdash and mdashmdashmdash 1984 Desarrollo larval de Cal-lianassa uncinata H Milne Edwards1837 (DecapodaCallianassidae) bajo condiciones de laboratoriomdashGayana Zooloacutegica 48 41ndash56
Felder D L 1978 Osmotic and ionic regulation in sev-eral western Atlantic Callianassidae (Crustacea De-capoda Thalassinidea)mdashBiological Bulletin 154409ndash429
mdashmdashmdash and R B Griffis 1994 Dominant infaunalcommunities at risk in shoreline habitats burrowingthalassinid CrustaceamdashOCS Study number MMS94ndash0007 United States Department of the InteriorMinerals Management Service Gulf of Mexico OCSRegional Office New Orleans Louisiana Pp 1ndash87
Forbes A T 1973 An unusual abbreviated larval lifehistory in the estuarine burrowing prawn Callianassakraussi (Crustacea Decapoda Thalassinidea)mdashMa-rine Biology 22 361ndash365
Konishi K 1989 Larval development of the mudshrimp Upogebia (Upogebia) major (de Haan) (Crus-tacea Thalassinidea Upogebiidae) under laboratoryconditions with comments on larval characters of tha-lassinid familiesmdashBulletin of the National ResearchInstitute of Aquaculture 15 1ndash17
mdashmdashmdash R Quintana and Y Fukuda 1990 A completedescription of larval stages of the ghost shrimp Cal-lianassa petalura Stimpson (Crustacea ThalassinideaCallianassidae) under laboratory conditionsmdashBulletinof the National Research Institute of Aquaculture 1727ndash49
Manning R B and D L Felder 1986 The status ofthe callianassid genus Callichirus Stimpson 1866(Crustacea Decapoda Thalassinidea)mdashProceedingsof the Biological Society of Washington 99 437ndash443
mdashmdashmdash and mdashmdashmdash 1991 Revision of the AmericanCallianassidae (Crustacea Decapoda Thalassinidea)mdashProceedings of the Biological Society of Washington104 764ndash794
Nates S F D L Felder and R Lemaitre 1997 Com-parative larval development in two species of the bur-rowing ghost shrimp genus Lepidophthalmus (Decap-oda Callianassidae)mdashJournal of Crustacean Biology17 497ndash519
Rodrigues S de A 1976 Sobre a reproduccedilatildeo embri-ologia e desenvolvimento larval de Callichirus majorSay 1818 (Crustacea Decapoda Thalassinidea)mdashBo-letim de Zoologia Universidade de Satildeo Paulo 185ndash104
mdashmdashmdash 1984 Desenvolvimento poacutes-embrionaacuterio deCallichirus mirim (Rodrigues 1971) obtido emcondiccedilotildees artificiais (Crustacea Decapoda Tha-lassinidea)mdashBoletim de Zoologia Universidade deSatildeo Paulo 8 239ndash256
STRASSER AND FELDER LARVAL DEVELOPMENT IN CALLICHIRUS ISLAGRANDE 117
Sankolli K N and S Shenoy 1975 Larval develop-ment of mud shrimp Callianassa (Callichirus) kewal-ramanii Sankolli in the laboratory (Crustacea Decap-oda)mdashBulletin of the Department of Marine ScienceUniversity of Cochin 4 704ndash720
Staton J L and D L Felder 1995 Genetic variationin populations of the ghost shrimp genus Callichirus(Crustacea Decapoda Thalassinoidea) in the westernAtlantic and Gulf of MexicomdashBulletin of Marine Sci-ence 56 523ndash536
Strasser K M and D L Felder 1998 Settlement cuesin successive developmental stages of the ghost shrimpCallichirus major and C islagrande (Crustacea De-capoda Thalassinidea)mdashMarine Biology
mdashmdashmdash and mdashmdashmdash 1999a Larval development of twopopulations of the ghost shrimp Callichirus major (De-capoda Thalassinidea)mdashJournal of Crustacean Biology
mdashmdashmdash and mdashmdashmdash 1999b Sand as a stimulus for set-tlement in the ghost shrimp Callichirus major (Say)and C islagrande (Schmitt) (Crustacea DecapodaThalassinidea)mdashJournal of Experimental Marine Bi-ology and Ecology 239 211ndash222
mdashmdashmdash and mdashmdashmdash 1999c Settlement cues in theFlorida Atlantic population of the ghost shrimp Cal-lichirus major (Crustacea Decapoda Thalassinidea)mdashMarine Ecology Progress Series
Thessalou-Legaki M 1990 Advanced larval develop-ment of Callianassa tyrrhena (Decapoda Tha-lassinidea) and the effect of environmental factorsmdashJournal of Crustacean Biology 10 659ndash666
RECEIVED 19 September 1998ACCEPTED 3 December 1998
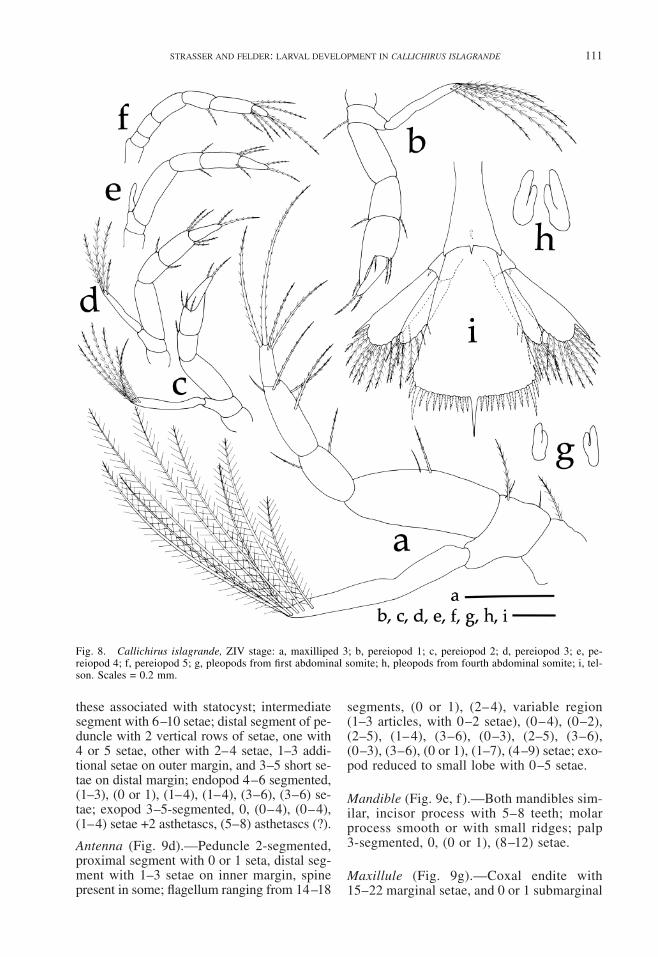
STRASSER AND FELDER LARVAL DEVELOPMENT IN CALLICHIRUS ISLAGRANDE 111
these associated with statocyst intermediatesegment with 6ndash10 setae distal segment of pe-duncle with 2 vertical rows of setae one with4 or 5 setae other with 2ndash4 setae 1ndash3 addi-tional setae on outer margin and 3ndash5 short se-tae on distal margin endopod 4ndash6 segmented(1ndash3) (0 or 1) (1ndash4) (1ndash4) (3ndash6) (3ndash6) se-tae exopod 3ndash5-segmented 0 (0ndash4) (0ndash4)(1ndash4) setae +2 asthetascs (5ndash8) asthetascs ()
Antenna (Fig 9d)mdashPeduncle 2-segmentedproximal segment with 0 or 1 seta distal seg-ment with 1ndash3 setae on inner margin spinepresent in some flagellum ranging from 14ndash18
segments (0 or 1) (2ndash4) variable region(1ndash3 articles with 0ndash2 setae) (0ndash4) (0ndash2)(2ndash5) (1ndash4) (3ndash6) (0ndash3) (2ndash5) (3ndash6)(0ndash3) (3ndash6) (0 or 1) (1ndash7) (4ndash9) setae exo-pod reduced to small lobe with 0ndash5 setae
Mandible (Fig 9e f )mdashBoth mandibles sim-ilar incisor process with 5ndash8 teeth molarprocess smooth or with small ridges palp3-segmented 0 (0 or 1) (8ndash12) setae
Maxillule (Fig 9g)mdashCoxal endite with15ndash22 marginal setae and 0 or 1 submarginal
Fig 8 Callichirus islagrande ZIV stage a maxilliped 3 b pereiopod 1 c pereiopod 2 d pereiopod 3 e pe-reiopod 4 f pereiopod 5 g pleopods from first abdominal somite h pleopods from fourth abdominal somite i tel-son Scales = 02 mm
112 JOURNAL OF CRUSTACEAN BIOLOGY VOL 20 NO 1 2000
seta basal endite with 3ndash5 plumose setae14ndash19 dentate spines 0ndash2 plumose setae oninner margin endopod reduced unsegmentedwith 0ndash8 setae
Maxilla (Fig 9h)mdashCoxal endite trilobed(17ndash21)+(7ndash10)+(6ndash9) setae basal enditebilobed 8ndash10 setae on proximal lobe 8ndash15setae on distal lobe endopod reduced with
Fig 9 Callichirus islagrande ZV stage a one example of the antenna b one example of pleopods Decapodidstage c antennule d antenna e right mandible f left mandible g maxillule h maxilla (plumose condition notshown) i maxilliped 1 j maxilliped 2 Scales = 02 mm
STRASSER AND FELDER LARVAL DEVELOPMENT IN CALLICHIRUS ISLAGRANDE 113
5ndash11 setae scaphognathite with 37ndash45 se-tae all setae plumose
Maxilliped 1 (Fig 9i)mdashCoxa with 8ndash12 se-tae basis highly setose with 27ndash39 setae en-dopod reduced with 0ndash11 setae exopod un-segmented 6ndash14 setae bilobed epipod with-out setae
Maxilliped 2 (Fig 9j)mdashCoxa partially fusedto basis with 1ndash4 setae basis with 3ndash5 se-
tae endopod 4-segmented (16ndash23) (0 or 1)(6ndash11) (4ndash8) setae exopod with 0ndash8 setaeepipod present but without setae
Maxilliped 3 (Fig 10a)mdashCoxa partiallyfused to basis with 0ndash3 setae basis with 1ndash4setae endopod 5-segmented (11ndash19) (9ndash15)(7ndash11) (8ndash21) (5ndash11) setae exopod usuallyreduced and without setae occasionally with2ndash9 setae epipod present
Fig 10 Callichirus islagrande decapodid stage a maxilliped 3 b pereiopod 1 c pereiopod 2 d pereiopod 3e pereiopod 4 f pereiopod 5 g pleopods from first abdominal somite h pleopods from fourth abdominal somitei telson Scales = 02 mm
114 JOURNAL OF CRUSTACEAN BIOLOGY VOL 20 NO 1 2000
Pereiopod 1 (Fig 10b)mdashChelate biramous7-segmented coxa with 0ndash6 setae basis with2ndash5 setae ischium with 4ndash7 setae and 0ndash3spines merus with 5ndash8 setae carpus with10ndash14 setae propodus with 27ndash37 setae and3ndash4 spines on inner margin of immovable fin-ger dactylus with 21ndash33 setae exopod usu-ally reduced and without setae occasionallywith 4ndash7 setae
Pereiopod 2 (Fig 10c)mdashChelate biramous7-segmented coxa with 2ndash5 setae basis with1ndash5 setae ischium with 4ndash10 setae meruswith 15ndash20 setae carpus with 8ndash12 setaepropodus with 30ndash40 setae and 1ndash4 spineson inner margin of immovable finger dacty-lus with 21ndash30 setae exopod usually reduced0ndash7 setae
Pereiopod 3 (Fig 10d)mdashBiramous 7-seg-mented coxa with 2ndash5 setae basis with 1ndash3setae ischium with 5ndash8 setae merus with6ndash9 setae carpus with 7ndash11 setae propodusbladelike with 25ndash34 setae 1 spine on innerdistal margin dactylus with 15ndash21 setae exo-pod usually reduced 0ndash5 setae
Pereiopod 4 (Fig 10e)mdashBiramous 7-seg-mented coxa with 2ndash7 setae basis with 1ndash4setae ischium with 3ndash8 setae merus with5ndash11 setae carpus with 4ndash9 setae propoduswith 27ndash38 setae 1 spine on inner distal mar-gin dactylus with 15ndash19 setae exopod usu-ally reduced with 0ndash2 setae
Pereiopod 5 (Fig 10f )mdashCheliform unira-mous 7-segmented coxa with 2ndash6 setae ba-sis with l or 2 setae ischium with 2ndash5 setaemerus with 5ndash9 setae carpus with 4ndash7 se-tae propodus with 22ndash30 setae 1 spine onimmovable finger dactylus with 6ndash10 setaeexopod absent
Pleopods (somite 2) (Fig 10g)mdashBiramouscoxa partially fused to basis without setaebasis with 0ndash3 setae endopod with 3ndash5hooked setae on appendix interna 10ndash17 mar-ginal plumose setae 0ndash2 submarginal nakedsetae exopod with 21ndash27 marginal plumosesetae and 0ndash1 submarginal naked seta
Pleopods (somites 3ndash5) (Fig 10h)mdashBiramouscoxa without setae basis with 3ndash6 marginalplumose setae endopod with 3ndash6 hooked se-tae on appendix interna 10ndash18 marginalplumose setae and 0ndash3 naked submarginal se-tae exopod with 20ndash29 marginal plumose se-tae and 0 or 1 submarginal naked seta
Telson and Uropods (Fig 10i)mdashWith 24ndash36processes on posterior margin median pro-cess usually absent or reduced uropods bi-ramous endopod with 19ndash25 plumose setaeexopod with 35ndash58 plumose setae protopodwith 3ndash5 setae
DISCUSSION
As in other species of Callichirus (see Ro-drigues 1976 Aste and Retamal 1983Strasser and Felder in press b) larval devel-opment of C islagrande included four or fivezoeal stages before metamorphosis to the de-capodid ZI larvae may be identified by thepresence of an elongate rounded rostrumsessile eyes and a lack of pereiopodspleopods or uropods (Fig 1a) At ZII lar-vae acquired biramous buds of pereiopods 1and 2 eyes were separated from the carapaceand the rostrum became flattened and serratedlaterally (Fig 1b) Buds of pleopods on ab-dominal segments 2ndash5 pereiopods 3ndash5 anduropods were developed at ZIII (Fig 1c) Inaddition to further development of structuresmentioned thus far ZIV larvae could be dis-tinguished by presence of a distal spine on theexopod and setae on the endopod of each uro-pod (Fig 1d) When it occurred ZV typicallyresembled ZIV but varied widely in mor-phology Appendages at ZV resembled thoseof ZIV the decapodid stage or some grada-tion between the two (see Figs 9a b for ex-amples) The decapodid stage was similar toother species of Callichirus in the reductionof larval characters such as the elongate ros-trum dorsal spines on the abdomen and ex-opods of the pereiopods (Fig le) Acquisitionof fan-like pleopods marked the transition toa primarily abdominal mode of swimming atthis stage
In the Florida Atlantic population of C ma-jor both salinity and presence of putative set-tlement cues at ZIV impacted the observednumber of zoeal stages and the duration ofZIV (Strasser and Felder in preparation1999c) In C islagrande the percentage oflarvae that passed through five versus fourzoeal stages varied (9ndash89) between parentalfemales but was not shown to depend onsalinity or presence of various putative set-tlement cues (Strasser and Felder 1998 inpreparation) Larvae that molted from ZIVto ZV were usually normal morphologicallyand had mortality similar to those that molteddirectly from ZIV to D In contrast larvae of
STRASSER AND FELDER LARVAL DEVELOPMENT IN CALLICHIRUS ISLAGRANDE 115
the Florida Atlantic C major that molted toZV had a higher incidence of mortality anddeformity at D than did those that had by-passed ZV (Strasser and Felder 1999a c)
The number of zoeal stages appears to beless variable in Callichirus garthi and theGulf of Mexico population of C major thanin congeneric populations studied to dateCallichirus garthi was reported to have fivezoeal stages and a long larval duration reach-ing the decapodid stage 48 days after hatch-ing (Aste and Retamal 1983) Developmentof the Gulf population of C major wasshorter in duration (decapodid reached about10 days after hatching) and typically includedfour zoeal stages (Strasser and Felder 1999a)animals rarely passed through three or fivezoeal stages before the molt to the decapodidAlthough the number of stages is reducedfrom that observed in C garthi developmentin the Gulf population of C major cannot beclassified as abbreviated There are species inboth the Callianassinae (Callianassa slkraussi Stebbing C sl kewalramanii and Csl tyrrhena (Petagna)) and Callichirinae(Sergio mirim Lepidophthalmus sinuensisand L louisianensis) that have only two zoealstages before the molt to the decapodid(Forbes 1973 Sankolli and Shenoy 1975Rodrigues 1984 Thessalou-Legaki 1990Nates et al 1997)
Compared to other species of the genusCallichirus C islagrande is morphologicallycloser to C garthi (from Chile) than to theGulf of Mexico population of C major whichoverlaps in distribution with C islagrande Inthe zoeal phase of C islagrande and Cgarthi the telson is concave and the outer-most process on the ventral margin is articu-lated In both C major and Callichirus sp theposterior margin of the telson is straight andthe outermost process is unarticulated duringthe zoeal stages (Rodrigues 1976 Strasserand Felder 1999a) There are also differencesin spination of the abdomen In zoeae of Cislagrande and C garthi the dorsal spine ofabdominal segment 3 is not longer than thosefound on segments 4 and 5 (Aste and Reta-mal 1983) In contrast the dorsal spine of ab-dominal segment 3 is approximately twice aslong as the dorsal spines on segments 4 and5 in early zoeal stages of C major and Cal-lichirus sp (Rodrigues 1976 Aste and Re-tamal 1983 Strasser and Felder 1999a) Thegreatest difference between these groups is
the number of pleopod pairs at zoeal and de-capodid stages Callichirus islagrande and Cgarthi have pleopods on abdominal segments2ndash5 while pleopods are present only on ab-dominal segments 3ndash5 in C major and Cal-lichirus sp (Rodrigues 1976 Aste and Re-tamal 1983 Strasser and Felder 1999a) Al-though Callichirus islagrande and C garthiare similar morphologically there are differ-ences between these two species Callichirusislagrande as well as C major and Cal-lichirus sp develops pleopod buds andchelate pereiopods at ZIII whereas C garthidoes not acquire these characters until ZIV
There are few callianassid species forwhich the larval development has been de-scribed (Sankolli and Shenoy 1975 Ro-drigues 1976 1984 Aste and Retamal 19831984 Konishi et al 1990 Nates et al 1997Strasser and Felder 1999a) all of which aremembers of the Callichirinae and Cal-lianassinae Among species there is a greatdeal of overlap in larval characters makingit difficult to draw distinctions between thesesubfamilies At present information on lar-val development of species in the Callichiri-nae is limited to larval descriptions of Cal-lichirus major C garthi Callichirus sp Cislagrande Sergio mirim Lepidophthalmuslouisianensis and L sinuensis (see Ro-drigues 1976 1984 Aste and Retamal 1983Nates et al 1997 Strasser and Felder1999a) As mentioned above species of Cal-lichirus have been shown to have four to fivezoeal stages while the other members exhibitabbreviated development that consists of onlytwo zoeal stages The most obvious differencebetween larvae of the species of Callichirinaewith abbreviated development is the numberof pleopod pairs Larvae of S mirim havepleopods on abdominal segments 3ndash5whereas L louisianensis and L sinuensishave pleopods on segments 2ndash5 (Rodrigues1984 Nates et al 1997)
Ghost shrimps of the genus Callichirus maybe distinguished from other genera of the Cal-lianassidae by the cuticular pattern on the dor-sal surface of the abdomen and by the strap-like shape of the uropodal endopods (Manningand Felder 1991) Whereas these charactersmay be used to distinguish adults of the genusCallichirus they are not present at zoeal anddecapodid stages Larvae may be distin-guished from all other callianassid genera byhaving four to five zoeal stages and numer-
116 JOURNAL OF CRUSTACEAN BIOLOGY VOL 20 NO 1 2000
ous processes (12ndash25+1+12ndash25) on the pos-terior margin of the telson during the earlyzoeal stages Larvae of Callichirus may alsodiffer from other callianassids with extendeddevelopment by the retention of the anomu-ran hair as the second process on the poste-rior margin of the telson in the late zoealstages Whereas this process is converted toa spine in Neotrypaea uncinata at ZV and Cal-lianassa sl petalura at ZIV the anomuranhair is retained at ZIV in Callichirus majorand C islagrande (see Rodrigues 1976Strasser and Felder 1999a) Although notmentioned specifically in the descriptions theanomuran hair appears to be present in illus-trations of Callichirus sp at ZIII and Cgarthii at ZV (Rodrigues 1976 fig 35 Asteand Retamal 1983 fig 8Q) A bilobed epi-pod on the first maxilliped of ZI is consistentlyfound in Callichirus but is also present at ZIin Lepidophthalmus sinuensis (see Nates etal 1997) Other callianassids acquire thischaracter at ZII (L louisianensis Nates et al1997 Callianassa sl kewlramanii Sankolliand Shenoy 1975) at ZV (Neotrypaea unci-nata Aste and Retamal 1984) or at the de-capodid stage (Sergio mirim Rodrigues 1984C sl petalura Konishi et al 1990) Thusthe number of zoeal stages and setation of thetelson may serve as the best characters to dis-tinguish larvae of Callichirus from other gen-era However since larval descriptions arelacking for many callianassid species it is un-known if these characters (in combination)will prove to be diagnostic until descriptionsfor other species are published
ACKNOWLEDGEMENTS
We thank Robert Bourgeois Barbara Hasek JimMaKinster Sergio Nates and Kurt Strasser who pro-vided assistance in the field and laboratory and MaryE Rice Director of the Smithsonian Marine StationLink Port Florida who facilitated access to station fa-cilities We also thank R Bauer R Jaeger R Lemaitreand G Watson for their early comments on this manu-script and Raymond B Manning who facilitated fund-ing and gave advice on collecting sites Direct supportfor this study was provided to Karen M Strasser by adoctoral fellowship from the University of Louisianaa graduate student research grant from the LUMCON(Louisiana Universities Marine Consortium) Founda-tion and a summer research award from the Smith-sonian Marine Station Additional support for field andlab activities was provided to D L Felder under Loui-siana Sea Grant College Program Grant RCFB-21 USFish and Wildlife Service Cooperative Agreement 14-16-0009-89-96 (Task Order No 6) US Department of En-ergy grant DE-FG02-97ER12220 and several small proj-ect grants from the Smithsonian Marine Station Views
expressed herein are those of the authors and do not nec-essarily reflect views of supporting government agenciesor subagencies mentioned above This is contributionNumber 466 of the Smithsonian Marine Station and con-tribution Number 67 of the ULL Laboratory for Crus-tacean Research All experiments conducted in this studycomply with current applicable state and federal laws
LITERATURE CITED
Aste A and M Retamal 1983 Desarrollo larval deCallianassa garthi Retamal 1975 bajo condiciones delaboratoriomdashCiencia y Tecnologiacutea del Mar CONA 75ndash26
mdashmdashmdash and mdashmdashmdash 1984 Desarrollo larval de Cal-lianassa uncinata H Milne Edwards1837 (DecapodaCallianassidae) bajo condiciones de laboratoriomdashGayana Zooloacutegica 48 41ndash56
Felder D L 1978 Osmotic and ionic regulation in sev-eral western Atlantic Callianassidae (Crustacea De-capoda Thalassinidea)mdashBiological Bulletin 154409ndash429
mdashmdashmdash and R B Griffis 1994 Dominant infaunalcommunities at risk in shoreline habitats burrowingthalassinid CrustaceamdashOCS Study number MMS94ndash0007 United States Department of the InteriorMinerals Management Service Gulf of Mexico OCSRegional Office New Orleans Louisiana Pp 1ndash87
Forbes A T 1973 An unusual abbreviated larval lifehistory in the estuarine burrowing prawn Callianassakraussi (Crustacea Decapoda Thalassinidea)mdashMa-rine Biology 22 361ndash365
Konishi K 1989 Larval development of the mudshrimp Upogebia (Upogebia) major (de Haan) (Crus-tacea Thalassinidea Upogebiidae) under laboratoryconditions with comments on larval characters of tha-lassinid familiesmdashBulletin of the National ResearchInstitute of Aquaculture 15 1ndash17
mdashmdashmdash R Quintana and Y Fukuda 1990 A completedescription of larval stages of the ghost shrimp Cal-lianassa petalura Stimpson (Crustacea ThalassinideaCallianassidae) under laboratory conditionsmdashBulletinof the National Research Institute of Aquaculture 1727ndash49
Manning R B and D L Felder 1986 The status ofthe callianassid genus Callichirus Stimpson 1866(Crustacea Decapoda Thalassinidea)mdashProceedingsof the Biological Society of Washington 99 437ndash443
mdashmdashmdash and mdashmdashmdash 1991 Revision of the AmericanCallianassidae (Crustacea Decapoda Thalassinidea)mdashProceedings of the Biological Society of Washington104 764ndash794
Nates S F D L Felder and R Lemaitre 1997 Com-parative larval development in two species of the bur-rowing ghost shrimp genus Lepidophthalmus (Decap-oda Callianassidae)mdashJournal of Crustacean Biology17 497ndash519
Rodrigues S de A 1976 Sobre a reproduccedilatildeo embri-ologia e desenvolvimento larval de Callichirus majorSay 1818 (Crustacea Decapoda Thalassinidea)mdashBo-letim de Zoologia Universidade de Satildeo Paulo 185ndash104
mdashmdashmdash 1984 Desenvolvimento poacutes-embrionaacuterio deCallichirus mirim (Rodrigues 1971) obtido emcondiccedilotildees artificiais (Crustacea Decapoda Tha-lassinidea)mdashBoletim de Zoologia Universidade deSatildeo Paulo 8 239ndash256
STRASSER AND FELDER LARVAL DEVELOPMENT IN CALLICHIRUS ISLAGRANDE 117
Sankolli K N and S Shenoy 1975 Larval develop-ment of mud shrimp Callianassa (Callichirus) kewal-ramanii Sankolli in the laboratory (Crustacea Decap-oda)mdashBulletin of the Department of Marine ScienceUniversity of Cochin 4 704ndash720
Staton J L and D L Felder 1995 Genetic variationin populations of the ghost shrimp genus Callichirus(Crustacea Decapoda Thalassinoidea) in the westernAtlantic and Gulf of MexicomdashBulletin of Marine Sci-ence 56 523ndash536
Strasser K M and D L Felder 1998 Settlement cuesin successive developmental stages of the ghost shrimpCallichirus major and C islagrande (Crustacea De-capoda Thalassinidea)mdashMarine Biology
mdashmdashmdash and mdashmdashmdash 1999a Larval development of twopopulations of the ghost shrimp Callichirus major (De-capoda Thalassinidea)mdashJournal of Crustacean Biology
mdashmdashmdash and mdashmdashmdash 1999b Sand as a stimulus for set-tlement in the ghost shrimp Callichirus major (Say)and C islagrande (Schmitt) (Crustacea DecapodaThalassinidea)mdashJournal of Experimental Marine Bi-ology and Ecology 239 211ndash222
mdashmdashmdash and mdashmdashmdash 1999c Settlement cues in theFlorida Atlantic population of the ghost shrimp Cal-lichirus major (Crustacea Decapoda Thalassinidea)mdashMarine Ecology Progress Series
Thessalou-Legaki M 1990 Advanced larval develop-ment of Callianassa tyrrhena (Decapoda Tha-lassinidea) and the effect of environmental factorsmdashJournal of Crustacean Biology 10 659ndash666
RECEIVED 19 September 1998ACCEPTED 3 December 1998

112 JOURNAL OF CRUSTACEAN BIOLOGY VOL 20 NO 1 2000
seta basal endite with 3ndash5 plumose setae14ndash19 dentate spines 0ndash2 plumose setae oninner margin endopod reduced unsegmentedwith 0ndash8 setae
Maxilla (Fig 9h)mdashCoxal endite trilobed(17ndash21)+(7ndash10)+(6ndash9) setae basal enditebilobed 8ndash10 setae on proximal lobe 8ndash15setae on distal lobe endopod reduced with
Fig 9 Callichirus islagrande ZV stage a one example of the antenna b one example of pleopods Decapodidstage c antennule d antenna e right mandible f left mandible g maxillule h maxilla (plumose condition notshown) i maxilliped 1 j maxilliped 2 Scales = 02 mm
STRASSER AND FELDER LARVAL DEVELOPMENT IN CALLICHIRUS ISLAGRANDE 113
5ndash11 setae scaphognathite with 37ndash45 se-tae all setae plumose
Maxilliped 1 (Fig 9i)mdashCoxa with 8ndash12 se-tae basis highly setose with 27ndash39 setae en-dopod reduced with 0ndash11 setae exopod un-segmented 6ndash14 setae bilobed epipod with-out setae
Maxilliped 2 (Fig 9j)mdashCoxa partially fusedto basis with 1ndash4 setae basis with 3ndash5 se-
tae endopod 4-segmented (16ndash23) (0 or 1)(6ndash11) (4ndash8) setae exopod with 0ndash8 setaeepipod present but without setae
Maxilliped 3 (Fig 10a)mdashCoxa partiallyfused to basis with 0ndash3 setae basis with 1ndash4setae endopod 5-segmented (11ndash19) (9ndash15)(7ndash11) (8ndash21) (5ndash11) setae exopod usuallyreduced and without setae occasionally with2ndash9 setae epipod present
Fig 10 Callichirus islagrande decapodid stage a maxilliped 3 b pereiopod 1 c pereiopod 2 d pereiopod 3e pereiopod 4 f pereiopod 5 g pleopods from first abdominal somite h pleopods from fourth abdominal somitei telson Scales = 02 mm
114 JOURNAL OF CRUSTACEAN BIOLOGY VOL 20 NO 1 2000
Pereiopod 1 (Fig 10b)mdashChelate biramous7-segmented coxa with 0ndash6 setae basis with2ndash5 setae ischium with 4ndash7 setae and 0ndash3spines merus with 5ndash8 setae carpus with10ndash14 setae propodus with 27ndash37 setae and3ndash4 spines on inner margin of immovable fin-ger dactylus with 21ndash33 setae exopod usu-ally reduced and without setae occasionallywith 4ndash7 setae
Pereiopod 2 (Fig 10c)mdashChelate biramous7-segmented coxa with 2ndash5 setae basis with1ndash5 setae ischium with 4ndash10 setae meruswith 15ndash20 setae carpus with 8ndash12 setaepropodus with 30ndash40 setae and 1ndash4 spineson inner margin of immovable finger dacty-lus with 21ndash30 setae exopod usually reduced0ndash7 setae
Pereiopod 3 (Fig 10d)mdashBiramous 7-seg-mented coxa with 2ndash5 setae basis with 1ndash3setae ischium with 5ndash8 setae merus with6ndash9 setae carpus with 7ndash11 setae propodusbladelike with 25ndash34 setae 1 spine on innerdistal margin dactylus with 15ndash21 setae exo-pod usually reduced 0ndash5 setae
Pereiopod 4 (Fig 10e)mdashBiramous 7-seg-mented coxa with 2ndash7 setae basis with 1ndash4setae ischium with 3ndash8 setae merus with5ndash11 setae carpus with 4ndash9 setae propoduswith 27ndash38 setae 1 spine on inner distal mar-gin dactylus with 15ndash19 setae exopod usu-ally reduced with 0ndash2 setae
Pereiopod 5 (Fig 10f )mdashCheliform unira-mous 7-segmented coxa with 2ndash6 setae ba-sis with l or 2 setae ischium with 2ndash5 setaemerus with 5ndash9 setae carpus with 4ndash7 se-tae propodus with 22ndash30 setae 1 spine onimmovable finger dactylus with 6ndash10 setaeexopod absent
Pleopods (somite 2) (Fig 10g)mdashBiramouscoxa partially fused to basis without setaebasis with 0ndash3 setae endopod with 3ndash5hooked setae on appendix interna 10ndash17 mar-ginal plumose setae 0ndash2 submarginal nakedsetae exopod with 21ndash27 marginal plumosesetae and 0ndash1 submarginal naked seta
Pleopods (somites 3ndash5) (Fig 10h)mdashBiramouscoxa without setae basis with 3ndash6 marginalplumose setae endopod with 3ndash6 hooked se-tae on appendix interna 10ndash18 marginalplumose setae and 0ndash3 naked submarginal se-tae exopod with 20ndash29 marginal plumose se-tae and 0 or 1 submarginal naked seta
Telson and Uropods (Fig 10i)mdashWith 24ndash36processes on posterior margin median pro-cess usually absent or reduced uropods bi-ramous endopod with 19ndash25 plumose setaeexopod with 35ndash58 plumose setae protopodwith 3ndash5 setae
DISCUSSION
As in other species of Callichirus (see Ro-drigues 1976 Aste and Retamal 1983Strasser and Felder in press b) larval devel-opment of C islagrande included four or fivezoeal stages before metamorphosis to the de-capodid ZI larvae may be identified by thepresence of an elongate rounded rostrumsessile eyes and a lack of pereiopodspleopods or uropods (Fig 1a) At ZII lar-vae acquired biramous buds of pereiopods 1and 2 eyes were separated from the carapaceand the rostrum became flattened and serratedlaterally (Fig 1b) Buds of pleopods on ab-dominal segments 2ndash5 pereiopods 3ndash5 anduropods were developed at ZIII (Fig 1c) Inaddition to further development of structuresmentioned thus far ZIV larvae could be dis-tinguished by presence of a distal spine on theexopod and setae on the endopod of each uro-pod (Fig 1d) When it occurred ZV typicallyresembled ZIV but varied widely in mor-phology Appendages at ZV resembled thoseof ZIV the decapodid stage or some grada-tion between the two (see Figs 9a b for ex-amples) The decapodid stage was similar toother species of Callichirus in the reductionof larval characters such as the elongate ros-trum dorsal spines on the abdomen and ex-opods of the pereiopods (Fig le) Acquisitionof fan-like pleopods marked the transition toa primarily abdominal mode of swimming atthis stage
In the Florida Atlantic population of C ma-jor both salinity and presence of putative set-tlement cues at ZIV impacted the observednumber of zoeal stages and the duration ofZIV (Strasser and Felder in preparation1999c) In C islagrande the percentage oflarvae that passed through five versus fourzoeal stages varied (9ndash89) between parentalfemales but was not shown to depend onsalinity or presence of various putative set-tlement cues (Strasser and Felder 1998 inpreparation) Larvae that molted from ZIVto ZV were usually normal morphologicallyand had mortality similar to those that molteddirectly from ZIV to D In contrast larvae of
STRASSER AND FELDER LARVAL DEVELOPMENT IN CALLICHIRUS ISLAGRANDE 115
the Florida Atlantic C major that molted toZV had a higher incidence of mortality anddeformity at D than did those that had by-passed ZV (Strasser and Felder 1999a c)
The number of zoeal stages appears to beless variable in Callichirus garthi and theGulf of Mexico population of C major thanin congeneric populations studied to dateCallichirus garthi was reported to have fivezoeal stages and a long larval duration reach-ing the decapodid stage 48 days after hatch-ing (Aste and Retamal 1983) Developmentof the Gulf population of C major wasshorter in duration (decapodid reached about10 days after hatching) and typically includedfour zoeal stages (Strasser and Felder 1999a)animals rarely passed through three or fivezoeal stages before the molt to the decapodidAlthough the number of stages is reducedfrom that observed in C garthi developmentin the Gulf population of C major cannot beclassified as abbreviated There are species inboth the Callianassinae (Callianassa slkraussi Stebbing C sl kewalramanii and Csl tyrrhena (Petagna)) and Callichirinae(Sergio mirim Lepidophthalmus sinuensisand L louisianensis) that have only two zoealstages before the molt to the decapodid(Forbes 1973 Sankolli and Shenoy 1975Rodrigues 1984 Thessalou-Legaki 1990Nates et al 1997)
Compared to other species of the genusCallichirus C islagrande is morphologicallycloser to C garthi (from Chile) than to theGulf of Mexico population of C major whichoverlaps in distribution with C islagrande Inthe zoeal phase of C islagrande and Cgarthi the telson is concave and the outer-most process on the ventral margin is articu-lated In both C major and Callichirus sp theposterior margin of the telson is straight andthe outermost process is unarticulated duringthe zoeal stages (Rodrigues 1976 Strasserand Felder 1999a) There are also differencesin spination of the abdomen In zoeae of Cislagrande and C garthi the dorsal spine ofabdominal segment 3 is not longer than thosefound on segments 4 and 5 (Aste and Reta-mal 1983) In contrast the dorsal spine of ab-dominal segment 3 is approximately twice aslong as the dorsal spines on segments 4 and5 in early zoeal stages of C major and Cal-lichirus sp (Rodrigues 1976 Aste and Re-tamal 1983 Strasser and Felder 1999a) Thegreatest difference between these groups is
the number of pleopod pairs at zoeal and de-capodid stages Callichirus islagrande and Cgarthi have pleopods on abdominal segments2ndash5 while pleopods are present only on ab-dominal segments 3ndash5 in C major and Cal-lichirus sp (Rodrigues 1976 Aste and Re-tamal 1983 Strasser and Felder 1999a) Al-though Callichirus islagrande and C garthiare similar morphologically there are differ-ences between these two species Callichirusislagrande as well as C major and Cal-lichirus sp develops pleopod buds andchelate pereiopods at ZIII whereas C garthidoes not acquire these characters until ZIV
There are few callianassid species forwhich the larval development has been de-scribed (Sankolli and Shenoy 1975 Ro-drigues 1976 1984 Aste and Retamal 19831984 Konishi et al 1990 Nates et al 1997Strasser and Felder 1999a) all of which aremembers of the Callichirinae and Cal-lianassinae Among species there is a greatdeal of overlap in larval characters makingit difficult to draw distinctions between thesesubfamilies At present information on lar-val development of species in the Callichiri-nae is limited to larval descriptions of Cal-lichirus major C garthi Callichirus sp Cislagrande Sergio mirim Lepidophthalmuslouisianensis and L sinuensis (see Ro-drigues 1976 1984 Aste and Retamal 1983Nates et al 1997 Strasser and Felder1999a) As mentioned above species of Cal-lichirus have been shown to have four to fivezoeal stages while the other members exhibitabbreviated development that consists of onlytwo zoeal stages The most obvious differencebetween larvae of the species of Callichirinaewith abbreviated development is the numberof pleopod pairs Larvae of S mirim havepleopods on abdominal segments 3ndash5whereas L louisianensis and L sinuensishave pleopods on segments 2ndash5 (Rodrigues1984 Nates et al 1997)
Ghost shrimps of the genus Callichirus maybe distinguished from other genera of the Cal-lianassidae by the cuticular pattern on the dor-sal surface of the abdomen and by the strap-like shape of the uropodal endopods (Manningand Felder 1991) Whereas these charactersmay be used to distinguish adults of the genusCallichirus they are not present at zoeal anddecapodid stages Larvae may be distin-guished from all other callianassid genera byhaving four to five zoeal stages and numer-
116 JOURNAL OF CRUSTACEAN BIOLOGY VOL 20 NO 1 2000
ous processes (12ndash25+1+12ndash25) on the pos-terior margin of the telson during the earlyzoeal stages Larvae of Callichirus may alsodiffer from other callianassids with extendeddevelopment by the retention of the anomu-ran hair as the second process on the poste-rior margin of the telson in the late zoealstages Whereas this process is converted toa spine in Neotrypaea uncinata at ZV and Cal-lianassa sl petalura at ZIV the anomuranhair is retained at ZIV in Callichirus majorand C islagrande (see Rodrigues 1976Strasser and Felder 1999a) Although notmentioned specifically in the descriptions theanomuran hair appears to be present in illus-trations of Callichirus sp at ZIII and Cgarthii at ZV (Rodrigues 1976 fig 35 Asteand Retamal 1983 fig 8Q) A bilobed epi-pod on the first maxilliped of ZI is consistentlyfound in Callichirus but is also present at ZIin Lepidophthalmus sinuensis (see Nates etal 1997) Other callianassids acquire thischaracter at ZII (L louisianensis Nates et al1997 Callianassa sl kewlramanii Sankolliand Shenoy 1975) at ZV (Neotrypaea unci-nata Aste and Retamal 1984) or at the de-capodid stage (Sergio mirim Rodrigues 1984C sl petalura Konishi et al 1990) Thusthe number of zoeal stages and setation of thetelson may serve as the best characters to dis-tinguish larvae of Callichirus from other gen-era However since larval descriptions arelacking for many callianassid species it is un-known if these characters (in combination)will prove to be diagnostic until descriptionsfor other species are published
ACKNOWLEDGEMENTS
We thank Robert Bourgeois Barbara Hasek JimMaKinster Sergio Nates and Kurt Strasser who pro-vided assistance in the field and laboratory and MaryE Rice Director of the Smithsonian Marine StationLink Port Florida who facilitated access to station fa-cilities We also thank R Bauer R Jaeger R Lemaitreand G Watson for their early comments on this manu-script and Raymond B Manning who facilitated fund-ing and gave advice on collecting sites Direct supportfor this study was provided to Karen M Strasser by adoctoral fellowship from the University of Louisianaa graduate student research grant from the LUMCON(Louisiana Universities Marine Consortium) Founda-tion and a summer research award from the Smith-sonian Marine Station Additional support for field andlab activities was provided to D L Felder under Loui-siana Sea Grant College Program Grant RCFB-21 USFish and Wildlife Service Cooperative Agreement 14-16-0009-89-96 (Task Order No 6) US Department of En-ergy grant DE-FG02-97ER12220 and several small proj-ect grants from the Smithsonian Marine Station Views
expressed herein are those of the authors and do not nec-essarily reflect views of supporting government agenciesor subagencies mentioned above This is contributionNumber 466 of the Smithsonian Marine Station and con-tribution Number 67 of the ULL Laboratory for Crus-tacean Research All experiments conducted in this studycomply with current applicable state and federal laws
LITERATURE CITED
Aste A and M Retamal 1983 Desarrollo larval deCallianassa garthi Retamal 1975 bajo condiciones delaboratoriomdashCiencia y Tecnologiacutea del Mar CONA 75ndash26
mdashmdashmdash and mdashmdashmdash 1984 Desarrollo larval de Cal-lianassa uncinata H Milne Edwards1837 (DecapodaCallianassidae) bajo condiciones de laboratoriomdashGayana Zooloacutegica 48 41ndash56
Felder D L 1978 Osmotic and ionic regulation in sev-eral western Atlantic Callianassidae (Crustacea De-capoda Thalassinidea)mdashBiological Bulletin 154409ndash429
mdashmdashmdash and R B Griffis 1994 Dominant infaunalcommunities at risk in shoreline habitats burrowingthalassinid CrustaceamdashOCS Study number MMS94ndash0007 United States Department of the InteriorMinerals Management Service Gulf of Mexico OCSRegional Office New Orleans Louisiana Pp 1ndash87
Forbes A T 1973 An unusual abbreviated larval lifehistory in the estuarine burrowing prawn Callianassakraussi (Crustacea Decapoda Thalassinidea)mdashMa-rine Biology 22 361ndash365
Konishi K 1989 Larval development of the mudshrimp Upogebia (Upogebia) major (de Haan) (Crus-tacea Thalassinidea Upogebiidae) under laboratoryconditions with comments on larval characters of tha-lassinid familiesmdashBulletin of the National ResearchInstitute of Aquaculture 15 1ndash17
mdashmdashmdash R Quintana and Y Fukuda 1990 A completedescription of larval stages of the ghost shrimp Cal-lianassa petalura Stimpson (Crustacea ThalassinideaCallianassidae) under laboratory conditionsmdashBulletinof the National Research Institute of Aquaculture 1727ndash49
Manning R B and D L Felder 1986 The status ofthe callianassid genus Callichirus Stimpson 1866(Crustacea Decapoda Thalassinidea)mdashProceedingsof the Biological Society of Washington 99 437ndash443
mdashmdashmdash and mdashmdashmdash 1991 Revision of the AmericanCallianassidae (Crustacea Decapoda Thalassinidea)mdashProceedings of the Biological Society of Washington104 764ndash794
Nates S F D L Felder and R Lemaitre 1997 Com-parative larval development in two species of the bur-rowing ghost shrimp genus Lepidophthalmus (Decap-oda Callianassidae)mdashJournal of Crustacean Biology17 497ndash519
Rodrigues S de A 1976 Sobre a reproduccedilatildeo embri-ologia e desenvolvimento larval de Callichirus majorSay 1818 (Crustacea Decapoda Thalassinidea)mdashBo-letim de Zoologia Universidade de Satildeo Paulo 185ndash104
mdashmdashmdash 1984 Desenvolvimento poacutes-embrionaacuterio deCallichirus mirim (Rodrigues 1971) obtido emcondiccedilotildees artificiais (Crustacea Decapoda Tha-lassinidea)mdashBoletim de Zoologia Universidade deSatildeo Paulo 8 239ndash256
STRASSER AND FELDER LARVAL DEVELOPMENT IN CALLICHIRUS ISLAGRANDE 117
Sankolli K N and S Shenoy 1975 Larval develop-ment of mud shrimp Callianassa (Callichirus) kewal-ramanii Sankolli in the laboratory (Crustacea Decap-oda)mdashBulletin of the Department of Marine ScienceUniversity of Cochin 4 704ndash720
Staton J L and D L Felder 1995 Genetic variationin populations of the ghost shrimp genus Callichirus(Crustacea Decapoda Thalassinoidea) in the westernAtlantic and Gulf of MexicomdashBulletin of Marine Sci-ence 56 523ndash536
Strasser K M and D L Felder 1998 Settlement cuesin successive developmental stages of the ghost shrimpCallichirus major and C islagrande (Crustacea De-capoda Thalassinidea)mdashMarine Biology
mdashmdashmdash and mdashmdashmdash 1999a Larval development of twopopulations of the ghost shrimp Callichirus major (De-capoda Thalassinidea)mdashJournal of Crustacean Biology
mdashmdashmdash and mdashmdashmdash 1999b Sand as a stimulus for set-tlement in the ghost shrimp Callichirus major (Say)and C islagrande (Schmitt) (Crustacea DecapodaThalassinidea)mdashJournal of Experimental Marine Bi-ology and Ecology 239 211ndash222
mdashmdashmdash and mdashmdashmdash 1999c Settlement cues in theFlorida Atlantic population of the ghost shrimp Cal-lichirus major (Crustacea Decapoda Thalassinidea)mdashMarine Ecology Progress Series
Thessalou-Legaki M 1990 Advanced larval develop-ment of Callianassa tyrrhena (Decapoda Tha-lassinidea) and the effect of environmental factorsmdashJournal of Crustacean Biology 10 659ndash666
RECEIVED 19 September 1998ACCEPTED 3 December 1998

STRASSER AND FELDER LARVAL DEVELOPMENT IN CALLICHIRUS ISLAGRANDE 113
5ndash11 setae scaphognathite with 37ndash45 se-tae all setae plumose
Maxilliped 1 (Fig 9i)mdashCoxa with 8ndash12 se-tae basis highly setose with 27ndash39 setae en-dopod reduced with 0ndash11 setae exopod un-segmented 6ndash14 setae bilobed epipod with-out setae
Maxilliped 2 (Fig 9j)mdashCoxa partially fusedto basis with 1ndash4 setae basis with 3ndash5 se-
tae endopod 4-segmented (16ndash23) (0 or 1)(6ndash11) (4ndash8) setae exopod with 0ndash8 setaeepipod present but without setae
Maxilliped 3 (Fig 10a)mdashCoxa partiallyfused to basis with 0ndash3 setae basis with 1ndash4setae endopod 5-segmented (11ndash19) (9ndash15)(7ndash11) (8ndash21) (5ndash11) setae exopod usuallyreduced and without setae occasionally with2ndash9 setae epipod present
Fig 10 Callichirus islagrande decapodid stage a maxilliped 3 b pereiopod 1 c pereiopod 2 d pereiopod 3e pereiopod 4 f pereiopod 5 g pleopods from first abdominal somite h pleopods from fourth abdominal somitei telson Scales = 02 mm
114 JOURNAL OF CRUSTACEAN BIOLOGY VOL 20 NO 1 2000
Pereiopod 1 (Fig 10b)mdashChelate biramous7-segmented coxa with 0ndash6 setae basis with2ndash5 setae ischium with 4ndash7 setae and 0ndash3spines merus with 5ndash8 setae carpus with10ndash14 setae propodus with 27ndash37 setae and3ndash4 spines on inner margin of immovable fin-ger dactylus with 21ndash33 setae exopod usu-ally reduced and without setae occasionallywith 4ndash7 setae
Pereiopod 2 (Fig 10c)mdashChelate biramous7-segmented coxa with 2ndash5 setae basis with1ndash5 setae ischium with 4ndash10 setae meruswith 15ndash20 setae carpus with 8ndash12 setaepropodus with 30ndash40 setae and 1ndash4 spineson inner margin of immovable finger dacty-lus with 21ndash30 setae exopod usually reduced0ndash7 setae
Pereiopod 3 (Fig 10d)mdashBiramous 7-seg-mented coxa with 2ndash5 setae basis with 1ndash3setae ischium with 5ndash8 setae merus with6ndash9 setae carpus with 7ndash11 setae propodusbladelike with 25ndash34 setae 1 spine on innerdistal margin dactylus with 15ndash21 setae exo-pod usually reduced 0ndash5 setae
Pereiopod 4 (Fig 10e)mdashBiramous 7-seg-mented coxa with 2ndash7 setae basis with 1ndash4setae ischium with 3ndash8 setae merus with5ndash11 setae carpus with 4ndash9 setae propoduswith 27ndash38 setae 1 spine on inner distal mar-gin dactylus with 15ndash19 setae exopod usu-ally reduced with 0ndash2 setae
Pereiopod 5 (Fig 10f )mdashCheliform unira-mous 7-segmented coxa with 2ndash6 setae ba-sis with l or 2 setae ischium with 2ndash5 setaemerus with 5ndash9 setae carpus with 4ndash7 se-tae propodus with 22ndash30 setae 1 spine onimmovable finger dactylus with 6ndash10 setaeexopod absent
Pleopods (somite 2) (Fig 10g)mdashBiramouscoxa partially fused to basis without setaebasis with 0ndash3 setae endopod with 3ndash5hooked setae on appendix interna 10ndash17 mar-ginal plumose setae 0ndash2 submarginal nakedsetae exopod with 21ndash27 marginal plumosesetae and 0ndash1 submarginal naked seta
Pleopods (somites 3ndash5) (Fig 10h)mdashBiramouscoxa without setae basis with 3ndash6 marginalplumose setae endopod with 3ndash6 hooked se-tae on appendix interna 10ndash18 marginalplumose setae and 0ndash3 naked submarginal se-tae exopod with 20ndash29 marginal plumose se-tae and 0 or 1 submarginal naked seta
Telson and Uropods (Fig 10i)mdashWith 24ndash36processes on posterior margin median pro-cess usually absent or reduced uropods bi-ramous endopod with 19ndash25 plumose setaeexopod with 35ndash58 plumose setae protopodwith 3ndash5 setae
DISCUSSION
As in other species of Callichirus (see Ro-drigues 1976 Aste and Retamal 1983Strasser and Felder in press b) larval devel-opment of C islagrande included four or fivezoeal stages before metamorphosis to the de-capodid ZI larvae may be identified by thepresence of an elongate rounded rostrumsessile eyes and a lack of pereiopodspleopods or uropods (Fig 1a) At ZII lar-vae acquired biramous buds of pereiopods 1and 2 eyes were separated from the carapaceand the rostrum became flattened and serratedlaterally (Fig 1b) Buds of pleopods on ab-dominal segments 2ndash5 pereiopods 3ndash5 anduropods were developed at ZIII (Fig 1c) Inaddition to further development of structuresmentioned thus far ZIV larvae could be dis-tinguished by presence of a distal spine on theexopod and setae on the endopod of each uro-pod (Fig 1d) When it occurred ZV typicallyresembled ZIV but varied widely in mor-phology Appendages at ZV resembled thoseof ZIV the decapodid stage or some grada-tion between the two (see Figs 9a b for ex-amples) The decapodid stage was similar toother species of Callichirus in the reductionof larval characters such as the elongate ros-trum dorsal spines on the abdomen and ex-opods of the pereiopods (Fig le) Acquisitionof fan-like pleopods marked the transition toa primarily abdominal mode of swimming atthis stage
In the Florida Atlantic population of C ma-jor both salinity and presence of putative set-tlement cues at ZIV impacted the observednumber of zoeal stages and the duration ofZIV (Strasser and Felder in preparation1999c) In C islagrande the percentage oflarvae that passed through five versus fourzoeal stages varied (9ndash89) between parentalfemales but was not shown to depend onsalinity or presence of various putative set-tlement cues (Strasser and Felder 1998 inpreparation) Larvae that molted from ZIVto ZV were usually normal morphologicallyand had mortality similar to those that molteddirectly from ZIV to D In contrast larvae of
STRASSER AND FELDER LARVAL DEVELOPMENT IN CALLICHIRUS ISLAGRANDE 115
the Florida Atlantic C major that molted toZV had a higher incidence of mortality anddeformity at D than did those that had by-passed ZV (Strasser and Felder 1999a c)
The number of zoeal stages appears to beless variable in Callichirus garthi and theGulf of Mexico population of C major thanin congeneric populations studied to dateCallichirus garthi was reported to have fivezoeal stages and a long larval duration reach-ing the decapodid stage 48 days after hatch-ing (Aste and Retamal 1983) Developmentof the Gulf population of C major wasshorter in duration (decapodid reached about10 days after hatching) and typically includedfour zoeal stages (Strasser and Felder 1999a)animals rarely passed through three or fivezoeal stages before the molt to the decapodidAlthough the number of stages is reducedfrom that observed in C garthi developmentin the Gulf population of C major cannot beclassified as abbreviated There are species inboth the Callianassinae (Callianassa slkraussi Stebbing C sl kewalramanii and Csl tyrrhena (Petagna)) and Callichirinae(Sergio mirim Lepidophthalmus sinuensisand L louisianensis) that have only two zoealstages before the molt to the decapodid(Forbes 1973 Sankolli and Shenoy 1975Rodrigues 1984 Thessalou-Legaki 1990Nates et al 1997)
Compared to other species of the genusCallichirus C islagrande is morphologicallycloser to C garthi (from Chile) than to theGulf of Mexico population of C major whichoverlaps in distribution with C islagrande Inthe zoeal phase of C islagrande and Cgarthi the telson is concave and the outer-most process on the ventral margin is articu-lated In both C major and Callichirus sp theposterior margin of the telson is straight andthe outermost process is unarticulated duringthe zoeal stages (Rodrigues 1976 Strasserand Felder 1999a) There are also differencesin spination of the abdomen In zoeae of Cislagrande and C garthi the dorsal spine ofabdominal segment 3 is not longer than thosefound on segments 4 and 5 (Aste and Reta-mal 1983) In contrast the dorsal spine of ab-dominal segment 3 is approximately twice aslong as the dorsal spines on segments 4 and5 in early zoeal stages of C major and Cal-lichirus sp (Rodrigues 1976 Aste and Re-tamal 1983 Strasser and Felder 1999a) Thegreatest difference between these groups is
the number of pleopod pairs at zoeal and de-capodid stages Callichirus islagrande and Cgarthi have pleopods on abdominal segments2ndash5 while pleopods are present only on ab-dominal segments 3ndash5 in C major and Cal-lichirus sp (Rodrigues 1976 Aste and Re-tamal 1983 Strasser and Felder 1999a) Al-though Callichirus islagrande and C garthiare similar morphologically there are differ-ences between these two species Callichirusislagrande as well as C major and Cal-lichirus sp develops pleopod buds andchelate pereiopods at ZIII whereas C garthidoes not acquire these characters until ZIV
There are few callianassid species forwhich the larval development has been de-scribed (Sankolli and Shenoy 1975 Ro-drigues 1976 1984 Aste and Retamal 19831984 Konishi et al 1990 Nates et al 1997Strasser and Felder 1999a) all of which aremembers of the Callichirinae and Cal-lianassinae Among species there is a greatdeal of overlap in larval characters makingit difficult to draw distinctions between thesesubfamilies At present information on lar-val development of species in the Callichiri-nae is limited to larval descriptions of Cal-lichirus major C garthi Callichirus sp Cislagrande Sergio mirim Lepidophthalmuslouisianensis and L sinuensis (see Ro-drigues 1976 1984 Aste and Retamal 1983Nates et al 1997 Strasser and Felder1999a) As mentioned above species of Cal-lichirus have been shown to have four to fivezoeal stages while the other members exhibitabbreviated development that consists of onlytwo zoeal stages The most obvious differencebetween larvae of the species of Callichirinaewith abbreviated development is the numberof pleopod pairs Larvae of S mirim havepleopods on abdominal segments 3ndash5whereas L louisianensis and L sinuensishave pleopods on segments 2ndash5 (Rodrigues1984 Nates et al 1997)
Ghost shrimps of the genus Callichirus maybe distinguished from other genera of the Cal-lianassidae by the cuticular pattern on the dor-sal surface of the abdomen and by the strap-like shape of the uropodal endopods (Manningand Felder 1991) Whereas these charactersmay be used to distinguish adults of the genusCallichirus they are not present at zoeal anddecapodid stages Larvae may be distin-guished from all other callianassid genera byhaving four to five zoeal stages and numer-
116 JOURNAL OF CRUSTACEAN BIOLOGY VOL 20 NO 1 2000
ous processes (12ndash25+1+12ndash25) on the pos-terior margin of the telson during the earlyzoeal stages Larvae of Callichirus may alsodiffer from other callianassids with extendeddevelopment by the retention of the anomu-ran hair as the second process on the poste-rior margin of the telson in the late zoealstages Whereas this process is converted toa spine in Neotrypaea uncinata at ZV and Cal-lianassa sl petalura at ZIV the anomuranhair is retained at ZIV in Callichirus majorand C islagrande (see Rodrigues 1976Strasser and Felder 1999a) Although notmentioned specifically in the descriptions theanomuran hair appears to be present in illus-trations of Callichirus sp at ZIII and Cgarthii at ZV (Rodrigues 1976 fig 35 Asteand Retamal 1983 fig 8Q) A bilobed epi-pod on the first maxilliped of ZI is consistentlyfound in Callichirus but is also present at ZIin Lepidophthalmus sinuensis (see Nates etal 1997) Other callianassids acquire thischaracter at ZII (L louisianensis Nates et al1997 Callianassa sl kewlramanii Sankolliand Shenoy 1975) at ZV (Neotrypaea unci-nata Aste and Retamal 1984) or at the de-capodid stage (Sergio mirim Rodrigues 1984C sl petalura Konishi et al 1990) Thusthe number of zoeal stages and setation of thetelson may serve as the best characters to dis-tinguish larvae of Callichirus from other gen-era However since larval descriptions arelacking for many callianassid species it is un-known if these characters (in combination)will prove to be diagnostic until descriptionsfor other species are published
ACKNOWLEDGEMENTS
We thank Robert Bourgeois Barbara Hasek JimMaKinster Sergio Nates and Kurt Strasser who pro-vided assistance in the field and laboratory and MaryE Rice Director of the Smithsonian Marine StationLink Port Florida who facilitated access to station fa-cilities We also thank R Bauer R Jaeger R Lemaitreand G Watson for their early comments on this manu-script and Raymond B Manning who facilitated fund-ing and gave advice on collecting sites Direct supportfor this study was provided to Karen M Strasser by adoctoral fellowship from the University of Louisianaa graduate student research grant from the LUMCON(Louisiana Universities Marine Consortium) Founda-tion and a summer research award from the Smith-sonian Marine Station Additional support for field andlab activities was provided to D L Felder under Loui-siana Sea Grant College Program Grant RCFB-21 USFish and Wildlife Service Cooperative Agreement 14-16-0009-89-96 (Task Order No 6) US Department of En-ergy grant DE-FG02-97ER12220 and several small proj-ect grants from the Smithsonian Marine Station Views
expressed herein are those of the authors and do not nec-essarily reflect views of supporting government agenciesor subagencies mentioned above This is contributionNumber 466 of the Smithsonian Marine Station and con-tribution Number 67 of the ULL Laboratory for Crus-tacean Research All experiments conducted in this studycomply with current applicable state and federal laws
LITERATURE CITED
Aste A and M Retamal 1983 Desarrollo larval deCallianassa garthi Retamal 1975 bajo condiciones delaboratoriomdashCiencia y Tecnologiacutea del Mar CONA 75ndash26
mdashmdashmdash and mdashmdashmdash 1984 Desarrollo larval de Cal-lianassa uncinata H Milne Edwards1837 (DecapodaCallianassidae) bajo condiciones de laboratoriomdashGayana Zooloacutegica 48 41ndash56
Felder D L 1978 Osmotic and ionic regulation in sev-eral western Atlantic Callianassidae (Crustacea De-capoda Thalassinidea)mdashBiological Bulletin 154409ndash429
mdashmdashmdash and R B Griffis 1994 Dominant infaunalcommunities at risk in shoreline habitats burrowingthalassinid CrustaceamdashOCS Study number MMS94ndash0007 United States Department of the InteriorMinerals Management Service Gulf of Mexico OCSRegional Office New Orleans Louisiana Pp 1ndash87
Forbes A T 1973 An unusual abbreviated larval lifehistory in the estuarine burrowing prawn Callianassakraussi (Crustacea Decapoda Thalassinidea)mdashMa-rine Biology 22 361ndash365
Konishi K 1989 Larval development of the mudshrimp Upogebia (Upogebia) major (de Haan) (Crus-tacea Thalassinidea Upogebiidae) under laboratoryconditions with comments on larval characters of tha-lassinid familiesmdashBulletin of the National ResearchInstitute of Aquaculture 15 1ndash17
mdashmdashmdash R Quintana and Y Fukuda 1990 A completedescription of larval stages of the ghost shrimp Cal-lianassa petalura Stimpson (Crustacea ThalassinideaCallianassidae) under laboratory conditionsmdashBulletinof the National Research Institute of Aquaculture 1727ndash49
Manning R B and D L Felder 1986 The status ofthe callianassid genus Callichirus Stimpson 1866(Crustacea Decapoda Thalassinidea)mdashProceedingsof the Biological Society of Washington 99 437ndash443
mdashmdashmdash and mdashmdashmdash 1991 Revision of the AmericanCallianassidae (Crustacea Decapoda Thalassinidea)mdashProceedings of the Biological Society of Washington104 764ndash794
Nates S F D L Felder and R Lemaitre 1997 Com-parative larval development in two species of the bur-rowing ghost shrimp genus Lepidophthalmus (Decap-oda Callianassidae)mdashJournal of Crustacean Biology17 497ndash519
Rodrigues S de A 1976 Sobre a reproduccedilatildeo embri-ologia e desenvolvimento larval de Callichirus majorSay 1818 (Crustacea Decapoda Thalassinidea)mdashBo-letim de Zoologia Universidade de Satildeo Paulo 185ndash104
mdashmdashmdash 1984 Desenvolvimento poacutes-embrionaacuterio deCallichirus mirim (Rodrigues 1971) obtido emcondiccedilotildees artificiais (Crustacea Decapoda Tha-lassinidea)mdashBoletim de Zoologia Universidade deSatildeo Paulo 8 239ndash256
STRASSER AND FELDER LARVAL DEVELOPMENT IN CALLICHIRUS ISLAGRANDE 117
Sankolli K N and S Shenoy 1975 Larval develop-ment of mud shrimp Callianassa (Callichirus) kewal-ramanii Sankolli in the laboratory (Crustacea Decap-oda)mdashBulletin of the Department of Marine ScienceUniversity of Cochin 4 704ndash720
Staton J L and D L Felder 1995 Genetic variationin populations of the ghost shrimp genus Callichirus(Crustacea Decapoda Thalassinoidea) in the westernAtlantic and Gulf of MexicomdashBulletin of Marine Sci-ence 56 523ndash536
Strasser K M and D L Felder 1998 Settlement cuesin successive developmental stages of the ghost shrimpCallichirus major and C islagrande (Crustacea De-capoda Thalassinidea)mdashMarine Biology
mdashmdashmdash and mdashmdashmdash 1999a Larval development of twopopulations of the ghost shrimp Callichirus major (De-capoda Thalassinidea)mdashJournal of Crustacean Biology
mdashmdashmdash and mdashmdashmdash 1999b Sand as a stimulus for set-tlement in the ghost shrimp Callichirus major (Say)and C islagrande (Schmitt) (Crustacea DecapodaThalassinidea)mdashJournal of Experimental Marine Bi-ology and Ecology 239 211ndash222
mdashmdashmdash and mdashmdashmdash 1999c Settlement cues in theFlorida Atlantic population of the ghost shrimp Cal-lichirus major (Crustacea Decapoda Thalassinidea)mdashMarine Ecology Progress Series
Thessalou-Legaki M 1990 Advanced larval develop-ment of Callianassa tyrrhena (Decapoda Tha-lassinidea) and the effect of environmental factorsmdashJournal of Crustacean Biology 10 659ndash666
RECEIVED 19 September 1998ACCEPTED 3 December 1998

114 JOURNAL OF CRUSTACEAN BIOLOGY VOL 20 NO 1 2000
Pereiopod 1 (Fig 10b)mdashChelate biramous7-segmented coxa with 0ndash6 setae basis with2ndash5 setae ischium with 4ndash7 setae and 0ndash3spines merus with 5ndash8 setae carpus with10ndash14 setae propodus with 27ndash37 setae and3ndash4 spines on inner margin of immovable fin-ger dactylus with 21ndash33 setae exopod usu-ally reduced and without setae occasionallywith 4ndash7 setae
Pereiopod 2 (Fig 10c)mdashChelate biramous7-segmented coxa with 2ndash5 setae basis with1ndash5 setae ischium with 4ndash10 setae meruswith 15ndash20 setae carpus with 8ndash12 setaepropodus with 30ndash40 setae and 1ndash4 spineson inner margin of immovable finger dacty-lus with 21ndash30 setae exopod usually reduced0ndash7 setae
Pereiopod 3 (Fig 10d)mdashBiramous 7-seg-mented coxa with 2ndash5 setae basis with 1ndash3setae ischium with 5ndash8 setae merus with6ndash9 setae carpus with 7ndash11 setae propodusbladelike with 25ndash34 setae 1 spine on innerdistal margin dactylus with 15ndash21 setae exo-pod usually reduced 0ndash5 setae
Pereiopod 4 (Fig 10e)mdashBiramous 7-seg-mented coxa with 2ndash7 setae basis with 1ndash4setae ischium with 3ndash8 setae merus with5ndash11 setae carpus with 4ndash9 setae propoduswith 27ndash38 setae 1 spine on inner distal mar-gin dactylus with 15ndash19 setae exopod usu-ally reduced with 0ndash2 setae
Pereiopod 5 (Fig 10f )mdashCheliform unira-mous 7-segmented coxa with 2ndash6 setae ba-sis with l or 2 setae ischium with 2ndash5 setaemerus with 5ndash9 setae carpus with 4ndash7 se-tae propodus with 22ndash30 setae 1 spine onimmovable finger dactylus with 6ndash10 setaeexopod absent
Pleopods (somite 2) (Fig 10g)mdashBiramouscoxa partially fused to basis without setaebasis with 0ndash3 setae endopod with 3ndash5hooked setae on appendix interna 10ndash17 mar-ginal plumose setae 0ndash2 submarginal nakedsetae exopod with 21ndash27 marginal plumosesetae and 0ndash1 submarginal naked seta
Pleopods (somites 3ndash5) (Fig 10h)mdashBiramouscoxa without setae basis with 3ndash6 marginalplumose setae endopod with 3ndash6 hooked se-tae on appendix interna 10ndash18 marginalplumose setae and 0ndash3 naked submarginal se-tae exopod with 20ndash29 marginal plumose se-tae and 0 or 1 submarginal naked seta
Telson and Uropods (Fig 10i)mdashWith 24ndash36processes on posterior margin median pro-cess usually absent or reduced uropods bi-ramous endopod with 19ndash25 plumose setaeexopod with 35ndash58 plumose setae protopodwith 3ndash5 setae
DISCUSSION
As in other species of Callichirus (see Ro-drigues 1976 Aste and Retamal 1983Strasser and Felder in press b) larval devel-opment of C islagrande included four or fivezoeal stages before metamorphosis to the de-capodid ZI larvae may be identified by thepresence of an elongate rounded rostrumsessile eyes and a lack of pereiopodspleopods or uropods (Fig 1a) At ZII lar-vae acquired biramous buds of pereiopods 1and 2 eyes were separated from the carapaceand the rostrum became flattened and serratedlaterally (Fig 1b) Buds of pleopods on ab-dominal segments 2ndash5 pereiopods 3ndash5 anduropods were developed at ZIII (Fig 1c) Inaddition to further development of structuresmentioned thus far ZIV larvae could be dis-tinguished by presence of a distal spine on theexopod and setae on the endopod of each uro-pod (Fig 1d) When it occurred ZV typicallyresembled ZIV but varied widely in mor-phology Appendages at ZV resembled thoseof ZIV the decapodid stage or some grada-tion between the two (see Figs 9a b for ex-amples) The decapodid stage was similar toother species of Callichirus in the reductionof larval characters such as the elongate ros-trum dorsal spines on the abdomen and ex-opods of the pereiopods (Fig le) Acquisitionof fan-like pleopods marked the transition toa primarily abdominal mode of swimming atthis stage
In the Florida Atlantic population of C ma-jor both salinity and presence of putative set-tlement cues at ZIV impacted the observednumber of zoeal stages and the duration ofZIV (Strasser and Felder in preparation1999c) In C islagrande the percentage oflarvae that passed through five versus fourzoeal stages varied (9ndash89) between parentalfemales but was not shown to depend onsalinity or presence of various putative set-tlement cues (Strasser and Felder 1998 inpreparation) Larvae that molted from ZIVto ZV were usually normal morphologicallyand had mortality similar to those that molteddirectly from ZIV to D In contrast larvae of
STRASSER AND FELDER LARVAL DEVELOPMENT IN CALLICHIRUS ISLAGRANDE 115
the Florida Atlantic C major that molted toZV had a higher incidence of mortality anddeformity at D than did those that had by-passed ZV (Strasser and Felder 1999a c)
The number of zoeal stages appears to beless variable in Callichirus garthi and theGulf of Mexico population of C major thanin congeneric populations studied to dateCallichirus garthi was reported to have fivezoeal stages and a long larval duration reach-ing the decapodid stage 48 days after hatch-ing (Aste and Retamal 1983) Developmentof the Gulf population of C major wasshorter in duration (decapodid reached about10 days after hatching) and typically includedfour zoeal stages (Strasser and Felder 1999a)animals rarely passed through three or fivezoeal stages before the molt to the decapodidAlthough the number of stages is reducedfrom that observed in C garthi developmentin the Gulf population of C major cannot beclassified as abbreviated There are species inboth the Callianassinae (Callianassa slkraussi Stebbing C sl kewalramanii and Csl tyrrhena (Petagna)) and Callichirinae(Sergio mirim Lepidophthalmus sinuensisand L louisianensis) that have only two zoealstages before the molt to the decapodid(Forbes 1973 Sankolli and Shenoy 1975Rodrigues 1984 Thessalou-Legaki 1990Nates et al 1997)
Compared to other species of the genusCallichirus C islagrande is morphologicallycloser to C garthi (from Chile) than to theGulf of Mexico population of C major whichoverlaps in distribution with C islagrande Inthe zoeal phase of C islagrande and Cgarthi the telson is concave and the outer-most process on the ventral margin is articu-lated In both C major and Callichirus sp theposterior margin of the telson is straight andthe outermost process is unarticulated duringthe zoeal stages (Rodrigues 1976 Strasserand Felder 1999a) There are also differencesin spination of the abdomen In zoeae of Cislagrande and C garthi the dorsal spine ofabdominal segment 3 is not longer than thosefound on segments 4 and 5 (Aste and Reta-mal 1983) In contrast the dorsal spine of ab-dominal segment 3 is approximately twice aslong as the dorsal spines on segments 4 and5 in early zoeal stages of C major and Cal-lichirus sp (Rodrigues 1976 Aste and Re-tamal 1983 Strasser and Felder 1999a) Thegreatest difference between these groups is
the number of pleopod pairs at zoeal and de-capodid stages Callichirus islagrande and Cgarthi have pleopods on abdominal segments2ndash5 while pleopods are present only on ab-dominal segments 3ndash5 in C major and Cal-lichirus sp (Rodrigues 1976 Aste and Re-tamal 1983 Strasser and Felder 1999a) Al-though Callichirus islagrande and C garthiare similar morphologically there are differ-ences between these two species Callichirusislagrande as well as C major and Cal-lichirus sp develops pleopod buds andchelate pereiopods at ZIII whereas C garthidoes not acquire these characters until ZIV
There are few callianassid species forwhich the larval development has been de-scribed (Sankolli and Shenoy 1975 Ro-drigues 1976 1984 Aste and Retamal 19831984 Konishi et al 1990 Nates et al 1997Strasser and Felder 1999a) all of which aremembers of the Callichirinae and Cal-lianassinae Among species there is a greatdeal of overlap in larval characters makingit difficult to draw distinctions between thesesubfamilies At present information on lar-val development of species in the Callichiri-nae is limited to larval descriptions of Cal-lichirus major C garthi Callichirus sp Cislagrande Sergio mirim Lepidophthalmuslouisianensis and L sinuensis (see Ro-drigues 1976 1984 Aste and Retamal 1983Nates et al 1997 Strasser and Felder1999a) As mentioned above species of Cal-lichirus have been shown to have four to fivezoeal stages while the other members exhibitabbreviated development that consists of onlytwo zoeal stages The most obvious differencebetween larvae of the species of Callichirinaewith abbreviated development is the numberof pleopod pairs Larvae of S mirim havepleopods on abdominal segments 3ndash5whereas L louisianensis and L sinuensishave pleopods on segments 2ndash5 (Rodrigues1984 Nates et al 1997)
Ghost shrimps of the genus Callichirus maybe distinguished from other genera of the Cal-lianassidae by the cuticular pattern on the dor-sal surface of the abdomen and by the strap-like shape of the uropodal endopods (Manningand Felder 1991) Whereas these charactersmay be used to distinguish adults of the genusCallichirus they are not present at zoeal anddecapodid stages Larvae may be distin-guished from all other callianassid genera byhaving four to five zoeal stages and numer-
116 JOURNAL OF CRUSTACEAN BIOLOGY VOL 20 NO 1 2000
ous processes (12ndash25+1+12ndash25) on the pos-terior margin of the telson during the earlyzoeal stages Larvae of Callichirus may alsodiffer from other callianassids with extendeddevelopment by the retention of the anomu-ran hair as the second process on the poste-rior margin of the telson in the late zoealstages Whereas this process is converted toa spine in Neotrypaea uncinata at ZV and Cal-lianassa sl petalura at ZIV the anomuranhair is retained at ZIV in Callichirus majorand C islagrande (see Rodrigues 1976Strasser and Felder 1999a) Although notmentioned specifically in the descriptions theanomuran hair appears to be present in illus-trations of Callichirus sp at ZIII and Cgarthii at ZV (Rodrigues 1976 fig 35 Asteand Retamal 1983 fig 8Q) A bilobed epi-pod on the first maxilliped of ZI is consistentlyfound in Callichirus but is also present at ZIin Lepidophthalmus sinuensis (see Nates etal 1997) Other callianassids acquire thischaracter at ZII (L louisianensis Nates et al1997 Callianassa sl kewlramanii Sankolliand Shenoy 1975) at ZV (Neotrypaea unci-nata Aste and Retamal 1984) or at the de-capodid stage (Sergio mirim Rodrigues 1984C sl petalura Konishi et al 1990) Thusthe number of zoeal stages and setation of thetelson may serve as the best characters to dis-tinguish larvae of Callichirus from other gen-era However since larval descriptions arelacking for many callianassid species it is un-known if these characters (in combination)will prove to be diagnostic until descriptionsfor other species are published
ACKNOWLEDGEMENTS
We thank Robert Bourgeois Barbara Hasek JimMaKinster Sergio Nates and Kurt Strasser who pro-vided assistance in the field and laboratory and MaryE Rice Director of the Smithsonian Marine StationLink Port Florida who facilitated access to station fa-cilities We also thank R Bauer R Jaeger R Lemaitreand G Watson for their early comments on this manu-script and Raymond B Manning who facilitated fund-ing and gave advice on collecting sites Direct supportfor this study was provided to Karen M Strasser by adoctoral fellowship from the University of Louisianaa graduate student research grant from the LUMCON(Louisiana Universities Marine Consortium) Founda-tion and a summer research award from the Smith-sonian Marine Station Additional support for field andlab activities was provided to D L Felder under Loui-siana Sea Grant College Program Grant RCFB-21 USFish and Wildlife Service Cooperative Agreement 14-16-0009-89-96 (Task Order No 6) US Department of En-ergy grant DE-FG02-97ER12220 and several small proj-ect grants from the Smithsonian Marine Station Views
expressed herein are those of the authors and do not nec-essarily reflect views of supporting government agenciesor subagencies mentioned above This is contributionNumber 466 of the Smithsonian Marine Station and con-tribution Number 67 of the ULL Laboratory for Crus-tacean Research All experiments conducted in this studycomply with current applicable state and federal laws
LITERATURE CITED
Aste A and M Retamal 1983 Desarrollo larval deCallianassa garthi Retamal 1975 bajo condiciones delaboratoriomdashCiencia y Tecnologiacutea del Mar CONA 75ndash26
mdashmdashmdash and mdashmdashmdash 1984 Desarrollo larval de Cal-lianassa uncinata H Milne Edwards1837 (DecapodaCallianassidae) bajo condiciones de laboratoriomdashGayana Zooloacutegica 48 41ndash56
Felder D L 1978 Osmotic and ionic regulation in sev-eral western Atlantic Callianassidae (Crustacea De-capoda Thalassinidea)mdashBiological Bulletin 154409ndash429
mdashmdashmdash and R B Griffis 1994 Dominant infaunalcommunities at risk in shoreline habitats burrowingthalassinid CrustaceamdashOCS Study number MMS94ndash0007 United States Department of the InteriorMinerals Management Service Gulf of Mexico OCSRegional Office New Orleans Louisiana Pp 1ndash87
Forbes A T 1973 An unusual abbreviated larval lifehistory in the estuarine burrowing prawn Callianassakraussi (Crustacea Decapoda Thalassinidea)mdashMa-rine Biology 22 361ndash365
Konishi K 1989 Larval development of the mudshrimp Upogebia (Upogebia) major (de Haan) (Crus-tacea Thalassinidea Upogebiidae) under laboratoryconditions with comments on larval characters of tha-lassinid familiesmdashBulletin of the National ResearchInstitute of Aquaculture 15 1ndash17
mdashmdashmdash R Quintana and Y Fukuda 1990 A completedescription of larval stages of the ghost shrimp Cal-lianassa petalura Stimpson (Crustacea ThalassinideaCallianassidae) under laboratory conditionsmdashBulletinof the National Research Institute of Aquaculture 1727ndash49
Manning R B and D L Felder 1986 The status ofthe callianassid genus Callichirus Stimpson 1866(Crustacea Decapoda Thalassinidea)mdashProceedingsof the Biological Society of Washington 99 437ndash443
mdashmdashmdash and mdashmdashmdash 1991 Revision of the AmericanCallianassidae (Crustacea Decapoda Thalassinidea)mdashProceedings of the Biological Society of Washington104 764ndash794
Nates S F D L Felder and R Lemaitre 1997 Com-parative larval development in two species of the bur-rowing ghost shrimp genus Lepidophthalmus (Decap-oda Callianassidae)mdashJournal of Crustacean Biology17 497ndash519
Rodrigues S de A 1976 Sobre a reproduccedilatildeo embri-ologia e desenvolvimento larval de Callichirus majorSay 1818 (Crustacea Decapoda Thalassinidea)mdashBo-letim de Zoologia Universidade de Satildeo Paulo 185ndash104
mdashmdashmdash 1984 Desenvolvimento poacutes-embrionaacuterio deCallichirus mirim (Rodrigues 1971) obtido emcondiccedilotildees artificiais (Crustacea Decapoda Tha-lassinidea)mdashBoletim de Zoologia Universidade deSatildeo Paulo 8 239ndash256
STRASSER AND FELDER LARVAL DEVELOPMENT IN CALLICHIRUS ISLAGRANDE 117
Sankolli K N and S Shenoy 1975 Larval develop-ment of mud shrimp Callianassa (Callichirus) kewal-ramanii Sankolli in the laboratory (Crustacea Decap-oda)mdashBulletin of the Department of Marine ScienceUniversity of Cochin 4 704ndash720
Staton J L and D L Felder 1995 Genetic variationin populations of the ghost shrimp genus Callichirus(Crustacea Decapoda Thalassinoidea) in the westernAtlantic and Gulf of MexicomdashBulletin of Marine Sci-ence 56 523ndash536
Strasser K M and D L Felder 1998 Settlement cuesin successive developmental stages of the ghost shrimpCallichirus major and C islagrande (Crustacea De-capoda Thalassinidea)mdashMarine Biology
mdashmdashmdash and mdashmdashmdash 1999a Larval development of twopopulations of the ghost shrimp Callichirus major (De-capoda Thalassinidea)mdashJournal of Crustacean Biology
mdashmdashmdash and mdashmdashmdash 1999b Sand as a stimulus for set-tlement in the ghost shrimp Callichirus major (Say)and C islagrande (Schmitt) (Crustacea DecapodaThalassinidea)mdashJournal of Experimental Marine Bi-ology and Ecology 239 211ndash222
mdashmdashmdash and mdashmdashmdash 1999c Settlement cues in theFlorida Atlantic population of the ghost shrimp Cal-lichirus major (Crustacea Decapoda Thalassinidea)mdashMarine Ecology Progress Series
Thessalou-Legaki M 1990 Advanced larval develop-ment of Callianassa tyrrhena (Decapoda Tha-lassinidea) and the effect of environmental factorsmdashJournal of Crustacean Biology 10 659ndash666
RECEIVED 19 September 1998ACCEPTED 3 December 1998

STRASSER AND FELDER LARVAL DEVELOPMENT IN CALLICHIRUS ISLAGRANDE 115
the Florida Atlantic C major that molted toZV had a higher incidence of mortality anddeformity at D than did those that had by-passed ZV (Strasser and Felder 1999a c)
The number of zoeal stages appears to beless variable in Callichirus garthi and theGulf of Mexico population of C major thanin congeneric populations studied to dateCallichirus garthi was reported to have fivezoeal stages and a long larval duration reach-ing the decapodid stage 48 days after hatch-ing (Aste and Retamal 1983) Developmentof the Gulf population of C major wasshorter in duration (decapodid reached about10 days after hatching) and typically includedfour zoeal stages (Strasser and Felder 1999a)animals rarely passed through three or fivezoeal stages before the molt to the decapodidAlthough the number of stages is reducedfrom that observed in C garthi developmentin the Gulf population of C major cannot beclassified as abbreviated There are species inboth the Callianassinae (Callianassa slkraussi Stebbing C sl kewalramanii and Csl tyrrhena (Petagna)) and Callichirinae(Sergio mirim Lepidophthalmus sinuensisand L louisianensis) that have only two zoealstages before the molt to the decapodid(Forbes 1973 Sankolli and Shenoy 1975Rodrigues 1984 Thessalou-Legaki 1990Nates et al 1997)
Compared to other species of the genusCallichirus C islagrande is morphologicallycloser to C garthi (from Chile) than to theGulf of Mexico population of C major whichoverlaps in distribution with C islagrande Inthe zoeal phase of C islagrande and Cgarthi the telson is concave and the outer-most process on the ventral margin is articu-lated In both C major and Callichirus sp theposterior margin of the telson is straight andthe outermost process is unarticulated duringthe zoeal stages (Rodrigues 1976 Strasserand Felder 1999a) There are also differencesin spination of the abdomen In zoeae of Cislagrande and C garthi the dorsal spine ofabdominal segment 3 is not longer than thosefound on segments 4 and 5 (Aste and Reta-mal 1983) In contrast the dorsal spine of ab-dominal segment 3 is approximately twice aslong as the dorsal spines on segments 4 and5 in early zoeal stages of C major and Cal-lichirus sp (Rodrigues 1976 Aste and Re-tamal 1983 Strasser and Felder 1999a) Thegreatest difference between these groups is
the number of pleopod pairs at zoeal and de-capodid stages Callichirus islagrande and Cgarthi have pleopods on abdominal segments2ndash5 while pleopods are present only on ab-dominal segments 3ndash5 in C major and Cal-lichirus sp (Rodrigues 1976 Aste and Re-tamal 1983 Strasser and Felder 1999a) Al-though Callichirus islagrande and C garthiare similar morphologically there are differ-ences between these two species Callichirusislagrande as well as C major and Cal-lichirus sp develops pleopod buds andchelate pereiopods at ZIII whereas C garthidoes not acquire these characters until ZIV
There are few callianassid species forwhich the larval development has been de-scribed (Sankolli and Shenoy 1975 Ro-drigues 1976 1984 Aste and Retamal 19831984 Konishi et al 1990 Nates et al 1997Strasser and Felder 1999a) all of which aremembers of the Callichirinae and Cal-lianassinae Among species there is a greatdeal of overlap in larval characters makingit difficult to draw distinctions between thesesubfamilies At present information on lar-val development of species in the Callichiri-nae is limited to larval descriptions of Cal-lichirus major C garthi Callichirus sp Cislagrande Sergio mirim Lepidophthalmuslouisianensis and L sinuensis (see Ro-drigues 1976 1984 Aste and Retamal 1983Nates et al 1997 Strasser and Felder1999a) As mentioned above species of Cal-lichirus have been shown to have four to fivezoeal stages while the other members exhibitabbreviated development that consists of onlytwo zoeal stages The most obvious differencebetween larvae of the species of Callichirinaewith abbreviated development is the numberof pleopod pairs Larvae of S mirim havepleopods on abdominal segments 3ndash5whereas L louisianensis and L sinuensishave pleopods on segments 2ndash5 (Rodrigues1984 Nates et al 1997)
Ghost shrimps of the genus Callichirus maybe distinguished from other genera of the Cal-lianassidae by the cuticular pattern on the dor-sal surface of the abdomen and by the strap-like shape of the uropodal endopods (Manningand Felder 1991) Whereas these charactersmay be used to distinguish adults of the genusCallichirus they are not present at zoeal anddecapodid stages Larvae may be distin-guished from all other callianassid genera byhaving four to five zoeal stages and numer-
116 JOURNAL OF CRUSTACEAN BIOLOGY VOL 20 NO 1 2000
ous processes (12ndash25+1+12ndash25) on the pos-terior margin of the telson during the earlyzoeal stages Larvae of Callichirus may alsodiffer from other callianassids with extendeddevelopment by the retention of the anomu-ran hair as the second process on the poste-rior margin of the telson in the late zoealstages Whereas this process is converted toa spine in Neotrypaea uncinata at ZV and Cal-lianassa sl petalura at ZIV the anomuranhair is retained at ZIV in Callichirus majorand C islagrande (see Rodrigues 1976Strasser and Felder 1999a) Although notmentioned specifically in the descriptions theanomuran hair appears to be present in illus-trations of Callichirus sp at ZIII and Cgarthii at ZV (Rodrigues 1976 fig 35 Asteand Retamal 1983 fig 8Q) A bilobed epi-pod on the first maxilliped of ZI is consistentlyfound in Callichirus but is also present at ZIin Lepidophthalmus sinuensis (see Nates etal 1997) Other callianassids acquire thischaracter at ZII (L louisianensis Nates et al1997 Callianassa sl kewlramanii Sankolliand Shenoy 1975) at ZV (Neotrypaea unci-nata Aste and Retamal 1984) or at the de-capodid stage (Sergio mirim Rodrigues 1984C sl petalura Konishi et al 1990) Thusthe number of zoeal stages and setation of thetelson may serve as the best characters to dis-tinguish larvae of Callichirus from other gen-era However since larval descriptions arelacking for many callianassid species it is un-known if these characters (in combination)will prove to be diagnostic until descriptionsfor other species are published
ACKNOWLEDGEMENTS
We thank Robert Bourgeois Barbara Hasek JimMaKinster Sergio Nates and Kurt Strasser who pro-vided assistance in the field and laboratory and MaryE Rice Director of the Smithsonian Marine StationLink Port Florida who facilitated access to station fa-cilities We also thank R Bauer R Jaeger R Lemaitreand G Watson for their early comments on this manu-script and Raymond B Manning who facilitated fund-ing and gave advice on collecting sites Direct supportfor this study was provided to Karen M Strasser by adoctoral fellowship from the University of Louisianaa graduate student research grant from the LUMCON(Louisiana Universities Marine Consortium) Founda-tion and a summer research award from the Smith-sonian Marine Station Additional support for field andlab activities was provided to D L Felder under Loui-siana Sea Grant College Program Grant RCFB-21 USFish and Wildlife Service Cooperative Agreement 14-16-0009-89-96 (Task Order No 6) US Department of En-ergy grant DE-FG02-97ER12220 and several small proj-ect grants from the Smithsonian Marine Station Views
expressed herein are those of the authors and do not nec-essarily reflect views of supporting government agenciesor subagencies mentioned above This is contributionNumber 466 of the Smithsonian Marine Station and con-tribution Number 67 of the ULL Laboratory for Crus-tacean Research All experiments conducted in this studycomply with current applicable state and federal laws
LITERATURE CITED
Aste A and M Retamal 1983 Desarrollo larval deCallianassa garthi Retamal 1975 bajo condiciones delaboratoriomdashCiencia y Tecnologiacutea del Mar CONA 75ndash26
mdashmdashmdash and mdashmdashmdash 1984 Desarrollo larval de Cal-lianassa uncinata H Milne Edwards1837 (DecapodaCallianassidae) bajo condiciones de laboratoriomdashGayana Zooloacutegica 48 41ndash56
Felder D L 1978 Osmotic and ionic regulation in sev-eral western Atlantic Callianassidae (Crustacea De-capoda Thalassinidea)mdashBiological Bulletin 154409ndash429
mdashmdashmdash and R B Griffis 1994 Dominant infaunalcommunities at risk in shoreline habitats burrowingthalassinid CrustaceamdashOCS Study number MMS94ndash0007 United States Department of the InteriorMinerals Management Service Gulf of Mexico OCSRegional Office New Orleans Louisiana Pp 1ndash87
Forbes A T 1973 An unusual abbreviated larval lifehistory in the estuarine burrowing prawn Callianassakraussi (Crustacea Decapoda Thalassinidea)mdashMa-rine Biology 22 361ndash365
Konishi K 1989 Larval development of the mudshrimp Upogebia (Upogebia) major (de Haan) (Crus-tacea Thalassinidea Upogebiidae) under laboratoryconditions with comments on larval characters of tha-lassinid familiesmdashBulletin of the National ResearchInstitute of Aquaculture 15 1ndash17
mdashmdashmdash R Quintana and Y Fukuda 1990 A completedescription of larval stages of the ghost shrimp Cal-lianassa petalura Stimpson (Crustacea ThalassinideaCallianassidae) under laboratory conditionsmdashBulletinof the National Research Institute of Aquaculture 1727ndash49
Manning R B and D L Felder 1986 The status ofthe callianassid genus Callichirus Stimpson 1866(Crustacea Decapoda Thalassinidea)mdashProceedingsof the Biological Society of Washington 99 437ndash443
mdashmdashmdash and mdashmdashmdash 1991 Revision of the AmericanCallianassidae (Crustacea Decapoda Thalassinidea)mdashProceedings of the Biological Society of Washington104 764ndash794
Nates S F D L Felder and R Lemaitre 1997 Com-parative larval development in two species of the bur-rowing ghost shrimp genus Lepidophthalmus (Decap-oda Callianassidae)mdashJournal of Crustacean Biology17 497ndash519
Rodrigues S de A 1976 Sobre a reproduccedilatildeo embri-ologia e desenvolvimento larval de Callichirus majorSay 1818 (Crustacea Decapoda Thalassinidea)mdashBo-letim de Zoologia Universidade de Satildeo Paulo 185ndash104
mdashmdashmdash 1984 Desenvolvimento poacutes-embrionaacuterio deCallichirus mirim (Rodrigues 1971) obtido emcondiccedilotildees artificiais (Crustacea Decapoda Tha-lassinidea)mdashBoletim de Zoologia Universidade deSatildeo Paulo 8 239ndash256
STRASSER AND FELDER LARVAL DEVELOPMENT IN CALLICHIRUS ISLAGRANDE 117
Sankolli K N and S Shenoy 1975 Larval develop-ment of mud shrimp Callianassa (Callichirus) kewal-ramanii Sankolli in the laboratory (Crustacea Decap-oda)mdashBulletin of the Department of Marine ScienceUniversity of Cochin 4 704ndash720
Staton J L and D L Felder 1995 Genetic variationin populations of the ghost shrimp genus Callichirus(Crustacea Decapoda Thalassinoidea) in the westernAtlantic and Gulf of MexicomdashBulletin of Marine Sci-ence 56 523ndash536
Strasser K M and D L Felder 1998 Settlement cuesin successive developmental stages of the ghost shrimpCallichirus major and C islagrande (Crustacea De-capoda Thalassinidea)mdashMarine Biology
mdashmdashmdash and mdashmdashmdash 1999a Larval development of twopopulations of the ghost shrimp Callichirus major (De-capoda Thalassinidea)mdashJournal of Crustacean Biology
mdashmdashmdash and mdashmdashmdash 1999b Sand as a stimulus for set-tlement in the ghost shrimp Callichirus major (Say)and C islagrande (Schmitt) (Crustacea DecapodaThalassinidea)mdashJournal of Experimental Marine Bi-ology and Ecology 239 211ndash222
mdashmdashmdash and mdashmdashmdash 1999c Settlement cues in theFlorida Atlantic population of the ghost shrimp Cal-lichirus major (Crustacea Decapoda Thalassinidea)mdashMarine Ecology Progress Series
Thessalou-Legaki M 1990 Advanced larval develop-ment of Callianassa tyrrhena (Decapoda Tha-lassinidea) and the effect of environmental factorsmdashJournal of Crustacean Biology 10 659ndash666
RECEIVED 19 September 1998ACCEPTED 3 December 1998

116 JOURNAL OF CRUSTACEAN BIOLOGY VOL 20 NO 1 2000
ous processes (12ndash25+1+12ndash25) on the pos-terior margin of the telson during the earlyzoeal stages Larvae of Callichirus may alsodiffer from other callianassids with extendeddevelopment by the retention of the anomu-ran hair as the second process on the poste-rior margin of the telson in the late zoealstages Whereas this process is converted toa spine in Neotrypaea uncinata at ZV and Cal-lianassa sl petalura at ZIV the anomuranhair is retained at ZIV in Callichirus majorand C islagrande (see Rodrigues 1976Strasser and Felder 1999a) Although notmentioned specifically in the descriptions theanomuran hair appears to be present in illus-trations of Callichirus sp at ZIII and Cgarthii at ZV (Rodrigues 1976 fig 35 Asteand Retamal 1983 fig 8Q) A bilobed epi-pod on the first maxilliped of ZI is consistentlyfound in Callichirus but is also present at ZIin Lepidophthalmus sinuensis (see Nates etal 1997) Other callianassids acquire thischaracter at ZII (L louisianensis Nates et al1997 Callianassa sl kewlramanii Sankolliand Shenoy 1975) at ZV (Neotrypaea unci-nata Aste and Retamal 1984) or at the de-capodid stage (Sergio mirim Rodrigues 1984C sl petalura Konishi et al 1990) Thusthe number of zoeal stages and setation of thetelson may serve as the best characters to dis-tinguish larvae of Callichirus from other gen-era However since larval descriptions arelacking for many callianassid species it is un-known if these characters (in combination)will prove to be diagnostic until descriptionsfor other species are published
ACKNOWLEDGEMENTS
We thank Robert Bourgeois Barbara Hasek JimMaKinster Sergio Nates and Kurt Strasser who pro-vided assistance in the field and laboratory and MaryE Rice Director of the Smithsonian Marine StationLink Port Florida who facilitated access to station fa-cilities We also thank R Bauer R Jaeger R Lemaitreand G Watson for their early comments on this manu-script and Raymond B Manning who facilitated fund-ing and gave advice on collecting sites Direct supportfor this study was provided to Karen M Strasser by adoctoral fellowship from the University of Louisianaa graduate student research grant from the LUMCON(Louisiana Universities Marine Consortium) Founda-tion and a summer research award from the Smith-sonian Marine Station Additional support for field andlab activities was provided to D L Felder under Loui-siana Sea Grant College Program Grant RCFB-21 USFish and Wildlife Service Cooperative Agreement 14-16-0009-89-96 (Task Order No 6) US Department of En-ergy grant DE-FG02-97ER12220 and several small proj-ect grants from the Smithsonian Marine Station Views
expressed herein are those of the authors and do not nec-essarily reflect views of supporting government agenciesor subagencies mentioned above This is contributionNumber 466 of the Smithsonian Marine Station and con-tribution Number 67 of the ULL Laboratory for Crus-tacean Research All experiments conducted in this studycomply with current applicable state and federal laws
LITERATURE CITED
Aste A and M Retamal 1983 Desarrollo larval deCallianassa garthi Retamal 1975 bajo condiciones delaboratoriomdashCiencia y Tecnologiacutea del Mar CONA 75ndash26
mdashmdashmdash and mdashmdashmdash 1984 Desarrollo larval de Cal-lianassa uncinata H Milne Edwards1837 (DecapodaCallianassidae) bajo condiciones de laboratoriomdashGayana Zooloacutegica 48 41ndash56
Felder D L 1978 Osmotic and ionic regulation in sev-eral western Atlantic Callianassidae (Crustacea De-capoda Thalassinidea)mdashBiological Bulletin 154409ndash429
mdashmdashmdash and R B Griffis 1994 Dominant infaunalcommunities at risk in shoreline habitats burrowingthalassinid CrustaceamdashOCS Study number MMS94ndash0007 United States Department of the InteriorMinerals Management Service Gulf of Mexico OCSRegional Office New Orleans Louisiana Pp 1ndash87
Forbes A T 1973 An unusual abbreviated larval lifehistory in the estuarine burrowing prawn Callianassakraussi (Crustacea Decapoda Thalassinidea)mdashMa-rine Biology 22 361ndash365
Konishi K 1989 Larval development of the mudshrimp Upogebia (Upogebia) major (de Haan) (Crus-tacea Thalassinidea Upogebiidae) under laboratoryconditions with comments on larval characters of tha-lassinid familiesmdashBulletin of the National ResearchInstitute of Aquaculture 15 1ndash17
mdashmdashmdash R Quintana and Y Fukuda 1990 A completedescription of larval stages of the ghost shrimp Cal-lianassa petalura Stimpson (Crustacea ThalassinideaCallianassidae) under laboratory conditionsmdashBulletinof the National Research Institute of Aquaculture 1727ndash49
Manning R B and D L Felder 1986 The status ofthe callianassid genus Callichirus Stimpson 1866(Crustacea Decapoda Thalassinidea)mdashProceedingsof the Biological Society of Washington 99 437ndash443
mdashmdashmdash and mdashmdashmdash 1991 Revision of the AmericanCallianassidae (Crustacea Decapoda Thalassinidea)mdashProceedings of the Biological Society of Washington104 764ndash794
Nates S F D L Felder and R Lemaitre 1997 Com-parative larval development in two species of the bur-rowing ghost shrimp genus Lepidophthalmus (Decap-oda Callianassidae)mdashJournal of Crustacean Biology17 497ndash519
Rodrigues S de A 1976 Sobre a reproduccedilatildeo embri-ologia e desenvolvimento larval de Callichirus majorSay 1818 (Crustacea Decapoda Thalassinidea)mdashBo-letim de Zoologia Universidade de Satildeo Paulo 185ndash104
mdashmdashmdash 1984 Desenvolvimento poacutes-embrionaacuterio deCallichirus mirim (Rodrigues 1971) obtido emcondiccedilotildees artificiais (Crustacea Decapoda Tha-lassinidea)mdashBoletim de Zoologia Universidade deSatildeo Paulo 8 239ndash256
STRASSER AND FELDER LARVAL DEVELOPMENT IN CALLICHIRUS ISLAGRANDE 117
Sankolli K N and S Shenoy 1975 Larval develop-ment of mud shrimp Callianassa (Callichirus) kewal-ramanii Sankolli in the laboratory (Crustacea Decap-oda)mdashBulletin of the Department of Marine ScienceUniversity of Cochin 4 704ndash720
Staton J L and D L Felder 1995 Genetic variationin populations of the ghost shrimp genus Callichirus(Crustacea Decapoda Thalassinoidea) in the westernAtlantic and Gulf of MexicomdashBulletin of Marine Sci-ence 56 523ndash536
Strasser K M and D L Felder 1998 Settlement cuesin successive developmental stages of the ghost shrimpCallichirus major and C islagrande (Crustacea De-capoda Thalassinidea)mdashMarine Biology
mdashmdashmdash and mdashmdashmdash 1999a Larval development of twopopulations of the ghost shrimp Callichirus major (De-capoda Thalassinidea)mdashJournal of Crustacean Biology
mdashmdashmdash and mdashmdashmdash 1999b Sand as a stimulus for set-tlement in the ghost shrimp Callichirus major (Say)and C islagrande (Schmitt) (Crustacea DecapodaThalassinidea)mdashJournal of Experimental Marine Bi-ology and Ecology 239 211ndash222
mdashmdashmdash and mdashmdashmdash 1999c Settlement cues in theFlorida Atlantic population of the ghost shrimp Cal-lichirus major (Crustacea Decapoda Thalassinidea)mdashMarine Ecology Progress Series
Thessalou-Legaki M 1990 Advanced larval develop-ment of Callianassa tyrrhena (Decapoda Tha-lassinidea) and the effect of environmental factorsmdashJournal of Crustacean Biology 10 659ndash666
RECEIVED 19 September 1998ACCEPTED 3 December 1998

STRASSER AND FELDER LARVAL DEVELOPMENT IN CALLICHIRUS ISLAGRANDE 117
Sankolli K N and S Shenoy 1975 Larval develop-ment of mud shrimp Callianassa (Callichirus) kewal-ramanii Sankolli in the laboratory (Crustacea Decap-oda)mdashBulletin of the Department of Marine ScienceUniversity of Cochin 4 704ndash720
Staton J L and D L Felder 1995 Genetic variationin populations of the ghost shrimp genus Callichirus(Crustacea Decapoda Thalassinoidea) in the westernAtlantic and Gulf of MexicomdashBulletin of Marine Sci-ence 56 523ndash536
Strasser K M and D L Felder 1998 Settlement cuesin successive developmental stages of the ghost shrimpCallichirus major and C islagrande (Crustacea De-capoda Thalassinidea)mdashMarine Biology
mdashmdashmdash and mdashmdashmdash 1999a Larval development of twopopulations of the ghost shrimp Callichirus major (De-capoda Thalassinidea)mdashJournal of Crustacean Biology
mdashmdashmdash and mdashmdashmdash 1999b Sand as a stimulus for set-tlement in the ghost shrimp Callichirus major (Say)and C islagrande (Schmitt) (Crustacea DecapodaThalassinidea)mdashJournal of Experimental Marine Bi-ology and Ecology 239 211ndash222
mdashmdashmdash and mdashmdashmdash 1999c Settlement cues in theFlorida Atlantic population of the ghost shrimp Cal-lichirus major (Crustacea Decapoda Thalassinidea)mdashMarine Ecology Progress Series
Thessalou-Legaki M 1990 Advanced larval develop-ment of Callianassa tyrrhena (Decapoda Tha-lassinidea) and the effect of environmental factorsmdashJournal of Crustacean Biology 10 659ndash666
RECEIVED 19 September 1998ACCEPTED 3 December 1998


























