Embed Size (px)
Citation preview

of June 12, 2018.This information is current as
Cell Activation , Is Required for TlckSH2 Domain of p56
Lad, an Adapter Protein Interacting with the
Young Bong Choi, Chan Ki Kim and Yungdae Yun
http://www.jimmunol.org/content/163/10/52421999; 163:5242-5249; ;J Immunol
Referenceshttp://www.jimmunol.org/content/163/10/5242.full#ref-list-1
, 31 of which you can access for free at: cites 54 articlesThis article
average*
4 weeks from acceptance to publicationFast Publication! •
Every submission reviewed by practicing scientistsNo Triage! •
from submission to initial decisionRapid Reviews! 30 days* •
Submit online. ?The JIWhy
Subscriptionhttp://jimmunol.org/subscription
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/About/Publications/JI/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/alertsReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists All rights reserved.Copyright © 1999 by The American Association of1451 Rockville Pike, Suite 650, Rockville, MD 20852The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on June 12, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
by guest on June 12, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

Lad, an Adapter Protein Interacting with the SH2 Domain ofp56lck, Is Required for T Cell Activation 1,2
Young Bong Choi,*† Chan Ki Kim,* and Yungdae Yun 3*†
T cell-specific Src family tyrosine kinase, p56lck, plays crucial roles in T cell differentiation, activation, and proliferation. Thesemultiple functions of p56lck are believed to be conducted through the protein-protein interactions with various cellular signalingproteins. To clarify the mechanisms through which p56lck contributes to T cell signaling, we identified the proteins binding to theSrc homology 2 (SH2) domain of p56lck through a tyrosine phosphorylation-dependent yeast two-hybrid screening. Subsequentcharacterization of positive clones revealed the presence of a protein of 366 aa named Lad (Lck-associated adapter protein), whichis a potential murine homologue of previously reported TSAd, a T cell-specific adapter protein. Lad contains several protein-protein interaction domains including a zinc-finger motif, an SH2 domain, a proline-rich SH3 binding motif, and several phos-photyrosine sites. Furthermore, Lad was tyrosine phosphorylated and associated with p56lck in vivo and redistributed fromcytoplasm to the plasma membrane in a T cell activation-dependent manner. Moreover in T cells, IL-2 promoter activity wasenhanced upon coexpression of Lad but was inhibited by the coexpression of antisense Lad RNA. These characteristics of Ladsuggest that Lad play an essential role as an adapter protein in p56lck-mediated T cell signaling. The Journal of Immunology,1999, 163: 5242–5249.
T he best-characterized lymphocyte-specific member of theSrc family tyrosine kinases, p56lck, plays essential roles inT cell signaling that regulates diverse T cell functions
such as development, activation, proliferation, and adhesion.Knock-out mice lacking p56lck show a pronounced thymic at-
rophy owing to blockade of the progression from CD42CD82
double negative to CD41CD81 double positive thymocytes (1).Transgenic mice harboring a dominant negative form of p56lck aredefective in allelic exclusion of the pre TCRb-chain gene whichpermits normal thymic selection (2, 3). Both animal studies indi-cate that p56lck is important for T cell development (4).
During Ag-induced T cell activation, p56lck transmits a positivesignal by interacting with the CD4/CD8 glycoproteins (5–7). Fur-thermore, genetic evidence using JCaM1 cells lacking p56lck
shows that p56lck is involved in TCR-mediated cell activation (8).During T cell proliferation by IL-2, p56lck associates with theIL-2R b-chain (9) and regulates c-fos/c-jungene expression (10,11). In addition, p56lck associates with other costimulatory adhe-sion molecules such as 4–1BB (12), CD2 (13), CD44 (14), andL-selectin (15) to enhance T cell responsiveness. These multiplefunctions of p56lck are believed to be conducted through interac-tion with various cellular signaling proteins.
Like other Src family protein tyrosine kinases (PTK),4 p56lck
consists of five domains: SH1 (Src homology domain 1), SH2,SH3, SH4, and NH2 unique domain. The SH1 is the enzymaticdomain of PTK that phosphorylates tyrosines on cellular proteinswith catalytic specificity (16). The N-terminal unique domain in-fluences substrate preference without the regulation of intrinsickinase activity (17) and regulates interaction with protein tyrosinephosphatases (18). The SH4 domain directs p56lck to the plasmamembrane by denoting sites for lipidation such as palmitoylationor myristoylation (19, 20), which enables p56lck to interact withGPI-anchored proteins such as CD59 (21). The SH3 domain neg-atively regulates the enzymatic activity of p56lck and is dispensablefor cell transformation by activated p56lck (F505) (22, 23). Severalgroups, however, reported that the SH3 domain interacts with sev-eral cellular signaling proteins including phosphatidylinositol 3-ki-nase (PI3K) (24, 25), p120 (26), and LckBP1 (27) through theirproline-rich motifs. The functional significance of the p56lck SH3domain in T cell signaling remains yet to be elucidated. Finally,the SH2 domain negatively or positively regulates the function ofp56lck (22). In the inactive form of p56lck, the SH2 domain inter-acts with its own phosphorylated Y505 (pY505), but in the activeform of p56lck, the SH2 domain interacts with other tyrosine phos-phorylated cellular signaling proteins (28) to transmit a positivesignal for T cell activation.
The importance of p56lck in T cell activation has been describedextensively and both the kinase and the regulatory domains havebeen shown to be required. A model was established in which,upon engagement with CD4/CD8, the kinase domain of p56lck
phosphorylates thez-chain of TCR and provides the binding sitefor another kinase, ZAP-70. These successive events lead to theamplification of TCR-mediated signaling (29). On the other hand,even though the kinase activity of p56lck is required for full T cell
*Signal Transduction Laboratory Mogam Biotechnology Research Institute,Koosungmyon, Yonginsi, Kyunggido, Korea; and†Department of Molecular LifeScience and Center for Cell Signaling Research, Ewha Women’s University, Seo-daemungu, Daehyundong 11-1, Seoul, 120-750, Korea.
Received for publication February 10, 1999. Accepted for publication August30, 1999.
The costs of publication of this article were defrayed in part by the payment of pagecharges. This article must therefore be hereby markedadvertisementin accordancewith 18 U.S.C. Section 1734 solely to indicate this fact.1 This work was supported in part by grants from the Korea Green Cross Co. and theMinistry of Science and Technology of Korea.2 The sequence reported in this article will appear in the GenBank database underaccession no. U69460.3 Address correspondence and requests to Dr. Y. Yun, Division of Molecular LifeScience and Center for Cell Signaling Research, Ewha Women’s University, Seoul,120-750, Korea. E-mail adddress: [email protected]
4 Abbreviations used in this paper: PTK, protein tyrosine kinase; SH1, Src homologydomain 1; PI3K, phosphatidylinositol 3-kinase; pY, phosphotyrosine; 59-RACE, 59-rapid amplification of cDNA ends; Lad, Lck-associated adapter protein; TSAd, Tcell-specific adapter protein; WT, wild type.
Copyright © 1999 by The American Association of Immunologists 0022-1767/99/$02.00
by guest on June 12, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

activation, a kinase-independent function of p56lck, mainly medi-ated by the SH2 domain was shown to independently contribute toT cell activation (7, 30–32). Subsequent efforts resulted in theidentification of ZAP-70 (33), CD45 (34, 35), and Sam68 (36) asbinding partners of the p56lck SH2 domain. However, this infor-mation does not fully explain the contribution of the SH2 domainto the multiple functions of p56lck in T cells.
Here, to understand the mechanisms by which p56lck acts in Tcell signaling and the role of the p56lck SH2 domain in this pro-cess, we identified the binding partners of the p56lck SH2 domainusing a tyrosine phosphorylation-dependent yeast two-hybrid sys-tem. As a result of the screening, a novel protein of 366 aa that wenamed Lad (Lck adapter) was isolated. Upon T cell activation, Ladcoimmunoprecipitated p56lck, was tyrosine phosphorylated, andacted as a substrate of p56lck tyrosine kinase. Moreover, overex-pression of dominant negative Lad blocked the IL-2 promoter-driven transcriptional activation following TCR stimulation.Taken together, these results indicate that Lad plays an essentialrole as an adapter protein in p56lck-mediated T cell signaling.
Materials and MethodsPlasmids
All bait plasmids encoding parts of murine p56lck and Lad were generatedby cloning the PCR-amplified fragment into pGBT9, a GAL4 DNA bind-ing domain vector (Clontech, Palo Alto, CA). Primers withEcoRI (59primer) or BamHI (39 primer) restriction sites were used to facilitate thesubcloning. The PCR products were digested withEcoRI/BamHI andcloned into the corresponding site of pGBT9. The mammalian expressionplasmids of p56lck wild type (WT), F505, A273, K154, Lad, Lad-antisense,and Lad-SH2 were constructed by PCR amplification and subsequent in-sertion into theEcoRI/XhoI site of pcDNAI/Amp (Invitrogen, San Diego,CA) using the same approach as described above. The point mutants ofp56lck were generated using a Quick mutagenesis kit (Stratagene, La Jolla,CA). The GST-fusion constructs of the SH2 domain of p56lck (aa 123–225)or the Lad C terminus (aa 208–366) were generated by PCR and subclonedinto pGEX-KG (Pharmacia, Piscataway, NJ) usingEcoRI orBamHI/EcoRIsites, respectively. The pGL3/IL-2-Luc contains the luciferase reportergene downstream from the IL-2 promoter region, including 548 bp 59 of thetranscriptional starting site, and was generated by subcloning aHindIIIfragment of pIL-2-CAT (37) into theHindIII site of pGL3-Basic (Promega,Madison, WI).
Yeast two hybrid screen
A cDNA fragment encoding the SH2 and kinase domain (the constitutivelyactive form, F505) of mouse p56lck was cloned into pGBT9. The resultingplasmid, Lck SH2K (F505) was used as the bait in the yeast two-hybridscreens of a murine T cell lymphoma cDNA library cloned into pACT(Clontech). The bait and library DNAs were cotransformed by the lithiumacetate method as previously described (38). Seventy-five out of 23 106
transformants grew in the absence of histidine and showed detectableb-ga-lactosidase staining within 2 h of incubation. To eliminate clones bindingto the bait in a tyrosine phosphorylation-independent manner, the plasmidsfrom each positive clone were cotransformed with control bait plasmidsencoding SH2 domain only (Lck SH2) or kinase domain only (Lck K(F505)). Approximately 70 clones, which did not bind to either of the twocontrol baits, were finally selected and subjected to partial sequence de-termination. The sequences revealed that seven clones contained a part ofthe same cDNA encoding a novel protein which we named Lad.
cDNA cloning
One of the Lad clones isolated from the yeast two hybrid screen was em-ployed as a probe to screen the mouse spleen cDNA library inlgt10 (Clon-tech). Three independent clones were obtained and subjected to nucleotidesequencing. In addition, 59-RACE was performed using the mouse spleen59 stretch cDNA template (Clontech). From the combination of both ap-proaches, the cDNA encompassing;1.6 kb in its entirety was obtained.
Cells, activation, and transfection
Jurkat and EL4 cells were obtained from the American Type Culture Col-lection (Manassas, VA) and maintained in RPMI 1640 medium supple-mented with 10% FBS, 5 mM 2-ME, and antibiotics. COS-1 cells were
grown in DMEM supplemented with 10% FBS and antibiotics. Thymo-cytes and splenocytes were isolated by passing mouse thymus and spleenthrough a sieve. Jurkat T cells were activated by cross-linking CD3 andCD4 with corresponding Abs, OKT3 and OKT4 (a gift of Dr. Shin, Har-vard Medical School, Boston, MA), respectively, at a saturated concentra-tion. EL4 cells were activated by CD3 cross-linking with 145-2C11 Ab.COS-1 cells were transfected using the standard DEAE-dextran method.
Abs, immunoprecipitation, and Western blot analysis
Anti-p56lck Ab was obtained from Transduction Laboratory (Lexington,KY) or generated by immunization of rabbits with GST fusion proteinsencompassing the SH3 and SH2 domain (aa 66–224). Antiphosphotyrosine(pY) Ab (4G10) and anti-SHP Ab were obtained from Upstate Biotech-nology (Lake Placid, NY). Anti-mouse CD3e (145-2C11), anti-CD90(Thy-1) (G7), anti-human CD3 (UCHT1), anti-hamster IgG, and anti-mouse IgG were obtained from PharMingen (San Diego, CA). Polyclonalantiserum against Lad or GST was raised in rabbits immunized with GST-Lad C terminus (aa 208–366) or GST, respectively. For immunoprecipi-tation or Western blot, cells were lysed with TNE buffer (50 mM Tris (pH8.0), 150 mM NaCl, 2 mM EDTA, 1% Nonidet P-40, 1 mM Na3VO4, 5mM NaF, 25mg/ml aprotinin, 1 mM PMSF, 25mg/ml leupeptin, and 1mg/ml BSA) for 1 h on ice. All bands analyzed by Western blot weredetected using the enhanced chemiluminescence protocol (Amersham, Ar-lington, Heights, IL).
Purification of GST fusion proteins and the binding assay
The GST fusion protein encompassing the SH2 domain of p56lck (aa 123–224) or the C terminus of Lad (aa 208–366) was expressed inEscherichiacoli and purified as previously described (39). For the binding study, 5mgof GST-p56lck SH2 fusion protein immobilized on glutathione-Sepharose4B beads was incubated with Jurkat T cell lysates prepared in TNE bufferfor 1 h at 4°C. After washing, samples were analyzed by Western blottingusing the antiserum against GST-LadC.
In vitro kinase assay
Purified GST-Lad C terminus or GST protein was incubated with 20 U ofpurified p56lck (Upstate Biotechnology) in a kinase buffer containing 50mM Tris (pH 7.5), 1 mM DTT, 10 mM MnCl2, and 50 mM NaCl in thepresence or absence of 100mM ATP. After 2 h at 30°C, the reactionmixtures were subjected to SDS-PAGE followed by Western blot analysisusing anti-pY Ab, 4G10 or anti-GST Ab.
Subcellular fractionation
All steps were performed as described (40). Approximately 23 107 EL4cells were activated by cross-linking CD3, resuspended in 0.5 ml of hy-potonic solution (25 mM Tris (pH 7.5), 5 mM EDTA, 5 mM EGTA, 250mM sucrose, 25mg/ml aprotinin, 1 mM PMSF, and 25mg/ml leupeptin),and then subjected to two successive freeze-thaw cycles. The cell suspen-sion was homogenized on ice using a Dounce homogenizer (40 strokes),and the salt concentration was adjusted to 150 mM NaCl. Nuclei and otherdebris in the cell lysates were removed by two rounds of centrifugation at4803 g for 5 min. The soluble and particulate fractions were separated bycentrifugation at 100,0003 g for 30 min. Fractionated proteins were re-solved by SDS-PAGE and Western blotted with the corresponding Abs.
Luciferase assay
A total of 5 3 106 Jurkat T cells were cotransfected with 2.5mg each ofIL-2-luc along with 2.5mg of pcDNAI/Lad, pcDNAI/Lad-antisense orpcDNAI/Lad-SH2 using Superfect (Qiagen, Chatswroth, CA). After incu-bation for 24 h with DNA-Superfect mixtures, cells were activated byincubation on anti-CD3 Ab plates coated with 5mg/ml UCHT1, by treat-ment of 10mg/ml PHA, or by treatment with 5 ng/ml PMA plus 0.5mg/mlA23187 for 14 h and were harvested.
ResultsIdentification of Lad as a phosphorylation-dependent bindingpartner of the p56lck SH2 domain
Based on the previous report that the binding specificity of phos-phopeptides to the p56lck SH2 domain overlaps with the substratespecificity of the p56lck kinase domain in a peptide library ap-proach (16), a part of p56lck encompassing the SH2 and kinasedomains (SH2K (F505) in Fig. 1A; aa 123–509) was used as a bait.
5243The Journal of Immunology
by guest on June 12, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

In this bait, Tyr505 was substituted for Phe505 to provide the con-stitutively active kinase form. In this system, presumably, the ty-rosine kinase activity of the bait will phosphorylate the bindingpartner proteins expressed from the cotransformed library, whichin turn will bind to the SH2 domain of the bait through pY to givea positive signal. From the screening of a total of 23 106 inde-pendent colonies of a murine T cell lymphoma cDNA library, 75strong positives were isolated and tested by retransformation witha series of control plasmids to confirm specificity and tyrosinephosphorylation-dependent binding (data not shown). Partial nu-cleotide sequencing of these cDNA fragments revealed that sevenclones encoded a portion of the same protein that we named Ladfor reasons that will be described in the latter part of this paper.
The binding specificity of Lad to p56lck was studied further us-ing the various control plasmids described in Fig. 1A. First, evenif the cloning vectors for Lad and SH2K (F505) were switched, theinteraction was not affected (Fig. 1B). Second, Lad interacted withSH2K (F505), but not with unrelated proteins such as GAL4 bind-ing domain only, p53 protein, and hepatitis B virus X-gene prod-uct. Third, Lad did not interact with ZAP-70 SH2-p56lck kinasefusion protein (ZAP-70 SH2K (F505)) or PI3K SH2-p56lck kinasefusion protein (PI3K SH2K (F505)) (Fig. 1B), suggesting that Ladmay preferentially bind to p56lck SH2 domain. Fourth, Lad did notinteract with either the SH2 domain or kinase domain (F505) only(Fig. 1C). This result supports the notion that the interaction be-tween SH2K (F505) and Lad involves cooperation between theSH2 domain and the active kinase domain rather than the individ-ual binding to either one domain. Finally, the pY-dependent inter-action was demonstrated further using the SH2 mutant (K154),which is incapable of binding to pY and the kinase mutant (A273),which is catalytcally inactive (Fig. 1C). Neither of these mutantsshowed any binding to Lad.
Structural characteristics of Lad
Next, a full open reading frame of Lad was obtained through thecombination of screening of a mouse lung cDNA library and 59-RACE from the mouse spleen 59stretch cDNA (Fig. 2A). The openreading frame of Lad encodes a 366-aa protein with homology toa T cell-specific adapter protein (TSAd) recently reported as aninducible human protein in activated T cells (41), even though thefunction of TSAd remained elusive. Lad is potentially a mousehomologue of TSAd and displays several interesting features of asignaling molecule. The structure of Lad and TSAd is conservedexcept for the presence of a zinc-finger motif in Lad (Fig. 2B). TheN terminus of Lad contains a CC-CC class of zinc-finger motif.Considering that some of the zinc finger motifs mediate protein-to-protein interaction, as exemplified by the association of ZPR1 tothe cytoplasmic tail of epidermal growth factor receptor (42), theCC-CC motif of Lad may act as an interface for signaling mole-cules. The central region contains an SH2 domain, which belongsto the 1b class of SH2 domains for which the amino acid atbD5position is tyrosine or phenylalanine (43). The highest identity isfound with the SH2 domains of GAPn, Csk, Grb2, and Src andfalls within the range of 25–35% (data not shown). The proline-rich motif of Lad exactly matches the consensus sequence1PPcPXKP (1, basic amino acids;c, hydrophobic amino acids)(44) preferred by class II cortactin SH3 domains found in cortactin,HS1, LckBP1, SH3P7, and SH3P8 (27, 45). The C terminus of Ladcontains four potential pY sites, one of which is an NPXpY292
motif known as the ligand for the pY binding domain (20). Over-all, these sequence characteristics suggest that Lad may be anadapter protein with several domains possibly involved in protein-protein interactions.
FIGURE 1. Binding properties of Lad to p56lck in a yeast two-hybridsystem.A, Schematic representation of the baits used in the yeast two-hybrid assay.B, Bait-specific interaction of Lad. In the swapping experi-ment, Lad was fused to GAL4 BD and SH2K (F505) was fused to thetranscriptional activation domain (AD) of GAL4. Interaction of Lad withunrelated proteins, such as GAL4 BD only, p53, hepatitis B Virus X-geneproduct (HBV-X), ZAP-70 SH2-p56lck kinase (ZAP-70 SH2K (F505)), andPI3K SH2-p56lck kinase (PI3K SH2K (F505))was tested for control.C,Tyrosine phosphorylation-dependent interaction of Lad to p56lck SH2domain. Binding affinity of SH2K (F505) to Lad was compared with thoseof other baits. Binding affinity was scored as111 (deep blue),11 (in-termediate blue),1 (pale blue), or2 (white) upon X-Gal (5-bromo-4-chloro-3-indolylb-D-galactoside) staining.
5244 Lck-BINDING ADAPTER PROTEIN: Lad
by guest on June 12, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

On Northern blot analysis, Lad cDNA detected messages of 1.7and 4.4 kb in spleen and lung of mouse and of 1.8 and 4.9 kb inspleen, thymus, and PBMC of human (Fig. 3A). The size of 1.7-kbmessage in mouse is consistent with that of assembled cDNA. Atthis stage, the identity of 4.4-kb and 4.9-kb message is not clear.
Next, an anti-Lad Ab was raised against the GST fusion proteincovering the C terminus of Lad (GST-LadC, aa 208–366). On
Western blot analysis with this Ab, Lad was detected as a 45-kDaprotein in EL4 mouse thymoma cells and mouse lymphocytes pu-rified from thymus and spleen, but not in NIH 3T3 cells (Fig. 3B).Notably, the expression level of Lad protein in thymus was muchhigher than that in spleen. In addition, size of in vitro translatedLad was identical to that in EL4 cells (Fig. 3C), even though thecalculated molecular mass was 40 kDa, indicating the authenticityof the predicted open reading frame.
Lad associates with p56lck upon TCR stimulation
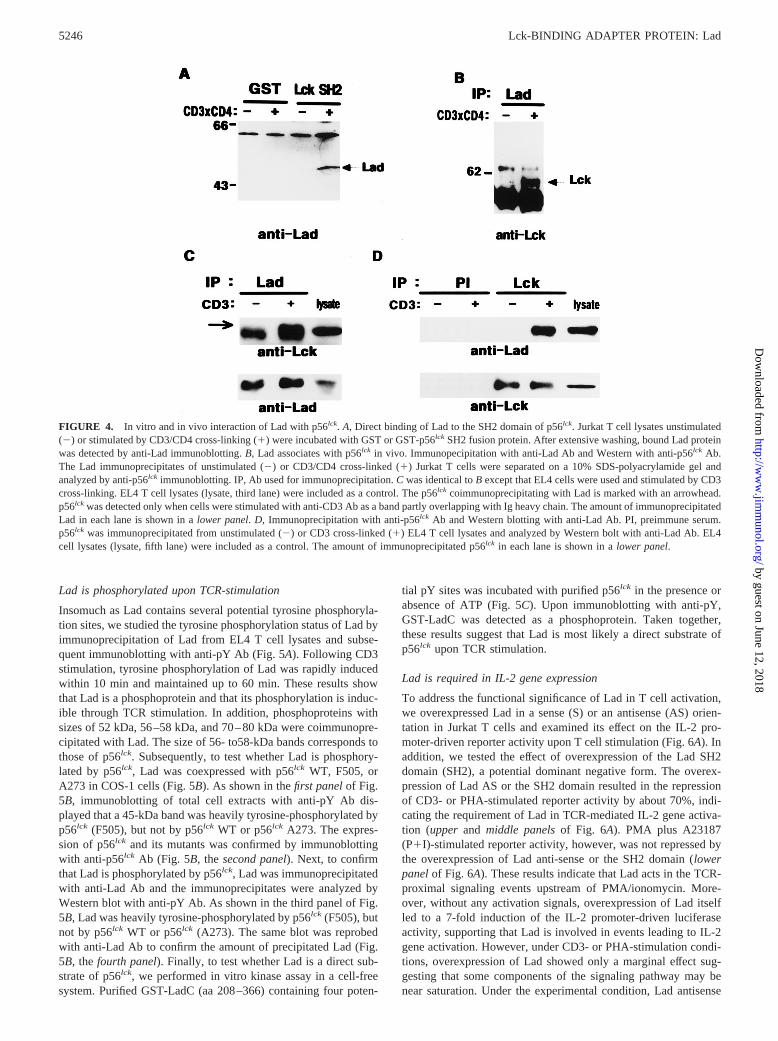
To confirm the binding of Lad with p56lck, GST fusion proteins ofp56lck SH2 domain bound to glutathione-Sepharose 4B beads wereincubated with Jurkat T cell lysates unstimulated/stimulated withanti-CD4 and CD3 mAbs (Fig. 4A). The surface expression ofCD3 and CD4 on Jurkat T cells were confirmed by FACS analysis.CD3 and CD4 were detected in;90% and 50% of the cells, re-spectively, and the total level of tyrosine phosphorylation was en-hanced by CD3/CD4 cross-linking compared with that by CD3cross-linking alone (data not shown). As shown in Fig. 4A, theGST-p56lck SH2 fusion protein precipitated a 52-kDa band, cor-responding to the m.w. of TSAd. The amount of precipitated Ladwas increased by;3-fold upon T cell activation.
Next, the in vivo interaction of Lad to p56lck was analyzed bycoimmunoprecipitation studies in Jurkat or EL4 cells. For the em-ployed EL4 cells, we observed that tyrosine phosphorylation of thecellular proteins was rapidly induced upon CD3 cross-linking (datanot shown). A mAb (145-2C11) raised against the CD3e of themurine TCR has been shown to induce IL-2 expression in EL4cells (46, 47) and was employed for stimulation of EL4 cells.p56lck could be readily detected in the Lad immunoprecipitates ofthe T cell lysates upon CD3/CD4 cross-linking of Jurkat T cells(Fig. 4B) or CD3 cross-linking of EL4 cells (Fig. 4C). Conversely,Lad was readily detected in the Lck immunoprecipitates upon CD3stimulation of EL4 cells (Fig. 4D). These results indicate that Ladinducibly binds to the SH2 domain of p56lck in vivo upon T cellactivation through TCR.
FIGURE 2. Structural characteristicsof Lad. A, Comparison of the deducedamino acid sequence of Lad with that ofTSAd. The amino acids conserved be-tween Lad and TSAd are marked by as-terisk (p). Potential protein-protein inter-action motifs, a zinc-finger motif, an SH2domain, a proline-rich SH3 binding mo-tif, and four pY sites are underlined.B,Schematic representation of the Ladstructure.
FIGURE 3. Expression of Lad.A, Tissue distribution of Lad mRNA.Northern blot analysis was performed with mouse and human MultipleTissue Northern blots (Clontech).B, Expression of Lad protein. Westernblot analysis was performed with EL4, NIH 3T3, and purified thymocytesand splenocytes at 53 106 cells/lane.C. Molecular weight of Lad. Lad isdetected as a 45-kDa protein upon in vitro translation and in T cell lysates.(2), control without Lad mRNA; Lad, in vitro translation product of LadmRNA; EL4, lysates of mouse EL4 T cell.
5245The Journal of Immunology
by guest on June 12, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
Lad is phosphorylated upon TCR-stimulation
Insomuch as Lad contains several potential tyrosine phosphoryla-tion sites, we studied the tyrosine phosphorylation status of Lad byimmunoprecipitation of Lad from EL4 T cell lysates and subse-quent immunoblotting with anti-pY Ab (Fig. 5A). Following CD3stimulation, tyrosine phosphorylation of Lad was rapidly inducedwithin 10 min and maintained up to 60 min. These results showthat Lad is a phosphoprotein and that its phosphorylation is induc-ible through TCR stimulation. In addition, phosphoproteins withsizes of 52 kDa, 56–58 kDa, and 70–80 kDa were coimmunopre-cipitated with Lad. The size of 56- to58-kDa bands corresponds tothose of p56lck. Subsequently, to test whether Lad is phosphory-lated by p56lck, Lad was coexpressed with p56lck WT, F505, orA273 in COS-1 cells (Fig. 5B). As shown in thefirst panelof Fig.5B, immunoblotting of total cell extracts with anti-pY Ab dis-played that a 45-kDa band was heavily tyrosine-phosphorylated byp56lck (F505), but not by p56lck WT or p56lck A273. The expres-sion of p56lck and its mutants was confirmed by immunoblottingwith anti-p56lck Ab (Fig. 5B, thesecond panel). Next, to confirmthat Lad is phosphorylated by p56lck, Lad was immunoprecipitatedwith anti-Lad Ab and the immunoprecipitates were analyzed byWestern blot with anti-pY Ab. As shown in the third panel of Fig.5B, Lad was heavily tyrosine-phosphorylated by p56lck (F505), butnot by p56lck WT or p56lck (A273). The same blot was reprobedwith anti-Lad Ab to confirm the amount of precipitated Lad (Fig.5B, thefourth panel). Finally, to test whether Lad is a direct sub-strate of p56lck, we performed in vitro kinase assay in a cell-freesystem. Purified GST-LadC (aa 208–366) containing four poten-
tial pY sites was incubated with purified p56lck in the presence orabsence of ATP (Fig. 5C). Upon immunoblotting with anti-pY,GST-LadC was detected as a phosphoprotein. Taken together,these results suggest that Lad is most likely a direct substrate ofp56lck upon TCR stimulation.
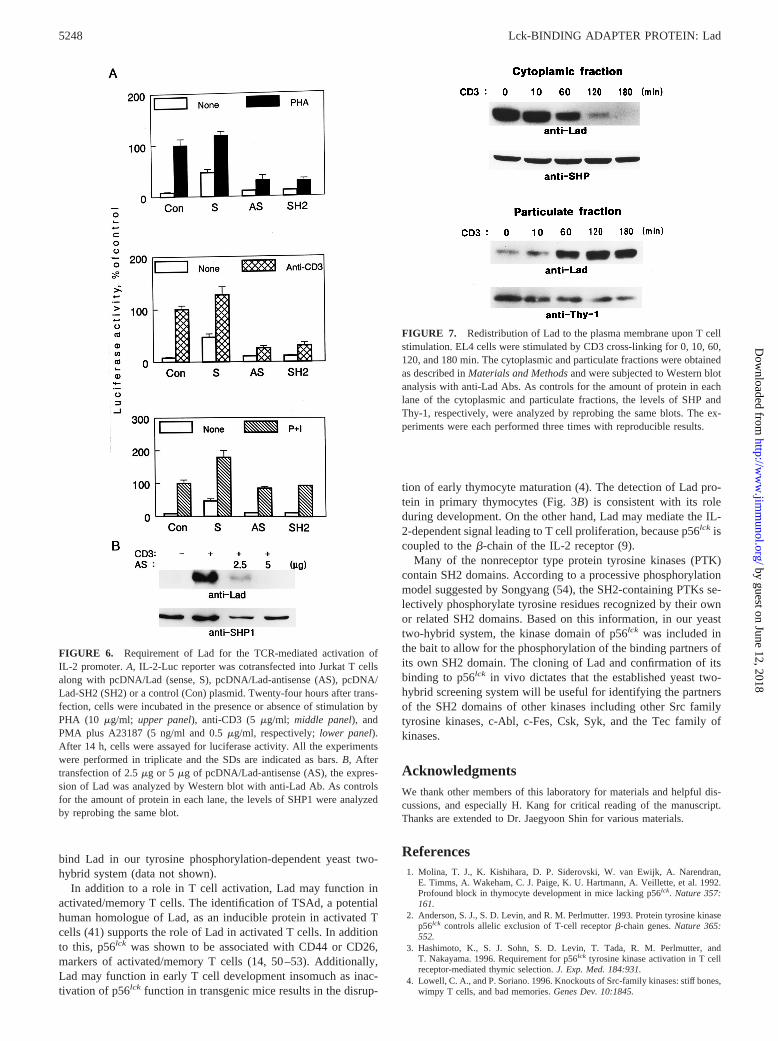
Lad is required in IL-2 gene expression
To address the functional significance of Lad in T cell activation,we overexpressed Lad in a sense (S) or an antisense (AS) orien-tation in Jurkat T cells and examined its effect on the IL-2 pro-moter-driven reporter activity upon T cell stimulation (Fig. 6A). Inaddition, we tested the effect of overexpression of the Lad SH2domain (SH2), a potential dominant negative form. The overex-pression of Lad AS or the SH2 domain resulted in the repressionof CD3- or PHA-stimulated reporter activity by about 70%, indi-cating the requirement of Lad in TCR-mediated IL-2 gene activa-tion (upper and middle panelsof Fig. 6A). PMA plus A23187(P1I)-stimulated reporter activity, however, was not repressed bythe overexpression of Lad anti-sense or the SH2 domain (lowerpanelof Fig. 6A). These results indicate that Lad acts in the TCR-proximal signaling events upstream of PMA/ionomycin. More-over, without any activation signals, overexpression of Lad itselfled to a 7-fold induction of the IL-2 promoter-driven luciferaseactivity, supporting that Lad is involved in events leading to IL-2gene activation. However, under CD3- or PHA-stimulation condi-tions, overexpression of Lad showed only a marginal effect sug-gesting that some components of the signaling pathway may benear saturation. Under the experimental condition, Lad antisense
FIGURE 4. In vitro and in vivo interaction of Lad with p56lck. A, Direct binding of Lad to the SH2 domain of p56lck. Jurkat T cell lysates unstimulated(2) or stimulated by CD3/CD4 cross-linking (1) were incubated with GST or GST-p56lck SH2 fusion protein. After extensive washing, bound Lad proteinwas detected by anti-Lad immunoblotting.B, Lad associates with p56lck in vivo. Immunopecipitation with anti-Lad Ab and Western with anti-p56lck Ab.The Lad immunoprecipitates of unstimulated (2) or CD3/CD4 cross-linked (1) Jurkat T cells were separated on a 10% SDS-polyacrylamide gel andanalyzed by anti-p56lck immunoblotting. IP, Ab used for immunoprecipitation.C was identical toB except that EL4 cells were used and stimulated by CD3cross-linking. EL4 T cell lysates (lysate, third lane) were included as a control. The p56lck coimmunoprecipitating with Lad is marked with an arrowhead.p56lck was detected only when cells were stimulated with anti-CD3 Ab as a band partly overlapping with Ig heavy chain. The amount of immunoprecipitatedLad in each lane is shown in alower panel.D, Immunoprecipitation with anti-p56lck Ab and Western blotting with anti-Lad Ab. PI, preimmune serum.p56lck was immunoprecipitated from unstimulated (2) or CD3 cross-linked (1) EL4 T cell lysates and analyzed by Western bolt with anti-Lad Ab. EL4cell lysates (lysate, fifth lane) were included as a control. The amount of immunoprecipitated p56lck in each lane is shown in alower panel.
5246 Lck-BINDING ADAPTER PROTEIN: Lad
by guest on June 12, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

effectively inhibited the expression of Lad protein induced uponCD3 stimulation in a dose-dependent manner (Fig. 6B). Takentogether, these results demonstrate that Lad is required for theTCR-mediated signaling pathway leading to IL-2 gene expression.
Activation-dependent plasma membrane localization of Lad
To examine the subcellular localization of Lad, we prepared thecytoplasmic and particulate fractions from CD3-stimulated EL4cells and studied the distribution of Lad by Western blot analysis(Fig. 7). In the absence of stimulation, the majority of Lad wasdetected in the cytoplasmic fraction. Upon stimulation by CD3cross-linking, the level of cytoplasmic Lad gradually diminishedand was almost nondetectable after 2 h, whereas the level of par-ticulate Lad gradually increased during that time. On the otherhand, the levels of SHP-1 and Thy-1, employed as controls, wereconsistent throughout the period (Fig. 7). In addition, redistributionof Lad to the plasma membrane was also observed in the immu-nofluorescence assay with anti-Lad Ab (data not shown). The ob-served redistribution to the plasma membrane provides Lad withan opportunity to interact with p56lck upon TCR engagement andsupports the model that Lad acts as a partner of p56lck at the mem-brane-proximal signaling pathway of TCR activation.
DiscussionIn this report, we have described the molecular cloning and char-acterization of a p56lck-binding protein, Lad. We have found that:1) Lad is expressed in T cells; 2) Lad binds to the SH2 domain ofp56lck in vitro and in vivo in a T cell activation-dependent manner;3) Lad is tyrosine phosphorylated upon TCR stimulation; 4) Lad is
involved in the up-regulation of the IL-2 gene expression upon Tcell activation; and 5) Lad is redistributed to the plasma membraneupon TCR stimulation. These results indicate that Lad physicallyinteracts with p56lck upon T cell activation and plays an importantrole in the p56lck-dependent T cell signaling.
Insomuch as Lad is directly phosphorylated by p56lck in intactcells and by purified p56lck in vitro (Fig. 5, B andC), it is mostlikely that the tyrosine phosphorylation of Lad upon T cell acti-vation is mediated by p56lck. However, we do not exclude thepossibility that the initial tyrosine phosphorylation is mediated byother tyrosine kinases (e.g., Csk, ZAP-70, Tec family, and p59fyn)known to be involved in T cell activation. Upon translocation tothe plasma membrane, the SH2 domain, proline-rich motif or zinc-finger motif of Lad may recruit additional signaling molecules tothe p56lck/TCR complex leading to the amplification of activationsignals.
Four potential pY sites are concentrated in the C terminus ofLad. Notably, the NPXpY292 and NXpY317 motifs match the se-quences known to be recognized by pY binding domains, whichare found in signaling proteins such as Shc and IRS-1 (NPXpY),or Cbl (NxpY) (48, 49). However, in our preliminary experiments,association of Shc to Lad was not detected upon T cell activation(data not shown). On the other hand, all four pY motifs(pY275TSP, pY292QEP, pY302AMG, and pY317AEV) may serve assubstrates for various SH2 domains. The pYXXP motif is pre-ferred by the SH2 domains of Crk, PLCg1 and c-Abl, and thepYXXV motif is preferred by the SH2 domains of Src familytyrosine kinases (43). Consistent with this possibility, we havepreliminarily observed that the SH2 domains of Grb-2 and PLC-g1
FIGURE 5. Analysis of Lad phosphorylation.A, Lad is tyrosine phosphorylated upon T cellactivation. Lad was immunoprecipitated fromEL4 T cell lysates activated by CD3 cross-link-ing for 0, 10, 30, and 60 min. The immunopre-cipitates were subjected to Western blot analysiswith anti-pY (4G10) Ab (upper panel) or anti-Lad Ab (lower panel).B, Phosphorylation ofLad by p56lck in COS-1 cells. COS-1 cells weretransfected with plasmids expressing Lad only(lane 1), Lad plus p56lck (WT) (lane 2), Lad plusp56lck (F505) (lane 3), and Lad plus p56lck
(A273) (lane 4). Total cell extracts were ana-lyzed by Western blot with 4G10 (first panel)and anti-p56lck Ab (second panel). Subse-quently, Lad was immunoprecipitated fromCOS-1 cell extracts and the immunoprecipitateswere analyzed by Western with 4G10 (third pan-el), or anti-Lad Ab (fourth panel).C, In vitrophosphorylation of Lad by p56lck. Upper panel,purified GST or GST-LadC (Lad C terminus, aa208–366) was incubated with purified p56lck inthe presence (1) or absence (2) of ATP. Thekinase reaction mixtures were analyzed with anti-pY immunoblotting. Thelower panelshows theamount of loaded GST or GST-LadC as acontrol.
5247The Journal of Immunology
by guest on June 12, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
bind Lad in our tyrosine phosphorylation-dependent yeast two-hybrid system (data not shown).
In addition to a role in T cell activation, Lad may function inactivated/memory T cells. The identification of TSAd, a potentialhuman homologue of Lad, as an inducible protein in activated Tcells (41) supports the role of Lad in activated T cells. In additionto this, p56lck was shown to be associated with CD44 or CD26,markers of activated/memory T cells (14, 50–53). Additionally,Lad may function in early T cell development insomuch as inac-tivation of p56lck function in transgenic mice results in the disrup-
tion of early thymocyte maturation (4). The detection of Lad pro-tein in primary thymocytes (Fig. 3B) is consistent with its roleduring development. On the other hand, Lad may mediate the IL-2-dependent signal leading to T cell proliferation, because p56lck iscoupled to theb-chain of the IL-2 receptor (9).
Many of the nonreceptor type protein tyrosine kinases (PTK)contain SH2 domains. According to a processive phosphorylationmodel suggested by Songyang (54), the SH2-containing PTKs se-lectively phosphorylate tyrosine residues recognized by their ownor related SH2 domains. Based on this information, in our yeasttwo-hybrid system, the kinase domain of p56lck was included inthe bait to allow for the phosphorylation of the binding partners ofits own SH2 domain. The cloning of Lad and confirmation of itsbinding to p56lck in vivo dictates that the established yeast two-hybrid screening system will be useful for identifying the partnersof the SH2 domains of other kinases including other Src familytyrosine kinases, c-Abl, c-Fes, Csk, Syk, and the Tec family ofkinases.
AcknowledgmentsWe thank other members of this laboratory for materials and helpful dis-cussions, and especially H. Kang for critical reading of the manuscript.Thanks are extended to Dr. Jaegyoon Shin for various materials.
References1. Molina, T. J., K. Kishihara, D. P. Siderovski, W. van Ewijk, A. Narendran,
E. Timms, A. Wakeham, C. J. Paige, K. U. Hartmann, A. Veillette, et al. 1992.Profound block in thymocyte development in mice lacking p56lck. Nature 357:161.
2. Anderson, S. J., S. D. Levin, and R. M. Perlmutter. 1993. Protein tyrosine kinasep56lck controls allelic exclusion of T-cell receptorb-chain genes.Nature 365:552.
3. Hashimoto, K., S. J. Sohn, S. D. Levin, T. Tada, R. M. Perlmutter, andT. Nakayama. 1996. Requirement for p56lck tyrosine kinase activation in T cellreceptor-mediated thymic selection.J. Exp. Med. 184:931.
4. Lowell, C. A., and P. Soriano. 1996. Knockouts of Src-family kinases: stiff bones,wimpy T cells, and bad memories.Genes Dev. 10:1845.
FIGURE 6. Requirement of Lad for the TCR-mediated activation ofIL-2 promoter.A, IL-2-Luc reporter was cotransfected into Jurkat T cellsalong with pcDNA/Lad (sense, S), pcDNA/Lad-antisense (AS), pcDNA/Lad-SH2 (SH2) or a control (Con) plasmid. Twenty-four hours after trans-fection, cells were incubated in the presence or absence of stimulation byPHA (10 mg/ml; upper panel), anti-CD3 (5 mg/ml; middle panel), andPMA plus A23187 (5 ng/ml and 0.5mg/ml, respectively;lower panel).After 14 h, cells were assayed for luciferase activity. All the experimentswere performed in triplicate and the SDs are indicated as bars.B, Aftertransfection of 2.5mg or 5 mg of pcDNA/Lad-antisense (AS), the expres-sion of Lad was analyzed by Western blot with anti-Lad Ab. As controlsfor the amount of protein in each lane, the levels of SHP1 were analyzedby reprobing the same blot.
FIGURE 7. Redistribution of Lad to the plasma membrane upon T cellstimulation. EL4 cells were stimulated by CD3 cross-linking for 0, 10, 60,120, and 180 min. The cytoplasmic and particulate fractions were obtainedas described inMaterials and Methodsand were subjected to Western blotanalysis with anti-Lad Abs. As controls for the amount of protein in eachlane of the cytoplasmic and particulate fractions, the levels of SHP andThy-1, respectively, were analyzed by reprobing the same blots. The ex-periments were each performed three times with reproducible results.
5248 Lck-BINDING ADAPTER PROTEIN: Lad
by guest on June 12, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

5. Turner, J. M., M. H. Brodsky, B. A. Irving, S. D. Levin, R. M. Perlmutter, andD. R. Littman. 1990. Interaction of the unique N-terminal region of tyrosinekinase p56lck with cytoplasmic domains of CD4 and CD8 is mediated by cysteinemotifs. Cell 60:755.
6. Glaichenhaus, N., N. Shastri, D. R. Littman, and J. M. Turner. 1991. Requirementfor association of p56lck with CD4 in antigen-specific signal transduction in Tcells.Cell 64:511.
7. Collins, T. L., and S. J. Burakoff. 1993. Tyrosine kinase activity of CD4-asso-ciated p56lck may not be required for CD4-dependent T-cell activation.Proc.Natl. Acad. Sci. USA 90:11885.
8. Straus, D. B., and A. Weiss. 1992. Genetic evidence for the involvement of thelck tyrosine kinase in signal transduction through the T cell antigen receptor.Cell70:585.
9. Hatakeyama, M., T. Kono, N. Kobayashi, A. Kawahara, S. D. Levin,R. M. Perlmutter, and T. Taniguchi. 1991. Interaction of the IL-2 receptor withthe src-family kinase p56lck: identification of novel intermolecular association.Science 252:1523.
10. Minami, Y., T. Kono, K. Yamada, N. Kobayashi, A. Kawahara, R. M. Perlmutter,and T. Taniguchi. 1993. Association of p56lck with IL-2 receptorb chain iscritical for the IL-2-induced activation of p56lck. EMBO J. 12:759.
11. Miyazaki, T., Z. J. Liu, A. Kawahara, Y. Minami, K. Yamada, Y. Tsujimoto,E. L. Barsoumian, R. M. Perlmutter, and T. Taniguchi. 1995. Three distinct IL-2signaling pathways mediated by bcl-2, c-myc, andlck cooperate in hematopoieticcell proliferation.Cell 81:223.
12. Kim, Y. J., K. E. Pollok, Z. Zhou, A. Shaw, J. B. Bohlen, M. Fraser, andB. S. Kwon. 1993. Novel T cell antigen 4–1BB associates with the protein ty-rosine kinase p56lck. J. Immunol. 151:1255.
13. Bell, G. M., J. Fargnoli, J. B. Bolen, L. Kish, and J. B. Imboden. 1996. The SH3domain of p56lck binds to proline-rich sequences in the cytoplasmic domain ofCD2. J. Exp. Med. 183:169.
14. Taher, T. E., L. Smit, A. W. Griffioen, E. J. Schilder-Tol, J. Borst, and S. T. Pals.1996. Signaling through CD44 is mediated by tyrosine kinases: association withp56lck in T lymphocytes.J. Biol. Chem. 271:2863.
15. Brenner, B., E. Gulbins, K. Schlottmann, U. Koppenhoefer, G. L. Busch,B. Walzog, M. Steinhausen, K. M. Coggeshall, O. Linderkamp, and F. Lang.1996. L-selectin activates the Ras pathway via the tyrosine kinase p56lck. Proc.Natl. Acad. Sci. USA 93:15376.
16. Zhou, S., R., K. L. Carraway, M. J. Eck, S. C. Harrison, R. A. Feldman,M. Mohammadi, J. Schlessinger, S. R. Hubbard, D. P. Smith, C. Eng, et al. 1995.Catalytic specificity of protein-tyrosine kinases is critical for selective signalling.Nature 373:536.
17. Carrera, A. C., H. Paradis, L. R. Borlado, T. M. Roberts, and C. Martinez. 1995.Lck unique domain influences Lck specificity and biological function.J. Biol.Chem. 270:3385.
18. Gervais, F. G., and A. Veillette. 1995. The unique amino-terminal domain ofp56lck regulates interactions with tyrosine protein phosphatases in T lymphocytes.Mol. Cell. Biol. 15:2393.
19. Paige, L. A., M. J. Nadler, M. L. Harrison, J. M. Cassady, and R. L. Geahlen.1993. Reversible palmitoylation of the protein-tyrosine kinase p56lck. J. Biol.Chem. 268:8669.
20. Kavanaugh, W. M., C. W. Turck, and L. T. Williams. 1995. PTB domain bindingto signaling proteins through a sequence motif containing phosphotyrosine.Sci-ence 268:1177.
21. Shenoy-Scaria, A. M., L. K. Gauen, J. Kwong, A. S. Shaw, and D. M. Lublin.1993. Palmitylation of an amino-terminal cysteine motif of protein tyrosine ki-nases p56lck and p59fyn mediates interaction with glycosyl-phosphatidylinositol-anchored proteins.Mol. Cell. Biol. 13:6385.
22. Veillette, A., L. Caron, M. Fournel, and T. Pawson. 1992. Regulation of theenzymatic function of the lymphocyte-specific tyrosine protein kinase p56lck bythe noncatalytic SH2 and SH3 domains.Oncogene 7:971.
23. Reynolds, P. J., T. R. Hurley, and B. M. Sefton. 1992. Functional analysis of theSH2 and SH3 domains of thelck tyrosine protein kinase.Oncogene 7:1949.
24. Prasad, K. V., R. Kapeller, O. Janssen, H. Repke, J. S. Duke-Cohan,L. C. Cantley, and C. E. Rudd. 1993. Phosphatidylinositol (PI) 3-kinase and PI4-kinase binding to the CD4–p56lck complex: the p56lck SH3 domain binds to PI3-kinase but not PI 4-kinase.Mol. Cell. Biol. 13:7708.
25. Vogel, L. B., and D. J. Fujita. 1993. The SH3 domain of p56lck is involved inbinding to phosphatidylinositol 39-kinase from T lymphocytes.Mol. Cell. Biol.13:7408.
26. Reedquist, K. A., T. Fukazawa, B. Druker, G. Panchamoorthy, S. E. Shoelson,and H. Band. 1994. Rapid T-cell receptor-mediated tyrosine phosphorylation ofp120, an Fyn/Lck Src homology 3 domain-binding protein.Proc. Natl. Acad. Sci.USA 91:4135.
27. Takemoto, Y., M. Furuta, X. K. Li, W. J. Strong-Sparks, and Y. Hashimoto.1995. LckBP1, a proline-rich protein expressed in haematopoietic lineage cells,directly associates with the SH3 domain of protein tyrosine kinase p56lck. EMBOJ. 14:3403.
28. Peri, K. G., F. G. Gervais, R. Weil, D. Davidson, G. D. Gish, and A. Veillette.1993. Interactions of the SH2 domain of lymphocyte-specific tyrosine proteinkinase p56lck with phosphotyrosine-containing proteins.Oncogene 8:2765.
29. Iwashima, M., B. A. Irving, N. S. van Oers, A. C. Chan, and A. Weiss. 1994.Sequential interactions of the TCR with two distinct cytoplasmic tyrosine ki-nases.Science 263:1136.
30. Xu, H., and D. R. Littman. 1993. A kinase-independent function of Lck in po-tentiating antigen-specific T cell activation.Cell 74:633.
31. Straus, D. B., A. C. Chan, B. Patai, and A. Weiss. 1996. SH2 domain function isessential for the role of the Lck tyrosine kinase in T cell receptor signal trans-duction.J. Biol. Chem. 271:9976.
32. Lewis, L. A., C. D. Chung, J. Chen, J. R. Parnes, M. Moran, V. P. Patel, andM. C. Miceli. 1997. The Lck SH2 phosphotyrosine binding site is critical forefficient TCR-induced processive tyrosine phosphorylation of thez-chain andIL-2 production.J. Immunol. 159:2292.
33. Duplay, P., M. Thome, F. Herve, and O. Acuto. 1994. p56lck interacts via its Srchomology 2 domain with the ZAP-70 kinase.J. Exp. Med. 179:1163.
34. Lee, J. M., M. Fournel, A. Veillette, and P. E. Branton. 1996. Association ofCD45 with Lck and components of the Ras signaling pathway in pervanadate-treated mouse T-cell lines.Oncogene 12:253.
35. Ng, D. H., J. D. Watts, R. Aebersold, and P. Johnson. 1996. Demonstration of adirect interaction between p56lck and the cytoplasmic domain of CD45 in vitro.J. Biol. Chem. 271:1295.
36. Fusaki, N., A. Iwamatsu, M. Iwashima, and J. i. Fujisawa. 1997. Interactionbetween Sam68 and Src family tyrosine kinases, Fyn and Lck, in T cell receptorsignaling.J. Biol. Chem. 272:6214.
37. Williams, T. M., J. E. Burlein, S. Ogden, L. J. Kricka, and J. A. Kant. 1989.Advantages of firefly luciferase as a reporter gene: application to the interleukin-2gene promoter.Anal. Biochem. 176:28.
38. Gietz, D., A. St. Jean, R. A. Woods, and R. H. Schiestl. 1992. Improved methodfor high efficiency transformation of intact yeast cells.Nucleic Acids Res. 20:1425.
39. Joung, I., T. Kim, L. A. Stolz, G. Payne, D. G. Winkler, C. T. Walsh,J. L. Strominger, and J. Shin. 1995. Modification of Ser59 in the unique N-terminal region of tyrosine kinase p56lck regulates specificity of its Src homology2 domain.Proc. Natl. Acad. Sci. USA 92:5778.
40. Kabouridis, P. S., A. I. Magee, and S. C. Ley. 1997. S-acylation of Lck proteintyrosine kinase is essential for its signaling function in T lymphocytes.EMBO J.16:4983.
41. Spurkland, A., J. E. Brinchmann, G. Markussen, F. Pedeutour, E. Munthe, T. Lea,F. Vartdal, and H. C. Aasheim. 1998. Molecular cloning of a T cell-specificadapter protein (TSAd) containing an Src homology (SH) 2 domain and putativeSH3 and phosphotyrosine binding sites.J. Biol. Chem. 273:4539.
42. Galcheva-Gargova, Z., K. N. Konstantinov, I. H. Wu, F. G. Klier, T. Barrett, andR. J. Davis. 1996. Binding of zinc finger protein ZPR1 to the epidermal growthfactor receptor.Science 272:1797.
43. Songyang, Z., S. E. Shoelson, M. Chaudhuri, G. Gish, T. Pawson, W. G. Haser,F. King, T. Roberts, S. Ratnofsky, R. J. Lechleider, et al. 1993. SH2 domainsrecognize specific phosphopeptide sequences.Cell 72:767.
44. Sparks, A. B., J. E. Rider, N. G. Hoffman, D. M. Fowlkes, L. A. Quillam, andB. K. Kay. 1996. Distinct ligand preferences of Src homology 3 domains fromSrc, Yes, Abl, Cortactin, p53bp2, PLCg, Crk, and Grb2.Proc. Natl. Acad. Sci.USA 93:1540.
45. Sparks, A. B., N. G. Hoffman, S. J. McConnell, D. M. Fowlkes, and B. K. Kay.1996. Cloning of ligand targets: systematic isolation of SH3 domain-containingproteins.Nat. Biotechnol. 14:741.
46. Rayter, S. I., M. Woodrow, S. C. Lucas, D. A. Cantrell, and J. Downward. 1992.p21ras mediates control of IL-2 gene promoter function in T cell activation.EMBO J. 11:4549.
47. Woodrow, M. A., S. Rayter, J. Downward, and D. A. Cantrell. 1993. p21ras
function is important for T cell antigen receptor and protein kinase C regulationof nuclear factor of activated T cells.J. Immunol. 150:3853.
48. Wolf, G., T. Trub, E. Ottinger, L. Groninga, A. Lynch, M. F. White,M. Miyazaki, J. Lee, and S. E. Shoelson. 1995. PTB domains of IRS-1 and Shchave distinct but overlapping binding specificities.J. Biol. Chem. 270:27407.
49. Lupher, M. L. Jr., Z. Songyang, S. E. Shoelson, L. C. Cantley, and H. Band. 1997.The Cbl phosphotyrosine-binding domain selects a D(N/D)XpY motif and bindsto the Tyr292 negative regulatory phosphorylation site of ZAP-70.J. Biol. Chem.272:33140.
50. Ulmer, A. J., T. Mattern, and H. D. Flad. 1992. Expression of CD26 (dipeptidylpeptidase IV) on memory and naive T lymphocytes.Scand. J. Immunol. 35:551.
51. Hegen, M., J. Kameoka, R. P. Dong, S. F. Schlossman, and C. Morimoto. 1997.Cross-linking of CD26 by antibody induces tyrosine phosphorylation and acti-vation of mitogen-activated protein kinase.Immunology 90:257.
52. Dutton, R. W., L. M. Bradley, and S. L. Swain. 1998. T cell memory.Annu. Rev.Immunol. 16:201.
53. Ilangumaran, S., A. Briol, and D. C. Hoessli. 1998. CD44 selectively associateswith active Src family protein tyrosine kinases Lck and Fyn in glycosphingolipid-rich plasma membrane domains of human peripheral blood lymphocytes.Blood91:3901.
54. Zhou, S., and L. C. Cantley. 1995. Recognition and specificity in protein tyrosinekinase-mediated signaling.Trends Biochem. Sci. 20:470.
5249The Journal of Immunology
by guest on June 12, 2018http://w
ww
.jimm
unol.org/D
ownloaded from





























