Embed Size (px)
Citation preview

UNIVERSIDAD NACIONAL DEL CENTRO DEL PERÙ
Presentado por: Sheyla Alvites Solís
Asignatura:Bioquímica
Docente:Ing. Rafael Pantoja Esquivel

TRANSPORTADORES DE LA
GLUCOSA EN LOS SERES VIVO
GLUT 1(SLC2A1): Un Glut de alta afinidad presente en tejidos que utilizan a la glucosa como combustible
principal
Encargado del transporte de glucosa y galactosa.
Se encuentra ampliamente distribuido en los
tejidos fetales, y en los adultos se encuentra
expresado en sus máximos niveles en eritrocitos y
en las células endoteliales del cerebro, en las
células neuronales (astrocitos), barreras
hematoencefalicas entre otras.
En el riñón se ha encontrado en todos los
segmentos de nefrones.
Función:
Su función principal sería la de mantener la
glucosa basal en la célula y posibilitar la entrada
de glucosa en reposo
Actúa como una puerta en la cual la proteína une
al azúcar en la superficie externa de la membrana
y sufre un cambio conformacional que conduce al
azúcar hacia el interior de la célula, donde se
desune.

GLUT 2 (SLC2A2): Un Glut con función glucosensora
Se expresa principalmente en las células
pancreáticas, en el hígado, en el riñón y en la
membrana basolateral del intestino delgado.
Transporta glucosa, galactosa y también fructosa.
Función:
Otro caso interesante es la intervención del Glut-2 en el
metabolismo hepático de la glucosa. Después de las comidas, el
hígado es capaz de incorporar la glucosa proveniente de los
alimentos gracias al Glut-2 para ser convertida rápidamente en
glucógeno. De forma inversa, durante el período post-pandrial
tardío (período comprendido de 6 a 8 horas después de las
comidas) el glucógeno sufre degradación generando moléculas
de glucosa que salen de la célula hepática a la sangre,
manteniendo así los niveles de glucosa plasmática dentro de
límites normales. De esta forma, es fácil notar que el Glut-2 es
un transportador de tipo bidireccional que puede transportar
glucosa desde la sangre al tejido o desde el tejido hacia la
sangre, hecho particularmente cierto a nivel hepático y renal
funcionando como sensor de la concentración plasmática de
glucosa y permitiendo su intercambio entre la sangre y el
hepatocito dependiendo de la condición alimentaria
predominante en el momento.
Actúa como un regulador que solo permite la entrada
de glucosa cuando está lo suficiente elevada en
plasma como para requerir la liberación de una
cantidad significativamente importante de insulina.

GLUT 3 (SLC2A3): El Glut de más alta afinidad por la glucosa
Función:
Presenta alta afinidad por la glucosa, se expresa en
tejidos que tienen un alto requerimiento de este azúcar,
aunque también transporta galactosa.
En el ser humano se presenta su mayor expresión en
el sistema nervioso central ya que es el principal
transportador de glucosa para las neuronas o las
células nerviosas, en la placenta, en el hígado, en el
riñón y en el corazón.
En el cerebro su función está acoplada al GLUT 1,
permitiendo el transporte vectorial de la glucosa
desde la sangre hasta las neuronas.
La deficiencia del GLUT 3 está relacionada con la
restricción del crecimiento intrauterino fetal, lo que
aumenta el riesgo de retraso mental y físico,
enfermedades cardiovasculares y diabetes tipo II.
La presencia de este transportador co-agregado con
el Glut-1 en tejido nervioso habla a favor de que este
transportador tenga funciones de mantenimiento del
nivel basal de glucosa en neuronas y placenta.

GLUT 4 (SLC2A4): Un Glut con gran movilidad
Función:
Se expresa fundamentalmente en tejido muscular
estriado, tejido muscular cardíaco y adipocito.
Es único en el sentido de la regulación de su
localización en el citosol o en la membrana por la
insulina. En condiciones basales, la vasta mayoría de
las moléculas de Glut-4 se encuentran localizadas
dentro de vesículas en el citosol que forman dos tipos
de compartimientos bien definidos, ya que un grupo de
estas vesículas responden a la señal de la insulina y
otro grupo responde fundamentalmente al estímulo que
representa la actividad física.
El GLUT-4 transporta las moléculas de glucosa dentro
de las adiposas. Tanto el sistema esquelético como las
células adiposas necesitan insulina y una proteína
transportadora de glucosa para absorber las moléculas
de glucosa del torrente sanguíneo. El páncreas libera
insulina que, posteriormente, se adhiere a receptores en
las membranas celulares esqueléticas y adiposas.
Debido a que el GLUT-4 es una proteína que responde
a la insulina, se alerta ante la presencia de insulina
unida a los receptores en la membrana celular. Luego,
la molécula GLUT-4 es capaz de transportar las
moléculas de glucosa a través de la membrana celular y
dentro de las células.

GLUT 5 (SLC2A5): Un Glut específico para la Fructosa
Función:
Transporta exclusivamente a la fructuosa, se
encuentra en el intestino delgado, en los
testículos y en los riñones. No muestra
afinidad por la glucosa.
Está formado por 501 aminoácidos y está
codificado por un gen localizado en el
cromosoma .
Su función principal es el transporte de
fructuosa.
Su expresión en el músculo esquelético
humano se relaciona a su capacidad de
utilizar la fructosa para la glucólisis y la
síntesis de glucógeno de forma
independiente a través de la incorporación
por medio del Glut-1 y el Glut-4.

RUTAS O DESTINOS CATABOLICOS DEL PIRUVATO
En los procesos catabólicos las moléculas orgánicas se van degradando, paso a paso, hasta formar otras moléculas
más simples y, finalmente, sustancias inorgánicas. Globalmente son procesos de oxidación en los que las moléculas
orgánicas van perdiendo electrones que, tras pasar por una cadena transportadora, son captados por una molécula
aceptara de electrones final. En esos procesos la energía liberada permite la formación de moléculas de ATP. }
El piruvato que se obtiene al final de la glucólisis se encuentra en un cruce metabólico en el que puede seguir dos
destinos, dependiendo de la disponibilidad de oxígeno y el tipo de célula: la vía anaerobia de las fermentaciones
(ausencia de oxigeno) o la vía aerobia de la respiración celular (presencia de oxigeno).
El piruvato formado en la glucólisis no siempre sigue la vía de la
respiración celular; en determinadas circunstancias puede pasar
a la vía alternativa de las fermentaciones.
Las fermentaciones genuinas son procesos anaerobios,
realizados por microorganismos que no toleran el oxígeno o por
ciertas células animales o vegetales cuando no disponen de
suficiente oxígeno. Son poco rentables desde el punto de vista
energético, ya que la oxidación de la materia orgánica es
incompleta y se forma mucho menos ATP que en la respiración
celular aerobia. En general, únicamente 2 ATP por cada
molécula de glucosa.
Dependiendo el producto final, se diferencian varios tipos de
fermentaciones. Las más importantes son: la fermentación
alcohólica y la fermentación láctica.
a) Vía anaerobia de las fermentaciones
Fermentación alcohólica: En la fermentación alcohólica o
etílica, el piruvato se transforma en etanol y se desprende
CO2.
Fermentación láctica: En ella el piruvato se transforma en
lactato. La realizan diversas bacterias (Lactobacillus...)

b) Vía aerobia de la respiración celular
La vía aerobia consiste en dos grupos de reacciones: el ciclo de Krebs, al que se incorpora una
molécula de acetil-CoA proveniente del piruvato de la glucólisis; y la cadena de transporte de
electrones, que va a servir para sintetizar ATP en un proceso de fosforilación oxidativa.
Antes de comenzar la vía aerobia el piruvato resultante de la glucólisis se tiene que transformar en
acetil-CoA mediante una descarboxilación oxidativa
Descarboxilación oxidativa del
piruvato
• El piruvato pasa al interior de la mitocondria, donde, mediante una reacción irreversible, seune a un coenzima y sufre una descarboxilación (pérdida de CO2) y una oxidación,formándose: CO2, NADH y acetil-CoA.
• El NADH liberará su poder reductor en la cadena respiratoria, mientras que el acetil-CoAentrará en el ciclo de Krebs.
El ciclo de Krebs
•Se liberan 2 moléculas de CO2, que se puede considerar que proceden del grupo acetilo del acetil-CoA, con lo cual seconsigue una oxidación completa de la materia orgánica. El CO2 será, por tanto, el producto residual último de la respiraciónaerobia.
•Los electrones más los protones (hidrógeno), que pierde el grupo acetilo al oxidarse, van a parar, en cuatro fases, atransportadores de electrones, formándose 3 moléculas de NADH + H+ y una molécula de FADH2. El poder reductor de estasmoléculas será transferido luego a través de la cadena respiratoria mitocondrial.
•En un paso del ciclo tiene lugar una fosforilación a nivel de sustrato que origina una molécula de GTP (equivalente desde elpunto de vista energético a un ATP).
La cadena respiratoria
•Los transportadores de electrones NADH y FADH2, originados fundamentalmente en el ciclo de Krebs,pero también en otros procesos catabólicos, albergan el poder reductor que les confieren los electrones“energéticos” que transportan. Esa energía será liberada, poco a poco, a lo largo de la cadenarespiratoria que tiene lugar en las crestas y en la membrana mitocondrial interna.
•Con fines prácticos, aunque no es del todo exacto, se considera que una molécula de NADH permite laformación de 3 moléculas de ATP, mientras que una de FADH2 sólo aportará 2 ATP.

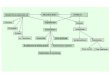
Glucosa-6-fosfato
3-Hidroxi-3metil-glutaril-CoA(HMG-CoA)
Oxalacetato
Colesterol Cuerposcetónicos
PIRUVATO
Lactato
Alanina
ACETIL-CoA
CO2
Acidos grasos
CicloKrebs
Biosíntesis
Degradación
C.K. CO2
CO2
Aminoácidos cetogénicos
Otros monosacáridos

Mecanismos de regulación de glicolisis
La regulación de este proceso está dirigida por tres enzimas: la fosfofructoquinasa, la fructosa 2,6-
Bisfosfato y la hexoquinasa las cuales manifestarán un comportamiento u otro en función de unos
factores específicos como pueden ser el pH o la concentración de una determinada sustancia.
A) La fosfofructoquinasa.
Su actividad está regulada principalmente por el nivel de energía: cuando se
observan niveles altos de ATP, el enzima se inhibe alostéricamente
disminuyendo la afinidad del enzima por la fructosa 6-Fosfato. La relación
inhibidora del ATP se contrarresta por el AMP, de manera que cuanto menor
sea la relación ATP/AMP, mayor será la actividad del enzima.
La fosfofructoquinasa también se ve afectada por el pH, esto es para evitar la
acumulación excesiva de lactato y la caída brusca del pH sanguíneo
provocando acidosis.

B) Regulación de la Fructosa 2,6-Bisfosfato
La concentración de fructosa 2,6-Bisfosfato está regulada por dos enzimas: la
fosfofructoquinasa2 (PKF2) que fosforila la fructosa 6-Fosfato y la fructosa
bisfosfatasa 2 (FBPasa2) que hidroliza la fructosa 2,6-Bisfosfato en fructosa 6-
Fosfato. Pero esto no es del todo correcto; ya que la PKF2 y la FBPasa2 son en
realidad la misma enzima. Forma parte de la misma cadena polipeptídica. Es una
enzima bifuncional con tres dominios: uno regulador en la región N-terminal, un
dominio quinasa y un dominio fosfatasa.
Las dos actividades de la enzima están reguladas por la fosforilación de un único
residuo de serina. Cuando la glucosa es baja en sangre, aumenta los niveles de
la hormona glucagón que favorece la fosforilación de la enzima inhibiendo la
actividad PFK2, lo que hace descender el nivel de F-2,6-BP. Y al revés, si la
glucosa es alta en sangre, la enzima pierde el fosfato unido activando la PFK2.

La hexoquinasa cataliza la primera etapa de la glicólisis y se inhibe con
su producto: la glucosa 6-fosfato. Altas concentraciones de esta indican
que la célula no precisa de más glucosa.
En el hígado se encuentra una isozima especializada de la
hexoquinasa: la glucoquinasa, la cual no se inhibe por la glucosa 6-
fosfato. Pero esta isozima de la hexoquinasa sólo es activa a altas
concentraciones de glucosa (tiene una afinidad 50 veces menor que la
hexoquinasa). Su función es suministrar glucosa 6-fosfato para la
síntesis de glucógeno como medio de almacenamiento de la glucosa.
C) Hexoquinasa y Glucoquinasa.