Embed Size (px)
Citation preview

Keratinocyte Growth Factor 1 Inhibits Wound Edge EpithelialCell Apoptosis In Vitro
James D. Firth and Edward E. PutninsDepartment of Oral Biological and Medical Sciences, University of British Columbia, Vancouver, British Columbia, Canada
The ability of keratinocyte growth factor 1 to modulate apoptosis in the absence of proliferation was studied
in vitro. A HaCaT scrape wound model was developed in which dense monolayers prior to wounding were cultured
to quiescence in defined media with hydroxyurea at concentrations that blocked proliferation without loss of cell
viability. Scrape wounding was then found to induce apoptosis, originating at the wound edge, but subsequently
radiating away over a 24 h period to encompass areas not originally damaged. Keratinocyte growth factor 1
inhibited this radial progression of apoptosis in a concentration-dependent manner up to 20 ng per mL with
induced migration present at the wound edge. The extent of this rescue was modulated by the concentration of
Ca2þ prior to wounding. In control wound cultures apoptotic bodies were found in cells adjacent to the wound
interface but were greatly reduced in keratinocyte-growth-factor-1-treated groups. Keratinocyte growth factor 1
receptor expression was significantly induced within two to three cell widths of the scraped wound edge, at levels
far exceeding those found at the leading edge of a nonwounded epithelial sheet. Tumor necrosis factor a (1–5 ng
per mL) or Escherichia coli lipopolysaccharide (10–50 ng per mL) exacerbated scrape-induced early apoptosis (1–4
h), but was largely ameliorated by coculture with keratinocyte growth factor 1. Keratinocyte growth factor 1
protection was associated with a reduction in both caspase-3 activation and cytokeratin-19 loss. Protected wound
edges were also associated with the maintenance of e-cadherin expression and induction of b1 integrin and actin
stress fiber organization. These results suggest that keratinocyte growth factor 1 may play a role in limiting
mechanically induced apoptotic processes at the epithelial wound edge in a manner that is distinct from its
proliferative function.
Key words: apoptosis/epithelia/keratinocyte growth factor 1/wound.J Invest Dermatol 122:222 –231, 2004
Apoptosis is programmed cell death that occurs duringtissue development, maintenance of homeostasis, in res-ponse to infection and during wound healing. During woundhealing the controlled elimination of damaged tissue pro-ceeds and facilitates the reparative processes of migra-tion, proliferation, and differentiation (Mohan et al, 2000).Apoptosis can be caused at the wound edge as a re-sult of direct mechanical trauma such as occurs in venousgrafts (Kalra and Miller, 2000), spinal cord injury (Yong et al,1998; Beattie et al, 2000), and corneal keratectomy (Wilson,2000). After corneal epithelial injury apoptosis eliminatesadjacent underlying keratocytes, which are subsequentlyreplenished within 3–4 d through proliferation and migrationof residual keratocytes (Wilson et al, 1996; Wilson and Kim,1998). Changes associated with cell survival includingapoptotic staining (Citron et al, 1997), cytokine receptorexpression (Ellis et al, 2001), and intracellular signalingevents (Dieckgraefe et al, 1997; Goke et al, 1998) arespecifically modulated near the edge of experimental
incisional wounds. Cytokines are key mediators in control-ling the onset and extent of apoptosis. Several such as Fasligand, interleukin-1a/b (IL-1a/b), and tumor necrosis factora/b (TNF-a/b) are released from injured epithelium andpenetrate into damaged anterior corneal stroma where theybind their respective keratocyte receptors (Wilson et al,1996; Helena et al, 1998; Wilson and Kim, 1998). Con-versely, epithelial growth factor (EGF) accelerates gastricepithelial healing due to the attenuation of apoptosis(Konturek et al, 2001). There appears to be considerableredundancy, however, in the expression of proapoptoticcytokines produced by the corneal epithelium. For exampleFas or Fas ligand null mice showed a significant decrease inkeratinocyte apoptosis after corneal epithelial wounding,but the overall response remains robust (Mohan et al, 1997).This suggests the possible involvement of other as yet un-identified cytokines in controlling proapoptotic and anti-apoptotic mechanisms.
Several reports have suggested that the cytokinekeratinocyte growth factor 1 (KGF-1) modulates cell survivalin several tissue types. Transient alveolar epithelial cellhyperplasia induced by KGF-1 pulse treatment in mice islater resolved in the absence of KGF-1 by apoptosis(Fehrenbach et al, 2000). Conversely, apoptosis inducedby vitamin D3 analog treatment of a benign prostate
Abbreviations: HB-EGF, heparin-binding epidermal-growth-factor-like growth factor; KGF-1, keratinocyte growth factor1; KGFr, KGF receptor; LPS, Escherichia coli lipopolysac-charide; MAPK, mitogen-activated protein kinase.
Copyright r 2003 by The Society for Investigative Dermatology, Inc.
222

hyperplasia cell line was partially prevented by administra-tion of KGF-1. KGF-1 protection was associated with itsinduction of proliferation (Crescioli et al, 2002). KGF-1attenuated H2O2-induced DNA damage in a human alveolarepithelial cell culture, however, by a mechanism thatprobably did not include increased proliferation, as thepercentage of cells in S phase did not change in the treatedgroup (Wu et al, 1998). Similarly, alveolar damage inducedby oxygen exposure in mice was prevented by intravenousKGF-1 treatment. Furthermore, during oxygen administra-tion proliferation was markedly reduced and KGF-1 wasunable to return it to normal levels, suggesting that KGF-1protective function may be independent of its proliferativeeffect. This study was unable to demonstrate a KGF-1reduction in the extent of internucleosomal degradation,however, so the antiapoptotic mechanism remained un-certain (Barazzone et al, 1999). When KGF-1 was given torats prior to isolation of alveolar epithelial cells into in vitroculture, enhanced repair of monolayer scratch woundsresulted. This process was shown not to include prolifera-tion, but rather KGF-1-induced migration of existing cells tofill the wound void (Atabai et al, 2002). Similarly, althoughKGF-1 does not affect endometrial proliferation or menstru-al sloughing in rhesus macaques it was found to inhibitapoptosis in the basalis zone (Slayden et al, 2000). KGF-1has also been proposed as a survival factor in skin. It wasfound to be associated with a less terminally differentiatedmorphology, including reduced envelope cross-linking,membrane transglutaminase activity, and internucleosomalDNA fragmentation. As KGF-1 sustains the proliferation ofhuman keratinocytes after cultures have achieved con-fluence, however, it was not apparent whether KGF-1antiapoptotic function in skin was distinct from its pro-liferative effects (Hines and Allen-Hoffmann, 1996).
The interrelationship and control of the multiple functionsof KGF-1, including proliferation, migration, and antiapop-totic signaling, are yet to be elucidated. Also, the role ofKGF-1 in regulating mechanically induced apoptosis in skinhas not been extensively studied, nor has a clear distinctionbeen made between its proliferative effect at the woundedge and other cellular effects. In this study we developed anovel in vitro model for studying the effect of KGF-1 inmediating the epithelial wound edge apoptotic processes.Specifically, KGF-1-induced proliferation was inhibited inorder that other effects might be discerned and studiedindependently. Previously, hydroxyurea has been used incell culture to inhibit proliferation while not itself inducingapoptosis over the experimental period or interfering withother cell behaviors at the wound edge, such as migration(Pukac et al, 1998; Providence et al, 2000). Here, cultureconditions were developed using HaCaT epithelial cells indefined media (Tsuboi et al, 1993), modified and supple-mented with hydroxyurea at concentrations that inhibitedKGF-1-induced proliferation but retained cell viability overthe experimental time period. Scratch wounds were sub-sequently created in the quiescent monolayers to mechani-cally induce apoptosis. The ability of KGF-1 to negateapoptosis was then examined and correlated to KGFreceptor (KGFr) expression at the wound edge. Concurrenteffects of the inflammatory mediator TNF-a and bacterialvirulence factor Escherichia coli lipopolysaccharide (LPS)
were also studied. The results suggest that KGF-1 protectswound edge epithelial cells from apoptosis independent ofits proliferative effects.
Results
Inhibition of KGF-1 induced proliferation A cell culturemodel was developed to study the role of KGF-1 ininhibiting wound edge apoptosis independent of its pro-liferative effects. A defined medium (Tsuboi et al, 1993) wasmodified (KBMþ ) and used to test the proliferativeresponse of HaCaT epithelial cells to KGF-1 (20 ng permL), compared to fully supplemented commercially pre-pared growth medium (KGM). KGF-1 induced the strongestproliferative response at 48 h over the 0 h (horizontalreference line) cell number (Fig 1a). Pretreatment (24 h) ofcultures with increasing concentrations of hydroxyurea (0–8mM) inhibited proliferation in a concentration-dependentmanner until no significant increase in cell number wasevident between 4 and 8 mM (po0.05). Increasing KGF-1concentrations up to 40 ng per mL did not reverse theinhibitory effect of hydroxyurea (data not shown). In notreatment group did cell number fall below that of the 0 htime point. In addition, visual inspection revealed normalmorphology and no anoikis. Relative to proliferating cells,(Fig 1b) inhibtion of proliferation at 4.0 mM hydroxyurea wasconfirmed (Fig 1c) by loss of proliferating cell nuclearantigen immunostaining. No differences in apoptotic bodieswere detected between the proliferating (Fig 1d) orhydroxyurea quiesced groups (Fig 1e). Subsequently, allexperiments were carried out in KBMþ , with up to 4 mMhydroxyurea, with or without KGF-1 (20 ng per mL).
KGF-1 protection of mechanically induced wounds Con-fluent monolayers cultured to quiescence in hydroxyurea(0–4.0 mM) were scratched, washed, and then furthercultured with the same medium with or without KGF-1 for24 h (Fig 2a). With increasing hydroxyurea concentrationsareas of lateral scraping were found to exhibit a zone ofapparent cell death characterized by a flattened cellmorphology (Shao et al, 1998) and reduced stainingintensity (arrow 1). KGF-1-treated cultures were found tobe partially protected from scrape-induced cell death (arrow2). Experiments were also performed with cultures pulsedwith elevated Ca2þ (0.3 mM, 24 h) prior to scratch wound-ing (Fig 2b) as previous reports had shown this to induceKGFr expression in keratinocytes (Capone et al, 2000).Onset of mechanically induced cell death began at higherhydroxyurea concentration (arrow 3). Moreover, KGF-1-in-duced protection was found to be almost complete (arrow4). Areas of scrape-induced G1 arrest morphology (arrows)with cells tightly apposed to the substrate were evidentradiating away from the wound edge at 18 h, whereas KGF-1-treated samples showed apparently normal morphologyalong the wound edge, with some migration (Fig 3a).Samples stained with Hoescht 33258 revealed the presenceof distinct late apoptotic body associated DNA condensa-tion most intensely along the control scrape wound edge(Fig 3b). KGF-1-treated samples showed a reduction andeven elimination of nuclear condensation. Control scratch
KGF-1 INHIBITS EPITHELIAL APOPTOSIS 223122 : 1 JANUARY 2004

samples immunostained for KGFr showed strong focal la-beling within 1–3 cell widths of the wound edge at 18 h(Fig 3c), whereas none was detectable at 0 h (data notshown). Also, after 18 h of KGF-1 stimulation reducedreceptor labeling, consistent with internalization wasidentified (Fig 3c). Non-scraped advancing monolayerswere also stained for KGF-1 receptor levels and foundlikewise to show elevated expression at the edge of the
advancing sheet, albeit at levels lower than those found inthe wounded cultures. As in scrape-wounded cultures,KGF-1 treatment decreased receptor labeling (Fig 3d).
KGF-1 inhibition of early and late apoptosis A two-stainsystem was used to distinguish increased cell membrane(early apoptosis) and nuclear (late apoptosis) permeability(Fig 4). Monolayers (4.0 mM hydroxyurea) were scratchwounded across the entire surface on a 1 mm grid aspreviously described (Ellis et al, 2001) to generate a uniformwound (Fig 4a). Harvested cells were analyzed by flowcytometry and dot blots (FSC vs SSC) used to define intactviable (R1) populations versus cellular debris (R2) (Fig 4b–d). R2 was consistently below 3.9% of total cell count in alltest groups. R1 populations were further analyzed forstaining intensity (Fig 4e–g). Autofluorescent (nonapoptotic)cells from nonscratch cultures were used to define aquadrant for statistical analysis (Fig 4e). Relative to this,46.46% of R1 cells from scratch wound cultures shifted intoearly (FL1, green) and 3.67% into late (FL2, red) apoptosisby 8 h (Fig 4f). KGF-1 treatment significantly reduced theearly (26.01%) and almost eliminated the late (0.41%)apoptotic shift (Fig 4g). Time course analysis showed thatKGF-1 reduction in the loss of viable cells was concentra-tion-dependent between 5–20 ng per ml over the test period(Fig 5a) with a corresponding decrease in the shift to earlyapoptosis (Fig 5b) while late apoptosis groups wereunchanged within 4 h (Fig 5c). Control wound culturesshowed a reduction of viable cells within 1 h which wasexacerbated by coculture with either bacterial virulencefactor LPS or the human inflammatory mediator TNF-a (Fig6a). KGF-1 at 20 ng per ml fully rescued 1 ng per ml TNF-abut was less effective at the higher TNF-a concentration(Fig 6b). KGF-1 was also relatively effective in rescuing10 ng per ml LPS exacerbated apoptosis with little rescue atthe higher concentration (Fig 6c).
Figure 1Hydroxyurea inhibits KGF-1-induced epithelial keratinocyte prolif-eration. Subconfluent HaCaT epithelial cells were cultured to quies-cence for 24 h in defined basal medium (KBMþ ) and variousconcentrations of hydroxyurea (0–8.0 mM). Samples were washedand further cultured for 48 h at the same hydroxyurea concentrations ineither KBMþ , KBMþ with 20 ng per mL KGF-1 (KGF-1), or commercialgrowth medium (KGM). Groups were colorimetrically assayed for cellnumber (mean � SD; n¼4) at 0 h (horizontal reference line) or after48 h. Asterisk indicates no significant difference between 0 h and 48 hKGF-1-treated samples (po0.05). Separate groups were stained withproliferating cell nuclear antigen either (b) before quiescence, or (c) after48 h quiescence in KBM+/4.0 mM hydroxyurea. Other cultures werestained with Hoescht 33258 to detect apoptotic bodies (d ) beforequiescence, or (e) after 48 h quiescence in KBM+/4.0 mM hydroxyurea.Representative of three independent experiments.
Figure2KGF-1 inhibits scrape-wound-induced cell death. Confluent mono-layers of HaCaT cells were cultured to quiescence in defined medium(KBMþ ) with increasing concentrations of hydroxyurea (0–4.0 mM) and0.1 mM Ca2þ for 24 h (a). Some samples were further pulsed at 0.3 mMCa2þ for an additional 2 h (b). Samples were then scraped, washed,and further cultured for 24 h in fresh KBMþ with the same hydroxyureaconcentrations all at 0.1 mM Ca2þ , with or without KGF-1 (20 ng permL). Samples were fixed and stained with crystal violet and thenanalyzed for areas of reduced staining indicating cell death (arrows).Representative of three independent experiments.
224 FIRTH AND PUTNINS THE JOURNAL OF INVESTIGATIVE DERMATOLOGY

KGF-1 antiapoptotic signal transduction Grid-scratchedsamples were subjected to western analysis to detectinactive (32 kDa) and active (19 kDa) levels of caspase-3, aterminal effector in apoptosis. In control scratch cultures(Fig 7a) activation of caspase-3 was discernible at 4 h andincreased over 24 h. In KGF-1-treated cultures (Fig 7b),active caspase-3 was minimal until 16 h and at 24 h was stillclearly less than controls. Degradation of intermediatefilaments, a late apoptotic event associated with caspase-3 activity (Dohmoto et al, 2001), was also tested (Fig 7c).Permeabilized cell suspensions from grid scratch cultureswere stained with antibodies to cytokeratin-8, cytokeratin-
18, and cytokeratin-19 and analyzed by flow cytometry.Cytokeratin-19 was found to be highly expressed inunscratched cultures (0 h) and rapidly reduced (69.62%)
Figure 3KGF-1 antiapoptotic effect acts specifically at the scrape woundedge. Nonproliferating, quiescent HaCaT epithelial monolayers (KBMþ ,4.0 mM hydroxyurea) were scrape wounded (a–c), and then cultured foran additional 18 h in the same medium (Control) or supplemented withKGF-1 (20 ng per mL). A nonwounded, subconfluent, advancingepithelial margin was included in the treatment groups (d). Sampleswere fixed and stained with (a) crystal violet to detect cell deathradiating from the wound edge (arrows), (b) Hoescht 33258 to detectapoptotic bodies, or (c), (d) anti-KGF-1 receptor antibody. Representa-tive of 10 fields from three independent experiments. Figure4
KGF-1 inhibits early and late apoptotic events in mechanicallyinduced wounds. Nonproliferating, quiescent HaCaT epithelial mono-layers (KBMþ , 4.0 mM hydroxyurea) were scrape wounded across theentire surface on a 1 mm grid (a). Samples were washed and furthercultured for 8 h in the same medium with or without KGF-1 (20 ng permL). Harvested cells were processed with a two-color apoptosis assayand flow cytometry (b–d) to detect intact cells (R1), which were furtheranalyzed with a two-color apoptosis assay and flow cytometry (e–g).Viable (autofluorescent) cell populations that did not stain were used todefine a quadrant to classify shifts in staining to early apoptotic (FL1,green) or late apoptotic (FL2, red); (b), (e) unscratched controls; (c), (f)scratched; (d), (g) scratched and KGF-1 treated. Representative ofthree independent experiments.
KGF-1 INHIBITS EPITHELIAL APOPTOSIS 225122 : 1 JANUARY 2004

on control grid-scratched samples at 24 h. Thereafter asmall recovery in levels was found. KGF-1 reduced the rateand total amount of cytokeratin-19 loss (50.60%) up to 24 h.By 48 h the KGF-1 samples showed a 26.14% induction ofcytokeratin-19 above the 0 h controls. Loss of cell�cellcontact associated with radial transmission of the apoptoticsignal was investigated by assaying for e-cadherin (Kuwa-
hara et al, 2001). The role of mechanical signal transductionin apoptosis was assayed using b1 integrin and b actin(Kainulainen et al, 2002). Immunostained control scrapewound samples showed a loss of all three molecules
Figure 5KGF-1 rescue of mechanically-induced apoptosis is concentra-tion-dependent. Nonproliferating, quiescent HaCaT epithelial mono-layers (KBMþ , 4.0 mM hydroxyurea) were scrape wounded across theentire surface on a 1 mm grid. Samples were washed, then furthercultured with fresh quiescence media alone for 1 to 4 h (Control), orsupplemented with various concentrations of KGF-1 (5–20 ng). Har-vested cell suspension were processed with a 2 color apoptosis assayand flow cytometry then categorized by quandrant analysis as either a)viable, b) early apoptotic, or (c) late apoptotic. Mean � sd; n¼4.
Figure6KGF-1 rescues TNF-a and LPS effects at mechanically inducedwounds. Nonproliferating, quiescent HaCaT epithelial monolayers(KBMþ , 4.0 mM hydroxyurea) were scrape wounded across the entiresurface on a 1 mm grid. Sample groups were washed and then furthercultured with fresh quiescent medium for 1–4 h: (a) fresh quiescencemedia alone or with TNF-a (5 ng per ml) or LPS (50 ng per ml); (b) KGF-1(20 ng per ml) plus or minus TNF-a (1 or 5 ng per ml), or (c) KGF-1 (20 ngper ml) plus or minus LPS (10 or 50 ng per ml). Harvested cellsuspensions were processed with two-color apoptosis assay and flowcytometry. Increase in viable cell number was quantified as a loss ofgreen fluorescence (FL1) associated with early apoptosis and analyzedas percentage rescue (mean � SD; n¼4).
226 FIRTH AND PUTNINS THE JOURNAL OF INVESTIGATIVE DERMATOLOGY
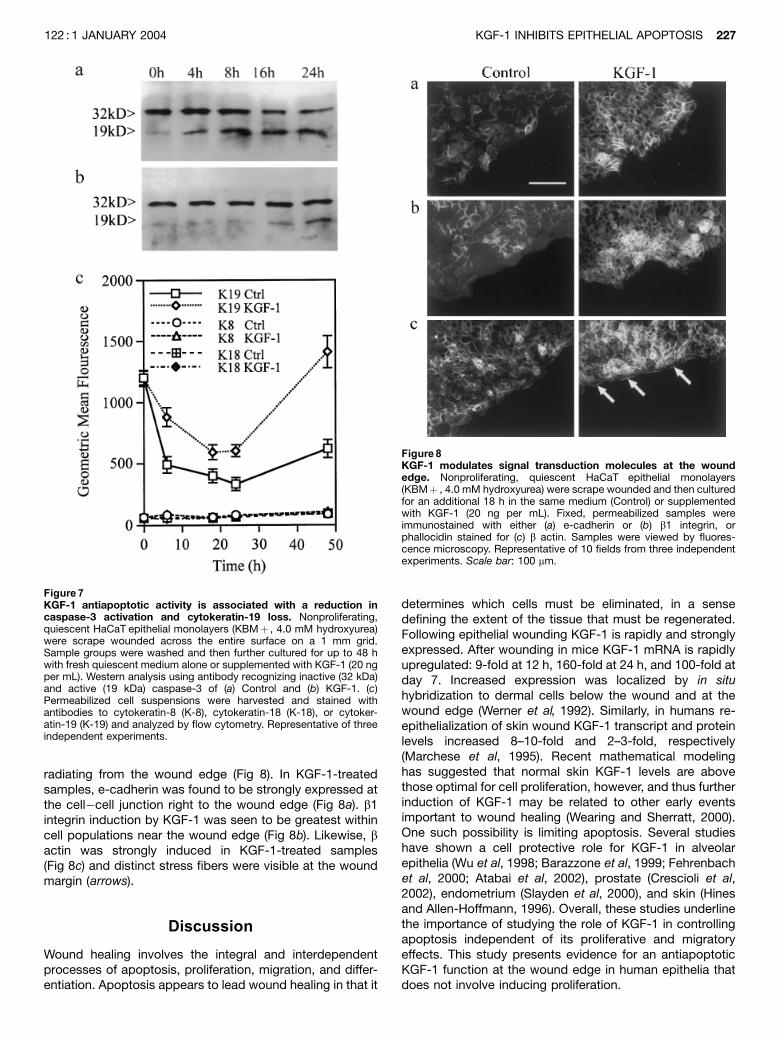
radiating from the wound edge (Fig 8). In KGF-1-treatedsamples, e-cadherin was found to be strongly expressed atthe cell�cell junction right to the wound edge (Fig 8a). b1integrin induction by KGF-1 was seen to be greatest withincell populations near the wound edge (Fig 8b). Likewise, bactin was strongly induced in KGF-1-treated samples(Fig 8c) and distinct stress fibers were visible at the woundmargin (arrows).
Discussion
Wound healing involves the integral and interdependentprocesses of apoptosis, proliferation, migration, and differ-entiation. Apoptosis appears to lead wound healing in that it
determines which cells must be eliminated, in a sensedefining the extent of the tissue that must be regenerated.Following epithelial wounding KGF-1 is rapidly and stronglyexpressed. After wounding in mice KGF-1 mRNA is rapidlyupregulated: 9-fold at 12 h, 160-fold at 24 h, and 100-fold atday 7. Increased expression was localized by in situhybridization to dermal cells below the wound and at thewound edge (Werner et al, 1992). Similarly, in humans re-epithelialization of skin wound KGF-1 transcript and proteinlevels increased 8–10-fold and 2–3-fold, respectively(Marchese et al, 1995). Recent mathematical modelinghas suggested that normal skin KGF-1 levels are abovethose optimal for cell proliferation, however, and thus furtherinduction of KGF-1 may be related to other early eventsimportant to wound healing (Wearing and Sherratt, 2000).One such possibility is limiting apoptosis. Several studieshave shown a cell protective role for KGF-1 in alveolarepithelia (Wu et al, 1998; Barazzone et al, 1999; Fehrenbachet al, 2000; Atabai et al, 2002), prostate (Crescioli et al,2002), endometrium (Slayden et al, 2000), and skin (Hinesand Allen-Hoffmann, 1996). Overall, these studies underlinethe importance of studying the role of KGF-1 in controllingapoptosis independent of its proliferative and migratoryeffects. This study presents evidence for an antiapoptoticKGF-1 function at the wound edge in human epithelia thatdoes not involve inducing proliferation.
Figure 7KGF-1 antiapoptotic activity is associated with a reduction incaspase-3 activation and cytokeratin-19 loss. Nonproliferating,quiescent HaCaT epithelial monolayers (KBMþ , 4.0 mM hydroxyurea)were scrape wounded across the entire surface on a 1 mm grid.Sample groups were washed and then further cultured for up to 48 hwith fresh quiescent medium alone or supplemented with KGF-1 (20 ngper mL). Western analysis using antibody recognizing inactive (32 kDa)and active (19 kDa) caspase-3 of (a) Control and (b) KGF-1. (c)Permeabilized cell suspensions were harvested and stained withantibodies to cytokeratin-8 (K-8), cytokeratin-18 (K-18), or cytoker-atin-19 (K-19) and analyzed by flow cytometry. Representative of threeindependent experiments.
Figure8KGF-1 modulates signal transduction molecules at the woundedge. Nonproliferating, quiescent HaCaT epithelial monolayers(KBMþ , 4.0 mM hydroxyurea) were scrape wounded and then culturedfor an additional 18 h in the same medium (Control) or supplementedwith KGF-1 (20 ng per mL). Fixed, permeabilized samples wereimmunostained with either (a) e-cadherin or (b) b1 integrin, orphallocidin stained for (c) b actin. Samples were viewed by fluores-cence microscopy. Representative of 10 fields from three independentexperiments. Scale bar: 100 mm.
KGF-1 INHIBITS EPITHELIAL APOPTOSIS 227122 : 1 JANUARY 2004

Cells located at the wound edge appear to play aspecific role in detecting and controlling the woundresponse. Previously, it was reported that scrape-woundingrat intestinal epithelial cell monolayers caused a rapidincrease in levels of heparin-binding epidermal-growth-factor-like growth factor (HB-EGF) mRNA, with a maximalresponse at approximately 1 h. The HB-EGF transcriptswere primarily localized to within a few cell widths along thewound edge. HB-EGF mRNA expression was preceded bycell survival p42/p44 mitogen-activated protein kinase(MAPK) activation, suggesting the involvement of as yetunidentified upstream activators (Ellis et al, 2001). Themechanism of wounding seems to affect the extent of theapoptotic response at the wound edge. In experimentalwounds of mouse skin it was found that the apoptotic indexin electrocautery wounds was higher than in laser-inducedwounds, but the opposite was the case for KGFr expres-sion, which was greater in laser wounds than in electro-cautery wounds (Fukuda et al, 2002). This suggests thatspecific types of wound mechanisms are most effective atupregulating KGFr expression, and furthermore that thisexpression may confer protection against apoptosis. Thisstudy detected the expression of KGFr at increased levelswithin a few cell lengths of the scratch wound edge and atlevels greater than those observed at the edge of anunwounded advancing monolayer. Furthermore, KGF-1protection was enhanced after preincubation at elevatedCa2þ consistent with a previous study that showed that suchtreatment induced KGFr expression (Capone et al, 2000).
Once an apoptotic response has been initiated at thewound edge certain signaling mechanisms can be detectedradiating away. Previously, it was found using mouse motorneurons in culture that sister cells undergo apoptosisadjacent to those at the ‘‘epicenter’’ of the scratch injury.This was restricted only to those neurons proximal to thescratch, early after injury, suggesting that a ‘‘death signal’’or wave of apoptosis is produced by injured cells movingoutward from the injury and not by factors diffusing from thewound edge cells (Citron et al, 1997). Signaling processeshave also been detected radiating from scrape-woundedintestinal epithelial cell cultures. Specifically, localizedactivation of the ERK MAPK was detected several cell-widths from the wound border in cells not originallydamaged (Dieckgraefe et al, 1997; Goke et al, 1998).Similarly, we found that cell death originating at the scrapewound edge radiated laterally over time. This was largelyprevented by KGF-1 treatment at the time of wounding,however. The transduction of apoptotic signal modulated byKGF-1 is yet to be elucidated. One candidate for involve-ment may be the cell�cell adhesion molecule e-cadherin.Loss of its expression has been assayed in mouseexcisional and incisional wounds radiating up to 500 mmaway from the wound edge by 48 h (Kuwahara et al, 2001).Consistent with this, we found e-cadherin expression atcell�cell contacts was reduced adjacent to control woundedges but strongly expressed in KGF-1-treated samples. Aswell, cytokeratin degradation is involved in the apoptoticprocess (Dohmoto et al, 2001). Conversely, antiapoptotictyrosine phosphorylation of cytokeratin 19 is mediated viathe p38 MAPK pathway (Feng et al, 1999). This study foundthe scrape wound associated with rapid and severe loss of
cytokeratin-19, associated with caspase-3 activation, whichwas largely reduced by KGF-1 treatment.
Mechanical induction of apoptosis appears to bemediated via the cytoskeleton. Apoptosis can be inducedin vascular smooth muscle cells by mechanical stretch andis accompanied by sustained activation of c-Jun and p38MAPK as well as clustering of TNF-a receptor 1 and itsassociation with TNF-a receptor-associated factor 2 (So-toudeh et al, 2002). Similarly, cyclic stretch of alveolar type IIcells induces the onset of apoptosis (Edwards et al, 1999).During tidal breathing this cell type may be protectedagainst apoptosis by protective factors present in the lung,including KGF-1 (Chelly et al, 1999; Edwards et al, 2000).Cells exposed to high amplitude static tensile forces appliedthrough integrins undergo apoptosis. This occurs, in part,due to membrane depolarization resulting from mechani-cally distorting the cell. Thus the tensile strength of thecytoskeleton is an important modulator of a cell’s suscept-ibility to mechanically induced apoptosis (Kainulainen et al,2002). KGF-1 has been shown to induce stress fibers in ahuman prostate epithelial cell line via the p38 MAPKpathway (Mehta et al, 2001). Also, we have previouslyshown that KGF-1 induced b1 integrin clustering inassociation with organization of cytoskeletal stress fibersand increased attachment avidity to extracellular matrix innormal human epithelial keratinocytes in vitro (Putnins et al,1999). Here we show that both b1 integrin and b actin areinduced at the wound edge by KGF-1. Increased attach-ment to extracellular matrix following KGF-1 treatment hasalso been shown in alveolar epithelial cells in vitro. Thissuggests that a possible KGF-1 antiapoptotic function mayinvolve stabilizing integrin/cytoskeleton organization. Thepossible role of p38 MAPK pathway signaling is worthy offurther investigation. It has been generally characterized asproapoptotic (Xia et al, 1995; Kummer et al, 1997). Its role incytokeratin-19 protection and stress fiber induction, how-ever, may be contradictory.
LPS, a component of the outer membrane of all Gram-negative bacteria, can induce apoptosis in human epithelialcells by a mechanism that requires either mRNA or proteinsynthesis (Choi et al, 1998; Bannerman et al, 2001). Inaddition to a direct role, LPS stimulates the production ofTNF-a, a proinflammatory cytokine that can also induceapoptosis (Thijs et al, 1996). TNF-a (Sanaie et al, 2002) andLPS (Putnins et al, 2002) treatment of human gingivalfibroblasts strongly induced KGF-1 protein expression.Apoptosis is mediated by death receptors belonging tothe TNF receptor family, including Fas (CD95), TNFR1,TRAMP, TRAIL, TRAIL-R2, and DR-6 (Wehrli et al, 2000).LPS-induced apoptosis can be blocked by pretreatmentwith anti-TNF-a antibody suggesting that its action ismediated through TNF-a and its interaction with TNFR1(Yokochi et al, 1998). It has been shown that KGF-1 protectsmurine hepatocytes from LPS-induced TNF-mediatedapoptosis (Senaldi et al, 1998). Conversely, membranereceptor FAS transfection induces apoptosis that KGF-1could not rescue (Freiberg et al, 1996). The TNFR1apoptotic signal pathway is mediated through caspase-8to the effector caspase-3, which in turn cleaves multiplecellular proteins, resulting in cell death (Rath and Aggarwal,1999). This study shows that addition of either TNF-a or LPS
228 FIRTH AND PUTNINS THE JOURNAL OF INVESTIGATIVE DERMATOLOGY

to the scratch wound model exacerbated the rapid wound-induced shift to early apoptosis but that KGF-1 could largelyameliorate the early effects. Activation of nuclear factor kBhas been found to be a key mechanism by which cells arerendered resistant to TNF-a-induced apoptosis (Beg andBaltimore, 1996; Van Antwerp et al, 1996) but the role ofKGF-1 in nuclear factor kB activity is unknown. Also, themechanisms that couple p38 MAPK activity to the TNFpathway are ambiguous (Baud and Karin, 2001); thus thisconnection may also be worthy of further investigation, asthis study implicates both in KGF-1 protection againstmechanically induced apoptosis.
In conclusion, an important role for KGF-1 in woundhealing has been established through its early and largeinduction at wound sites, and its ability to induce epithelialproliferation, migration, and an antiapoptotic function. Thisstudy demonstrates a model for studying mechanicallyinduced apoptosis at the epithelial wound edge. Using thismodel KGF-1 was found to inhibit wound edge apoptosiswithout the participation of proliferation. KGF-1 antiapopto-tic protection was found to be associated with elevated b1integrin expression, stress fiber formation, a short-termreduction in cytokeratin-19 loss, which was associated withreduced caspase-3 activation, and a longer-term cytoker-atin-19 induction.
Materials and Methods
Cell culture Cells from the epidermal keratinocyte cell line HaCaT,a generous gift from Dr Hubert Fusenig, German Cancer Center,Heidelberg, Germany, were maintained in a-modified Eagle’smedium/10% fetal bovine serum (Gibco, Rockville, MD). All re-agents were purchased from Sigma (St Louis, MO) unless other-wise stated. A defined medium (Tsuboi et al, 1993) modified forexperiments (KBMþ ) included keratinocyte basal medium (KBM,Clonetics, San Diego, CA), antibiotics (100 mg per mL streptomycinsulfate, 100 U per mL penicillin), antimycotic (amphotericin B,0.25 mg per mL), 5.0 mg per mL insulin, 0.5 mM hydrocortisone, 0.1mM ethanolamine, 0.1 mM phosphoethanolamine, and 10 mg per mLfibronectin. Some experiments were performed in KGM (Clonetics)fully supplemented with the manufacturer’s growth additives.Calcium concentrations were varied between 0.1 and 0.3 mM.
Proliferation assay Cells were plated (5 � 103 per well) in 96-welltiter plates, allowed to attach overnight in KGM, and then furthercultured to quiescence in KBMþ for 8 h. Cells were then switchedto KGM or KBMþ with 0–40 ng per mL KGF-1 (UpstateBiotechnology, Lake Placid, NY) for an additional 24 h. All mediawere supplemented with varying concentrations of hydroxyurea (0–8 mm). At 24 h, cultures were assayed for viable cell number usinga tetrazolium salt assay (CellTiter 96, Promega, Madison, MI)according to the manufacturer’s instructions. Relative cell numberswere assayed colorimetrically at 0 and 48 h using an opticaldensity plate reader (Titertek, Huntsville, AL) at 570 nm. Todemonstrate statistical significance of enhanced cell proliferationor reduced apoptosis, the unpaired Student’s t test was applied.
Scrape wound Cells plated (5 � 104 per well) in 24-well cultureplates or on glass coverslips were grown to dense confluence inKGM and then further cultured to quiescence in KBMþ for 8 h.Culture surfaces were scraped once across the point of maximumdiameter with a transfer pipette, and washed with KBMþ . FreshKBMþ supplemented with various concentrations of hydroxyurea(0–4 mM) with or without KGF-1 (20 ng per mL) were added andcultures were incubated an additional 18–36 h. Cultures on plasticwere then washed with phosphate-buffered saline (PBS), fixed with
4% formaldehyde, 5% sucrose, PBS, stained with 0.5% crystalviolet in 20% methanol, and washed. Cultures on glass wereterminated in Carnoy’s fixative (glacial acetic acid:methanol, 1 : 3)and stained with Hoescht 33258/Hank’s balanced salt solution.Images were digitally recorded with either tungsten (plastic) orultraviolet (glass) illumination.
Flow cytometry Cells plated (2 � 105) in 35 cm culture disheswere grown to dense confluence in KGM and then further culturedto quiescence for 8 h in KBMþ /4.0 mM hydroxyurea. Culturesurfaces were then scratched with a fragment of a fine tooth haircomb in a 1 mm grid pattern covering the entire surface aspreviously described (Ellis et al, 2001). Samples were washed andincubated in the same medium with various concentrations ofKGF-1 (0–20 ng per mL), TNF-a (1–5 ng per mL), or LPS (10–50 ngper mL) for different periods (1–8 h). Cells were then trypsinized,incubated with trypsin neutralizing solution (Clonetics), pelleted,and resuspended in PBS. Samples were treated with VybrantApoptosis assay kit (Molecular Probes, Eugene, OR) according tothe manufacturer’s instructions. Cells (2 � 104) were assayed forearly apoptosis (green fluorescence at 488 nm (FL1)) and lateapoptosis/necrosis (red fluorescence at 546 nm (FL2)) by flowcytometry, and dot blots of FL1 versus FL2 were subjected toquadrant analysis using Cell Quest software (Becton Dickinson,Franklin Lakes, NJ). In other experiments resuspended cells werestained with anti-cytokeratin-8, anti-cytokeratin-18, or anti-cyto-keratin-19 primary antibodies (Chemicon International, Temecula,CA), washed, stained using an Alexa-488 (Molecular Probes)secondary antibody, and then subjected to flow cytometry at 488nm. Samples stained only with secondary antibody were includedto detect autofluorescence.
Immunostaining Experiments performed on glass coverslips asabove were fixed (2% paraformaldehyde, 5% sucrose, PBS),permeabilized (0.5% Triton X-100, PBS, 4 min), and then washedfive times in PBS. Cells were quenched (fresh 0.05% NaBH4, PBS)and blocked (3.0 mg per mL bovine serum albumin (BSA), 1 mg permL glycine, PBS, 30 min). Cultures were then incubated in theblocking solution with primary antibody for either KGF receptor(Bek C-17; Santa Cruz Biotechnology, Santa Cruz, CA), b1 integrin,or e-cadherin (Chemicon) or proliferating cell nuclear antibody(DAKO, Glostrup, Denmark). overnight at 41C. Samples werewashed five times (1 mg per mL BSA, PBS) and then incubated inwash solution with 1:50 dilution of RITC secondary antibody(Boehringer, Mannheim, Germany) for 1 h at room temperature inthe dark. Cultures were washed twice with PBS and then mountedin Vectashield (Vector Laboratories, Burlington, CA). In otherexperiments, fixed and blocked samples were stained withBodipy-phallocidin (Molecular Probes), then avidly washed inPBS, and mounted. Preparations were digitally recorded underlaser epifluorescence at 488 nm.
Western analysis Comb scratch experiments were performed asabove at various time points. Cultures were extracted with poly-acrylamide gel loading buffer, sonicated 3 � 1 s, and then centri-fuged for 30 min at 13,000 � g. Denatured, soluble protein (40 mg)was fractionated by 10% sodium dodecyl sulfate polyacrylamidegel electrophoresis and blotted to Immobilon-P membranes(Millipore, Bedford, MA). The filter was blocked at room tempera-ture for 1 h in 5% nonfat milk powder in PBS Tween-20 (25 mMTris�HCl, pH 8.0, 144 mM NaCl, 0.05% Tween-20). The filter waswashed three times, incubated for 2 h with 2 mg per mL primaryantibody (caspase-3, Chemicon), blocked in 0.1% BSA�PBS,washed, and incubated for 1 h at room temperature with 1:5000horseradish peroxidase secondary antibody (Amersham Pharma-cia, Little Chalfont, UK) in blocking buffer. Filters were then washedfive times PBS Tween-20. Antigen was detected using enhancedchemiluminescence kit as per the manufacturer’s instructions(Amersham Pharmacia Biotech). Relative change in signal amount
KGF-1 INHIBITS EPITHELIAL APOPTOSIS 229122 : 1 JANUARY 2004

was calculated by comparing scanned western images to Ponceaustained filters using NIH Image 1.62 (NIH, Bethesda, MD).
This project was supported by a grant from the Canadian Institutes ofHealth Research to Edward E. Putnins.
DOI: 10.1046/j.0022-202X.2003.22124.x
Manuscript received June 17, 2003; revised August 4, 2003; acceptedfor publication September 8, 2003
Address correspondence to: Dr Edward E. Putnins, Department of OralBiological and Medical Sciences, Faculty of Dentistry, University ofBritish Columbia, 2199 Wesbrook Mall, Vancouver, British Columbia,Canada V6T 1Z3; Email: [email protected]
References
Atabi K, Ishigaki M, Geiser T, Ueki I, Matthay MA, Ware LB: Keratinocyte growth
factor can enhance alveolar epithelial repair by nonmitogenic mechan-
isms. Am J Physiol Lung Cell Mol Physiol 283:L163–L169, 2002
Bannerman DD, Tupper JC, Ricketts WA, Bennett CF, Winn RK, Harlan JM: A
constitutive cytoprotective pathway protects endothelial cells from
lipopolysaccharide-induced apoptosis. J Biol Chem 276:14924–14932,
2001
Barazzone C, Donati YR, Rochat AF, Vesin C, Kan CD, Pache JC, Piguet PF:
Keratinocyte growth factor protects alveolar epithelium and endothelium
from oxygen-induced injury in mice. Am J Pathol 154:1479–1487, 1999
Baud V, Karin M: Signal transduction by tumor necrosis factor and its relatives.
Trends Cell Biol 11:372–377, 2001
Beattie MS, Farooqui AA, Bresnahan JC: Review of current evidence for
apoptosis after spinal cord injury. J Neurotrauma 17:915–925, 2000
Beg AA, Baltimore D: An essential role for NF-kB in preventing TNF-a-induced
cell death. Science 274:782–784, 1996
Capone A, Visco V, Belleudi F, et al: Up-modulation of the expression of functional
keratinocyte growth factor receptors induced by high cell density in the
human keratinocyte HaCaT cell line. Cell Growth Differ 11:607–614, 2000
Chelly N, Mouhieddine-Gueddiche OB, Barlier-Mur AM, Chailley-Heu B, Bourbon
JR: Keratinocyte growth factor enhances maturation of fetal rat lung type
II cells. Am J Respir Cell Mol Biol 20:423–432, 1999
Choi KB, Wong F, Harlan JM, Chaudhary PM, Hood L, Karsan A: Lipopolysac-
charide mediates endothelial apoptosis by a FADD-dependent pathway. J
Biol Chem 273:20185–20188, 1998
Citron BA, Zhang SX, Smirnova IV, Festoff BW: Apoptotic, injury-induced cell
death in cultured mouse murine motor neurons. Neurosci Lett 230:25–28,
1997
Crescioli C, Maggi M, Luconi M, et al: Vitamin D3 analogue inhibits keratinocyte
growth factor signaling and induces apoptosis in human prostate cancer
cells. Prostate 50:15–26, 2002
Dieckgraefe BK, Weems DM, Santoro SA, Alpers DH: ERK and p38 MAP kinase
pathways are mediators of intestinal epithelial wound-induced signal
transduction. Biochem Biophys Res Commun 233:389–394, 1997
Dohmoto K, Hojo S, Fujita J, et al: The role of caspase 3 in producing cytokeratin
19 fragment (CYFRA21-1) in human lung cancer cell lines. Int J Cancer
15:468–473, 2001
Edwards YS, Sutherland LM, Power JHT, Nicholas TE, Murray AW: Cyclic stretch
induces both apoptosis and secretion in rat alveolar type II cells. FEBS
Lett 448:127–130, 1999
Edwards YS, Sutherland LM, Murray AW: NO protects alveolar type II cells from
stretch-induced apoptosis. A novel role for macrophages in the lung. Am
J Physiol Lung Cell Mol Physiol 279:L1236–L1242, 2000
Ellis PD, Hadfield KM, Pascall JC, Brown KD: Heparin-binding epidermal-growth-
factor-like growth factor gene expression is induced by scrape-wounding
epithelial cell monolayers: Involvement of mitogen-activated protein
kinase cascades. Biochem J 354:99–106, 2001
Fehrenbach H, Kasper M, Koslowski R, Pan T, Schuh D, Muller M, Mason RJ:
Alveolar epithelial type II cell apoptosis in vivo during resolution of
keratinocyte growth factor-induced hyperplasia in the rat. Histochem Cell
Biol 114:49–61, 2000
Feng L, Zhou X, Liao J, Omary MB: Pervanadate-mediated tyrosine phosphor-
ylation of keratins 8 and 19 via a p38 mitogen-activated protein kinase-
dependent pathway. J Cell Sci 112:2081–2090, 1999
Freiberg RA, Spencer DM, Choate KA, Peng PD, Schreiber SL, Crabtree GR,
Khavari PA: Specific triggering of the Fas signal transduction pathway in
normal human keratinocytes. J Biol Chem 271:31666–31669, 1996
Fukuda Y, Ito Y, Azumi H, et al: Cell death and proliferation in Nd-YAG laser,
electrocautery, and scalpel wounds on mice skin. J Dermatol Sci 28:106–
118, 2002
Goke M, Kanai M, Lynch-Devaney K, Podolsky DK: Rapid mitogen-activated
protein kinase activation by transforming growth factor a in wounded rat
intestinal epithelial cells. Gastroenterology 114:697–705, 1998
Helena MC, Baerveldt F, Kim WJ, Wilson SE: Keratocyte apoptosis after corneal
surgery. Invest Ophthalmol Vis Sci 39:276–283, 1998
Hines MD, Allen-Hoffmann BL: Keratinocyte growth factor inhibits cross-linked
envelope formation and nucleosomal fragmentation in cultured human
keratinocytes. J Biol Chem 271:6245–6251, 1996
Kainulainen T, Pender A, D’Addario M, Feng Y, Lekic P, McCulloch CA: Cell death
and mechanoprotection by Filamin A in connective tissues after challenge
by applied tensile forces. J Biol Chem 277:21998–22009, 2002
Kalra M, Miller VM: Early remodeling of saphenous vein grafts: Proliferation,
migration and apoptosis of adventitial and medial cells occur simulta-
neously with changes in graft diameter and blood flow. J Vasc Res
37:576–584, 2000
Konturek PC, Brzozowski T, Duda A, et al: Epidermal growth factor and
prostaglandin E(2) accelerate mucosal recovery from stress-induced
gastric lesions via inhibition of apoptosis. J Physiol Paris 95:361–367,
2001
Kummer JL, Rao PK, Heidenreich KA: Apoptosis induced by withdrawal of
trophic factors is mediated by p38 mitogen-activated protein kinase. J
Biol Chem 272:20490–20494, 1997
Kuwahara M, Hatoko M, Tada H, Tanaka A: E-cadherin expression in wound
healing of mouse skin. J Cutan Pathol 28:191–194, 2001
Marchese C, Chedid M, Dirsch OR, et al: Modulation of keratinocyte growth
factor and its receptor in reepithelializing human skin. J Exp Med
182:1369–1376, 1995
Mehta PB, Robson CN, Neal DE, Leung HY: Keratinocyte growth factor activates
p38 MAPK to induce stress fibre formation in human prostate DU145 cell.
Oncogene 20:5359–5365, 2001
Mohan RR, Liang Q, Kim WJ, Helena MC, Baerveldt F, Wilson SE: Apoptosis in
the cornea: Further characterization of Fas/Fas ligand system. Exp Eye
Res 65:575–589, 1997
Mohan RR, Mohan RR, Kim WJ, Stark GR, Wilson SE: Defective keratocyte
apoptosis in response to epithelial injury in stat 1 null mice. Exp Eye Res
70:485–491, 2000
Providence KM, Kutz SM, Staiano-Coico L, Higgins PJ: PAI-1 gene expression is
regionally induced in wounded epithelial cell monolayers and required for
injury repair. J Cell Physiol 182:269–280, 2000
Pukac L, Huangpu J, Karnovsky MJ: Platelet-derived growth factor-BB, insulin-
like growth factor-I, and phorbol ester activate different signaling
pathways for stimulation of vascular smooth muscle cell migration. Exp
Cell Res 242:548–560, 1998
Putnins EE, Firth JD, Lohachitranont A, Uitto V-J, Larjava H: Keratinocyte growth
factor (KGF) promotes keratinocyte cell attachment and migration on
collagen and fibronectin. Cell Adhes Commun 7:211–221, 1999
Putnins EE, Sanaie AR, Wu Q, Firth JD: Induction of keratinocyte growth factor 1
expression by lipopolysaccharide is regulated by CD-14 and Toll-like
receptors 2 and 4. Infect Immun 70:6541–6548, 2002
Rath PC, Aggarwal BB: TNF-induced signaling in apoptosis. J Clin Immunol
19:350–364, 1999
Sanaie AR, Firth JD, Uitto V-J, Putnins EE: Keratincyte growth factor (KGF) -1 and
-2 protein and gene expression in human gingival fibroblasts. J Periodont
Res 37:66–74, 2002
Senaldi G, Shaklee CL, Simon B, Rowan CG, Lacey DL, Hartung T: Keratinocyte
growth factor protects murine hepatocytes from tumor necrosis factor-
induced apoptosis in vivo and in vitro. Hepatology 27:1584–1591, 1998
Shao N, Chai Y, Cui JQ, Wang N, Aysola K, Reddy ES, Rao VN: Induction of
apoptosis by Elk-1 and dElk-1 proteins. Oncogene 17:527–532, 1998
Slayden OD, Rubin JS, Lacey DL, Brenner RM: Effects of keratinocyte growth
factor in the endometrium of rhesus macaques during the luteal�follicular
transition. J Clin Endocrinol Metab 85:275–285, 2000
Sotoudeh M, Li Y-S, Yajima N, et al: Induction of apoptosis in vascular smooth
muscle cells by mechanical stretch. Am J Physiol Heart Circ Physiol
282:H1709–H1716, 2002
Thijs LG, Groeneveld AB, Hack CE: Multiple organ failure in septic shock. Curr
Top Microbiol Immunol 216:209–237, 1996
Tsuboi R, Sato C, Kurita Y, Ron D, Rubin JS, Ogawa H: Keratinocyte growth
factor (FGF-7) stimulaes migration and plaminogen activator activity of
normal human kertinocytes. J Invest Dermtol 101:49–53, 1993
Van Antwerp DJ, Martin S, Kafri T, Green DR, Verma IM: Suppression of TNF-a-
induced apoptosis by NF-kB. Science 274:787–789, 1996
230 FIRTH AND PUTNINS THE JOURNAL OF INVESTIGATIVE DERMATOLOGY

Wearing HJ, Sherratt JA: Keratinocyte growth factor signaling: A mathematical
model of dermal–epidermal interaction in epidermal wound healing. Math
Biosci 165:41–62, 2000
Wehrli P, Viard I, Bullani R, Tschopp J, French LE: Death receptors in cutaneous
biology and disease. J Invest Dermatol 115:141–148, 2000
Werner S, Peters KG, Longaker MT, Fuller-Pace F, Banda MJ, Williams LT: Large
induction of keratinocyte growth factor expression in the dermis during
wound healing. Proc Nat’l Acad Sci USA 89: 6896–6900, 1992
Wilson SE: Role of apoptosis in wound healing in the cornea. Cornea 19:s7–s12,
2000
Wilson SE, Kim W-J: Keratocyte apoptosis: Implications on corneal wound
healing, tissue organization, and disease. Invest Ophthalmol Vis Sci
39:220–226, 1998
Wilson SE, He YG, Weng J, Li Q, McDowall AW, Vital M, Chwang EL: Epithelial
injury induces keratocyte apoptosis: Hypothesized role for the interleukin-
1 system in the modulation of corneal tissue organization and wound
healing. Exp Eye Res 62:325–327, 1996
Wu KI, Pollack N, Panos RJ, Sporn PH, Kamp DW: Keratinocyte growth factor
promotes alveolar epithelial cell DNA repair after H2O2 exposure. Am J
Physiol 275:L732–L787, 1998
Xia Z, Dickens M, Raingeaud J, Davis RJ, Greenberg ME: Opposing effects of
ERK and JNK-p38 MAP kinases on apoptosis. Science 270:1326–1331,
1995
Yokochi T, Morikawa A, Kato Y, Sugiyama T, Koide N: Apoptotic cell death in
response to LPS. Prog Clin Biol Res 397:235–242, 1998
Yong C, Arnold PM, Zoubine MN, Citron BA, Watanabe I, Berman NE, Festoff
BW: Apoptosis in cellular compartments of rat spinal cord after severe
contusion injury. J Neurotrauma 15:459–472, 1998
KGF-1 INHIBITS EPITHELIAL APOPTOSIS 231122 : 1 JANUARY 2004





























