Embed Size (px)
Citation preview

Journal Club 20-‐June-‐2014
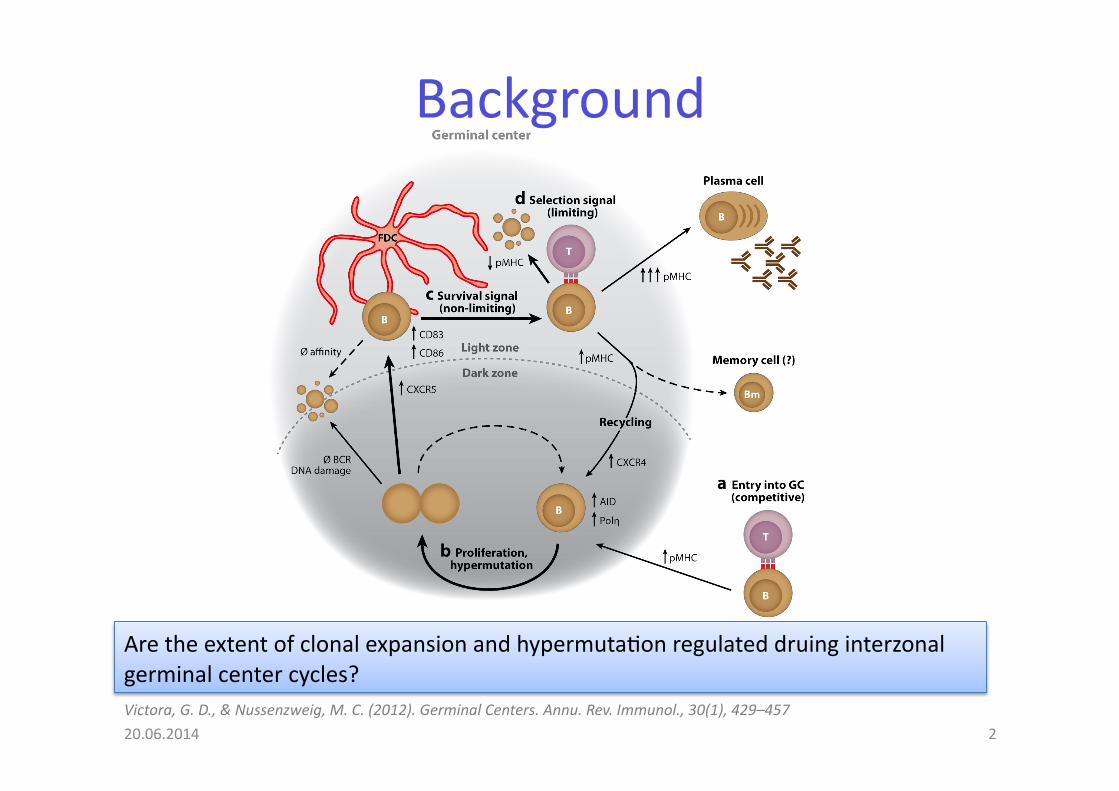
Background
Victora, G. D., & Nussenzweig, M. C. (2012). Germinal Centers. Annu. Rev. Immunol., 30(1), 429–457
Are the extent of clonal expansion and hypermuta@on regulated druing interzonal germinal center cycles?
20.06.2014 2

Does the amount of AG internalized by GC B cells steer clonal expansion?
HC specific for 4-‐hydroxy-‐3-‐nitrophenyl acetyl (NP) with Igλ LC. Transferred # 1.5-‐5x106 (about 10% NP specific)
10ug popliteal lymph node
from spleen
Ova
αDEC205-‐OVA
B1-‐8hi
DEC205+/+ (CD45.1+)
B1-‐8hi
DEC205-‐/-‐ (CD45.1+
CD45.2+)
5% 95% α DEC205-‐CS Plasmodium falciparum circumsporozite protein-‐> irrelavant AG
CS
20.06.2014 3
50ug
25 ug
10ug into footpads

AG amount and capture regulates GC B cell expansion
Fig1
10ug lymph node
from spleen
Suppl Fig1 B1-‐8hi DEC205+/+
B1-‐8hi DEC205-‐/-‐
Fig1
20.06.2014 4

AG amount and capture regulates GC B cell expansion
Fig1
GC
DZ
LZ
Increased amount of cognate AG presented by GC B cell subset to Tb cells leads to their selec@ve expansion at expense of GC B cells that present less AG.
B220+ FAS+ CD38- CD86- CXCR4+
B220+ FAS+ CD38- CD86+ CXCR4-
B220+ FAS+
20.06.2014 5

T-‐cell help regulates the number of GC B-‐cell divisions
Suppl2
20.06.2014 6
B1-8hi tTA-H2B-mCh
36h or 84h DOX 1.6mg ip and 0.2mg in hind footpad 2mg/ml in drinking water (+10mg/ml sucrose)
B220+ lymphocytes of peripheral blood

T-‐cell help regulates the number of GC B-‐cell divisions
Fig 2
20.06.2014 7
tTA-‐H2B-‐mCh can be used to monitor cell division in GC (only prime/boost, no αDEC205-‐OVA)
B1-‐8hi tTA-‐H2B-‐mCh
60h post DOX
! AG capture + presenta@on" !cell division by GC B-‐cells. Change in zonal distribu@on: aDEC-‐OVA targeted GC B cells almost exclusively in DZ.
72h αDEC205-Ova 36h DOX Control cells 2:1 in DZ αDEC205-Ova: 90% DZ
control=αDEC-CS and PBS

Higher AG capture increased S phase ini@a@on in DZ of GC B-‐cells
20.06.2014 8
Fig 3
!AG captured and presented to TFH" !propor@on of cells ini@a@ng S phase in DZ. GC B cells that express !! AG-‐> ini@ate addi@onal cell divisions in DZ before returning to LZ.
αDEC205-Ova PBS

Longer DZ residence @me during selec@ve expansion of GC B-‐cells
Fig 3
20.06.2014 9 DZ: B220+ FAS+ CD38- CD86- CXCR4+
LZ: B220+ FAS+ CD38- CD86+ CXCR4-

Increased cell division in polyclonal GCs -‐> !Ig affinity and SHM
Fig 4
20.06.2014 10
tTA-H2B-mCh NP OVA 50ug ip + 12.5ug hind foot pad
DOX
d12.5
FACS sort mChHi + mChLo GC B cells
36h
VH 186.2 family genes analysed for high affinity anti-NP W33L mutation
! rates of prolifera@on and !muta@on rate of high-‐affinity GC B cells in a polyclonal response.
Analyze intron downstream of JH4 for SHM (region is target for SHM but not subject to selection)
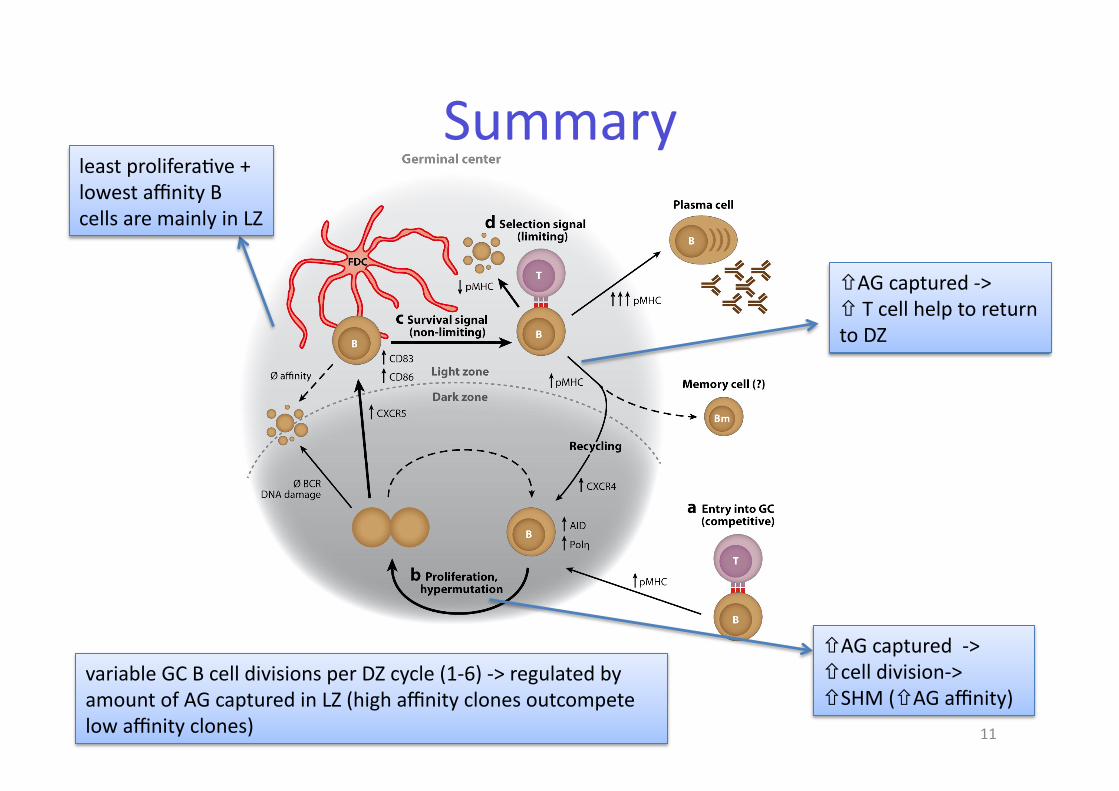
Summary
20.06.2014 11
least prolifera@ve + lowest affinity B cells are mainly in LZ
!AG captured -‐> #!cell division-‐> ! SHM (!AG affinity)
!AG captured -‐> ! T cell help to return to DZ
variable GC B cell divisions per DZ cycle (1-‐6) -‐> regulated by amount of AG captured in LZ (high affinity clones outcompete low affinity clones)

20.06.2014 12

Background
• At 20 weeks of gesta@on T cells in fetus begin to colonize the periphery (blood, secondary lymphoid organs.)
• So far the neonatal T cell compartment was supposed to only contain naive T cells (TN) and Treg for feto-‐maternal tolerance (tolerant to noninherited maternal AG -‐>NIMA).
• Fetal T cell compartmet was thought to be devoid of memory T cells TEM (adults: 50% of T cells in blood are TEM)
• The placenta harbors a nonpathogenic microbiota and is not sterile as previously thought. Aagaard, K., et al Science TranslaMonal Medicine, 6(237), 237ra65–237ra65
20.06.2014 13
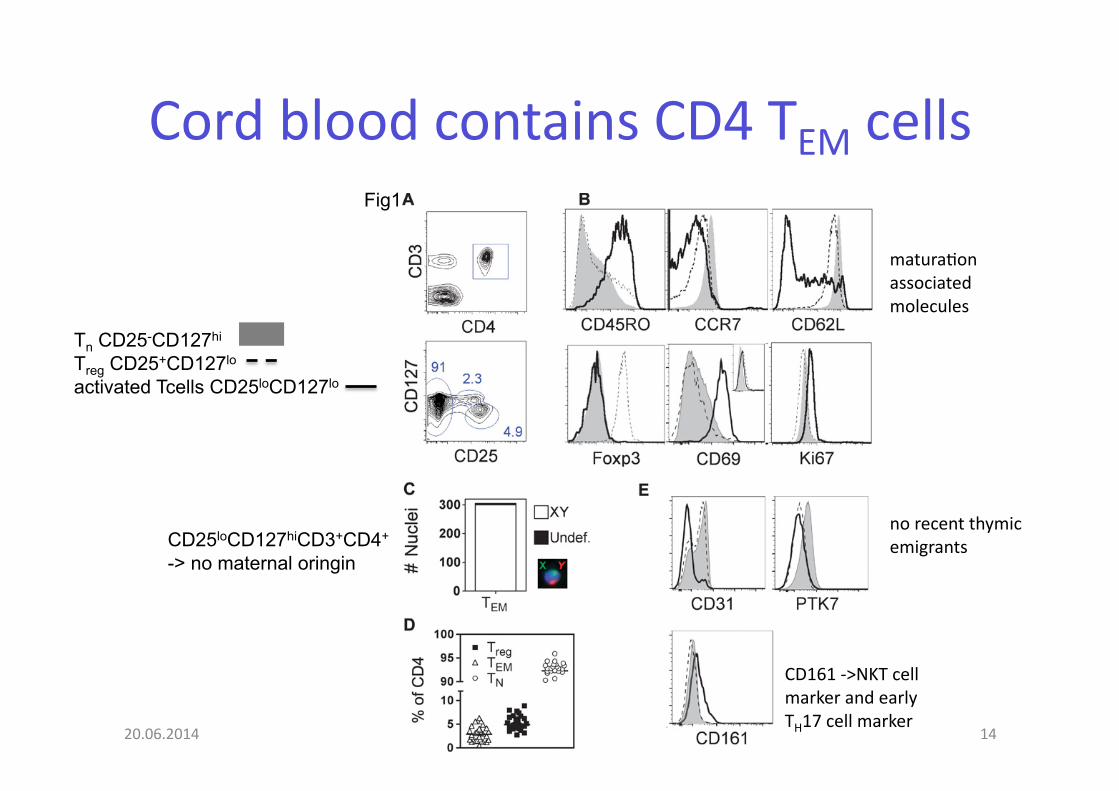
Cord blood contains CD4 TEM cells
20.06.2014 14
Fig1
Tn CD25-CD127hi
Treg CD25+CD127lo activated Tcells CD25loCD127lo
no recent thymic emigrants
CD161 -‐>NKT cell marker and early TH17 cell marker
matura@on associated molecules
CD25loCD127hiCD3+CD4+
-> no maternal oringin

Neonatal TEM are TCRαβ T cells with a polyclonal TCR repertoire
20.06.2014 15
Fig2

Neonatal TEM are TCRαβ T cells with a polyclonal TCR repertoire
20.06.2014 16
Fig2
TCRBV germline genes cluster in 24 families according to level of homology-‐> they selected 10 for analysis (ca 70% of adult T cell repertoire). Wide expression of 10 Vb genes and variable CDR3 region-‐> polylonal TCR repertoire.
TCRBVb repertoire analysis
+Immunoscope technology for CD3 length
CD3 length
fluor
esce
nce
in
tens
ity qPCR (cDNA)
Run Off (PCR product)
DNA Sequencer separated acc. to length

Neonatal TEM display TH1 and TH2 func@ons
20.06.2014 17
Fig 3 A+B FACS sorted, 8h PMA/Ionomycin
20h anti-CD3/CD28
72h anti-CD3/CD28
n=13
TEM cells can readily secrete TH1/TH2 cytokines. Under the same condi@ons this was not true for Treg and TN cells
TN
TEM
TREG
TEM-> same in adults
TEM in adults also IL-17

Chemokine receptor analysis shows a large variety of neonatal CD4 T cells
20.06.2014 18
Fig4
- CXCR3 on TH1 cells - CRTH2 onTH2 cells - CCR6 on TH17 cells - CCR4 not specific for TH2 but associated with CRTH2 in adults
9 combinatorial phenotypes of chemokine receptors in neonates. Chemokine rec. expression confirms TH1 popula@on but does not discriminate other TH func@ons.
CXCR3+ 35% CCR6+ 20% (1/3 alsoCXCR3+)

Chemokine receptor expression paoern defines molecularly different cord blood TEM subsets
20.06.2014 19
Fig 5 n=3 male donors
sequential gating strategy for different chemokine rec. Rare CRTH2 population was not included
35%
10-15%
45-50%
CD45RA+
Func@onal studies (before) show Th1 and Th2 func@ons of TEM. Microarray analysis of four TEM phenotypes, show close affilia@on of CXCR3+ TEM to TH1 and CCR6+ to TH17.
B+C) 24h anti-CD3/CD28
fold change rel. to TN
trend for TH2

Microarray analysis discriminates between different TEM subsets
20.06.2014 20
Fig 6 24h anti-CD3/CD28
Dis@nct gene clusters between TEM subgroups. CCR-‐ uplregulate several genes-‐> poten@ally s@ll capable of acquiring different phenotypes (intermediate between TN and TEM?).

Assessment of TH17 poten@al for CCR6+ TEM cells
20.06.2014 21
Fig7
fold change of CCR6+ TEM compared to TN
Receptor for IL-1 and IL-23
4-‐6d an@-‐CD3/CD28 +/-‐ IL-‐1 and IL-‐23
IL-‐22 secre@on indep. of IL-‐1 and IL-‐23-‐> indicates no associa@on with TH17 response.
Neonatal TEM cells can develop into TH17 cells.

Discussion • Iden@fica@on of memory type CD4 T cells in neonatal cord blood
(CD25loCD127hi), 1-‐3% of total CD4 T cells.
• Func@onal studies: Upon ac@va@on TH1 and TH2 like func@ons and also poten@al for TH17 when s@mulated with IL-‐1 and IL-‐23.
• Microarray: CXCR3+ TEM cells express IFNγ transcripts-‐> inflammatory TH1 cells early in life w/o infec@on at steady state.
• CCR6+ TEM cells express TH17 related genes w/o the secre@on of IL-‐17
• Neonatal TEM are highly diverse.
• Do TEM develop in response to maternal AG? -‐> no response to NIMA (non inherited maternal AG) in vitro.
• Do TEM develop in response to mild/asympoma@c infec@on or vaccina@on of mother?
• Signal for immune matura@on from commensal bacteria in placenta?
• An@gen Specificity/Self-‐reac@vity?-‐> origin and role of those cells? Role in Vaccines and infec@on?
20.06.2014 22












![RESEARCHARTICLE HigherFrequencyofNKandCD4 T-Cellsin ...bibliotecadigital.udea.edu.co/bitstream/10495/... · aslipopolysaccharides (LPS),from intestinal lumentosystemic circulation[3].Asaresult,](https://img.dokumen.tips/doc/110x75/5e2ab1d36b477e57ef2baee8/researcharticle-higherfrequencyofnkandcd4-t-cellsin-aslipopolysaccharides-lpsfrom.jpg)



![#CapCom16 : AT20 - [Étude] Communiquer autour du sport](https://img.dokumen.tips/doc/110x75/58734ddb1a28ab56378b5a31/capcom16-at20-etude-communiquer-autour-du-sport.jpg)







![Why did Europeans colonize Africa?. African Trade [15c-17c]](https://img.dokumen.tips/doc/110x75/56649ce55503460f949b2ece/why-did-europeans-colonize-africa-african-trade-15c-17c.jpg)




