Embed Size (px)
Citation preview

Journal of Theoretics
Journal Home Page
Application of BSM Atomic Models for Theoretical Analysis of Biomolecules
S. Sarg
Abstract: The proteins have amazing properties of guiding the energy along their long chains and preserving own three-dimensional structure in proper environments. These features cannot obtain satisfactory explanations by the quantum mechanical models of the atoms. A wide range interdisciplinary study, using a different theoretical approach, indicates that the quantum mechanical models do not describe the full physical features of the real atoms. Based on an alternative concept about the physical vacuum, a recently developed Basic Structures of Matter (BSM) theory allows unveiling the physical structures of the elementary particles and their spatial arrangement in the atomic nuclei. The obtained physical models of the atoms exhibit the same energy levels as the quantum mechanical models, while the positions of the electron orbits are defined by the nuclear structure. The derived atomic models allow studying the physical conditions behind the structural and bond restrictions of the atoms connected in molecules. The existing data base about the atomic structural compositions of the organic and biomolecules provides an excellent opportunity for test and validation of the derived physical models of the atoms. The presented article demonstrates a new original method for theoretical analysis of biomolecules. The analysis of DNA molecule, leads to a formulation of hypotheses about an energy storage mechanism in proteins and a possible involvement of DNA in cell cycle synchronization. The analysis of tRNA molecule leads to formulation of hypothesis about a binary decoding mechanism behind the 20 flavors of the complex of the aminoacyle-tRNA synthetases with tRNA. Keywords: Structure of biomolecules, secondary structure of proteins, molecular structure of DNA, Levinthal’s paradox, C-value paradox, twenty flavors, aminoacyl-tRNA synthetases.
Journal Home Page

Total pages: 40
1 30 Apr 2003
1. IntroductionNote: The numbering of the equations and figuresgiven in square brackets corresponds to their num-bering in the monograph1 where a detailed analy-sis and derivation is provided.
Despite the huge number of possible config-urations of the atoms in the proteins, according tothe Quantum mechanics, they fold reliably andquickly to their native state. From a point of viewof the Quantum mechanical considerations, this ef-fect, known as a Levinthal’s paradox, is not ex-plainable. According to them, a protein molecule of2,000 atoms, for example, should possess an astro-nomical number of degrees of freedom. The obser-vations show that this number is drasticallyreduced by some strong structural restrictions, suchas bond lengths, and restricted angle range of bond
connections and rotations. The stable appearanceof the secondary and tertiary structures of the pro-teins indicates also that some additional restrictionstake place in a proper environment. All this restric-tions could not get satisfactory explanation by theQuantum mechanical models of the atoms. Whilethese models rely heavily on the the uncertaintyprinciple, the quite deterministic structure and be-haviour of the complex molecules like proteins donot show its signature.
An extensive interdisciplinary study1 fromdifferent fields of physics indicates that the Quan-tum mechanical models of the atoms are rathermathematical models than physical ones, so theyare not able to provide all the features that the realatoms possess. The same study also shows that theatomic models are strongly dependable of the con-cept of the physical vacuum. This concept has been
Application of BSM atomic models for theoretical analysis of biomolecules. Hypotheses: 1. Energy storage mechanism in biomolecules; 2. DNA involve-
ment in cell cycle synchronization; 3. Binary decoding mechanism behind the twenty flavors in aminoacyl-tRNA synthetases.
Abstract The proteins have amazing properties of guiding the energy along their long chains andpreserving own three-dimensional structure in proper environments. These features cannot obtainsatisfactory explanations by the quantum mechanical models of the atoms. A wide range interdis-ciplinary study, using a different theoretical approach, indicates that the quantum mechanical mod-els do not describe the full physical features of the real atoms. Based on an alternative conceptabout the physical vacuum, a recently developed Basic Structures of Matter (BSM) theory allowsunveiling the physical structures of the elementary particles and their spatial arrangement in theatomic nuclei. The obtained physical models of the atoms exhibit the same energy levels as thequantum mechanical models, while the positions of the electron orbits are defined by the nuclearstructure. The derived atomic models allow studying the physical conditions behind the structuraland bond restrictions of the atoms connected in molecules. The existing data base about the atomicstructural compositions of the organic and biomolecules provides an excellent opportunity for testand validation of the derived physical models of the atoms. The presented article demonstrates anew original method for theoretical analysis of biomolecules. The analysis of DNA molecule,leads to a formulation of hypotheses about an energy storage mechanism in proteins and a possibleinvolvement of DNA in cell cycle synchronization. The analysis of tRNA molecule leads to for-mulation of hypothesis about a binary decoding mechanism behind the 20 flavors of the complexof the aminoacyle-tRNA synthetases with tRNA.
Keywords: Structure of biomolecules, secondary structure of proteins, molecular structure ofDNA, Levinthal’s paradox, C-value paradox, twenty flavors, aminoacyl-tRNA synthetases.

Total pages: 40
2 30 Apr 2003
changed four times in the history of the physics2,3 .The adopted by the contemporary physics conceptstill could not be considered as a final truth. The re-sults of the study led to development of an originalunified field theory called Basic Structures of Mat-ter (BSM)1. In order to build such theory, however,the concept of the vacuum had to be reconsideredonce again. The developed BSM theory from itshand allowed derivation of quite different physicalmodels of the atoms, possessing rich physicalstructures, while characterized by the same energylevel of excitation. The unveiled structural featuresof the atoms are not apparent from the quantummechanical atomic models, operating only by ener-gy levels, while assuming that the physical vacuumis an empty space, not possessing a matter.
The purpose of the initial part of this article(sections 2 to 9) is to acquaint very briefly the sci-entist from the field of molecular biology with theconcept of the BSM theory and particularly withthe derived physical models of the atoms.
The second part of the article (sections 10 to11) shows the potential application of the BSMconcept and the derived atomic models for study-ing the properties of the biomolecules. The theoret-ical analysis of some features of DNA and tRNAleads to formulation of three hypotheses.
While the BSM concept is quite original, thereader who want becoming familiar in details withits arguments outputs should be acquainted withthe article4 “Brief introduction to BSM theory andderived atomic models” (Journal of Theoretics, on-line access)) or with the theory1.
2. The new point of view of the BSM theoryExtensive analysis of phenomena from differentfields of physics, indicates that the vacuum is nota void space, but possessing a underlying gridstructure of sub-elementary particles arranged innodes. These particles called twisted prisms areformed of two types super dens intrinsic mattersubstances. Prisms of the same type are attracted ina pure void space by Intrinsic Gravitational (IG)forces, FIG, that are inverse proportional to a cubeof the distance.
[(2.1)]
where: mo1 and mo2 are intrinsic masses (of this su-perdense particles), Go is the Intrinsic Gravitation-al constant valid for pure void space.
It is assumed that the IG force is related tothe well known physical parameter called Planck’sfrequency,
(1)
This assumption is in agreement with thetheoretical work of H. E. Puthoff5 (1989) showinga derivation of the Newton’s law of gravitation,based on the Plank’s frequency and using one hy-pothesis of Sakharov.
According to BSM concept, every node ofthe underlying vacuum grid is comprised of fourprisms of same substance held by IG forces, so itsgeometry is flexible. The prisms possess axial IGanisotropy with a twisting component, due to alower level structure, so they are called twistedprisms. The Intrinsic Gravitation (IG) appears as atype of energy interaction between intrinsic matterat lower level of matter organization involved in anintrinsic energy balance. The gaps between the al-ternatively arranged nodes of different types arealso kept by an intrinsic energy balance. The esti-mated distance between neighbouring nodes of thegrid is in order of (1 ~ 2)E-20 (m), while the matterdensity of the prisms is about 1E13 times higherthan the density of the average atomic matter. It iscalled a Cosmic Lattice (CL), so the physical vac-uum in BSM theory is referenced as a CL space.
The self-sustainable CL structure is sup-ported by super strong interactions between thehighly dens intrinsic matter from which the prismsare built. The elementary particles are build by thesame prisms and comprised of spatially ordered he-lical shells called helical structures possessing alsointernal lattice formed from the same prisms. Theonly forces responsible for holding the twistedprisms in both: the CL structure and the structureof the elementary particles are the IG forces. TheCL structure of the physical vacuum penetrates notonly in all aggregate states of the atomic matter butpartly in the elementary particles as well. The New-tonian mass (the mass we are familiar with) is ex-pressible as a CL static pressure exercised on theinternal volume structure of the particle that is notpenetrative to the CL node structure. The Newto-
FIG G0mo1mo2
r3-------------------=
ωPL
ωPL2πc5
G------------=

Total pages: 40
3 30 Apr 2003
nian gravitation is a propagation of the IG fieldthrough the CL space in a distance much longerthan the CL node distance (by the self alignedprisms of the neighbouring CL nodes). At closedistances between particles, atoms and molecules,however, the IG forces leak through CL spacecausing an attraction stronger than the Newton’sgravity (Casimir forces and some of Van der Wall’sforces. Such effect is explainable when taking intoaccount that the matter density in the prisms isabout 1E13 times higher than the average atomicmatter.
A massive object such as a star, a planet ora moon, has an ability to hold the CL space struc-ture up to some extend, defining in such way theconditions of General relativity. CL space has spe-cific features of possessing distributed Zero PointEnergy and conditions for gravitational, electricaland magnetic field. Another specific features of theCL space is the ability of the CL nodes to fold andpass through the grid structure of a normal CLspace when a less massive object moves in a CLspace of a more massive one. This feature is behindthe inertial properties of the atomic matter in CLspace and the equivalence between gravitationaland inertial mass. The prisms are formed in anunique formation process existing in a hiddenphase of a galaxy evolution govern by IG forcesand fundamental matter-energy process.
Fig. 2.20 illustrates a geometry of a singlenode in position of geometrical equilibrium withtwo sets of axes, denoted as abcd and xyz.
Fig. 2.20 CL node in geometrical equilibrium positionThe two sets of axes are: abcd and xyz
The CL node has two sets of axes: one setof 4 axes along anyone of the prisms called abcdaxes, and another set of 3 orthogonal axes called
xyz axes. In geometrical equilibrium the angles be-tween anyone of abcd axes is 109.5o. The abcdaxes define a tetrahedron. The xyz axes passthrough the middle of every two opposite edges ofthe tetrahedron. In the same time, the orthogonalxyz axes of the neighbouring CL nodes are aligned.Such arrangement gives conditions for complexnode oscillations under the inverse cubic law of In-trinsic Gravitation. The return forces (of inversecubic law) acting on deviated from the central po-sition CL node exhibits set of minimums. Theseminimums can be associated with energy wells.Two symmetrical minimums appear along any-one of xyz axes and one minimum along the pos-itive direction of anyone of abcd axes. These setof minimums provide conditions for complex os-cillations of the CL node. From a dynamical aspectthese minimums contribute to the total energy wellof the CL node (Zero Point Energy of vacuum).Fig. [2.24] illustrates the return forces along thetwo sets of axes and the associated with them ener-gy wells. The right vertical axis indicates specificenergy points. The energy level EC2 corresponds tothe filled energy wells or the Zero Point Energy ofthe vacuum.
The complex oscillation of the CL node iswell described by two vectors: a Node ResonanceMomentum (NRM) and a Spatial Precession Mo-mentum (SPM). The NRM vector is related to thelight velocity expressed by one resonance cycle ofCL node per node distance. The SPM vector is re-lated to the permeability and permittivity of thevacuum. The frequency of the SPM vector is thewell known Compton frequency. Analysing the dy-namics and mutual interactions of the CL nodes(§2.9 of BSM theory), it is possible to understandsome of the fundamental physical parameters, such
Fig. [2.24]

Total pages: 40
4 30 Apr 2003
as: unite charge, magnetic field, Planck constant,Zero Point Energy, photon wavetrain structure,light velocity, permeability and permittivity of vac-uum.
From a BSM point of view, the interpreta-tion of the scattering experiments does not providecorrect real dimensions of the elementary particles,because their structures and the CL space structureof the vacuum are not taken into account. BSManalysis found that the stable particles, such asproton, neutron and electron (and positron) possesstructures with well defined spatial geometry anddenser internal lattices. They are comprised ofcomplex but understandable three-dimensionalhelical structures whose elementary buildingblocks are the same as those involved in the vacu-um grid - the two types of prisms. Analysing theinteractions between the vacuum space and ele-mentary particles, but from a new point of view, theBSM theory allowed a derivation of number of use-ful equations, such as a light velocity equation - ex-pressed by the CL space parameters, a massequation (the mass we are familiar with), an equa-tion about the vacuum energy (zero point energy),and some relations between CL space parametersand the known physical constants. The BSM pro-vides also an understandable physical explanationof what is an elementary electrical charge and whyit is constant.
The motion analysis of the smallest chargeparticle - the electron from a new point of view(Chapter 3 and 4 of BSM) allows to unveil its phys-ical structure and intrinsic properties. The electronis a system comprised of three helical structures asillustrated by Fig. 2. Two of its helical structurespossess denser lattices located in the internal spaceof the helix envelopes (not shown in Fig. 2).
The physical dimensions of this structuresare: RC - Compton radius of electron (known), re -
a small electron radius, rp a small positron radius,se - helix step.
External helical structure with internaldenser lattice (from right handed prisms, for exam-ple) is referenced in BSM as external electron shell.It is responsible for the modulation of the CL spacearound the electron that we detect as an electricalcharge. The internal helical structure with internaldenser lattice (from a left-handed prisms, respec-tively) with a central core (from right handedprisms) is an internal positron. Regarded as a 3body oscillating system, the electron has two prop-er frequencies:
- a first proper frequency: between the ex-ternal electron shell and the internal positron
- a second proper frequency: between theinternal positive shell and the central negative core.
It is found that the first proper frequency ofthe electron is equal to the frequency of the SPMvector of the oscillating CL node. The frequency ofthe SPM vector appears to be the well knownCompton frequency.
It conditions of screw-like motion of the elec-tron with tangential velocity equal to the light ve-locity, the phase of the first proper frequency of therotating electron matches the phase of the SPMvector of CL nodes. They both oscillate with aCompton frequency. In the same time, the internalcore oscillation (with a proper frequency of threetimes the Compton frequency) provides a third har-monic feature for this motion. As a result the rotat-ing and oscillating electron exhibit a maximuminteraction with the CL space - a kind of quantuminteraction. The electron axial velocity for this caseis , corresponding to kinetic energy of 13.6eV. In such type of motion the helical step of theelectron, se, is estimated by the following relations:
(m) [(3.9)]
[(3.12.a)]where: Rc - is the Compton radius, - is the finestructure constant, ge - is the gyromagnetic factor,
- is the Compton wavelength (CL space param-eter).
From the analysis of the Fractional QuantumHall experiments in Chapter 4 of BSM, it is foundthat: . Then all the of geometrical param-eters of the electron are determined.
Fig. 2
Oscillatingelectron
Vax αc=
se2πRcα
1 α2–-------------------
λcα
1 α2–------------------- 1.7706 14–×10= = =
se gere 2.002319re= =α
λc
rp/re 2/3=

Total pages: 40
5 30 Apr 2003
The confined screw like motion of the oscil-lating electron in CL space is characterized bystrong quantum interactions with the oscillating CLnodes. This effect is contributed by two conditions:a phase match between the involved cycles (dis-cussed above) and conditions of integer number ofCompton wavelengths for boundary conditions ofthe induced magnetic field from the rotatingelectron4 (more details in Chapter 3 of BSM).These two conditions allow strong quantum effectsto appear at particular velocities of screw-like mo-tion of the electron, corresponding to the energylevels of 13.6 eV, 3.4 eV, 1.51 eV, 0.85 eV and soon.
The magnetic radius rmb in a plane normalto Vax is defined from the conditions that the ro-tating IG field of the internal lattice of the elec-tron helical structure (that modulates the CLspace) could not exceed the light velocity.
The magnetic radius for 13.6 eV is verifiedfrom the analysis of the quantum magnetic field(see §3.11 in Chapter 3 of BSM thesis): The accurate value of rmb for 13.6 eV is almostequal to Rc, but slightly larger due to a finite thick-ness of the electron helical structure.
If relating the above energy levels with thenumber of full rotations of the electron one obtains:
13. 6 eV - 1 rotation per SPM cycle3.4 eV - 1/2 rotations per SPM cycle1.51 eV - 1/3 rotations per SPM cycle 0.85 eV = 1/4 rotations per SPM cycle1 SPM cycle = Compton time BSM uses a parameter called subharmonic
number, n, in order to notify the quantum motionconditions of the electron. This number is related tothe electron axial velocity by the expression
. In the same time the subharmonicnumber matches the quantum number of the elec-tron orbit in Bohr atomic model. A quantum mo-tion with a first harmonic velocity corresponds to13.6 eV; a motion with a second subharmonics -3.4 eV; a motion with a third subharmonics - 1.51eV and so on. The term subharmonic number ischosen because it annotates the spin rotation of theelectron in its confined motion. The strength of thequantum interaction with the CL space decreasewith increasing of n.
Analysing the efficiency of the quantum in-teractions between the confined moving electron
and CL space at relativistic velocities the relativis-tic gamma factor is derived in §3.11.A. (BSMChapter 3). The analysis provides a physical expla-nation of the relativistic effect of mass increase. Itis a result of the finite rate at which the CL nodecould be folded and displaced. A limiting factor ofthis rate is the resonance frequency of CL node, es-timated in §2.11.3, Chapter 2 of BSM, as:
(Hz) [(2.55)]
3. Quantum loops and possible orbits for elec-tron with optimal confined velocity. Embedded signature of the fine structure constant.
3.1 Quantum motion of the electron in closed loop trajectories.
The motion of the electron is always a resultof external forces. The orbital motion of the elec-tron could be regarded as motion in closed loop,whose trajectory follows equipotential surface ofelectrical field defined by one or more positivecharges. All this conditions are ideal for a quantummotion of the electron in a closed loop.
The analysis of the confined motion of the os-cillating electron in closed loop4, leads to the con-clusion:
(A) The number of cycles of the first andsecond proper frequency of the electron in theorbital trace is defined only by the fine structureconstant.
In the same article4 it is shown that the condi-tions of accurate phase match between the first andsecond proper frequency of the electron in its con-fined quantum motion in a closed loop is satisfiedfor 2573380 full turns of the electron in which thethe expression is valid
The CODATA 98 is
the closest value of the fine structure constant sat-isfying the above expression.
The introduced by BSM theory subharmonicnumber (n) of the moving and oscillation electroncorresponds to the principal quantum number ofBohr model.
Table 1 shows the quantum motion parame-
ters of the electron in a quantum loop for differentnumber n.
Table 1
Φ0 h/qo=
Vax αc/n=
νR 1.092646 29×10=
1/α3 integer 2573380= =α 7.2973525 27( ) 10 3–×=

Total pages: 40
6 30 Apr 2003
------------------------------------------------------------------------No E (eV) Vax Vt rmb lql Lq (A)------------------------------------------------------------------------- -------------------------------------------------------------------------where: E - is the electron energy, Vax - is the axial velocity, Vt- is the tangential velocity of the rotating electron structure,rmb - is the value of the boundary electron magnetic radius ina plane normal to Vax vector, c - is a light velocity, Rc - is theCompton radius, ao - is the Bohr radius, lql - is the trace lengthfor a motion in closed loop (single quantum loop), Lq - is thelength size of the quantum loop as Hippoped curve with pa-rameter .
The introduced parameter subharmonicnumber shows the rotational rate of the wholeelectron structure. The rotational rate decreaseswith the consecutive increase of this number, butthe number of the first and second proper frequen-cy cycles is not changed. This is very importantfeature formulated by the conclusion (B).
(B) The number of first and second properfrequency cycles of the electron in closed loopwith any subharmonic number is a constant.
From the analysis provided in “Brief intro-duction to BSM theory”4 it is evident that the elec-tron makes 18778.362 rotations for one quantumloop. One quantum orbit may contain one ore morequantum loops. From Table 1 we see that for a con-fined moving electron the circumference length ofthe boundary of the electron’s magnetic radius in aplane normal to Vax is equal to a whole number of
3.2. Quantum orbits. Emission and absorption of photon.
A stable quantum loop is defined by the re-peatable motion of oscillating electron. The shapeof such loop is defined by external conditions. Suchconditions exist in the following simple cases:
- a quantum loop obtained between particleswith equal but opposite charges and same mass, asin the case of positronium (see Chapter 3 of BSM)
- a quantum loop obtained between particlesof opposite charges and different masses, as in caseof the hydrogen atom.
In both cases the loops are repeatable and wemay call them quantum loops. One or more quan-tum loops in series form a quantum orbit. It is usu-ally a three dimensional closed curve. In case ofhydrogen, any possible quantum orbits obtain afixed position in respect to the proton.
The unveiled conditions of the electron mo-tion in a quantum loop allow definition of any pos-sible quantum orbit in the atoms and molecules. InChapter 7 of BSM, the possible quantum orbits inHydrogen are discussed and a model of Balmer se-ries is suggested. Its analysis confirms the conceptof confined motion of the electron. The trace lengthof the quantum orbit of Balmer series match thequantum loop No 2 from Table 1. The same quan-tum loop exists also in Deuteron. The BSM modelof Hydrogen is different from the Bohr model, butall types of quantum numbers are identifiable. Thequantum levels are obtainable by considering awhole number of Compton wavelengths but in aspecific conditions defined by factors, such as theproximity distributed E-field of the proton, and itsIntrinsic Gravitation field.
It appears that the limiting orbit has a lengthof and all other quantum orbits are inferior.This is valid not only for Balmer series in Hydro-gen but also for all possible quantum orbits in dif-ferent atoms, if they are able to provide linespectra. Therefore, the suggested physical modelprovides a solution of the boundary conditionsproblem of the electron orbits for the Hydrogenand for all other atomic models suggested byBSM theory.
The analysis of interactions between the os-cillating electron in confined motion and oscillat-ing CL nodes allows unveiling the process of thephoton generation. According to the BSM concept,the emission of a photon, contributing to line spec-tra, is a result of pumping of CL nodes from theorbiting and simultaneously rotating and oscillat-ing electron. In this process more than one orbitalcycles are involved. The emission of a photon oc-curs from the surrounding CL space in the mo-ment when the electron drops from higher tolower energy orbit. This explains why the Quan-tum mechanical model needs an uncertaintyprinciple, while the BSM model does not needsuch.
1 13.6 αc c ~Rc 2πa0 1.36262 3.4 αc/2 c/2 2Rc 2πa0/2 0.68133 1.51 αc/3 c/3 3Rc 2πa0/3 0.45424 0.85 αc/4 c/4 4Rc 2πa0/4 0.34065 0.544 αc/5 c/5 5Rc 2πa0/5 0.2725
a 3=
λc
2πa0

Total pages: 40
7 30 Apr 2003
In Quantum Mechanical models the CL sub-stance is missing and the process of the photon gen-eration has to be directly connected to the orbitingelectron. In such case, it seems that the electron po-sition could not be located, so a concept of “elec-tron cloud” has been introduced. In BSM model theelectron in the atom has well defined orbit, whilethe concept of CL space allow analysis of the elec-tron motion in any portion of the orbit.
3.3 Lifetime of the orbital motion of the elec-tronIn section 3.1 it was mentioned that the conditionfor phase repetition of the two proper frequenciesof the electron with velocity (13.6 eV) aremet for 2573380 electron turns (about 137 orbitalcycles ) and approximately met for three orbital cy-cles. The full travel of the rotating electron in bothcases can be expressed by the product of thenumber of turns and helical step. Taking into ac-count the relativistic gamma correction the fulltravel is . In the same time the quantumvelocity of the electron is is known. Then dividingthe full travel by the velocity we obtain the orbitallifetime.
(s) (7)
For a second harmonic quantum loops thenumber of electron turns is twice smaller, so its ve-locity and travel length are also twice smaller, sothe lifetime appears the same. Consequently theobtained equation (7) appears valid for a quan-tum orbit with any subharmonic number, n,comprised of single quantum loop as shown inTable 2. It is quite reasonable to consider this to bethe lifetime for spontaneous emission. For quan-tum orbit comprised of m number of quantum loopsthe lifetime will be m times larger. Such conditionsappear for the higher order series of Hydrogen inrespect to the Balmer series.
In case of simulated emission, the lifetime ofthe orbiting electron could be shortened. It is inter-esting to find what could be the shortest lifetime inwhich a photon generation is still possible. FromEq. [(3.43.h)] we see that this could be the lifetimecorresponding to three orbital cycles. Proceeding ina similar way as for the spontaneous emission wearrive to the result
(s) (8)
Note: In the derived equations (7) and (8) therelativistic consideration are not taken into ac-count.
Summary:• The orbital lifetime for spontaneous emission is
the time for which the first electron frequency makes 2573380 cycles
• The shorter lifetime for simulated emission is the time for which the first electron frequency make 18778.3 cycles (the secondary electron frequency makes 56335 cycles)
4. External shape and geometry of proton and neutronThe considerations leading to unveiling the struc-ture and shape of the stable particles neutron andproton are given in the article “Brief introduction toBSM theory4, while the detailed analysis is pre-sented in the BSM theory. Both, the proton and theneutron are comprised by one and a same hardwarecompositions of helical structures, whose singleturn element is the same as the electron structure.The proton and neutron are distinguished only bytheir external shape and the slight twisting differ-ence in their helical structures that is responsiblefor their Newton’s mass differences. The shapes ofthe proton and neutron are shown in Fig. 3.
Fig. 3. Geometrical parameters of the proton and neutron. The magnified view shows only the external helical shell without the internal structures of pions and kaon (extraction from Atlas of ANS).
The proton is a twisted torus with a shapeclose to a figure 8, while the neutron is a same
V αc=
1/α3( ) se×
τsp3λc
α3c 1 α2–----------------------------- 6.248 14–×10= =
τmin3λc
α2c 1 α2–----------------------------- 4.5596 16–×10= =

Total pages: 40
8 30 Apr 2003
structure but in shape of double folded torus. Moreaccurately the plane projection of the proton enve-lope is quite close to a Hippoped curve with a pa-rameter . The twisting (and folding)direction is strongly defined by the underlinedstructures of the pions and kaon inside the proton(neutron) envelope (see Chapter 6 of BSM). Con-sequently all protons (and neutrons) involved in theatomic nuclei have one and a same handedness.
The estimated geometrical parameters of theelectron, proton and neutron are given in Table 2.
Physical dimensions of stable elementary particles Table 2===========================================Parameter Value Description Reference BSM (m) Chapter No.------------------------------------------------------------------------- 1.6277E-10 p and n core length 5, 6 0.667E-10 p length 6, 7, 8, 9 0.19253E-10 p width 6, 7, 8, 9 8.8428E-15 small radius of e- 3, 4, 6 5.8952E-15 small radius of e+ 3, 4, 6 1.7706E-14 e- (e+) helix step 3 3.86159E-13 e- Compton radius Known
7.8411E-13 p and n thickness 6, 7, 8, 9------------------------------------------------------------------------- Notations: p - proton, n - neutron, e- - electron, e+ - positron
The following question may arise: why the
proton possesses a charge while the neutron - not.The answer is: The electrical charge is a result ofthe modulation of CL nodes by the internal latticeof the helical structures4. In case of neutron, all hel-ical structures get overall symmetry in respect to itsaxis. For a such shape, the modulation of CL spacein the far field is compensated and the electricalfield is a zero. In case of proton, the overall torus istwisted and the axial symmetry of RL(T) is de-stroyed. Therefore, it is able to modulate the CLnodes also in the far field.
In fact the modulation symmetry for the neu-tron is not perfect in the near field as in the far field,so the neutron still exhibits some modulation of theCL nodes and consequently an electrical field, butonly in a proximity range. This field is locked bythe stronger IG field in the proximity range to itsenvelope so it is not detectable in the far field. Thelocking mechanism of IG field, although, does notwork well when the neutron is in a confined motionin CL space, so it exhibits a magnetic moment (the
magnetic moment of the neutron as a neutral parti-cle has not been satisfactorily explained by themodern physics so far). The electrical field of theproton is always unlocked due to its different over-all shape. Therefore, in the far range the electricalfield of the proton appears as emerging from apoint, but in the near field, it is distributed over theproton’s envelop.
Fig 3 shows the spatial geometry of theDeuteron, where: p - is the proton and n - is theneutron. The neutron is centred over the protonsaddle and kept by the Intrinsic Gravitation (IG)field and the proximity electrical fields of the neu-tron and proton. In such conditions the position ofthe neutron is stable with some rotational freedom,but it is not able to unfold and convert to proton.
Fig. 4 illustrates the protons and neutronsarrangement in the nucleus of He. In such close dis-tance, the internal lattices of the proton’s helicalstructures are kept by IG forces that are inverseproportional to the cube of the distance. The nucle-us of helium is the most compact atomic structure.Therefore, its influence on the CL space parame-ters is the strongest one. As a result, the helium nu-cleus possesses the largest binding energy betweenthe involved protons and neutrons.
.
When taking into account the two features ofthe proton: a finite geometrical size and the distrib-uted proximity electrical field it is evident that theCoulomb law is valid down to some limit, definedby the finite size of the proton structure. This is ver-ified by the model of Balmer series in Hydrogen
a 3=
LPCLPWPrerpseRc
2 Rc rp+( )
Fig. 4. Deuteron withelectron in Balmer seriesaccording to BSM physi-cal model
Fig. 5. Helium nucleusaccording to BSM phy-sical model

Total pages: 40
9 30 Apr 2003
presented in Chapter 7. The idealized shape ofBalmer series orbit is shown in Fig. [7.7].
Fig. [7.7]. Idealized shape of Balmer series orbit. Rc- is the Compton radius, rqm - is a magnetic radius of electronat sub optimal quantum velocity. The Compton wavelength
shown as standing waves is is not in scale
5. Atlas of Atomic Nuclear Structures
5.1. Physical atomic models according to BSM concept.
One of the most useful results of BSM theorywith practical importance is the Atlas of AtomicNuclear Structures6,7 (ANS). The analysis leadingto unveiling the spatial arrangement of the protonsand neutrons in atomic nuclei is provided in Chap-ter 8 of BSM. It shows that the protons and neu-trons follow a strict spatial order with well definedbuilding tendency related to the Z number of the el-ements. The signature of this tendency matchesquite well the row-column pattern of the Periodictable, the Hund’s rules and the Pauli exclusionprinciple. The Atlas of ANS provides nuclear con-figurations of the elements from Hydrogen to Law-rencium (Z = 103). For drawing simplification ofthe nuclear structures, the protons and neutrons arepresented by simplified patterns reminding theirshape. The left part of Fig. 6 shows the patternsused for the proton, deuteron, tritii and helium,
while the right part shows the most common shapesand possible dimension of the quantum orbits. Thedimensions of the quantum orbits and the protonand neutron are given in one and a same scale.
In the Atlas of ANS, the pattern of proton issymbolized by arrow in order to simplify the draw-ings. Additional symbols are also used for the samereason.
For any atomic nucleus, a polar axis can beidentified. It is defined by the long symmetricalaxis of one or more He nuclei in the middle ofatomic nucleus. The atomic nuclei posses alsotwisting features due to the proton twisting, but it isnot shown in the drawings. In the Atlas of ANS ad-ditional symbolic notations are used for the un-veiled types of proton’s bonds and pairing in whichIG and EM fields are involved.
5.2. Three-dimensional structure of atomic nuclei and limited angular freedom of the valence protons.
The Atlas of Atomic Nuclear Structures pro-vides the nuclear configurations of the stable iso-topes. One or more He nuclei are in the nuclearcentre, aligned with the polar axis of symmetry.The peripheral building blocks of the lighter ele-ments are usually deuterons, while the elementtritii appears more frequently in the nuclei of heav-ier elements. The positions of the protons are de-fined by the consecutive number of proton’s shelland by the type of the bonds between the protons.
Fig. [8.2] a. - polar structure; b. - polar-chain structure
Fig [8.2] illustrates the backbone structure ofnucleus allowing the most dens pack of the protonsand neutrons, having in mind the repulsive forces
λc
Fig. 6
Patterns forsimple ele-ments and quantum orbits

Total pages: 40
10 30 Apr 2003
from the proximity fields of the protons. The polarchain structure appears for the atoms with Z > 18(after Argon).
The unveiled type of bonds in the atomic nu-cleus are listed in Table 8.1.
Bonds in the atomic nuclear structure Table 8.1
===========================================Bond notation Description------------------------------------------------------------------------- GB Gravitational bond by IG forces GBpa polar attached GB GBpc polar clamped GB GBclp (proton) club proximity GB GBnp neutron to proton GB EB electronic bond (weak bond)-------------------------------------------------------------------------
The gravitational bonds are held by the IGfield of the intrinsic matter (more explicitly the IGfield of the internal lattices of the helical structuresfrom which the proton and neutron are built). TheIG field controls also the proximity E-field of theproton (and proximity locked E-field of neutron)and its unity charge appearance. For this reason, allGB types of bonds are very stable.
The four types of the gravitational bonds andone type of electronic bond are illustrated in Fig.[8.4], where the positions of some quantum orbitsare also shown.
Fig. [8.4]The left-side structure is 7Li nucleus,. The right-sidestructure is only a portion of nucleus showing the different type of bonds, according to Table [8.1]. The planes of the shown quantum orbits in fact are normal to the drawing plane
The GBpa and GBclp bonds can not haveany freedom of motion. The GBpa bonds for va-lence protons have an angular restricted freedom of
motion in a plane close to the polar section. The EBtype of bonds are valid only for the valence protonsbut they are provided by electron orbit pairing, cor-responding to the Hund rules. There are few typesof such pairings. The two of them are more impor-tant: a first type - two orbits in separated parallel or-bital planes; a second type - two electrons withdifferent QM spins, according to a Pauli exclusionprinciple (their circling directions are oppositeeach other). The second type of Hund rule appearsvalid for EB bonds.
The structural restrictions of the positions ofthe atoms in the molecules comes from two factors:
- stable structural arrangements of the bondedprotons of the nucleus
- angular restrictions of the valence protons. The GB type bonds could not be broken in
any type of chemical reaction, but only in nuclearreactions where quite large energies are involved.The EB type of bond however is weak. Bonds ofsuch type begin from the raw 13 of the Periodic ta-ble while in raw 18 (noble gazes) they are convert-ed to GBclp bonds. The EB bonds normallyexclude the external shell protons from chemicalvalence, so they play a role for the principal va-lence of the elements from group 13 to 17. This isvalid for rows 1, 2, 3 of the Periodical table. Insome conditions, however, the EB bonds could bebroken, so the element may exhibit multiple oxida-tion numbers.
For some chemical compounds between ele-ments with large number of valence protons, not allfree valence protons can be connected by electronicbonds. This is a result of the finite nuclear size ofthe atoms and the angular restriction of the valenceprotons.
It is evident from the nuclear structure thatthe positions of the electron orbits are strictly deter-mined by the positions of the protons with theirproximity electrical fields and the conditions ofquantum orbits provided by Table 1. While the, theelectron orbits are not shown in the Atlas of ANS,their positions are easily identifiable. It becomesapparent that: The first ionization potential, ob-tained experimentally and used in the QuantumMechanical models of the atoms could be ex-plained by the different positions of the electronquantum orbits in the atomic nuclei and the influ-

Total pages: 40
11 30 Apr 2003
ence of the nuclear IG field (from protons and neu-trons).
6. Electronic bonds between atoms in moleculesIt is apparent that the BSM atomic models al-
lows identification of the orbital planes and chem-ical bond orientation of the atoms in the chemicalcompounds. Additionally the quantum mechanicalspin of the electron circling in orbit around the pro-ton is also identifiable. The proton envelope istwisted torus, so it possesses a well-defined hand-edness along any one of its axes of symmetry.Then, the electron in the quantum orbit shown inFig. 4 has an option to circle in two different direc-tion in respect to the proton direction of twisting.This will corresponds to two slightly different en-ergy levels. Its signature is a fine structure split-ting of the spectral line.
The intrinsic conditions of the quantum orbitsdefined by the two proper frequencies of the elec-tron and CL node dynamics are valid also for thebonding electrons in molecules. Let consider amost simple case of H2 molecule identified as anortho-I state. Its shape is illustrated in Fig. [(19.2)]In this figure the three-dimensional shape of theproton is replaced, for simplicity, by a 2-dimen-sional Hippoped curve with parameter . Themolecular vibration in such simple system is of lin-ear type. The long axes of the protons and quantumorbit are aligned with the molecular vibrational ax-is.
Fig. [9.12] Structure of H2 - ortho-I state molecule
Lp - is a proton lengthLq(1) is a long side of a first harmonic quantum orbitrn - is the distance between the Hydrogen atomsr - distance between the electron and the proton’s core in the circular section (for the most external orbit
Note: The quantum orbit quasiplane is perpendicular to the quasiplane of the protons. However, they both are shown in one plane for simplification
of the drawing
The both electrons circle in a commonquantum loop (orbit) but in opposite directions (op-posite QM spins). The quantum orbit crosses theHippoped curves of the protons in the locus points.We may assume that every orbiting electron is ableto neutralize one charge, by interconnecting its E-filed lines to the proton E-field. Let considering themoment, when both electron are in the locus pointsof the Hippoped curves, representing the protons.Their velocity vectors in this case are perpendicularto the direction of the molecular vibration and donot contribute to the momentum energy of the sys-tem. Then, their moment interaction can be esti-mated by considering only two unit charges atdistance rn. Now, let assume that the left proton andthe right electron are both missing. The system en-ergy in this case is [eV]. The same consid-erations and results are valid also for the othersymmetrical case. Adding the energies from thetwo symmetrical cases we get the full system ener-gy.
[(9.4)]
where: the factor 0.6455 defines the distance of thelocus from the central symmetrical point of theHippoped curve with factor .The obtained value of 16.06 eV is pretty close tothe experimentally measured vertical ionizationpotential of the photoelectron spectrum of H2(15.98 eV).
7. Vibrational motion of atoms connected in molecule by electronic bonds.
The Intrinsic Gravitation (IG) is importantfeature of the new concept about the vacuum. IGforces are involved in the following two cases
(a) strong attraction forces valid for small dis-tances
(b) providing the energy of the electricalcharge
The case (a) is valid not only for the prismsand nodes in empty space, but for elementary par-ticles, atoms and molecules in CL space at smalldistances. Examples of the manifestation of the In-trinsic Gravitation are some of the Van der Wallforces between molecules and the Casimir forcesbetween solid polished bodies in close proximity.
a 3=
protonr
rn
0.65Lp 0.65Lp
Lp
Lq(1)
quantum orbit
q/4πεorn
E 2q4πε0 Lq 1 ) 0.6455Lp+( )[ ]--------------------------------------------------------------- 16.06 eV= =
a 3=

Total pages: 40
12 30 Apr 2003
In case (b): BSM analysis unveils the physi-cal meaning of the electrical charge as modulationof CL nodes by the IG field of the internal RLstructures of the helical structures from which theparticles are built4. In such aspect the energy of theelectrical charge could be regarded as a part of theIG energy of the particle in CL space environment.The proximity field of the charge (in approximaterange in order of Bohr radius) possesses a spatialconfiguration, while it appears as a point charge inthe far field. The study of vibrational motion of twoatoms connected in molecule by electronic bondsunveils one important feature. The vibration causesspatial modulation of the near electrical field creat-ed by the protons involved in the electronic bond.This feature is analysed by using a total energy ofthe system that includes:
- the IG energy- the energy of the electrical charge- the kinetic energy of the involved particles- the energy of the emitted or absorbed pho-
ton- the vibrational energy The energy of the IG field is presented as an
integration of IG forces from some initial value toinfinity, but practically the integration could betruncated at finite distance because the IG fieldfalls too fast with the distance
[(9.13)]
where: mpo is the Intrinsic mass of the proton, Gois the intrinsic gravitational constant
- IG factorNote: The factor 2 in front of the integral
comes from the two arm branches (along abcd ax-es) of the CL space cell unit. They both are includ-ed in the xyz cell unit to which all the CL spaceparameters are referenced. All equations using CIGfactor in the following analysis confirms the needof factor 2.
The principle of energy conservation is uni-versally valid. The new concept, however, allowsto see that the IG energy balance is quite fast. Thismeans that the momentary total energy balanceshould be investigated. Based on this approach thevibrational motion of the most simple diatomicmolecule H2 (shown in Fig. [9.12]) is analysed.
The total momentum energy balance at theequilibrium point is given by
(eV) [(9.18)]
where: - is the energy of the twoelectrical charges (for two protons);
- is the kinetic energy of the twoelectrons.
The analysis unveils the vibrational statesand one metastable state of H2 ortho-I molecule.For the vibrational levels with good for identifica-tion accuracy, the following expression is obtained:
(eV) (9.23)
(9.23.a)
where: v - is the vibrational level, vmax is the maxvibrational level identified by the photoelectronspectrum.
The small value of 6.26 ev in Eq. (9.23) islikely a constant due to integration in Eq. (9.13)and its value is obtained by fitting the calculated vi-brational levels to identified optical transitions.
The photoelectron and optical spectrum areanalysed and the corresponding transitions and vi-brational levels are identified. The calculated vi-brational levels are compared to the identifiedvibrational levels from the optical spectrum. Thedependence of the vibrational levels from vibra-tional quantum number fits quite well after adjust-ing one small energy value that is a constant fromintegration in Eq. [(9.13)] This value, however, isabout 10E-6 time smaller in comparison to the IGenergy and could be omitted. Therefore, the analy-sis allows to determine the important parameterCIG.
[(9.17)]
Any disturbance of this balance is related
with emission or absorption of a photon. This isan important conclusion from the analysis.
Fig. [9.24] shows the energy levels Ev , calcu-lated by Eq. [(9.23)] and vibrational levels of theoptical transitions E(0-ν’’) and E(1-ν’’) taken fromexperimental data8 (I. Dabrowsky, 1984). The op-
EIG CP( ) 2–Gompo
2
r3---------------- rd
rne
∞
∫CIG
Lq 1( ) 0.6455Lp+( )2--------------------------------------------------= =
CIG Gompo2=
CIG
q Lq 1( ) 06455Lp+( )2----------------------------------------------------
2Eqq
---------2EK
q----------+=
2Eq/q = 511 KeV
2EK/q = 2 x 13.6 eV
EυCIG
qr2---------
2Eqq
---------–2EK
q----------– 6.26+=
r Lq 1( )[ ] 1 πα4 υm υ–( )2–( )[ ] 0.6455Lp+=
CIG 2hνc hνcα2+( ) Lq 1 ) 0.6455+( )Lp( )=
CIG G0mn02 5.2651 33–×10= =

Total pages: 40
13 30 Apr 2003
tical spectrum is from a H2 system known as a Ly-man system.
Fig. [9.24]. Energy levels Ev, (eV) calculated by Eq.(9.23) and vibrational levels of the optical transitions E(0-ν’’)and E(1-ν’’), corresponding to two QM spin values. The cal-culated levels are shown by step line, while the optical transi-tions by diamonds.
The shown vibrational levels corresponds tooptical spectrum in UV range. They are result oftransitions between these levels (including other“rotational levels”) and one metastable state of H2molecule, the physical configuration of which isunveiled (discussed in Chapter 9).
The fractional error between the calculatedlevels (Ev) by Eq. [(9.230] and the optical data iswithin +/- 0.035%.
Similar analysis is provided also for D2 mol-ecule since it is a more typical building element inthe atomic nuclei. Applying further analysis for di-atomic molecules an analytical expression for vi-brational levels, of homonuclear molecules isobtained, allowing a determination of the approxi-mate internuclear distance of a homonuclear dia-tomic molecule.
(9)
where: EB(n) is the momentum total balance ener-gy of the electronic bond given by Eq. (9.23), A - isthe atomic mass of participated atom in mass units,n - is the subharmonic number of the quantum or-bit, p - is the number of connected protons (valencenumber)
The conditions defining the intermolecularquantum orbits are different from those of atomicquantum orbits. For atomic orbits, the definitionconditions are referenced to the home nucleus andnot influenced by another nucleus. For intermo-lecular quantum orbits, however, the definitionconditions are influenced by the nuclear motion ofinvolved atoms, in which the IG field interactionsare also involved. The IG forces are able to modu-late the spatial configuration of the proximity E-field of the protons involved in the chemical bond.As a result, the vibrational quantum conditionsoccur at intrinsically small deviations from theinternuclear distance (see Chapter 9). For H2ortho-I molecule, for example, the vibrationalrange is only 4E-16 (m) while the internuclear dis-tance is 2.23E-10 (m). This effect in fact facilitatesthe identification of the possible configuration of asimple molecule by combination of two methods:an internuclear distance calculation and a drawingmethod4. The second one is based on the physicaldimensions of the proton (and neutron) and thepossible quantum orbit.
8. Combined method for determination of the possible configuration for diatomic molecules
The possible molecular configurations of adiatomic molecule can be unveiled by using thefollowing approach:
- a drawing method: using the spatial config-uration of the involved nuclei and a selected possi-ble orbit from the quantum orbit set
- a theoretical calculation of the correspond-ing internuclear distance by Eq. (9).
- matching the calculated data with identifiedspectral bands from the optical and photoelectronspectra.
Examples of using the above approach areshown in the article Brief introduction to BSMtheory4. Some of results for diatomic molecules areshown in Fig. [9.43], [9.45], [9.54], [9.56]. In thesedrawings the protons and neutrons in the centralpolar section of the atomic nucleus are only shown.
∆E
rn n A p, ,( ) A p–( )2αCIGpEB n( )------------------=

Total pages: 40
14 30 Apr 2003
Fig. [9.43] Possible configurations of O2 molecule indifferent states (D) and O2(A). (structural states {D} and{A} denoted by BSM model?). The orbital planes of elec-trons do not lie in the drawing plane, but they are shown inthis way for drawing simplification. The number in a bracketindicates the subharmonic number of the quantum orbit.
Fig. [9.45] A possible configuration of O2 molecule in {E}state
Fig. [9.54] Configuration of (OH)- ion. Every electronicbond orbit contains two electrons with opposite QM spins(the planes of bonding orbits are at 90 deg, in respect to theprotons equivalent planes)
Fig. 9.56 One view of CO2 molecule. The CO2 mole-cule possesses rotational symmetry about the polar axis dueto the 90 deg rotational symmetry of C atom. If rotating 90deg around zz axis the view of the left side will change withthe view of the right side. .
Fig. [9.59] Water molecule
9. Structural and angular restrictions of the chemical bonds of electronic type
9.1. Restrictions imposed by the nuclear config-uration of the involved atoms
The provided considerations in §5, §6 and §7demonstrate that the structural and angular restric-tions reduce significantly the degree of freedom ofthe atoms connected by electronic bonds. The samerestrictions are also responsible for the molecularbending. These restricting features are not apparentfrom the Quantum mechanical models of the at-oms.
The mentioned considerations are not validfor ionic bonds where the atoms are not connectedby electronic bonds but by attractive forces be-tween oppositely charged ions. The internucleardistances in ionic bonds are also larger and suchmolecules exhibit different physical properties.Consequently they are not able to possess vibra-tional motion in which the quantum orbits play im-portant roles. For this reasons the ionic compoundsdon’t have vibrational rotational spectra. Only sep-arated ions are able to provide ionic line spectra.

Total pages: 40
15 30 Apr 2003
The following conclusion is valid only forchemical compounds with electronic bonds, butnot for compounds with ionic bonds.• The degrees of freedom of connected atoms
in molecules by electronic bonds are reduced by structural restrictions and limited angu-lar freedom of the valence protons. This restrictions are defined by the nuclear con-figurations of the involved atoms.
9.2. Restrictions from spin-orbital interactionsIf not taking into account the implemented
He nucleus in the atomic nucleus and the polarelectrons, every single proton in the nucleus hasown electron, connected to the free club of the pro-ton (one that is not polar attached). Some of theseproton’s clubs, however, are GBclp or EB bondedin pairs. For EB bonded protons the two electronswith opposite QM spins circulate in a common or-bit whose plane is almost parallel to the polar atom-ic axis. For GBclp bonded protons the twoelectrons with opposite spins circulate in a com-mon orbit whose plane is almost perpendicular tothe polar axis. In both cases, the common quantumorbit passes through the clubs of the protons. Theorbital trajectory is well defined by the proximityE-field of the involved protons. The positions ofGBclp protons are strongly fixed, while the EBprotons are weakly fixed by the quantum orbits.Any fixed orbit may not lye in a plane but it is quiteclose to a fixed equivalent orbital plane that gets aproper symmetrical position in respect to the polarnuclear axis. From above mentioned considerationit is evident that the quantum orbits connected toBGps, GBclp and EB type of bonds have fixed ori-entation of their equivalent planes. These fixed po-sitions are held much stronger than the orbitalplane positions of the chemical bonds.
Table 2 provides the number of different typeproton bonds in the nuclei of some elements andthe total electrons with commonly aligned equiva-lent orbital planes.
Table 2.===========================================
Atom GBclp EB Ne-------------------------------------------------------------------------
C 0 0 2N 0 1 2O 2 0 6P 4 1 10S 4 2 10Cl 4 3 10Ca 8 0 18Fe 8 0 18Cu 8 4 18
-------------------------------------------------------------------------where: Ne - is the number of electrons with
aligned equivalent orbital planes.In the analysis provided in Chapter 8 and 9 of
BSM it is found that two of the external shell pro-tons of the oxygen atom are GBclp bonded. Thesignature indicating such type of bonding appearsin the first ionization potential trend as a functionof Z-number and also in the photoionization poten-tial of the oxygen (known as autoionization fea-ture). The nuclear structure of Oxygen is illustratedin Fig. 7.
Fig. 7. Nuclear structure of the Oxygen atom
In the right part of the drawing, the commonpositions of the fixed electron orbits are shown asviewed from the polar axis. The projections pat-terns of the two polar orbit electrons (1s electronsaccording to QM model) are shown by differentcolours. Their similar patterns exhibit a small an-gular displacement around the polar axis due to thetwisting of the atomic nucleus. This feature, validfor all atoms, is a result of the proton twisting. TheGBclp bonded protons (deuterons) are shown in theplane of drawing, while the valence protons (deu-terons) are closely aligned to a perpendicular plane,

Total pages: 40
16 30 Apr 2003
but shown at oblique angles for a drawing simplifi-cation purpose.
The stability of such nuclear configurationwith two GBclp pairs is evidently a result of the nu-clear symmetry in which the two polar electrons(from 1s shell) have strong influence. This config-uration provides much larger angular freedom ofthe two valence protons that may explain the largechemical activity of the Oxygen atom.
10. Application of the BSM models for analysis of biomolecules with identified structure and composition.
10.1. General considerationsThe 3D structures and atomic compositions
of many biomolecules now are well known. In suchstructures, the individual atoms are identified asnodes with known coordinates. The angular coordi-nates of their chemical bonds are also known. Thisinformation is sufficient to allow a replacement ofthe nodes in the 3D structure of any large moleculewith the physical models of the atoms according tothe BSM theory. If the BSM atomic models arecorrect, their three-dimensional structure and angu-lar bond restrictions should match the three-dimen-sional structure of the biomolecules. Once theBSM models are validated and corrected if neces-sary, the properties of the complex biomoleculesand any macromolecule with known shape couldbe studied and analysed from a new point of view.The application of the BSM models, for example,allows an identification of the positions of all or-bits. This includes the internal shell and the chem-ical bonds electronic orbits, as well. The conditionsfor possible interactions, modifications and energytransfer could be analysed at atomic level.
10.2. Ring atomic structures in organic mole-cules.
Most of the organic molecules contain ringatomic structures. The molecule of benzene couldbe considered as a simple example of a ring struc-ture. The biomolecules usually possess a largenumber of ring atomic structures. Fig. 8. shows the3D molecular structure of aspirin where the ringstructure of 6 carbon atoms is similar as in benzene.
Fig. 8 3D structure of the molecule of aspirin (PDB file aspirin visualized by Chime software)
Fig. 9 shows the same structure of aspirin atatomic level by application of BSM atomic models.
Fig. 9 3D structure of aspirin by BSM atomic models
The single atoms Deuteron, Oxygen and Car-bon and the size of quantum orbit of second subhar-

Total pages: 40
17 30 Apr 2003
monic are shown in the left upper corner. Thevalence protons (deuterons) of the oxygen atom arein fact in a plane perpendicular to the plane of EBbonded protons (deuterons) but they are shownwith reduced dimensions in order to imitate an ob-lique angle in a 3D view. The same is valid also forthe valence protons of the carbon atom.
In Fig. 9 the electronic orbits providingchemical bonds are only shown. For molecule withknown 3D structure and composition the commonpositions of all electronic orbits with their equiva-lent orbital planes are identifiable. It is clearly ap-parent that the 3D structure of the molecule isdefined by the following conditions:
(a) a finite size of the involved atomic nuclei(b) an angular restricted freedom of valence
protons(c) a finite orbital trace length defined by the
quantum conditions of the orbiting electron(d) orbital interactions(e) a QM spin of the electron (the motion di-
rection of the electron in respect to the proton twist-ing)
The QM mechanical models of atoms aremathematical models in which the features (c), (d),and (e) are directly involved, while the features (a)and (b) are indirectly involved by the selection ofproper wavefunctions. In this process howeversome of the spatial and almost all angular restric-tions are lost. Let emphasize now the difference be-tween the suggested BSM models of atoms andmolecules from one side and and the QM modelsfrom the other.
- QM model: the electrons participating inchemical bonds are orbiting around both point-likenuclei, i. e. they are not localised between the nu-clei
- BSM model: the electrons involved in thechemical bonds are localised between the nuclei
-QM model: the chemical bond lengths areestimated from the electron microscopy assumingthe planetary atomic model in which the largerelectron concentrations are centred around thepoinlike nucleus
- BSM model: the chemical bond length mayneed re-estimation, because the orbits of the chem-ical bond electrons do not encircle the bound atom-ic nuclei.
- QM model: The length of C=C double va-lence bond is estimated as 1.34 A (angstrom),while for a single valence C-C - as 1.54 A. Howev-er, these lengths show a small variation in the samering groups included in different biomolecules.
- BSM model: the length of single C-C bondmay vary only by the subharmonic number ofquantum orbit, while the length in a double C=Cbond is additionally dependent of the angular posi-tions of the valence protons.
The adopted and existed so far concept of cir-cling delocalised electrons for explanation of theequality between single and double bonds in ben-zene molecule is not logical from a point of view ofBSM model. This is quite important for unveilingsome of the specific properties of the ring struc-tures in the biomolecules.
Ring structures are very abundant in manybiomolecules and they are very often arranged inparticular order along the long chain structure ofthe molecule. DNA and proteins contain largenumber of ring structures. Fig. 10 shows the spatialarrangement of ring atomic structures in a portionof β−type DNA. The positions of some (O+4C)rings from the deoxyribose molecule that is in-volved in the helical backbone strands of DNA arepointed by arrows.
Fig. 10 Part of DNA structure with indicated positionsof some of (O+5C) atomic rings
Fig 11. shows the ring atomic structure
(O+4C) from the DNA strand. The deuterons in-volved in the ring structure shown in Fig. 11 prac-tically have some small twisting, but the quantumorbit of single valence bond also could be twisted.This feature gives some freedom for formation ofring structures of different atoms. The rotationalfreedom of the single valence bonds, however, may

Total pages: 40
18 30 Apr 2003
be accompanied by some stiffness that increaseswith the degree of the orbital twisting.
Fig. 11. Ring atomic structure from the deoxyribose molecule involved in DNA strand
In DNA molecule, some of the atoms of thering structure are also connected to other externalatoms. All this considerations provide explanationwhy the ring structure (O+5C) connected to theDNA strand is not flat but curved.
10.3 Weak hydrogen bondsIt is known that a weak hydrogen bond is pos-
sible between two atoms, one of which does notpossess a free valence. The bond connection is a re-sult of orbital interactions. In such aspects, the hy-drogen bonds connecting the purines topyramidines in DNA molecule are of two types: <N-H...O> and <N-H...N>, where the single va-lence electronic bond is denoted by “-” and the H-bond is denoted by “...”. The BSM concept allowsto find the possible orbital orientation for such typeof bond. This is illustrated by Fig. 12.
Fig. 12. Two types of hydrogen bonds
In a hydrogen bond of type N-H...O theplane of electronic orbit of hydrogen appears al-most parallel to the commonly oriented nuclear or-bits of oxygen atom in which six electrons areinvolved (see Table 2 and Fig. 6). In a hydrogenbond of type N-H...N the plane of electronic orbitof hydrogen is almost parallel to the equivalentplanes of the two polar orbits of N in which twoelectrons are involved.
It is evident that the hydrogen bond is charac-terized by the following features:
- the connection is a result of common orbitalorientation
- the H-bond requires critical range of dis-tance
- the H-bond allows a rotational freedom in alimited angular range.
This three features allows the DNA moleculeto possess excellent folding properties.
10.4. Hypothesis of energy storage mechanism in molecules possessing ring atomic structures.
It is well known from the atomic spectra, thatonly the alkali metals (Group I) and the positiveatomic ions with a single valence electron possessatomic spectrum that could be described by theBohr atomic model. For elements with more thanone valence electron, the principal quantum num-bers exhibit more than one energy level (degener-ate levels), due to the spin orbital interactions. The

Total pages: 40
19 30 Apr 2003
signature of this feature is apparent from the Grot-rian diagrams of the atomic spectra. The spin-orbit-al interaction from a point view of BSM atomicmodels is discussed in Chapter 8 of BSM. Theanalysis leads to a conclusion that the pumped CLspace energy is not emitted in full, after an orbitingelectron is dropped to a lower quantum level. Partof the pumped energy is preserved by the atomicnucleus and redirected to the valence proton,whose quantum orbital plane is parallel to the orbit-al plane of the consideration. The physics of thiseffect is explainable if considering the total energybalance including IG field. The latter controls theproton’s proximity E-fields distribution, that fromhis hand defines the orbital conditions of the elec-tron. Therefore, the redirected energy providessome shift of the energy levels, but the effect isstronger for the lower states, closer to the groundstate of the series (Balmer series has own groundstate, according to BSM model). The physical ex-planation of this effect allows making a conclusionthat the released energy prior to formation of a pho-ton is preferentially guided by connected struc-tures of protons (deuterons). Applying the sameconsiderations for the ring atomic structures theremust therefore be a guiding energy process be-tween the atomic nuclei or protons involved in thering. Such consideration leads to the followingconclusion:
In proper environments, the ring atomicstructures in organic molecules may have abilityto store energy as an exited state rotating in thering loop.
The effect of the rotating excited state is pos-sible due to the consecutive re-excitation of theelectrons in the separate bonding orbits in the ring.This effect is not apparent by the Quantum me-chanical model, where the wavefunctions are com-plex envelope around the whole nuclei of theinvolved atoms. However, it is a known fact thatthe bonding strengths between atoms involved in aring atomic structure are stronger than betweensame atoms when not participating in such struc-ture.
Evidently, the condition of rotating excitedstate in a ring structure could be obtained only forequal energy level differences. Such concept al-lows considering excited states not only from samevalence bonds but also from single and second va-
lence bonds as well. From the other side, for a ringstructure containing more than two bonds of samevalence, excited states may preferentially exists be-tween the same valence bonds. In case of aspirin,for example, such conditions exists for three pairsof orbits of second valence and three pairs of orbitsof single valence bonds. If considering also thefixed nuclear orbits of the atoms in the ring thentwelve polar electrons could be also involved in aring storage effect. Theoretically they may store amuch larger energy not only due to their number,but also due to the larger transition energies.
In proper environments, the stored energy inthe ring structures of the biomolecules may havethe following features:
- a stable cycle of exited state rotation dueto a stable finite time of single excited state
- a possibility for interactions with proper-ly oriented neighbouring ring structures in themoment between two consecutive excited states(conditions for synchronization between the ro-tating states of neighbouring rings)
- a cascade type of energy transferMany of the building blocks of the biomole-
cules or reagents contain number of single or at-tached rings. For example Adenine (2 attachedrings), Guanine (2 attached rings). Vitamin D con-tains one single and two attached rings. Alpha andBeta tubulins contain groups of: GDP (one singleand two attached rings), GTP (one single and twoattached rings), TAXOL (4 single rings). The ster-oids hormones contain usually four attached atom-ic rings. The ATP, an important energy carrier inthe cells contains one single and two attachedrings. It is quite logical to consider that the energyrotating cycles in the attached rings are mutuallydependable so they must be synchronized. Then itis logically to expect that the attached rings mayhave an increased ability to hold a stored energy incase of environment change.

Total pages: 40
20 30 Apr 2003
10.5 Hypothesis of energy flow through the chain structure of the biomolecule.
10.5.1. Energy flow in DNA molecule and its effect on the higher order structural character-istics.
For long chain biomolecules, like DNA, thering atomic structures are characterised by few ad-ditional features:
(a) a strong repeating order(b) a strong orientation in respect to the host
strand of DNA(c) a strong orientational order of the neigh-
bouring rings along the helixThese features are well known and can be
easily visualized when rotating the 3D structure ofDNA (by programs like: “chime” “Rasmol”, “pro-tein explorer” etc.).
The consideration of cascade type of excitedstate transfer could be applied not only for a ringatomic structure but also for a long chain moleculebuilt of repeatable atomic structures connected byelectronic bonds. In this case some more compli-cated but mutually dependable mechanisms are in-volved. The following analysis tries to unveil suchmechanisms. Let considering for this purpose oneof the backbone strands of DNA. Fig. 13. illustratesthe connection path of the electronic bonds in thestrand.
Fig. 13. Bond connection path through a DNA strand The connection path corresponding to one cascadeof the nucleotide is denoted by a thick green line
Three important features are apparent forevery repeatable cascade:
(d) the bond connection path is formed bydeuterons or protons connected by electronic orbits
(e) the bond connection path passes throughone C-C bond from the (O+5C) ring.
(f) all bonds involved in the bond connectionpath are single valence
It is reasonable to expect that the long chainof single valence bonds may provide conditions forcascade excited state transfer in one direction. Thetime of every excited state is determined by a quan-tum mechanical consideration - the lifetime of thespontaneous emission. Having in mind the smalldistance between the neighbouring electronicbonds, the transfer time between two consecutiveexcited states (with a light velocity) is practicallyalmost a zero. Then the time dependence betweenthe two energy process (a cascade energy transferand cycle period of energy rotation in the ring) iseasily obtainable. The bonding path of one cascadecontains six bonds total in which one C-C bondfrom the (O+4C) ring is included. This ring in-volves five bonds. Then one may consider that thefollowing condition is valid:
(9)where: tc - is the time transfer interval for one cas-cades of nucleotide estimated by the sum of life-times of excited states in involved bonds, TR - isthe cycle time of the rotating state in the ring, -is the average lifetime for a single bond.
The expression (9) means that the cascadetransfer and ring cycle are mutually time dependa-ble processes, so they should have proper phasesynchronization. Additionally, all parameters ofEq. (9) are dependable from the temperature, but ina different way. This will impose a limit tempera-ture range for successful phase synchronization.
Keeping in mind the features (a), (b), and (c),the rotating energy states in the ring could be com-monly dependable.
The whole mechanism will be characterisedby the following features:
(k) the rotated excited states in (O+5C) ringswill possess one and a same handedness deter-mined by the direction of cascade energy flowthrough the strand to which they are attached
(l) the rotating energy states are phase syn-chronized along the DNA strand
0 TR tc–( ) τav< <
τav

Total pages: 40
21 30 Apr 2003
(m) the rotating energy states sustain the ten-dency of unidirectional cascade energy flowthrough the DNA strand
(j) the whole mechanism will work at opti-mum temperature and limited temperature rangeboth defined by the conditions of optimal phase be-tween the cascade energy transfer and the ring en-ergy cycle.
It is apparent that the commonly dependablefeatures (g), (h), (h) and (j) will lead to a self-sus-tainable mechanism.
Let analyse now the conditions that may sup-port the tendency of unidirectional energy transfer.For this reason we will consider a small portion ofDNA ignoring it supercoiling. In such case it couldbe regarded as a linear type DNA. It is important toemphasize two structural features of Beta typeDNA that might be related to the tendency of uni-directional energy transfer:
- It is well known that the nucleotide arrange-ment in DNA is antiparallel, so the same definitionis valid, also, for the bonding paths through the twostrands.
- The DNA double helix is characterised by aminor and major grove. This means that one of thehelix is slightly axially shifted in respect to the oth-er.
The concept of unidirectional energy flowthrough the DNA strand could be investigated if as-sociating it with the magnetic field of a solenoid. Insuch approach, the double helix configuration ofDNA could be regarded as two parallel solenoidswith a common axis. Now let consider that the cas-cade energy flows through the both strands are inopposite (aniparallel) directions. This will corre-sponds to opposite currents through both solenoids.In such case, the magnetic lines in the internal re-gion of the solenoids will have antiparallel direc-tion, while the external magnetic lines will beclosed in the proximity of both ends of the sole-noids.
Let call this type of field a “complimentarycompensated solenoids type”. The magnetic linesof such field are schematically illustrated by Fig.14. The two solenoids that simulate the two strandsare shown by green and red. Their field lines shownas dashed lines are antiparallel inside the solenoids,
while they are connected in proximity at the bothends.
Fig. 15. Association of energy flow conditions between DNA strands with complimentary compensated solenoids. The magnetic field linesclose in a proximity at both ends of the solenoids. The current directions in both solenoids are opposite(shown by black arrows)
If attempting to separate the both solenoids,they will opposed.
The analysis of magnetic field with such spa-tial configuration leads to the following additionalfeatures:
(h) The configuration of the associated mag-netic field is independent of the secondary (super-coiling) shape of DNA
(i) Such type of magnetic field will providean additional strength of the connections betweenthe both strands. It will oppose the separation be-tween the strands because this leads to increase ofthe close path lengths of the magnetic lines
(j) For a small portion of DNA molecule thecomplimentary compensated solenoids type offield is axially symmetrical.
The provided considerations may put also alight about the hydrophobic mechanism existing inthe space between the two DNA strands. The twobond angles of the water molecule are illustrated inFig. [9.59] where the positions of the orbits of thetwo valence electrons are also shown. If a watermolecule is placed inside the symmetrical field ofthe compensated solenoids, the angular positionsof the valence electron orbits evidently will be in aconflict with the solenoids field. The interaction ofthe orbiting electrons with such field may provideexpelling forces for such type of molecule. Thismight explain the hydrophobic environments of theinternal region of DNA between the two strands.The hydrophobic environment is quite importantfor H-bondings between the purines and pyrimi-dines.

Total pages: 40
22 30 Apr 2003
The analysis of compensated double solenoidmodel for the energy flow through DNA leads tothe following conclusion:
(A). The DNA double helix molecule couldbe easily folded in any shape under influence ofexternal factors.
The external factors could be different kindof proteins.
10.5.2. Magnetic field conditions for proteins.It is well known, that in proper environments
the proteins, possessing usually a complex tertiarystructure, preserve their native shape. The linearDNA, from the other side, when it is free of bend-ing proteins, does not exhibit such a feature. Themain reason for the different behaviours of the pro-teins and the DNA of linear type perhaps is a resultof the structural differences between them. Theprotein is a long single strand molecular chainwith diversified sequences of aminoacids. The pro-tein backbone does not contain low order repeata-ble structures as the DNA nucleotide. It maycontain, however, higher order repeats, that mayform some helices with small number of turns orother spatial configurations. In such arrangementthe conditions for H-bonding are significantly re-duced. All these structural differences indicate thatwe could not apply the concept of the complimen-tary compensated solenoids for the proteins, as forthe analysis of DNA.
Fig. 16. tRNA molecule. (a) - real shape, (b) planedrawing, showing the loops and base pairs
The conclusion, that the diversified sequenceof the aminoacids is one of the reason for the dif-ferent higher order shape of the proteins is support-ed by the analysis of the shape of the tRNAmolecule. Its shape and atomic composition isshown in Fig. 16.
The tRNA molecule possesses a single strandof base pairs but the repeating order of the nucle-otides is similar as in the DNA molecule. Its singlestrand arrangement obtains a stable shape of “clo-verleaf” demonstrating in such way a tendency forarranging parts of the long single strand in proxim-ity.
In such arrangement, the unidirectional mag-netic field through the single chain of tRNA be-comes closer to the magnetic field of thecomplimentary compensated solenoid, so the prin-ciple of shortest magnetic lines is satisfied. Themagnetic lines through the strand are connected inthe proximity at the open ends and this is the areawhere the tRNA molecule attaches a proper aminoacid.
The proteins synthesized according to theDNA code have different sequences of amino acidsand their taxonomy exhibits a large diversity.While the model of the complementary compensat-ed solenoids is not valid for the proteins, they stillcould be analysed by a single solenoid model. If ac-cepting that a unidirectional cascade state transferis possible through the bonding path of the molec-ular chain, then a single solenoid model could beused for the helices of the protein. In such aspect,the analysis of the secondary and tertiary structuresof the proteins leads to the following considera-tions:
(a) Different types of secondary structures ashelices, sheets, beta turns, bulges and so on providedifferent structures of magnetic fields.
(a) The magnetic field from the unidirection-al cascade state transfer may influence the commonpositions of the secondary structures
(c) The bond angular positions betweenneighbouring atoms are restricted by the strongconditions of finite nuclear structure and restrictedangular freedom of the chemical bonds
(d) Discrete type of rotational freedom withsome finite deviation is possible between single va-lence bonds

Total pages: 40
23 30 Apr 2003
(e) The above mentioned considerations andspatial restrictions lead to a conclusion that anoverall asymmetry of the magnetic filed from thesecondary structures (mainly from the helices) mayexist. Then the tertiary structure may try to restorethe asymmetry of magnetic fields arisen from thediversified secondary structures by obtaining anoverall symmetry at some higher structural order.Simultaneously, this will be accompanied with atendency of shortest (as possible) magnetic lines.
(f) The cascade energy transfer through thebonding path of the protein chain might be accom-panied with some ion current, whose energy couldbe also involved in the total energy balance.
The taxonomy of 3D protein structures iswell described in the book Advances in ProteinChemistry9 (1981) by C. B. Anfinsen, J. T. Edsalland F. M. Richards.
The stiffness of the secondary order structureis larger than the stiffness of the tertiary one. Thismatches the strength of the mechanism involved inthe restoration of the overall field symmetry. As aresult every protein may have a sustainable nativeshape when placed in proper environments (tem-perature, pH, ATP).
The provided analysis helps to explain one ofthe problems in molecular biology, known as aLevinthal’s paradox: “Why the proteins folds reli-ably and quickly to their native state despite the as-tronomical number of the possible quantum statesaccording to Quantum mechanics?”.
Now let find out how the protein may react ifthe condition of shortest magnetic lines is tempo-rally disturbed by some external factor, for exam-ple, by addition of Adenosine triphosphate (ATP).The ATP carries stored energy in its rings. Whenthe chemical reaction occursthe stored energy might be induced in some ringsin the protein under consideration. This maychange the conditions of the compensated magnet-ic field (related to the principle of shortest magnet-ic lines). Then the reaction leading to restoration ofthe energy balance (involving the magnetic energy)could involve some kind of change of the shape ofthe protein that could invoke some motion. Thismay eventually explain the protein motility inproper environments (temperature, pH, ATPconcentration) that is related with a temporarychange of its higher order shape.
10.5.3. Environment considerations for preser-vation of the native shape of the proteins.
Many proteins have a complex 3D structurescomprised of secondary helix and tertiary struc-tures. They exhibit amazing tendency to preservetheir 3D shape in proper environment conditions inwhich the temperature is one of the important fac-tors.
The electronic bonds and hydrogen bonds al-low some freedom of the 3D structure of biomole-cules. It is known from the Quantum mechanicsthat the excitation of particular vibrational-rota-tional band of the molecular spectra is temperaturedependant. Translated to BSM model this depend-ence means activation of proper subharmonicnumber from the available set of the quantumorbit for each electronic bond.
Consequently the complex 3D shape of thebiomolecules is dependable of two main features:
(a) a proper subharmonic and quantumnumber of every bonding electron
(b) a same subharmonic and quantum numberof the similar interatomic bonds along the molecu-lar chain
The feature (a) puts a restriction for the ab-solute temperature range in order to preserve thenative state of a long chain molecule, while the fea-ture (b) puts a strict requirement for a temperatureuniformity along the molecular chain. This, ofcause is not the only factor. Additional factors arethe interactions between the highly oriented atomicring structures discussed in previous paragraphsand the conditions for H-bondings.
Another environmental conditions that arealso important, for example, are such as the pH fac-tor and ATP concentration. The first one may as-sure the proper ion current conditions (accordingconsideration (f) in section 10.5.2, while the sec-ond one - the necessary energy.
10.5.4. Magnetic field involved in the higher order structures of DNA.
The DNA molecules of the single cell organ-isms are usually of circular type with a supercoil-ing. The DNA molecules of eukaryotes, however,are of linear type with much more base pairs andcomplicated secondary and third order structures.
ATP H2O ADP P+→+

Total pages: 40
24 30 Apr 2003
According to the principle of the shortest magneticlines and compensated magnetic field, the circularDNA should be more resistant, because the mag-netic line paths are enclosed inside of the structure.This condition might be partly disturbed only dur-ing the transcription. The linear DNA, however,possesses some additional properties that may fa-cilitate the process of intermolecular DNA commu-nications, according to a hypotheses presentedbelow.
The length to diameter ratio of DNA is verylarge. It is extremely large especially for eukarioticDNA. For human chromosomes, for example, theaverage ratio is in order of 1x107. In order to beheld in a tiny space of cell nucleus such long mol-ecule have higher order structure, known as super-coiling. The mechanisms involved in the formationand sustaining of such a structure has not beencompletely understood so far. The following anal-ysis put some light about the possible mechanisms.
Let presenting briefly some of the experi-mental observations about supercoiling features ofa long DNA. In a paper10 “DNA-Inspired Electro-statics” in Physics Today by W. M Gelbart et al.(2000), the authors provide a summary about oneimportant feature of DNA. They say: “Under phys-ical conditions (a 0.1 molar solution of NaCl), aDNA molecule takes on the form of a disorder coilwith a radius of gyration of several micrometers; ifany lengths of the molecule come within 1 nm ofthe other, they strongly repel. But under differentconditions-in a highly diluted aqueous solution thatalso contain a small concentration of polyvalentcations - the same DNA molecule condenses into atightly packed, circumferentially wound torus.”Fig. 17 shows the toroidal shape of DNA from thesame paper10. The figure has been adopted from OLamber et al., Proceedings of the National Acade-my of Sciences (USA), vol. 97, p.7248, (2000).
Fig. 17 Toroidal DNA condensates, Courtesy of O.Lambert et al. Proceedings of the National Academy of Sci-ences (USA), vol. 97, p.7248, (2000).
The DNA molecule is usually negativecharged, so the repels between the different part ofthe long chain molecule in close distance (aboutone nm) is understandable. However, why theDNA folds in such packed toroidal structure in apresence of proper polyvalent cations? This effectgets reasonable physical explanation by BSM the-ory if analysing the magnetic field conditions at aCL node level.
Fig. 18. Electrical field and SPM Quasisphere elonga-tion between two parallel parts of DNA
Fig. 18 shows the electrical field intensity be-tween two parallel parts of DNA. The left verticalscale axis shows the electrical field intensity, while

Total pages: 40
25 30 Apr 2003
the right vertical axis shows the elongation of theSPM quasispheres in the plane of drawing. Onespecific feature is apparent from the drawing: theelongation of SPM quasispheres becomes zero inthe middle between the parallel strands. (see “Briefintroduction to BSM theory..”4 or Chapter 2 ofBSM for more details about the Magnetic and andElectrical quasispheres of SPM vector). This ispossible only due to the strong spatial orientationof the Electrical Quasispheres (EQ), contributed tothe E-field along the parallel DNA parts. Conse-quently, the elongation of all SPM quasispheres inthe plane passing through the parallel DNA axes isreduced to zero. Then these SPM quasispheres be-come of Magnetic Quasisphere (MQ) type. In thesame time the SPM vector of these MQs might besynchronized, because the interacting EQs from theparallel long chains could be easily synchronized.Then the obtained in such way MQs provides ex-cellent conditions for a permanent magnetic field.In order the its direction to be stabilized, however,it needs some complementary interactions (havingsome inertial features) with some external currentinvolving charge particles. Such current may beprovided by some heavy and polyvalent ions (be-cause the masses of the light ions Na and Cl are notso different and their charge is not enough large.The currents from the positive and negative ionswith overall helical trajectories are expected tohave different paths. This could happen if theyhave different masses, because they will get differ-ent centripetal acceleration in a helical trajectory.The supercoiling shape of the long DNA will pro-vide a necessary condition for such trajectory. Inthe same time the condition of shorter magneticlines will keep the supercoiled DNA in compactconfiguration, while the negative charge of DNAstrands will keep the proper distance between thedifferent parallel parts of the molecule.
The explanation of the DNA supercoiling ineukariotic cells in a natural cell environments re-quires some additional considerations. The humanDNA, for example, is supercoiled around a proteincalled histon octamer. Presently, the shapes of thehigher order structures of DNA as a nucleosomeformation, a Chromatin and a Chromosome arewell known. They are illustrated by Fig. 19.
Fig. 19. Higher order structures of DNA in Chromatinand Chromosome
The conclusion (A) in §10.5.1 provides anexplanation why the DNA molecule is easily fold-ed as a secondary helix around the histone octamer.The latter, evidently, provides a higher order helicalshape conditions. According to the considerationsin §10.5.1 and §10.5.2, the histones may also havefeatures of energy flow. If examining the atomicstructures of the histones (by using the Protein dataBank) we will find that two type of atomic rings:(6C) and (N+4C) are involved into the buildingblocks. A possible energy flow through the spatial-ly arranged rings may play a role in the DNA bend-ing and coiling around the histone octamer.
The Chromatin, known as a chromatin fibber(with diameter of 30 nm) is additionally coiled intoa daisy flower shape, called a chromosome mini-band. The chromosome miniband, shown in the leftbottom side of Fig. 17 contains 18 loops of daisyflower shape. A stack of daisy flower minibands ar-ranged in a superhelix with diameter about 840 nmforms a chromosome.
The discussed effect of magnetic field crea-tion with sustainable ion current might be part ofthe complex mechanism sustaining the supercoiledstructure of the human chromosome. The balancebetween the magnetic field, the ion current and theelectrochemical potential will depends from the in-ternal cell environments as in the case of the pro-teins. Simultaneously, the principles of the shortestmagnetic lines and compensated magnetic field

Total pages: 40
26 30 Apr 2003
should also play some role. When one of the bothends of the chromosome is attached to the cell theion current and the magnetic field will be stabi-lized, so the supercoiled shape will be stable. If thechromosome, however, is not attached by somereason, a small change in the ion current will causea reaction of the magnetic field. This may invoke ashape change and a motility of the chromosome.
10.6. Hypothesis about a DNA involvement in a cell cycle synchronization mechanism
10.6.1 General considerationsThe cell cycle synchronization is an impor-
tant factor for a normal organ formation in thegrowing multi-cellular organism. One simple ex-ample demonstrating its importance is the eye for-mation. Some not synchronized cell division inearly phase of eye formation may lead to signifi-cant defects.But, how the enormous number of in-dividual cell know when exactly to enter intomitosis? If assuming that an autonomous clockmechanisms in the cells are involved, they could beasynchronized after a number of cycles. A synchro-nization by chemical messengers passing throughthe bulk of enormous amount of cells is also notquite convincing for synchronized growing of or-gan like eye, for example, consisting of many thou-sands cells.
It is reasonable to expect that DNA might beinvolved in the cell cycle synchronization. Theanalysis of DNA at atomic level by BSM conceptled to formulation of hypothesis about electromag-netic intercommunication capability of DNA mol-ecules located in different cells of a same type. Theproposed hypothesis relies on a pure physicalmechanism some of the features of which has beendiscussed.
Let use the concept of complimentary com-pensated solenoids described in §10.5.1., but re-garding the both complimentary parts of thissystem as separate helices. In this case, they couldbe considered as two solenoids serially connectedin circle. Then the direction of cascade energytransfer through both strands of DNA (correspond-ing to the two separated solenoids) could be aclockwise or counter clockwise. This will definetwo different directions of the magnetic lines. Letsuppose, that the obtained magnetic field is in-
volved also in some interaction processes in thecell, so it could not be reversed spontaneously. Thismeans that the direction of the cascade energy flowwill be kept stable in respect to the helical direc-tion. Let denote the direction of a stable energy cas-cade through one strand to be +z. This directionregarded as axis +z in fact is not a straight line, buta helix geometrically centred along one of theDNA strands. Obviously, the stable energy flowthrough this strand will influence the direction ofthe rotating energy states (handedness) in the at-tached (O+5C) rings. Thus we may accept that theyall have a clockwise direction coinciding with therotating direction of the running energy cascadethrough DNA strand along +z axis. In other wordsall stored energies in (O+5C) rings have the samehandedness. Note that the definition of handednessof the rotating energy states in the rings is refer-enced to the direction of the cascade energy flow.
Let now pay attention about the energy stor-age capability of Purines and Pyrimidines involvedin DNA. The pyrimidins Cytozyn (C) and Thymin(T) have single rings (2N+3C) with electronic bondconnection to the DNA strands. The Purines ade-nine (A) and Guanine (G) both have two attachedring structures (2N+3C) and (2N+4C) that are alsoconnected to the DNA strands. Because, the singlerings of C and T are the same as the attached ringsof A and G, we may accept that they carry one anda same amount of stored energy.
The Purines and Pyrimidines have strongerbond connections to their own strand, than betweenthemselves as base pairs. The handedness of theirstored energies could be also influenced directly bythe handedness of the energies in the (O+5C) ringsfrom the strands. Using the same logic, the(2N+3C) rings should get the proper handednessfrom the strands to which they are strongly con-nected. The attached to them (2N+4C) rings how-ever will get a complimentary oppositehandedness.
Now let consider the normal situation, whenthe both strands of DNA are closely spaced in ashape of double helix. If aligning the DNA along anew defined axis +Z, the rotated energies in therings connected to one selected strand, say a firstone, will have a clockwise rotation, while from thesecond one - a counter clockwise. Consequently,when introducing a common direction axis (+Z) for

Total pages: 40
27 30 Apr 2003
both strands, the energies from (O+5C) connectedto the two strands of DNA will look as they havedifferent handedness. (In the further analysis, thecommon +Z axis will be considered for bothstrands).
Table 3 provides the handedness of the storedenergies for all types of rings in DNA molecule, asa result of the above analysis. The handedness ofstored energy in any kind of ring is referenced to acommon axis Z+ of the DNA molecule.
Energy and handedness states Table 3===================================strand A strand B Energy states for------------------------------------------------------ES/|\ ES\|/ (O+5C) ring (E1/|\)(E2\|/) ------- (E1\|/) A---T and G---C (E1/|\) ------- (E2/|\)(E1\|/) T ---A and C---G-----------------------------------------------------Notations:
/|\ and \|/ - denotes two states of handednessES - a rotating state energy in (O+5C) ringE1 - a rotating state energy in (2N+4C) ringE2 - a rotating state energy in (2N+3C) ring----- a connection by hydrogen bond(E1/|\)(E2\|/) - energy states with complimen-
tary handedness in attached ringsLet examine, now, how the stored energies in
the rings of different types could be influencedfrom some change of cascade sequence. The ener-gies of (O+5C) rings can be easily affected by achange in the energy cascade flow through thestrand, because the strand bonding path passesthrough C=C bond of every ring. The stored ener-gies in single (2N+3C) rings could be affected bythe change of the cascade type energy flow through(O+5C) rings. The stored energies in the attachedrings (2N+3C) and (2N+4C), however, is of com-plimentary type, so it is more resistant to the men-tioned above energy flaw changes.
Summarizing the above considerations wemay conclude:
(1) The stored energy sequence in (O+5C)rings (with unit value of ES) is strongly influencedby a change in the cascade energy flow throughDNA strands
(2) The complementary energies (E1/|\)(E2\|/)in the attached rings are more stable than the ener-gy E1 in the single ring.
(3) A change of energy sequence in (O+5C)rings could influence stronger the stored energiesin the single (2N+3C) rings than in the (2N +4C)rings.
(4) The energies E1 and E2 from connectedby H bonds A--T and G--C have always the samehandedness.
Now, let consider that the DNA strands areopened in one end only. In such case, the paths ofthe magnetic lines is increased at this end. It isknown from the physics that if some alternativefield components appear in such conditions, someenergy will be emitted as EM waves. At this point,however, we must consider some specific featuresof the double helix structure DNA:
The (O+4C) rings, arranged in a helix, con-tains equally spaced gaps. Having in mind the enor-mous number of these rings, the cascade energytransfer will have a very large unidirectional com-ponent but quite a small alternative component forone clock. Then it is reasonable to accept that thegenerated magnetic field will have the same unidi-rectional feature like the field from the energy cas-cade through the strands. This means that everyemitted pulse will have energy of 2ES.
When considering the energies of singlerings (N2+C4), however, the emission conditionsare different. They do not form uninterrupted se-quence along the chain as the (O+4C) rings, so thesequence of the released E1 energies can not pos-sess an own permanent component. This might af-fect the preferred direction of the magnetic fieldgeneration. The helicity of DNA and higher orderhelicity of chromatin and chromosome with somecomplimentary ion currents may also provide con-ditions of energy release from (O+4C) rings in oneand a same direction for both strands. It is reasona-ble to accept that the emitting direction could fol-low the direction of emptying the energies of(O+4C) rings. Let considering that this directioncorresponds to the introduced +Z axes. Fig. 20 il-lustrates the sequence of emptying the stored ener-gies in (O+4C) rings (with individual energy ES)and (2N+4C) rings (with energy E1). The energystatus and the direction of energy emission areshown for two consecutive clocks (i) and (i+1). Theenergies stored in the two attached rings (2N+4C)and (2N +3C) are not shown in the figure, but theyare always complimentary to the (O+4C) rings.

Total pages: 40
28 30 Apr 2003
Fig. 20 Energy status and direction of energytransfer for two consecutive clocks. Theemitting direction is along +Z axis
The handedness of all ES energies in respectto the direction of the permanent magnetic field(defined by the cascade energy transfer through thebonding path) indicated by arrows in a dashed lineis one and a same. When emitted by the common+Z direction the corresponding ES energies con-tribute to a common 2ES pulses. For E1 energies,however, the emission conditions are different be-cause they do not have a stable unidirectional se-quence. Then considering the energy emptyingdirection +Z the handedness from E1 belonging tothe different strands are different (clockwise orcounterclockwise in respect to +Z axis). Let con-sidering:
(a) The E1 energies with clockwise handed-ness in respect to +Z axis provide a photon se-quence with a clockwise phase difference betweenthe photons.
(b) The E1 energies with a counter clockwisehandedness in respect to +Z axis provide a photonsequence with a counterclockwise phase differencebetween the photons.
(c) The emitted photons of E1 energies fromthe same strand are entangled with preserved phasedifference. The phase advance in one strand (with+Z orientation of its cascade energy) will have aclockwise phase sequence, while from the otherstrand it will have a counter clockwise.
(d) Anyone of the both sequences of entan-gled photons contains embedded information aboutthe A--T and G--C sequence of DNA referenced toone strand.
(e) The energy of entangled photon sequenceis less dispersive in conditions of intercellulartransmission.
(f) The probability of absorption of the entan-gled photons is much higher if meeting a similarspatially arranged atomic rings.
(g) If the process of DNA energy release isinvoked not by internal factor, but from externallyinduced synchronization, it might have some phasedelay, due to a missing of the initial synchroniza-tion sequence.
The photon entanglement, as a physical phe-nomena) is observed in lasers. Firstly, two entan-gled photons have been observed few years ago.Observation of three entangled photons has beenpublished by Zelinger group11,12 (D. Bouwmeesteret al, 1999). The authors express opinion for “en-tanglement between many more particles”. Thesame group later reported observation of four en-tangled photons and provided also experiment ofstimulated emission of polarized-entangledphotons12 (A. Lamas_Linares, J. C. Howell & D.Bouwmeester, 2001). In the conclusions of latterarticle they write:” We consider that entaglementrobustness-together with the rotational symmetryof the state created by stimulated polarization en-tanglement - opens the way to many applications inquantum information, and provides a powerful toolfor studying the almost unexplored area betweenthe discrete and the macroscopic optical quantumcorrelation experiments.”
Experiments with entangled photons showthat the entangled states fight the diffraction limitvalid for a single photon. This may provide expla-nation of the mentioned above feature (e).

Total pages: 40
29 30 Apr 2003
It is reasonable to consider that conditions ofuniform temperature and proper spatial arrange-ment of the ring structures in DNA may provide thepossibility for multiple photon entanglements inthe described above mechanism. It is known fromthe single mode narrow line emitting lasers thattheir coherence time is quite long. Let for exampleaccept that the coherence time in the DNA emittingprocess is in order of 1E-12 sec. If an average life-time for a single bond is about 1E-14 sec, then thetime between two ES clocks is 6E-14 sec (sixbonds). In this case, the entangled sequence will in-clude a binary code corresponding to 600 ESclocks. Such binary code corresponds to 600 bp(base pairs) and involves 200 codons.
Now let pay attention about one particularproblem related to DNA investigation, known as aC value paradox.
A substantial fraction of the genomes ofmany eukariotes is comprised of repetitive DNAin which short sequences are tandemly repeatedin small to huge arrays.
Tandemly repetitive sequences, known as“satellite” DNA’s are classified into three majorgroups;
-satellites - with repetitive lengths from oneto several thousands base pairs.
- minisatellites - repetitive arrays of 9 to 100bp, but usually 15 bp, generally involved in meanarrays lengths of 0.5 to 30 kb.
- microsatellites - repetitive arrays of short 2to 6 bp found in vertebrate, insect and plant ge-nomes.
The code of repetitive sequence does not en-code amino acids. The percentage of this non-in-formative DNA increases significantly withorganism complexity but depends also of other fac-tors. Among them are the living environment con-ditions of the species.
From a point of view of proposed hypothesis,the C value paradox obtains quite logical explana-tion:
(h) The repetitive sequences in DNA codeprovide repetitive synchronization code thatmay increase reliability of the DNA intercellularcommunication.
A large number of not discussed here diver-sified features of the DNA redundancy in different
organisms and species could find a logical explana-tion from the point of view of feature (h).
For example, the shown below repetitive se-quence in DNA from Drosophilia corresponds tothe following binary codes embedded in two entan-gled photons, related to the two strands:
(AATAT)n, where n - is number of repeatsfrom DNA strand 1: (00101)nfrom DNA strand 2: (11010)nIf the emitted entangled photon from strand 1
contains CW circular phase sequence of the code,the emitted entangled photon from strand 2 willcontain CCW circular phase sequence of the com-plimentary code.
It is evident that the entangled photons carryembedded code that is dependable of both: the ami-no acid code and the redundancy code. This de-pendence may eventually play a role in theimmunological response to transplanted tissues ororgans. The redundancy code greatly increases theprobability of successful synchronization. Then itis reasonable to expect a large abundance of repeat-able code near the end side of the linear DNA in eu-kariotic cells that is an observed fact.
10.6.2. Time sequence in the energy read-out process of DNA and its possible relation to the cell cycle synchronization.
Let consider some initial state of DNA mole-cule when all rings are charged with their normalenergies ES, E1 and E2. In some moment invokedby internal or external triggering, the stored ener-gies start to clock out with synchronization se-quence ES and two entangled photon sequences.Ones this process is started it will be self sustaineduntil all ES energies states are read-out. We maycall this effect a read-out process of DNA. Thetime duration (ttotal) for DNA read-out should be
(10)
where: tav - is the average lifetime of excited statesin the bonding path of DNA strand, 0.34 nm is thedistance between the rings of the neighbouringbase pairs.
The total read-out time is very short. ForDNA with length of 1 m and tav ~ 1E-14 s the read-out time is only 0.18 ms.
ttotal 6tavDNA length
0.34 nm-----------------------------≈

Total pages: 40
30 30 Apr 2003
After the DNA is read-out, all ES energies areemitted. The E1 energies only of the T and C areemitted, but the complementary state energies E1and E2 of A and G are preserved due to the attachedrings. This invokes some type of asymmetry be-tween connected Purines and Pyrimidines by H-bonds. Such conditions may help for separation ofthe DNA strands for initiating of the replicationprocess.
Now, let analyse the possible involvement ofDNA read-out process in the cell cycle synchroni-zation in eukariotes. We will not discuss here thecomplex processes of cell cycle regulation in whichthe proteins are involved, but only the conditions ofsuccessful triggering and read-out of DNA. Wemay assume only that every individual cell pos-sesses some kind of local triggering mechanism.Such mechanism is necessary but not enough con-dition in order to initiate the read-out process. TheDNA read-out can be successful only if conditionsfor emission of EM energy exist. Consequently itwill depend of the following conditional states ofDNA molecule:
(a) The two strands of one end of DNA mustbe separated (or unbalanced by promoter) in orderthe EM quanta (photons) to be emitted)
(b) The two strands (not counting the endconditions mentioned in (a)) must be completelysymmetrical in order the read-out process to beself-sustainable.
The requirements (a) and (b) excludes themRNA attachment to DNA or any other regulatoryprotein, so the transcription process and regulatorymechanisms should be completed. Then the opti-mum conditions for development of synchronizedread-out as an avalanche process should be thephase S (start of DNA replication).
Now let assuming that the internal triggeringmechanism provides triggering clocks with periodmuch shorter than the cell cycle. They will not pro-vide successful triggering until the conditions (a)and (b) are satisfied. However, once they are satis-fied the DNA read-out will be successful. Then theemitted entangled photons with a large commonenergy and encoded sequence possess an increasedprobability for activation of similar read-out proc-esses in the DNA of the neighbouring cells. The ex-tremely fast read-out process could lead to similarread-out process in many DNA. In such case, the
emitted entangled photons may additionally inter-fere and contribute to the avalanche process of thesynchronization. The avalanche process, however,will be contributed only of DNA molecules that donot have significant differences. In case of tissueand organ transplantation, the avalanche processwill not work between the DNA from different spe-cies whose synchronization sequences are too dif-ferent and even the not decoding pieces (related tothe C value paradox) might play an important role.
10.6.3. Environment considerations for the effi-ciency of the avalanche process
The phase accuracy of the cell cycle synchro-nization evidently depends of the individual cellcycle phases and environmental conditions.
10.6.3.1 Phase accuracy dependence of the cell cycle period
If considering the triggering of the synchroni-zation from internal cell mechanism in a properphase of the cell cycle, the period between two con-secutive synchronization bursts will depend of thetime lag between the local clock triggering events.Evidently the period of these events should bemuch smaller than the cell cycle period.
Let accept that the individual cell cycle regu-lation mechanisms provides a Gaussian type distri-bution of the cell cycle period from many cells.Then the optimum conditions for avalanche read-out should be expected in some moment that isclosed but not overpassing the maximum of theGaussian curve. Once it is initiated, the Gaussiancurve will be cut down for all cell contributing tothe avalanche. Those who had performed a pre-liminary read-out and those not activated by thesynchronization will not contribute. The cells inwhich the transcription and regulation processesare not completed in this moment will also be ex-cluded and their mitosis will not be in harmonywith the synchronized cells.
Additionally we may not expect that all thesynchronized cells will be activated in the very be-ginning of the read-out of the first activated cells.The repeatable sequences may provide conditionsfor larger number of entangled photons. Such pho-tons posses also an increased probability for ab-sorption and activation of similar repeats from

Total pages: 40
31 30 Apr 2003
DNA in other cells. Consequently the tandemlyrepetitive sequences of DNA increase the proba-bility for avalanche development of DNA read-out.
10.6.3.2 Physical factors of cell environmentThe most important environmental factor for
successful EM synchronization is the temperature.It is reasonable to assume that the cascade en-
ergy transfer through the bonding path of the DNAstrand is phase synchronized by the energy rotationcycle in the (O+5C) rings. The duration of this cy-cle is:
(11)where: t0 and tc are respectively the lifetimes of theexcited states in oxygen and carbon
The period, TS, of the synchronization pulsesis defined by the sum of the consecutive lifetimesthrough the bonding path of the strand, shown inFig. 13
(12)where: tav is the averaged lifetime value of
the bonding electrons involved in the bonding pathof the strand.
Then the condition for mentioned abovephase synchronization is expressed by the relation:
(13)The expression (13) is a constant for all in-
volved DNA molecules if the same energy levels ofthe excited states are involved. This is a quantummechanical condition that will depend only of thetemperature. The successful generation of entan-gled photons, however, is additionally dependentof the correct spatial arrangement of the (2N+4C)rings. If the DNA molecule is sharply bent or it isin proximity to a protein involving magnetic fieldasymmetry, the necessary conditions for genera-tion of multiple photon entanglement will be dis-turbed. This will affect the efficiency of thesynchronization process. Formations of Cruci-forms, for example, may affect the emitted se-quence. Z-type DNA inclusions and any externalinfluences causing a helical non-uniformity of Betatype DNA may also block the successful read-outprocess.
The synchronization is possible in limitedtemperature range and good temperature uni-formity along the DNA chain. The synchroniza-
tion efficiency is dependent of the bendingconditions of DNA and the spatial and helicaluniformity along its chain. External factorscausing any modification of the spatial parame-ters of the DNA double helix may inhibit or dis-turb the readout process.
10.7. Established features and experimentalresults supporting the hypothesis of the DNAinvolvement in the cell cycle synchronization
Many properties of the DNA molecules in eu-kariotes are in good agreement with the features ofDNA according to the provided analysis.
10.7.1 Absorption properties of DNA.DNA absorbs ultraviolet light in the range of
240 to 280 nm with a maximum at about 260 nm.Increasing the temperature destabilizes the doublehelix. Experimental observation leads to a conclu-sion that the thermal stability of DNA is a functionof base stacking, and not only of the hydrogenbonding13 (Saenger, 1984). This is in agreementwith the suggested mechanism of stabilizing mag-netic field due to the cascade energy flow throughthe bonding paths of the DNA strands.
The mechanism of energy store and releasesuggested by the presented hypothesis is in agree-ment with the observations, reported by Joseph La-kowiccz et al. (2001) in a paper14 “IntrinsicFluorescence from DNA can Be Enhanced by Me-tallic Particles”. They use silver particles of sizeabout 4 nm put in the surface of two quartz plateswith distance between them of 1 - 1.5 um. TheDNA placed in this gap has been excited at 287 nmby pulse sequences from a dye laser with a 100 pspulse with. The intrinsic emission following suchexcitation in the range from about 330 to 350 nm isincreased about 80 times around the metal parti-cles. The detected radiation they believed to befrom adenine and guanine.
The authors explain the effect by a decreasedlifetime in relation with the SERS effect (Surface-enhanced Ramman spectroscopy). Nevertheless,they also acknowledge the existence of another lessunderstood effect. The obtained results of time de-pendent intensity decay are shown in Fig. 21 (cor-responding to Fig 5 of their publication).
TR tO 5tC+=
TS tC 5tav+=
TS TR– t0 4tC 5tav–– const= =

Total pages: 40
32 30 Apr 2003
Fig. 21 Time-dependent intensity decay of DNA with-out metal (-) and between silver island film (Adapted from J.R. Lakowicz et al., 2001).
The explanation of the observed effect from a
point of view of BSM theory and suggested hy-pothesis is a following. The laser pulse charges theenergy storage rings of DNA. The silver metal is-lands introduce not uniformity field conditions inthe proximity of DNA. In such conditions, themechanism that may keep the stored energy in therings is disturbed. As a result, in the DNA portionsthat are in close proximity to the metal islands aspontaneous read-out process may occur sporadi-cally. This will appear in a shorter time interval be-tween the excited pulse and the fluorescenceemission in comparison to the case when DNA isnot in proximity with such particles. This corre-sponds to the observed decreased lifetime. It is in-teresting to note that the fluorescence lifetime isstill much longer than the lifetime in a single elec-tronic bond. This means that the stored energies inthe rings may exist not for one but but for numberof rotational cycles.
10.7.2 Increased binding ability of some simple organic molecules
The proteins that bind to DNA are usuallyvery large molecules containing thousands of at-oms. But some comparatively simple organic mol-ecules also have an increased ability for binding.When examining such molecules we see that theyposses attached atomic rings. As was mentionedbefore, the energy in such rings is more stable. Thisgives them additional ability to interact with DNAdue to magnetic field interactions. Such moleculesfor example are:
Trimethylpsoralen: 3 attached ringsEthidium Bromide: 3 attached rings + 1 singleChloroquine: 2 attached rings.
10.7.3 Effects of the salt concentration in helical winding and DNA supercoiling
As the concentration of a monovalent cati-on (Na+) or divalent cation (Mg+2) increases tohigh levels, the DNA double helix becomes woundless tightly.
According to the suggested hypothesis, theion concentration influences the ion currents in-volved in the stabilisation of the magnetic field ofDNA, responsible for its supercoiling shape.
10.7.4 Role of the intermolecular triplexes in the genetic recombination
Regions of triplex DNA formations havebeen identified near sites involved in genetic re-combinations. They usually have a mirror symme-try repeats of base pairs (as AGGAG). Kohwi andPanchenko15 (1993) found that the formation of in-tramolecular triplex structures in DNA in vivo caninduce genetic recombination between two directrepeats flanking the triplex formed sequence.
According to the proposed hypothesis, thetriplex formation in DNA disturbs the symmetry ofthe compensated field solenoid of DNA. Instead offlowing in the internal region of DNA double helix,some of the magnetic lines are forced to passthrough the external region. In conditions of chro-mosome recombination in vivo, the read-out proc-ess may be more often, than in the regulated cellcycle. Then the emitted sequence of entangled pho-tons from one chromosome could be directly in-duced into the other. This may influence theircommon position and orientation. In the same timethe magnetic field of the cascade energy transferthrough the bonding path of the DNA strand is notcompensated in the triplex region. When combin-ing with a similar region from another chromosomethe complex magnetic field may become compen-sated and stable.
10.7.5. Electronic properties of DNA This subject is of increased interest of phys-
icists and chemists. Despite the subject is far fromnew, it is very controversial. A recent overview pa-

Total pages: 40
33 30 Apr 2003
per about this topic is presented by C. Dekker16 inPhysics World, 2001. The experimental results de-viate from good insulator to good conductor, whilemany researchers consider that DNA is able to pro-vide some kind of energy flow.
Currently, two possible mechanisms ofcharge transfer are accepted: a coherent process ofsingle step electron tunnelling and a thermal hop-ing. It is interesting that a signature of cascadetransfer exists in both suggested processes. Whilethis theoretically suggested mechanisms seam rea-sonable, direct electrical measurements by numberof physics groups provide conflicting results. Theresults reported by Hans-Werner Fink and Chris-tian Schonenberger (1999), for 1 long DNA invacuum indicate a good conductivity17. However,this contradicts to the accepted theoretical consid-erations where DNA is expected to behave as asemiconductor with a large energy gap between thevalence and conduction bands. The experiments ofPorath and co-workers (2000), for example, on aparticular type of DNA - called poly(gG)-poly(dC)DNA and show that it behaves as a large gapsemiconductor18.
One important fact that may influence the ex-perimental results is the different environmentalconditions at which the experiments are performed.Additionally, in number of experiments the elec-tronic properties are not directly measured, but de-rivable from the transient absorption spectra.
Let try to explain the possible electrical prop-erties of DNA from the point of view of BSM mod-els. In order to have a stable energy cascadetransfer, the DNA must be in proper environments(pH, temperature, ATP) and to be enough long. Innormal environments, it is considered a negativelycharged. This charge, however, is uniformly dis-tributed, so it may not influence the energy cascadetransfer. In such case, the stored ring energies aresynchronized and have a proper handedness. Forsmall voltage along DNA, the electrons do not quittheir spatially distributed orbits. For large voltages,however, the proximity proton field defining theirorbits are biased, end the valence electron couldjump synchronously. This transition, however, isexpected to interact with the rotating energy statesin the rings. The signature of such interaction mightbe the effect of “single step electron-tunnellingmechanism”. It is evident that such conditions may
occur only if a stable energy cascade transferthrough the DNA strand exist, but the latter effectis possible only at proper environments.
11. Hypothesis of decoding process in some of the complexes aminoacyle-tRNA synthetases - tRNA.
11.1. General considerations and code analysisThe prototype of tRNA molecule is coded in
the gene. The current estimate for number of tRNAin animals and plants is up to 50. The typical shapeof the tRNA is shown in Fig. 16. The tRNA bindsto the proper enzyme that charges to its end one ofthe 20 amino acids corresponding to the anticodon.Aminoacyl-tRNA Synthetase is a family of 20 en-zymes whose structures are pretty well known. Theenzymes usually recognize the correspondingtRNA using the anticodon.
Let analyse the coding of the amino acidsbased on the anticodons used by tRNA. Instead ofanticodons we may use for convenience the mostpopular RNA codons, given by Table 12.1 (in ageneral case the anticodon code is directly obtaina-ble from the corresponding codon code). Theirnumber is 64. One exclusion from that rule is theanticodon of Alanine, where the three codes GCU,GCC and GCA are presented by one codon CGI ( I- Ionosine). This reduces the total number of anti-codons to 61.
One of the features of the amino acid coding,that is evident from the table, is the code redun-dancy. Most of the amino acids are coded by morethan one codon. One additional surprise comesfrom the genome analysis. Some organisms don’thave genes for all twenty aminoacyl-tRNA syn-thetases, but they still use all twenty amino acids tobuild their proteins.
The synthetase mechanism of the Aminoa-cyl-tRNA synthetases involves two steps. In thefirst step they form an aminoacyl-adenylate inwhich the carboxyl of the amino acid is linked tothe alpha-phosphate of ATP by displacing pyro-phosphate. In the second step, if only a correcttRNA is bound, the aminoacyl group of the ami-noacyle-adenylate is transferred to 2’ or 3’ terminalOH of the tRNA.
µm

Total pages: 40
34 30 Apr 2003
Table 12. 1
The second step is conditional. This meansthat if a wrong enzyme is bound to tRNA the out-put will be zero (not charging the proper amino ac-ids). If assuming that the process of binding thecorrect complex of enzyme - tRNA is occasional,then the number of possible combination for all 20amino acid codes is . This means suchnumber of produced tRNA and enzymes, so it isunreasonable. Another possible option is the en-zymes to recognize or at least to increase the prob-ability for binding to the correct tRNA in whichcase the above number could be significantly re-duced.
One of the currently discussed problems ishow these enzymes (aminoacyle-tRNA synthetas-es) recognize 20 different flavours. It is estimatedthat they admit intrinsically small number of errors- about 1 in 10,000. The unmistakeably recognitionof 20 different flavours looks as quite intelligenttask for a biomolecular structure of only few thou-sands atoms.
The above task could not be resolved unlessthe enzymes have some sensors for preliminary de-tection before binding and some memory. Havingin mind that the enzyme contains only a few thou-sands atoms it is apparent that such capabilityshould be built very economically using some basicphysical properties at atomic and molecular levels.Then the sensing should be based on some simpli-fied detection mechanism of energy states, while
the memory should be based on basic physicalstates of binary type. Let concentrate firstly on thememory feature. The possible binary physicalstates for a molecule of complexity of the enzymesare following:
(1) Quantum mechanical spin of the electron(2) direction of magnetic field (S-N or N-S)(3) handedness of rotating energy states (ac-
cording to BSM)(4) electrical charge ( + and -)The option (4) could be excluded because its
realization requires complex structure of semicon-ductor type. The most reasonable option is (3) withsome combination of options (1) and (2).
Let evaluate the required memory for decod-ing the codon of any amino acid using the Booleanalgebra without minimization. Any codon is athree-digit code. Anyone of these digits needs 4states in order to present one of the four letters(A,B,C,D). Then the three-digit code of the codoncould be presented of 12 bit binary code. This cor-responds to a memory map of 4096 bits. Suchmemory hardly be achieved by any enzyme whosemolecule usually includes a few thousands atoms.Additionally the enzyme must have remote sensorsfor recognition of tRNA before binding and toolsfor test of the anticodon.
Obviously, the above general approach fordecoding of 20 flavours is not feasible. We mustlook for some economic natural code hidden in the
UUUUUCUUAUUG
Phenylalanine
Leucine
UCUUCCUCAUCG
SerineUAUUACUAAUAG
Tyrosine
StopStop
UGUUGCUGAUGG
Cysteine
StopTryptophan
CUUCUCCUACUG
LeucineCCUCCCCCACCG
ProlineCAUCACCAACAG
Histidine
Glutamine
CGUCGCCGACGG
Arginine
AUUAUCAUAAUG
Isoleucine
Start; Methionine
ACUACCACAACG
ThreonineAAUAACAAAAAG
Asparagine
Lysine
AGUAGCAGAAGG
Serine
Arginine
GUUGUCGUAGUG
ValineGCUGCCGCAGCG
AlanineGAUGACGAAGAG
Aspartic acidGlutamicacid
GGUGGCGGAGGG
Glycine
220 1048576=
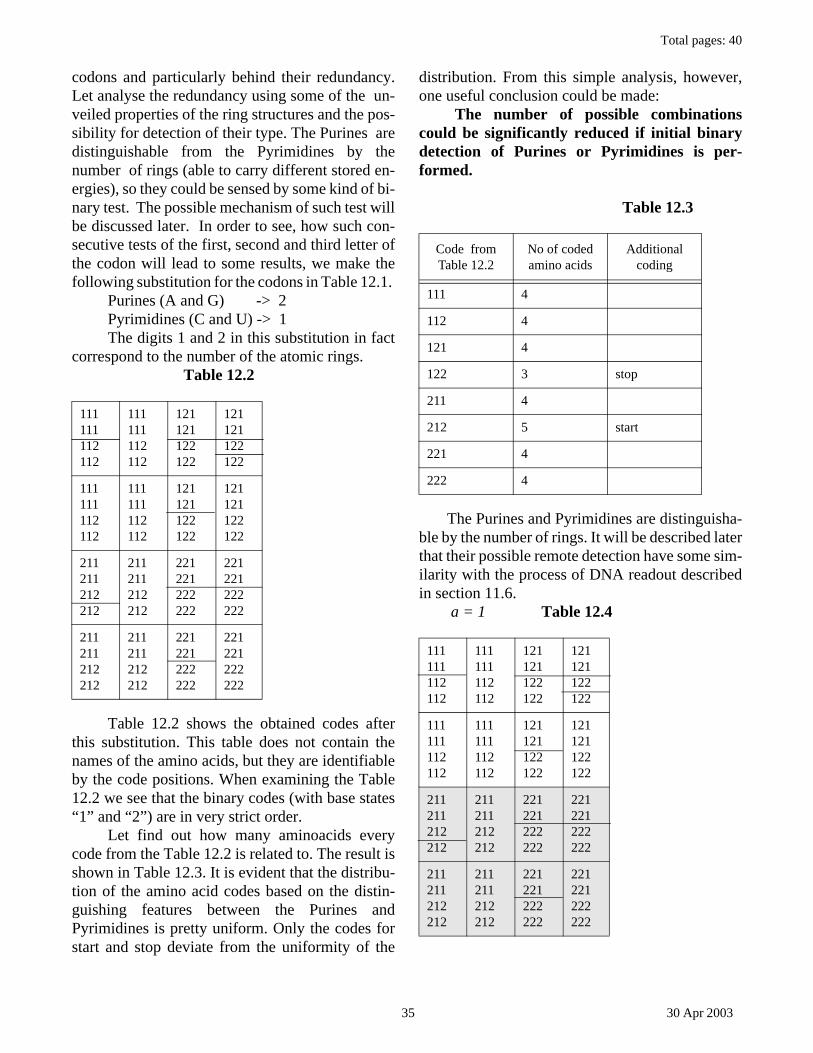
Total pages: 40
35 30 Apr 2003
codons and particularly behind their redundancy.Let analyse the redundancy using some of the un-veiled properties of the ring structures and the pos-sibility for detection of their type. The Purines aredistinguishable from the Pyrimidines by thenumber of rings (able to carry different stored en-ergies), so they could be sensed by some kind of bi-nary test. The possible mechanism of such test willbe discussed later. In order to see, how such con-secutive tests of the first, second and third letter ofthe codon will lead to some results, we make thefollowing substitution for the codons in Table 12.1.
Purines (A and G) -> 2Pyrimidines (C and U) -> 1The digits 1 and 2 in this substitution in fact
correspond to the number of the atomic rings. Table 12.2
Table 12.2 shows the obtained codes afterthis substitution. This table does not contain thenames of the amino acids, but they are identifiableby the code positions. When examining the Table12.2 we see that the binary codes (with base states“1” and “2”) are in very strict order.
Let find out how many aminoacids everycode from the Table 12.2 is related to. The result isshown in Table 12.3. It is evident that the distribu-tion of the amino acid codes based on the distin-guishing features between the Purines andPyrimidines is pretty uniform. Only the codes forstart and stop deviate from the uniformity of the
distribution. From this simple analysis, however,one useful conclusion could be made:
The number of possible combinationscould be significantly reduced if initial binarydetection of Purines or Pyrimidines is per-formed.
Table 12.3
The Purines and Pyrimidines are distinguisha-ble by the number of rings. It will be described laterthat their possible remote detection have some sim-ilarity with the process of DNA readout describedin section 11.6. a = 1 Table 12.4
111111112112
111111112112
121121122122
121121122122
111111112112
111111112112
121121122122
121121122122
211211212212
211211212212
221221222222
221221222222
211211212212
211211212212
221221222222
221221222222
Code from Table 12.2
No of coded amino acids
Additional coding
111 4
112 4
121 4
122 3 stop
211 4
212 5 start
221 4
222 4
111111112112
111111112112
121121122122
121121122122
111111112112
111111112112
121121122122
121121122122
211211212212
211211212212
221221222222
221221222222
211211212212
211211212212
221221222222
221221222222

Total pages: 40
36 30 Apr 2003
Let denote the tRNA anticodon by the tripletabc, but using instead the corresponding codons ofRNA. The following example shows the reductionof the possible combinations in three consecutivebinary tests (Purines or Pyrimidines) applied forthe codes of Table 12.2. Let the first test for a givesvalue: a = 1. This means that all codons startingwith 2 are excluded from the following tests, whilethose starting with 1 are enabled. Only 32 from ofall 64 combinations are left. This is illustrated inTable 12.4., where the excluded combinations aremasked by a grey colour.
Let apply the same test for b and the result forexample is b = 2. The available combinations areadditionally reduced in half as shown in Table12.5.
b = 2 Table 12.5
In a similar way the third test of c, for exam-ple, with a result c = 2 will lead to reduction of theavailable combinations to 8 as shown in Table12.6.
c = 2 Table 12.6
We see that by three consecutive tests theavailable combinations are reduced from 64 to 8,while they address only 4 amino acids. If assumingthat this test is performed prior to binding of the en-zyme to the tRNA, then the required combinationsfor a probable correct match are only . Con-sequently a possible remote sensing will reduce theamount of the necessary number of enzymes andtRNA by a factor of 65536.
The additional tests for identification of thecorrect amino acids require distinguishing of Ade-nine from Uracile and Guanine from Cytozine forevery letter of the triplet abc. The detection forsuch identification, however, could not be remoteand perhaps is performed in the second step of thesynthetase mechanism. It is known, for example,that in glutaminyl-tRNA synthetase with its tRNAthe enzyme firmly grips the anticodon, spreadingthe three bases widely apart. Let assuming that theenzyme keeps in its memory the binary results ofthe previous remote detection (Purines or Pyrimi-dines) for every letter. Then the following test afterthe enzyme has grabbed the anticodon (for everyletter of the codon) must be also a binary test of atype: [Adenine (A) or Guanine (G)] and [Cytozine(C) or Uracile (U)]. How could they be distin-guished, while having the same number of rings?When examining their structure we see that thelarger ring of Adenine includes three bonds of sin-
111111112112
111111112112
121121122122
121121122122
111111112112
111111112112
121121122122
121121122122
211211212212
211211212212
221221222222
221221222222
211211212212
211211212212
221221222222
221221222222
111111112112
111111112112
122122
122122
111111112112
111111112112
122122
122122
211211212212
211211212212
221221222222
221221222222
211211212212
211211212212
221221222222
221221222222
24 16=
121 121121 121
121 121121 121

Total pages: 40
37 30 Apr 2003
gle valence, while the larger ring of Guanine in-cludes four bonds of single valence. The ring ofCytosine includes two bonds of second valence,while the ring of Uracile includes one bond of sec-ond valence. These differences of the ring struc-tures may cause significant differences in theirspectral emission-absorption properties. The verynarrow temperature range will define a pretty nar-row range of the population of the excited states.This may simplify the task for recognizing the dif-ferences between (A and G) and (C and U) by theirspectral signatures.
After all these binary tests are performed, itseems that the correct amino acid should be decod-ed. However, we must not forget that the letters abcmust be read in a correct order (from the most to theless significant digit). If this rule is not observed,the following code pairs could not be distin-guished:
UUA - AUUUUC - CUUUUG - GUUUCC - CCUCCA - ACCAAG - GAAUGG - GGUAssuming that the memory map of the en-
zyme is naturally minimized, the correct code rea-dout obviously must be initially detected prior tothe remote sensing described above. In order tofind out the possible detection mechanism for thecorrect code readout direction, we may examinethe structure of tRNA that has been already shownin Fig. 16. In the left side of the figure, the realshape of the backbone structure is shown. In theright side of the same figure, the tRNA is presentedlike a flat curve in order to show more clearly someof its structural features such as, the folding of thesingle strand in cloverleaf shape, the position ofloops, the H-bonds, the position of the anticodonand the asymmetrical ends.
The tRNA strand is similar as the DNA strandand contains regularly attached (O+4C) rings. Inproper environment the tRNA could be energized,so a cascade energy transfer may occur through itsstrand in a similar way as in the DNA and proteins.The asymmetrically terminated end of tRNA mayassure the proper direction of the cascade energy
transfer. If such process is stable enough for a shorttime, the rotating energies in (O+4C) rings can besynchronized, obtaining in such way a properhandedness. The same is valid for the H-bondingbase pairs and consequently for the codon loop.While the tRNA is not so long (75 - 90 nucleotides)the energy cascade process could not be so stablelike in the DNA and the proteins. The disruption ofthe energy cascade will lead to synchronized read-out of the rotating energy states in (O+4C) ringsthat could be emitted as a coherent sequence ofphoton pulse whose parameters (phase sequence)will carry the direction of codon readout. This re-adout of (O+4C) energies from its side might pro-voke a readout process from the base pairs in asimilar way as in the DNA. The conditions foremission from tRNA, however, are different fromthose of DNA due to the different shape of thetRNA molecule. Let assuming a hypothetical axis(not exactly linear), defined by the twisting shape(known as a secondary structure) of tRNA andpassing from the anticodon loop of tRNA to its ter-minating ends. The two major side loops of tRNAcould be regarded as approximately symmetricalpairs in respect to the introduced axis. The super-coiling and tertiary structure may also influence thesymmetry of the mentioned pairs, but will not in-fluence the symmetrical features of the anticodonloop. In the same time, all the loops are free of H-bonds. If regarding the emission process as EMfiled, it is evident that the two side loops are com-plimentary and will have a comparatively smallerEM emission capability (because the magneticlines appears closed in a circle around the intro-duced tRNA axis). The anticodon loop, however,does not have complimentary symmetry withanother loop, so its emission capability could bemuch larger. Then the emitted photons from theanticodon loop could be detected remotely by thecorrect enzyme. The burst of photon sequence from(O+4C) rings readout may serve as a synchroniza-tion that prepares the enzyme for detection of theanticodon. In such process the detection could be ofsynchronized type, so it could allow an increasedprobability for detection of the anticodon sequence.The tRNA could be re-energize and the readout se-quence could be performed a few times. In suchway, the anticodon could be detected by number ofcorresponding enzymes.

Total pages: 40
38 30 Apr 2003
The remotely detected sequence from antico-don loop (carrying information for Purines or Pyri-midines) could be regarded as very fast consecutivetests of the letters in the triplet abc prior to bindingof the correct enzyme to tRNA. The test results,however, will be additionally needed for the con-secutive tests (A or G and C or U) that will be per-formed after the binding of the enzyme to thetRNA. Let assuming that these test results are di-rectly passed to some kind of binary decoder im-plemented in the enzyme. Then after this remotetest the number of codons from 64 is reduced to 8,corresponding to 4 amino acids. This significantlyincreases the probability of correct binding be-tween tRNA and the corresponding enzyme.
11.2 Decoding algorithmFig. 22 presents the decoding algorithm ac-
cording to the analysis and considerations dis-cussed in the previous paragraph.
The remote sensing includes tests 1, 2, 3,(shown in the figure) while the other tests are per-formed after the tRNA is bound to the correct en-zyme. The numbers in circle show the enabledcodon combinations after each test. Not all the de-coding tree for all 20 aminoacides is shown, but thesuggested possible algorithm for their decoding isthe same. It is evident, that maximum of 6 tests arenecessary for decoding of anyone of the aminoac-ids, but some of them are decoded even at test No5. (Leucine, for example). Two of the stop codonsare also decoded at test No. 5. This provides theopportunity to use the test No. 6 for additional truetest, that should increase the decoding reliability.The other stop codon UGA, however is decoded af-ter the test No. 6. Then it could be interesting tostudy the statistics of UAA and UAG stop codonsin comparison with the UGA stop codon (in boththe RNA and DNA sequences).
According to the suggested algorithm, thetests from the remote sensing increase the probabil-ity for correct binding between tRNA and the prop-er enzyme, but does not exclude completely wrongbindings. It is logical to expect that only a correctbinding will lead to attachment of amino acid afterthe tests 3, 4 and 5. The wrong binding will providea zero result.
Discussion. The first version of the theoretical analysis
of the biomolecules with suggested threehypotheses19 has been archived in the National Li-brary of Canada in 2002. The reader of this articlewho has not been acquainted with the BSM theorymay put one reasonable question: Why the suggest-ed atomic models, pretending to match the physicalreality, are not widely discussed and promoted bythe physical society? The answer is: The alternativeconcept of the vacuum space is a fundamentalchange in physics that may affect a number of high-ly abstracted and mathematically sophisticated the-ories. This may cause a process of adaptation thatcould probably take years. In the fields of structuralchemistry, organic molecules and nanotechnolo-gy, however, the suggested models of the atoms,could be tested and applied right away because thecriterion for their validation comes simultaneouslywith their applications. The large data base of or-ganic and biomolecules with known structure andatomic composition provides an excellent opportu-nity for reliable test and validation.
The Quantum mechanical models of the at-oms are very useful for their ability to provide ac-curate calculation of the energy levels andinteraction probabilities. The models, suggested byBSM thesis, do not intend to replace or underminethe useful features of the quantum mechanicalmodels but to enrich the knowledge about the phys-ical structures of the atoms and the quantum proc-esses at subatomic level.

Total pages: 40
39 30 Apr 2003
Fig. 22. Decoding algorithm according to the hypothesis of decoding process in some of the complexes tRNA - enzyme.

References: 1. S. Sarg, Basic Structures of Matter, monograph, (2001), http://www.helical-structures.org also in: http://collection.nlc-bnc.ca/amicus/index-e.html (AMICUS No. 27105955) 2. T. H. Boyer, The Classical Vacuum, Scientific American, Aug. 1985, p. 70-78. 3. H. E. Puthoff, Can the Vacuum be Engineered for Spaceflight applications, NASA Breakthrough Propulsion Physics, conference at Lewis Res. Center, 1977 4. S. Sarg, Brief introduction to Basic Structures of Matter theory and derived atomic models, Journal of Theoretics, 2003 www.journaloftheoretics.com/Links/Papers/Sarg.pdf 5. H. E. Puthoff, Gravity as a zero-point-fluctuation force, Physical review A, v. 39, No 5, 1989, p. 2333-2342. 6. S. Sarg, Atlas of Atomic Nuclear Structures according to Basic Structures of Matter theory, Journal of theoretics, 2003 www.journaloftheoretics.com/Links/Papers/Sarg2.pdf 7 S. Sarg, Atlas of Atomic Nuclear Structures (2001), National Library of Canada. http://collection.nlc-bnc.ca/amicus/index-e.html (AMICUS No. 27106037) also in http://www.helical-structures.org 8. I. Dabrowski, Can. J. Phys. 62, 1639 (1984) 9. C. B. Anfinsen, J, T. Edsall and F. M. Richards, Advances in protein chemistry, vol. 34, 1981, Academic Press. 10. W. M. Gelbart, R. F. Bruinsma, P. A. Pincus and V. A. Parsegian, DNA-inspired electrostatics, Physics Today, 38-44, September (2000). 11. D. Bouwmeester et al, Observation of Three-Photon Greenberger-Horne_Zeilinger Entaglement, Phys. Rev. Lett., v. 82, No 7, 1345-1349, (1991) 12. A. Lamas-Linares, J. C. Howell & D. Bouwmeester, Stimulated emission of polarization-entangled photons, Nature, vol. 412, 887-890, (2001). 13. W. Saenger, Principles of Nucleic Acid Structure, Springer-Verlab, Ney York, (1984) 14.J. R. Lakowicz, B. Shen, Z. Gryczynski, S. D’Auria and I. Gryczynski, Intrinsic fluorescence from DNA can be enhanced by metalic particles, Biochemical and Biophysical Res. Comm., 286, 875-879 (2001) 15. Y. Kohwi and Y. Panchenko, Transcription-dependent recombination induced by triple-helix formation. Genes Dev. 7, 1766-1778, (1993) 16. C. Dekker, Electronic properties of DNA, Physics World, Aug. 2001 http://physicsweb.org/article/world/14/8/8 17. H-W Fink and C. Schonenberger, Electrical conduction through DNA molecule, Nature, 398, 407, (1999) 18. D. Porath et al., Direct measurement of electrical transport through DNA molecules, Nature, 403, 635, (2000) 19. S. Sarg, Theoretical analysis of biomolecules using BSM models (2002), archived in the National Library of Canada, http://collection.nlc-bnc.ca/amicus/index-e.html AMICUS No. 27749841







![Journal of Mechanics of Materials and Structures - MSP · JOURNAL OF MECHANICS OF MATERIALS AND STRUCTURES ... Lee–Tarver ignition and growth model, ... [Tarver and Hallquist 1981]](https://img.dokumen.tips/doc/110x75/5b5b85047f8b9ac7498e4557/journal-of-mechanics-of-materials-and-structures-msp-journal-of-mechanics.jpg)





![Journal of Theoretics · Journal of Theoretics On Newton's Paradoxes Carlo Santagata carlo.santagata@tin.it Abstract: Reading the Principia [1] it is easy to realize that Newton,](https://img.dokumen.tips/doc/110x75/5caba87088c993453d8d8560/journal-of-journal-of-theoretics-on-newtons-paradoxes-carlo-santagata-carlosantagatatinit.jpg)















