Embed Size (px)
Citation preview

Joe Mychaleckyj
Slide 1
Linkage Disequilibrium
Joe MychaleckyjCenter for Public Health
Genomics982-1107

Slide 2
Joe Mychaleckyj
Today we’ll cover…
• Haplotypes• Linkage Disequilibrium• Visualizing LD• HapMap

Slide 3
Joe Mychaleckyj
References
Principles of Population Genetics, Fourth Edition (Hardcover) by Daniel L. Hartl, Andrew G. Clark (Author)
xx
x
Genetic Data Analysis II Bruce S WeirQuickTime™ and a
TIFF (Uncompressed) decompressorare needed to see this picture.

Slide 4
Joe Mychaleckyj
References
Statistical Genetics: Gene Mapping Through Linkage and Association Eds Benjamin M. Neale, Manuel A.R. Ferreira, Sarah E. Medland, Danielle Posthuma

Slide 5
Joe Mychaleckyj
SNP1 SNP2 SNP3
[A / T] [C / G] [A / G]
A C G
A C A
T G G
2N (ie very large diversity possible)
Haplotype: specific combination of alleles occurring (cis) on the same chromosome (segment of chromosome)
N SNPs - How many Haplotypes are possible ?

Slide 6
Joe Mychaleckyj
Terminology
• Haplotype: Specific combination (phasing) of alleles occurring (cis) on the same chromosomal segment
• Linkage/Linked Markers: Physical co-location of markers on the same chromosome
• Diplotype: Haplogenotype ie pair of phased haplotypes one maternally, one paternally inherited

Slide 7
Joe Mychaleckyj
SNP2 [ B / b ]SNP1 [ A / a ]
Major Allele Freq: p(A) p(B)
Minor Allele Freq: p(a) p(b)
Independently segregating SNPs:
Haplotype Frequency p(ab) = p(a) x p(b)
LINKAGE DISEQUILIBRIUM
Haplotype Frequency p(ab)≠ p(a) x p(b)
LINKAGE EQUILIBRIUM
(How many haplotypes in total ?)

Slide 8
Joe Mychaleckyj
Linkage Disequilibrium
• Non-random assortment of alleles at 2 (or more) loci
• The closer the markers, the stronger the LD since recombination will have occurred at a low rate
• Markers co-segregate within and between families
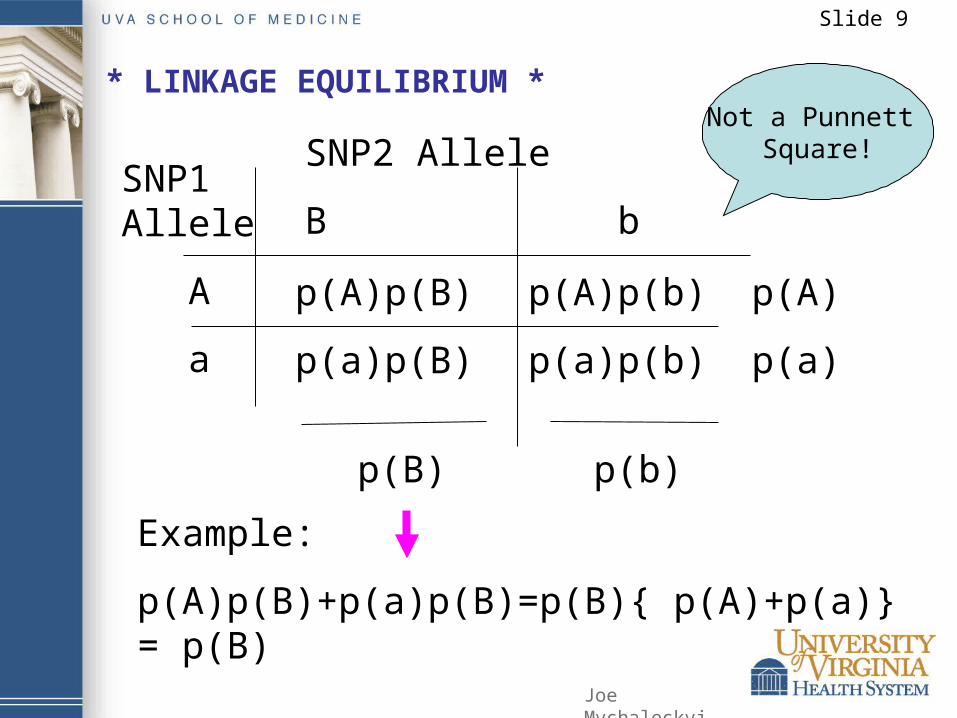
Slide 9
Joe Mychaleckyj
SNP1 Allele
A
a
SNP2 Allele
B b
p(A)p(B)
p(a)p(B)
p(A)p(b) p(A)
p(a)p(b) p(a)
p(B) p(b)
Example:
p(A)p(B)+p(a)p(B)=p(B){ p(A)+p(a)} = p(B)
* LINKAGE EQUILIBRIUM *Not a Punnett
Square!

Slide 10
Joe Mychaleckyj
SNP2 [ B / b ]SNP1 [ A / a ]
Major Allele Freq: p(A) p(B)
Minor Allele Freq: p(a) p(b)
LINKAGE DISEQUILIBRIUM
Haplotype Frequency p(ab) = p(a) p(b) + D
(sign of D is generally arbitrary, unless comparing D values between populations or studies)
D: Lewontin’s LD Parameter (Lewontin 1960)

Slide 11
Joe Mychaleckyj
SNP1 Allele
A
a
SNP2 Allele
B b
p(A)p(B)+D
p(a)p(B)-D
p(A)p(b)-D p(A)
p(a)p(b)+D p(a)
p(B) p(b)
p(A)p(B)+D + p(a)p(B)-D =p(B){ p(A)+p(a)} = p(B)
* LINKAGE DISEQUILIBRIUM *

Slide 12
Joe Mychaleckyj
0.16 0.04
0.14 0.66
a
A
b B
p(a)=0.20
p(B)=0.80
p(b)=0.30 p(B)=0.70
What is the LD ?
≠ 0
p(ab) ≠ p(a) p(b)
p(ab) = p(a) p(b) + D
0.16 = 0.2 x 0.3 + D
D = 0.1
Since p(ab) = p(a)p(b)+ D
+D was used and D is +ve here, but arbitrary
eg can relabel alleles A,B as minor

Slide 13
Joe Mychaleckyj
Range of D values (-ve to +ve)
D has a minimum and maximum value that depends on the allele frequencies of the markers
Since haplotype frequencies cannot be -ve
p(aB) = p(a)p(B) - D ≥ 0 D ≤ p(a)p(B)
p(Ab) = p(A)p(b) - D ≥ 0 D ≤ p(A)p(b)
These cannot both be true, so D ≤ min( p(a)p(B), p(A)p(b) )
p(ab) = p(a)p(b) + D ≥ 0 D ≥ -p(a)p(b)
p(AB) = p(A)p(B) + D ≥ 0 D ≥ -p(A)p(B)
These cannot both be true, so D ≥ max( -p(a)p(b), -p(A)p(B) )
* Similar equations if we had defined p(ab) = p(a)p(b) - D

Slide 14
Joe Mychaleckyj
Limits of D LD Parameter
Limits of D are a function of allele frequencies
Standardize D by rescaling to a proportion of its maximal value for the given allele frequencies (D') D’ = D
Dmax

Slide 15
Joe Mychaleckyj
D’ (Lewontin, 1964)
D’ = D / Dmax
Dmax = min (p(A)p(B), p(a)p(b)) D < 0
Dmax = min (p(A)p(b), p(a)p(B)) D > 0Again, sign of D’ depends on definition
D’ = 1 or -1 if one of p(A)p(B), p(A)p(b), p(a)p(B), p(a)p(b) = 0
= Complete LD (ie only 3 haplotypes seen)D’=1 or -1 suggests that no recombination has
taken place between markersBeware rare markers - may not have enough
power/sample size to detect 4th haplotype

Slide 16
Joe Mychaleckyj
D’ Interpretation
0.06 0.14
0.24 0.56
a
A
b B
p(a)=0.20p(A)=0.80
p(b)=0.30 p(B)=0.70
0.2 0
0.1 0.7
a
A
b B
p(a)=0.20P(A)=0.80
p(b)=0.30 p(B)=0.70
D=0 ; Dmax undefined D=Dmax =0.14 ; D’ = +1
p(a) = 0.2
p(b)= 0.3
D’=1 (perfect LD using D’ measure - No recombination between marker - Only 3 haplotypes are seen

Slide 17
Joe Mychaleckyj
Creation of LD
• Easiest to understand when markers are physically linked
• Creation of LD– Mutation– Founder effect– Admixture– Inbreeding / non-random mating– Selection– Population bottleneck or stratification– Epistatic interaction
• LD can occur between unlinked markers• Gametic phase disequilibrium is a more
general term

Slide 18
Joe Mychaleckyj
A
a
A B
A b
a B
SNP1
SNP1
SNP2
Recombinationn=2 haplotypesn=2 haplotypes
n=3 haplotypesn=3 haplotypes
SNP1
SNP2
A B
A b
a B
a b
n=4 haplotypesn=4 haplotypes

Slide 19
Joe Mychaleckyj
Destruction of LD
• Main force is recombination • Gene conversion may also act at
short distances (~ 100-1,000 bases)
• LD decays over time (generations of interbreeding)

Slide 20
Joe Mychaleckyj
Initial LD between SNP1 - SNP2: D0
After 1 generation
Preservation of LD:D1 = D0(1-θ)
After t generations:Dt = D0 (1- θ)t
SNP1 SNP2 Probability Recombination occurs = θ
Probability Recombination does not occur = 1-θ
NB: Overly simple model - does not account for allele frequency drift over time

Slide 21
Joe Mychaleckyj
QuickTime™ and aTIFF (Uncompressed) decompressor
are needed to see this picture.
Dt = D0 (1-θ)t

Slide 22
Joe Mychaleckyj
r2 LD Parameter (Hill & Robertson, 1968)
• Squared correlation coefficient varies 0 - 1
• Frequency dependent• Better LD measure for allele correlation
between markers - predictive power of SNP1 alleles for those at SNP2
• Used extensively in disease gene or phenotype mapping through association testing
r2 = D2
p(a)p(b)p(A)p(B)

Slide 23
Joe Mychaleckyj
r2 Interpretation
0.06 0.14
0.24 0.56
a
A
b B
p(a)=0.20p(A)=0.80
p(b)=0.30 p(B)=0.70
0.2 0
0.1 0.7
a
A
b B
D=0 ; Dmax undefined D=Dmax =0.14 ; D’ = +1
r2 = 0 r2 = 0.14/0.24 = 0.58
p(a) = 0.2
p(b) = 0.3r2 ≠ 1 Correlation is not perfect, even
though D’ = 1
r2 = 1 if D’ = 1 and p(a) = p(b) = 0.3
p(a)=0.20p(A)=0.80
p(b)=0.30 p(B)=0.70

Slide 24
Joe Mychaleckyj
r2 Interpretationp(a) = 0.3
p(b) = 0.3Only 2 haplotypes:
r2 = 1 Correlation is perfect
D’ =1 (less than 4 haplotypes)
p(a) = p(b) (= 0.3 in this example)
• r2=1 when there is perfect correlation between markers and one genotype predicts the other exactly
– Only 2 haplotypes present
• D’ = 1 ≠> r2 = 1• No recombination AND markers must have
identical allele frequency– SNPs are of similar age
• Corollary– Low r2 values do not necessarily = high recombination– Discrepant allele frequencies

Slide 25
Joe Mychaleckyj
-1 D’ 1
0 r2 1
Common Measures of Linkage Disequilibrium
Recombination
Correlation
Other LD Measures exist, less common usage

Joe Mychaleckyj
Slide 26
Visualizing LD metrics

Slide 27
Joe Mychaleckyj
SNP1
SNP2
SNP3
SNP4
SNP5
SNP6
SNP1 2 3 4 5 6
0.2
0.6
0.8
1.0
0
| D’ |
Not usually worried about sign of D’

Slide 28
Joe Mychaleckyj

Slide 29
Joe Mychaleckyj
Haploview: TCN2 (r2)

Slide 30
Joe Mychaleckyj
Launched October 2002
http://www.hapmap.org

Slide 31
Joe Mychaleckyj
International HapMap Project• Initiated Oct 2002• Collaboration of scientists worldwide• Goal: describe common patterns of human
DNA sequence variation• Identify LD and haplotype distributions• Populations of different ancestry
(European, African, Asian)– Identify common haplotypes and population-specific differences
• Has had major impact on:– Understanding of human popualtion history as reflected in genetic
diversity and similarity– Design and analysis of genetic association studies

Slide 32
Joe Mychaleckyj
HapMap samples
• 90 Yoruba individuals (30 parent-parent-offspring trios) from Ibadan, Nigeria (YRI)
• 90 individuals (30 trios) of European descent from Utah (CEU)
• 45 Han Chinese individuals from Beijing (CHB)
• 44 Japanese individuals from Tokyo (JPT)

Slide 33
Joe Mychaleckyj
Project feasible because of:
• The availability of the human genome sequence• Databases of common SNPs (subsequently
enriched by HapMap) from which genotyping assays could be designed
• Development of inexpensive, accurate technologies for highthroughput SNP genotyping
• Web-based tools for storing and sharing data• Frameworks to address associated ethical and
cultural issues

Slide 34
Joe Mychaleckyj
HapMap goals
• Define patterns of genetic variation across human genome
• Guide selection of SNPs efficiently to “tag” common variants
• Public release of all data (assays, genotypes)• Phase I: 1.3 M markers in 269 people
1 SNP/5kb (1.3M markers)
Minor allele frequency (MAF) >5%
• Phase II: +2.8 M markers in 270 people

Slide 35
Joe Mychaleckyj
http://www.hapmap.org/

Slide 36
Joe Mychaleckyj
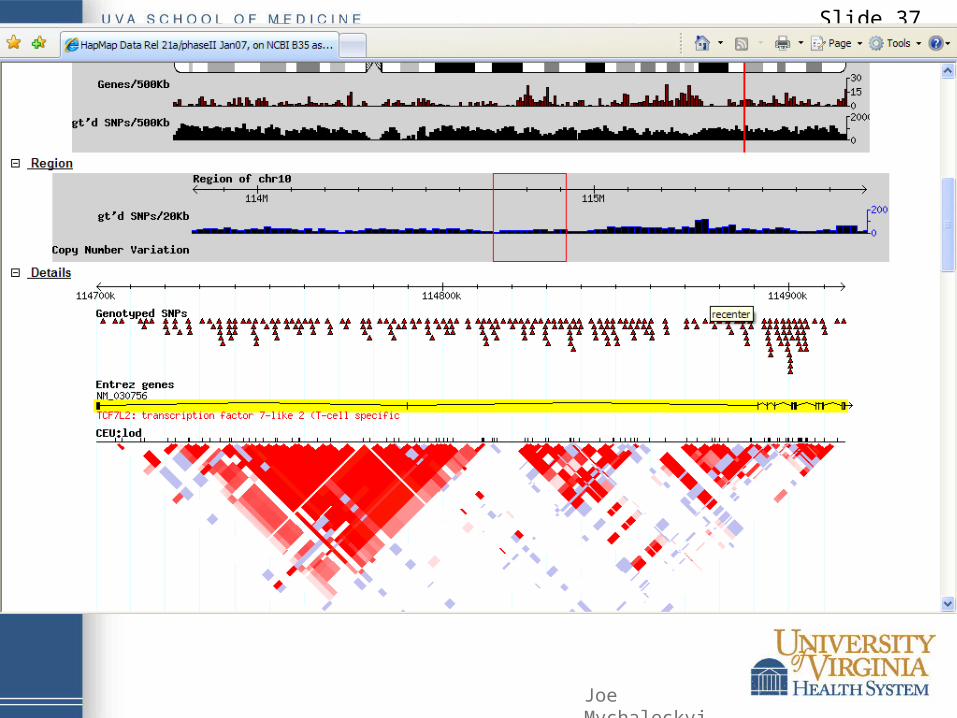
Slide 37
Joe Mychaleckyj

Slide 38
Joe Mychaleckyj
HapMap publications
• The International HapMap Consortium. A Haplotype Map of the Human Genome. Nature 437, 1299-1320. 2005.
• The International HapMap Consortium. The International HapMap Project. Nature 426, 789-796. 2003.
• The International HapMap Consortium. Integrating Ethics and Science in the International HapMap Project. Nature Reviews Genetics 5, 467 -475. 2004.
• Thorisson, G.A., Smith, A.V., Krishnan, L., and Stein, L.D. The International HapMap Project Web site. Genome Research,15:1591-1593. 2005.

Slide 39
Joe Mychaleckyj
ENCODE project
• Aim: To compare the genome-wide resource to a more complete database of common variation—one in which all common SNPs and many rarer ones have been discovered and tested
• Selected a representative collection of ten regions, each 500 kb in length
• Each 500-kb region was sequenced in 48 individuals, and all SNPs in these regions (discovered or in dbSNP) were genotyped in the complete set of 269 DNA samples

Slide 40
Joe Mychaleckyj
Comparison of linkage disequilibrium and recombination for two ENCODE regions
Nature 437, 1299-1320. 2005

Joe Mychaleckyj
Slide 41
LD in Human Populations

Slide 42
Joe Mychaleckyj
Haplotype Blocks
N SNPs = 2N Haplotypes possible, ie very large diversity possible
But: we do not see the full extent of haplotype diversity in human populations
Extensive LD especially at short distances eg ~20kbases.
Haplotypes are broken into blocks of markers with high mutual LD separated by recombination hotspots
Non-uniform LD across genome

Slide 43
Joe Mychaleckyj
Haplotype Blocks
Haplotype blocks: at least 80% of observed haplotypes with frequency >= 5% could be grouped into common patterns
Whole Genome Patterns of Common DNA Variation in Three Human Populations, Science 2005, Hinds et al.

Slide 44
Joe Mychaleckyj
Length of LD spans
r2

Slide 45
Joe Mychaleckyj
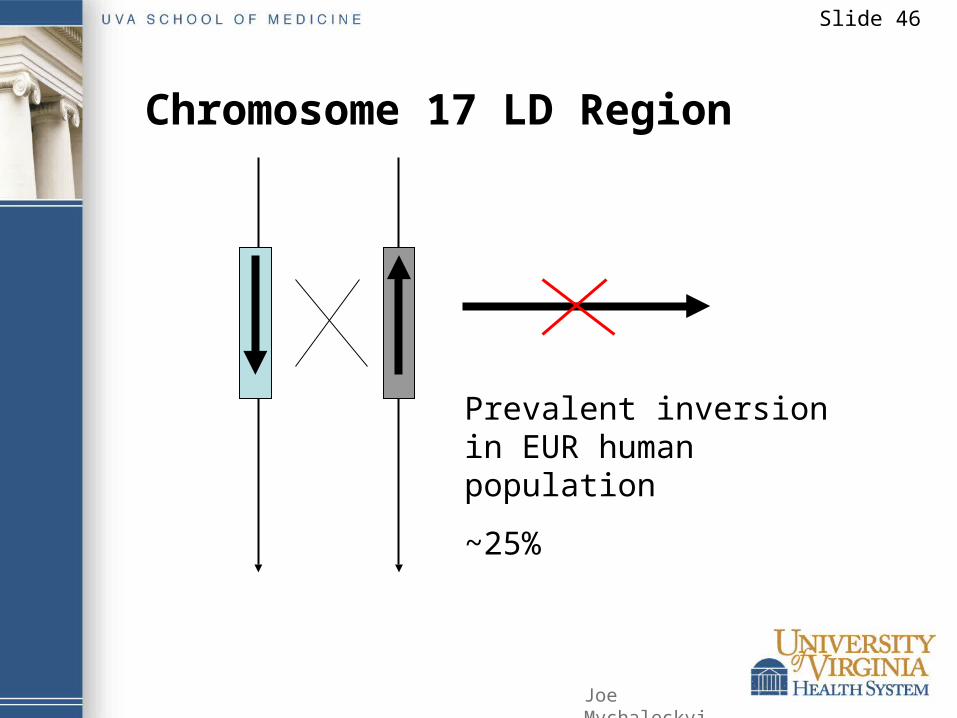
Example: Large block of LD on chromosome 17Cluster of common (frequent SNPs In high LD)518 SNPs, spanning 800 kb25% in EUR, 9% in AFR, missing in CHNGenes:
Microtubule-associated protein tauMutations associated with a variety of neurodegeneartive disordersGene coding for a protease similar to presenilinsMutations result in Alzheimer’s diseaseGene for corticotropin-releasing hormone receptor
• Immune, endocrine, autonomic, behavioral response to stress
Slide 46
Joe Mychaleckyj
Chromosome 17 LD Region
Prevalent inversion in EUR human population
~25%





























