Embed Size (px)
Citation preview

Research ArticleJin Fu Kang Oral Liquid Inhibits Lymphatic Endothelial CellsFormation and Migration
Hai-Lang He1 Dan Wang1 Jie Tang1 Xian-Mei Zhou1 Jian-Xin Li2 and Ling Xu34
1Department of Respiratory Medicine Affiliated Jiangsu Province Hospital of Traditional Chinese MedicineNanjing University of Chinese Medicine Nanjing 210029 China2State Key Laboratory of Analytical Chemistry for Life Science and Collaborative Innovation Center of Chemistry for Life SciencesSchool of Chemistry and Chemical Engineering Nanjing University Nanjing 210023 China3Tumor Institute of Traditional Chinese Medicine Shanghai University of Traditional Chinese Medicine725 South Wanping Road Shanghai 200032 China4Department of Oncology Yueyang Hospital of Integrated Traditional Chinese and Western MedicineShanghai University of Traditional Chinese Medicine 110 Ganhe Road Shanghai 200437 China
Correspondence should be addressed to Xian-Mei Zhou zhouxianmeijsaliyuncomJian-Xin Li lijxnjunjueducn and Ling Xu xulq67aliyuncom
Received 19 March 2016 Accepted 30 June 2016
Academic Editor Janelle Wheat
Copyright copy 2016 Hai-Lang He et al This is an open access article distributed under the Creative Commons Attribution Licensewhich permits unrestricted use distribution and reproduction in any medium provided the original work is properly cited
Lung cancer is the leading cause of cancer-related deaths worldwide Jin Fu Kang (JFK) an oral liquid prescription of Chineseherbal drugs has been clinically available for the treatment of non-small cell lung cancer (NSCLC) Lymphangiogenesis is aprimary event in the process of cancer development and metastasis and the formation and migration of lymphatic endothelialcells (LECs) play a key role in the lymphangiogenesis To assess the activity of stromal cell-derived factor-1 (SDF-1) and the coeffectof SDF-1 and vascular endothelial growth factor-C (VEGF-C) on the formation and migration of LECs and clarify the inhibitoryeffects of JFK on the LECs the LECs were differentiated from CD34+VEGFR-3+ endothelial progenitor cells (EPCs) and JFK-containing serums were prepared from rats SDF-1 and VEGF-C both induced the differentiation of CD34+VEGFR-3+ EPCstowards LECs and enhanced the LECs migration Couse of SDF-1 and VEGF-C displayed an additive effect on the LECs formationbut not on their migration JFK inhibited the formation and migration of LECs and the inhibitory effects were most probably viaregulation of the SDF-1CXCR4 andVEGF-CVEGFR-3 axesThe current finding suggested that JFKmight inhibit NSCLC throughantilymphangiogenesis and also provided a potential to discover antilymphangiogenesis agents from natural resources
1 Introduction
Lung cancer as the leading cause of cancer-related deathsworldwide is the most common cancer affecting both menandwomen and holds approximately 27 of all cancer deathsin the United States [1] Non-small cell lung cancer (NSCLC)accounts for gt80 of all lung cancer cases [2] The cancercell migration to distant tissues occurs through blood andlymphatic vessels and is essential for tumor growth andmetastasis [3] Cancer metastasis is a very important event incancer development and accounts for approximately 90 oftreatment failure and related deaths for all cancer However
effective approaches to inhibiting cancer metastasis have notyet been developed
Lymphatic metastasis to regional lymph nodes has beenfocused on as a major indicator for the staging and the prog-nosis of most human cancers and accurate lymph node stag-ing is one of the most important factors in the NSCLC treat-ment and prognosis [4] Growing evidences revealed that thelymphatic vasculature and tumors interact with each otherand promote metastasis formation [5] Lymphatic metastasisalso closely relates to the tumor-induced formation andgrowth of new lymphatic vessels named as lymphangiogene-sis an important initial event in tumor growth and spread
Hindawi Publishing CorporationEvidence-Based Complementary and Alternative MedicineVolume 2016 Article ID 3635209 10 pageshttpdxdoiorg10115520163635209
2 Evidence-Based Complementary and Alternative Medicine
[6] Tumor-induced lymphangiogenesis plays a key role inpromoting the initial spread of malignant tumor cells andresearches designed to block lymphangiogenesis are beingcarried out in the hope of arresting and reversing tumordevelopment [7] Therefore the idea of blocking lymphan-giogenesis might be a useful therapeutic strategy to restrictmetastatic spread [8]
Lymphatic endothelial cells (LECs) play a vital role in reg-ulation of lymphaticmetastasis and lymphangiogenesis inhi-bition of LECs formation and migration might reduce lymphnode and organ metastasis [9 10] Circulating endothelialprogenitor cells (EPCs) have the capacity to contribute toneovessel formation in the presence of proper stimuli [11]Vascular endothelial growth factor-C (VEGF-C) and stromalcell-derived factor-1 (SDF-1 or CXCL12) are two criticalfactors in LECs formation andmigration VEGF-C stimulatescord blood-derived CD34 and vascular endothelial growthfactor receptor-3-positive (CD34+VEGFR-3+) EPCs to dif-ferentiate into LECs that express lymphatic vessel endothelialhyaluronan receptor-1 (LYVE-1) a lymphatic endothelial-specific marker [10] Furthermore although there are nodirect evidences for the effect of SDF-1 on LECs formationand migration SDF-1 closely relates to the tumor lymphan-giogenesis and lymphatic metastasis [12] Therefore bothVEGF-C and SDF-1might be potential targets for therapeuticintervention on cancer [13]
Jin Fu Kang oral liquid (JFK) a Chinese herbal prescrip-tion has been approved by China Food and Drug Adminis-tration and clinically available for the treatment of NSCLC[14 15] Studies have shown that JFK prevents tumor growthand progression and inhibits tumor angiogenesis in NSCLCpatients The possible mechanism might be via inhibition ofthe tumor cells to secrete VEGF [15 16] However whether itsantitumor effect correlates with inhibitory activity on LECsformation and lymphatic metastasis is still unclear
In the present study aiming at clarifying the activity ofSDF-1 and coeffect of SDF-1 and VEGF-C on LECs formationand migration CD34+VEGFR-3+ EPCs were isolated fromcord blood and the LECs formation and migration wereinduced in the presences of SDF-1 and VEGF-C Further-more the JFK-containing serums were prepared from JFK-dosed rats and the inhibitory effects of JFK on formation andmigration of LECs were investigated
2 Materials and Methods
21 Animals and Ethics Statement Twenty healthy male SDrats weighing 200 plusmn 20 g were purchased from the Exper-imental Animal Center of Zhejiang Province The animalswere housed in plastic cages with room temperature of 23 plusmn1∘C under a 12 h light-dark cycle and were given standardlaboratory food and water freely The rats were acclimatizedfor 3 days before the experiment
The animal study was approved by the Jiangsu AnimalCare and Use Committee and all of the protocols compliedwith the national and institutional rules regarding animalexperiments
22 Reagents FITC-conjugated mouse anti-human CD34and mouse IgG1 K isotype control-FITC were from eBio-science (CA USA) APC-conjugated mouse anti-humanVEGFR-3 and mouse IgG1 isotype control-APC were fromRampD systems (USA) Anti-LYVE-1 antibody was fromAbcam FITC-conjugated secondary antibodies were fromKeyGEN Biotechnology Company (Nanjing China)
Ficoll-Hypaque solution was purchased from HaoyangBiotechnology Company (Tianjin China) Goat serumand 4 paraformaldehyde were from Boster Biotechnol-ogy Company (Wuhan China) VEGF-C and SDF-1 werefrom PeproTech Lenvatinib (E7080) was from SelleckchemAMD3100 was from Sigma-Aldrich Dulbeccorsquos modifiedEaglersquos medium (DMEM) and fetal bovine serum (FBS)were from Hyclone Complete growth medium EGM-2MVwas fromLonza 410158406-Diamidino-2-phenylindole (DAPI)wasfrom Beyotime Biotechnology Company (Shanghai China)JFK (Lot number 130202) was obtained from Jilin JinfukangPharmaceutical Co Ltd (Jilin China)
23 Preparation of Blank and JFK-Containing SerumsTwentymale SD rats were randomly divided into four groups5 rats per group Rats in the blank groupwere given 36mLkgof distillated water rats in the three JFK groups were orallygiven JFK at doses of 18 36 and 72 gkg three times per dayfor three successive days respectively After 1 h of final admin-istration the blood samples were collected from the abdomi-nal aorta under ether anaesthesia The blank or JFK-contain-ing serums were acquired by centrifugation of the bloodsamples at 3000 rpm for 15 minutes at 4∘C All serum sampleswere then sterilized by suction with a 022 120583mMillipore filterand finally stored at minus80∘C for experiment use
24 Collection of Mononuclear Cells from Cord Blood Col-lection of cord blood was approved by Ethical Committee ofNanjing Health Hospital for Women and Children and per-formed from placentas of healthy delivery women Mononu-clear cells (MNCs) in cord blood were isolated by density-gradient centrifugation with ficoll solution at the density of1077 After centrifugation for 30min at 400 g the fractionof the mononuclear cells was collected Then the cells weregently washed with PBS in centrifugation for two times [17]
25 Flow Cytometry For identification of CD34+VEGFR-3+ cells the mononuclear cells were incubated with combi-nations of FITC-conjugated mouse anti-human CD34 andAPC-conjugated mouse anti-human VEGFR-3 for 30 min-utes at 4∘C subsequently the cells were washed twice withPBS The immunoglobulin G isotype antibodies were usedas negative controls Then the number of CD34+VEGFR-3+cells was determinedwith FACSort flow cytometer (BeckmanBiosciences) All the staining was carried out according tomanufacturerrsquos protocols
26 Sorting of EPCs from MNCs For sorting of CD34+VEGFR-3+ cells the mononuclear cells were suspended withPBS containing 2 FBS and adjusted to 1 times 107 cellsmLAfter centrifugation the cells were incubated with FITC-conjugated mouse anti-human CD34 and APC-conjugated
Evidence-Based Complementary and Alternative Medicine 3
mouse anti-human VEGFR-3 for 30 minutes at 4∘C Sub-sequently the cells were washed with PBS containing 2FBS and then resuspended with EGM-2MV medium ina 5mL centrifuge tube [18] CD34+VEGFR-3+ cells werecollected by cell sorting with a Beckman MoFlo XDPFACS (fluorescence-activated cell sorter Beckman CoulterFullerton CA USA)
27 Induction of Cell Differentiation Firstly the LECs wereinduced with SDF-1 The freshly sorted CD34+VEGFR-3+ cells were suspended in EGM-2MV medium contain-ing 60 ngmL SDF-1 20 FBS 100UmL penicillin and100 120583gmL streptomycin and seeded in a 24-well plate at adensity of 1 times 105 cellswell The cells were incubated for 14days in a humidified incubator at 37∘C 5 CO
2 and the
medium was changed every 3 daysThe LECs were also induced with VEGF-C and SDF-1 +
VEGF-C with the same protocol as described above exceptthat VEGF-C (60 ngmL) or SDF-1 (100 ngmL) + VEGF-C(60 ngmL) were used instead of SDF-1
28 Transmission Electron Microscopy After induction withSDF-1 VEGF-C or SDF-1 +VEGF-C for 14 days the differen-tiated cells were fixed with 25 glutaraldehyde overnight at4∘Cand thenpostfixedwith 1osmium tetroxideAfter beingdehydrated with gradient alcohol the cells were soaked withanhydrous acetone and Spurr resin and embeddedwith Spurrresin Ultrathin sections were stained with 3 uranyl acetateand lead citrate [10]The ultrastructural characteristics of thecells were examined by using a JEOL JEM-1010 transmissionelectron microscope
29 Immunofluorescence All the LECs differentiated fromthe CD34+VEGFR-3+ cells in the presences of SDF-1 VEGF-C or SDF-1 + VEGF-C and each JFK-containing serumwere identified with expression of lymphatic endotheliumspecific markers LYVE-1 as reported method [19] Brieflyafter induction for 14 days as described above the cellswere fixed by 4 paraformaldehyde and then incubated withrabbit anti-human LYVE-1 antibody (1 100) overnight at 4∘CAfter washing the cells were incubated with FITC-labelledgoat anti-rabbit IgG (1 200) for 60 minutes at 37∘C Thenuclei were counterstained with DAPIThe cells were viewedby using confocal laser scanning microscope Fluorescenceimages and integral optical density (IOD) values were furtheranalyzed with Image-Pro Plus 60 software
210 Transwell Migration Assay The migration efficiency ofLECs was assessed using 8mm pore Transwell filter mem-brane (Corning) following reported method [20] Briefly theLECs were seeded at a density of 2 times 105 cellsmL in theupper chamber and in the lower chamber 100 ngmL of SDF-1 was added After incubation for 16 h the cells on two sidesof the membrane were dried in air and then fixed with 4paraformaldehyde for 15min Subsequently the cells werestained with DAPI for 10min The cells that migrated to thelower sides of the membrane were quantified by countingin 5 fields under a Nikon TE2000 inverted fluorescencemicroscope [21] The same protocol was also performed in
the presences of VEGF-C (60 ngmL) or SDF-1 (100 ngmL) +VEGF-C (60 ngmL)
211 MTT Assay To assess the cytotoxicity of the JFK-containing serums MTT [3-(45-dimethylthiazol-2-yl)-25-diphenyl-2H-tetrazoliumbromide] assaywas performedTheCD34+VEGFR-3+ cells solution (100120583L) containing 10(vv) of the JFK serumswas seeded in a 96-well plate at a den-sity of 1times 104 cellswell and three parallel wells for each groupThe cells were cultured for 48 h as described above After that10 120583L of MTT dye solution (5mgmL in phosphate bufferedsaline pH 74) was added to each well and the plates werefurther incubated for 4 h After incubation the supernatantwas aspirated and the formazan crystals were dissolved inDMSOand the optical density (OD)wasmeasured at 490 nm
212 Inhibition of JFK on LECs Formation To test the effect ofJFK the CD34+VEGFR-3+ cells were divided into 5 groupscontrol group the cells were cultured with blank serum JFK-1 JFK-2 and JFK-3 groups the cells were cultured with JFK-containing serums prepared with JFK at doses of 54 108and 216 gkgday respectively and 100 120583L of each serum wasadded in 09mL of culture medium and AMD group thecells were culturedwith AMD3100 (10 120583M) a SDF-1 inhibitoras a positive control SDF-1 (100 ngmL) was added to allgroups as described previously
The same protocol as in the above was also conducted inwhich VEGF-C (60 ngmL) or SDF-1 (100 ngmL) + VEGF-C(60 ngmL) were used instead of SDF-1 Lenvatinib (10 120583M)a VEGF-C inhibitor or AMD3000 + Lenvatinib (each 10 120583M)were used as positive controls respectively
213 Inhibition of JFK on LECs Migration To test the effectof JFK-containing serums Transwell migration assay wasapplied as described previously In the lower chamber SDF-1(100 ngmL) was addedThe LECs were divided into 5 groupsas described above and pretreated with the blank serumJFK-containing serums (10 vv of culture medium) andAMD3000 (10 120583M) for 30 minutes respectively
The same protocol as in the above was also performedand only VEGF-C (60 ngmL) or SDF-1 (100 ngmL) +VEGF-C (60 ngmL) were added in the lower chamberinstead of SDF-1 Lenvatinib (10 120583M) and AMD3000 +Lenvatinib (each 10 120583M) were used as positive controls
214 Statistical Analysis Data were expressed as means plusmn SDDifferences between experimental groups were assessed bythe two-tailed 119905-test using SPSS 190 software All analyseswere performed using the SPSS 1150 software package andprobability values of 005 or less were considered to bestatistically significant
3 Results
31 Preparation of JFK-Containing Serums As describedpreviously JFK has been used for the treatment of NSCLC inChina JFK is made from 12 Chinese herbal drugs (Table 1)In order to get real effective compositions of JFK workingin vivo an animal experiment using rats was conducted to
4 Evidence-Based Complementary and Alternative Medicine
083
FITC-CD34
APC
-VEG
FR-3
103
102
101
100
100
101
102 10
3
(a)
FITC-IgG
APC
-IgG
103
102
101
100
100
101
102 10
3
(b)
Figure 1 Sorting of CD34+VEGFR-3+ cellsThemononuclear cells were analyzed for the expressions of CD34 andVEGFR-3 with dual-colorflow cytometry (a) and percentage of the CD34+VEGFR-3+ cells was compared with isotype control (b)
Table 1 Composition of Jin Fu Kang oral liquid (JFK)
Herbal drug Part usedAstragalus membranaceus RootGlehnia littoralis RootAsparagus cochinchinensis RootLigustrum lucidum FruitSelaginella doederleinii Whole plantParis polyphylla RootEpimedium sagittatum LeafGynostemma pentaphyllum LeafCornus officinalis FruitSalvia chinensis Whole plantOphiopogon japonicus RootTrigonella foenum graecum Seed
prepare JFK-containing serums Three doses of 18 36 and72 gkg (equivalent to raw herbal drugs) were applied for theserum preparations and the dose of 36 gkg was equal toclinical used one
32 Sorting and Detecting of CD34+VEGFR-3+ Cells Themononuclear cells isolated from cord blood are round havinground- ellipse- or horseshoe-shaped nuclei After beingstained with FITC-conjugated mouse anti-human CD34 andAPC-conjugated mouse anti-human VEGFR-3 the diameterof the cells coexpressing CD34 and VEGFR-3 was 10ndash12 120583mBy double-color flow cytometric analysis the frequency ofCD34+VEGFR-3+ cells was 083 plusmn 021 in the mononu-clear cells (Figures 1(a) and 1(b)) and the freshly sortedCD34+VEGFR-3+ EPCs were round or oval
33 SDF-1 and VEGF-C Additively Induced LECs FormationDifferentiation of CD34+VEGFR-3+ cells towards LECs was
(a) (b)
(c)
Figure 2Morphological changes of CD34+VEGFR-3+ cells duringdifferentiation At day 7 after induction with SDF-1 most cellsbecame spindle-shaped (a) At day 14 the confluentmonolayer of thecells demonstrated a typical cobblestone appearance (b) There aremore mitochondria (white arrow) in the cytoplasm Weibel-Paladebody (black arrow) enwrapped by the membrane is observable (c)
firstly induced in the presence of SDF-1 As described abovethe fresh sorted CD34+VEGFR-3+ cells were round oroval while at day 3 after induction with SDF-1 some cellsrepresented spindle shape or polygon shape At day 7 themost cells were long spindle-shaped (Figure 2(a)) At the endof the experiment (day 14) the cells displayed polygonal shapeand grew into confluent monolayer (Figure 2(b))
To understand the ultrastructural characteristics of thecells a transmission electron microscope examination was
Evidence-Based Complementary and Alternative Medicine 5
performed The observation revealed that the nuclei of thecells were large and there are more mitochondria andphagocytic vesicles in the cytoplasm Furthermore as canbe seen in Figure 2(c) Weibel-Palade body the unique rod-shaped secretory organelle of vascular endothelial cells wasclearly exhibited
As lymphatic vessel endothelial hyaluronan receptor-1 (LYVE-1) is a specific marker of LECs an immunoflu-orescence analysis was conducted with LYVE-1 antibodyThe results showed that after induction for 2 weeks in thepresence of SDF-1 almost all of the cells were positive forLYVE-1 immunostaining (Figure 3(b)) while without SDF-1stimulation (control) only a few LYVE-1 positive LECs wereobserved (Figure 3(a))
VEGF-C was also used for the formation of LECs fromCD34+VEGFR-3+ cellsThe result demonstrated thatVEGF-C induced the differentiation of CD34+VEGFR-3+ EPCstowards LECs while the number of LECs was a little bit morecompared with that of SDF-1 (Figure 3(c))
As shown in Figure 3(d) the LYVE-1 positive cell numberin SDF-1 + VEGF-C group was significantly greater thanthose of SDF-1 or VEGF-C aloneThe integral optical density(IOD) value of LYVE-1 fluorescence in SDF-1 + VEGF-C group (391828) was almost the sum of those in SDF-1(195995) and VEGF-C (236695) groups (Figure 3(e)) Theresults revealed that compared with SDF-1 or VEGF-C alonecostimulation with SDF-1 and VEGF-C displayed an additiveeffect on induction of LECs formation
34 SDF-1 and VEGF-C Promote Migration of LECs but NotAdditively In cell migration assay LECs were seeded in theupper chamber of a Transwell insert and firstly SDF-1 wasadded in the lower chamber As shown in Figures 4(a) and4(b) in the presence of SDF-1 the number of the LECs thatmigrated from the upper chamber to the lower chamber wassignificantly greater comparedwith that in no addition groupThis result clearly indicated that SDF-1 induced the LECsmigration VEGF-C treatment also displayed the same effectas SDF-1 (Figure 4(c))
However when SDF-1 and VEGF-C were cousedalthough a little bitmore LECsmigrated to the lower chambercomparedwith those in SDF-1 orVEGF-C alone (Figures 4(d)and 4(e)) the cell number of SDF-1 + VEGF-C (638 cells) wasfar less than the sum of SDF-1 (511 cells) and VEGF-C (471cells) The results clearly indicated that costimulation withSDF-1 and VEGF-C did not result in an additive effect on thecell transmigration
35 Cytotoxicity of JFK-Containing Serums To verify thecytotoxicity of JFK-containing serums a MTT assay wasperformed As shown in Figure 5 blank serum showed noeffects on EPCs and all the JFK-containing serum samples(10 concentration vv serum to culture medium) exhibitedno cytotoxicity on the CD34+VEGFR-3+ EPCs during 2daysrsquo culture period Thus the 10 concentration of JFK-containing serums was used for the following bioassays
36 Inhibitory Effects of JFK on the LECs Formation As JFKwas used for the treatment of NSCLC to understand whether
(a) (b)
(c) (d)
0
100000
Control SDF-1 VEGF-C VEGF-C + SDF-1
200000
300000
400000
500000IO
Dlowast
lowastlowast
(e)
Figure 3 LECs with positive LYVE-1 immunostaining differen-tiated from CD34+VEGFR-3+ EPCs The CD34+VEGFR-3+ cellswere cultured in the presence of SDF-1 (100 ngmL) and VEGF-C (60 ngmL) The cells expressed LYVE-1 marker after inductionwith no addition ((a) control) SDF-1 (b) VEGF-C (c) and SDF-1 +VEGF-C (d) for two weeks Integral optical density (IOD) of LYVE-1 fluorescence of each group (e) Values are expressed as mean plusmn SD119899 = 3 lowast119901 lt 001 significant increase versus control group
JFK possesses the inhibitory effect on LECs formation JFK-containing serums were testedThe result revealed that blankserum (BS) showed no impact on LECs formation (data notshown) As can be seen in Figure 6 and Table 2 in the positivecontrol AMD3100 (AMD) a SDF-1 inhibitor significantlysuppressed the IODvalues of LYVE-1 about 9373 comparedwith SDF-1 only (BS) As expected all of the JFK-containingserums decreased IOD of LYVE-1 in particular the serumof 72 gkg reduced IOD value about 8279 These resultssuggested JFK-containing serums inhibited LECs formation
The same as SDF-1 the LECs formations induced byVEGF-C and SDF-1 + VEGF-C were also suppressed by JFK-containing serums
6 Evidence-Based Complementary and Alternative Medicine
(a) (b)
(c) (d)
Control SDF-1 VEGF-C VEGF-C + SDF-10
200
400
600
800
The m
igra
ting
cells
lowast
lowastlowast
(e)
Figure 4 Transmigrated LECs in the lower chamber In the lower chamber no addition ((a) control) SDF-1 (100 ngmL (b)) VEGF-C (60 ngmL (c)) and SDF-1 (100 ngmL) + VEGF-C (100 ngmL) (d) were used The migrated cells number in each group (e) Data areexpressed as mean plusmn SD 119899 = 3 lowast119901 lt 001 significant increase versus control group
Table 2 Inhibitory effect of JFK on LECs formation
Group Inhibition rate ()SDF-1 VEGF-C SDF-1 + VEGF-C
BS 000 plusmn 1440 000 plusmn 561 000 plusmn 1829
JFK-1 4410 plusmn 680lowast
3889 plusmn 630lowast
5882 plusmn 1237lowast
JFK-2 6057 plusmn 451lowast
7971 plusmn 131lowast
7606 plusmn 954lowast
JFK-3 8279 plusmn 644lowast
8914 plusmn 143lowast
9116 plusmn 231lowast
AMD 9373 plusmn 060lowast
Len 8718 plusmn 580lowast
Len + AMD 9633 plusmn 060lowast
BS blank serum JFK-1 JFK-2 and JFK-3 the serums prepared from ratsby oral administration of JFK at doses of 18 36 and 72 gkg respectivelyAMD AMD3100 Len Lenvatinib The inhibition rates of BS groups werepegged as 000 and others were calculated relative to it Data wereexpressed as mean plusmn SD 119899 = 3 lowast119901 lt 001 compared with the BS grouprespectively
37 Inhibitory Effects of JFK on the LECsMigration As tumorcell migration is an extremely important process in cancermetastasis the effect of JFK was further evaluated on the
Table 3 Inhibitory effect of JFK on LECs migration
Group Inhibition rate ()SDF-1 VEGF-C VEGF-C + SDF-1
BS 000 plusmn 547 000 plusmn 728 000 plusmn 552
JFK-1 4419 plusmn 503lowast
3281 plusmn 1136lowast
3496 plusmn 541lowast
JFK-2 6603 plusmn 282lowast
6987 plusmn 308lowast
6706 plusmn 549lowast
JFK-3 8049 plusmn 527lowast
8222 plusmn 570lowast
7204 plusmn 277lowast
AMD 8705 plusmn 379lowast
Len 8469 plusmn 390lowast
Len + AMD 8482 plusmn 209lowast
BS blank serum JFK-1 JFK-2 and JFK-3 the serums prepared from ratsby oral administration of JFK at doses of 18 36 and 72 gkg respectivelyAMD AMD3100 Len Lenvatinib The inhibition rates of BS groups werepegged as 000 and others were calculated relative to it Data wereexpressed as mean plusmn SD 119899 = 3 lowast119901 lt 001 compared with the BS grouprespectively
LECs migration As shown in Figure 7 and Table 3 the cellmigration assay data clearly exhibited that JFK-containingserums prepared at all doses demonstrated a significant
Evidence-Based Complementary and Alternative Medicine 7
Control BS JFK-1 JFK-2 JFK-340
60
80
100
120
Cel
l sur
viva
l rat
e (
)
Figure 5 Cytotoxicity of JFK-containing serums The EPCs werecultured with no addition (control) blank serum (BS) and JFK-1JFK-2 and JFK-3 JFK-1 JFK-2 and JFK-3 represented the serumsprepared from rats by oral administration of JFK at doses of 18 36and 72 gkg respectively and 10 (vv) of the serums were used
inhibitory effect on the SDF-1 and VEGF-C induced migra-tions Furthermore the LECs migration induced with SDF-1+ VEGF-C was also markedly suppressed by JFK-containingserums in a dose dependent manner Blank serum showedno effect on the migration (data not shown) The SDF-1 andVEGF-C inducedmigrations were markedly blocked by theirinhibitors AMD3100 or Lenvatinib respectively
4 Discussion
In traditional bioassays in vitro on traditional Chinesemedicines generally extracts or simply purified componentsof crude drugs are directly added to the culture system [22]However the pharmacological actions of traditional Chinesemedicines are rather complicated due to their complexcompositions More important thing is that many of thecomponents contained in the crud drugs do not directlyexhibit their pharmacological actions until they experience aseries of biotransformation during absorption distributionand metabolite procedures Therefore without biotransfor-mation the results from the bioassays in vitromight not trulyreflect the pharmacological actions in vivo Furthermore theserum drug concentration directly reflects the drug efficacyin most cases Therefore serum pharmacology in which thedrug-containing serum was prepared in vivo was used forbioactivity evaluation and research on traditional Chinesemedicines in vitro and the results were more scientific andexact compared with traditional pharmacology [23] In thecurrent study the JFK-containing serums were prepared withrats and applied for the in vitro assays
EPCs have been extensively studied as a potential markerfor endothelial regeneration ability and shown to be atherapeutic potential by directly enhancing angiogenesis
or forming new vessels through vasculogenesis EPCs alsoexhibit a character to be highly migratory to ischemicareas [24] It is reported that circulating CD34+VEGFR-3+lymphaticvascular endothelial progenitor cells correlatewithlymph node metastasis in patients with epithelial ovariancancer [25] and further study revealed that CD34+VEGFR-3+ EPCs in human cord blood could differentiate into lym-phatic endothelial cells [10] Flow cytometry is widely usedto quantify and study circulating EPCs while EPC-specificmarkers are essential [11] Therefore in the current studyCD34 and VEGFR-3 were applied for the sorting of the targetEPCs from human cord blood and the results demonstratedthat the frequency of the CD34+VEGFR-3+ cells was 083in the mononuclear cells which was consistent with thereported data [18]
It is well accepted that LECs contribute to lymphangio-genesis that plays an active role in the induction of metastasisto the lymph nodes in cancers CXC chemokine receptor 4(CXCR4) plays a central role in tumor cell dissemination andmetastasis development in more than 75 of all cancers [26]SDF-1 also known as chemokine CXC ligand binds to theCXCR4 receptor The SDF-1CXCR4 axis is essential for themigration of progenitor cells tissue regeneration and vascu-larization [27] Furthermore SDF-1CXCR4 axis significantlyassociates with lymph node metastasis and promotes lym-phangiogenesis in tumors [28] VEGF-CVEGFR-3 signalingis a keymodulator of the lymphatic system andVEGF-C levelcorrelates with lymph node metastasis and poor prognosis[9] VEGF-C promotes LEC proliferation migration andtube formation [10] VEGFR-3 siRNA effectively inhibitsdifferentiation and lymphangiogenesis of CD34+VEGFR-3+EPCs [29] Therefore in the present study we used SDF-1and VEGF-C to induce the LECs formation and migrationOur results clearly demonstrated that SDF-1 and VEGF-Cpromoted the CD34+VEGFR-3+ EPCs to differentiate to theLECs that showed positive LYVE-1 staining a specific LECmarker and also enhanced the LECs migration
Although it is reported that VEGF-C stimulation upreg-ulates the expression level of CXCR4 on lymphangiogenicendothelial cells SDF-1CXCR4 axis exerts its function ina VEGFR-3 independent pattern [12] More interestinglytargeting both SDF-1 and VEGF-C by neutralizing antibodiesresulted in an additive inhibitory effect on both tumor lym-phangiogenesis and lymphatic metastasis [12] Furthermoreit is hypothesized that SDF-1 and VEGF may have syner-getic effect on endothelial differentiation through a positivefeedback loop with mutual promotion [30] Thus to clarifythe coeffect of SDF-1 and VEGF-C we added both in theculture system and the data revealed that costimulation withSDF-1 and VEGF-C displayed an additive effect on the LECsdifferentiation however no additive effect was observedon the LECs migration which needs a further mechanismclarification
Studies have shown that JFK can prevent tumor growthand progression and inhibit tumor angiogenesis [16] How-ever there is no data onwhether JFK inhibits tumor lymphan-giogenesis The present work provided the first experimentalevidence that JFK suppressed the formation and directionalmigration of LECs in vitro which closely related to the tumor
8 Evidence-Based Complementary and Alternative Medicine
JFK-3BSSD
F-1
SDF-
1SD
F-1
JFK-1 JFK-2 AMDM
erge
DA
PILY
VE-
1
(a1) (a2) (a3) (a4) (a5)
(b1) (b2) (b3) (b4) (b5)
(c1) (c2) (c3) (c4) (c5)JFK-3BS
VEG
F-C
VEG
F-C
VEG
F-C
JFK-1 JFK-2 LEN
(A1)
(B1)
(C1) (C2) (C3) (C4) (C5)
(B2) (B3) (B4) (B5)
(A2) (A3) (A4) (A5)
Mer
geD
API
LYV
E-1
(a1) (a2) (a3) (a4) (a5)
(b1) (b2) (b3) (b4) (b5)
(A1)
(B1) (B2) (B3) (B4) (B5)
(A2) (A3) (A4) (A5)
100120583m
100120583m
100120583m
Figure 6 Effect of JFK on LYVE-1 positive LECs differentiation The LECs formation was induced from CD34+VEGFR-3+ EPCs withSDF-1 (100 ngmL) and VEGF-C (60 ngmL) respectively BS blank serum JFK-1 JFK-2 and JFK-3 the serums prepared from rats by oraladministration of JFK at doses of 18 36 and 72 gkg respectively and 10 (vv) of the serums were added AMD AMD3100 (10 120583M) LenLenvatinib (10 120583M)
Evidence-Based Complementary and Alternative Medicine 9
AMDV
EGF-
CSD
F-1
JFK-2JFK-1 JFK-3BS
LenJFK-2JFK-1 JFK-3BS
200120583m
200120583m
(A1)
(B1) (B2) (B3) (B4) (B5)
(A2) (A3) (A4) (A5)
Figure 7 Effect of JFK on LECs transmigration The LECs migration was induced with SDF-1 (100 ngmL) and VEGF-C (60 ngmL)respectively The LECs were pretreated with blank serum (BS) the serums (each 10 vv) prepared from rats by oral administration ofJFK at doses of 18 36 and 72 gkg (JFK-1 JFK-2 and JFK-3 resp) AMD3100 (AMD 10 120583M) Lenvatinib (Len 10 120583M)
lymphangiogenesis Furthermore the inhibitory effect mightbe via regulation of SDF-1CXCR4 and VEGF-CVEGFR-3axes As JFK is composed of 12 herbal drugs further efforton chemical isolation and structure identification of JFK isessential to clarify the bioactive components responsible forthe effect and might provide a potential to discover the anti-lymphangiogenesis agents from the natural resources
5 Conclusion
This study demonstrated that SDF-1 and VEGF-C inducedCD34+VEGFR-3+ EPCs to differentiate towards LECs andenhanced the LECsmigration SDF-1 andVEGF-C exerted anadditive role on the LECs formation but not on themigrationJFK inhibited the differentiation and migration of LECs viaregulation of the SDF-1CXCR4 and VEGF-CVEGFR-3 axesat least The current finding suggested that JFK might inhibitNSCLC through antilymphangiogenesis and also provided apotential to discover antilymphangiogenesis agents Furtherresearch on the inhibitory effect of JFK on lymphangiogenesisin vivo and clarification of its bioactive components areundergoing in our lab
Competing Interests
The authors declare that they have no conflict of interests
Acknowledgments
This work was supported by Research Project for PracticeDevelopment of National TCM Clinical Research Bases(JDZX2012117) and the Project of Key Discipline for TCMConstruction of Jiangsu Province China (no JS1302)
References
[1] R A Smith D Manassaram-Baptiste D Brooks et al ldquoCancerscreening in the United States 2015 a review of currentAmerican Cancer Society guidelines and current issues incancer screeningrdquo CA A Cancer Journal for Clinicians vol 65no 1 pp 30ndash54 2015
[2] T K Kilvaer E-E Paulsen S M Hald et al ldquoLymphangiogenicmarkers and their impact on nodal metastasis and survival innon-small cell lung cancermdasha structured review with meta-analysisrdquo PLoS ONE vol 10 no 8 Article ID e0132481 2015
[3] F G Gomes F Nedel A M Alves J E Nor and S BC Tarquinio ldquoTumor angiogenesis and lymphangiogenesistumorendothelial crosstalk and cellularmicroenvironmentalsignaling mechanismsrdquo Life Sciences vol 92 no 2 pp 101ndash1072013
[4] Y Wang S Ma M Dong Y Yao K Liu and J Zhou ldquoEvalua-tion of the factors affecting the maximum standardized uptakevalue of metastatic lymph nodes in different histological typesof non-small cell lung cancer on PET-CTrdquo BMC PulmonaryMedicine vol 15 no 1 article 20 2015
[5] J P Sleeman and W Thiele ldquoTumor metastasis and the lym-phatic vasculaturerdquo International Journal of Cancer vol 125 no12 pp 2747ndash2756 2009
[6] S Li and Q Li ldquoCancer stem cells lymphangiogenesis andlymphatic metastasisrdquo Cancer Letters vol 357 no 2 pp 438ndash447 2015
[7] M Bzowska R Męzyk-Kopec T Prochnicki M KuleszaT Klaus and J Bereta ldquoAntibody-based antiangiogenic andantilymphangiogenic therapies to prevent tumor growth andprogressionrdquo Acta Biochimica Polonica vol 60 no 3 pp 263ndash275 2013
[8] S A Stacker S P Williams T Karnezis R Shayan S B Foxand M G Achen ldquoLymphangiogenesis and lymphatic vesselremodelling in cancerrdquo Nature Reviews Cancer vol 14 no 3pp 159ndash172 2014
[9] J Yu X Zhang P M Kuzontkoski et al ldquoSlit2N and Robo4regulate lymphangiogenesis through the VEGF-CVEGFR-3
10 Evidence-Based Complementary and Alternative Medicine
pathwayrdquo Cell Communication and Signaling vol 12 no 1article 25 pp 1ndash15 2014
[10] Y-Z Tan H-J Wang M-H Zhang Z Quan T Li and Q-Z He ldquoCD34+VEGFR-3+ progenitor cells have a potential todifferentiate towards lymphatic endothelial cellsrdquo Journal ofCellular and Molecular Medicine vol 18 no 3 pp 422ndash4332014
[11] M Moschetta Y Mishima I Sahin et al ldquoRole of endothelialprogenitor cells in cancer progressionrdquo Biochimica et BiophysicaActamdashReviews on Cancer vol 1846 no 1 pp 26ndash39 2014
[12] WZhuo L JiaN Song et al ldquoTheCXCL12ndashCXCR4 chemokinepathway a novel axis regulates lymphangiogenesisrdquo ClinicalCancer Research vol 18 no 19 pp 5387ndash5398 2012
[13] B Li W Bai P Sun B Zhou B Hu and J Ying ldquoThe effectof CXCL12 on endothelial progenitor cells potential target forangiogenesis in intracerebral hemorrhagerdquo Journal of Interferonand Cytokine Research vol 35 no 1 pp 23ndash31 2015
[14] J Liu M Pan Y Li D Ye and Y Guo ldquoClinical study of oralliquid Jin Fu Kang for the treatment of primary non-small celllung cancerrdquo Tumor vol 21 no 6 pp 463ndash465 2001
[15] J Liu Z Shi H Li et al ldquoClinical observation on 271 cases ofnon-small lung cancer treated with Yifei Kangliu Yin (Jin-Fu-Kang)rdquo Chinese Journal of Integrated Traditional and WesternMedicine vol 7 no 4 pp 247ndash250 2001
[16] Z CWang J L Zeng H L Zhang and JW Li ldquoJin FuKang ontumor growth and metastasis and vascular endothelial growthfactor of patients with advanced non-small cell lung cancerrdquoPractical Journal of Clinical Medicine vol 8 no 4 pp 148ndash1492011
[17] P Salven A Orpana and H Joensuu ldquoLeukocytes and plateletsof patients with cancer contain high levels of vascular endothe-lial growth factorrdquo Clinical Cancer Research vol 5 no 3 pp487ndash491 1999
[18] H-J Wang D Zhang Y-Z Tan and T Li ldquoAutophagy inendothelial progenitor cells is cytoprotective in hypoxic con-ditionsrdquo American Journal of PhysiologymdashCell Physiology vol304 no 7 pp C617ndashC626 2013
[19] M Tawada S-I Hayashi S Osada S Nakashima and KYoshida ldquoHuman gastric cancer organizes neighboring lym-phatic vessels via recruitment of bone marrow-derived lym-phatic endothelial progenitor cellsrdquo Journal of Gastroenterologyvol 47 no 9 pp 1057ndash1060 2012
[20] AMasuda H Yasuoka T Satoh Y Okazaki Y Yamaguchi andM Kuwana ldquoVersican is upregulated in circulating monocytesin patients with systemic sclerosis and amplifies a CCL2-mediated pathogenic looprdquo Arthritis Research andTherapy vol15 article R74 2013
[21] J Goldman T X Le M Skobe andM A Swartz ldquoOverexpres-sion of VEGF-C causes transient lymphatic hyperplasia but notincreased lymphangiogenesis in regenerating skinrdquo CirculationResearch vol 96 no 11 pp 1193ndash1199 2005
[22] S Nishida and H Satoh ldquoMechanisms for the vasodilationsinduced by Ginkgo biloba extract and its main constituentbilobalide in rat aortardquo Life Sciences vol 72 no 23 pp 2659ndash2667 2003
[23] B Wang L Zhu and Q Chen ldquoPrimary study on the applica-tion of Serum Pharmacology in Chinese traditional medicinerdquoColloids and Surfaces B Biointerfaces vol 43 no 3-4 pp 194ndash197 2005
[24] F Ma A Morancho J Montaner and A Rosell ldquoEndothelialprogenitor cells and revascularization following strokerdquo BrainResearch vol 1623 pp 150ndash159 2015
[25] H Qiu L Cao D Wang H Xu and Z Liang ldquoHigh levelsof circulatingCD34+VEGFR3+ lymphaticvascular endothelialprogenitor cells is correlated with lymph node metastasis inpatients with epithelial ovarian cancerrdquo Journal of Obstetrics andGynaecology Research vol 39 no 7 pp 1268ndash1275 2013
[26] B Furusato A Mohamed M Uhlen and J S Rhim ldquoCXCR4and cancer review Articlerdquo Pathology International vol 60 no7 pp 497ndash505 2010
[27] E Karshovska D Zagorac A Zernecke C Weber and ASchober ldquoA small molecule CXCR4 antagonist inhibits neoin-tima formation and smoothmuscle progenitor cellmobilizationafter arterial injuryrdquo Journal ofThrombosis andHaemostasis vol6 no 10 pp 1812ndash1815 2008
[28] A C Chiang and J Massague ldquoMolecular basis of metastasisrdquoTheNew England Journal of Medicine vol 359 no 26 pp 2814ndash2823 2008
[29] T Li G-DWang Y-Z Tan andH-JWang ldquoInhibition of lym-phangiogenesis of endothelial progenitor cells with VEGFR-3siRNA delivered with PEI-alginate nanoparticlesrdquo InternationalJournal of Biological Sciences vol 10 no 2 pp 160ndash170 2014
[30] M Wang and Z Zou ldquoMultiple mechanisms of SDF-1 promot-ing VEGF-induced endothelial differentiation of mesenchymalstem cellsrdquo International Journal of Cardiology vol 177 no 3pp 1098ndash1099 2014

2 Evidence-Based Complementary and Alternative Medicine
[6] Tumor-induced lymphangiogenesis plays a key role inpromoting the initial spread of malignant tumor cells andresearches designed to block lymphangiogenesis are beingcarried out in the hope of arresting and reversing tumordevelopment [7] Therefore the idea of blocking lymphan-giogenesis might be a useful therapeutic strategy to restrictmetastatic spread [8]
Lymphatic endothelial cells (LECs) play a vital role in reg-ulation of lymphaticmetastasis and lymphangiogenesis inhi-bition of LECs formation and migration might reduce lymphnode and organ metastasis [9 10] Circulating endothelialprogenitor cells (EPCs) have the capacity to contribute toneovessel formation in the presence of proper stimuli [11]Vascular endothelial growth factor-C (VEGF-C) and stromalcell-derived factor-1 (SDF-1 or CXCL12) are two criticalfactors in LECs formation andmigration VEGF-C stimulatescord blood-derived CD34 and vascular endothelial growthfactor receptor-3-positive (CD34+VEGFR-3+) EPCs to dif-ferentiate into LECs that express lymphatic vessel endothelialhyaluronan receptor-1 (LYVE-1) a lymphatic endothelial-specific marker [10] Furthermore although there are nodirect evidences for the effect of SDF-1 on LECs formationand migration SDF-1 closely relates to the tumor lymphan-giogenesis and lymphatic metastasis [12] Therefore bothVEGF-C and SDF-1might be potential targets for therapeuticintervention on cancer [13]
Jin Fu Kang oral liquid (JFK) a Chinese herbal prescrip-tion has been approved by China Food and Drug Adminis-tration and clinically available for the treatment of NSCLC[14 15] Studies have shown that JFK prevents tumor growthand progression and inhibits tumor angiogenesis in NSCLCpatients The possible mechanism might be via inhibition ofthe tumor cells to secrete VEGF [15 16] However whether itsantitumor effect correlates with inhibitory activity on LECsformation and lymphatic metastasis is still unclear
In the present study aiming at clarifying the activity ofSDF-1 and coeffect of SDF-1 and VEGF-C on LECs formationand migration CD34+VEGFR-3+ EPCs were isolated fromcord blood and the LECs formation and migration wereinduced in the presences of SDF-1 and VEGF-C Further-more the JFK-containing serums were prepared from JFK-dosed rats and the inhibitory effects of JFK on formation andmigration of LECs were investigated
2 Materials and Methods
21 Animals and Ethics Statement Twenty healthy male SDrats weighing 200 plusmn 20 g were purchased from the Exper-imental Animal Center of Zhejiang Province The animalswere housed in plastic cages with room temperature of 23 plusmn1∘C under a 12 h light-dark cycle and were given standardlaboratory food and water freely The rats were acclimatizedfor 3 days before the experiment
The animal study was approved by the Jiangsu AnimalCare and Use Committee and all of the protocols compliedwith the national and institutional rules regarding animalexperiments
22 Reagents FITC-conjugated mouse anti-human CD34and mouse IgG1 K isotype control-FITC were from eBio-science (CA USA) APC-conjugated mouse anti-humanVEGFR-3 and mouse IgG1 isotype control-APC were fromRampD systems (USA) Anti-LYVE-1 antibody was fromAbcam FITC-conjugated secondary antibodies were fromKeyGEN Biotechnology Company (Nanjing China)
Ficoll-Hypaque solution was purchased from HaoyangBiotechnology Company (Tianjin China) Goat serumand 4 paraformaldehyde were from Boster Biotechnol-ogy Company (Wuhan China) VEGF-C and SDF-1 werefrom PeproTech Lenvatinib (E7080) was from SelleckchemAMD3100 was from Sigma-Aldrich Dulbeccorsquos modifiedEaglersquos medium (DMEM) and fetal bovine serum (FBS)were from Hyclone Complete growth medium EGM-2MVwas fromLonza 410158406-Diamidino-2-phenylindole (DAPI)wasfrom Beyotime Biotechnology Company (Shanghai China)JFK (Lot number 130202) was obtained from Jilin JinfukangPharmaceutical Co Ltd (Jilin China)
23 Preparation of Blank and JFK-Containing SerumsTwentymale SD rats were randomly divided into four groups5 rats per group Rats in the blank groupwere given 36mLkgof distillated water rats in the three JFK groups were orallygiven JFK at doses of 18 36 and 72 gkg three times per dayfor three successive days respectively After 1 h of final admin-istration the blood samples were collected from the abdomi-nal aorta under ether anaesthesia The blank or JFK-contain-ing serums were acquired by centrifugation of the bloodsamples at 3000 rpm for 15 minutes at 4∘C All serum sampleswere then sterilized by suction with a 022 120583mMillipore filterand finally stored at minus80∘C for experiment use
24 Collection of Mononuclear Cells from Cord Blood Col-lection of cord blood was approved by Ethical Committee ofNanjing Health Hospital for Women and Children and per-formed from placentas of healthy delivery women Mononu-clear cells (MNCs) in cord blood were isolated by density-gradient centrifugation with ficoll solution at the density of1077 After centrifugation for 30min at 400 g the fractionof the mononuclear cells was collected Then the cells weregently washed with PBS in centrifugation for two times [17]
25 Flow Cytometry For identification of CD34+VEGFR-3+ cells the mononuclear cells were incubated with combi-nations of FITC-conjugated mouse anti-human CD34 andAPC-conjugated mouse anti-human VEGFR-3 for 30 min-utes at 4∘C subsequently the cells were washed twice withPBS The immunoglobulin G isotype antibodies were usedas negative controls Then the number of CD34+VEGFR-3+cells was determinedwith FACSort flow cytometer (BeckmanBiosciences) All the staining was carried out according tomanufacturerrsquos protocols
26 Sorting of EPCs from MNCs For sorting of CD34+VEGFR-3+ cells the mononuclear cells were suspended withPBS containing 2 FBS and adjusted to 1 times 107 cellsmLAfter centrifugation the cells were incubated with FITC-conjugated mouse anti-human CD34 and APC-conjugated
Evidence-Based Complementary and Alternative Medicine 3
mouse anti-human VEGFR-3 for 30 minutes at 4∘C Sub-sequently the cells were washed with PBS containing 2FBS and then resuspended with EGM-2MV medium ina 5mL centrifuge tube [18] CD34+VEGFR-3+ cells werecollected by cell sorting with a Beckman MoFlo XDPFACS (fluorescence-activated cell sorter Beckman CoulterFullerton CA USA)
27 Induction of Cell Differentiation Firstly the LECs wereinduced with SDF-1 The freshly sorted CD34+VEGFR-3+ cells were suspended in EGM-2MV medium contain-ing 60 ngmL SDF-1 20 FBS 100UmL penicillin and100 120583gmL streptomycin and seeded in a 24-well plate at adensity of 1 times 105 cellswell The cells were incubated for 14days in a humidified incubator at 37∘C 5 CO
2 and the
medium was changed every 3 daysThe LECs were also induced with VEGF-C and SDF-1 +
VEGF-C with the same protocol as described above exceptthat VEGF-C (60 ngmL) or SDF-1 (100 ngmL) + VEGF-C(60 ngmL) were used instead of SDF-1
28 Transmission Electron Microscopy After induction withSDF-1 VEGF-C or SDF-1 +VEGF-C for 14 days the differen-tiated cells were fixed with 25 glutaraldehyde overnight at4∘Cand thenpostfixedwith 1osmium tetroxideAfter beingdehydrated with gradient alcohol the cells were soaked withanhydrous acetone and Spurr resin and embeddedwith Spurrresin Ultrathin sections were stained with 3 uranyl acetateand lead citrate [10]The ultrastructural characteristics of thecells were examined by using a JEOL JEM-1010 transmissionelectron microscope
29 Immunofluorescence All the LECs differentiated fromthe CD34+VEGFR-3+ cells in the presences of SDF-1 VEGF-C or SDF-1 + VEGF-C and each JFK-containing serumwere identified with expression of lymphatic endotheliumspecific markers LYVE-1 as reported method [19] Brieflyafter induction for 14 days as described above the cellswere fixed by 4 paraformaldehyde and then incubated withrabbit anti-human LYVE-1 antibody (1 100) overnight at 4∘CAfter washing the cells were incubated with FITC-labelledgoat anti-rabbit IgG (1 200) for 60 minutes at 37∘C Thenuclei were counterstained with DAPIThe cells were viewedby using confocal laser scanning microscope Fluorescenceimages and integral optical density (IOD) values were furtheranalyzed with Image-Pro Plus 60 software
210 Transwell Migration Assay The migration efficiency ofLECs was assessed using 8mm pore Transwell filter mem-brane (Corning) following reported method [20] Briefly theLECs were seeded at a density of 2 times 105 cellsmL in theupper chamber and in the lower chamber 100 ngmL of SDF-1 was added After incubation for 16 h the cells on two sidesof the membrane were dried in air and then fixed with 4paraformaldehyde for 15min Subsequently the cells werestained with DAPI for 10min The cells that migrated to thelower sides of the membrane were quantified by countingin 5 fields under a Nikon TE2000 inverted fluorescencemicroscope [21] The same protocol was also performed in
the presences of VEGF-C (60 ngmL) or SDF-1 (100 ngmL) +VEGF-C (60 ngmL)
211 MTT Assay To assess the cytotoxicity of the JFK-containing serums MTT [3-(45-dimethylthiazol-2-yl)-25-diphenyl-2H-tetrazoliumbromide] assaywas performedTheCD34+VEGFR-3+ cells solution (100120583L) containing 10(vv) of the JFK serumswas seeded in a 96-well plate at a den-sity of 1times 104 cellswell and three parallel wells for each groupThe cells were cultured for 48 h as described above After that10 120583L of MTT dye solution (5mgmL in phosphate bufferedsaline pH 74) was added to each well and the plates werefurther incubated for 4 h After incubation the supernatantwas aspirated and the formazan crystals were dissolved inDMSOand the optical density (OD)wasmeasured at 490 nm
212 Inhibition of JFK on LECs Formation To test the effect ofJFK the CD34+VEGFR-3+ cells were divided into 5 groupscontrol group the cells were cultured with blank serum JFK-1 JFK-2 and JFK-3 groups the cells were cultured with JFK-containing serums prepared with JFK at doses of 54 108and 216 gkgday respectively and 100 120583L of each serum wasadded in 09mL of culture medium and AMD group thecells were culturedwith AMD3100 (10 120583M) a SDF-1 inhibitoras a positive control SDF-1 (100 ngmL) was added to allgroups as described previously
The same protocol as in the above was also conducted inwhich VEGF-C (60 ngmL) or SDF-1 (100 ngmL) + VEGF-C(60 ngmL) were used instead of SDF-1 Lenvatinib (10 120583M)a VEGF-C inhibitor or AMD3000 + Lenvatinib (each 10 120583M)were used as positive controls respectively
213 Inhibition of JFK on LECs Migration To test the effectof JFK-containing serums Transwell migration assay wasapplied as described previously In the lower chamber SDF-1(100 ngmL) was addedThe LECs were divided into 5 groupsas described above and pretreated with the blank serumJFK-containing serums (10 vv of culture medium) andAMD3000 (10 120583M) for 30 minutes respectively
The same protocol as in the above was also performedand only VEGF-C (60 ngmL) or SDF-1 (100 ngmL) +VEGF-C (60 ngmL) were added in the lower chamberinstead of SDF-1 Lenvatinib (10 120583M) and AMD3000 +Lenvatinib (each 10 120583M) were used as positive controls
214 Statistical Analysis Data were expressed as means plusmn SDDifferences between experimental groups were assessed bythe two-tailed 119905-test using SPSS 190 software All analyseswere performed using the SPSS 1150 software package andprobability values of 005 or less were considered to bestatistically significant
3 Results
31 Preparation of JFK-Containing Serums As describedpreviously JFK has been used for the treatment of NSCLC inChina JFK is made from 12 Chinese herbal drugs (Table 1)In order to get real effective compositions of JFK workingin vivo an animal experiment using rats was conducted to
4 Evidence-Based Complementary and Alternative Medicine
083
FITC-CD34
APC
-VEG
FR-3
103
102
101
100
100
101
102 10
3
(a)
FITC-IgG
APC
-IgG
103
102
101
100
100
101
102 10
3
(b)
Figure 1 Sorting of CD34+VEGFR-3+ cellsThemononuclear cells were analyzed for the expressions of CD34 andVEGFR-3 with dual-colorflow cytometry (a) and percentage of the CD34+VEGFR-3+ cells was compared with isotype control (b)
Table 1 Composition of Jin Fu Kang oral liquid (JFK)
Herbal drug Part usedAstragalus membranaceus RootGlehnia littoralis RootAsparagus cochinchinensis RootLigustrum lucidum FruitSelaginella doederleinii Whole plantParis polyphylla RootEpimedium sagittatum LeafGynostemma pentaphyllum LeafCornus officinalis FruitSalvia chinensis Whole plantOphiopogon japonicus RootTrigonella foenum graecum Seed
prepare JFK-containing serums Three doses of 18 36 and72 gkg (equivalent to raw herbal drugs) were applied for theserum preparations and the dose of 36 gkg was equal toclinical used one
32 Sorting and Detecting of CD34+VEGFR-3+ Cells Themononuclear cells isolated from cord blood are round havinground- ellipse- or horseshoe-shaped nuclei After beingstained with FITC-conjugated mouse anti-human CD34 andAPC-conjugated mouse anti-human VEGFR-3 the diameterof the cells coexpressing CD34 and VEGFR-3 was 10ndash12 120583mBy double-color flow cytometric analysis the frequency ofCD34+VEGFR-3+ cells was 083 plusmn 021 in the mononu-clear cells (Figures 1(a) and 1(b)) and the freshly sortedCD34+VEGFR-3+ EPCs were round or oval
33 SDF-1 and VEGF-C Additively Induced LECs FormationDifferentiation of CD34+VEGFR-3+ cells towards LECs was
(a) (b)
(c)
Figure 2Morphological changes of CD34+VEGFR-3+ cells duringdifferentiation At day 7 after induction with SDF-1 most cellsbecame spindle-shaped (a) At day 14 the confluentmonolayer of thecells demonstrated a typical cobblestone appearance (b) There aremore mitochondria (white arrow) in the cytoplasm Weibel-Paladebody (black arrow) enwrapped by the membrane is observable (c)
firstly induced in the presence of SDF-1 As described abovethe fresh sorted CD34+VEGFR-3+ cells were round oroval while at day 3 after induction with SDF-1 some cellsrepresented spindle shape or polygon shape At day 7 themost cells were long spindle-shaped (Figure 2(a)) At the endof the experiment (day 14) the cells displayed polygonal shapeand grew into confluent monolayer (Figure 2(b))
To understand the ultrastructural characteristics of thecells a transmission electron microscope examination was
Evidence-Based Complementary and Alternative Medicine 5
performed The observation revealed that the nuclei of thecells were large and there are more mitochondria andphagocytic vesicles in the cytoplasm Furthermore as canbe seen in Figure 2(c) Weibel-Palade body the unique rod-shaped secretory organelle of vascular endothelial cells wasclearly exhibited
As lymphatic vessel endothelial hyaluronan receptor-1 (LYVE-1) is a specific marker of LECs an immunoflu-orescence analysis was conducted with LYVE-1 antibodyThe results showed that after induction for 2 weeks in thepresence of SDF-1 almost all of the cells were positive forLYVE-1 immunostaining (Figure 3(b)) while without SDF-1stimulation (control) only a few LYVE-1 positive LECs wereobserved (Figure 3(a))
VEGF-C was also used for the formation of LECs fromCD34+VEGFR-3+ cellsThe result demonstrated thatVEGF-C induced the differentiation of CD34+VEGFR-3+ EPCstowards LECs while the number of LECs was a little bit morecompared with that of SDF-1 (Figure 3(c))
As shown in Figure 3(d) the LYVE-1 positive cell numberin SDF-1 + VEGF-C group was significantly greater thanthose of SDF-1 or VEGF-C aloneThe integral optical density(IOD) value of LYVE-1 fluorescence in SDF-1 + VEGF-C group (391828) was almost the sum of those in SDF-1(195995) and VEGF-C (236695) groups (Figure 3(e)) Theresults revealed that compared with SDF-1 or VEGF-C alonecostimulation with SDF-1 and VEGF-C displayed an additiveeffect on induction of LECs formation
34 SDF-1 and VEGF-C Promote Migration of LECs but NotAdditively In cell migration assay LECs were seeded in theupper chamber of a Transwell insert and firstly SDF-1 wasadded in the lower chamber As shown in Figures 4(a) and4(b) in the presence of SDF-1 the number of the LECs thatmigrated from the upper chamber to the lower chamber wassignificantly greater comparedwith that in no addition groupThis result clearly indicated that SDF-1 induced the LECsmigration VEGF-C treatment also displayed the same effectas SDF-1 (Figure 4(c))
However when SDF-1 and VEGF-C were cousedalthough a little bitmore LECsmigrated to the lower chambercomparedwith those in SDF-1 orVEGF-C alone (Figures 4(d)and 4(e)) the cell number of SDF-1 + VEGF-C (638 cells) wasfar less than the sum of SDF-1 (511 cells) and VEGF-C (471cells) The results clearly indicated that costimulation withSDF-1 and VEGF-C did not result in an additive effect on thecell transmigration
35 Cytotoxicity of JFK-Containing Serums To verify thecytotoxicity of JFK-containing serums a MTT assay wasperformed As shown in Figure 5 blank serum showed noeffects on EPCs and all the JFK-containing serum samples(10 concentration vv serum to culture medium) exhibitedno cytotoxicity on the CD34+VEGFR-3+ EPCs during 2daysrsquo culture period Thus the 10 concentration of JFK-containing serums was used for the following bioassays
36 Inhibitory Effects of JFK on the LECs Formation As JFKwas used for the treatment of NSCLC to understand whether
(a) (b)
(c) (d)
0
100000
Control SDF-1 VEGF-C VEGF-C + SDF-1
200000
300000
400000
500000IO
Dlowast
lowastlowast
(e)
Figure 3 LECs with positive LYVE-1 immunostaining differen-tiated from CD34+VEGFR-3+ EPCs The CD34+VEGFR-3+ cellswere cultured in the presence of SDF-1 (100 ngmL) and VEGF-C (60 ngmL) The cells expressed LYVE-1 marker after inductionwith no addition ((a) control) SDF-1 (b) VEGF-C (c) and SDF-1 +VEGF-C (d) for two weeks Integral optical density (IOD) of LYVE-1 fluorescence of each group (e) Values are expressed as mean plusmn SD119899 = 3 lowast119901 lt 001 significant increase versus control group
JFK possesses the inhibitory effect on LECs formation JFK-containing serums were testedThe result revealed that blankserum (BS) showed no impact on LECs formation (data notshown) As can be seen in Figure 6 and Table 2 in the positivecontrol AMD3100 (AMD) a SDF-1 inhibitor significantlysuppressed the IODvalues of LYVE-1 about 9373 comparedwith SDF-1 only (BS) As expected all of the JFK-containingserums decreased IOD of LYVE-1 in particular the serumof 72 gkg reduced IOD value about 8279 These resultssuggested JFK-containing serums inhibited LECs formation
The same as SDF-1 the LECs formations induced byVEGF-C and SDF-1 + VEGF-C were also suppressed by JFK-containing serums
6 Evidence-Based Complementary and Alternative Medicine
(a) (b)
(c) (d)
Control SDF-1 VEGF-C VEGF-C + SDF-10
200
400
600
800
The m
igra
ting
cells
lowast
lowastlowast
(e)
Figure 4 Transmigrated LECs in the lower chamber In the lower chamber no addition ((a) control) SDF-1 (100 ngmL (b)) VEGF-C (60 ngmL (c)) and SDF-1 (100 ngmL) + VEGF-C (100 ngmL) (d) were used The migrated cells number in each group (e) Data areexpressed as mean plusmn SD 119899 = 3 lowast119901 lt 001 significant increase versus control group
Table 2 Inhibitory effect of JFK on LECs formation
Group Inhibition rate ()SDF-1 VEGF-C SDF-1 + VEGF-C
BS 000 plusmn 1440 000 plusmn 561 000 plusmn 1829
JFK-1 4410 plusmn 680lowast
3889 plusmn 630lowast
5882 plusmn 1237lowast
JFK-2 6057 plusmn 451lowast
7971 plusmn 131lowast
7606 plusmn 954lowast
JFK-3 8279 plusmn 644lowast
8914 plusmn 143lowast
9116 plusmn 231lowast
AMD 9373 plusmn 060lowast
Len 8718 plusmn 580lowast
Len + AMD 9633 plusmn 060lowast
BS blank serum JFK-1 JFK-2 and JFK-3 the serums prepared from ratsby oral administration of JFK at doses of 18 36 and 72 gkg respectivelyAMD AMD3100 Len Lenvatinib The inhibition rates of BS groups werepegged as 000 and others were calculated relative to it Data wereexpressed as mean plusmn SD 119899 = 3 lowast119901 lt 001 compared with the BS grouprespectively
37 Inhibitory Effects of JFK on the LECsMigration As tumorcell migration is an extremely important process in cancermetastasis the effect of JFK was further evaluated on the
Table 3 Inhibitory effect of JFK on LECs migration
Group Inhibition rate ()SDF-1 VEGF-C VEGF-C + SDF-1
BS 000 plusmn 547 000 plusmn 728 000 plusmn 552
JFK-1 4419 plusmn 503lowast
3281 plusmn 1136lowast
3496 plusmn 541lowast
JFK-2 6603 plusmn 282lowast
6987 plusmn 308lowast
6706 plusmn 549lowast
JFK-3 8049 plusmn 527lowast
8222 plusmn 570lowast
7204 plusmn 277lowast
AMD 8705 plusmn 379lowast
Len 8469 plusmn 390lowast
Len + AMD 8482 plusmn 209lowast
BS blank serum JFK-1 JFK-2 and JFK-3 the serums prepared from ratsby oral administration of JFK at doses of 18 36 and 72 gkg respectivelyAMD AMD3100 Len Lenvatinib The inhibition rates of BS groups werepegged as 000 and others were calculated relative to it Data wereexpressed as mean plusmn SD 119899 = 3 lowast119901 lt 001 compared with the BS grouprespectively
LECs migration As shown in Figure 7 and Table 3 the cellmigration assay data clearly exhibited that JFK-containingserums prepared at all doses demonstrated a significant
Evidence-Based Complementary and Alternative Medicine 7
Control BS JFK-1 JFK-2 JFK-340
60
80
100
120
Cel
l sur
viva
l rat
e (
)
Figure 5 Cytotoxicity of JFK-containing serums The EPCs werecultured with no addition (control) blank serum (BS) and JFK-1JFK-2 and JFK-3 JFK-1 JFK-2 and JFK-3 represented the serumsprepared from rats by oral administration of JFK at doses of 18 36and 72 gkg respectively and 10 (vv) of the serums were used
inhibitory effect on the SDF-1 and VEGF-C induced migra-tions Furthermore the LECs migration induced with SDF-1+ VEGF-C was also markedly suppressed by JFK-containingserums in a dose dependent manner Blank serum showedno effect on the migration (data not shown) The SDF-1 andVEGF-C inducedmigrations were markedly blocked by theirinhibitors AMD3100 or Lenvatinib respectively
4 Discussion
In traditional bioassays in vitro on traditional Chinesemedicines generally extracts or simply purified componentsof crude drugs are directly added to the culture system [22]However the pharmacological actions of traditional Chinesemedicines are rather complicated due to their complexcompositions More important thing is that many of thecomponents contained in the crud drugs do not directlyexhibit their pharmacological actions until they experience aseries of biotransformation during absorption distributionand metabolite procedures Therefore without biotransfor-mation the results from the bioassays in vitromight not trulyreflect the pharmacological actions in vivo Furthermore theserum drug concentration directly reflects the drug efficacyin most cases Therefore serum pharmacology in which thedrug-containing serum was prepared in vivo was used forbioactivity evaluation and research on traditional Chinesemedicines in vitro and the results were more scientific andexact compared with traditional pharmacology [23] In thecurrent study the JFK-containing serums were prepared withrats and applied for the in vitro assays
EPCs have been extensively studied as a potential markerfor endothelial regeneration ability and shown to be atherapeutic potential by directly enhancing angiogenesis
or forming new vessels through vasculogenesis EPCs alsoexhibit a character to be highly migratory to ischemicareas [24] It is reported that circulating CD34+VEGFR-3+lymphaticvascular endothelial progenitor cells correlatewithlymph node metastasis in patients with epithelial ovariancancer [25] and further study revealed that CD34+VEGFR-3+ EPCs in human cord blood could differentiate into lym-phatic endothelial cells [10] Flow cytometry is widely usedto quantify and study circulating EPCs while EPC-specificmarkers are essential [11] Therefore in the current studyCD34 and VEGFR-3 were applied for the sorting of the targetEPCs from human cord blood and the results demonstratedthat the frequency of the CD34+VEGFR-3+ cells was 083in the mononuclear cells which was consistent with thereported data [18]
It is well accepted that LECs contribute to lymphangio-genesis that plays an active role in the induction of metastasisto the lymph nodes in cancers CXC chemokine receptor 4(CXCR4) plays a central role in tumor cell dissemination andmetastasis development in more than 75 of all cancers [26]SDF-1 also known as chemokine CXC ligand binds to theCXCR4 receptor The SDF-1CXCR4 axis is essential for themigration of progenitor cells tissue regeneration and vascu-larization [27] Furthermore SDF-1CXCR4 axis significantlyassociates with lymph node metastasis and promotes lym-phangiogenesis in tumors [28] VEGF-CVEGFR-3 signalingis a keymodulator of the lymphatic system andVEGF-C levelcorrelates with lymph node metastasis and poor prognosis[9] VEGF-C promotes LEC proliferation migration andtube formation [10] VEGFR-3 siRNA effectively inhibitsdifferentiation and lymphangiogenesis of CD34+VEGFR-3+EPCs [29] Therefore in the present study we used SDF-1and VEGF-C to induce the LECs formation and migrationOur results clearly demonstrated that SDF-1 and VEGF-Cpromoted the CD34+VEGFR-3+ EPCs to differentiate to theLECs that showed positive LYVE-1 staining a specific LECmarker and also enhanced the LECs migration
Although it is reported that VEGF-C stimulation upreg-ulates the expression level of CXCR4 on lymphangiogenicendothelial cells SDF-1CXCR4 axis exerts its function ina VEGFR-3 independent pattern [12] More interestinglytargeting both SDF-1 and VEGF-C by neutralizing antibodiesresulted in an additive inhibitory effect on both tumor lym-phangiogenesis and lymphatic metastasis [12] Furthermoreit is hypothesized that SDF-1 and VEGF may have syner-getic effect on endothelial differentiation through a positivefeedback loop with mutual promotion [30] Thus to clarifythe coeffect of SDF-1 and VEGF-C we added both in theculture system and the data revealed that costimulation withSDF-1 and VEGF-C displayed an additive effect on the LECsdifferentiation however no additive effect was observedon the LECs migration which needs a further mechanismclarification
Studies have shown that JFK can prevent tumor growthand progression and inhibit tumor angiogenesis [16] How-ever there is no data onwhether JFK inhibits tumor lymphan-giogenesis The present work provided the first experimentalevidence that JFK suppressed the formation and directionalmigration of LECs in vitro which closely related to the tumor
8 Evidence-Based Complementary and Alternative Medicine
JFK-3BSSD
F-1
SDF-
1SD
F-1
JFK-1 JFK-2 AMDM
erge
DA
PILY
VE-
1
(a1) (a2) (a3) (a4) (a5)
(b1) (b2) (b3) (b4) (b5)
(c1) (c2) (c3) (c4) (c5)JFK-3BS
VEG
F-C
VEG
F-C
VEG
F-C
JFK-1 JFK-2 LEN
(A1)
(B1)
(C1) (C2) (C3) (C4) (C5)
(B2) (B3) (B4) (B5)
(A2) (A3) (A4) (A5)
Mer
geD
API
LYV
E-1
(a1) (a2) (a3) (a4) (a5)
(b1) (b2) (b3) (b4) (b5)
(A1)
(B1) (B2) (B3) (B4) (B5)
(A2) (A3) (A4) (A5)
100120583m
100120583m
100120583m
Figure 6 Effect of JFK on LYVE-1 positive LECs differentiation The LECs formation was induced from CD34+VEGFR-3+ EPCs withSDF-1 (100 ngmL) and VEGF-C (60 ngmL) respectively BS blank serum JFK-1 JFK-2 and JFK-3 the serums prepared from rats by oraladministration of JFK at doses of 18 36 and 72 gkg respectively and 10 (vv) of the serums were added AMD AMD3100 (10 120583M) LenLenvatinib (10 120583M)
Evidence-Based Complementary and Alternative Medicine 9
AMDV
EGF-
CSD
F-1
JFK-2JFK-1 JFK-3BS
LenJFK-2JFK-1 JFK-3BS
200120583m
200120583m
(A1)
(B1) (B2) (B3) (B4) (B5)
(A2) (A3) (A4) (A5)
Figure 7 Effect of JFK on LECs transmigration The LECs migration was induced with SDF-1 (100 ngmL) and VEGF-C (60 ngmL)respectively The LECs were pretreated with blank serum (BS) the serums (each 10 vv) prepared from rats by oral administration ofJFK at doses of 18 36 and 72 gkg (JFK-1 JFK-2 and JFK-3 resp) AMD3100 (AMD 10 120583M) Lenvatinib (Len 10 120583M)
lymphangiogenesis Furthermore the inhibitory effect mightbe via regulation of SDF-1CXCR4 and VEGF-CVEGFR-3axes As JFK is composed of 12 herbal drugs further efforton chemical isolation and structure identification of JFK isessential to clarify the bioactive components responsible forthe effect and might provide a potential to discover the anti-lymphangiogenesis agents from the natural resources
5 Conclusion
This study demonstrated that SDF-1 and VEGF-C inducedCD34+VEGFR-3+ EPCs to differentiate towards LECs andenhanced the LECsmigration SDF-1 andVEGF-C exerted anadditive role on the LECs formation but not on themigrationJFK inhibited the differentiation and migration of LECs viaregulation of the SDF-1CXCR4 and VEGF-CVEGFR-3 axesat least The current finding suggested that JFK might inhibitNSCLC through antilymphangiogenesis and also provided apotential to discover antilymphangiogenesis agents Furtherresearch on the inhibitory effect of JFK on lymphangiogenesisin vivo and clarification of its bioactive components areundergoing in our lab
Competing Interests
The authors declare that they have no conflict of interests
Acknowledgments
This work was supported by Research Project for PracticeDevelopment of National TCM Clinical Research Bases(JDZX2012117) and the Project of Key Discipline for TCMConstruction of Jiangsu Province China (no JS1302)
References
[1] R A Smith D Manassaram-Baptiste D Brooks et al ldquoCancerscreening in the United States 2015 a review of currentAmerican Cancer Society guidelines and current issues incancer screeningrdquo CA A Cancer Journal for Clinicians vol 65no 1 pp 30ndash54 2015
[2] T K Kilvaer E-E Paulsen S M Hald et al ldquoLymphangiogenicmarkers and their impact on nodal metastasis and survival innon-small cell lung cancermdasha structured review with meta-analysisrdquo PLoS ONE vol 10 no 8 Article ID e0132481 2015
[3] F G Gomes F Nedel A M Alves J E Nor and S BC Tarquinio ldquoTumor angiogenesis and lymphangiogenesistumorendothelial crosstalk and cellularmicroenvironmentalsignaling mechanismsrdquo Life Sciences vol 92 no 2 pp 101ndash1072013
[4] Y Wang S Ma M Dong Y Yao K Liu and J Zhou ldquoEvalua-tion of the factors affecting the maximum standardized uptakevalue of metastatic lymph nodes in different histological typesof non-small cell lung cancer on PET-CTrdquo BMC PulmonaryMedicine vol 15 no 1 article 20 2015
[5] J P Sleeman and W Thiele ldquoTumor metastasis and the lym-phatic vasculaturerdquo International Journal of Cancer vol 125 no12 pp 2747ndash2756 2009
[6] S Li and Q Li ldquoCancer stem cells lymphangiogenesis andlymphatic metastasisrdquo Cancer Letters vol 357 no 2 pp 438ndash447 2015
[7] M Bzowska R Męzyk-Kopec T Prochnicki M KuleszaT Klaus and J Bereta ldquoAntibody-based antiangiogenic andantilymphangiogenic therapies to prevent tumor growth andprogressionrdquo Acta Biochimica Polonica vol 60 no 3 pp 263ndash275 2013
[8] S A Stacker S P Williams T Karnezis R Shayan S B Foxand M G Achen ldquoLymphangiogenesis and lymphatic vesselremodelling in cancerrdquo Nature Reviews Cancer vol 14 no 3pp 159ndash172 2014
[9] J Yu X Zhang P M Kuzontkoski et al ldquoSlit2N and Robo4regulate lymphangiogenesis through the VEGF-CVEGFR-3
10 Evidence-Based Complementary and Alternative Medicine
pathwayrdquo Cell Communication and Signaling vol 12 no 1article 25 pp 1ndash15 2014
[10] Y-Z Tan H-J Wang M-H Zhang Z Quan T Li and Q-Z He ldquoCD34+VEGFR-3+ progenitor cells have a potential todifferentiate towards lymphatic endothelial cellsrdquo Journal ofCellular and Molecular Medicine vol 18 no 3 pp 422ndash4332014
[11] M Moschetta Y Mishima I Sahin et al ldquoRole of endothelialprogenitor cells in cancer progressionrdquo Biochimica et BiophysicaActamdashReviews on Cancer vol 1846 no 1 pp 26ndash39 2014
[12] WZhuo L JiaN Song et al ldquoTheCXCL12ndashCXCR4 chemokinepathway a novel axis regulates lymphangiogenesisrdquo ClinicalCancer Research vol 18 no 19 pp 5387ndash5398 2012
[13] B Li W Bai P Sun B Zhou B Hu and J Ying ldquoThe effectof CXCL12 on endothelial progenitor cells potential target forangiogenesis in intracerebral hemorrhagerdquo Journal of Interferonand Cytokine Research vol 35 no 1 pp 23ndash31 2015
[14] J Liu M Pan Y Li D Ye and Y Guo ldquoClinical study of oralliquid Jin Fu Kang for the treatment of primary non-small celllung cancerrdquo Tumor vol 21 no 6 pp 463ndash465 2001
[15] J Liu Z Shi H Li et al ldquoClinical observation on 271 cases ofnon-small lung cancer treated with Yifei Kangliu Yin (Jin-Fu-Kang)rdquo Chinese Journal of Integrated Traditional and WesternMedicine vol 7 no 4 pp 247ndash250 2001
[16] Z CWang J L Zeng H L Zhang and JW Li ldquoJin FuKang ontumor growth and metastasis and vascular endothelial growthfactor of patients with advanced non-small cell lung cancerrdquoPractical Journal of Clinical Medicine vol 8 no 4 pp 148ndash1492011
[17] P Salven A Orpana and H Joensuu ldquoLeukocytes and plateletsof patients with cancer contain high levels of vascular endothe-lial growth factorrdquo Clinical Cancer Research vol 5 no 3 pp487ndash491 1999
[18] H-J Wang D Zhang Y-Z Tan and T Li ldquoAutophagy inendothelial progenitor cells is cytoprotective in hypoxic con-ditionsrdquo American Journal of PhysiologymdashCell Physiology vol304 no 7 pp C617ndashC626 2013
[19] M Tawada S-I Hayashi S Osada S Nakashima and KYoshida ldquoHuman gastric cancer organizes neighboring lym-phatic vessels via recruitment of bone marrow-derived lym-phatic endothelial progenitor cellsrdquo Journal of Gastroenterologyvol 47 no 9 pp 1057ndash1060 2012
[20] AMasuda H Yasuoka T Satoh Y Okazaki Y Yamaguchi andM Kuwana ldquoVersican is upregulated in circulating monocytesin patients with systemic sclerosis and amplifies a CCL2-mediated pathogenic looprdquo Arthritis Research andTherapy vol15 article R74 2013
[21] J Goldman T X Le M Skobe andM A Swartz ldquoOverexpres-sion of VEGF-C causes transient lymphatic hyperplasia but notincreased lymphangiogenesis in regenerating skinrdquo CirculationResearch vol 96 no 11 pp 1193ndash1199 2005
[22] S Nishida and H Satoh ldquoMechanisms for the vasodilationsinduced by Ginkgo biloba extract and its main constituentbilobalide in rat aortardquo Life Sciences vol 72 no 23 pp 2659ndash2667 2003
[23] B Wang L Zhu and Q Chen ldquoPrimary study on the applica-tion of Serum Pharmacology in Chinese traditional medicinerdquoColloids and Surfaces B Biointerfaces vol 43 no 3-4 pp 194ndash197 2005
[24] F Ma A Morancho J Montaner and A Rosell ldquoEndothelialprogenitor cells and revascularization following strokerdquo BrainResearch vol 1623 pp 150ndash159 2015
[25] H Qiu L Cao D Wang H Xu and Z Liang ldquoHigh levelsof circulatingCD34+VEGFR3+ lymphaticvascular endothelialprogenitor cells is correlated with lymph node metastasis inpatients with epithelial ovarian cancerrdquo Journal of Obstetrics andGynaecology Research vol 39 no 7 pp 1268ndash1275 2013
[26] B Furusato A Mohamed M Uhlen and J S Rhim ldquoCXCR4and cancer review Articlerdquo Pathology International vol 60 no7 pp 497ndash505 2010
[27] E Karshovska D Zagorac A Zernecke C Weber and ASchober ldquoA small molecule CXCR4 antagonist inhibits neoin-tima formation and smoothmuscle progenitor cellmobilizationafter arterial injuryrdquo Journal ofThrombosis andHaemostasis vol6 no 10 pp 1812ndash1815 2008
[28] A C Chiang and J Massague ldquoMolecular basis of metastasisrdquoTheNew England Journal of Medicine vol 359 no 26 pp 2814ndash2823 2008
[29] T Li G-DWang Y-Z Tan andH-JWang ldquoInhibition of lym-phangiogenesis of endothelial progenitor cells with VEGFR-3siRNA delivered with PEI-alginate nanoparticlesrdquo InternationalJournal of Biological Sciences vol 10 no 2 pp 160ndash170 2014
[30] M Wang and Z Zou ldquoMultiple mechanisms of SDF-1 promot-ing VEGF-induced endothelial differentiation of mesenchymalstem cellsrdquo International Journal of Cardiology vol 177 no 3pp 1098ndash1099 2014

Evidence-Based Complementary and Alternative Medicine 3
mouse anti-human VEGFR-3 for 30 minutes at 4∘C Sub-sequently the cells were washed with PBS containing 2FBS and then resuspended with EGM-2MV medium ina 5mL centrifuge tube [18] CD34+VEGFR-3+ cells werecollected by cell sorting with a Beckman MoFlo XDPFACS (fluorescence-activated cell sorter Beckman CoulterFullerton CA USA)
27 Induction of Cell Differentiation Firstly the LECs wereinduced with SDF-1 The freshly sorted CD34+VEGFR-3+ cells were suspended in EGM-2MV medium contain-ing 60 ngmL SDF-1 20 FBS 100UmL penicillin and100 120583gmL streptomycin and seeded in a 24-well plate at adensity of 1 times 105 cellswell The cells were incubated for 14days in a humidified incubator at 37∘C 5 CO
2 and the
medium was changed every 3 daysThe LECs were also induced with VEGF-C and SDF-1 +
VEGF-C with the same protocol as described above exceptthat VEGF-C (60 ngmL) or SDF-1 (100 ngmL) + VEGF-C(60 ngmL) were used instead of SDF-1
28 Transmission Electron Microscopy After induction withSDF-1 VEGF-C or SDF-1 +VEGF-C for 14 days the differen-tiated cells were fixed with 25 glutaraldehyde overnight at4∘Cand thenpostfixedwith 1osmium tetroxideAfter beingdehydrated with gradient alcohol the cells were soaked withanhydrous acetone and Spurr resin and embeddedwith Spurrresin Ultrathin sections were stained with 3 uranyl acetateand lead citrate [10]The ultrastructural characteristics of thecells were examined by using a JEOL JEM-1010 transmissionelectron microscope
29 Immunofluorescence All the LECs differentiated fromthe CD34+VEGFR-3+ cells in the presences of SDF-1 VEGF-C or SDF-1 + VEGF-C and each JFK-containing serumwere identified with expression of lymphatic endotheliumspecific markers LYVE-1 as reported method [19] Brieflyafter induction for 14 days as described above the cellswere fixed by 4 paraformaldehyde and then incubated withrabbit anti-human LYVE-1 antibody (1 100) overnight at 4∘CAfter washing the cells were incubated with FITC-labelledgoat anti-rabbit IgG (1 200) for 60 minutes at 37∘C Thenuclei were counterstained with DAPIThe cells were viewedby using confocal laser scanning microscope Fluorescenceimages and integral optical density (IOD) values were furtheranalyzed with Image-Pro Plus 60 software
210 Transwell Migration Assay The migration efficiency ofLECs was assessed using 8mm pore Transwell filter mem-brane (Corning) following reported method [20] Briefly theLECs were seeded at a density of 2 times 105 cellsmL in theupper chamber and in the lower chamber 100 ngmL of SDF-1 was added After incubation for 16 h the cells on two sidesof the membrane were dried in air and then fixed with 4paraformaldehyde for 15min Subsequently the cells werestained with DAPI for 10min The cells that migrated to thelower sides of the membrane were quantified by countingin 5 fields under a Nikon TE2000 inverted fluorescencemicroscope [21] The same protocol was also performed in
the presences of VEGF-C (60 ngmL) or SDF-1 (100 ngmL) +VEGF-C (60 ngmL)
211 MTT Assay To assess the cytotoxicity of the JFK-containing serums MTT [3-(45-dimethylthiazol-2-yl)-25-diphenyl-2H-tetrazoliumbromide] assaywas performedTheCD34+VEGFR-3+ cells solution (100120583L) containing 10(vv) of the JFK serumswas seeded in a 96-well plate at a den-sity of 1times 104 cellswell and three parallel wells for each groupThe cells were cultured for 48 h as described above After that10 120583L of MTT dye solution (5mgmL in phosphate bufferedsaline pH 74) was added to each well and the plates werefurther incubated for 4 h After incubation the supernatantwas aspirated and the formazan crystals were dissolved inDMSOand the optical density (OD)wasmeasured at 490 nm
212 Inhibition of JFK on LECs Formation To test the effect ofJFK the CD34+VEGFR-3+ cells were divided into 5 groupscontrol group the cells were cultured with blank serum JFK-1 JFK-2 and JFK-3 groups the cells were cultured with JFK-containing serums prepared with JFK at doses of 54 108and 216 gkgday respectively and 100 120583L of each serum wasadded in 09mL of culture medium and AMD group thecells were culturedwith AMD3100 (10 120583M) a SDF-1 inhibitoras a positive control SDF-1 (100 ngmL) was added to allgroups as described previously
The same protocol as in the above was also conducted inwhich VEGF-C (60 ngmL) or SDF-1 (100 ngmL) + VEGF-C(60 ngmL) were used instead of SDF-1 Lenvatinib (10 120583M)a VEGF-C inhibitor or AMD3000 + Lenvatinib (each 10 120583M)were used as positive controls respectively
213 Inhibition of JFK on LECs Migration To test the effectof JFK-containing serums Transwell migration assay wasapplied as described previously In the lower chamber SDF-1(100 ngmL) was addedThe LECs were divided into 5 groupsas described above and pretreated with the blank serumJFK-containing serums (10 vv of culture medium) andAMD3000 (10 120583M) for 30 minutes respectively
The same protocol as in the above was also performedand only VEGF-C (60 ngmL) or SDF-1 (100 ngmL) +VEGF-C (60 ngmL) were added in the lower chamberinstead of SDF-1 Lenvatinib (10 120583M) and AMD3000 +Lenvatinib (each 10 120583M) were used as positive controls
214 Statistical Analysis Data were expressed as means plusmn SDDifferences between experimental groups were assessed bythe two-tailed 119905-test using SPSS 190 software All analyseswere performed using the SPSS 1150 software package andprobability values of 005 or less were considered to bestatistically significant
3 Results
31 Preparation of JFK-Containing Serums As describedpreviously JFK has been used for the treatment of NSCLC inChina JFK is made from 12 Chinese herbal drugs (Table 1)In order to get real effective compositions of JFK workingin vivo an animal experiment using rats was conducted to
4 Evidence-Based Complementary and Alternative Medicine
083
FITC-CD34
APC
-VEG
FR-3
103
102
101
100
100
101
102 10
3
(a)
FITC-IgG
APC
-IgG
103
102
101
100
100
101
102 10
3
(b)
Figure 1 Sorting of CD34+VEGFR-3+ cellsThemononuclear cells were analyzed for the expressions of CD34 andVEGFR-3 with dual-colorflow cytometry (a) and percentage of the CD34+VEGFR-3+ cells was compared with isotype control (b)
Table 1 Composition of Jin Fu Kang oral liquid (JFK)
Herbal drug Part usedAstragalus membranaceus RootGlehnia littoralis RootAsparagus cochinchinensis RootLigustrum lucidum FruitSelaginella doederleinii Whole plantParis polyphylla RootEpimedium sagittatum LeafGynostemma pentaphyllum LeafCornus officinalis FruitSalvia chinensis Whole plantOphiopogon japonicus RootTrigonella foenum graecum Seed
prepare JFK-containing serums Three doses of 18 36 and72 gkg (equivalent to raw herbal drugs) were applied for theserum preparations and the dose of 36 gkg was equal toclinical used one
32 Sorting and Detecting of CD34+VEGFR-3+ Cells Themononuclear cells isolated from cord blood are round havinground- ellipse- or horseshoe-shaped nuclei After beingstained with FITC-conjugated mouse anti-human CD34 andAPC-conjugated mouse anti-human VEGFR-3 the diameterof the cells coexpressing CD34 and VEGFR-3 was 10ndash12 120583mBy double-color flow cytometric analysis the frequency ofCD34+VEGFR-3+ cells was 083 plusmn 021 in the mononu-clear cells (Figures 1(a) and 1(b)) and the freshly sortedCD34+VEGFR-3+ EPCs were round or oval
33 SDF-1 and VEGF-C Additively Induced LECs FormationDifferentiation of CD34+VEGFR-3+ cells towards LECs was
(a) (b)
(c)
Figure 2Morphological changes of CD34+VEGFR-3+ cells duringdifferentiation At day 7 after induction with SDF-1 most cellsbecame spindle-shaped (a) At day 14 the confluentmonolayer of thecells demonstrated a typical cobblestone appearance (b) There aremore mitochondria (white arrow) in the cytoplasm Weibel-Paladebody (black arrow) enwrapped by the membrane is observable (c)
firstly induced in the presence of SDF-1 As described abovethe fresh sorted CD34+VEGFR-3+ cells were round oroval while at day 3 after induction with SDF-1 some cellsrepresented spindle shape or polygon shape At day 7 themost cells were long spindle-shaped (Figure 2(a)) At the endof the experiment (day 14) the cells displayed polygonal shapeand grew into confluent monolayer (Figure 2(b))
To understand the ultrastructural characteristics of thecells a transmission electron microscope examination was
Evidence-Based Complementary and Alternative Medicine 5
performed The observation revealed that the nuclei of thecells were large and there are more mitochondria andphagocytic vesicles in the cytoplasm Furthermore as canbe seen in Figure 2(c) Weibel-Palade body the unique rod-shaped secretory organelle of vascular endothelial cells wasclearly exhibited
As lymphatic vessel endothelial hyaluronan receptor-1 (LYVE-1) is a specific marker of LECs an immunoflu-orescence analysis was conducted with LYVE-1 antibodyThe results showed that after induction for 2 weeks in thepresence of SDF-1 almost all of the cells were positive forLYVE-1 immunostaining (Figure 3(b)) while without SDF-1stimulation (control) only a few LYVE-1 positive LECs wereobserved (Figure 3(a))
VEGF-C was also used for the formation of LECs fromCD34+VEGFR-3+ cellsThe result demonstrated thatVEGF-C induced the differentiation of CD34+VEGFR-3+ EPCstowards LECs while the number of LECs was a little bit morecompared with that of SDF-1 (Figure 3(c))
As shown in Figure 3(d) the LYVE-1 positive cell numberin SDF-1 + VEGF-C group was significantly greater thanthose of SDF-1 or VEGF-C aloneThe integral optical density(IOD) value of LYVE-1 fluorescence in SDF-1 + VEGF-C group (391828) was almost the sum of those in SDF-1(195995) and VEGF-C (236695) groups (Figure 3(e)) Theresults revealed that compared with SDF-1 or VEGF-C alonecostimulation with SDF-1 and VEGF-C displayed an additiveeffect on induction of LECs formation
34 SDF-1 and VEGF-C Promote Migration of LECs but NotAdditively In cell migration assay LECs were seeded in theupper chamber of a Transwell insert and firstly SDF-1 wasadded in the lower chamber As shown in Figures 4(a) and4(b) in the presence of SDF-1 the number of the LECs thatmigrated from the upper chamber to the lower chamber wassignificantly greater comparedwith that in no addition groupThis result clearly indicated that SDF-1 induced the LECsmigration VEGF-C treatment also displayed the same effectas SDF-1 (Figure 4(c))
However when SDF-1 and VEGF-C were cousedalthough a little bitmore LECsmigrated to the lower chambercomparedwith those in SDF-1 orVEGF-C alone (Figures 4(d)and 4(e)) the cell number of SDF-1 + VEGF-C (638 cells) wasfar less than the sum of SDF-1 (511 cells) and VEGF-C (471cells) The results clearly indicated that costimulation withSDF-1 and VEGF-C did not result in an additive effect on thecell transmigration
35 Cytotoxicity of JFK-Containing Serums To verify thecytotoxicity of JFK-containing serums a MTT assay wasperformed As shown in Figure 5 blank serum showed noeffects on EPCs and all the JFK-containing serum samples(10 concentration vv serum to culture medium) exhibitedno cytotoxicity on the CD34+VEGFR-3+ EPCs during 2daysrsquo culture period Thus the 10 concentration of JFK-containing serums was used for the following bioassays
36 Inhibitory Effects of JFK on the LECs Formation As JFKwas used for the treatment of NSCLC to understand whether
(a) (b)
(c) (d)
0
100000
Control SDF-1 VEGF-C VEGF-C + SDF-1
200000
300000
400000
500000IO
Dlowast
lowastlowast
(e)
Figure 3 LECs with positive LYVE-1 immunostaining differen-tiated from CD34+VEGFR-3+ EPCs The CD34+VEGFR-3+ cellswere cultured in the presence of SDF-1 (100 ngmL) and VEGF-C (60 ngmL) The cells expressed LYVE-1 marker after inductionwith no addition ((a) control) SDF-1 (b) VEGF-C (c) and SDF-1 +VEGF-C (d) for two weeks Integral optical density (IOD) of LYVE-1 fluorescence of each group (e) Values are expressed as mean plusmn SD119899 = 3 lowast119901 lt 001 significant increase versus control group
JFK possesses the inhibitory effect on LECs formation JFK-containing serums were testedThe result revealed that blankserum (BS) showed no impact on LECs formation (data notshown) As can be seen in Figure 6 and Table 2 in the positivecontrol AMD3100 (AMD) a SDF-1 inhibitor significantlysuppressed the IODvalues of LYVE-1 about 9373 comparedwith SDF-1 only (BS) As expected all of the JFK-containingserums decreased IOD of LYVE-1 in particular the serumof 72 gkg reduced IOD value about 8279 These resultssuggested JFK-containing serums inhibited LECs formation
The same as SDF-1 the LECs formations induced byVEGF-C and SDF-1 + VEGF-C were also suppressed by JFK-containing serums
6 Evidence-Based Complementary and Alternative Medicine
(a) (b)
(c) (d)
Control SDF-1 VEGF-C VEGF-C + SDF-10
200
400
600
800
The m
igra
ting
cells
lowast
lowastlowast
(e)
Figure 4 Transmigrated LECs in the lower chamber In the lower chamber no addition ((a) control) SDF-1 (100 ngmL (b)) VEGF-C (60 ngmL (c)) and SDF-1 (100 ngmL) + VEGF-C (100 ngmL) (d) were used The migrated cells number in each group (e) Data areexpressed as mean plusmn SD 119899 = 3 lowast119901 lt 001 significant increase versus control group
Table 2 Inhibitory effect of JFK on LECs formation
Group Inhibition rate ()SDF-1 VEGF-C SDF-1 + VEGF-C
BS 000 plusmn 1440 000 plusmn 561 000 plusmn 1829
JFK-1 4410 plusmn 680lowast
3889 plusmn 630lowast
5882 plusmn 1237lowast
JFK-2 6057 plusmn 451lowast
7971 plusmn 131lowast
7606 plusmn 954lowast
JFK-3 8279 plusmn 644lowast
8914 plusmn 143lowast
9116 plusmn 231lowast
AMD 9373 plusmn 060lowast
Len 8718 plusmn 580lowast
Len + AMD 9633 plusmn 060lowast
BS blank serum JFK-1 JFK-2 and JFK-3 the serums prepared from ratsby oral administration of JFK at doses of 18 36 and 72 gkg respectivelyAMD AMD3100 Len Lenvatinib The inhibition rates of BS groups werepegged as 000 and others were calculated relative to it Data wereexpressed as mean plusmn SD 119899 = 3 lowast119901 lt 001 compared with the BS grouprespectively
37 Inhibitory Effects of JFK on the LECsMigration As tumorcell migration is an extremely important process in cancermetastasis the effect of JFK was further evaluated on the
Table 3 Inhibitory effect of JFK on LECs migration
Group Inhibition rate ()SDF-1 VEGF-C VEGF-C + SDF-1
BS 000 plusmn 547 000 plusmn 728 000 plusmn 552
JFK-1 4419 plusmn 503lowast
3281 plusmn 1136lowast
3496 plusmn 541lowast
JFK-2 6603 plusmn 282lowast
6987 plusmn 308lowast
6706 plusmn 549lowast
JFK-3 8049 plusmn 527lowast
8222 plusmn 570lowast
7204 plusmn 277lowast
AMD 8705 plusmn 379lowast
Len 8469 plusmn 390lowast
Len + AMD 8482 plusmn 209lowast
BS blank serum JFK-1 JFK-2 and JFK-3 the serums prepared from ratsby oral administration of JFK at doses of 18 36 and 72 gkg respectivelyAMD AMD3100 Len Lenvatinib The inhibition rates of BS groups werepegged as 000 and others were calculated relative to it Data wereexpressed as mean plusmn SD 119899 = 3 lowast119901 lt 001 compared with the BS grouprespectively
LECs migration As shown in Figure 7 and Table 3 the cellmigration assay data clearly exhibited that JFK-containingserums prepared at all doses demonstrated a significant
Evidence-Based Complementary and Alternative Medicine 7
Control BS JFK-1 JFK-2 JFK-340
60
80
100
120
Cel
l sur
viva
l rat
e (
)
Figure 5 Cytotoxicity of JFK-containing serums The EPCs werecultured with no addition (control) blank serum (BS) and JFK-1JFK-2 and JFK-3 JFK-1 JFK-2 and JFK-3 represented the serumsprepared from rats by oral administration of JFK at doses of 18 36and 72 gkg respectively and 10 (vv) of the serums were used
inhibitory effect on the SDF-1 and VEGF-C induced migra-tions Furthermore the LECs migration induced with SDF-1+ VEGF-C was also markedly suppressed by JFK-containingserums in a dose dependent manner Blank serum showedno effect on the migration (data not shown) The SDF-1 andVEGF-C inducedmigrations were markedly blocked by theirinhibitors AMD3100 or Lenvatinib respectively
4 Discussion
In traditional bioassays in vitro on traditional Chinesemedicines generally extracts or simply purified componentsof crude drugs are directly added to the culture system [22]However the pharmacological actions of traditional Chinesemedicines are rather complicated due to their complexcompositions More important thing is that many of thecomponents contained in the crud drugs do not directlyexhibit their pharmacological actions until they experience aseries of biotransformation during absorption distributionand metabolite procedures Therefore without biotransfor-mation the results from the bioassays in vitromight not trulyreflect the pharmacological actions in vivo Furthermore theserum drug concentration directly reflects the drug efficacyin most cases Therefore serum pharmacology in which thedrug-containing serum was prepared in vivo was used forbioactivity evaluation and research on traditional Chinesemedicines in vitro and the results were more scientific andexact compared with traditional pharmacology [23] In thecurrent study the JFK-containing serums were prepared withrats and applied for the in vitro assays
EPCs have been extensively studied as a potential markerfor endothelial regeneration ability and shown to be atherapeutic potential by directly enhancing angiogenesis
or forming new vessels through vasculogenesis EPCs alsoexhibit a character to be highly migratory to ischemicareas [24] It is reported that circulating CD34+VEGFR-3+lymphaticvascular endothelial progenitor cells correlatewithlymph node metastasis in patients with epithelial ovariancancer [25] and further study revealed that CD34+VEGFR-3+ EPCs in human cord blood could differentiate into lym-phatic endothelial cells [10] Flow cytometry is widely usedto quantify and study circulating EPCs while EPC-specificmarkers are essential [11] Therefore in the current studyCD34 and VEGFR-3 were applied for the sorting of the targetEPCs from human cord blood and the results demonstratedthat the frequency of the CD34+VEGFR-3+ cells was 083in the mononuclear cells which was consistent with thereported data [18]
It is well accepted that LECs contribute to lymphangio-genesis that plays an active role in the induction of metastasisto the lymph nodes in cancers CXC chemokine receptor 4(CXCR4) plays a central role in tumor cell dissemination andmetastasis development in more than 75 of all cancers [26]SDF-1 also known as chemokine CXC ligand binds to theCXCR4 receptor The SDF-1CXCR4 axis is essential for themigration of progenitor cells tissue regeneration and vascu-larization [27] Furthermore SDF-1CXCR4 axis significantlyassociates with lymph node metastasis and promotes lym-phangiogenesis in tumors [28] VEGF-CVEGFR-3 signalingis a keymodulator of the lymphatic system andVEGF-C levelcorrelates with lymph node metastasis and poor prognosis[9] VEGF-C promotes LEC proliferation migration andtube formation [10] VEGFR-3 siRNA effectively inhibitsdifferentiation and lymphangiogenesis of CD34+VEGFR-3+EPCs [29] Therefore in the present study we used SDF-1and VEGF-C to induce the LECs formation and migrationOur results clearly demonstrated that SDF-1 and VEGF-Cpromoted the CD34+VEGFR-3+ EPCs to differentiate to theLECs that showed positive LYVE-1 staining a specific LECmarker and also enhanced the LECs migration
Although it is reported that VEGF-C stimulation upreg-ulates the expression level of CXCR4 on lymphangiogenicendothelial cells SDF-1CXCR4 axis exerts its function ina VEGFR-3 independent pattern [12] More interestinglytargeting both SDF-1 and VEGF-C by neutralizing antibodiesresulted in an additive inhibitory effect on both tumor lym-phangiogenesis and lymphatic metastasis [12] Furthermoreit is hypothesized that SDF-1 and VEGF may have syner-getic effect on endothelial differentiation through a positivefeedback loop with mutual promotion [30] Thus to clarifythe coeffect of SDF-1 and VEGF-C we added both in theculture system and the data revealed that costimulation withSDF-1 and VEGF-C displayed an additive effect on the LECsdifferentiation however no additive effect was observedon the LECs migration which needs a further mechanismclarification
Studies have shown that JFK can prevent tumor growthand progression and inhibit tumor angiogenesis [16] How-ever there is no data onwhether JFK inhibits tumor lymphan-giogenesis The present work provided the first experimentalevidence that JFK suppressed the formation and directionalmigration of LECs in vitro which closely related to the tumor
8 Evidence-Based Complementary and Alternative Medicine
JFK-3BSSD
F-1
SDF-
1SD
F-1
JFK-1 JFK-2 AMDM
erge
DA
PILY
VE-
1
(a1) (a2) (a3) (a4) (a5)
(b1) (b2) (b3) (b4) (b5)
(c1) (c2) (c3) (c4) (c5)JFK-3BS
VEG
F-C
VEG
F-C
VEG
F-C
JFK-1 JFK-2 LEN
(A1)
(B1)
(C1) (C2) (C3) (C4) (C5)
(B2) (B3) (B4) (B5)
(A2) (A3) (A4) (A5)
Mer
geD
API
LYV
E-1
(a1) (a2) (a3) (a4) (a5)
(b1) (b2) (b3) (b4) (b5)
(A1)
(B1) (B2) (B3) (B4) (B5)
(A2) (A3) (A4) (A5)
100120583m
100120583m
100120583m
Figure 6 Effect of JFK on LYVE-1 positive LECs differentiation The LECs formation was induced from CD34+VEGFR-3+ EPCs withSDF-1 (100 ngmL) and VEGF-C (60 ngmL) respectively BS blank serum JFK-1 JFK-2 and JFK-3 the serums prepared from rats by oraladministration of JFK at doses of 18 36 and 72 gkg respectively and 10 (vv) of the serums were added AMD AMD3100 (10 120583M) LenLenvatinib (10 120583M)
Evidence-Based Complementary and Alternative Medicine 9
AMDV
EGF-
CSD
F-1
JFK-2JFK-1 JFK-3BS
LenJFK-2JFK-1 JFK-3BS
200120583m
200120583m
(A1)
(B1) (B2) (B3) (B4) (B5)
(A2) (A3) (A4) (A5)
Figure 7 Effect of JFK on LECs transmigration The LECs migration was induced with SDF-1 (100 ngmL) and VEGF-C (60 ngmL)respectively The LECs were pretreated with blank serum (BS) the serums (each 10 vv) prepared from rats by oral administration ofJFK at doses of 18 36 and 72 gkg (JFK-1 JFK-2 and JFK-3 resp) AMD3100 (AMD 10 120583M) Lenvatinib (Len 10 120583M)
lymphangiogenesis Furthermore the inhibitory effect mightbe via regulation of SDF-1CXCR4 and VEGF-CVEGFR-3axes As JFK is composed of 12 herbal drugs further efforton chemical isolation and structure identification of JFK isessential to clarify the bioactive components responsible forthe effect and might provide a potential to discover the anti-lymphangiogenesis agents from the natural resources
5 Conclusion
This study demonstrated that SDF-1 and VEGF-C inducedCD34+VEGFR-3+ EPCs to differentiate towards LECs andenhanced the LECsmigration SDF-1 andVEGF-C exerted anadditive role on the LECs formation but not on themigrationJFK inhibited the differentiation and migration of LECs viaregulation of the SDF-1CXCR4 and VEGF-CVEGFR-3 axesat least The current finding suggested that JFK might inhibitNSCLC through antilymphangiogenesis and also provided apotential to discover antilymphangiogenesis agents Furtherresearch on the inhibitory effect of JFK on lymphangiogenesisin vivo and clarification of its bioactive components areundergoing in our lab
Competing Interests
The authors declare that they have no conflict of interests
Acknowledgments
This work was supported by Research Project for PracticeDevelopment of National TCM Clinical Research Bases(JDZX2012117) and the Project of Key Discipline for TCMConstruction of Jiangsu Province China (no JS1302)
References
[1] R A Smith D Manassaram-Baptiste D Brooks et al ldquoCancerscreening in the United States 2015 a review of currentAmerican Cancer Society guidelines and current issues incancer screeningrdquo CA A Cancer Journal for Clinicians vol 65no 1 pp 30ndash54 2015
[2] T K Kilvaer E-E Paulsen S M Hald et al ldquoLymphangiogenicmarkers and their impact on nodal metastasis and survival innon-small cell lung cancermdasha structured review with meta-analysisrdquo PLoS ONE vol 10 no 8 Article ID e0132481 2015
[3] F G Gomes F Nedel A M Alves J E Nor and S BC Tarquinio ldquoTumor angiogenesis and lymphangiogenesistumorendothelial crosstalk and cellularmicroenvironmentalsignaling mechanismsrdquo Life Sciences vol 92 no 2 pp 101ndash1072013
[4] Y Wang S Ma M Dong Y Yao K Liu and J Zhou ldquoEvalua-tion of the factors affecting the maximum standardized uptakevalue of metastatic lymph nodes in different histological typesof non-small cell lung cancer on PET-CTrdquo BMC PulmonaryMedicine vol 15 no 1 article 20 2015
[5] J P Sleeman and W Thiele ldquoTumor metastasis and the lym-phatic vasculaturerdquo International Journal of Cancer vol 125 no12 pp 2747ndash2756 2009
[6] S Li and Q Li ldquoCancer stem cells lymphangiogenesis andlymphatic metastasisrdquo Cancer Letters vol 357 no 2 pp 438ndash447 2015
[7] M Bzowska R Męzyk-Kopec T Prochnicki M KuleszaT Klaus and J Bereta ldquoAntibody-based antiangiogenic andantilymphangiogenic therapies to prevent tumor growth andprogressionrdquo Acta Biochimica Polonica vol 60 no 3 pp 263ndash275 2013
[8] S A Stacker S P Williams T Karnezis R Shayan S B Foxand M G Achen ldquoLymphangiogenesis and lymphatic vesselremodelling in cancerrdquo Nature Reviews Cancer vol 14 no 3pp 159ndash172 2014
[9] J Yu X Zhang P M Kuzontkoski et al ldquoSlit2N and Robo4regulate lymphangiogenesis through the VEGF-CVEGFR-3
10 Evidence-Based Complementary and Alternative Medicine
pathwayrdquo Cell Communication and Signaling vol 12 no 1article 25 pp 1ndash15 2014
[10] Y-Z Tan H-J Wang M-H Zhang Z Quan T Li and Q-Z He ldquoCD34+VEGFR-3+ progenitor cells have a potential todifferentiate towards lymphatic endothelial cellsrdquo Journal ofCellular and Molecular Medicine vol 18 no 3 pp 422ndash4332014
[11] M Moschetta Y Mishima I Sahin et al ldquoRole of endothelialprogenitor cells in cancer progressionrdquo Biochimica et BiophysicaActamdashReviews on Cancer vol 1846 no 1 pp 26ndash39 2014
[12] WZhuo L JiaN Song et al ldquoTheCXCL12ndashCXCR4 chemokinepathway a novel axis regulates lymphangiogenesisrdquo ClinicalCancer Research vol 18 no 19 pp 5387ndash5398 2012
[13] B Li W Bai P Sun B Zhou B Hu and J Ying ldquoThe effectof CXCL12 on endothelial progenitor cells potential target forangiogenesis in intracerebral hemorrhagerdquo Journal of Interferonand Cytokine Research vol 35 no 1 pp 23ndash31 2015
[14] J Liu M Pan Y Li D Ye and Y Guo ldquoClinical study of oralliquid Jin Fu Kang for the treatment of primary non-small celllung cancerrdquo Tumor vol 21 no 6 pp 463ndash465 2001
[15] J Liu Z Shi H Li et al ldquoClinical observation on 271 cases ofnon-small lung cancer treated with Yifei Kangliu Yin (Jin-Fu-Kang)rdquo Chinese Journal of Integrated Traditional and WesternMedicine vol 7 no 4 pp 247ndash250 2001
[16] Z CWang J L Zeng H L Zhang and JW Li ldquoJin FuKang ontumor growth and metastasis and vascular endothelial growthfactor of patients with advanced non-small cell lung cancerrdquoPractical Journal of Clinical Medicine vol 8 no 4 pp 148ndash1492011
[17] P Salven A Orpana and H Joensuu ldquoLeukocytes and plateletsof patients with cancer contain high levels of vascular endothe-lial growth factorrdquo Clinical Cancer Research vol 5 no 3 pp487ndash491 1999
[18] H-J Wang D Zhang Y-Z Tan and T Li ldquoAutophagy inendothelial progenitor cells is cytoprotective in hypoxic con-ditionsrdquo American Journal of PhysiologymdashCell Physiology vol304 no 7 pp C617ndashC626 2013
[19] M Tawada S-I Hayashi S Osada S Nakashima and KYoshida ldquoHuman gastric cancer organizes neighboring lym-phatic vessels via recruitment of bone marrow-derived lym-phatic endothelial progenitor cellsrdquo Journal of Gastroenterologyvol 47 no 9 pp 1057ndash1060 2012
[20] AMasuda H Yasuoka T Satoh Y Okazaki Y Yamaguchi andM Kuwana ldquoVersican is upregulated in circulating monocytesin patients with systemic sclerosis and amplifies a CCL2-mediated pathogenic looprdquo Arthritis Research andTherapy vol15 article R74 2013
[21] J Goldman T X Le M Skobe andM A Swartz ldquoOverexpres-sion of VEGF-C causes transient lymphatic hyperplasia but notincreased lymphangiogenesis in regenerating skinrdquo CirculationResearch vol 96 no 11 pp 1193ndash1199 2005
[22] S Nishida and H Satoh ldquoMechanisms for the vasodilationsinduced by Ginkgo biloba extract and its main constituentbilobalide in rat aortardquo Life Sciences vol 72 no 23 pp 2659ndash2667 2003
[23] B Wang L Zhu and Q Chen ldquoPrimary study on the applica-tion of Serum Pharmacology in Chinese traditional medicinerdquoColloids and Surfaces B Biointerfaces vol 43 no 3-4 pp 194ndash197 2005
[24] F Ma A Morancho J Montaner and A Rosell ldquoEndothelialprogenitor cells and revascularization following strokerdquo BrainResearch vol 1623 pp 150ndash159 2015
[25] H Qiu L Cao D Wang H Xu and Z Liang ldquoHigh levelsof circulatingCD34+VEGFR3+ lymphaticvascular endothelialprogenitor cells is correlated with lymph node metastasis inpatients with epithelial ovarian cancerrdquo Journal of Obstetrics andGynaecology Research vol 39 no 7 pp 1268ndash1275 2013
[26] B Furusato A Mohamed M Uhlen and J S Rhim ldquoCXCR4and cancer review Articlerdquo Pathology International vol 60 no7 pp 497ndash505 2010
[27] E Karshovska D Zagorac A Zernecke C Weber and ASchober ldquoA small molecule CXCR4 antagonist inhibits neoin-tima formation and smoothmuscle progenitor cellmobilizationafter arterial injuryrdquo Journal ofThrombosis andHaemostasis vol6 no 10 pp 1812ndash1815 2008
[28] A C Chiang and J Massague ldquoMolecular basis of metastasisrdquoTheNew England Journal of Medicine vol 359 no 26 pp 2814ndash2823 2008
[29] T Li G-DWang Y-Z Tan andH-JWang ldquoInhibition of lym-phangiogenesis of endothelial progenitor cells with VEGFR-3siRNA delivered with PEI-alginate nanoparticlesrdquo InternationalJournal of Biological Sciences vol 10 no 2 pp 160ndash170 2014
[30] M Wang and Z Zou ldquoMultiple mechanisms of SDF-1 promot-ing VEGF-induced endothelial differentiation of mesenchymalstem cellsrdquo International Journal of Cardiology vol 177 no 3pp 1098ndash1099 2014

4 Evidence-Based Complementary and Alternative Medicine
083
FITC-CD34
APC
-VEG
FR-3
103
102
101
100
100
101
102 10
3
(a)
FITC-IgG
APC
-IgG
103
102
101
100
100
101
102 10
3
(b)
Figure 1 Sorting of CD34+VEGFR-3+ cellsThemononuclear cells were analyzed for the expressions of CD34 andVEGFR-3 with dual-colorflow cytometry (a) and percentage of the CD34+VEGFR-3+ cells was compared with isotype control (b)
Table 1 Composition of Jin Fu Kang oral liquid (JFK)
Herbal drug Part usedAstragalus membranaceus RootGlehnia littoralis RootAsparagus cochinchinensis RootLigustrum lucidum FruitSelaginella doederleinii Whole plantParis polyphylla RootEpimedium sagittatum LeafGynostemma pentaphyllum LeafCornus officinalis FruitSalvia chinensis Whole plantOphiopogon japonicus RootTrigonella foenum graecum Seed
prepare JFK-containing serums Three doses of 18 36 and72 gkg (equivalent to raw herbal drugs) were applied for theserum preparations and the dose of 36 gkg was equal toclinical used one
32 Sorting and Detecting of CD34+VEGFR-3+ Cells Themononuclear cells isolated from cord blood are round havinground- ellipse- or horseshoe-shaped nuclei After beingstained with FITC-conjugated mouse anti-human CD34 andAPC-conjugated mouse anti-human VEGFR-3 the diameterof the cells coexpressing CD34 and VEGFR-3 was 10ndash12 120583mBy double-color flow cytometric analysis the frequency ofCD34+VEGFR-3+ cells was 083 plusmn 021 in the mononu-clear cells (Figures 1(a) and 1(b)) and the freshly sortedCD34+VEGFR-3+ EPCs were round or oval
33 SDF-1 and VEGF-C Additively Induced LECs FormationDifferentiation of CD34+VEGFR-3+ cells towards LECs was
(a) (b)
(c)
Figure 2Morphological changes of CD34+VEGFR-3+ cells duringdifferentiation At day 7 after induction with SDF-1 most cellsbecame spindle-shaped (a) At day 14 the confluentmonolayer of thecells demonstrated a typical cobblestone appearance (b) There aremore mitochondria (white arrow) in the cytoplasm Weibel-Paladebody (black arrow) enwrapped by the membrane is observable (c)
firstly induced in the presence of SDF-1 As described abovethe fresh sorted CD34+VEGFR-3+ cells were round oroval while at day 3 after induction with SDF-1 some cellsrepresented spindle shape or polygon shape At day 7 themost cells were long spindle-shaped (Figure 2(a)) At the endof the experiment (day 14) the cells displayed polygonal shapeand grew into confluent monolayer (Figure 2(b))
To understand the ultrastructural characteristics of thecells a transmission electron microscope examination was
Evidence-Based Complementary and Alternative Medicine 5
performed The observation revealed that the nuclei of thecells were large and there are more mitochondria andphagocytic vesicles in the cytoplasm Furthermore as canbe seen in Figure 2(c) Weibel-Palade body the unique rod-shaped secretory organelle of vascular endothelial cells wasclearly exhibited
As lymphatic vessel endothelial hyaluronan receptor-1 (LYVE-1) is a specific marker of LECs an immunoflu-orescence analysis was conducted with LYVE-1 antibodyThe results showed that after induction for 2 weeks in thepresence of SDF-1 almost all of the cells were positive forLYVE-1 immunostaining (Figure 3(b)) while without SDF-1stimulation (control) only a few LYVE-1 positive LECs wereobserved (Figure 3(a))
VEGF-C was also used for the formation of LECs fromCD34+VEGFR-3+ cellsThe result demonstrated thatVEGF-C induced the differentiation of CD34+VEGFR-3+ EPCstowards LECs while the number of LECs was a little bit morecompared with that of SDF-1 (Figure 3(c))
As shown in Figure 3(d) the LYVE-1 positive cell numberin SDF-1 + VEGF-C group was significantly greater thanthose of SDF-1 or VEGF-C aloneThe integral optical density(IOD) value of LYVE-1 fluorescence in SDF-1 + VEGF-C group (391828) was almost the sum of those in SDF-1(195995) and VEGF-C (236695) groups (Figure 3(e)) Theresults revealed that compared with SDF-1 or VEGF-C alonecostimulation with SDF-1 and VEGF-C displayed an additiveeffect on induction of LECs formation
34 SDF-1 and VEGF-C Promote Migration of LECs but NotAdditively In cell migration assay LECs were seeded in theupper chamber of a Transwell insert and firstly SDF-1 wasadded in the lower chamber As shown in Figures 4(a) and4(b) in the presence of SDF-1 the number of the LECs thatmigrated from the upper chamber to the lower chamber wassignificantly greater comparedwith that in no addition groupThis result clearly indicated that SDF-1 induced the LECsmigration VEGF-C treatment also displayed the same effectas SDF-1 (Figure 4(c))
However when SDF-1 and VEGF-C were cousedalthough a little bitmore LECsmigrated to the lower chambercomparedwith those in SDF-1 orVEGF-C alone (Figures 4(d)and 4(e)) the cell number of SDF-1 + VEGF-C (638 cells) wasfar less than the sum of SDF-1 (511 cells) and VEGF-C (471cells) The results clearly indicated that costimulation withSDF-1 and VEGF-C did not result in an additive effect on thecell transmigration
35 Cytotoxicity of JFK-Containing Serums To verify thecytotoxicity of JFK-containing serums a MTT assay wasperformed As shown in Figure 5 blank serum showed noeffects on EPCs and all the JFK-containing serum samples(10 concentration vv serum to culture medium) exhibitedno cytotoxicity on the CD34+VEGFR-3+ EPCs during 2daysrsquo culture period Thus the 10 concentration of JFK-containing serums was used for the following bioassays
36 Inhibitory Effects of JFK on the LECs Formation As JFKwas used for the treatment of NSCLC to understand whether
(a) (b)
(c) (d)
0
100000
Control SDF-1 VEGF-C VEGF-C + SDF-1
200000
300000
400000
500000IO
Dlowast
lowastlowast
(e)
Figure 3 LECs with positive LYVE-1 immunostaining differen-tiated from CD34+VEGFR-3+ EPCs The CD34+VEGFR-3+ cellswere cultured in the presence of SDF-1 (100 ngmL) and VEGF-C (60 ngmL) The cells expressed LYVE-1 marker after inductionwith no addition ((a) control) SDF-1 (b) VEGF-C (c) and SDF-1 +VEGF-C (d) for two weeks Integral optical density (IOD) of LYVE-1 fluorescence of each group (e) Values are expressed as mean plusmn SD119899 = 3 lowast119901 lt 001 significant increase versus control group
JFK possesses the inhibitory effect on LECs formation JFK-containing serums were testedThe result revealed that blankserum (BS) showed no impact on LECs formation (data notshown) As can be seen in Figure 6 and Table 2 in the positivecontrol AMD3100 (AMD) a SDF-1 inhibitor significantlysuppressed the IODvalues of LYVE-1 about 9373 comparedwith SDF-1 only (BS) As expected all of the JFK-containingserums decreased IOD of LYVE-1 in particular the serumof 72 gkg reduced IOD value about 8279 These resultssuggested JFK-containing serums inhibited LECs formation
The same as SDF-1 the LECs formations induced byVEGF-C and SDF-1 + VEGF-C were also suppressed by JFK-containing serums
6 Evidence-Based Complementary and Alternative Medicine
(a) (b)
(c) (d)
Control SDF-1 VEGF-C VEGF-C + SDF-10
200
400
600
800
The m
igra
ting
cells
lowast
lowastlowast
(e)
Figure 4 Transmigrated LECs in the lower chamber In the lower chamber no addition ((a) control) SDF-1 (100 ngmL (b)) VEGF-C (60 ngmL (c)) and SDF-1 (100 ngmL) + VEGF-C (100 ngmL) (d) were used The migrated cells number in each group (e) Data areexpressed as mean plusmn SD 119899 = 3 lowast119901 lt 001 significant increase versus control group
Table 2 Inhibitory effect of JFK on LECs formation
Group Inhibition rate ()SDF-1 VEGF-C SDF-1 + VEGF-C
BS 000 plusmn 1440 000 plusmn 561 000 plusmn 1829
JFK-1 4410 plusmn 680lowast
3889 plusmn 630lowast
5882 plusmn 1237lowast
JFK-2 6057 plusmn 451lowast
7971 plusmn 131lowast
7606 plusmn 954lowast
JFK-3 8279 plusmn 644lowast
8914 plusmn 143lowast
9116 plusmn 231lowast
AMD 9373 plusmn 060lowast
Len 8718 plusmn 580lowast
Len + AMD 9633 plusmn 060lowast
BS blank serum JFK-1 JFK-2 and JFK-3 the serums prepared from ratsby oral administration of JFK at doses of 18 36 and 72 gkg respectivelyAMD AMD3100 Len Lenvatinib The inhibition rates of BS groups werepegged as 000 and others were calculated relative to it Data wereexpressed as mean plusmn SD 119899 = 3 lowast119901 lt 001 compared with the BS grouprespectively
37 Inhibitory Effects of JFK on the LECsMigration As tumorcell migration is an extremely important process in cancermetastasis the effect of JFK was further evaluated on the
Table 3 Inhibitory effect of JFK on LECs migration
Group Inhibition rate ()SDF-1 VEGF-C VEGF-C + SDF-1
BS 000 plusmn 547 000 plusmn 728 000 plusmn 552
JFK-1 4419 plusmn 503lowast
3281 plusmn 1136lowast
3496 plusmn 541lowast
JFK-2 6603 plusmn 282lowast
6987 plusmn 308lowast
6706 plusmn 549lowast
JFK-3 8049 plusmn 527lowast
8222 plusmn 570lowast
7204 plusmn 277lowast
AMD 8705 plusmn 379lowast
Len 8469 plusmn 390lowast
Len + AMD 8482 plusmn 209lowast
BS blank serum JFK-1 JFK-2 and JFK-3 the serums prepared from ratsby oral administration of JFK at doses of 18 36 and 72 gkg respectivelyAMD AMD3100 Len Lenvatinib The inhibition rates of BS groups werepegged as 000 and others were calculated relative to it Data wereexpressed as mean plusmn SD 119899 = 3 lowast119901 lt 001 compared with the BS grouprespectively
LECs migration As shown in Figure 7 and Table 3 the cellmigration assay data clearly exhibited that JFK-containingserums prepared at all doses demonstrated a significant
Evidence-Based Complementary and Alternative Medicine 7
Control BS JFK-1 JFK-2 JFK-340
60
80
100
120
Cel
l sur
viva
l rat
e (
)
Figure 5 Cytotoxicity of JFK-containing serums The EPCs werecultured with no addition (control) blank serum (BS) and JFK-1JFK-2 and JFK-3 JFK-1 JFK-2 and JFK-3 represented the serumsprepared from rats by oral administration of JFK at doses of 18 36and 72 gkg respectively and 10 (vv) of the serums were used
inhibitory effect on the SDF-1 and VEGF-C induced migra-tions Furthermore the LECs migration induced with SDF-1+ VEGF-C was also markedly suppressed by JFK-containingserums in a dose dependent manner Blank serum showedno effect on the migration (data not shown) The SDF-1 andVEGF-C inducedmigrations were markedly blocked by theirinhibitors AMD3100 or Lenvatinib respectively
4 Discussion
In traditional bioassays in vitro on traditional Chinesemedicines generally extracts or simply purified componentsof crude drugs are directly added to the culture system [22]However the pharmacological actions of traditional Chinesemedicines are rather complicated due to their complexcompositions More important thing is that many of thecomponents contained in the crud drugs do not directlyexhibit their pharmacological actions until they experience aseries of biotransformation during absorption distributionand metabolite procedures Therefore without biotransfor-mation the results from the bioassays in vitromight not trulyreflect the pharmacological actions in vivo Furthermore theserum drug concentration directly reflects the drug efficacyin most cases Therefore serum pharmacology in which thedrug-containing serum was prepared in vivo was used forbioactivity evaluation and research on traditional Chinesemedicines in vitro and the results were more scientific andexact compared with traditional pharmacology [23] In thecurrent study the JFK-containing serums were prepared withrats and applied for the in vitro assays
EPCs have been extensively studied as a potential markerfor endothelial regeneration ability and shown to be atherapeutic potential by directly enhancing angiogenesis
or forming new vessels through vasculogenesis EPCs alsoexhibit a character to be highly migratory to ischemicareas [24] It is reported that circulating CD34+VEGFR-3+lymphaticvascular endothelial progenitor cells correlatewithlymph node metastasis in patients with epithelial ovariancancer [25] and further study revealed that CD34+VEGFR-3+ EPCs in human cord blood could differentiate into lym-phatic endothelial cells [10] Flow cytometry is widely usedto quantify and study circulating EPCs while EPC-specificmarkers are essential [11] Therefore in the current studyCD34 and VEGFR-3 were applied for the sorting of the targetEPCs from human cord blood and the results demonstratedthat the frequency of the CD34+VEGFR-3+ cells was 083in the mononuclear cells which was consistent with thereported data [18]
It is well accepted that LECs contribute to lymphangio-genesis that plays an active role in the induction of metastasisto the lymph nodes in cancers CXC chemokine receptor 4(CXCR4) plays a central role in tumor cell dissemination andmetastasis development in more than 75 of all cancers [26]SDF-1 also known as chemokine CXC ligand binds to theCXCR4 receptor The SDF-1CXCR4 axis is essential for themigration of progenitor cells tissue regeneration and vascu-larization [27] Furthermore SDF-1CXCR4 axis significantlyassociates with lymph node metastasis and promotes lym-phangiogenesis in tumors [28] VEGF-CVEGFR-3 signalingis a keymodulator of the lymphatic system andVEGF-C levelcorrelates with lymph node metastasis and poor prognosis[9] VEGF-C promotes LEC proliferation migration andtube formation [10] VEGFR-3 siRNA effectively inhibitsdifferentiation and lymphangiogenesis of CD34+VEGFR-3+EPCs [29] Therefore in the present study we used SDF-1and VEGF-C to induce the LECs formation and migrationOur results clearly demonstrated that SDF-1 and VEGF-Cpromoted the CD34+VEGFR-3+ EPCs to differentiate to theLECs that showed positive LYVE-1 staining a specific LECmarker and also enhanced the LECs migration
Although it is reported that VEGF-C stimulation upreg-ulates the expression level of CXCR4 on lymphangiogenicendothelial cells SDF-1CXCR4 axis exerts its function ina VEGFR-3 independent pattern [12] More interestinglytargeting both SDF-1 and VEGF-C by neutralizing antibodiesresulted in an additive inhibitory effect on both tumor lym-phangiogenesis and lymphatic metastasis [12] Furthermoreit is hypothesized that SDF-1 and VEGF may have syner-getic effect on endothelial differentiation through a positivefeedback loop with mutual promotion [30] Thus to clarifythe coeffect of SDF-1 and VEGF-C we added both in theculture system and the data revealed that costimulation withSDF-1 and VEGF-C displayed an additive effect on the LECsdifferentiation however no additive effect was observedon the LECs migration which needs a further mechanismclarification
Studies have shown that JFK can prevent tumor growthand progression and inhibit tumor angiogenesis [16] How-ever there is no data onwhether JFK inhibits tumor lymphan-giogenesis The present work provided the first experimentalevidence that JFK suppressed the formation and directionalmigration of LECs in vitro which closely related to the tumor
8 Evidence-Based Complementary and Alternative Medicine
JFK-3BSSD
F-1
SDF-
1SD
F-1
JFK-1 JFK-2 AMDM
erge
DA
PILY
VE-
1
(a1) (a2) (a3) (a4) (a5)
(b1) (b2) (b3) (b4) (b5)
(c1) (c2) (c3) (c4) (c5)JFK-3BS
VEG
F-C
VEG
F-C
VEG
F-C
JFK-1 JFK-2 LEN
(A1)
(B1)
(C1) (C2) (C3) (C4) (C5)
(B2) (B3) (B4) (B5)
(A2) (A3) (A4) (A5)
Mer
geD
API
LYV
E-1
(a1) (a2) (a3) (a4) (a5)
(b1) (b2) (b3) (b4) (b5)
(A1)
(B1) (B2) (B3) (B4) (B5)
(A2) (A3) (A4) (A5)
100120583m
100120583m
100120583m
Figure 6 Effect of JFK on LYVE-1 positive LECs differentiation The LECs formation was induced from CD34+VEGFR-3+ EPCs withSDF-1 (100 ngmL) and VEGF-C (60 ngmL) respectively BS blank serum JFK-1 JFK-2 and JFK-3 the serums prepared from rats by oraladministration of JFK at doses of 18 36 and 72 gkg respectively and 10 (vv) of the serums were added AMD AMD3100 (10 120583M) LenLenvatinib (10 120583M)
Evidence-Based Complementary and Alternative Medicine 9
AMDV
EGF-
CSD
F-1
JFK-2JFK-1 JFK-3BS
LenJFK-2JFK-1 JFK-3BS
200120583m
200120583m
(A1)
(B1) (B2) (B3) (B4) (B5)
(A2) (A3) (A4) (A5)
Figure 7 Effect of JFK on LECs transmigration The LECs migration was induced with SDF-1 (100 ngmL) and VEGF-C (60 ngmL)respectively The LECs were pretreated with blank serum (BS) the serums (each 10 vv) prepared from rats by oral administration ofJFK at doses of 18 36 and 72 gkg (JFK-1 JFK-2 and JFK-3 resp) AMD3100 (AMD 10 120583M) Lenvatinib (Len 10 120583M)
lymphangiogenesis Furthermore the inhibitory effect mightbe via regulation of SDF-1CXCR4 and VEGF-CVEGFR-3axes As JFK is composed of 12 herbal drugs further efforton chemical isolation and structure identification of JFK isessential to clarify the bioactive components responsible forthe effect and might provide a potential to discover the anti-lymphangiogenesis agents from the natural resources
5 Conclusion
This study demonstrated that SDF-1 and VEGF-C inducedCD34+VEGFR-3+ EPCs to differentiate towards LECs andenhanced the LECsmigration SDF-1 andVEGF-C exerted anadditive role on the LECs formation but not on themigrationJFK inhibited the differentiation and migration of LECs viaregulation of the SDF-1CXCR4 and VEGF-CVEGFR-3 axesat least The current finding suggested that JFK might inhibitNSCLC through antilymphangiogenesis and also provided apotential to discover antilymphangiogenesis agents Furtherresearch on the inhibitory effect of JFK on lymphangiogenesisin vivo and clarification of its bioactive components areundergoing in our lab
Competing Interests
The authors declare that they have no conflict of interests
Acknowledgments
This work was supported by Research Project for PracticeDevelopment of National TCM Clinical Research Bases(JDZX2012117) and the Project of Key Discipline for TCMConstruction of Jiangsu Province China (no JS1302)
References
[1] R A Smith D Manassaram-Baptiste D Brooks et al ldquoCancerscreening in the United States 2015 a review of currentAmerican Cancer Society guidelines and current issues incancer screeningrdquo CA A Cancer Journal for Clinicians vol 65no 1 pp 30ndash54 2015
[2] T K Kilvaer E-E Paulsen S M Hald et al ldquoLymphangiogenicmarkers and their impact on nodal metastasis and survival innon-small cell lung cancermdasha structured review with meta-analysisrdquo PLoS ONE vol 10 no 8 Article ID e0132481 2015
[3] F G Gomes F Nedel A M Alves J E Nor and S BC Tarquinio ldquoTumor angiogenesis and lymphangiogenesistumorendothelial crosstalk and cellularmicroenvironmentalsignaling mechanismsrdquo Life Sciences vol 92 no 2 pp 101ndash1072013
[4] Y Wang S Ma M Dong Y Yao K Liu and J Zhou ldquoEvalua-tion of the factors affecting the maximum standardized uptakevalue of metastatic lymph nodes in different histological typesof non-small cell lung cancer on PET-CTrdquo BMC PulmonaryMedicine vol 15 no 1 article 20 2015
[5] J P Sleeman and W Thiele ldquoTumor metastasis and the lym-phatic vasculaturerdquo International Journal of Cancer vol 125 no12 pp 2747ndash2756 2009
[6] S Li and Q Li ldquoCancer stem cells lymphangiogenesis andlymphatic metastasisrdquo Cancer Letters vol 357 no 2 pp 438ndash447 2015
[7] M Bzowska R Męzyk-Kopec T Prochnicki M KuleszaT Klaus and J Bereta ldquoAntibody-based antiangiogenic andantilymphangiogenic therapies to prevent tumor growth andprogressionrdquo Acta Biochimica Polonica vol 60 no 3 pp 263ndash275 2013
[8] S A Stacker S P Williams T Karnezis R Shayan S B Foxand M G Achen ldquoLymphangiogenesis and lymphatic vesselremodelling in cancerrdquo Nature Reviews Cancer vol 14 no 3pp 159ndash172 2014
[9] J Yu X Zhang P M Kuzontkoski et al ldquoSlit2N and Robo4regulate lymphangiogenesis through the VEGF-CVEGFR-3
10 Evidence-Based Complementary and Alternative Medicine
pathwayrdquo Cell Communication and Signaling vol 12 no 1article 25 pp 1ndash15 2014
[10] Y-Z Tan H-J Wang M-H Zhang Z Quan T Li and Q-Z He ldquoCD34+VEGFR-3+ progenitor cells have a potential todifferentiate towards lymphatic endothelial cellsrdquo Journal ofCellular and Molecular Medicine vol 18 no 3 pp 422ndash4332014
[11] M Moschetta Y Mishima I Sahin et al ldquoRole of endothelialprogenitor cells in cancer progressionrdquo Biochimica et BiophysicaActamdashReviews on Cancer vol 1846 no 1 pp 26ndash39 2014
[12] WZhuo L JiaN Song et al ldquoTheCXCL12ndashCXCR4 chemokinepathway a novel axis regulates lymphangiogenesisrdquo ClinicalCancer Research vol 18 no 19 pp 5387ndash5398 2012
[13] B Li W Bai P Sun B Zhou B Hu and J Ying ldquoThe effectof CXCL12 on endothelial progenitor cells potential target forangiogenesis in intracerebral hemorrhagerdquo Journal of Interferonand Cytokine Research vol 35 no 1 pp 23ndash31 2015
[14] J Liu M Pan Y Li D Ye and Y Guo ldquoClinical study of oralliquid Jin Fu Kang for the treatment of primary non-small celllung cancerrdquo Tumor vol 21 no 6 pp 463ndash465 2001
[15] J Liu Z Shi H Li et al ldquoClinical observation on 271 cases ofnon-small lung cancer treated with Yifei Kangliu Yin (Jin-Fu-Kang)rdquo Chinese Journal of Integrated Traditional and WesternMedicine vol 7 no 4 pp 247ndash250 2001
[16] Z CWang J L Zeng H L Zhang and JW Li ldquoJin FuKang ontumor growth and metastasis and vascular endothelial growthfactor of patients with advanced non-small cell lung cancerrdquoPractical Journal of Clinical Medicine vol 8 no 4 pp 148ndash1492011
[17] P Salven A Orpana and H Joensuu ldquoLeukocytes and plateletsof patients with cancer contain high levels of vascular endothe-lial growth factorrdquo Clinical Cancer Research vol 5 no 3 pp487ndash491 1999
[18] H-J Wang D Zhang Y-Z Tan and T Li ldquoAutophagy inendothelial progenitor cells is cytoprotective in hypoxic con-ditionsrdquo American Journal of PhysiologymdashCell Physiology vol304 no 7 pp C617ndashC626 2013
[19] M Tawada S-I Hayashi S Osada S Nakashima and KYoshida ldquoHuman gastric cancer organizes neighboring lym-phatic vessels via recruitment of bone marrow-derived lym-phatic endothelial progenitor cellsrdquo Journal of Gastroenterologyvol 47 no 9 pp 1057ndash1060 2012
[20] AMasuda H Yasuoka T Satoh Y Okazaki Y Yamaguchi andM Kuwana ldquoVersican is upregulated in circulating monocytesin patients with systemic sclerosis and amplifies a CCL2-mediated pathogenic looprdquo Arthritis Research andTherapy vol15 article R74 2013
[21] J Goldman T X Le M Skobe andM A Swartz ldquoOverexpres-sion of VEGF-C causes transient lymphatic hyperplasia but notincreased lymphangiogenesis in regenerating skinrdquo CirculationResearch vol 96 no 11 pp 1193ndash1199 2005
[22] S Nishida and H Satoh ldquoMechanisms for the vasodilationsinduced by Ginkgo biloba extract and its main constituentbilobalide in rat aortardquo Life Sciences vol 72 no 23 pp 2659ndash2667 2003
[23] B Wang L Zhu and Q Chen ldquoPrimary study on the applica-tion of Serum Pharmacology in Chinese traditional medicinerdquoColloids and Surfaces B Biointerfaces vol 43 no 3-4 pp 194ndash197 2005
[24] F Ma A Morancho J Montaner and A Rosell ldquoEndothelialprogenitor cells and revascularization following strokerdquo BrainResearch vol 1623 pp 150ndash159 2015
[25] H Qiu L Cao D Wang H Xu and Z Liang ldquoHigh levelsof circulatingCD34+VEGFR3+ lymphaticvascular endothelialprogenitor cells is correlated with lymph node metastasis inpatients with epithelial ovarian cancerrdquo Journal of Obstetrics andGynaecology Research vol 39 no 7 pp 1268ndash1275 2013
[26] B Furusato A Mohamed M Uhlen and J S Rhim ldquoCXCR4and cancer review Articlerdquo Pathology International vol 60 no7 pp 497ndash505 2010
[27] E Karshovska D Zagorac A Zernecke C Weber and ASchober ldquoA small molecule CXCR4 antagonist inhibits neoin-tima formation and smoothmuscle progenitor cellmobilizationafter arterial injuryrdquo Journal ofThrombosis andHaemostasis vol6 no 10 pp 1812ndash1815 2008
[28] A C Chiang and J Massague ldquoMolecular basis of metastasisrdquoTheNew England Journal of Medicine vol 359 no 26 pp 2814ndash2823 2008
[29] T Li G-DWang Y-Z Tan andH-JWang ldquoInhibition of lym-phangiogenesis of endothelial progenitor cells with VEGFR-3siRNA delivered with PEI-alginate nanoparticlesrdquo InternationalJournal of Biological Sciences vol 10 no 2 pp 160ndash170 2014
[30] M Wang and Z Zou ldquoMultiple mechanisms of SDF-1 promot-ing VEGF-induced endothelial differentiation of mesenchymalstem cellsrdquo International Journal of Cardiology vol 177 no 3pp 1098ndash1099 2014

Evidence-Based Complementary and Alternative Medicine 5
performed The observation revealed that the nuclei of thecells were large and there are more mitochondria andphagocytic vesicles in the cytoplasm Furthermore as canbe seen in Figure 2(c) Weibel-Palade body the unique rod-shaped secretory organelle of vascular endothelial cells wasclearly exhibited
As lymphatic vessel endothelial hyaluronan receptor-1 (LYVE-1) is a specific marker of LECs an immunoflu-orescence analysis was conducted with LYVE-1 antibodyThe results showed that after induction for 2 weeks in thepresence of SDF-1 almost all of the cells were positive forLYVE-1 immunostaining (Figure 3(b)) while without SDF-1stimulation (control) only a few LYVE-1 positive LECs wereobserved (Figure 3(a))
VEGF-C was also used for the formation of LECs fromCD34+VEGFR-3+ cellsThe result demonstrated thatVEGF-C induced the differentiation of CD34+VEGFR-3+ EPCstowards LECs while the number of LECs was a little bit morecompared with that of SDF-1 (Figure 3(c))
As shown in Figure 3(d) the LYVE-1 positive cell numberin SDF-1 + VEGF-C group was significantly greater thanthose of SDF-1 or VEGF-C aloneThe integral optical density(IOD) value of LYVE-1 fluorescence in SDF-1 + VEGF-C group (391828) was almost the sum of those in SDF-1(195995) and VEGF-C (236695) groups (Figure 3(e)) Theresults revealed that compared with SDF-1 or VEGF-C alonecostimulation with SDF-1 and VEGF-C displayed an additiveeffect on induction of LECs formation
34 SDF-1 and VEGF-C Promote Migration of LECs but NotAdditively In cell migration assay LECs were seeded in theupper chamber of a Transwell insert and firstly SDF-1 wasadded in the lower chamber As shown in Figures 4(a) and4(b) in the presence of SDF-1 the number of the LECs thatmigrated from the upper chamber to the lower chamber wassignificantly greater comparedwith that in no addition groupThis result clearly indicated that SDF-1 induced the LECsmigration VEGF-C treatment also displayed the same effectas SDF-1 (Figure 4(c))
However when SDF-1 and VEGF-C were cousedalthough a little bitmore LECsmigrated to the lower chambercomparedwith those in SDF-1 orVEGF-C alone (Figures 4(d)and 4(e)) the cell number of SDF-1 + VEGF-C (638 cells) wasfar less than the sum of SDF-1 (511 cells) and VEGF-C (471cells) The results clearly indicated that costimulation withSDF-1 and VEGF-C did not result in an additive effect on thecell transmigration
35 Cytotoxicity of JFK-Containing Serums To verify thecytotoxicity of JFK-containing serums a MTT assay wasperformed As shown in Figure 5 blank serum showed noeffects on EPCs and all the JFK-containing serum samples(10 concentration vv serum to culture medium) exhibitedno cytotoxicity on the CD34+VEGFR-3+ EPCs during 2daysrsquo culture period Thus the 10 concentration of JFK-containing serums was used for the following bioassays
36 Inhibitory Effects of JFK on the LECs Formation As JFKwas used for the treatment of NSCLC to understand whether
(a) (b)
(c) (d)
0
100000
Control SDF-1 VEGF-C VEGF-C + SDF-1
200000
300000
400000
500000IO
Dlowast
lowastlowast
(e)
Figure 3 LECs with positive LYVE-1 immunostaining differen-tiated from CD34+VEGFR-3+ EPCs The CD34+VEGFR-3+ cellswere cultured in the presence of SDF-1 (100 ngmL) and VEGF-C (60 ngmL) The cells expressed LYVE-1 marker after inductionwith no addition ((a) control) SDF-1 (b) VEGF-C (c) and SDF-1 +VEGF-C (d) for two weeks Integral optical density (IOD) of LYVE-1 fluorescence of each group (e) Values are expressed as mean plusmn SD119899 = 3 lowast119901 lt 001 significant increase versus control group
JFK possesses the inhibitory effect on LECs formation JFK-containing serums were testedThe result revealed that blankserum (BS) showed no impact on LECs formation (data notshown) As can be seen in Figure 6 and Table 2 in the positivecontrol AMD3100 (AMD) a SDF-1 inhibitor significantlysuppressed the IODvalues of LYVE-1 about 9373 comparedwith SDF-1 only (BS) As expected all of the JFK-containingserums decreased IOD of LYVE-1 in particular the serumof 72 gkg reduced IOD value about 8279 These resultssuggested JFK-containing serums inhibited LECs formation
The same as SDF-1 the LECs formations induced byVEGF-C and SDF-1 + VEGF-C were also suppressed by JFK-containing serums
6 Evidence-Based Complementary and Alternative Medicine
(a) (b)
(c) (d)
Control SDF-1 VEGF-C VEGF-C + SDF-10
200
400
600
800
The m
igra
ting
cells
lowast
lowastlowast
(e)
Figure 4 Transmigrated LECs in the lower chamber In the lower chamber no addition ((a) control) SDF-1 (100 ngmL (b)) VEGF-C (60 ngmL (c)) and SDF-1 (100 ngmL) + VEGF-C (100 ngmL) (d) were used The migrated cells number in each group (e) Data areexpressed as mean plusmn SD 119899 = 3 lowast119901 lt 001 significant increase versus control group
Table 2 Inhibitory effect of JFK on LECs formation
Group Inhibition rate ()SDF-1 VEGF-C SDF-1 + VEGF-C
BS 000 plusmn 1440 000 plusmn 561 000 plusmn 1829
JFK-1 4410 plusmn 680lowast
3889 plusmn 630lowast
5882 plusmn 1237lowast
JFK-2 6057 plusmn 451lowast
7971 plusmn 131lowast
7606 plusmn 954lowast
JFK-3 8279 plusmn 644lowast
8914 plusmn 143lowast
9116 plusmn 231lowast
AMD 9373 plusmn 060lowast
Len 8718 plusmn 580lowast
Len + AMD 9633 plusmn 060lowast
BS blank serum JFK-1 JFK-2 and JFK-3 the serums prepared from ratsby oral administration of JFK at doses of 18 36 and 72 gkg respectivelyAMD AMD3100 Len Lenvatinib The inhibition rates of BS groups werepegged as 000 and others were calculated relative to it Data wereexpressed as mean plusmn SD 119899 = 3 lowast119901 lt 001 compared with the BS grouprespectively
37 Inhibitory Effects of JFK on the LECsMigration As tumorcell migration is an extremely important process in cancermetastasis the effect of JFK was further evaluated on the
Table 3 Inhibitory effect of JFK on LECs migration
Group Inhibition rate ()SDF-1 VEGF-C VEGF-C + SDF-1
BS 000 plusmn 547 000 plusmn 728 000 plusmn 552
JFK-1 4419 plusmn 503lowast
3281 plusmn 1136lowast
3496 plusmn 541lowast
JFK-2 6603 plusmn 282lowast
6987 plusmn 308lowast
6706 plusmn 549lowast
JFK-3 8049 plusmn 527lowast
8222 plusmn 570lowast
7204 plusmn 277lowast
AMD 8705 plusmn 379lowast
Len 8469 plusmn 390lowast
Len + AMD 8482 plusmn 209lowast
BS blank serum JFK-1 JFK-2 and JFK-3 the serums prepared from ratsby oral administration of JFK at doses of 18 36 and 72 gkg respectivelyAMD AMD3100 Len Lenvatinib The inhibition rates of BS groups werepegged as 000 and others were calculated relative to it Data wereexpressed as mean plusmn SD 119899 = 3 lowast119901 lt 001 compared with the BS grouprespectively
LECs migration As shown in Figure 7 and Table 3 the cellmigration assay data clearly exhibited that JFK-containingserums prepared at all doses demonstrated a significant
Evidence-Based Complementary and Alternative Medicine 7
Control BS JFK-1 JFK-2 JFK-340
60
80
100
120
Cel
l sur
viva
l rat
e (
)
Figure 5 Cytotoxicity of JFK-containing serums The EPCs werecultured with no addition (control) blank serum (BS) and JFK-1JFK-2 and JFK-3 JFK-1 JFK-2 and JFK-3 represented the serumsprepared from rats by oral administration of JFK at doses of 18 36and 72 gkg respectively and 10 (vv) of the serums were used
inhibitory effect on the SDF-1 and VEGF-C induced migra-tions Furthermore the LECs migration induced with SDF-1+ VEGF-C was also markedly suppressed by JFK-containingserums in a dose dependent manner Blank serum showedno effect on the migration (data not shown) The SDF-1 andVEGF-C inducedmigrations were markedly blocked by theirinhibitors AMD3100 or Lenvatinib respectively
4 Discussion
In traditional bioassays in vitro on traditional Chinesemedicines generally extracts or simply purified componentsof crude drugs are directly added to the culture system [22]However the pharmacological actions of traditional Chinesemedicines are rather complicated due to their complexcompositions More important thing is that many of thecomponents contained in the crud drugs do not directlyexhibit their pharmacological actions until they experience aseries of biotransformation during absorption distributionand metabolite procedures Therefore without biotransfor-mation the results from the bioassays in vitromight not trulyreflect the pharmacological actions in vivo Furthermore theserum drug concentration directly reflects the drug efficacyin most cases Therefore serum pharmacology in which thedrug-containing serum was prepared in vivo was used forbioactivity evaluation and research on traditional Chinesemedicines in vitro and the results were more scientific andexact compared with traditional pharmacology [23] In thecurrent study the JFK-containing serums were prepared withrats and applied for the in vitro assays
EPCs have been extensively studied as a potential markerfor endothelial regeneration ability and shown to be atherapeutic potential by directly enhancing angiogenesis
or forming new vessels through vasculogenesis EPCs alsoexhibit a character to be highly migratory to ischemicareas [24] It is reported that circulating CD34+VEGFR-3+lymphaticvascular endothelial progenitor cells correlatewithlymph node metastasis in patients with epithelial ovariancancer [25] and further study revealed that CD34+VEGFR-3+ EPCs in human cord blood could differentiate into lym-phatic endothelial cells [10] Flow cytometry is widely usedto quantify and study circulating EPCs while EPC-specificmarkers are essential [11] Therefore in the current studyCD34 and VEGFR-3 were applied for the sorting of the targetEPCs from human cord blood and the results demonstratedthat the frequency of the CD34+VEGFR-3+ cells was 083in the mononuclear cells which was consistent with thereported data [18]
It is well accepted that LECs contribute to lymphangio-genesis that plays an active role in the induction of metastasisto the lymph nodes in cancers CXC chemokine receptor 4(CXCR4) plays a central role in tumor cell dissemination andmetastasis development in more than 75 of all cancers [26]SDF-1 also known as chemokine CXC ligand binds to theCXCR4 receptor The SDF-1CXCR4 axis is essential for themigration of progenitor cells tissue regeneration and vascu-larization [27] Furthermore SDF-1CXCR4 axis significantlyassociates with lymph node metastasis and promotes lym-phangiogenesis in tumors [28] VEGF-CVEGFR-3 signalingis a keymodulator of the lymphatic system andVEGF-C levelcorrelates with lymph node metastasis and poor prognosis[9] VEGF-C promotes LEC proliferation migration andtube formation [10] VEGFR-3 siRNA effectively inhibitsdifferentiation and lymphangiogenesis of CD34+VEGFR-3+EPCs [29] Therefore in the present study we used SDF-1and VEGF-C to induce the LECs formation and migrationOur results clearly demonstrated that SDF-1 and VEGF-Cpromoted the CD34+VEGFR-3+ EPCs to differentiate to theLECs that showed positive LYVE-1 staining a specific LECmarker and also enhanced the LECs migration
Although it is reported that VEGF-C stimulation upreg-ulates the expression level of CXCR4 on lymphangiogenicendothelial cells SDF-1CXCR4 axis exerts its function ina VEGFR-3 independent pattern [12] More interestinglytargeting both SDF-1 and VEGF-C by neutralizing antibodiesresulted in an additive inhibitory effect on both tumor lym-phangiogenesis and lymphatic metastasis [12] Furthermoreit is hypothesized that SDF-1 and VEGF may have syner-getic effect on endothelial differentiation through a positivefeedback loop with mutual promotion [30] Thus to clarifythe coeffect of SDF-1 and VEGF-C we added both in theculture system and the data revealed that costimulation withSDF-1 and VEGF-C displayed an additive effect on the LECsdifferentiation however no additive effect was observedon the LECs migration which needs a further mechanismclarification
Studies have shown that JFK can prevent tumor growthand progression and inhibit tumor angiogenesis [16] How-ever there is no data onwhether JFK inhibits tumor lymphan-giogenesis The present work provided the first experimentalevidence that JFK suppressed the formation and directionalmigration of LECs in vitro which closely related to the tumor
8 Evidence-Based Complementary and Alternative Medicine
JFK-3BSSD
F-1
SDF-
1SD
F-1
JFK-1 JFK-2 AMDM
erge
DA
PILY
VE-
1
(a1) (a2) (a3) (a4) (a5)
(b1) (b2) (b3) (b4) (b5)
(c1) (c2) (c3) (c4) (c5)JFK-3BS
VEG
F-C
VEG
F-C
VEG
F-C
JFK-1 JFK-2 LEN
(A1)
(B1)
(C1) (C2) (C3) (C4) (C5)
(B2) (B3) (B4) (B5)
(A2) (A3) (A4) (A5)
Mer
geD
API
LYV
E-1
(a1) (a2) (a3) (a4) (a5)
(b1) (b2) (b3) (b4) (b5)
(A1)
(B1) (B2) (B3) (B4) (B5)
(A2) (A3) (A4) (A5)
100120583m
100120583m
100120583m
Figure 6 Effect of JFK on LYVE-1 positive LECs differentiation The LECs formation was induced from CD34+VEGFR-3+ EPCs withSDF-1 (100 ngmL) and VEGF-C (60 ngmL) respectively BS blank serum JFK-1 JFK-2 and JFK-3 the serums prepared from rats by oraladministration of JFK at doses of 18 36 and 72 gkg respectively and 10 (vv) of the serums were added AMD AMD3100 (10 120583M) LenLenvatinib (10 120583M)
Evidence-Based Complementary and Alternative Medicine 9
AMDV
EGF-
CSD
F-1
JFK-2JFK-1 JFK-3BS
LenJFK-2JFK-1 JFK-3BS
200120583m
200120583m
(A1)
(B1) (B2) (B3) (B4) (B5)
(A2) (A3) (A4) (A5)
Figure 7 Effect of JFK on LECs transmigration The LECs migration was induced with SDF-1 (100 ngmL) and VEGF-C (60 ngmL)respectively The LECs were pretreated with blank serum (BS) the serums (each 10 vv) prepared from rats by oral administration ofJFK at doses of 18 36 and 72 gkg (JFK-1 JFK-2 and JFK-3 resp) AMD3100 (AMD 10 120583M) Lenvatinib (Len 10 120583M)
lymphangiogenesis Furthermore the inhibitory effect mightbe via regulation of SDF-1CXCR4 and VEGF-CVEGFR-3axes As JFK is composed of 12 herbal drugs further efforton chemical isolation and structure identification of JFK isessential to clarify the bioactive components responsible forthe effect and might provide a potential to discover the anti-lymphangiogenesis agents from the natural resources
5 Conclusion
This study demonstrated that SDF-1 and VEGF-C inducedCD34+VEGFR-3+ EPCs to differentiate towards LECs andenhanced the LECsmigration SDF-1 andVEGF-C exerted anadditive role on the LECs formation but not on themigrationJFK inhibited the differentiation and migration of LECs viaregulation of the SDF-1CXCR4 and VEGF-CVEGFR-3 axesat least The current finding suggested that JFK might inhibitNSCLC through antilymphangiogenesis and also provided apotential to discover antilymphangiogenesis agents Furtherresearch on the inhibitory effect of JFK on lymphangiogenesisin vivo and clarification of its bioactive components areundergoing in our lab
Competing Interests
The authors declare that they have no conflict of interests
Acknowledgments
This work was supported by Research Project for PracticeDevelopment of National TCM Clinical Research Bases(JDZX2012117) and the Project of Key Discipline for TCMConstruction of Jiangsu Province China (no JS1302)
References
[1] R A Smith D Manassaram-Baptiste D Brooks et al ldquoCancerscreening in the United States 2015 a review of currentAmerican Cancer Society guidelines and current issues incancer screeningrdquo CA A Cancer Journal for Clinicians vol 65no 1 pp 30ndash54 2015
[2] T K Kilvaer E-E Paulsen S M Hald et al ldquoLymphangiogenicmarkers and their impact on nodal metastasis and survival innon-small cell lung cancermdasha structured review with meta-analysisrdquo PLoS ONE vol 10 no 8 Article ID e0132481 2015
[3] F G Gomes F Nedel A M Alves J E Nor and S BC Tarquinio ldquoTumor angiogenesis and lymphangiogenesistumorendothelial crosstalk and cellularmicroenvironmentalsignaling mechanismsrdquo Life Sciences vol 92 no 2 pp 101ndash1072013
[4] Y Wang S Ma M Dong Y Yao K Liu and J Zhou ldquoEvalua-tion of the factors affecting the maximum standardized uptakevalue of metastatic lymph nodes in different histological typesof non-small cell lung cancer on PET-CTrdquo BMC PulmonaryMedicine vol 15 no 1 article 20 2015
[5] J P Sleeman and W Thiele ldquoTumor metastasis and the lym-phatic vasculaturerdquo International Journal of Cancer vol 125 no12 pp 2747ndash2756 2009
[6] S Li and Q Li ldquoCancer stem cells lymphangiogenesis andlymphatic metastasisrdquo Cancer Letters vol 357 no 2 pp 438ndash447 2015
[7] M Bzowska R Męzyk-Kopec T Prochnicki M KuleszaT Klaus and J Bereta ldquoAntibody-based antiangiogenic andantilymphangiogenic therapies to prevent tumor growth andprogressionrdquo Acta Biochimica Polonica vol 60 no 3 pp 263ndash275 2013
[8] S A Stacker S P Williams T Karnezis R Shayan S B Foxand M G Achen ldquoLymphangiogenesis and lymphatic vesselremodelling in cancerrdquo Nature Reviews Cancer vol 14 no 3pp 159ndash172 2014
[9] J Yu X Zhang P M Kuzontkoski et al ldquoSlit2N and Robo4regulate lymphangiogenesis through the VEGF-CVEGFR-3
10 Evidence-Based Complementary and Alternative Medicine
pathwayrdquo Cell Communication and Signaling vol 12 no 1article 25 pp 1ndash15 2014
[10] Y-Z Tan H-J Wang M-H Zhang Z Quan T Li and Q-Z He ldquoCD34+VEGFR-3+ progenitor cells have a potential todifferentiate towards lymphatic endothelial cellsrdquo Journal ofCellular and Molecular Medicine vol 18 no 3 pp 422ndash4332014
[11] M Moschetta Y Mishima I Sahin et al ldquoRole of endothelialprogenitor cells in cancer progressionrdquo Biochimica et BiophysicaActamdashReviews on Cancer vol 1846 no 1 pp 26ndash39 2014
[12] WZhuo L JiaN Song et al ldquoTheCXCL12ndashCXCR4 chemokinepathway a novel axis regulates lymphangiogenesisrdquo ClinicalCancer Research vol 18 no 19 pp 5387ndash5398 2012
[13] B Li W Bai P Sun B Zhou B Hu and J Ying ldquoThe effectof CXCL12 on endothelial progenitor cells potential target forangiogenesis in intracerebral hemorrhagerdquo Journal of Interferonand Cytokine Research vol 35 no 1 pp 23ndash31 2015
[14] J Liu M Pan Y Li D Ye and Y Guo ldquoClinical study of oralliquid Jin Fu Kang for the treatment of primary non-small celllung cancerrdquo Tumor vol 21 no 6 pp 463ndash465 2001
[15] J Liu Z Shi H Li et al ldquoClinical observation on 271 cases ofnon-small lung cancer treated with Yifei Kangliu Yin (Jin-Fu-Kang)rdquo Chinese Journal of Integrated Traditional and WesternMedicine vol 7 no 4 pp 247ndash250 2001
[16] Z CWang J L Zeng H L Zhang and JW Li ldquoJin FuKang ontumor growth and metastasis and vascular endothelial growthfactor of patients with advanced non-small cell lung cancerrdquoPractical Journal of Clinical Medicine vol 8 no 4 pp 148ndash1492011
[17] P Salven A Orpana and H Joensuu ldquoLeukocytes and plateletsof patients with cancer contain high levels of vascular endothe-lial growth factorrdquo Clinical Cancer Research vol 5 no 3 pp487ndash491 1999
[18] H-J Wang D Zhang Y-Z Tan and T Li ldquoAutophagy inendothelial progenitor cells is cytoprotective in hypoxic con-ditionsrdquo American Journal of PhysiologymdashCell Physiology vol304 no 7 pp C617ndashC626 2013
[19] M Tawada S-I Hayashi S Osada S Nakashima and KYoshida ldquoHuman gastric cancer organizes neighboring lym-phatic vessels via recruitment of bone marrow-derived lym-phatic endothelial progenitor cellsrdquo Journal of Gastroenterologyvol 47 no 9 pp 1057ndash1060 2012
[20] AMasuda H Yasuoka T Satoh Y Okazaki Y Yamaguchi andM Kuwana ldquoVersican is upregulated in circulating monocytesin patients with systemic sclerosis and amplifies a CCL2-mediated pathogenic looprdquo Arthritis Research andTherapy vol15 article R74 2013
[21] J Goldman T X Le M Skobe andM A Swartz ldquoOverexpres-sion of VEGF-C causes transient lymphatic hyperplasia but notincreased lymphangiogenesis in regenerating skinrdquo CirculationResearch vol 96 no 11 pp 1193ndash1199 2005
[22] S Nishida and H Satoh ldquoMechanisms for the vasodilationsinduced by Ginkgo biloba extract and its main constituentbilobalide in rat aortardquo Life Sciences vol 72 no 23 pp 2659ndash2667 2003
[23] B Wang L Zhu and Q Chen ldquoPrimary study on the applica-tion of Serum Pharmacology in Chinese traditional medicinerdquoColloids and Surfaces B Biointerfaces vol 43 no 3-4 pp 194ndash197 2005
[24] F Ma A Morancho J Montaner and A Rosell ldquoEndothelialprogenitor cells and revascularization following strokerdquo BrainResearch vol 1623 pp 150ndash159 2015
[25] H Qiu L Cao D Wang H Xu and Z Liang ldquoHigh levelsof circulatingCD34+VEGFR3+ lymphaticvascular endothelialprogenitor cells is correlated with lymph node metastasis inpatients with epithelial ovarian cancerrdquo Journal of Obstetrics andGynaecology Research vol 39 no 7 pp 1268ndash1275 2013
[26] B Furusato A Mohamed M Uhlen and J S Rhim ldquoCXCR4and cancer review Articlerdquo Pathology International vol 60 no7 pp 497ndash505 2010
[27] E Karshovska D Zagorac A Zernecke C Weber and ASchober ldquoA small molecule CXCR4 antagonist inhibits neoin-tima formation and smoothmuscle progenitor cellmobilizationafter arterial injuryrdquo Journal ofThrombosis andHaemostasis vol6 no 10 pp 1812ndash1815 2008
[28] A C Chiang and J Massague ldquoMolecular basis of metastasisrdquoTheNew England Journal of Medicine vol 359 no 26 pp 2814ndash2823 2008
[29] T Li G-DWang Y-Z Tan andH-JWang ldquoInhibition of lym-phangiogenesis of endothelial progenitor cells with VEGFR-3siRNA delivered with PEI-alginate nanoparticlesrdquo InternationalJournal of Biological Sciences vol 10 no 2 pp 160ndash170 2014
[30] M Wang and Z Zou ldquoMultiple mechanisms of SDF-1 promot-ing VEGF-induced endothelial differentiation of mesenchymalstem cellsrdquo International Journal of Cardiology vol 177 no 3pp 1098ndash1099 2014
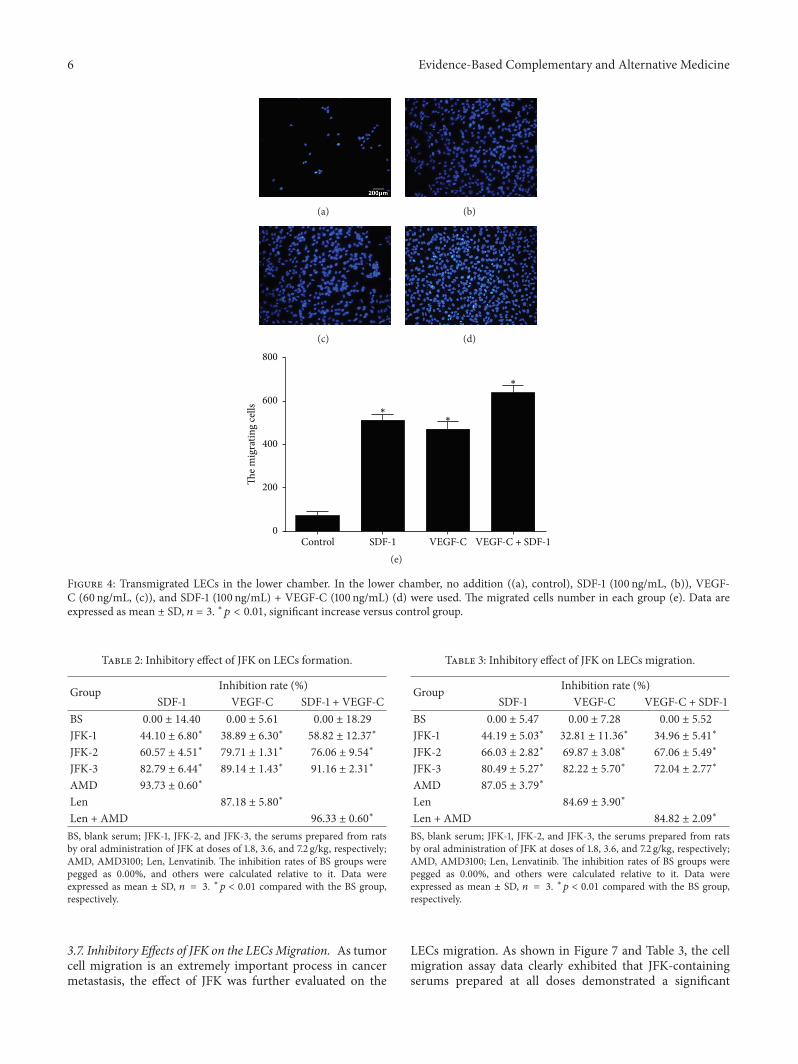
6 Evidence-Based Complementary and Alternative Medicine
(a) (b)
(c) (d)
Control SDF-1 VEGF-C VEGF-C + SDF-10
200
400
600
800
The m
igra
ting
cells
lowast
lowastlowast
(e)
Figure 4 Transmigrated LECs in the lower chamber In the lower chamber no addition ((a) control) SDF-1 (100 ngmL (b)) VEGF-C (60 ngmL (c)) and SDF-1 (100 ngmL) + VEGF-C (100 ngmL) (d) were used The migrated cells number in each group (e) Data areexpressed as mean plusmn SD 119899 = 3 lowast119901 lt 001 significant increase versus control group
Table 2 Inhibitory effect of JFK on LECs formation
Group Inhibition rate ()SDF-1 VEGF-C SDF-1 + VEGF-C
BS 000 plusmn 1440 000 plusmn 561 000 plusmn 1829
JFK-1 4410 plusmn 680lowast
3889 plusmn 630lowast
5882 plusmn 1237lowast
JFK-2 6057 plusmn 451lowast
7971 plusmn 131lowast
7606 plusmn 954lowast
JFK-3 8279 plusmn 644lowast
8914 plusmn 143lowast
9116 plusmn 231lowast
AMD 9373 plusmn 060lowast
Len 8718 plusmn 580lowast
Len + AMD 9633 plusmn 060lowast
BS blank serum JFK-1 JFK-2 and JFK-3 the serums prepared from ratsby oral administration of JFK at doses of 18 36 and 72 gkg respectivelyAMD AMD3100 Len Lenvatinib The inhibition rates of BS groups werepegged as 000 and others were calculated relative to it Data wereexpressed as mean plusmn SD 119899 = 3 lowast119901 lt 001 compared with the BS grouprespectively
37 Inhibitory Effects of JFK on the LECsMigration As tumorcell migration is an extremely important process in cancermetastasis the effect of JFK was further evaluated on the
Table 3 Inhibitory effect of JFK on LECs migration
Group Inhibition rate ()SDF-1 VEGF-C VEGF-C + SDF-1
BS 000 plusmn 547 000 plusmn 728 000 plusmn 552
JFK-1 4419 plusmn 503lowast
3281 plusmn 1136lowast
3496 plusmn 541lowast
JFK-2 6603 plusmn 282lowast
6987 plusmn 308lowast
6706 plusmn 549lowast
JFK-3 8049 plusmn 527lowast
8222 plusmn 570lowast
7204 plusmn 277lowast
AMD 8705 plusmn 379lowast
Len 8469 plusmn 390lowast
Len + AMD 8482 plusmn 209lowast
BS blank serum JFK-1 JFK-2 and JFK-3 the serums prepared from ratsby oral administration of JFK at doses of 18 36 and 72 gkg respectivelyAMD AMD3100 Len Lenvatinib The inhibition rates of BS groups werepegged as 000 and others were calculated relative to it Data wereexpressed as mean plusmn SD 119899 = 3 lowast119901 lt 001 compared with the BS grouprespectively
LECs migration As shown in Figure 7 and Table 3 the cellmigration assay data clearly exhibited that JFK-containingserums prepared at all doses demonstrated a significant
Evidence-Based Complementary and Alternative Medicine 7
Control BS JFK-1 JFK-2 JFK-340
60
80
100
120
Cel
l sur
viva
l rat
e (
)
Figure 5 Cytotoxicity of JFK-containing serums The EPCs werecultured with no addition (control) blank serum (BS) and JFK-1JFK-2 and JFK-3 JFK-1 JFK-2 and JFK-3 represented the serumsprepared from rats by oral administration of JFK at doses of 18 36and 72 gkg respectively and 10 (vv) of the serums were used
inhibitory effect on the SDF-1 and VEGF-C induced migra-tions Furthermore the LECs migration induced with SDF-1+ VEGF-C was also markedly suppressed by JFK-containingserums in a dose dependent manner Blank serum showedno effect on the migration (data not shown) The SDF-1 andVEGF-C inducedmigrations were markedly blocked by theirinhibitors AMD3100 or Lenvatinib respectively
4 Discussion
In traditional bioassays in vitro on traditional Chinesemedicines generally extracts or simply purified componentsof crude drugs are directly added to the culture system [22]However the pharmacological actions of traditional Chinesemedicines are rather complicated due to their complexcompositions More important thing is that many of thecomponents contained in the crud drugs do not directlyexhibit their pharmacological actions until they experience aseries of biotransformation during absorption distributionand metabolite procedures Therefore without biotransfor-mation the results from the bioassays in vitromight not trulyreflect the pharmacological actions in vivo Furthermore theserum drug concentration directly reflects the drug efficacyin most cases Therefore serum pharmacology in which thedrug-containing serum was prepared in vivo was used forbioactivity evaluation and research on traditional Chinesemedicines in vitro and the results were more scientific andexact compared with traditional pharmacology [23] In thecurrent study the JFK-containing serums were prepared withrats and applied for the in vitro assays
EPCs have been extensively studied as a potential markerfor endothelial regeneration ability and shown to be atherapeutic potential by directly enhancing angiogenesis
or forming new vessels through vasculogenesis EPCs alsoexhibit a character to be highly migratory to ischemicareas [24] It is reported that circulating CD34+VEGFR-3+lymphaticvascular endothelial progenitor cells correlatewithlymph node metastasis in patients with epithelial ovariancancer [25] and further study revealed that CD34+VEGFR-3+ EPCs in human cord blood could differentiate into lym-phatic endothelial cells [10] Flow cytometry is widely usedto quantify and study circulating EPCs while EPC-specificmarkers are essential [11] Therefore in the current studyCD34 and VEGFR-3 were applied for the sorting of the targetEPCs from human cord blood and the results demonstratedthat the frequency of the CD34+VEGFR-3+ cells was 083in the mononuclear cells which was consistent with thereported data [18]
It is well accepted that LECs contribute to lymphangio-genesis that plays an active role in the induction of metastasisto the lymph nodes in cancers CXC chemokine receptor 4(CXCR4) plays a central role in tumor cell dissemination andmetastasis development in more than 75 of all cancers [26]SDF-1 also known as chemokine CXC ligand binds to theCXCR4 receptor The SDF-1CXCR4 axis is essential for themigration of progenitor cells tissue regeneration and vascu-larization [27] Furthermore SDF-1CXCR4 axis significantlyassociates with lymph node metastasis and promotes lym-phangiogenesis in tumors [28] VEGF-CVEGFR-3 signalingis a keymodulator of the lymphatic system andVEGF-C levelcorrelates with lymph node metastasis and poor prognosis[9] VEGF-C promotes LEC proliferation migration andtube formation [10] VEGFR-3 siRNA effectively inhibitsdifferentiation and lymphangiogenesis of CD34+VEGFR-3+EPCs [29] Therefore in the present study we used SDF-1and VEGF-C to induce the LECs formation and migrationOur results clearly demonstrated that SDF-1 and VEGF-Cpromoted the CD34+VEGFR-3+ EPCs to differentiate to theLECs that showed positive LYVE-1 staining a specific LECmarker and also enhanced the LECs migration
Although it is reported that VEGF-C stimulation upreg-ulates the expression level of CXCR4 on lymphangiogenicendothelial cells SDF-1CXCR4 axis exerts its function ina VEGFR-3 independent pattern [12] More interestinglytargeting both SDF-1 and VEGF-C by neutralizing antibodiesresulted in an additive inhibitory effect on both tumor lym-phangiogenesis and lymphatic metastasis [12] Furthermoreit is hypothesized that SDF-1 and VEGF may have syner-getic effect on endothelial differentiation through a positivefeedback loop with mutual promotion [30] Thus to clarifythe coeffect of SDF-1 and VEGF-C we added both in theculture system and the data revealed that costimulation withSDF-1 and VEGF-C displayed an additive effect on the LECsdifferentiation however no additive effect was observedon the LECs migration which needs a further mechanismclarification
Studies have shown that JFK can prevent tumor growthand progression and inhibit tumor angiogenesis [16] How-ever there is no data onwhether JFK inhibits tumor lymphan-giogenesis The present work provided the first experimentalevidence that JFK suppressed the formation and directionalmigration of LECs in vitro which closely related to the tumor
8 Evidence-Based Complementary and Alternative Medicine
JFK-3BSSD
F-1
SDF-
1SD
F-1
JFK-1 JFK-2 AMDM
erge
DA
PILY
VE-
1
(a1) (a2) (a3) (a4) (a5)
(b1) (b2) (b3) (b4) (b5)
(c1) (c2) (c3) (c4) (c5)JFK-3BS
VEG
F-C
VEG
F-C
VEG
F-C
JFK-1 JFK-2 LEN
(A1)
(B1)
(C1) (C2) (C3) (C4) (C5)
(B2) (B3) (B4) (B5)
(A2) (A3) (A4) (A5)
Mer
geD
API
LYV
E-1
(a1) (a2) (a3) (a4) (a5)
(b1) (b2) (b3) (b4) (b5)
(A1)
(B1) (B2) (B3) (B4) (B5)
(A2) (A3) (A4) (A5)
100120583m
100120583m
100120583m
Figure 6 Effect of JFK on LYVE-1 positive LECs differentiation The LECs formation was induced from CD34+VEGFR-3+ EPCs withSDF-1 (100 ngmL) and VEGF-C (60 ngmL) respectively BS blank serum JFK-1 JFK-2 and JFK-3 the serums prepared from rats by oraladministration of JFK at doses of 18 36 and 72 gkg respectively and 10 (vv) of the serums were added AMD AMD3100 (10 120583M) LenLenvatinib (10 120583M)
Evidence-Based Complementary and Alternative Medicine 9
AMDV
EGF-
CSD
F-1
JFK-2JFK-1 JFK-3BS
LenJFK-2JFK-1 JFK-3BS
200120583m
200120583m
(A1)
(B1) (B2) (B3) (B4) (B5)
(A2) (A3) (A4) (A5)
Figure 7 Effect of JFK on LECs transmigration The LECs migration was induced with SDF-1 (100 ngmL) and VEGF-C (60 ngmL)respectively The LECs were pretreated with blank serum (BS) the serums (each 10 vv) prepared from rats by oral administration ofJFK at doses of 18 36 and 72 gkg (JFK-1 JFK-2 and JFK-3 resp) AMD3100 (AMD 10 120583M) Lenvatinib (Len 10 120583M)
lymphangiogenesis Furthermore the inhibitory effect mightbe via regulation of SDF-1CXCR4 and VEGF-CVEGFR-3axes As JFK is composed of 12 herbal drugs further efforton chemical isolation and structure identification of JFK isessential to clarify the bioactive components responsible forthe effect and might provide a potential to discover the anti-lymphangiogenesis agents from the natural resources
5 Conclusion
This study demonstrated that SDF-1 and VEGF-C inducedCD34+VEGFR-3+ EPCs to differentiate towards LECs andenhanced the LECsmigration SDF-1 andVEGF-C exerted anadditive role on the LECs formation but not on themigrationJFK inhibited the differentiation and migration of LECs viaregulation of the SDF-1CXCR4 and VEGF-CVEGFR-3 axesat least The current finding suggested that JFK might inhibitNSCLC through antilymphangiogenesis and also provided apotential to discover antilymphangiogenesis agents Furtherresearch on the inhibitory effect of JFK on lymphangiogenesisin vivo and clarification of its bioactive components areundergoing in our lab
Competing Interests
The authors declare that they have no conflict of interests
Acknowledgments
This work was supported by Research Project for PracticeDevelopment of National TCM Clinical Research Bases(JDZX2012117) and the Project of Key Discipline for TCMConstruction of Jiangsu Province China (no JS1302)
References
[1] R A Smith D Manassaram-Baptiste D Brooks et al ldquoCancerscreening in the United States 2015 a review of currentAmerican Cancer Society guidelines and current issues incancer screeningrdquo CA A Cancer Journal for Clinicians vol 65no 1 pp 30ndash54 2015
[2] T K Kilvaer E-E Paulsen S M Hald et al ldquoLymphangiogenicmarkers and their impact on nodal metastasis and survival innon-small cell lung cancermdasha structured review with meta-analysisrdquo PLoS ONE vol 10 no 8 Article ID e0132481 2015
[3] F G Gomes F Nedel A M Alves J E Nor and S BC Tarquinio ldquoTumor angiogenesis and lymphangiogenesistumorendothelial crosstalk and cellularmicroenvironmentalsignaling mechanismsrdquo Life Sciences vol 92 no 2 pp 101ndash1072013
[4] Y Wang S Ma M Dong Y Yao K Liu and J Zhou ldquoEvalua-tion of the factors affecting the maximum standardized uptakevalue of metastatic lymph nodes in different histological typesof non-small cell lung cancer on PET-CTrdquo BMC PulmonaryMedicine vol 15 no 1 article 20 2015
[5] J P Sleeman and W Thiele ldquoTumor metastasis and the lym-phatic vasculaturerdquo International Journal of Cancer vol 125 no12 pp 2747ndash2756 2009
[6] S Li and Q Li ldquoCancer stem cells lymphangiogenesis andlymphatic metastasisrdquo Cancer Letters vol 357 no 2 pp 438ndash447 2015
[7] M Bzowska R Męzyk-Kopec T Prochnicki M KuleszaT Klaus and J Bereta ldquoAntibody-based antiangiogenic andantilymphangiogenic therapies to prevent tumor growth andprogressionrdquo Acta Biochimica Polonica vol 60 no 3 pp 263ndash275 2013
[8] S A Stacker S P Williams T Karnezis R Shayan S B Foxand M G Achen ldquoLymphangiogenesis and lymphatic vesselremodelling in cancerrdquo Nature Reviews Cancer vol 14 no 3pp 159ndash172 2014
[9] J Yu X Zhang P M Kuzontkoski et al ldquoSlit2N and Robo4regulate lymphangiogenesis through the VEGF-CVEGFR-3
10 Evidence-Based Complementary and Alternative Medicine
pathwayrdquo Cell Communication and Signaling vol 12 no 1article 25 pp 1ndash15 2014
[10] Y-Z Tan H-J Wang M-H Zhang Z Quan T Li and Q-Z He ldquoCD34+VEGFR-3+ progenitor cells have a potential todifferentiate towards lymphatic endothelial cellsrdquo Journal ofCellular and Molecular Medicine vol 18 no 3 pp 422ndash4332014
[11] M Moschetta Y Mishima I Sahin et al ldquoRole of endothelialprogenitor cells in cancer progressionrdquo Biochimica et BiophysicaActamdashReviews on Cancer vol 1846 no 1 pp 26ndash39 2014
[12] WZhuo L JiaN Song et al ldquoTheCXCL12ndashCXCR4 chemokinepathway a novel axis regulates lymphangiogenesisrdquo ClinicalCancer Research vol 18 no 19 pp 5387ndash5398 2012
[13] B Li W Bai P Sun B Zhou B Hu and J Ying ldquoThe effectof CXCL12 on endothelial progenitor cells potential target forangiogenesis in intracerebral hemorrhagerdquo Journal of Interferonand Cytokine Research vol 35 no 1 pp 23ndash31 2015
[14] J Liu M Pan Y Li D Ye and Y Guo ldquoClinical study of oralliquid Jin Fu Kang for the treatment of primary non-small celllung cancerrdquo Tumor vol 21 no 6 pp 463ndash465 2001
[15] J Liu Z Shi H Li et al ldquoClinical observation on 271 cases ofnon-small lung cancer treated with Yifei Kangliu Yin (Jin-Fu-Kang)rdquo Chinese Journal of Integrated Traditional and WesternMedicine vol 7 no 4 pp 247ndash250 2001
[16] Z CWang J L Zeng H L Zhang and JW Li ldquoJin FuKang ontumor growth and metastasis and vascular endothelial growthfactor of patients with advanced non-small cell lung cancerrdquoPractical Journal of Clinical Medicine vol 8 no 4 pp 148ndash1492011
[17] P Salven A Orpana and H Joensuu ldquoLeukocytes and plateletsof patients with cancer contain high levels of vascular endothe-lial growth factorrdquo Clinical Cancer Research vol 5 no 3 pp487ndash491 1999
[18] H-J Wang D Zhang Y-Z Tan and T Li ldquoAutophagy inendothelial progenitor cells is cytoprotective in hypoxic con-ditionsrdquo American Journal of PhysiologymdashCell Physiology vol304 no 7 pp C617ndashC626 2013
[19] M Tawada S-I Hayashi S Osada S Nakashima and KYoshida ldquoHuman gastric cancer organizes neighboring lym-phatic vessels via recruitment of bone marrow-derived lym-phatic endothelial progenitor cellsrdquo Journal of Gastroenterologyvol 47 no 9 pp 1057ndash1060 2012
[20] AMasuda H Yasuoka T Satoh Y Okazaki Y Yamaguchi andM Kuwana ldquoVersican is upregulated in circulating monocytesin patients with systemic sclerosis and amplifies a CCL2-mediated pathogenic looprdquo Arthritis Research andTherapy vol15 article R74 2013
[21] J Goldman T X Le M Skobe andM A Swartz ldquoOverexpres-sion of VEGF-C causes transient lymphatic hyperplasia but notincreased lymphangiogenesis in regenerating skinrdquo CirculationResearch vol 96 no 11 pp 1193ndash1199 2005
[22] S Nishida and H Satoh ldquoMechanisms for the vasodilationsinduced by Ginkgo biloba extract and its main constituentbilobalide in rat aortardquo Life Sciences vol 72 no 23 pp 2659ndash2667 2003
[23] B Wang L Zhu and Q Chen ldquoPrimary study on the applica-tion of Serum Pharmacology in Chinese traditional medicinerdquoColloids and Surfaces B Biointerfaces vol 43 no 3-4 pp 194ndash197 2005
[24] F Ma A Morancho J Montaner and A Rosell ldquoEndothelialprogenitor cells and revascularization following strokerdquo BrainResearch vol 1623 pp 150ndash159 2015
[25] H Qiu L Cao D Wang H Xu and Z Liang ldquoHigh levelsof circulatingCD34+VEGFR3+ lymphaticvascular endothelialprogenitor cells is correlated with lymph node metastasis inpatients with epithelial ovarian cancerrdquo Journal of Obstetrics andGynaecology Research vol 39 no 7 pp 1268ndash1275 2013
[26] B Furusato A Mohamed M Uhlen and J S Rhim ldquoCXCR4and cancer review Articlerdquo Pathology International vol 60 no7 pp 497ndash505 2010
[27] E Karshovska D Zagorac A Zernecke C Weber and ASchober ldquoA small molecule CXCR4 antagonist inhibits neoin-tima formation and smoothmuscle progenitor cellmobilizationafter arterial injuryrdquo Journal ofThrombosis andHaemostasis vol6 no 10 pp 1812ndash1815 2008
[28] A C Chiang and J Massague ldquoMolecular basis of metastasisrdquoTheNew England Journal of Medicine vol 359 no 26 pp 2814ndash2823 2008
[29] T Li G-DWang Y-Z Tan andH-JWang ldquoInhibition of lym-phangiogenesis of endothelial progenitor cells with VEGFR-3siRNA delivered with PEI-alginate nanoparticlesrdquo InternationalJournal of Biological Sciences vol 10 no 2 pp 160ndash170 2014
[30] M Wang and Z Zou ldquoMultiple mechanisms of SDF-1 promot-ing VEGF-induced endothelial differentiation of mesenchymalstem cellsrdquo International Journal of Cardiology vol 177 no 3pp 1098ndash1099 2014

Evidence-Based Complementary and Alternative Medicine 7
Control BS JFK-1 JFK-2 JFK-340
60
80
100
120
Cel
l sur
viva
l rat
e (
)
Figure 5 Cytotoxicity of JFK-containing serums The EPCs werecultured with no addition (control) blank serum (BS) and JFK-1JFK-2 and JFK-3 JFK-1 JFK-2 and JFK-3 represented the serumsprepared from rats by oral administration of JFK at doses of 18 36and 72 gkg respectively and 10 (vv) of the serums were used
inhibitory effect on the SDF-1 and VEGF-C induced migra-tions Furthermore the LECs migration induced with SDF-1+ VEGF-C was also markedly suppressed by JFK-containingserums in a dose dependent manner Blank serum showedno effect on the migration (data not shown) The SDF-1 andVEGF-C inducedmigrations were markedly blocked by theirinhibitors AMD3100 or Lenvatinib respectively
4 Discussion
In traditional bioassays in vitro on traditional Chinesemedicines generally extracts or simply purified componentsof crude drugs are directly added to the culture system [22]However the pharmacological actions of traditional Chinesemedicines are rather complicated due to their complexcompositions More important thing is that many of thecomponents contained in the crud drugs do not directlyexhibit their pharmacological actions until they experience aseries of biotransformation during absorption distributionand metabolite procedures Therefore without biotransfor-mation the results from the bioassays in vitromight not trulyreflect the pharmacological actions in vivo Furthermore theserum drug concentration directly reflects the drug efficacyin most cases Therefore serum pharmacology in which thedrug-containing serum was prepared in vivo was used forbioactivity evaluation and research on traditional Chinesemedicines in vitro and the results were more scientific andexact compared with traditional pharmacology [23] In thecurrent study the JFK-containing serums were prepared withrats and applied for the in vitro assays
EPCs have been extensively studied as a potential markerfor endothelial regeneration ability and shown to be atherapeutic potential by directly enhancing angiogenesis
or forming new vessels through vasculogenesis EPCs alsoexhibit a character to be highly migratory to ischemicareas [24] It is reported that circulating CD34+VEGFR-3+lymphaticvascular endothelial progenitor cells correlatewithlymph node metastasis in patients with epithelial ovariancancer [25] and further study revealed that CD34+VEGFR-3+ EPCs in human cord blood could differentiate into lym-phatic endothelial cells [10] Flow cytometry is widely usedto quantify and study circulating EPCs while EPC-specificmarkers are essential [11] Therefore in the current studyCD34 and VEGFR-3 were applied for the sorting of the targetEPCs from human cord blood and the results demonstratedthat the frequency of the CD34+VEGFR-3+ cells was 083in the mononuclear cells which was consistent with thereported data [18]
It is well accepted that LECs contribute to lymphangio-genesis that plays an active role in the induction of metastasisto the lymph nodes in cancers CXC chemokine receptor 4(CXCR4) plays a central role in tumor cell dissemination andmetastasis development in more than 75 of all cancers [26]SDF-1 also known as chemokine CXC ligand binds to theCXCR4 receptor The SDF-1CXCR4 axis is essential for themigration of progenitor cells tissue regeneration and vascu-larization [27] Furthermore SDF-1CXCR4 axis significantlyassociates with lymph node metastasis and promotes lym-phangiogenesis in tumors [28] VEGF-CVEGFR-3 signalingis a keymodulator of the lymphatic system andVEGF-C levelcorrelates with lymph node metastasis and poor prognosis[9] VEGF-C promotes LEC proliferation migration andtube formation [10] VEGFR-3 siRNA effectively inhibitsdifferentiation and lymphangiogenesis of CD34+VEGFR-3+EPCs [29] Therefore in the present study we used SDF-1and VEGF-C to induce the LECs formation and migrationOur results clearly demonstrated that SDF-1 and VEGF-Cpromoted the CD34+VEGFR-3+ EPCs to differentiate to theLECs that showed positive LYVE-1 staining a specific LECmarker and also enhanced the LECs migration
Although it is reported that VEGF-C stimulation upreg-ulates the expression level of CXCR4 on lymphangiogenicendothelial cells SDF-1CXCR4 axis exerts its function ina VEGFR-3 independent pattern [12] More interestinglytargeting both SDF-1 and VEGF-C by neutralizing antibodiesresulted in an additive inhibitory effect on both tumor lym-phangiogenesis and lymphatic metastasis [12] Furthermoreit is hypothesized that SDF-1 and VEGF may have syner-getic effect on endothelial differentiation through a positivefeedback loop with mutual promotion [30] Thus to clarifythe coeffect of SDF-1 and VEGF-C we added both in theculture system and the data revealed that costimulation withSDF-1 and VEGF-C displayed an additive effect on the LECsdifferentiation however no additive effect was observedon the LECs migration which needs a further mechanismclarification
Studies have shown that JFK can prevent tumor growthand progression and inhibit tumor angiogenesis [16] How-ever there is no data onwhether JFK inhibits tumor lymphan-giogenesis The present work provided the first experimentalevidence that JFK suppressed the formation and directionalmigration of LECs in vitro which closely related to the tumor
8 Evidence-Based Complementary and Alternative Medicine
JFK-3BSSD
F-1
SDF-
1SD
F-1
JFK-1 JFK-2 AMDM
erge
DA
PILY
VE-
1
(a1) (a2) (a3) (a4) (a5)
(b1) (b2) (b3) (b4) (b5)
(c1) (c2) (c3) (c4) (c5)JFK-3BS
VEG
F-C
VEG
F-C
VEG
F-C
JFK-1 JFK-2 LEN
(A1)
(B1)
(C1) (C2) (C3) (C4) (C5)
(B2) (B3) (B4) (B5)
(A2) (A3) (A4) (A5)
Mer
geD
API
LYV
E-1
(a1) (a2) (a3) (a4) (a5)
(b1) (b2) (b3) (b4) (b5)
(A1)
(B1) (B2) (B3) (B4) (B5)
(A2) (A3) (A4) (A5)
100120583m
100120583m
100120583m
Figure 6 Effect of JFK on LYVE-1 positive LECs differentiation The LECs formation was induced from CD34+VEGFR-3+ EPCs withSDF-1 (100 ngmL) and VEGF-C (60 ngmL) respectively BS blank serum JFK-1 JFK-2 and JFK-3 the serums prepared from rats by oraladministration of JFK at doses of 18 36 and 72 gkg respectively and 10 (vv) of the serums were added AMD AMD3100 (10 120583M) LenLenvatinib (10 120583M)
Evidence-Based Complementary and Alternative Medicine 9
AMDV
EGF-
CSD
F-1
JFK-2JFK-1 JFK-3BS
LenJFK-2JFK-1 JFK-3BS
200120583m
200120583m
(A1)
(B1) (B2) (B3) (B4) (B5)
(A2) (A3) (A4) (A5)
Figure 7 Effect of JFK on LECs transmigration The LECs migration was induced with SDF-1 (100 ngmL) and VEGF-C (60 ngmL)respectively The LECs were pretreated with blank serum (BS) the serums (each 10 vv) prepared from rats by oral administration ofJFK at doses of 18 36 and 72 gkg (JFK-1 JFK-2 and JFK-3 resp) AMD3100 (AMD 10 120583M) Lenvatinib (Len 10 120583M)
lymphangiogenesis Furthermore the inhibitory effect mightbe via regulation of SDF-1CXCR4 and VEGF-CVEGFR-3axes As JFK is composed of 12 herbal drugs further efforton chemical isolation and structure identification of JFK isessential to clarify the bioactive components responsible forthe effect and might provide a potential to discover the anti-lymphangiogenesis agents from the natural resources
5 Conclusion
This study demonstrated that SDF-1 and VEGF-C inducedCD34+VEGFR-3+ EPCs to differentiate towards LECs andenhanced the LECsmigration SDF-1 andVEGF-C exerted anadditive role on the LECs formation but not on themigrationJFK inhibited the differentiation and migration of LECs viaregulation of the SDF-1CXCR4 and VEGF-CVEGFR-3 axesat least The current finding suggested that JFK might inhibitNSCLC through antilymphangiogenesis and also provided apotential to discover antilymphangiogenesis agents Furtherresearch on the inhibitory effect of JFK on lymphangiogenesisin vivo and clarification of its bioactive components areundergoing in our lab
Competing Interests
The authors declare that they have no conflict of interests
Acknowledgments
This work was supported by Research Project for PracticeDevelopment of National TCM Clinical Research Bases(JDZX2012117) and the Project of Key Discipline for TCMConstruction of Jiangsu Province China (no JS1302)
References
[1] R A Smith D Manassaram-Baptiste D Brooks et al ldquoCancerscreening in the United States 2015 a review of currentAmerican Cancer Society guidelines and current issues incancer screeningrdquo CA A Cancer Journal for Clinicians vol 65no 1 pp 30ndash54 2015
[2] T K Kilvaer E-E Paulsen S M Hald et al ldquoLymphangiogenicmarkers and their impact on nodal metastasis and survival innon-small cell lung cancermdasha structured review with meta-analysisrdquo PLoS ONE vol 10 no 8 Article ID e0132481 2015
[3] F G Gomes F Nedel A M Alves J E Nor and S BC Tarquinio ldquoTumor angiogenesis and lymphangiogenesistumorendothelial crosstalk and cellularmicroenvironmentalsignaling mechanismsrdquo Life Sciences vol 92 no 2 pp 101ndash1072013
[4] Y Wang S Ma M Dong Y Yao K Liu and J Zhou ldquoEvalua-tion of the factors affecting the maximum standardized uptakevalue of metastatic lymph nodes in different histological typesof non-small cell lung cancer on PET-CTrdquo BMC PulmonaryMedicine vol 15 no 1 article 20 2015
[5] J P Sleeman and W Thiele ldquoTumor metastasis and the lym-phatic vasculaturerdquo International Journal of Cancer vol 125 no12 pp 2747ndash2756 2009
[6] S Li and Q Li ldquoCancer stem cells lymphangiogenesis andlymphatic metastasisrdquo Cancer Letters vol 357 no 2 pp 438ndash447 2015
[7] M Bzowska R Męzyk-Kopec T Prochnicki M KuleszaT Klaus and J Bereta ldquoAntibody-based antiangiogenic andantilymphangiogenic therapies to prevent tumor growth andprogressionrdquo Acta Biochimica Polonica vol 60 no 3 pp 263ndash275 2013
[8] S A Stacker S P Williams T Karnezis R Shayan S B Foxand M G Achen ldquoLymphangiogenesis and lymphatic vesselremodelling in cancerrdquo Nature Reviews Cancer vol 14 no 3pp 159ndash172 2014
[9] J Yu X Zhang P M Kuzontkoski et al ldquoSlit2N and Robo4regulate lymphangiogenesis through the VEGF-CVEGFR-3
10 Evidence-Based Complementary and Alternative Medicine
pathwayrdquo Cell Communication and Signaling vol 12 no 1article 25 pp 1ndash15 2014
[10] Y-Z Tan H-J Wang M-H Zhang Z Quan T Li and Q-Z He ldquoCD34+VEGFR-3+ progenitor cells have a potential todifferentiate towards lymphatic endothelial cellsrdquo Journal ofCellular and Molecular Medicine vol 18 no 3 pp 422ndash4332014
[11] M Moschetta Y Mishima I Sahin et al ldquoRole of endothelialprogenitor cells in cancer progressionrdquo Biochimica et BiophysicaActamdashReviews on Cancer vol 1846 no 1 pp 26ndash39 2014
[12] WZhuo L JiaN Song et al ldquoTheCXCL12ndashCXCR4 chemokinepathway a novel axis regulates lymphangiogenesisrdquo ClinicalCancer Research vol 18 no 19 pp 5387ndash5398 2012
[13] B Li W Bai P Sun B Zhou B Hu and J Ying ldquoThe effectof CXCL12 on endothelial progenitor cells potential target forangiogenesis in intracerebral hemorrhagerdquo Journal of Interferonand Cytokine Research vol 35 no 1 pp 23ndash31 2015
[14] J Liu M Pan Y Li D Ye and Y Guo ldquoClinical study of oralliquid Jin Fu Kang for the treatment of primary non-small celllung cancerrdquo Tumor vol 21 no 6 pp 463ndash465 2001
[15] J Liu Z Shi H Li et al ldquoClinical observation on 271 cases ofnon-small lung cancer treated with Yifei Kangliu Yin (Jin-Fu-Kang)rdquo Chinese Journal of Integrated Traditional and WesternMedicine vol 7 no 4 pp 247ndash250 2001
[16] Z CWang J L Zeng H L Zhang and JW Li ldquoJin FuKang ontumor growth and metastasis and vascular endothelial growthfactor of patients with advanced non-small cell lung cancerrdquoPractical Journal of Clinical Medicine vol 8 no 4 pp 148ndash1492011
[17] P Salven A Orpana and H Joensuu ldquoLeukocytes and plateletsof patients with cancer contain high levels of vascular endothe-lial growth factorrdquo Clinical Cancer Research vol 5 no 3 pp487ndash491 1999
[18] H-J Wang D Zhang Y-Z Tan and T Li ldquoAutophagy inendothelial progenitor cells is cytoprotective in hypoxic con-ditionsrdquo American Journal of PhysiologymdashCell Physiology vol304 no 7 pp C617ndashC626 2013
[19] M Tawada S-I Hayashi S Osada S Nakashima and KYoshida ldquoHuman gastric cancer organizes neighboring lym-phatic vessels via recruitment of bone marrow-derived lym-phatic endothelial progenitor cellsrdquo Journal of Gastroenterologyvol 47 no 9 pp 1057ndash1060 2012
[20] AMasuda H Yasuoka T Satoh Y Okazaki Y Yamaguchi andM Kuwana ldquoVersican is upregulated in circulating monocytesin patients with systemic sclerosis and amplifies a CCL2-mediated pathogenic looprdquo Arthritis Research andTherapy vol15 article R74 2013
[21] J Goldman T X Le M Skobe andM A Swartz ldquoOverexpres-sion of VEGF-C causes transient lymphatic hyperplasia but notincreased lymphangiogenesis in regenerating skinrdquo CirculationResearch vol 96 no 11 pp 1193ndash1199 2005
[22] S Nishida and H Satoh ldquoMechanisms for the vasodilationsinduced by Ginkgo biloba extract and its main constituentbilobalide in rat aortardquo Life Sciences vol 72 no 23 pp 2659ndash2667 2003
[23] B Wang L Zhu and Q Chen ldquoPrimary study on the applica-tion of Serum Pharmacology in Chinese traditional medicinerdquoColloids and Surfaces B Biointerfaces vol 43 no 3-4 pp 194ndash197 2005
[24] F Ma A Morancho J Montaner and A Rosell ldquoEndothelialprogenitor cells and revascularization following strokerdquo BrainResearch vol 1623 pp 150ndash159 2015
[25] H Qiu L Cao D Wang H Xu and Z Liang ldquoHigh levelsof circulatingCD34+VEGFR3+ lymphaticvascular endothelialprogenitor cells is correlated with lymph node metastasis inpatients with epithelial ovarian cancerrdquo Journal of Obstetrics andGynaecology Research vol 39 no 7 pp 1268ndash1275 2013
[26] B Furusato A Mohamed M Uhlen and J S Rhim ldquoCXCR4and cancer review Articlerdquo Pathology International vol 60 no7 pp 497ndash505 2010
[27] E Karshovska D Zagorac A Zernecke C Weber and ASchober ldquoA small molecule CXCR4 antagonist inhibits neoin-tima formation and smoothmuscle progenitor cellmobilizationafter arterial injuryrdquo Journal ofThrombosis andHaemostasis vol6 no 10 pp 1812ndash1815 2008
[28] A C Chiang and J Massague ldquoMolecular basis of metastasisrdquoTheNew England Journal of Medicine vol 359 no 26 pp 2814ndash2823 2008
[29] T Li G-DWang Y-Z Tan andH-JWang ldquoInhibition of lym-phangiogenesis of endothelial progenitor cells with VEGFR-3siRNA delivered with PEI-alginate nanoparticlesrdquo InternationalJournal of Biological Sciences vol 10 no 2 pp 160ndash170 2014
[30] M Wang and Z Zou ldquoMultiple mechanisms of SDF-1 promot-ing VEGF-induced endothelial differentiation of mesenchymalstem cellsrdquo International Journal of Cardiology vol 177 no 3pp 1098ndash1099 2014

8 Evidence-Based Complementary and Alternative Medicine
JFK-3BSSD
F-1
SDF-
1SD
F-1
JFK-1 JFK-2 AMDM
erge
DA
PILY
VE-
1
(a1) (a2) (a3) (a4) (a5)
(b1) (b2) (b3) (b4) (b5)
(c1) (c2) (c3) (c4) (c5)JFK-3BS
VEG
F-C
VEG
F-C
VEG
F-C
JFK-1 JFK-2 LEN
(A1)
(B1)
(C1) (C2) (C3) (C4) (C5)
(B2) (B3) (B4) (B5)
(A2) (A3) (A4) (A5)
Mer
geD
API
LYV
E-1
(a1) (a2) (a3) (a4) (a5)
(b1) (b2) (b3) (b4) (b5)
(A1)
(B1) (B2) (B3) (B4) (B5)
(A2) (A3) (A4) (A5)
100120583m
100120583m
100120583m
Figure 6 Effect of JFK on LYVE-1 positive LECs differentiation The LECs formation was induced from CD34+VEGFR-3+ EPCs withSDF-1 (100 ngmL) and VEGF-C (60 ngmL) respectively BS blank serum JFK-1 JFK-2 and JFK-3 the serums prepared from rats by oraladministration of JFK at doses of 18 36 and 72 gkg respectively and 10 (vv) of the serums were added AMD AMD3100 (10 120583M) LenLenvatinib (10 120583M)
Evidence-Based Complementary and Alternative Medicine 9
AMDV
EGF-
CSD
F-1
JFK-2JFK-1 JFK-3BS
LenJFK-2JFK-1 JFK-3BS
200120583m
200120583m
(A1)
(B1) (B2) (B3) (B4) (B5)
(A2) (A3) (A4) (A5)
Figure 7 Effect of JFK on LECs transmigration The LECs migration was induced with SDF-1 (100 ngmL) and VEGF-C (60 ngmL)respectively The LECs were pretreated with blank serum (BS) the serums (each 10 vv) prepared from rats by oral administration ofJFK at doses of 18 36 and 72 gkg (JFK-1 JFK-2 and JFK-3 resp) AMD3100 (AMD 10 120583M) Lenvatinib (Len 10 120583M)
lymphangiogenesis Furthermore the inhibitory effect mightbe via regulation of SDF-1CXCR4 and VEGF-CVEGFR-3axes As JFK is composed of 12 herbal drugs further efforton chemical isolation and structure identification of JFK isessential to clarify the bioactive components responsible forthe effect and might provide a potential to discover the anti-lymphangiogenesis agents from the natural resources
5 Conclusion
This study demonstrated that SDF-1 and VEGF-C inducedCD34+VEGFR-3+ EPCs to differentiate towards LECs andenhanced the LECsmigration SDF-1 andVEGF-C exerted anadditive role on the LECs formation but not on themigrationJFK inhibited the differentiation and migration of LECs viaregulation of the SDF-1CXCR4 and VEGF-CVEGFR-3 axesat least The current finding suggested that JFK might inhibitNSCLC through antilymphangiogenesis and also provided apotential to discover antilymphangiogenesis agents Furtherresearch on the inhibitory effect of JFK on lymphangiogenesisin vivo and clarification of its bioactive components areundergoing in our lab
Competing Interests
The authors declare that they have no conflict of interests
Acknowledgments
This work was supported by Research Project for PracticeDevelopment of National TCM Clinical Research Bases(JDZX2012117) and the Project of Key Discipline for TCMConstruction of Jiangsu Province China (no JS1302)
References
[1] R A Smith D Manassaram-Baptiste D Brooks et al ldquoCancerscreening in the United States 2015 a review of currentAmerican Cancer Society guidelines and current issues incancer screeningrdquo CA A Cancer Journal for Clinicians vol 65no 1 pp 30ndash54 2015
[2] T K Kilvaer E-E Paulsen S M Hald et al ldquoLymphangiogenicmarkers and their impact on nodal metastasis and survival innon-small cell lung cancermdasha structured review with meta-analysisrdquo PLoS ONE vol 10 no 8 Article ID e0132481 2015
[3] F G Gomes F Nedel A M Alves J E Nor and S BC Tarquinio ldquoTumor angiogenesis and lymphangiogenesistumorendothelial crosstalk and cellularmicroenvironmentalsignaling mechanismsrdquo Life Sciences vol 92 no 2 pp 101ndash1072013
[4] Y Wang S Ma M Dong Y Yao K Liu and J Zhou ldquoEvalua-tion of the factors affecting the maximum standardized uptakevalue of metastatic lymph nodes in different histological typesof non-small cell lung cancer on PET-CTrdquo BMC PulmonaryMedicine vol 15 no 1 article 20 2015
[5] J P Sleeman and W Thiele ldquoTumor metastasis and the lym-phatic vasculaturerdquo International Journal of Cancer vol 125 no12 pp 2747ndash2756 2009
[6] S Li and Q Li ldquoCancer stem cells lymphangiogenesis andlymphatic metastasisrdquo Cancer Letters vol 357 no 2 pp 438ndash447 2015
[7] M Bzowska R Męzyk-Kopec T Prochnicki M KuleszaT Klaus and J Bereta ldquoAntibody-based antiangiogenic andantilymphangiogenic therapies to prevent tumor growth andprogressionrdquo Acta Biochimica Polonica vol 60 no 3 pp 263ndash275 2013
[8] S A Stacker S P Williams T Karnezis R Shayan S B Foxand M G Achen ldquoLymphangiogenesis and lymphatic vesselremodelling in cancerrdquo Nature Reviews Cancer vol 14 no 3pp 159ndash172 2014
[9] J Yu X Zhang P M Kuzontkoski et al ldquoSlit2N and Robo4regulate lymphangiogenesis through the VEGF-CVEGFR-3
10 Evidence-Based Complementary and Alternative Medicine
pathwayrdquo Cell Communication and Signaling vol 12 no 1article 25 pp 1ndash15 2014
[10] Y-Z Tan H-J Wang M-H Zhang Z Quan T Li and Q-Z He ldquoCD34+VEGFR-3+ progenitor cells have a potential todifferentiate towards lymphatic endothelial cellsrdquo Journal ofCellular and Molecular Medicine vol 18 no 3 pp 422ndash4332014
[11] M Moschetta Y Mishima I Sahin et al ldquoRole of endothelialprogenitor cells in cancer progressionrdquo Biochimica et BiophysicaActamdashReviews on Cancer vol 1846 no 1 pp 26ndash39 2014
[12] WZhuo L JiaN Song et al ldquoTheCXCL12ndashCXCR4 chemokinepathway a novel axis regulates lymphangiogenesisrdquo ClinicalCancer Research vol 18 no 19 pp 5387ndash5398 2012
[13] B Li W Bai P Sun B Zhou B Hu and J Ying ldquoThe effectof CXCL12 on endothelial progenitor cells potential target forangiogenesis in intracerebral hemorrhagerdquo Journal of Interferonand Cytokine Research vol 35 no 1 pp 23ndash31 2015
[14] J Liu M Pan Y Li D Ye and Y Guo ldquoClinical study of oralliquid Jin Fu Kang for the treatment of primary non-small celllung cancerrdquo Tumor vol 21 no 6 pp 463ndash465 2001
[15] J Liu Z Shi H Li et al ldquoClinical observation on 271 cases ofnon-small lung cancer treated with Yifei Kangliu Yin (Jin-Fu-Kang)rdquo Chinese Journal of Integrated Traditional and WesternMedicine vol 7 no 4 pp 247ndash250 2001
[16] Z CWang J L Zeng H L Zhang and JW Li ldquoJin FuKang ontumor growth and metastasis and vascular endothelial growthfactor of patients with advanced non-small cell lung cancerrdquoPractical Journal of Clinical Medicine vol 8 no 4 pp 148ndash1492011
[17] P Salven A Orpana and H Joensuu ldquoLeukocytes and plateletsof patients with cancer contain high levels of vascular endothe-lial growth factorrdquo Clinical Cancer Research vol 5 no 3 pp487ndash491 1999
[18] H-J Wang D Zhang Y-Z Tan and T Li ldquoAutophagy inendothelial progenitor cells is cytoprotective in hypoxic con-ditionsrdquo American Journal of PhysiologymdashCell Physiology vol304 no 7 pp C617ndashC626 2013
[19] M Tawada S-I Hayashi S Osada S Nakashima and KYoshida ldquoHuman gastric cancer organizes neighboring lym-phatic vessels via recruitment of bone marrow-derived lym-phatic endothelial progenitor cellsrdquo Journal of Gastroenterologyvol 47 no 9 pp 1057ndash1060 2012
[20] AMasuda H Yasuoka T Satoh Y Okazaki Y Yamaguchi andM Kuwana ldquoVersican is upregulated in circulating monocytesin patients with systemic sclerosis and amplifies a CCL2-mediated pathogenic looprdquo Arthritis Research andTherapy vol15 article R74 2013
[21] J Goldman T X Le M Skobe andM A Swartz ldquoOverexpres-sion of VEGF-C causes transient lymphatic hyperplasia but notincreased lymphangiogenesis in regenerating skinrdquo CirculationResearch vol 96 no 11 pp 1193ndash1199 2005
[22] S Nishida and H Satoh ldquoMechanisms for the vasodilationsinduced by Ginkgo biloba extract and its main constituentbilobalide in rat aortardquo Life Sciences vol 72 no 23 pp 2659ndash2667 2003
[23] B Wang L Zhu and Q Chen ldquoPrimary study on the applica-tion of Serum Pharmacology in Chinese traditional medicinerdquoColloids and Surfaces B Biointerfaces vol 43 no 3-4 pp 194ndash197 2005
[24] F Ma A Morancho J Montaner and A Rosell ldquoEndothelialprogenitor cells and revascularization following strokerdquo BrainResearch vol 1623 pp 150ndash159 2015
[25] H Qiu L Cao D Wang H Xu and Z Liang ldquoHigh levelsof circulatingCD34+VEGFR3+ lymphaticvascular endothelialprogenitor cells is correlated with lymph node metastasis inpatients with epithelial ovarian cancerrdquo Journal of Obstetrics andGynaecology Research vol 39 no 7 pp 1268ndash1275 2013
[26] B Furusato A Mohamed M Uhlen and J S Rhim ldquoCXCR4and cancer review Articlerdquo Pathology International vol 60 no7 pp 497ndash505 2010
[27] E Karshovska D Zagorac A Zernecke C Weber and ASchober ldquoA small molecule CXCR4 antagonist inhibits neoin-tima formation and smoothmuscle progenitor cellmobilizationafter arterial injuryrdquo Journal ofThrombosis andHaemostasis vol6 no 10 pp 1812ndash1815 2008
[28] A C Chiang and J Massague ldquoMolecular basis of metastasisrdquoTheNew England Journal of Medicine vol 359 no 26 pp 2814ndash2823 2008
[29] T Li G-DWang Y-Z Tan andH-JWang ldquoInhibition of lym-phangiogenesis of endothelial progenitor cells with VEGFR-3siRNA delivered with PEI-alginate nanoparticlesrdquo InternationalJournal of Biological Sciences vol 10 no 2 pp 160ndash170 2014
[30] M Wang and Z Zou ldquoMultiple mechanisms of SDF-1 promot-ing VEGF-induced endothelial differentiation of mesenchymalstem cellsrdquo International Journal of Cardiology vol 177 no 3pp 1098ndash1099 2014

Evidence-Based Complementary and Alternative Medicine 9
AMDV
EGF-
CSD
F-1
JFK-2JFK-1 JFK-3BS
LenJFK-2JFK-1 JFK-3BS
200120583m
200120583m
(A1)
(B1) (B2) (B3) (B4) (B5)
(A2) (A3) (A4) (A5)
Figure 7 Effect of JFK on LECs transmigration The LECs migration was induced with SDF-1 (100 ngmL) and VEGF-C (60 ngmL)respectively The LECs were pretreated with blank serum (BS) the serums (each 10 vv) prepared from rats by oral administration ofJFK at doses of 18 36 and 72 gkg (JFK-1 JFK-2 and JFK-3 resp) AMD3100 (AMD 10 120583M) Lenvatinib (Len 10 120583M)
lymphangiogenesis Furthermore the inhibitory effect mightbe via regulation of SDF-1CXCR4 and VEGF-CVEGFR-3axes As JFK is composed of 12 herbal drugs further efforton chemical isolation and structure identification of JFK isessential to clarify the bioactive components responsible forthe effect and might provide a potential to discover the anti-lymphangiogenesis agents from the natural resources
5 Conclusion
This study demonstrated that SDF-1 and VEGF-C inducedCD34+VEGFR-3+ EPCs to differentiate towards LECs andenhanced the LECsmigration SDF-1 andVEGF-C exerted anadditive role on the LECs formation but not on themigrationJFK inhibited the differentiation and migration of LECs viaregulation of the SDF-1CXCR4 and VEGF-CVEGFR-3 axesat least The current finding suggested that JFK might inhibitNSCLC through antilymphangiogenesis and also provided apotential to discover antilymphangiogenesis agents Furtherresearch on the inhibitory effect of JFK on lymphangiogenesisin vivo and clarification of its bioactive components areundergoing in our lab
Competing Interests
The authors declare that they have no conflict of interests
Acknowledgments
This work was supported by Research Project for PracticeDevelopment of National TCM Clinical Research Bases(JDZX2012117) and the Project of Key Discipline for TCMConstruction of Jiangsu Province China (no JS1302)
References
[1] R A Smith D Manassaram-Baptiste D Brooks et al ldquoCancerscreening in the United States 2015 a review of currentAmerican Cancer Society guidelines and current issues incancer screeningrdquo CA A Cancer Journal for Clinicians vol 65no 1 pp 30ndash54 2015
[2] T K Kilvaer E-E Paulsen S M Hald et al ldquoLymphangiogenicmarkers and their impact on nodal metastasis and survival innon-small cell lung cancermdasha structured review with meta-analysisrdquo PLoS ONE vol 10 no 8 Article ID e0132481 2015
[3] F G Gomes F Nedel A M Alves J E Nor and S BC Tarquinio ldquoTumor angiogenesis and lymphangiogenesistumorendothelial crosstalk and cellularmicroenvironmentalsignaling mechanismsrdquo Life Sciences vol 92 no 2 pp 101ndash1072013
[4] Y Wang S Ma M Dong Y Yao K Liu and J Zhou ldquoEvalua-tion of the factors affecting the maximum standardized uptakevalue of metastatic lymph nodes in different histological typesof non-small cell lung cancer on PET-CTrdquo BMC PulmonaryMedicine vol 15 no 1 article 20 2015
[5] J P Sleeman and W Thiele ldquoTumor metastasis and the lym-phatic vasculaturerdquo International Journal of Cancer vol 125 no12 pp 2747ndash2756 2009
[6] S Li and Q Li ldquoCancer stem cells lymphangiogenesis andlymphatic metastasisrdquo Cancer Letters vol 357 no 2 pp 438ndash447 2015
[7] M Bzowska R Męzyk-Kopec T Prochnicki M KuleszaT Klaus and J Bereta ldquoAntibody-based antiangiogenic andantilymphangiogenic therapies to prevent tumor growth andprogressionrdquo Acta Biochimica Polonica vol 60 no 3 pp 263ndash275 2013
[8] S A Stacker S P Williams T Karnezis R Shayan S B Foxand M G Achen ldquoLymphangiogenesis and lymphatic vesselremodelling in cancerrdquo Nature Reviews Cancer vol 14 no 3pp 159ndash172 2014
[9] J Yu X Zhang P M Kuzontkoski et al ldquoSlit2N and Robo4regulate lymphangiogenesis through the VEGF-CVEGFR-3
10 Evidence-Based Complementary and Alternative Medicine
pathwayrdquo Cell Communication and Signaling vol 12 no 1article 25 pp 1ndash15 2014
[10] Y-Z Tan H-J Wang M-H Zhang Z Quan T Li and Q-Z He ldquoCD34+VEGFR-3+ progenitor cells have a potential todifferentiate towards lymphatic endothelial cellsrdquo Journal ofCellular and Molecular Medicine vol 18 no 3 pp 422ndash4332014
[11] M Moschetta Y Mishima I Sahin et al ldquoRole of endothelialprogenitor cells in cancer progressionrdquo Biochimica et BiophysicaActamdashReviews on Cancer vol 1846 no 1 pp 26ndash39 2014
[12] WZhuo L JiaN Song et al ldquoTheCXCL12ndashCXCR4 chemokinepathway a novel axis regulates lymphangiogenesisrdquo ClinicalCancer Research vol 18 no 19 pp 5387ndash5398 2012
[13] B Li W Bai P Sun B Zhou B Hu and J Ying ldquoThe effectof CXCL12 on endothelial progenitor cells potential target forangiogenesis in intracerebral hemorrhagerdquo Journal of Interferonand Cytokine Research vol 35 no 1 pp 23ndash31 2015
[14] J Liu M Pan Y Li D Ye and Y Guo ldquoClinical study of oralliquid Jin Fu Kang for the treatment of primary non-small celllung cancerrdquo Tumor vol 21 no 6 pp 463ndash465 2001
[15] J Liu Z Shi H Li et al ldquoClinical observation on 271 cases ofnon-small lung cancer treated with Yifei Kangliu Yin (Jin-Fu-Kang)rdquo Chinese Journal of Integrated Traditional and WesternMedicine vol 7 no 4 pp 247ndash250 2001
[16] Z CWang J L Zeng H L Zhang and JW Li ldquoJin FuKang ontumor growth and metastasis and vascular endothelial growthfactor of patients with advanced non-small cell lung cancerrdquoPractical Journal of Clinical Medicine vol 8 no 4 pp 148ndash1492011
[17] P Salven A Orpana and H Joensuu ldquoLeukocytes and plateletsof patients with cancer contain high levels of vascular endothe-lial growth factorrdquo Clinical Cancer Research vol 5 no 3 pp487ndash491 1999
[18] H-J Wang D Zhang Y-Z Tan and T Li ldquoAutophagy inendothelial progenitor cells is cytoprotective in hypoxic con-ditionsrdquo American Journal of PhysiologymdashCell Physiology vol304 no 7 pp C617ndashC626 2013
[19] M Tawada S-I Hayashi S Osada S Nakashima and KYoshida ldquoHuman gastric cancer organizes neighboring lym-phatic vessels via recruitment of bone marrow-derived lym-phatic endothelial progenitor cellsrdquo Journal of Gastroenterologyvol 47 no 9 pp 1057ndash1060 2012
[20] AMasuda H Yasuoka T Satoh Y Okazaki Y Yamaguchi andM Kuwana ldquoVersican is upregulated in circulating monocytesin patients with systemic sclerosis and amplifies a CCL2-mediated pathogenic looprdquo Arthritis Research andTherapy vol15 article R74 2013
[21] J Goldman T X Le M Skobe andM A Swartz ldquoOverexpres-sion of VEGF-C causes transient lymphatic hyperplasia but notincreased lymphangiogenesis in regenerating skinrdquo CirculationResearch vol 96 no 11 pp 1193ndash1199 2005
[22] S Nishida and H Satoh ldquoMechanisms for the vasodilationsinduced by Ginkgo biloba extract and its main constituentbilobalide in rat aortardquo Life Sciences vol 72 no 23 pp 2659ndash2667 2003
[23] B Wang L Zhu and Q Chen ldquoPrimary study on the applica-tion of Serum Pharmacology in Chinese traditional medicinerdquoColloids and Surfaces B Biointerfaces vol 43 no 3-4 pp 194ndash197 2005
[24] F Ma A Morancho J Montaner and A Rosell ldquoEndothelialprogenitor cells and revascularization following strokerdquo BrainResearch vol 1623 pp 150ndash159 2015
[25] H Qiu L Cao D Wang H Xu and Z Liang ldquoHigh levelsof circulatingCD34+VEGFR3+ lymphaticvascular endothelialprogenitor cells is correlated with lymph node metastasis inpatients with epithelial ovarian cancerrdquo Journal of Obstetrics andGynaecology Research vol 39 no 7 pp 1268ndash1275 2013
[26] B Furusato A Mohamed M Uhlen and J S Rhim ldquoCXCR4and cancer review Articlerdquo Pathology International vol 60 no7 pp 497ndash505 2010
[27] E Karshovska D Zagorac A Zernecke C Weber and ASchober ldquoA small molecule CXCR4 antagonist inhibits neoin-tima formation and smoothmuscle progenitor cellmobilizationafter arterial injuryrdquo Journal ofThrombosis andHaemostasis vol6 no 10 pp 1812ndash1815 2008
[28] A C Chiang and J Massague ldquoMolecular basis of metastasisrdquoTheNew England Journal of Medicine vol 359 no 26 pp 2814ndash2823 2008
[29] T Li G-DWang Y-Z Tan andH-JWang ldquoInhibition of lym-phangiogenesis of endothelial progenitor cells with VEGFR-3siRNA delivered with PEI-alginate nanoparticlesrdquo InternationalJournal of Biological Sciences vol 10 no 2 pp 160ndash170 2014
[30] M Wang and Z Zou ldquoMultiple mechanisms of SDF-1 promot-ing VEGF-induced endothelial differentiation of mesenchymalstem cellsrdquo International Journal of Cardiology vol 177 no 3pp 1098ndash1099 2014

10 Evidence-Based Complementary and Alternative Medicine
pathwayrdquo Cell Communication and Signaling vol 12 no 1article 25 pp 1ndash15 2014
[10] Y-Z Tan H-J Wang M-H Zhang Z Quan T Li and Q-Z He ldquoCD34+VEGFR-3+ progenitor cells have a potential todifferentiate towards lymphatic endothelial cellsrdquo Journal ofCellular and Molecular Medicine vol 18 no 3 pp 422ndash4332014
[11] M Moschetta Y Mishima I Sahin et al ldquoRole of endothelialprogenitor cells in cancer progressionrdquo Biochimica et BiophysicaActamdashReviews on Cancer vol 1846 no 1 pp 26ndash39 2014
[12] WZhuo L JiaN Song et al ldquoTheCXCL12ndashCXCR4 chemokinepathway a novel axis regulates lymphangiogenesisrdquo ClinicalCancer Research vol 18 no 19 pp 5387ndash5398 2012
[13] B Li W Bai P Sun B Zhou B Hu and J Ying ldquoThe effectof CXCL12 on endothelial progenitor cells potential target forangiogenesis in intracerebral hemorrhagerdquo Journal of Interferonand Cytokine Research vol 35 no 1 pp 23ndash31 2015
[14] J Liu M Pan Y Li D Ye and Y Guo ldquoClinical study of oralliquid Jin Fu Kang for the treatment of primary non-small celllung cancerrdquo Tumor vol 21 no 6 pp 463ndash465 2001
[15] J Liu Z Shi H Li et al ldquoClinical observation on 271 cases ofnon-small lung cancer treated with Yifei Kangliu Yin (Jin-Fu-Kang)rdquo Chinese Journal of Integrated Traditional and WesternMedicine vol 7 no 4 pp 247ndash250 2001
[16] Z CWang J L Zeng H L Zhang and JW Li ldquoJin FuKang ontumor growth and metastasis and vascular endothelial growthfactor of patients with advanced non-small cell lung cancerrdquoPractical Journal of Clinical Medicine vol 8 no 4 pp 148ndash1492011
[17] P Salven A Orpana and H Joensuu ldquoLeukocytes and plateletsof patients with cancer contain high levels of vascular endothe-lial growth factorrdquo Clinical Cancer Research vol 5 no 3 pp487ndash491 1999
[18] H-J Wang D Zhang Y-Z Tan and T Li ldquoAutophagy inendothelial progenitor cells is cytoprotective in hypoxic con-ditionsrdquo American Journal of PhysiologymdashCell Physiology vol304 no 7 pp C617ndashC626 2013
[19] M Tawada S-I Hayashi S Osada S Nakashima and KYoshida ldquoHuman gastric cancer organizes neighboring lym-phatic vessels via recruitment of bone marrow-derived lym-phatic endothelial progenitor cellsrdquo Journal of Gastroenterologyvol 47 no 9 pp 1057ndash1060 2012
[20] AMasuda H Yasuoka T Satoh Y Okazaki Y Yamaguchi andM Kuwana ldquoVersican is upregulated in circulating monocytesin patients with systemic sclerosis and amplifies a CCL2-mediated pathogenic looprdquo Arthritis Research andTherapy vol15 article R74 2013
[21] J Goldman T X Le M Skobe andM A Swartz ldquoOverexpres-sion of VEGF-C causes transient lymphatic hyperplasia but notincreased lymphangiogenesis in regenerating skinrdquo CirculationResearch vol 96 no 11 pp 1193ndash1199 2005
[22] S Nishida and H Satoh ldquoMechanisms for the vasodilationsinduced by Ginkgo biloba extract and its main constituentbilobalide in rat aortardquo Life Sciences vol 72 no 23 pp 2659ndash2667 2003
[23] B Wang L Zhu and Q Chen ldquoPrimary study on the applica-tion of Serum Pharmacology in Chinese traditional medicinerdquoColloids and Surfaces B Biointerfaces vol 43 no 3-4 pp 194ndash197 2005
[24] F Ma A Morancho J Montaner and A Rosell ldquoEndothelialprogenitor cells and revascularization following strokerdquo BrainResearch vol 1623 pp 150ndash159 2015
[25] H Qiu L Cao D Wang H Xu and Z Liang ldquoHigh levelsof circulatingCD34+VEGFR3+ lymphaticvascular endothelialprogenitor cells is correlated with lymph node metastasis inpatients with epithelial ovarian cancerrdquo Journal of Obstetrics andGynaecology Research vol 39 no 7 pp 1268ndash1275 2013
[26] B Furusato A Mohamed M Uhlen and J S Rhim ldquoCXCR4and cancer review Articlerdquo Pathology International vol 60 no7 pp 497ndash505 2010
[27] E Karshovska D Zagorac A Zernecke C Weber and ASchober ldquoA small molecule CXCR4 antagonist inhibits neoin-tima formation and smoothmuscle progenitor cellmobilizationafter arterial injuryrdquo Journal ofThrombosis andHaemostasis vol6 no 10 pp 1812ndash1815 2008
[28] A C Chiang and J Massague ldquoMolecular basis of metastasisrdquoTheNew England Journal of Medicine vol 359 no 26 pp 2814ndash2823 2008
[29] T Li G-DWang Y-Z Tan andH-JWang ldquoInhibition of lym-phangiogenesis of endothelial progenitor cells with VEGFR-3siRNA delivered with PEI-alginate nanoparticlesrdquo InternationalJournal of Biological Sciences vol 10 no 2 pp 160ndash170 2014
[30] M Wang and Z Zou ldquoMultiple mechanisms of SDF-1 promot-ing VEGF-induced endothelial differentiation of mesenchymalstem cellsrdquo International Journal of Cardiology vol 177 no 3pp 1098ndash1099 2014