Embed Size (px)
Citation preview

Vol. 150, No. 1JOURNAL OF BACTERIOLOGY, Apr. 1982, p. 36-450021-9193/82/040036-10$02.00/0
Isoelectric Focusing and Crossed Immunoelectrophoresis ofHeme Proteins in the Escherichia coli Cytoplasmic Membrane
ROBERT G. KRANZ1 AND ROBERT B. GENNIS' 2*Departments ofBiochemistry' and Chemistry,2 University ofIllinois, Urbana, Illinois 61801
Received 9 July 1981/Accepted 18 November 1981
Isoelectric focusing (IEF), agarose electrophoresis, and crossed immunoelec-trophoresis (CIE) were used to resolve the heme-containing proteins of theEscherichia coli cytoplasmic membrane after solubilization by Triton X-100. Twobands in IEF stained for heme with pl values of 4.7 and 5.3. One of the bands,with an isoelectric point ofpH 5.3, was present only when the cells were grown tolate log or stationary phase and possessed N,N,N,'N'-tetramethyl-p-phenylene-diamine (TMPD) oxidase activity. The pI 4.7 band was present in cells harvestedin both mid-log and stationary phases. Agarose electrophoresis, using largersamples, revealed the same two components apparent by IEF, and, in addition, athird component. The heme-containing fractions were extracted after agaroseelectrophoresis and subjected to further study. The component which was presentin cells grown to stationary phase contained hemes b, a,, and d. The other twofractions contained only b heme. One of these corresponded to the componentwith pl 4.7 in IEF and had catalase activity. Antisera were raised against Triton X-100-solubilized cytoplasmic membranes and against the focused TMPD oxidasecomplex. With these anti-sera, CIE in the presence of Triton X-100 revealed fourprecipitin complexes containing heme. Three of these corresponded to thecomponents identified by IEF and agarose electrophoresis. We demonstrate thatthe combined use of IEF and CIE is valuable for analysis of membrane proteins.In particular, this work represents a substantial initial step toward a structuralelucidation of the E. coli aerobic respiratory chain.
The cytochromes present in the Escherichiacoli cytoplasmic membrane have been exten-sively studied by spectroscopic and potentio-metric techniques. Shipp (34), using fourth-or-der finite-difference analysis of spectra, reportedat least seven cytochromes, including two ccytochromes, three b cytochromes, cytochromea1, and cytochrome d (previously a2). Subse-quent workers have interpreted potentiometrictitration data in terms of either two (29) or three(16, 17, 30) b cytochromes in addition to cyto-chromes a, and d. Both cytochrome d and a bcytochrome, designated cytochrome o, bind toCO and are presumed to function as terminaloxidases (3, 30). There is conflicting evidenceover the possible role of cytochrome a1 as aterminal oxidase (7, 11). A minimal scheme,certainly oversimplified, for the E. coli respira-tory system appears to involve two parallelelectron transport chains:
Given the uncertainty as to even the composi-tion of the respiratory system, it is evident thatsuch a scheme is useful only as a crude workingmodel (for reviews, see references 12 and 18).
Little biochemistry has been attempted withthe membrane-bound cytochrome componentsof the E. coli respiratory chain. Earlier workersreported some success at solubilization and ini-tial attempts at biochemical resolution (1, 15).Kita and co-workers (20) reported the purifica-tion of the component referred to as cytochromeb556. Deeb and Hager (5), in earlier work, report-ed the purification of a b cytochrome (designat-ed cytochrome b1) from E. coli W strain. Therelationship between this cytochrome and thosewhich have been spectroscopically character-ized is unclear. In addition to strain differences,a further complication to biochemical studies isthat the cytochrome composition of E. coli ishighly dependent on the conditions under which
cyt b556 cyt o
Dehydrogenases -* quinones 02
cyt b558 --+cyt d
36
on May 22, 2020 by guest
http://jb.asm.org/
Dow
nloaded from

HEME PROTEINS IN E. COLI MEMBRANE 37
the cells are grown. However, by carefully con-trolling the growth conditions, the system can begreatly simplified for biochemical investigations(10, 28, 30, 32).One purpose of this paper is to demonstrate
the utility of both isoelectric focusing (IEF) andcrossed immunoelectrophoresis (CIE) for theresolution and analysis of the E. coli cyto-chromes. Both techniques have been previouslyapplied successfully to the study of membraneproteins (14, 26, 27, 36). In our study, fourheme-containing protein complexes were re-solved under nondenaturing conditions in thepresence of Triton X-100. We show that one ofthe components, probably the cytochrome dterminal oxidase, has N,N,N',N'-tetramethyl-p-phenylenediamine (TMPD) oxidase activity. Asecond component contains only b heme andpossesses catalase activity.
MATERIALS AND METHODS
Chemicals. Triton X-100 was obtained from Amer-sham Corp. TMPD was obtained from Marion Scien-tific Corp. as a 1% solution. 3,3,5',5'-Tetramethylben-zidine (TMBZ) was obtained from Sigma ChemicalCo.
Bacteria. Most of the work was performed with anE. coli K-12 strain carrying the F152 episome (denotedF2-gal in previous nomenclature). This strain, MR43L/F152, was obtained from W. Shipp and has beenpreviously described (35). This strain was also used byKita et al. for purifying cytochrome b556 (20). Thepresence of the episome results in enhanced levels(two- or threefold) of apparently all of the cyto-chromes. Strain SHSP19 (33), which requires exoge-nous -amino-levulinic acid (ALA) for heme biosyn-thesis, was made gal by mutagenesis with ethylmethane sulfonate. The F152 episome, which containsthe gal operon, was then transferred into SHSP19 bythe procedure described by Miller (25). The resultingstrain, denoted ALA/F152, has two- to threefold-enhanced levels of all of the cytochromes and stillrequires exogenous ALA for heme biosynthesis.Growth conditios. Cells were grown at 37°C on the
basal medium of Cohen and Rickenberg (4) supple-mented with 1 mM MgSO4, 0.1 mM CaCl2, 1.5 g ofCasamino Acids (Difco Laboratories) per liter, and 5 gof D,L-sodium lactate per liter. The ALA/F152 strainwas grown on glucose as the sole carbon source whenALA was not added.Two types of growth conditions were used. Cells
grown under conditions of high aeration in a 38-literNew Brunswick fermentor were harvested in early ormid-exponential growth phase. These cells have a pinkcolor. The air flow rate was 25 liters/min, and theagitation was 350 rpm. Cells grown under conditions oflow aeration produce large amounts of cytochrome d,causing the cells to have a dark green color. The airflow rate for these cells was 5 liters/min, and agitationwas at 200 rpm. These cells are harvested 5 to 10 h intostationary phase. The inoculum was a 100-ml culturegrown overnight in Penassay broth.
Absorption spectra. Spectrophotometric measure-
ments were made with an Aminco DW2 spectropho-tometer. A small amount of sodium dithionite wasused as the reductant, and potassium ferricyanide wasused as the oxidant for reduced-minus-oxidized spec-tra.
Preparations used for IEF. Cytoplasmic membranesof both mid-log and stationary-phase cells were isolat-ed by the method of Yamato et al. (39). All IEFpreparations used cytoplasmic membranes as thestarting point for solubilization. These membranes(approximately 4 mg of protein per ml) were solubi-lized by making the solution 1% Triton X-100, using a10%o Triton X-100 stock solution. The mixture wasincubated for 30 min at room temperature. After a 1-hcentrifugation at 90,000 x g at 5°C, the solubilizedsupernatant was used directly for IEF.As an alternative method, the inner membranes
were made 1% with sodium cholate, using a 20%osodium cholate stock solution, pH 7.2. The finalprotein concentration was approximately 4 mg/ml.This was saturated to 20%o (NH4)2SO4, using an 80%osaturated (NH4)2SO4 stock solution containing 0.05 Mpotassium phosphate, pH 7.0, and slowly stirred for 30min on ice. After centrifugation at 78,000 x g for 2 h at5°C, the supernatant was made 1% with Triton X-100,using a 1Oo Triton X-100 stock solution. This wasdialyzed overnight at 5°C against 1,000 volumes of0.2% Triton X-100, pH 7, and then centrifuged at78,000 x g for one h at 5°C. The supernatant was usedfor IEF and had a final protein concentration ofapproximately 1 mg/ml. A comparison of both thespectral characteristics and potentiometric titrationproffles of the intact membranes and the Triton X-100extracts demonstrated that the cytochromes were sim-ilar in the two preparations (not shown). This indicatedthat the detergent was apparently not selectively re-moving specific cytochromes.
IEF. IEF was carried out by using an LKB multi-phor flatbed system. All focusing was done at 5°C in 2-mm-thick polyacrylamide gels. The gels contained10%o glycerol, 1% Triton X-100, and 5% acrylamide. A3.5 to 9.5 pH gradient was made according to themanufacturer, using LKB ampholines. Gels were pre-focused at 300 v for 2 h. Application wells werecarefully cut and removed (approximately 1.5 mm by 3cm). The samples were applied in the wells andfocused for approximately 1.5 h before more samplewas applied to each well. Reapplication up to twotimes could be done. Focusing was carried out at 200V for 8 to 10 h after the final sample application. Thesample wells were cut near the cathode (approximate-ly pH 8.0). Variations of gel composition, applicationmethods, and focusing conditions were attempted, andthe conditions described are critical.
Preparations for CIE. The following procedure wasused for obtaining preparations to be used for CIE andagarose electrophoresis. Cells (5 g, wet weight) weresuspended in 40 ml of 0.05 M potassium phosphatebuffer-25% sucrose at pH 7.2. EDTA (0.4 ml of a 1 Mstock solution) and lysozyme (4 mg) were added, andcells were spheroplasted for 1 h at room temperature.After centrifugation at 11,000 x g for 30 min, thespheroplasts were suspended in 35 ml with 0.05 Mpotassium phosphate buffer, pH 7.2. Approximately 2mg of DNase was added, and the samples weresonicated at 5°C for 26 min (50%o pulse at 250 W).Whole cells were removed by centrifugation at 10,000
VOL. 150, 1982
on May 22, 2020 by guest
http://jb.asm.org/
Dow
nloaded from

38 KRANZ AND GENNIS
x g for 10 min. Membranes were pelleted at 160,000 xg in 2 to 4 h. The crude membrane pellet was homoge-nized in 8 ml of 0.05 M potassium phosphate buffer,pH 7.2. MgCl2 was then added to a final concentrationof 10 mM, using a 1 M stock solution. The membraneswere solubilized by adding 8 ml of an 8% Triton X-100solution and stirred slowly at room temperature for 30min. After centrifugation at 90,000 x g for 1 h at 50C,the supernatant was used for CIE. With radioactivelylabeled cells, the preparation was carried out as de-scribed above, but scaled down proportionately.CIE. CIE was carried out in 1% agarose in a pH 8.6
Tris-barbital buffer system (obtained from LKB).Electrophoresis in the first dimension was for 3.5 h at 5mA per plate, using 84 by 94-mm Gel Bond film (FMCCorp.). Rectangular sample application wells heldapproximately 60 ,l of sample. After the first-dimen-sion electrophoresis, plates were then sectioned ifnecessary to run only the proteins of interest againstthe selected antisera (as described below). Second-dimension electrophoresis was at 5 mA per plate (84by 94 mm) and run overnight. Plates were pressed,washed in 0.1 M NaCl, and pressed again, each for 20min. Heme staining was performed before, and proteinstaining was done after, plate drying.
Preparation of antiserum. Rabbits were injected withapproximately 5 mg of inner membrane protein fromstationary-phase cells solubilized in 4% Triton X-100.The preparation was emulsified with complete Freundadjuvant for the primary injection and with incompleteFreund adjuvant for monthly boosters. Sera werecollected weekly after the first booster, and 500 ml ofserum was treated according to Watt et al. (38) withsodium dextran sulfate, followed by ammonium sul-fate precipitation. The ammonium sulfate-precipitatedimmunoglobins were redissolved in 1/10 the originalserum volume. This was used for CIE and is referredto as the "anticytoplasmic membrane" serum.For the "anti-TMPD oxidase" serum, a rabbit was
immunized according to the following procedure. Thepl 5.3 band in IEF which stained for both heme andTMPD oxidase activity was sliced out. Several sam-ples were combined, and approximately 10 cm of thepolyacrylamide strips containing the TMPD oxidasewas homogenized through a 20-gauge needle in asolution consisting of 1 ml of 1% Triton X-100 plus 1ml of complete Freund adjuvant for the primary injec-tion or 1 ml of incomplete Freund adjuvant for themonthly booster injections. The rabbit was bled week-ly after the first booster, and 400 ml of the pooledserum was processed as above.Heme, TMPD, and protein staining. Heme staining
was carried out with TMBZ according to Thomes et al.(37). Protein staining was done with Coomassie bril-liant blue. Staining with TMPD was performed byusing a 1% TMPD solution obtained directly fromMarion Scientific Corp.
Protein determinations. Protein determinations werecarried out by using a modification of the method ofLowry et al. (21). All samples were made 1% withsodium dodecyl sulfate.
RESULTSFigure 1 shows the reduced-minus-oxidized
spectra of cells grown under conditions of high
A
t&Aa-0J15T
628590
520 560 600 640X (nm)
B
AA.AO5[ 568
5r.
FIG. 1. Reduced-minus-oxidized spectra of E. colicells recorded at room temperature. (A) Cells grownwith high aeration and harvested at early log phase(approximately 3 mg of protein per ml). (B) Cellsgrown with low aeration and harvested in stationaryphase (approximately 6 mg of protein per ml).
aeration (early log phase) and low aeration (sta-tionary phase). These data are consistent withprevious results from other laboratories (e.g.,30). Cells grown with high aeration and harvest-ed at early exponential growth phase containedprimarily b-type cytochromes. The predominantcytochrome present is reported to be cyto-chrome b556 (20). When the cells were grownwith low aeration and harvested in stationaryphase, several additional cytochromes ap-peared, including cytochromes a1, d, and b558.The spectral contribution due to cytochrome a,was always relatively small. Table 1 summarizesthe cytochrome contents of the cytoplasmicmembranes isolated from cells grown underthese two conditions.
Figure 2 shows the IEF profiles of solubilizedcytoplasmic membrane preparations. It is worthnoting that the procedure of initially solubilizingthe membranes in cholate plus (NH4)2SO4 andthen dialyzing in the presence of Triton X-100significantly improved the resolution obtainedby IEF. Possibly, some aggregated outer mem-brane contaminant was eliminated by the proce-dure which otherwise would cause "trailing"and streaking in the gel. Possibly for this samereason, it is important for best results to usecytoplasmic membranes as opposed to envelopepreparations containing outer membrane.
In defining the conditions required for suc-
J. BACTERIOL.
on May 22, 2020 by guest
http://jb.asm.org/
Dow
nloaded from
HEME PROTEINS IN E. COLI MEMBRANE 39
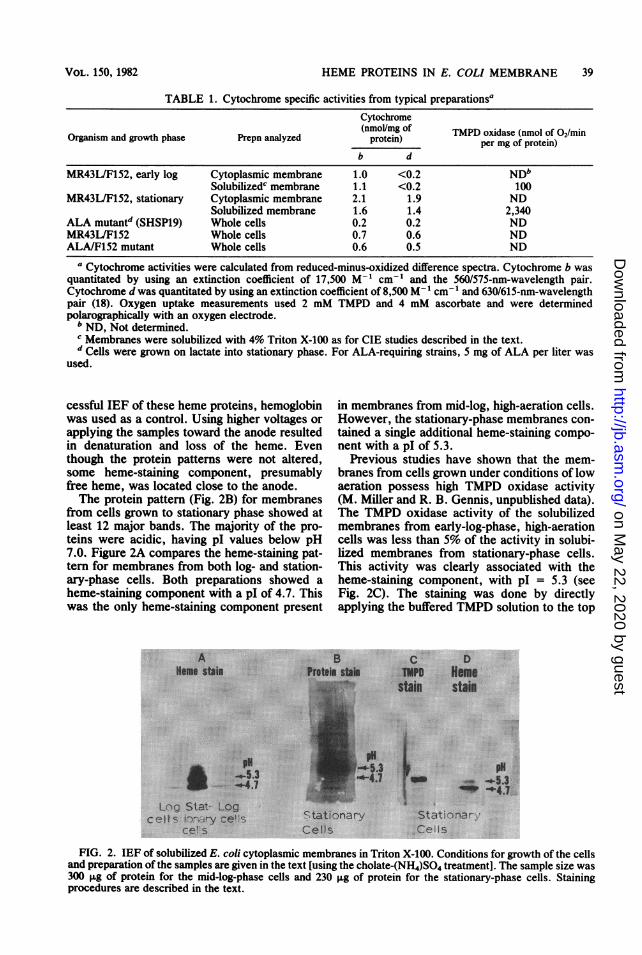
TABLE 1. Cytochrome specific activities from typical preparationsaCytochrome
Organism and growth phase Prepn analyzed (pmollmg of per mg of protein)b d
MR43L/F152, early log Cytoplasmic membrane 1.0 <0.2 NDbSolubilizedc membrane 1.1 <0.2 100
MR43L/F152, stationary Cytoplasmic membrane 2.1 1.9 NDSolubilized membrane 1.6 1.4 2,340
ALA mutantd (SHSP19) Whole cells 0.2 0.2 NDMR43L/F152 Whole cells 0.7 0.6 NDALA/F152 mutant Whole cells 0.6 0.5 ND
a Cytochrome activities were calculated from reduced-minus-oxidized difference spectra. Cytochrome b wasquantitated by using an extinction coefficient of 17,500 M-1 cm-' and the 560/575-nm-wavelength pair.Cytochrome d was quantitated by using an extinction coefficient of 8,500 M` cm-' and 630/615-nm-wavelengthpair (18). Oxygen uptake measurements used 2 mM TMPD and 4 mM ascorbate and were determinedpolarographically with an oxygen electrode.bND, Not determined.c Membranes were solubilized with 4% Triton X-100 as for CIE studies described in the text.d Cells were grown on lactate into stationary phase. For ALA-requiring strains, 5 mg of ALA per liter was
used.
cessful IEF of these heme proteins, hemoglobinwas used as a control. Using higher voltages orapplying the samples toward the anode resultedin denaturation and loss of the heme. Eventhough the protein patterns were not altered,some heme-staining component, presumablyfree heme, was located close to the anode.The protein pattern (Fig. 2B) for membranes
from cells grown to stationary phase showed atleast 12 major bands. The majority of the pro-teins were acidic, having pl values below pH7.0. Figure 2A compares the heme-staining pat-tern for membranes from both log- and station-ary-phase cells. Both preparations showed aheme-staining component with a pl of 4.7. Thiswas the only heme-staining component present
AHNM stab'.
Log Stat- LogceIIsi5ary cei's
ce's
in membranes from mid-log, high-aeration cells.However, the stationary-phase membranes con-tained a single additional heme-staining compo-nent with a pI of 5.3.
Previous studies have shown that the mem-branes from cells grown under conditions of lowaeration possess high TMPD oxidase activity(M. Miller and R. B. Gennis, unpublished data).The TMPD oxidase activity of the solubilizedmembranes from early-log-phase, high-aerationcells was less than 5% of the activity in solubi-lized membranes from stationary-phase cells.This activity was clearly associated with theheme-staining component, with pI = 5.3 (seeFig. 2C). The staining was done by directlyapplying the buffered TMPD solution to the top
Bb $
StationarCells
C D
staiN
y Stationar-v,,
Cells
FIG. 2. IEF of solubilized E. coli cytoplasmic membranes in Triton X-100. Conditions for growth of the cellsand preparation of the samples are given in the text [using the cholate-(NH4)SO4 treatment]. The sample size was300 ,g of protein for the mid-log-phase cells and 230 ,ug of protein for the stationary-phase cells. Stainingprocedures are described in the text.
VOL. 150, 1982
on May 22, 2020 by guest
http://jb.asm.org/
Dow
nloaded from

40 KRANZ AND GENNIS
pl 5.3TMaP Dox idase
pl4.7HemeCononiener
FIG. 3. Agarose electrophoresis of 4membranes in Triton X-100. Strain ALA/Flexogenous ALA (5 mg/liter) for heme biEThis strain was grown on glucose under loto stationary phase. Strain MR43L/F152 walactate. Sample size was 300 ,.g of protein inStaining procedures and sample preparatiscribed in the text.
of the IEF gel. Upon oxidation, the ddeep blue, and the activity was easilyAgarose electrophoresis in the pr
Triton X-100 could also be used to se]heme-staining components in E. coliby IEF (Fig. 3). This corresponds t(dimension of CIE. To verify that tioxidase activity and the heme-staininties of the two components were depthe presence of heme, a mutant whiciexogenous ALA for heme biosynttstudied. This mutant, ALA/F152, wasglucose to stationary phase both without exogenous ALA. When ALAifrom the growth medium, no cytochropresent by spectroscopic criteria (noand the cells had no respiratory activiTwo regions of the analytical ag
stained for heme when Triton X-100-sstationary-phase membranes of straitF152 were resolved electrophoreticallThe band with low mobility stained ioxidase activity. When these cells wEon glucose instead of lactate, the TOVplex appeared unchanged, but the amcsecond component appeared diminishown). When membranes from strF152 were grown in the presence ofglucose, the TMPD complex was cleent. However, growth in the absencresulted in the elimination of both I
staining and TMPD oxidase activities in theagarose gel. Clearly, heme was required forthese activities.The agarose electrophoresis was scaled up to
a preparative level to permit further analysis.Samples containing as much as 30 mg of protein
HEME were loaded onto the preparative gel. The hemeSTA IN
stain and TMPD oxidase stain revealed the samecomponents observed by the analytical proce-dure. In addition, a third component, with inter-mediate mobility, was evident in samples pre-pared from early-log-phase cells. This
TMPD component (b in Fig. 4) did not stain for heme,STAIN but was visible as a colored region in the prepar-
ative gel. Regions of the agarose gels wereexcised, and the proteins were extracted. Re-duced-minus-oxidized difference spectra wererecorded, and assays were made for TMPDoxidase and catalase. The heme-containing com-ponent with the lowest mobility present in sta-
solubilized tionary-phase cells contained hemes b, a1, and d52 requires (Fig. 4a). In addition, this fraction possessed allsyntesios of the TMPD oxidase activity in the membranes
)wgraton (Table 2). Although the electrophoresis resolvedeach laneo this component from other heme proteins, the
ion are de- specific activity did not indicate substantial puri-fication.The component in early-log-phase cells which
did not stain for heme contained only b heme[ye turned and did not have TMPD oxidase activity (Fig. 4;located. Table 2). This was the major heme protein inesence of cells grown under conditions of high aeration toparate the the early log phase. The component with fastestobserved mobility, present in both log- and stationary-
D the first phase membranes, was a minor component,he TMPD representing only about 5% of the total b hemeig proper- present in log-phase cells (Fig. 4). It containedendent on only b heme and, in addition, appeared to haveh requires high catalase activity. Solubilized E. coli cyto-hesis was plasmic membranes possessed catalase activitygrown on (Table 2). The catalase activity was present inand with- the membranes and not altered by solubilization
is omitted in Triton X-100. It is reasonable to assume that)mes were the heme-containing component is responsibleit shown), for this activity.ity. The E. coli K-12 strain we used has elevated,arose gel cytochrome levels and was chosen to increase;olubilized the chances of obtaining antibodies directedn MR43L/ against these components. Antiserum was pro-y (Fig. 3). duced from rabbits challenged with solubilizedFor TMPD cytoplasmic membranes from stationary-phaseere grown cells. CIE was performed with solubilized early-4PD com- log and stationary-phase membranes, using thisBunt of the antiserum. Figure 5 shows the results with sta-ished (not tionary-phase membranes. In both cases thereain ALA/ appeared to be at least 50 precipitin arcs visiblefALA on by Coomassie brillant blue staining for protein.arly pres- However, both early-log and stationary-phasee of ALA membranes resulted in only one precipitin arcthe heme- which stained for heme (Fig. 5c). This corre-
J. BACTERIOL.
on May 22, 2020 by guest
http://jb.asm.org/
Dow
nloaded from
~~HEME PROTEINS IN E. COLI MEMBRANE 41
....... .....
I \ t
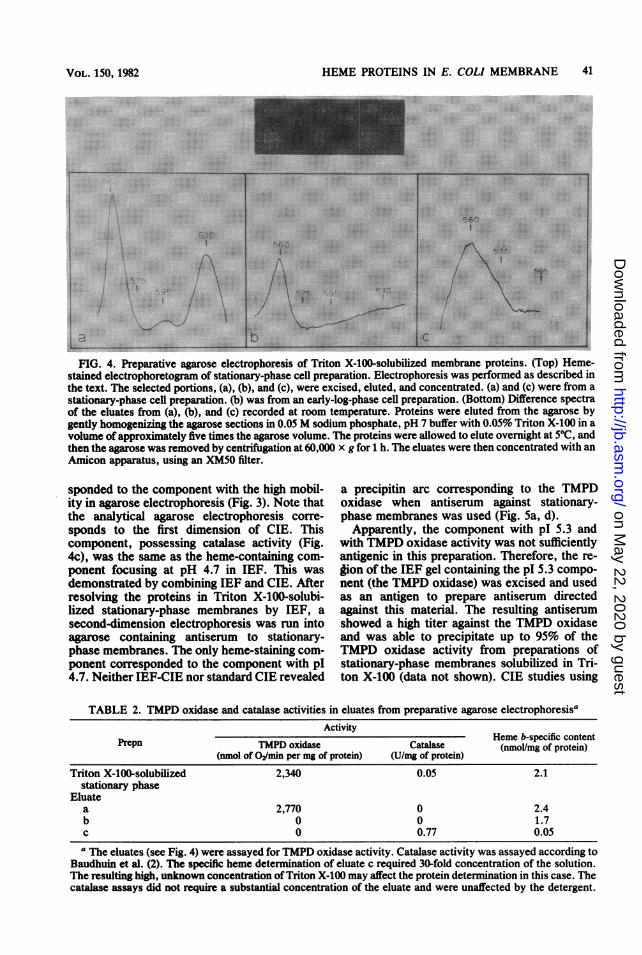
./ \;.FIG. 4. Preparative agarose electrophoresis of Triton X-100-solubilized membrane proteins. (Top) Heme-
stained electrophoretogram of stationary-phase cell preparation. Electrophoresis was performed as described in
the text. The selected portions, (a), (b), and (c), were excised, eluted, and concentrated. (a) and (c) were from a
stationary-phase cell preparation. (b) was from an early-log-phase cell preparation. (Bottom) Difference spectra
of the eluates from (a), (b), and (c) recorded at room temperature. Proteins were eluted from the agarose by
gently homogenizing the agarose sections in 0.05 M sodium phosphate, pH 7 buffer with 0.05% Triton X-100 in a
volume of approximately five times the agarose volume. The proteins were allowed to elute overnight at 50C, and
then the agarose was removed by centrifugation at 60,000 x g for h. The eluates were then concentrated with an
Amicon apparatus, using an XM50 ifter.
sponded to the component with the high mobil-
ity in agarose electrophoresis (Fig. 3). Note that
the analytical agarose electrophoresis corre-
sponds to the first dimension of CIE. This
component, possessing catalase activity (Fig.
4c), was the same as the heme-containing com-
ponent focusing at pH 4.7 in IEF. This was
demonstrated by combining IEF and CIE. After
resolving the proteins in Triton X-100-solubi-
lized stationary-phase membranes by IEF, a
second-dimension electrophoresis was run into
agarose containing antiserum to stationary-
phase membranes. The only heme-staining com-
ponent corresponded to the component with pI4.7. Neither IEF-CIE nor standard CIE revealed
a precipitin arc corresponding to the TMPD
oxidase when antiserum against stationary-
phase membranes was used (Fig. 5a, d).
Apparently, the component with pI 5.3 and
with TMPD oxidase activity was not sufficiently
antigenic in this preparation. Therefore, the re-
lfion of the TEE gel containing the pl 5.3 compo-
nent (the TMPD oxidase) was excised and used
as an antigen to prepare antiserum directed
against this material. The resulting antiserum
showed a high titer against the TMPD oxidase
and was able to precipitate up to 95% of the
TMPD oxidase activity from preparations of
stationary-phase membranes solubilized in Tri-
ton X-100 (data not shown). CIE studies using
TABLE 2. TMPD oxidase and catalase activities in eluates from preparative agarose electrophoresisaActivity
Prepn TMDoiaeaaseHeme b-specific contentTMPDoxidase Catalase ~~~(nmol/mg of protein)(nmol of 02/min per mg of protein) (U/mg of protein)
Triton X-100-solubilized 2,340 0.05 2.1stationary phase
Eluatea 2,770 0 .2.4b 0 0 1.7c 0 0.77 0.05a The eluates (see Fig. 4) were assayed for TMPD oxidase activity. Catalase activity was assayed according to
Baudhuin et al. (2). The specific heme determination of eluate c required 30-fold concentration of the solution.The resulting high, unknown concentration of Triton X-100 may affect the protein determination in this case. Thecatalase assays did not require a substantial concentration of the eluate and were unaffected by the detergent.
VOL. 150, 1982
on May 22, 2020 by guest
http://jb.asm.org/
Dow
nloaded from

42 KRANZ AND GENNIS
Protein stain HEME stain I TMPD oxidase stain
pH 4.7
FIG. 5. CIE of Triton X-100-solubilized membrane proteins from stationary-phase cell preparations, usinganticytoplasmic membrane serum. (a) IEF in the first dimension. (b), (c), and (d) are typical patterns of the righthalf of the first-dimension electrophoresis profiles (similar to Fig. 3) run against 1 ml of prepared antiserum perlarge (84 by 94 mm) CIE plate. Approximately 250 tg of membrane protein was loaded. Staining was withCoomassie brilliant blue (b), TMBZ for heme (a and c), and TMPD (d).
this anti-TMPD oxidase serum and solubilizedstationary-phase membranes showed that two ofthe major precipitin arcs stained for heme (Fig.6). Of these, only one arc stained with TMPD.These two heme-staining precipitin arcs wereabsent from CIE patterns run with solubilizedmembranes which were isolated from cells har-vested in early exponential growth (Fig. 6).We also demonstrated that all three of the
heme-staining precipitin arcs described con-tained heme. The ALA/F152 strain was grown inmedium containing [3H]ALA, and the presenceof heme was detected on the CIE plates byautoradiography. Figure 7 shows the two hemearcs which were observed after using the anti-TMPD oxidase serum and solubilized station-ary-phase membranes. These arcs, and no oth-ers, contained the radioactive label. Similarly,the precipitin arc corresponding to the pl 4.7component also was shown to contain heme bythis criterion, using anticytoplasmic membraneserum (not shown). In addition, another compo-nent, also very faint, was visible, correspondingto a component with a lower mobility in the firstdimension than that of the pI 4.7 component.This was probably due to the presence of a lowtiter of antibodies directed against the cyto-chrome observed in agarose electrophoresiswhich did not stain for heme (Fig. 4b).
DISCUSSIONThe E. coli aerobic respiratory chain is poorly
characterized from a biochemical standpoint.Our purpose here was to demonstrate the analyt-ical utility of IEF and CIE in resolving the heme-containing components in the E. coli cytoplas-mic membrane. At this point, four components
have been resolved in membranes solubilized inTriton X-100. Two of the heme-containing com-ponents are not present when the cells are grownunder conditions of high aeration and are har-vested at early-log growth phase. One of thesecomponents is a TMPD oxidase and is mostlikely the cytochrome d terminal oxidase (31).These two components, which are resolved byCIE, are not split by IEF and are both containedin the band with pI 5.3.One of the important conclusions of this work
is the demonstration that a single heme-contain-ing component is responsible for the observedTMPD oxidase activity in the membranes. Thisimplies that TMPD is a selective electron donorto the cytochrome d-containing branch of therespiratory system. Hence, TMPD can be usedto select for mutants deficient in cytochrome d,in a manner similar to that already reported forthe selection of cytochrome-deficient mutants inother bacteria (e.g., 23, 24). A preliminary re-port of such work has already been published(G. N. Green, R. Faiman, and R. B. Gennis,Fed. Proc. 40:1669, 1981).The heme-staining band with pI 4.7, which is
present in both the log- and stationary-phasemembranes, appears to be a single component.It corresponds to the only heme-staining precipi-tin arc which is present in CIE using serumdirected against cytoplasmic membranes. Theidentity of this component is not known. It is arelatively minor heme-containing protein and isapparently responsible for the catalase activityof E. coli membranes. Experiments to test itsfunction are in progress.
It is important to note that although the heme-containing protein with catalase activity is byspectroscopic analysis only a minor component,
J. BACTERIOL.
on May 22, 2020 by guest
http://jb.asm.org/
Dow
nloaded from

FIG. 6. CIE of Triton X-100-solubilized membrane proteins from stationary- and early-log-phase cells, usinganti-TMPD oxidase serum (as described in the text). These were stained with TMBZ, TMPD, and Coomassiebrilliant blue. The TMPD oxidase staining was performed by adding a few drops of a 1% TMPD solution over theCIE plate and then pressing the plate. Approximately 250 pLg of membrane protein was loaded and 400 ,ul ofprepared antiserum per large (84 by 94 mm) CIE plate. (A) Heme stain; (B) TMPD oxidase stain; (C) Proteinstain. SC, Stationary-phase cells; LC, log-phase cells.
on May 22, 2020 by guest
http://jb.asm.org/
Dow
nloaded from

44 KRANZ AND GENNIS
.::i i: ...j_
o.....
*: s: :..,i....... ..
:
::::
.:i.. :: . :: &i_
::;:.t....*, . '! ::. .
si .:e'.:
FIG. 7. CIEofsolubiliALA/F152 mutant growDplates were run as descrilwas visualized by autoramutant was grown in 100ml Klett flask shaken atstationary phase. Unlabr3y r-vA v A atd_% 9%1
formate dehydrogenase (8, 9) may be weaklyassociated with b cytochromes.IEF gels can also be stained for enzymatic
activities. Succinate dehydrogenase was clearlyfocused and was not associated with either ofthe heme-staining bands in the gel. The samefocusing protocol has been applied successfullyto Rhodospeudomonas sphaeroides cytoplasmicmembranes, as well as to purified E. coli D-lactate dehydrogenase and bacteriorhodopsinfrom Halobacterium halobium. The use of IEF,given specific staining and detection procedures,is of clear general value for studying membraneproteins and can be especially useful for produc-ing high-titer antisera toward specific proteins(6).One of the goals of this research is the produc-
tion of monospecific antibodies directed towardeach ofthe heme proteins in the E. coli cytoplas-
zed membrane proteins from mic membrane. The use of both IEF and CIEi with [3H]ALA. The CIE methods in achieving this goal is essential. Suchbed in Fig. 6. The [3H]ALA a library of antisera will be of great value,diography. The ALA/F152 especially in conjunction with preparative bio-ml of lactate media in a 250- chemical methods and genetics, in deciphering200 rpm and harvested in both the structure and function of the E. colieled ALA (0.25 mg) and aerobic respiratory chain.
l-'HJALA (0.05 mg, 0.3 mCi) were added to the mediafor optimal growth conditions.
it stains very heavily with the heme-specificTMBZ stain. In contrast, a major b-type cyto-chrome, present at a much higher concentrationin early-log-phase cells than the ("catalase,"does not stain for heme in this procedure. TheTMBZ stain is based on peroxidase activity and,clearly, heme proteins differ widely in theirefficiency in catalyzing this reaction. On theother hand, the only proteins which did stainwith TMBZ were shown to, in fact, containheme, using the ALA/F152 strain and [3H]ALAfor detecting these heme proteins.
In addition to staining the CIE plates forheme, zymograms were also made for the mostcommon dehydrogenases, as previously report-ed (27, 36). Precipitin arcs were identified whichcontained residual D-lactate, succinate, andNADH dehydrogenase activities. In no case didthese correspond to the major arcs containingheme. This is interesting in view of the recentreports that in Bacillus subtilis, succinate dehy-drogenase, solubilized in Triton X-100, is associ-ated with a b-type cytochrome (14). In beefheartmitochondria, the succinate-ubiquinone oxido-reductase (complex II) is also associated with aunique b cytochrome (13). This may not be thecase for any of the common dehydrogenases inaerobically grown E. coli. Further work is beingdirected at verifying this point. For anaerobical-ly grown E. coli, both nitrate reductase (22) and
ACKNOWLEDGMENTSWe thank John Cronan for providing us with the ALA
requiring strain and W. Shipp for supplying strain MR43L/F152. We are also grateful to Neil Green and Michael Millerfor helpful discussions and suggestions.
This prqject was supported by grants from the Public HealthService (HL 16101 from the National Institutes of Health) andthe Department of Energy (DE AC02-80ER10682). R.B.G.was supported in part by a Sloan fellowship, and R.G.K. wassupported in part as an National Institutes of Health trainee(Public Health Service grant IT32GM07283).
LITERATURE CITED1. _ae, R. D., C. Hon, and P. D. Bran. 1971. The
preparation and properties of a solubilized respiratorycomplex from Escherichia coli. Biochim. Biophys. Acta234:46-56.
2. Bandbuin, P., H. Beaufay, Y. Rhmn-I, 0. Z. Sdelnger,R. Watdaux, P. Jacques, and C. deDuve. 1964. Tissuefractionation studies. 17. Intracellular distribution ofmonoamine oxidase, aspartate aminotransferase, alanineaminotransferase, D-amino acid oxidase and catalase inrat-liver tissue. Biochem. J. 92:179-184.
3. Castor, L. N., and B. Chance. 1959. Oxidase of bacteria. J.Biol. Chem. 234:1587-1592.
4. Cobhe, G. N., and H. W. Rlckebelrg. 1956. Concentrationspecifique des aminoacides chez Escherichia coli. Ann.Inst. Pasteur Paris 91:693-720.
5. Deeb, S. S., and L. P. Hagr. 1964. Crystalline cyto-chrome b, from Escherichia coli. J. Biol. Chem. 239:1024.
6. DeMey, J., F. Vandeande, and K. Dlerkx. 1974. Identifi-cation of neurophysin producing cells. Cell Tissue Res.153:531-543.
7. Edwards, C., S. Beer, A. Sivram, and B. Chance. 1981.Photochemical action spectra of bacterial a- and o-typeoxidases using a dye laser. FEBS Lett. 128:205-207.
8. Enoch, H. G., and R. L. Lester. 1975. The purification offormate dehydrogenase and nitrate reductase from Esche-richia coli. J. Biol. Chem. 250:6693-6705.
9. Graham, A., and D. H. Boxer. 1981. The organization of
J. BACTERIOL.
on May 22, 2020 by guest
http://jb.asm.org/
Dow
nloaded from

HEME PROTEINS IN E. COLI MEMBRANE 45
formate dehydrogenase in the cytoplasmic membrane ofEscherichia coli. Biochem. J. 195:627-637.
10. Gray, C. T., J. W. T. Wimpenny, D. E. Hughes, and M. R.Mossman. 1966. Regulation of metabolism in facultativebacteria. Biochim. Biophys. Acta 117:22-32.
11. Haddock, B. A., J. A. Downie, and P. B. Garland. 1976.Kinetic characterization of the membrane-bound cyto-chromes of Escherichia coli grown under a variety ofconditions by using a stopped-flow dual-wavelength spec-trophotometer. Biochem. J. 154:285-294.
12. Haddock, B. A., and C. W. Jones. 1977. Bacterial respira-tion. Bacteriol. Rev. 41:47-99.
13. Hatefi, Y., and Y. M. Galante. 1980. Isolation of cyto-chrome b5w from complex II (succinate ubiquinone oxido-reductase) and its reconstitution with succinate dehydro-genase. J. Biol. Chem. 255:5530-5537.
14. Hederstedt, L., E. Holngren, and L. Rutberg. 1979.Characterization of a succinate dehydrogenase complexsolubilized from the cytoplasmic membrane of Bacillussubtilis with the nonionic detergent Triton X-100. J.Bacteriol. 138:370-376.
15. Hendler, R. W., and A. H. Burgess. 1972. Respiration andprotein synthesis in Escherichia coli membrane-envelopefragments. VI. Solubilization and characterization of theelectron transport chain. J. Cell Biol. 55:266-281.
16. Hendler, R. W., and R. I. Shrager. 1979. Potentiometricanalysis of Escherichia coli cytochromes in the opticalabsorbance range of 500 nm to 700 nm. J. Biol. Chem.254:11288-11299.
17. Hendler, R. W., D. W. Towne, and R. I. Shrager. 1975.Redox properties of b-type cytochromes in Escherichiacoli and rat liver mitochondria and techniques for theiranalysis. Biochim. Biophys. Acta 376:42-62.
18. Jones, C. W. 1977. Aerobic respiratory systems in bacte-ria, p. 23-60. In B. A. Haddock and W. A. Hamilton (ed.),Microbial energetics. Cambridge University Press, NewYork.
19. Jones, C. W., and E. R. Redfearn. 1966. Electron trans-port in Actobacter vinelandii. Biochim. Biophys. Acta113:467-481.
20. Kita, K., I. Yamato, and Y. Anraku. 1978. Purification andproperties of Cytochrome b556 in the respiratory chain ofaerobically grown Escherichia coli K12. J. Biol. Chem.253:8910-8915.
21. Lowry, 0. H., N. J. Rosebrough, A. L. Farr, and R. J.Randall. 1951. Protein measurement with the Folin phenolreagent. J. Biol. Chem. 193:265-275.
22. MacGregor, C. H. 1975. Anaerobic cytochrome b, inEscherichia coli: association with and regulation of nitratereductase. J. Bacteriol. 121:1111-1116.
23. Mam, B., and H. Gest. 1973. Genetic mutations affectingthe respiratory electron-transport system of the photosyn-thetic bacterium Rhodopseudomonas capsulata. J. Bac-
teriol. 114:1045-1051.24. Mkchels, P. A. M., and B. A. Haddock. 1980. Cytochrome
c deficient mutants of Rhodopseudomonas capsulata.FEMS Microbiol. Lett. 7:327-331.
25. Miller, J. H. 1972. Experiments in molecular genetics.Cold Spring Harbor Laboratory, Cold Spring Harbor,N.Y.
26. O'Farrell, P. H. 1975. High resolution two-dimensionalelectrophoresis of proteins. J. Biol. Chem. 250:4007-4021.
27. Owen, P., G. J. Kaczorowski, and H. R. Kaback. 1980.Resolution and identification of iron-containing antigensin membrane vesicles from Escherichia coli. Biochemistry19:596-600.
28. Pudek, M. R., and P. D. Bragg. 1974. Inhibition bycyanide of the respiratory chain oxidases of Escherichiacoli. Arch. Biochem. Biophys. 164:682-693.
29. Pudek, M. R., and P. D. Bragg. 1976. Redox potentials ofthe cytochromes in the respiratory chain of aerobicallygrown Escherichia coli. Arch. Biochem. Biophys.174:546-552.
30. Reid, G. A., and W. J. Ingeldew. 1979. Characterizationand phenotypic control of the cytochrome content ofEscherichia coli. Biochem. J. 182:465-472.
31. Reid, G. A., and W. J. Ingeldew. 1980. The purification ofa respiratory oxidase complex from Escherichia coli.FEBS Lett. 109:1-4.
32. Rke, C. W., and W. P. Hempling. 1978. Oxygen-limitedcontinuous culture and respiratory energy conservation inEscherichia coli. J. Bacteriol. 134:115-124.
33. Sasarman, A., M. Surdeanu, and T. Horodniceanu. 1968.Locus determining the synthesis of -aminolevulinic acidin Escherichia coli K-12. J. Bacteriol. 96:1882-1884.
34. Shipp, W. S. 1972. Cytochromes of Escherichia coli.Arch. Biochem. Biophys. 150:459-472.
35. Shipp, W. S., M. Plotrowski, and A. E. Friedman. 1972.Apparent cytochrome gene dose effects in F-lac and F-galheterogenotes of Escherichia coli. ArLh. Biochem.Biophys. 150:473-481.
36. Smyth, C. J., J. Siegel, M. R. J. Salton, and P. Owen.1978. Immunochemical analysis of inner and outer mem-branes of Escherichia coli by crossed immunoelectropho-resis. J. Bacteriol. 133:306-319.
37. Thomes, P. E., D. Ryan, and W. Levin. 1976. An improvedstaining procedure for the detection of the peroxidaseactivity of cytochrome p-450 on sodium dodecyl sulfatepolyacrylamide gels. Anal. Biochem. 75:168-176.
38. Watt, R. M., J. Herron, and E. M. Voss. 1980. First orderdissociation rates between a subpopulation of high affinityrabbit anti-fluorescyl IgG antibody and homologous li-gand. Mol. Immunol. 17:1237-1243.
39. Yamato, I., Y. Anraku, and K. Hhrosawa. 1975. Cytoplas-mic membrane vesicles of Escherichia coli. J. Biochem.77:705-718.
VOL. lS0, 1982
on May 22, 2020 by guest
http://jb.asm.org/
Dow
nloaded from






![CALCULATION OF ISOELECTRIC POINTS. · 808 Calculation of Isoelectric Points Except in the special case where the isoelectric point is at the “neutral” point of water [H+] does](https://img.dokumen.tips/doc/110x75/5f0a52187e708231d42b1422/calculation-of-isoelectric-808-calculation-of-isoelectric-points-except-in-the.jpg)




















![[Group 5] electrochemistry, electrophoresis, isoelectric focusing](https://img.dokumen.tips/doc/110x75/55c5bdefbb61eb5a3b8b458a/group-5-electrochemistry-electrophoresis-isoelectric-focusing.jpg)