Embed Size (px)
Citation preview

Írta:
NÉMETH ÁRON
Lektorálta:
REZESSYNÉ SZABÓ JUDIT
IPARI MIKROBIOLÓGIA Egyetemi tananyag
2011
Budapesti Műszaki és Gazdaságtudományi Egyetem Vegyészmérnöki és Biomérnöki Kar Alkalmazott Biotechnológia és Élelmiszertudományi Tanszék

COPYRIGHT: 2011-2016, Dr. Németh Áron, BME Vegyészmérnöki és Biomérnöki Kar
Alkalmazott Biotechnológia és Élelmiszertudományi Tanszék
LEKTORÁLTA: Dr. Rezessyné Szabó Judit, Budapesti Corvinus Egyetem
Creative Commons NonCommercial-NoDerivs 3.0 (CC BY-NC-ND 3.0)
A szerző nevének feltüntetése mellett nem kereskedelmi céllal szabadon
másolható, terjeszthető, megjelentethető és előadható, de nem módosítható.
TÁMOGATÁS:
Készült a TÁMOP-4.1.2-08/2/A/KMR-2009-0028 számú, „Multidiszciplináris, modulrendszerű, digitális
tananyagfejlesztés a vegyészmérnöki, biomérnöki és vegyész alapképzésben” című projekt keretében.
KÉSZÜLT: a Typotex Kiadó gondozásában
FELELŐS VEZETŐ: Votisky Zsuzsa
AZ ELEKTRONIKUS KIADÁST ELŐKÉSZÍTETTE: Benkő Márta
ISBN 978-963-279-478-5
KULCSSZAVAK:
biotechnológia, ipari mikrobiológia, alkalmazott mikrobiológia, mikrobiológiai műveletek, mikrobiális
anyagcsere, mikrobiális rendszertan, ipari baktériumok, ipari gombák, ipari algák, kevert tenyészetek
alkalmazása.
ÖSSZEFOGLALÁS:
Az elmúlt években vitathatatlan biotechnológiai fejlődésnek voltunk és vagyunk tanúi, amely fejlesztések
során mindig feltűnnek a mikroorganizmusok, akár mint termelő biokatalizátorok, akár mint termékek, vagy
egyre gyakrabban mint gén/enzim források. Ennek köszönhetően a biotechnológiai/biomérnöki képzésben az
Általános mikrobiológiát egy specifikus Ipari mikrobiológia követi, amelynek célja rendszerezetten
összefoglalni a mikrobák nyújtotta lehetőségeket, potenciálokat, illetve az ezek kiaknázásához szükséges
alapismereteket.
Ezt szem előtt tartva a jegyzet először a bevezetés után a mikroorganizmusokkal és kezelésükkel kapcsolatos
műveleteket és alapismerteket (fiziológiai, biokémiai) tekinti át, majd pedig a mikrobiális rendszertant
követve – abból kiemelve az ipari vonatkozású törzseket – bemutatja a mikrobákban rejlő ipari
lehetőségeket, és utal a már működő technológiákra.

Tartalom 3
© Németh Áron, BME © www.tankonyvtar.hu
TARTALOM
1. ALAPISMERETEK, IPARI MIKROBIOLÓGIAI MÓDSZEREK ................................................ 5 1.1. Bevezetés az ipari mikrobiológiába ............................................................................................. 5
Vörös biotechnológia .................................................................................................................. 5 Fehér biotechnológia ................................................................................................................... 6 Zöld biotechnológia .................................................................................................................... 6 Esettanulmány 1. – Lizingyártás ................................................................................................ 7 Esettanulmány 2. – 1,3-propándiolgyártás .................................................................................. 9
1.2. (Ipari) Mikrobiológiai műveletek ............................................................................................... 14 Törzsizolálás ............................................................................................................................. 14 Törzsfenntartás .......................................................................................................................... 20 Törzs-screening ......................................................................................................................... 21 Identifikáció .............................................................................................................................. 25 Törzsfejlesztés ........................................................................................................................... 26
1.3. Mikroorganizmusok ipari alkalmazása ...................................................................................... 31 Az oxigén szerepe ..................................................................................................................... 31 Aerobok tenyésztése ................................................................................................................. 32 Anaerob fermentációk (erjesztések) .......................................................................................... 32 Szubmerz kontra felületi fermentáció ....................................................................................... 37
2. AZ IPARI MIKROBIOLÓGIA BIOKÉMIAI ÉS FIZIOLÓGIAI HÁTTERE ............................. 40 2.1. Szénforrások hasznosulása, különleges metabolizmusok .......................................................... 40
C1-források hasznosulása ........................................................................................................ 40 C2-metabolizmus ...................................................................................................................... 41 C3-metabolizmus ...................................................................................................................... 42 C4-metabolizmus ...................................................................................................................... 43 C5-metabolizmus ...................................................................................................................... 43 C6-metabolizmus ...................................................................................................................... 44 C7-metabolizmus ...................................................................................................................... 44 Speciális energianyerő metabolizmusok (biometallurgy) ......................................................... 44
2.2. Primer metabolitok ..................................................................................................................... 46 2.3. Szekunder metabolitok ............................................................................................................... 46
De novo szekundermetabolit-termelés ...................................................................................... 46 Szteroid- (szekunder metabolit) konverzió ............................................................................... 50
3. AZ IPARI MIKROORGANIZMUSOK ÉS TECHNOLÓGIÁK TÁRGYALÁSA
TÖRZSRENDSZERTANBA ILLESZTVE .................................................................................. 57 3.1. Mikrobiális rendszertan .............................................................................................................. 57
Klasszikus rendszertan .............................................................................................................. 58 Filogenetikus rendszertan ......................................................................................................... 58 Bakteriális filogentika .............................................................................................................. 59 A baktériumok törzsének elágazási sorrendje ........................................................................... 60 Az indelmodell megbízhatósága és prediktív lehetőségei ........................................................ 61
3.2. Ipari szempontból fontos (ős)baktériumok ................................................................................. 65 1. Archeák ................................................................................................................................. 65 2. Firmicutes ............................................................................................................................. 68 3. Actinobacterek ...................................................................................................................... 72 4. Thermotoga ........................................................................................................................... 79 5. Fusobacteriumok ................................................................................................................... 80 6.-7. Deinococcus-Thermus ...................................................................................................... 80 8. Chloroflexi ............................................................................................................................ 81 9. Cyanobacterek ....................................................................................................................... 81 10. Spirohaeták ......................................................................................................................... 82 11., 12,. 13. Chlorobik, Fibrobacterek, Bacteroidesek .............................................................. 82

4 1. Alapismeretek, ipari mikrobiológiai módszerek
© Németh Áron, BME © www.tankonyvtar.hu
14., 15., 16. Planctomyceta-k, Verrucomicrobiumok, Chlamydiák .......................................... 83 17. Aquificae ............................................................................................................................. 83 18., 19., 20., 21., 22. Proteobacterek ......................................................................................... 84
3.3. Ipari szempontból fontos gombák .............................................................................................. 95 Basidomycotak .......................................................................................................................... 97 Ascomycotak ............................................................................................................................. 99 Zygomycotak .......................................................................................................................... 100
3.4. Iparilag jelentős algák .............................................................................................................. 101 3.5. Vegyes tenyészetek alkalmazása .............................................................................................. 102 3.6. A vakcinagyártás mikrobiológiája ........................................................................................... 103
4. A MIKROORGANIZMUSOK IPARI FELHASZNÁLÁSÁNAK ETIKAI ÉS KOCKÁZATI
KÉRDÉSEI .................................................................................................................................. 106 5. KITEKINTÉS .............................................................................................................................. 109 6. SZÓSZEDET ............................................................................................................................... 110 7. ÖNELLENŐRZŐ KÉRDÉSEK AZ IPARI MIKROBIOLÓGIA CÍMŰ JEGYZETHEZ ............ 118
Tesztmegoldó kulcs ......................................................................................................................... 128 8. HIVATKOZÁSOK ...................................................................................................................... 129 ÁBRÁK, ANIMÁCIÓK, VIDEÓK, TÁBLÁZATOK JEGYZÉKE ................................................... 138
Ábrák, animációk, videók ............................................................................................................... 138 Táblázatok ....................................................................................................................................... 139

1. Alapismeretek, ipari mikrobiológiai módszerek 5
© Németh Áron, BME © www.tankonyvtar.hu
1. ALAPISMERETEK, IPARI MIKROBIOLÓGIAI
MÓDSZEREK
1.1. Bevezetés az ipari mikrobiológiába
Az 1990-es években sok olyan jóslat látott napvilágot, melyek szerint a XXI. sz. a biotechnológiai
forradalmat követően a biotársadalommal lesz jellemezhető. Valóban, az utóbbi évtizedek mikro-
biológiai, biokémiai, genetikai, molekuláris biológiai és biotechnológiai fejlesztései lassan a hétköz-
napi emberhez is ilyen vagy olyan formában eljutnak, és különösen a környezetvédelmen keresztül
szélesebb társadalmi csoportokban is érezhetővé válnak.
A biotechnológiát – amely tehát a „Biotársadalom” meghatározó része – sokféleképpen defi-
niálták már. Talán a legegyszerűbb úgy megadni, mint egy olyan fogalom, amely a biológiai
rendszerek emberi célú (gyakran termelés orientált) felhasználását jelenti.
Annak bemutatására, hogy az utóbbi évtizedekben mekkorára duzzadtak a biotechnológia kutatási és
alkalmazási területei az 1.1.1. táblázatban került összefoglalásra a különböző szakterületeket jelző
színkódrendszer.
1.1.1. táblázat: Színkódrendszer a Biotechnológiai területek jelölésére
Szín Biotechnológiai kutatási és alkalmazási terület
Vörös Egészségügyi, orvosi, diagnosztikai biotechnológia
Fehér Ipari biotechnológia
Zöld Mezőgazdasági, környezetvédelmi biotechnológia
Kék Vízparti és tengeri biotechnológia
Barna Száraz, sivatagi biotechnológia
Fekete Bioterror, biológiai fegyverek
Bíbor Biotechnológiai találmányok, szabadalmak
Sárga Élelmiszer- és táplálkozástani biotechnológia
Arany Bioinformatika, nanotechnológia
Szürke Klasszikus fermentáció és Bioprocess
Noha az egyes színek használata még nem széleskörűen elterjedt, és olykor diszkrepanciával
terhelt, a táblázatban szereplő első három területet már szinte egy évtizede e három színkóddal jelölik,
ezért ezeket hasonlítjuk össze az alábbiakban az ágazatot jellemző termékek, mikrobiológiai,
biokémiai és technológiai igények szempontjából.
Vörös biotechnológia
Mivel a vörös biotechnológia termékeit az egészségügyben gyógyászati célokra használják (diagnosz-
tikumok, vakcinák, gyógyszerek), ezért termékoldalról „kis mennyiség – nagy érték” (low volume –
high value) jellemzi őket. Ez azt jelenti, hogy az éves világtermelés termékenként néhány száz tonna,
amelyek ára viszont igen magas (penicillin: 10-12 $/kg, vakcinák: 20-30 $/dózis). A vörös bio-
technológiai termékek előállításakor – legyenek azok (rekombionáns) fehérjék vagy biokonverzióval
nyert másodlagos anyagcseretermékek (ld. később) – enzimeket vagy nyugvó-, illetve aktívan
szaporodó tenyészeteket használnak. Biokémiai szempontból a termelő törzsekkel szemben támasztott
követelmény, hogy jó hozammal, kevés szennyező melléktermék kíséretében termeljék a célterméket.
Ezt gyakran törzsfejlesztéssel érik el, illetve a termelő mikroorganizmussal szemben támasztott másik
mikrobiológiai elvárás gyakran a jó expressziós képesség (megfelelő fehérjeszerkezet kialakítása mind
a 4 szinten1, lehetőleg extracellulárisan). Technológiai oldalról talán a legmeghatározóbb a minőség-
1 Fehérjeszerkezet 4 szintje: Elsődleges szerkezet: aminosavsorrend; másodlagos sz.: hosszabb szakaszok
térrendeződése a-hélix vagy b-redő formába; harmadlagos szerkezet: 3D „feltekeredés”, amelyet másodlagos
kötések stabilizálnak; negyedleges: alegységek kapcsolódási szerkezete (pl.: 222)

6 Ipari mikrobiológia
© Németh Áron, BME © www.tankonyvtar.hu
biztosítás, amelynek szinte mindent alárendelnek, és amely a költségeket is jelentősen befolyásolja
(=mindig ugyanolyan kiváló minőség).
Fehér biotechnológia
A fehér biotechnológia célja a „vegyipar kifehérítése”, azaz környezetkímélőbbé tétele. Ez céljaiban
hasonló a széleskörűen használt „zöldkémia” kifejezéssel, ám míg ez utóbbi a kémiai szintézisek
környezetkímélőbbé tételét kívánja elérni, addig a fehér biotechnológia a kémiai szintézisek mellett
biológiai gyártásokat kíván a vegyiparban meghonosítani, mivel ezek energiaigénye és környezet-
terhelése általában kisebb az azonos terméket előállító vegyipari eljárásnál. A vegyipar jelenleg is
fennálló struktúrája kőolajalapú, azaz ásványi olajból nyerik finomítással az elsődleges alapanyagokat,
illetve az energiát is. Ennek analógiájára a fehér biotechnológia eredményeit felhasználva kb. egy
évtizede terjed a biofinomító elnevezés, amely tehát olyan biokombinát, amely megújuló alapanyagból
fedezi a nyersanyag- és energiaigényét is.
A biofinomítókhoz szorosan kapcsolható a platformalkotó vegyület fogalma, amelyet olyan
vegyületekre használunk, amelyeknek egy sor piacképes, értékes származékát konvencionális vegy-
ipari folyamatokkal elő lehet állítani. A platformvegyületeket általában szénatomszám szerint csopor-
tosítják, így léteznek C3-C4-C5-C6-C7 platformok. A biofinomítók gyakran biológiai úton megújuló
nyersanyagból ilyen platform kemikáliát állítanak elő.
A fehér biotechnológia termékeit is a vegyipari termékekhez hasonlóan – amelyek más iparágak
(kozmetikai, oldószer, polimer iparok) viszonylag olcsó tömeges alapanyagaiként szolgálnak – a
„nagy tömeg – kis érték” (high amount – low value) jellemzi, és általában de novo fermentációs
eljárással készülnek (pl.: oldószerek 0,4-1,4 $/kg).
Biokémiai szempontból az olcsó tömegtermékek miatt a jó szubsztráthasznosulás még fontosabb,
illetve a megfelelő metabolikus utak aktiválása és a többi inaktiválása is csökkenti a feldolgozási
(downstream) és termékkinyerési költségeket. Mikrobiológiai szempontból a termelő törzs termék-
toleranciája fontos, és a törzsfejlesztésnek itt is fontos szerepe van. Technológiai aspektusból az
abszolút költséghatékonyság a cél mindenekfelett (olcsó up/downstream). A kis termelői és keres-
kedelmi árrés miatt a hatékony technológia mellett nagy mennyiségek előállítása, azaz nagy
termelőkapacitások szükségesek minimális műveleti lépés mellett.
Zöld biotechnológia
A zöld biotechnológiába korábban beleértették a környezetvédelmi, mezőgazdasági és élelmiszer-ipari
ágazatokat, ám ezek lassan külön színkódot kapnak. Az viszont mindegyikre igaz, hogy a termékeik
nagy tömegben kerülnek előállításra kicsi vagy közepes értékkel (takarmányok, étrend- kiegészítők,
biopeszticidek, talajjavítók stb.).
Biokémiai szempontból a speciális ízek kialakításhoz az egy termékű de novo fermentációk
helyett a heterofermentatív anyagcseréjűek kedvezőbbek, illetve a speciális tápértékek kialakításához
különböző anyagok (fehérjék, vitaminok) akkumulációjára képes mikrobák kerülnek felhasználásra.
Olykor enzimes technológia is előfordul (izocukorgyártás). Ehhez mikrobiológiai szempontból
különösen a biopeszticidek gyártásakor szükséges továbbá a haszonnövény-károkozó viszonyának, és
a károkozás mechanizmusának ismerete. Technológia szempontból – különösen az élelmiszer-ipari
termékeknél – a minőségbiztosítás ismét jelentős szerephez jut, amely erősen felhasználó centrikus
(kedvező színű, szagú, halmazállapotú termék).
A fenti BIOTECHNOLÓGIAI példákon igyekeztünk demonstrálni, hogy noha a biotechnológia
mára egy sok területet átölető ágazattá vált, az egyes szakterületek közös pontja a MIKROBA, amely
az egyes technológiákat és termékeket alapvetően befolyásolja. Például egy tejsav-fermentációnál az
alkalmazott tápközegnek olcsónak kell lennie, de a mikroba valamennyi igényét (C, N, vitaminforrás)
ki kell elégítenie úgy, hogy a feldolgozási műveletek a lehető legegyszerűbben (és legolcsóbban)
nyerhessék ki a terméket. Ahhoz, hogy a biotechnológia kulcsszereplőjét, a mikrobát megismerjük, és
céljainknak megfelelően a leghatékonyabban kihasználjuk, a mikrobatudománnyal, de legalább annak
ipari mikrobiológiai részével kell tisztában lennünk.

1. Alapismeretek, ipari mikrobiológiai módszerek 7
© Németh Áron, BME © www.tankonyvtar.hu
E jegyzet célja, hogy az iparban használatos mikrobiológiai műveleteket, élettani jelenségeket,
potenciálokat bemutassa. A napjainkban egyre nagyobb méretet öltő szövettenyésztési eljárások
azonban nem kerülnek tárgyalásra.
Amint azt a vörös/fehér/zöld biotechnológia rövid bemutatásánál láttuk, az ipari mikrobiológia 3
tudományterület között elhelyezkedő interdiszciplináris tudomány: Biokémia, Mikrobiológia, Mérnöki
tudományok (1.1.1. ábra).
1.1.1. ábra: Az ipari mikrobiológia helye a tudományban
Az ipari mikrobiológia tárgya tehát szükségképpen a termelő mikroorganizmusok (természetes és
rekombináns törzsek, illetve kevert tenyészetek), valamint a hozzájuk kapcsolódó (mikrobiológiai)
műveletek.
Bevezetésként következik két esettanulmány az ipari mikrobiológia eredményeinek demonstrálására.
Esettanulmány 1. – Lizingyártás [1]
OH
O
NH2
NH2
1.1.2. ábra: A lizin képlete
A lizin egy esszenciális aminosav (1.1.2. ábra), ezért előállítása mind az állati takarmányozás, mind a
humán élelmezés szempontjából fontos. A lizinpiac évi 600 000 t terméket produkál, amely évente
további 5-7%-kal bővül (2005).
Az előállítás első kulcslépése a fermentáció. Ezt glükóz/szaharóz- vagy melaszalapon végzik, és
30-40%-os hozammal képződik a lizin (100–150 g/L koncentrációban).
A fermentációt Corynebacterium glutamicum-mal végzik, amelyet először 1956-ban izoláltak egy
korabeli „új” módszer segítségével (bioautográf technika) glutaminsavtermelés céljából. A módszer
lényege, hogy az izolátumokat Petri-csészében lévő agar tápközegen kinövesztik, replika módszerrel a
Petri-csészéről másolatot készítenek, majd a másolaton kinőtt telepeket UV-val elölik, végül föléje egy

8 Ipari mikrobiológia
© Németh Áron, BME © www.tankonyvtar.hu
újabb agaros tápközeget rétegeznek, amelybe a keresett metabolitra hiánymutáns mikrobát
szuszpendálnak, és újra inkubálják. Ennek következtében az elölt izolátumok már nem, a
hiánymutánsok pedig csak ott fognak kinőni a 2. agar felszínén, ahol az alsó agaron kinövesztett
izolátum a hiányzó metabolitot termelte (1.1.3. animáció) [2]. Számos izolátum vizsgálatából kiderült,
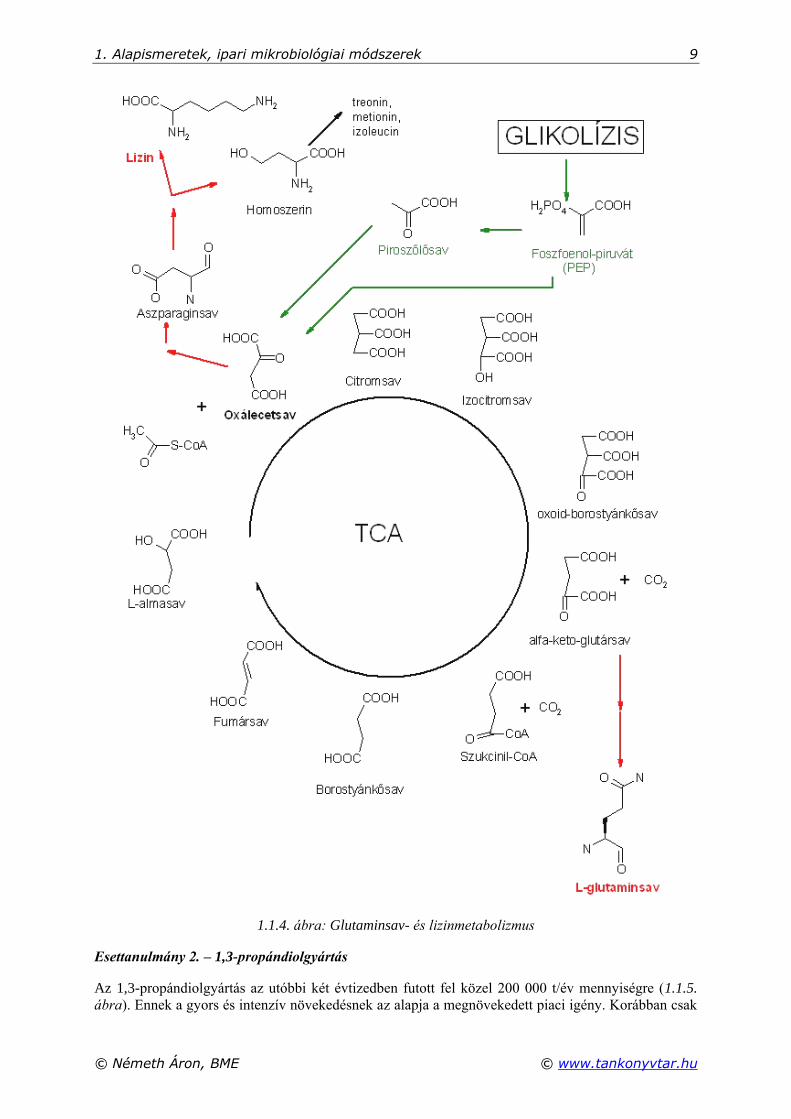
hogy a glutaminsav-termelők lizintermelésre is alkalmasak, mert mindkét aminosav a citrátkör (TCA
ciklus) intermedieréből eredeztethető (1.1.4. ábra).
1.1.3. animáció: A bioautográf módszer
A mikroorganizmus által szénforrásként elfogyasztott cukrok a glikolízisen keresztül bomlanak le
piroszőlősav – foszfoenolpiruvát – acetil-koenzim-A útvonalon. Az utóbbi molekula az acetilcso-
portját egy oxálecetsavnak átadva citromsavat hoz létre, amely két lépésben dekarboxileződik (2 szén-
dioxid keletkezése közben) és oxálecetsav marad vissza, a következő acetil-csoport eloxidálására. A
körfolyamat egyes intermedierjei azonban az aminosavfermentáció esetén beépülnek az előállítandó
aminosavba, így a körfolyamat megszakadna. Ennek ellensúlyozására működnek az anaplerotikus
(feltöltő) reakciók, amelyek szintén a cukrokból a piruváton át vagy akár a foszfoenolpiruvátból
közvetlenül is oxálecetsavat termelnek. Az 1.1.4. ábrán az is látható, hogy a TCA-ból a lizin felé
vezető metabolikus út elágazik, és homoszerinen keresztül további aminosavak (treonin, metionin,
izoleucin) képződnek, csökkentve ezzel a bevitt cukrok lizinre történő hasznosulását (hozamát).
Ennek okán a kezdeti (’60-as évek, 20%-os hozam [1]) törzsfejlesztések homoszerin útvonal
nélküli törzsek kialakítására irányultak, amelyek előállításához a klasszikus random mutáció –
szelekció ciklikus ismétléses módszerét alkalmazták. A sejtek élettani funkcióihoz azonban szükség
volt azokra az aminosavakra is, amelyek a homoszerin útvonalon képződtek, így a homoszerin
hiánymutáns (Hom-) törzsek termelőképessége nem az elvárt mértékben javult. Ennek kiküszöbölésére
a hiányzó aminosavakat a tápközegbe kellett adagolni, ami növelte az eljárás költségeit. Ennek
elkerülésére kifejlesztették a „leaky mutáns” (csepegő, áteresztő) módszert, amely olyan mutánsok
szelektálását célozta meg, amelyekben nem eltűnt, hanem jelentősen lecsökkent egy útvonal kulcs-
enzimének aktivitása. Így a mellékútvonal csak kis anyagáramot (fluxust) „pocsékolt” el, viszont
egyetlen aminosavból sem volt teljes hiánya a sejteknek.
A következő, még mindig klasszikus törzsfejlesztéssel kivitelezett cél olyan regulációs mutánsok
létrehozása volt, ahol a negatív visszacsatolások eliminálódtak. Például a képződött lizin az
aszparagin-kináz enzimet magasabb koncentrációban már gátolja, amely így az aszparagin – lizin
útvonal leállásához vezet. Mivel az ipar számára a költséghatékonyabb termelés és léfeldolgozás a cél
– azaz a magas termékkoncentráció az előnyös –, az ipari fermentáció számára fontos volt, hogy az ezt
akadályozó regulációs mechanizmusokat kiiktassák. A „leaky mutánsok” szabályozási (regulációs)
szempontból is jelentősnek bizonyultak, mivel a csökkent fluxus kisebb koncentrációkat eredmé-
nyezett a melléktermék-útvonalon, így a melléktermékek negatív feedback mechanizmusa elhanya-
golhatóvá vált.
A modern kori (’80-as évek) törzsfejlesztések során új módszereket vetettek be. Ezek közé
tartozik a metabolikus fluxus analízis (MFA), amelyet izotópos nyomkövetéssel is alátámasztottak.
Ezekkel a módszerekkel megállapítást nyert, hogy anyagáram szempontjából az anaplerotikus
reakciók jelentik a szűk keresztmetszetet. Megállapították azt is, hogy a két lehetséges feltöltő
mechanizmus közül a piruvátkarboxiláz enzimen keresztül vezető út felel az anaplerotikus aktivitás
90%-áért.
A fenti törzsfejlesztések eredményeképpen napjainkban 150 g/L végtitert és kb. 50%-os hozamot lehet
elérni a lizinfermentáció során.
1. Alapismeretek, ipari mikrobiológiai módszerek 9
© Németh Áron, BME © www.tankonyvtar.hu
1.1.4. ábra: Glutaminsav- és lizinmetabolizmus
Esettanulmány 2. – 1,3-propándiolgyártás
Az 1,3-propándiolgyártás az utóbbi két évtizedben futott fel közel 200 000 t/év mennyiségre (1.1.5.
ábra). Ennek a gyors és intenzív növekedésnek az alapja a megnövekedett piaci igény. Korábban csak

10 Ipari mikrobiológia
© Németh Áron, BME © www.tankonyvtar.hu
oldószerként, kenőanyagként és fagyállóként alkalmazták, ám a ’90-es évek óta a műanyagipar
használja fel legnagyobb mennyiségben a poli-trimetilén-tereftalát (PTT) előállítására. A PTT
különleges szerkezettel rendelkezik a hasonló célokra hagyományosan használt polietilén-tereftaláthoz
(PET) és polibutilén-tereftaláthoz (PBT) képest. (1.1.6. ábra).
0
50.000
100.000
150.000
200.000
250.000
1990 1995 2000 2005 2010
PD
[t/
év
]
1.1.5. ábra: Az 1,3-propándiol-előállítás alakulása az utóbbi két évtizedben
1.1.6. ábra: A propándiol okozta különleges szerkezetű PTT polimer
(PET: polietilén-tereftalát, PTT: poli-trimetilén-tereftalát, PBT: polibutilén-tereftalát,
ET: etilén, PR: propilén, BU: butilén, TFS: tereftálsav)
Az ábráról látszik, hogy a propándiollal alkotott kopolimer szerkezete különleges hajtogatással
rendelkezik. Ennek oka az, hogy míg mind az etilén, mind a butilén esetében a diol OH csoportok
szimmetrikusak egymáshoz képest és tökéletes cisz vagy transz helyzetűek, addig a propándiol
esetében az OH csoportok hidrogénjei (amelyek helyére a ftálsav köt) egymással szöget zárnak be
(1.1.7. animáció). Ez a különleges szerkezet igen előnyös tulajdonságokat kölcsönöz a PTT-nek,
amelynek következtében egyre kiterjedtebben használják fel textiliák, szálak, szövetek, ecsetszőrök
gyártásában.
1.1.7. animáció: 1,3-PD különleges szerkezete

1. Alapismeretek, ipari mikrobiológiai módszerek 11
© Németh Áron, BME © www.tankonyvtar.hu
Noha mindkét monomert napjainkig javarészt szintetikusan állítják elő kőolaj-alapanyagon (PD:
akrolein hidrolízisével vagy etilénoxid hidroformilezésével), az 1,3 propándiolt fermentációs úton is
előállítják, így a belőle készített műanyagok „megújulóvá” váltak (a fermentáció alapjául szolgáló
cukrot növényekből nyerik).
A szintetikus eljárások esetén nagy nyomást és hőmérsékletet, valamint nehézfém-katalizátorokat
használnak (1.1.8. ábra), ugyanakkor a DuPont és a Tate & Lyle által közösen felépített fermentációs
üzem pedig ehhez képest 30%-kal kevesebb energiát igényel és 55%-kal kevesebb üvegházhatású gázt
termel az enyhébb körülményeknek köszönhetően.
O
H
OH O
H
O OH OH
1,3-propándiol
H2O, H+
50-70°C
H2, Ni
135bar, 75-140°C
Akrolein
Etilénoxid
+ CO + H2100°C, 100bar
katalizátor
Degussa:
Shell Chemicals:
Dupont+Tate&Lyle: kukorica mag keményítõ glükóz glicerin55-90°C 25-30°C
E.coli
1.1.8. ábra: az 1,3-propándiol-előállító technológiák
Ipari mikrobiológiai szempontból a termelő törzset érdemes megvizsgálni.
A természetben előforduló mikroorganizmusok közül a lactobacillusok, klebsiellák, citrobacterek,
enterobacterek, clostridiumok [5] képesek glicerin anaerob (levegő kizárt) fermentációjával 1,3-
propándiolt előállítani.
Amint az az 1.1.9. ábrán látható, a fenti mikrobák a glicerint egy oxidációs lépést követően
(glicerin-dehidrogenáz enzim EC 1.1.1.6) a glikolízisen és a citrátkörön keresztül hasznosítják. Mind a
glikolízis, mind pedig a TCA-ciklus redukált koenzimet termel (NADH2), amely aerob esetben a
terminális oxidációban a légköri oxigén segítségével víz keletkezése mellett oxidálódik vissza
(regenerálódik). A felsorolt mikrobák egyik része fakultatív anaerob anyagcseréjű, azaz levegő
jelenlétében is és anaerob körülmények között is képes szaporodni, a másik részük pedig obligált
anaerob metabolizmussal rendelkezik. 1,3-propándiol azonban kizárólag az anaerob körülmények
mellett jelenik meg, mert élettani szerepe a terminális oxidáció pótlása, azaz a koenzim vissza-
oxidálása anaerob körülmények mellett.
Mindezek alapján a következő megfontolások tehetők:
1) a természetes termelő törzsek nem alkalmasak ipari termelésre, mert A) patogének, B) sok
más metabolit mellett kevés 1,3-propándiolt termelnek C) szubsztrát inhibíció vagy
termékgátlás áll fent, D) csak glicerinből képesek 1,3-propándiol előállítására (a glicerin mint
alapanyag csak a biodízelgyártás elterjedése és a szennyezéstűrő – metanol, sók, katalizátorok
– mikrobák kifejlesztésével vált jelentőssé).
2) A biotechnológusok „háziasított” mikrobája, az Escherichia coli csak valamelyik természetes
propándiol termelő törzs enzimjeinek segítségével tehető képessé a propándiol-előállításra, ám
ehhez még szükséges valamilyen mikrobiális (Saccharomyces) gén és géntermék, amelyekkel
a glükózból a glicerin előállítható.
3) Jól termelő törzs kifejlesztéséhez tehát ezúttal nem a klasszikus izolálás-screening-mutáció
screening műveletsorral, hanem rekombináns technikával jutottak.
A természetes 1,3-propándiol-termelő törzsekből 4 kulcsenzimet volt szükséges az Escherichia coli-ba
átvinni: 1) gliceirn-dehidrogenáz (GDH, EC 1.1.1.6, NAD+ koenzim segítségével a glicerint 1,3-

12 Ipari mikrobiológia
© Németh Áron, BME © www.tankonyvtar.hu
dihidroxiacetonná alakítja; 2) glicerin dehidratáz (GDHt, EC 4.2.1.30, amely B12 koenzim segítségével
gyökös reakcióban vizet hasít le a glicerinről); 3) 1,3-propándiol-oxidoreduktáz (PDOR, EC 1.1.1.202,
amely redukált NADH2 koenzim segítségével redukálja a vízkilépéssel kapott 3-hidroxi-
propionaldehidet 1,3-propándiollá (1.1.10. ábra). Ezáltal a sejten belül a glicerin diszproporciója
valósul meg.
1.1.9. ábra: Az anaerob glicerin metabolizmus

1. Alapismeretek, ipari mikrobiológiai módszerek 13
© Németh Áron, BME © www.tankonyvtar.hu
CH2
CH
CH2
OH
OH
OH
CH2
CH
CH2
OH
OH
OH
CH2
CH2
OH
OH
O
CH2
CH2
CH O
OH CH2
CH2
CH2
OH
OH
NADH2NAD+
Glicerin HPA
Glicerin
PD
DHA
DHA-foszfát Glikolízis
H2O
GDH
PDORGDHt
1.1.10. ábra: A glicerinmetabolizmus enzimei: GDH, GDHt, PDOR,
(DHA: dihidroxiaceton, HPA: hidroxipropionaldehid, PD: 1,3-propándiol)
A 4. kulcsenzim (ez idáig „rendszertanba” nem sorolt) a GDHt kulcsenzim reaktivációjához
szükséges, mivel a GDHt minden molekula átalakítása után a B12 koenzimével komplexben marad,
ahonnan ez az enzim szabadítja fel mind a koenzimet, mind a kulcsenzimet ATP energia segítségével.
Ahhoz, hogy glükózalapon lehessen 1,3-propándiolt előállítani, még további 2 enzimre volt szükség:
G3DH (glicerin-3-foszfát-dehidrogenáz) és GPP (glicerin-3-foszfatáz).
Ezekkel a génátvitelekkel már képessé tették az E.coli-t az 1,3-propándiol-termelésre, de még
ipari használatra nem volt alkalmas. A gyártásba való bevonás előtt a termékinhibíció hatását ki kellett
küszöbölni (terméktoleráns mutánsok), illetve a melléktermékeket (további metabolitokat) termelő
útvonalakat kellett minimalizálni. Mindezek végére az elkészült törzs 160 g/L PD-t is képes
előállítani, ha a B12 koenzimet adagolják (ezt nem termli az E. coli). Az 1,3-PD technológiája
követhető nyomon az 1.1.11 hiperpicture-ön.
1.1.11. hyperpicture: A rekombináns PD fermentációs technológia
A technológia első lépése a kis léptékű oltóanyag-előállítás 2L-es rázatott lombikban. A
laboratóriumi steril 2YT tápközeget oltják be a termelő E.coli fenntartó (ld. Törzsfenntartás, ferde
agaros kémcső, Petri-csészés tenyészet, liofilizált ampulla) tenyészetéről. A fermentáció 34°C-on
zajlik, és egyéb C-forrás hiányában a szerves nitrogénforrások széntartalmából (tripton, élesztő)
épülnek fel a kultúra sejtjei. A rázatás a keverés mellett a levegőztetést is biztosítja. Mintegy 10h
tenyészidő után kerül az oltóanyag az előkészített (tápközeggel feltöltött lesterilezett) 1500 L-es
oltóreaktorba. A rekombináns technológia alkalmazásának egyik hátránya, hogy a termelő hasznos
mutánsok megtartásának érdekében állandó szelekciós nyomást kell fenntartani, és ezt ez esetben is –
mint általában – antibiotikum hozzáadásával oldják meg, mivel az antibiotikum-rezisztencia a
heterológ génekhez van kapcsolva. Ebben a léptékben tehát már antibiotikum (Spectinomycin
250g/10m3) hozzáadásával védik a termelőképesség megmaradását, illetve tápközegnek nem egy
hagyományos laboratóriumi, hanem a termelő törzsre optimált tápközeget használnak. A fermentációt

14 Ipari mikrobiológia
© Németh Áron, BME © www.tankonyvtar.hu
33-34°C-on és pH=6,8-on végzik 0,3VVM (DO=10%) levegőztetés mellett kevert levegőztetett
bioreaktorban, szakaszos technikával 21h hosszan.
A termelő reaktorban ugyanezen körülmények és 34%-os hozam mellett történik glükózalapon
42h hosszan az 1,3-propándiol fermentációja. A termékképzést B12 koenzim hozzáadásával (két
részletben) indukálják, mivel a gazdaszervezet (E.coli) nem képes ennek szintézisére, ugyanakkor a
GDHt (2. kulcsenzim) aktivitásához szükséges. A termelő reaktorban azonban a glükózt rátáplálással
biztosítják, és mintegy 10g/L körüli értéken tartják.
Az elkészült fermentlevet „visszasterilezés” után (75°C-os inaktiválás 45 p-en keresztül) átadják a
feldolgozóüzemnek, ahol első lépésként az elölt és részben flokkulált sejtektől mikroszűréssel
szabadulnak meg (termék a permeátumban található). További membrán műveletekkel (ultra-, majd
nanoszűrés) és ipari kromatográfiával szabadulnak meg előbb a fehérjék nagy részétől, később
valamennyi kísérő szennyező anyagtól.
A fenti két példa megmutatta, hogy a klasszikus véletlenszerű mutáció-szelekció folyamat és
rekombináns DNS technika miként hasznosulhat technológia szinten, illetve hogyan térülhetnek meg a
mikrobiológiai törzsfejlesztésre fordított források.
1.2. (Ipari) Mikrobiológiai műveletek
Minden iparban használt mikroba „őse” egy természetből szándékkal vagy véletlenül kiválasztódott
(=izolált) mikroba. Az ipari termékspektrum, illetve termelési hatékonyság növelése érdekében fontos
további hasznosítható metabolitok, illetve mikrobák felkutatása, valamint gyakran a korábbinál
hatékonyabb termelők keresése. E folyamat három lépésből áll: kinyerés (izolálás), szűrővizsgálat
(screening), azonosítás (identifikáció). E három művelet igen szorosan épül egymásra, hiszen egy
fajokban dús természetes élőhelyről nagyon sok fajta mikroba tenyészthető ki, ezért a nagyobb
hatékonyság érdekében az izolálást már előszűréssel kombinálják, vagyis az adott feladatnak
megfelelő tulajdonságokra szelektíven történik az izolálás.
Törzsizolálás
A mikroorganizmusok legnagyobb számban és diverzitásban a különböző talajok felső rétegeiben
fordulnak elő, de természetesen (tenger)vízből és levegőből is lehet mikrobákat izolálni. A talaj
kulcsfontosságát a mikrobák izolálásában mi sem tükrözi jobban, mint hogy a törzskereséssel és
fejlesztéssel foglalkozó intézmények (törzsgyűjtemények, gyógyszergyárak) gyakran őriznek
talajmintákat a világ legkülönbözőbb helyeiről, és ezekről indítják az izolálást.
Az izolálási technika függ attól, hogy A) célzottan, egy a szakirodalom alapján megfelelőnek vélt
mikrobát találjunk, vagy B) minden potenciális mikrobát szeretnénk kinyerni, amelyek egy adott
tulajdonsággal rendelkeznek.
Szilárd mintából
A szilárd mintából/ról történő izolálást foglalja össze az 1.2.1. ábra.
A) Ha egy célmikrobát szeretnénk izolálni, akkor az első szelektív lépés lehet a minta különböző
intenzitású szárítása, amelynek következtében először a vegetatív sejtek (hifák, baktériumok)
pusztulnak el, aztán az egyre kitartóbb spórák is.
B) Ha minden kedvező tulajdonságú (pl. enzimtermelő) mikroba potenciális kiválasztott lehet,
akkor nem célszerű szárítással „frakcionálni” a talajminta mikroflóráját.
A mintát tehát előszárítással vagy anélkül 1) folyékony tápközegbe juttatjuk, és hígítás után vagy
anélkül szélesztjük (soil-dilution-plate method), vagy 2) Petri-csészés agarlemez felszínére szórjuk, és
steril vízzel nedvesítjük, majd szétkenjük (soil-plate method), vagy 3)steril pálcával a mintafelszínéről
veszünk mintát, és ezt érintkeztetjük Petri-csészés vagy folyékony tápközeggel.

1. Alapismeretek, ipari mikrobiológiai módszerek 15
© Németh Áron, BME © www.tankonyvtar.hu
1.2.1. ábra: Izolálás szilárd mintáról
Talajmintából történő izolálást mutat be az 1.2.1. videofilm.
1.2.1. videó: Izolálás
Bármelyik módon is került a minta érintkezésbe a tenyésztő tápközeggel, általában a homogenitás
a cél, illetve inkubálással kinöveszteni az izolátumo(ka)t.
Az izolálás során preszelekció céljából a tápközeget különböző adalékokkal egészíthetik ki. Az
adalékok célja lehet a másfajtájú mikrobák elnyomása (pl. baktériumok izolálásakor gombák,
penészek visszaszorítása), illetve a célmikroba iránti szelektivitást lehet így növelni (speciális
szubsztrátok sók alkalmazása).
A klasszikus ipari mikrobiológia során szintén talajmintából izolálták a főbb antibiotikum termelő
törzseket. Mivel az antibiotikumok általában gyógyszerként kerülnek piacra, ezért az izolálástól a
termék- (és profit) termelésig hosszú út vezet, amely során a tiszta tenyészet izolálását követően

16 Ipari mikrobiológia
© Németh Áron, BME © www.tankonyvtar.hu
fermentációs fejlesztéseket (tápközeg-optimálás, pH-, hőmérséklet-, cukorprofil stb. meghatározása),
biológiai-, majd kémiaiaktivitás-teszteket végeznek, végül toxikológiai és klinikai vizsgálatok (3
fázisban) után kerül sor a piaci bevezetésre.
Folyadékmintából
A folyadék fázisú minta előnye, hogy könnyen érintkeztethető a tápoldattal, viszont általában
kevesebb csírát tartalmaz, ezért első lépésként dúsításra van szükség. Ennek érdekében vagy 1) magát
a mintát inkubálják, és ezt követően adják a tápoldathoz, majd az újabb inkubáció után oltják
agarlemezre, vagy 2) tápoldattal együtt inkubálják, és erről oltanak szilárd táptalajra, vagy 3) steril
szűrőmembránon átszűrik a mintát, és a membránt inokulumként adják a tápközeghez (oldatba vagy
agarlemezre).
Levegő-(gáz)mintából
Ez esetben a legkisebb a csíraszám, ezért itt 1) intenzív dúsítást alkalmaznak a gáz átszűrésével vagy
2) nyitott Petri-csészés agarlemezzel „exponálnak”. Ilyenkor a nyitott és tápközeggel töltött agarlemez
érintkezik a gáztérrel, és a benne lévő mikrobák inkubáció után megjelenhetnek az agaron.
Az alábbiakban különböző rendszertani csoportok izolálásakor alkalmazott körülményeket mutatjuk
be.
1. Fonalas baktériumok (Actinomyces-ek) izolálása
Előfordulás: leggyakrabban a Streptomyces és Micromonospora fajok fordulnak elő mezőgazdasági
talajokban, tavak folyók és tenger üledékeiben. Termofil fajták silókban és komposztban
is megtalálhatók. Az Actinplanes fajok (antibiotikum ((teichoplanin)) termelő) avarból és
talajból izolálhatóak, különlegességük, hogy aktívan mozgó spórákkal rendelkeznek,
amelyek kemotaxissal keresik meg a kedvezőbb körülményeket (huminsav, KCl, kitin).
Streptosporangiumok (szintén antibiotikumtermelők) száraz, alkalikus talajból izolál-
hatóak. Az Actinomycesek közé tartoznak a Corynebacterium-ok is (ld. Lizingyártás,
illetve 3. fejezet Actinomyceták).
Mintavétel: a talaj felső 4-5cm-es részéből történik, a minta tárolása ideálisan -20–30°C között
lehetséges. A minta kíméletes szárításával a konkurens Gram-negatív baktériumok
elölhetőek, magasabb hőmérséklet (120°C, 1h) alkalmazásával pedig minden vegetatív
baktérium és gomba eliminálódik.
Tápközeg: gyakran gombaölő szerekkel (cikloheximid, nystatin) akadályozzák meg az izolátum
penészesedését. Az adalékolással az izolálás céljaira a kemotaxis is kiaknázható.
Szelektív szénforrásként kolloid kitint vagy sók, vitaminok kíséretében huminsavat
szoktak alkalmazni.
Kitenyésztés: a mezofilek (Topt.:25-30°C) 7-14 nap alatt nőnek ki, a termofilek (Topt.: 45-50°C) 2-5 nap
alatt.
2. Anaerobok izolálása
Előfordulás: levegőtől elzárt, gyakran extrém élőhelyeken, mint például bendőben (biogáztermelő
metanogének), mélytengeri üledékekben (Streptomyces griseus B12-gyártáshoz), gejzírek-
ben, hőforrásokban (termostabil enzimek Archea baktériumokból (=”Ősbaktériumok”)
Mintavétel: „Hungate” technikával, amely részletesen a www.dsmz.de weboldalon olvasható (szep-
tumos tenyésztő edényt injekciós fecskendőbe zárt inert gázzal öblítik át, majd szintén a
tűn keresztül oltják be, illetve vesznek mintát).
A szigorúan anaerob mikrobák érzékenyek a légköri oxigénre (amely reaktív peroxid
gyököket okoz a sejtben), ezért a minták kezelésekor a levegő teljes kizárására kell
törekedni.

1. Alapismeretek, ipari mikrobiológiai módszerek 17
© Németh Áron, BME © www.tankonyvtar.hu
Tápközeg: szükséges oxigéninikátor (pl.: resazurin), és gyakran redukált komponensek alkalmazása
(ezek oxidálódjanak a sejtkomponensek helyett), mint például: NaSH, DiTioTreitol
(DTT), cisztein stb.
Kitenyésztés: Anaerob boksz gázadszorbenssel vagy „gáztér átöblítés” oxigénmentesített (oxigén-
mentesített) (lugol oldaton átbuborékoltatott) inert (N2) gázzal (1.2.2. ábra).
A gázadszorbens működhet úgy, hogy citromsavat, mészkövet és diatóma földet kevernek
össze, és nedvesítés után zárják be az anaerob tenyésztőedénybe. Ekkor a diatóma föld
megköti az oxigént, a savas közegben a karbonát CO2-t bocsát ki, azaz kicserélődik a
levegő oxigénje CO2-ra.
Egy másik esetben az adszorbensből H2 és CO2 fejlődik, és előbbi palládiumkatalizátor
mellett vízzé reagáltatja az oxigént.
Az inert gázzal történő átöblítést az edény másik csonkjának megvákuumozásával is elő
lehet segíteni.
1.2.2. ábra: Anaerob tenyésztőrendszerek
3. Fonalas gombák izolálása
Előfordulás: heterotróf szaprofita fajaik talajban, illetve rothadó szerves anyagokon találhatók meg
minden égövben. Egyes fajaik szimbionita életformában zuzmókban, illetve mikorrhi-
zában fordulnak elő, de vannak paraziták is, amelyek a gazdanövényről, rovarról, halról,
emlősről stb. izolálhatóak.
Mintavétel: mivel szinte mindenhol előfordulnak, kétféle megoldás használatos: 1) „Ökológiai
megközelítés”: a kívánt tulajdonságnak megfelelő élőhelyről próbálnak izolálni (pl.:
termostabil gombákat hőforrásokból, vegyszerlebontókat szennyezett talajból stb.) 2)
„Taxonómiai (rendszertani) megközelítés”: rokonok tulajdonságai alapján pl.: citromsav-
termelő faj rokonait is be lehet vonni izolálás+screening programba jobb citrom-
savtermelő keresésekor. 3) „Csalétek módszer”: lebontandó anyagot párás melegbe helye-
zik, így az „megpenészedik”, azaz olyan fonalas gombák kolonizálják, amelyek képesek
lebontani és felhasználni a „csalétket”.
Előkészítés: kíméletes szárítással vagy hűtéssel (+5– -18 °C).
Tápközeg: széles irodalommal rendelkeznek. Általában enyhén savanyú (pH=4,5-6), magas C-
forrás- tartalmú tápközegeket használnak, amelyekben a N-forrás gyakran csak ammónia
vagy nitrát, de a leggyakoribb fonalasgomba-táptalajok szerves nitrogénforrást is
tartalmaznak (pl.: PDA, kukoricaliszt agar, malátakivonat stb.). Ezek a tápközegek
szelektívvé vagy félszelektívvé tehetők (100-200 mg/l) antibiotikum kiegészítéssel,
illetve a gyorsan kolonizáló („burjánzó”) penészek ellen adalékokkal kiegészíthetőek.
18 Ipari mikrobiológia
© Németh Áron, BME © www.tankonyvtar.hu
Fonalas gombák izolálásánál és tenyésztésénél komoly gondot okozhatnak az atkák,
amelyek a gombával táplálkoznak, és keresztbe fertőzhetik a Petri-csészés kultúrákat.
Inkubálás: mezofilek: 25-30°C 7–14 nap, termofilek: 45-50°C 2–5 nap
A fonalas gombák különböző rendszertani egységei annyira különböznek egymástól, hogy
izolálásukban is eltérhetnek:
A) Ascomycetes, Deuteromycetes (Imperfekt): ubiquiterek (széles körű előfordulás), jól
sporulálnak, ezért könnyű az izolálásuk (aspergillusok, penicilliumok, fusariumok).
B) Basidomycetes:
makroszkopikusak: (kalapos, pöffeteg, tapló) termőtestből, mikorrhizából
történhet az izolálás
mikroszkopikusak: (rozsda és üszöggombák ) mikorrhizából, de nehéz
fenntartani, mert csak az élő gazdanövényen szaporodik.
Jól tűrik a benomylt (gombaellenes növényvédőszer),
ezért 0,5-5mg/l benomyl melletti izolálás esetén javarészt
ezek nőnek ki, a többi gomba visszaszorul.
C) Zygomycetes: ezek is ubiquiterek, és a legburjánzóbb növekedésűek. A Petri-csészén általában
az első kinövők (Mucor, Rhizopus, Trichoderma)
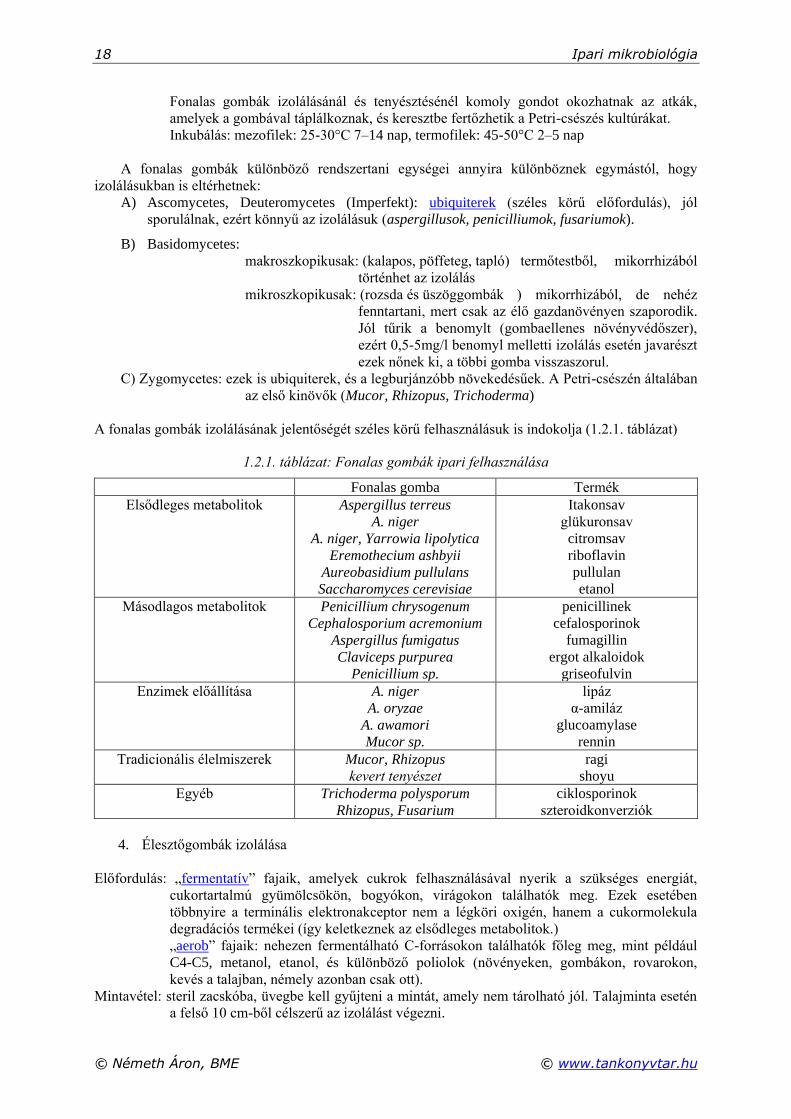
A fonalas gombák izolálásának jelentőségét széles körű felhasználásuk is indokolja (1.2.1. táblázat)
1.2.1. táblázat: Fonalas gombák ipari felhasználása
Fonalas gomba Termék
Elsődleges metabolitok Aspergillus terreus
A. niger
A. niger, Yarrowia lipolytica
Eremothecium ashbyii
Aureobasidium pullulans
Saccharomyces cerevisiae
Itakonsav
glükuronsav
citromsav
riboflavin
pullulan
etanol
Másodlagos metabolitok Penicillium chrysogenum
Cephalosporium acremonium
Aspergillus fumigatus
Claviceps purpurea
Penicillium sp.
penicillinek
cefalosporinok
fumagillin
ergot alkaloidok
griseofulvin
Enzimek előállítása A. niger
A. oryzae
A. awamori
Mucor sp.
lipáz
α-amiláz
glucoamylase
rennin
Tradicionális élelmiszerek Mucor, Rhizopus
kevert tenyészet
ragi
shoyu
Egyéb Trichoderma polysporum
Rhizopus, Fusarium
ciklosporinok
szteroidkonverziók
4. Élesztőgombák izolálása
Előfordulás: „fermentatív” fajaik, amelyek cukrok felhasználásával nyerik a szükséges energiát,
cukortartalmú gyümölcsökön, bogyókon, virágokon találhatók meg. Ezek esetében
többnyire a terminális elektronakceptor nem a légköri oxigén, hanem a cukormolekula
degradációs termékei (így keletkeznek az elsődleges metabolitok.)
„aerob” fajaik: nehezen fermentálható C-forrásokon találhatók főleg meg, mint például
C4-C5, metanol, etanol, és különböző poliolok (növényeken, gombákon, rovarokon,
kevés a talajban, némely azonban csak ott).
Mintavétel: steril zacskóba, üvegbe kell gyűjteni a mintát, amely nem tárolható jól. Talajminta esetén
a felső 10 cm-ből célszerű az izolálást végezni.

1. Alapismeretek, ipari mikrobiológiai módszerek 19
© Németh Áron, BME © www.tankonyvtar.hu
Előkészítés: dúsítás lehet szükséges: pH=3,5-4 rázatott lombikban (levegőztetve), így előszaporítva.
Tápközeg: Malátakivonat (maltextract), krumplifőzet (potatoe-dextrose-agar, PDA), de a pH 3,8 kell
legyen, azaz savasabb, mint a fonalas gombákénál. 50–200 mg/l kloramfenikol
hozzáadásával baktériummentes izolátum nyerhető. Általában ozmotoleránsok, azaz 30-
40% glükóz mellett is tenyészthetők
Inkubálás: mezofileket 20-25°C-on, a pszichrofileket 10-12°C-on szüksées tenyészteni.
A következő két csoport rendszertantól független, ezért egyaránt tartalmaz baktériumokat, gombákat
stb-t.
5. Alkalofilek izolálása
Az alkalofilek lúgos pH optimumú mikrobák, jelentőségük az antibiotikumok, mosószerenzimek
gyártásában van.
Előfordulás: alkalikus (szikes) talajok
Mintavétel: szilárd mintából/ról
Előkészítés: vizes szuszpenzió
Tápközeg: glükóz vagy keményítő, peptonnal élesztőkivonattal, sókkal kiegészítve, pH=8,5-12
Kitenyésztés: általában mezofilek, így 30-37°C-on 2-3 nap alatt érhető el.
6. Termofilek izolálása
A termofilek magas hőmérsékleti optimummal (55-75°C) rendelkező mikrobák.
A termofilhőmérséklet-optimum a mezofil és az extrém termofil között helyezkedik el, és
különbözik a termotoleráns fogalomtól: míg előbbieknél az optimális növekedési hőmérséklet alapján
történt a besorolás, addig a termotoleráns csak elviseli (esetleg növekszik is) a magasabb hőmér-
sékleten, de messze nem maximális sebességgel szaporodik és termel. A termofilek között bakté-
riumok, gombák és ősbaktériumok is megtalálhatók, mint ipari termelő vagy enzim-, illetve génforrás
mikroorganizmusok:
- fonalas gombák: mivel 55-60°C között számos gomba izolálható, de 62°C-on már egy sem,
úgy tűnik, hogy az eukarióták növekedésének 60°C körül korlátja van, amelynek oka
feltehetően, hogy nem tudnak termostabil jól funkcionáló belső membránokat létrehozni
[166]. Termofil fonalas gombára példa a Thermomyces lanuginosus (50°C optimummal
[167]), a Sporotrichum thermophile (45-50°C [168]), és a Rhizomucor miehei (35-45°C
[169]).
- baktériumok: thiobacillusok (acidophilek), bíborbakt: (fotoszint), cianobakt, actinomiceták.
- továbbá algák, protozoák, ősbaktériumok
Előfordulás: geotermikus hőforrások (gejzírek), spontán melegedő szubsztrátok (siló), fűtött környezet
(erőművi hűtővíz), talaj, édesvízi és tengeri üledékek (1.2.2. táblázat)
Mintavétel: gyakran obligát anaerobok (pl.: Thermotoga maritima), hiszen vagy az élőhelyük
levegőtől elzárt, vagy a magas hőmérsékleten nem oldódik be az oxigén a vizes közegbe,
ezért a mintavételhez Hungate-technika (ld. Anaerobok izolálása) javasolt.
Előkészítés: párologtató szükséges, hogy ne száradjon ki a tenyészet a magas hőmérsékleten
Tápközeg: komplex tápközeg ajánlott, amelynek összetétele az eredeti élőhelynek megfelelő
(mélytengeri, vulkanikus stb.). A szokásos szilárdító ágens (agar-agar) legfeljebb 65°C-ig
használható, efelett gellán gumi (100°C) vagy gelrite (+Ca2+ és Mg2+ sók) használata
javasolt a magasabb olvadáspont miatt.
Inkubálás: párologtató, esetleg anaerob kamra lehet szükséges.

20 Ipari mikrobiológia
© Németh Áron, BME © www.tankonyvtar.hu
1.2.2. táblázat: Termofil mikroorganizmusok lelőhelyei
Lelőhely faj
Geotermikus hőforrások
(savas-semleges-alkalikus)
Thermoproteus spp.
Thiobacillus spp.
Clostridium spp.
Thermoanaerobium spp.
Thermus spp.
Bacillus spp.
Thermotoga spp.
baktériumok
baktériumok
baktériumok
baktériumok
baktériumok
baktériumok
ősbaktériumok
Spontán melegedő szubsztrátok
(silók, komposztok)
Bacillus spp.
Thermoactinomyces spp.
Streptomyces spp.
Thermomyces spp
Talaromyces spp.
Thermoascus spp.
Clostridium spp.
Thermomonospora spp.
baktérium
baktérium
fonalas baktériumok
fonalas gomba
fonalas gomba
fonalas gomba
baktérium
baktérium
További fűtött környezetek
(erőművi hűtővíz, konzervek stb.)
Thermus spp.
Dactilarya spp.
baktérium
fonalas gomba
Nem fűtött szubsztrátok
(talaj, por, levegő, üledék)
Methanospirillium spp.
Methanogenium spp.
Thiobacillus spp.
Bacillus spp.
Lactobacillus spp.
Clostridium spp.
ősbaktériumok
baktériumok
baktériumok
baktériumok
baktériumok
baktériumok
Törzsfenntartás
A természetből izolált és/vagy mutációkkal nyert színtenyészetek fenntartása szintén kiemelkedő
fontosságú területe az ipari mikrobiológiának. Gondoljuk csak meg, hogy egy működő üzemnél, ahol
lehetőleg saját izolálású vagy fejlesztésű termelő mikrobát alkalmaznak, micsoda következménye
lenne a termelő törzs elvesztésének, tönkremenetelének. Ennek szem előtt tartásával tehát a
törzsfenntartás célja, hogy eredeti állapotában, azaz ugyanolyan szaporodó és termelőképességű
formában tartsa fenn a termelő törzset az üzem indításától a bezárásáig, kedvező esetben akár
évtizedekig (persze közben mindig kicsit újabb és újabb fejlesztésű sejtvonalat használnak, de a törzs
általában azonos marad).
Az izolátumok eltartására kétféle mód létezik: A) inaktív, és B) aktív formában.
Az aktív formában történő törzsfenntartást mutatja be az 1.2.3. videó.
1.2.3. videó: Törzsfenntartás aktív formában
Az inaktív forma jelenti a természetes szaporítóképletek (pl.: spórák) eltartását különböző
megoldások segítségével (folyadékszuszpenzióban, szilárd hordozóra adszorbeáltatva, micéliummal
együtt megszárítva stb.).
Az aktív forma jelenti a nagyobb kihívást, hiszen ilyenkor a vegetatív sejtek fenntartásáról
beszélünk, amelyek érzékenyek a környezeti hatásokra, és bár lassan, de szaporodnak, ami pedig a
genetikai változás (mutációk) veszélyét is magában hordozza. A vegetatív sejtek legjobban eltartható

1. Alapismeretek, ipari mikrobiológiai módszerek 21
© Németh Áron, BME © www.tankonyvtar.hu
aktív formája a liofilezést (fagyasztva szárítást) követően kapott sejtpor. Az eljárás lényege, hogy
kíméletes fagyasztást követően a tápközegből és a sejtekből egyre nagyobb vákuum alkalmazásával
elszublimáltatják a jeget, és végül csak a szárazanyag-tartalomnak megfelelő por marad vissza. A
kíméletes fagyasztás következtében azonban a sejtmembrán nem sérül, így steril folyadék (víz,
fiziológiás sóoldat, puffer vagy tápközeg) hozzáadásával a sejtek rehidratálhatóak, és ismét
tenyészhetőek. Ezt az eljárást hosszabb távú (évtizedes) tárolás esetén alkalmazzák, például a
törzsgyűjteményekben, így az onnan beszerzett mikrobák rendszerint ilyen formában kerülnek a
megrendelőhöz.
A mindennapi használat során azonban aktívan növő tenyészeteket használnak, amelyek
szaporodási sebessége a hűtőszekrény 6-10°C-os környezetében rendkívül lelassul, így általában kb. 1
hónapig eltarthatók túlöregedés nélkül. Ehhez Petri-csészébe öntött, megszilárdított (pl.: agaros)
tápközeget lehet alkalmazni, vagy helytakarékosság okán, illetve anaerob esetekben kémcsőben és
esetenként ferdén szilárdított tápközegen történik az eltartás. Az anaerob esetekben a tenyészet fölé
paraffinolaj rétegezése is bevett szokás, illetve gyakran a szokásos agarfelszínre oltás helyett oltótűvel
szúrt tenyészeteket hoznak létre (1.2.4. ábra).
1.2.4. ábra: Ferdeagaros, anaerob és szúrt tenyészetek, Petri-csészék és oltókacsok
Az ilyen formában tárolt tenyészetekről való átoltással egyrészt a termelő fermentációk
inokulumai kerülnek beoltásra, másrészt az öregedés ellen is a tárolt tenyészetet át kell időről időre
oltani. Az átoltást steril fülke alatt vörös izzásig leégetett steril oltókaccsal végzik autoklávban (vagy
egyéb módon) sterilezett tápközegre. Az átoltás után inkubálni kell a tenyészetet, amíg megjelennek a
telepek és/vagy a csíkozott tenyészetek, majd ezt követően kerülnek vissza a hűtőszekrénybe újabb 1
hónapos fenntartásra, vagy esetleg időközben fermentáció indításához való felhasználásra.
Törzs-screening
Az előbbiekben bemutatott mikrobaizolálási eljárásokkal különböző fajtájú, képességű, eredetű és
tulajdonságú mikrobák tucatjaira tehetünk szert létrehozva egy törzsgyűjteményt. Amikor azonban
ezek között keresünk egy adott tulajdonsággal rendelkezőt, akkor ezt a több tucat mikrobát kell
szűrővizsgálat alá vetni. Az ilyen vizsgálatok jellemzője, hogy adott tulajdonságot keres, lehetőleg
gyorsan, több száz minta analízisén keresztül. Az ipari mikrobiológiában a szűrővizsgálatot jobb
termelőképességű egyedek keresésére is használják, ilyenkor tehát nemcsak a kvalitatív szűrő-
vizsgálatot, hanem a kvantitatívot is el kell végezni, azaz nem elég kiválogatni a termelő- képes
mikrobákat, hanem a legjobb termelőképességűeket kell kiválasztani (a legnagyobb hozamút,
produktivitásút, végtiterűt.
A szűrővizsgálat tehát nagyfokú automatizáltságot kívánna meg, mivel nagyszámú mintát
lehetőleg gyorsan és – kvantitatív esetben – pontosan kell elemezni. Ugyanakkor minden feladat más
természetű, más indikációjú, tehát nem lehet rutinfeladatként végrehajtani. A szűrővizsgálatok első
lépcsőjében (általános vizsgálat nem optimált körülmények mellett) még minden potenciális jelöltet ki
kell szelektálni, a bizonytalanokat is, amelyek a későbbi lépcsőkön esetleg elvéreznek, de az is lehet,
hogy optimált körülmények között azok lesznek a legjobb termelők. Ezért szükséges az automatizált
szűrővizsgálatok manuális felügyelete, esetleg felülbírálata is.

22 Ipari mikrobiológia
© Németh Áron, BME © www.tankonyvtar.hu
A nagy hatékonyságú és kapacitású analitikával ellátott szűrőrendszereket HTS-nek (High
Throughput System) nevezzük.
A szűrővizsgálati módszer nagyban függ attól, hogy milyen céltulajdonságot keresünk (pl.: 1.
adott enzimtermelő mikrobát, 2. primermetabolit-termelőt, 3. valamely szekunder metabolitot termelő
mikrobát).
A klasszikus szűrővizsgálatok során enzimtermelő mikrobák keresésekor alapvetően az
enzimaktivitás-méréssel (assay) azonosítják a termelőket, illetve szelektív tápközeggel segítik ezek
izolálását is. A primermetabolit-termelőket klasszikusan szelektív tápközegen vizsgálják, és vala-
milyen nagy kapacitású és hatékonyságú analitikai módszerrel (GC, HPLC) követik nyomon a
metabolitok keletkezését. Ugyanez igaz a klasszikus szekunder metabolitot termelők screeningjére is,
azzal kiegészítve, hogy esetükben további analitikai vizsgálatokat is végeznek, mint például a
biológiai aktivitás és MIC- (minimal inhibitory concentration) meghatározás.
A modern eljárások a screening vizsgálatokat miniatürizálták, és egy kb. 10x16cm-es műanyag
lap 96 furatában párhuzamosan végezhetővé tették. Ehhez szükséges, hogy valamilyen indikációja
legyen a pozitív eredménynek (pl.: színreakció), amelyet egy plate-reader (lemezleolvasó)
fotométerrel és a hozzákapcsolt megfelelő szoftverrel lehet kiértékelni (1.2.5. ábra).
1.2.5. ábra: Microplate reader – HTS szűrővizsgálathoz
Klasszikus módszerek a szűrővizsgálatokhoz
A) Enzimtermelők szűrése
Proteázok termeléséhez a proteolitikus aktivitás indikációja szükséges, ezért a táptalajba
kicsapott fehérjét kevernek. A pozitiv eredményt a telep körül megjelenő tisztulási zóna adja,
mivel a kicsapott turbid fehérjét feloldja a proteáz termelő izolátum.

1. Alapismeretek, ipari mikrobiológiai módszerek 23
© Németh Áron, BME © www.tankonyvtar.hu
A keményítőbontó amiláztermelő mikrobák izolálása hasonló elven történik, de a kicsapott
fehérjék helyett vízben nem oldodó (=emiatt turbid) keményítőt alkalmaznak, és az amiláz-
termelő mikrobák ezt oldják fel szintén tisztulási zónát eredményezve a telep körül.
Az ipari szempontból érdekes további két enzim (celluláz és lipáz) esetében is ugyanezt az
elvet alkalmazzák, előbbi esetében cellulózrostok, utóbbi esetében olaj emulzió „feloldása”
eredményezi a tisztulási zónát a pozitív telepek körül.
A legnagyobb enzimgyártók egyike a Novo, amelyik az 1970-es években mosószerenzimek
előállítása céljából keresett termostabil alkalikus proteázokat. Ennek okán az alkalikus
talajmintákat kíméletesen megszárították, így a vegetatív sejteket elölték, csak a spórás
mikrobák maradtak izolálhatóak. Az ezután kinövő telepeket pH=9-10-re állított, kicsapott
haemoglobint tartalmazó agarra vitték, és 45-50°C-on inkubálták. A kinőtt telepek közül a
kioltási (tisztulási) zónával rendelkezőket tovább vizsgálták. Mivel a lombikos fermentációk
során kapott proteolitikus aktivitás nem korrelált a tisztulási zóna méretével, a kisebb zónájú
telepeket is érdemes volt tovább vizsgálni.
B) Primermetabolit-termelők keresése
Az elsődleges anyagcseretermékeket hatékonyan előállító mikrobák izolálásakor két módszert
használnak. Az egyik (ld. bioautográf módszer 1.1.3. animáció) szerint a keresett metabolitra
hiánymutáns mikrobát hoznak először létre random mutagenezissel, majd ezt szuszpendálják a
táptalajba. Az izolátumok ráoltása után inkubálják, majd az izolátumokat UV-val elölik, és
újabb inkubáció következik a hiánymutánsok tenyésztésére. Ezek csak ott tudnak kinőni, ahol
az izolátum a hiányzó keresett metabolitot előzőleg megtermelte, így itt nem tisztulási zónát,
hanem új telepet kerestek. Ennek a módszernek egyszerűbb formája, ha létezik olyan
természetes törzs, amely a keresett metabolitot tovább hasznosítani tudja, így nem kell
hiánymutánst létrehozni. Erre példa a tejsavtermelők keresésekor propionsav baktériumok
használata (ld. 2.1. C3 metabolizmus).
A másik módszer a szerves savak nem specifikus indikációjára alkalmas. Ebben az esetben
mészkőport (CaCO3) kevernek a táptalajba, amelyet a képződött szerves savak feloldanak
ismét tisztulási zónát eredményezve.
C) Szekundermetabolit-termelők keresése
A fermentációs ipar történetének legtöbb szekunder metabolitja az antibiotikumok közé
tartozik. Az ezek előállításhoz szükséges termelő mikrobák keresésekor a táptalajba a
szenzitív mikrobát szuszpendálták, amely ellen hatékony antibiotikumot kerestek. Az
antibiotikum termelő izolátumok körül kioltási zónát lehet tapasztalni.
Az antibiotikumok detekátlására agarkorongos módszert szoktak használni, amely módszert
szintén lehet screening célra adaptálni. Az agarkorongos módszer során egy steril
papírkorongot átitatnak az antibiotikummal, és olyan táptalajra helyezik, amelybe a szenzitív
mikrobát előre szuszpendálták, és ismét kioltási zónát vizsgálnak inkubáció után.
Modern HTS szűrővizsgálatok
A) Enzimtermelők szűrése megoldható kapcsolt enzimes reakcióval, ahol a kapcsolt reakció
szubsztrátja vagy koenzime (pl.: NAD+/NADH2 340nm-en mérhető) nyomon követhető
fotometriásan.
Hasonlóan optikailag követhető a kromogén vagy fluorogén szubsztát vagy szubsztrát
analógok használata a célenzimet termelő mikrobák esetében. Ilyenkor a fent bemutatott
készülékben (1.2.5. ábra) nagyszámú izoltáumot lehet nyomon követni speciális lemezeken
(plate), amelyek furataiba a kromogén/fluorogén szubsztrátot/szubsztrátanalógot előre
adszorbeáltatják, így a pozitív választ adó izoltáumokat a platereader (lemezolvasó) készülék
azonosítani tudja.
B) Esetenként kromogén/fluorogén szenzorokat alkalmaznak az előbbihez hasonló reakciók
kimutatására mikrotiterlemez-olvasó készülék helyett.

24 Ipari mikrobiológia
© Németh Áron, BME © www.tankonyvtar.hu
C) Szintén a modern módszerek közé tartozik a „microarray” technika. Ehhez fejlett
bioinformatikai adatbázisra van szükség, mivel a módszer elve, hogy a keresett tulajdonságot
kódoló gén klompenter DNS-ét kromogén/fluorogén jelzéssel ellátva adszorbeáltatják egy
mikrolemezre („DNS chip”), és az izolátumokkal együtt inkubálják. Ezt követően a nem
kötődött, jelzett DNS-t eltávolítják, és a kötött DNS-eket detektálják (pozitív izolátumok
esete).
D) Gyakran nincs lehetőség a keresett tulajdonság azonnali in-situ indikációjára az inkubációval
paralell, ilyenkor a HTS rendszerek csak a nagyszámú kis térfogatú minta tenyésztésére
használhatók, és a tenyésztés végén online vagy off line analitika (GC, HPLC, MS,
infraspektroszkópia, NMR stb.) szükséges.
Az alábbiakban néhány ipari példát mutatunk be termelő törzs szűrővizsgálatban történt szelekciójára.
1. Antibiotikum-termelők szűrésére bevált módszer, hogy a potenciális termelőtörzzsel egy
agarlemez kb. 1/3-ad nagyságú körszeletét oltják be, és az inkubáció során a biomassza-
képződés mellett az esetlegesen termelt antibiotikum az agar be nem oltott részei felé is
diffundál. Az első inkubációt követően tesztorganizmusokat oltanak a be nem oltott 2/3-adra,
és ismét inkubálják az agarlemezeket. A tesztorganizmus(ok) növekedéséből megállapítható,
hogy volt-e és milyen mértékű antibiotikum-termelés (több antibiotikum messzebbre
diffundál, rövidebb teszttelep 1.2.6. ábra).
1.2.6. ábra: Antibiotikum-termelők szűrővizsgálati tesztje
2. Egy japán gyógyszergyárban új enzimtermelő mikrobát kerestek szűrővizsgálattal. A cél a 7-
aminocephalosporánsav (7-ACA) oldallánc-módosításával félszintetikus cephalosporin-
származékok előállítása volt, amihez 7-ACA-deacetiláz enzimre volt szükség. A 7-ACADA
enzimtermelők kiválogatása érdekében a táptalajba 7-ACA-t adtak egyedüli szénforrásként,
így csak az ennek hasítására képes mikrobák szaporodhattak el. A vizsgált különböző fajok
közül a Rhodotorula glutinis élesztő került ki „győztesen” a feladatra.
3. Enziminhibítorok keresésére sikerrel alkalmazták a mikrotiter módszert, amelynek feltétele
valamilyen kis léptékben is alkalmazható színreakció. A β-laktamáz inhibítorok keresésekor
az enzimaktivitás-csökkenést követték nyomon valamilyen színreakcióval (pl.: penicillin
esetében a β-laktamáz enzimaktivitásának eredményeként keletkező penicilloinsav sztöhio-
metrikusan oxidálható jóddal, így a keményítő-jód kék komplexe elszíntelenedik, β-laktamáz
inhibítor jelenlétében kék marad).

1. Alapismeretek, ipari mikrobiológiai módszerek 25
© Németh Áron, BME © www.tankonyvtar.hu
4. A gyógyszergyártás során gyakran szükséges a gyógyítás eléréséhez valamely receptor
blokkolása vagy egy anyag receptorhoz kötésének megakadályozása kompetitiv receptor
antagonistával. Az erre alkalamas metabolitok keresésekor általában a receptort kipreparálják
és mikrotiterlemezre immbolizálják színreakcióval kombinálva.
5. Új biotranszformációkhoz szükséges mikrobák szűrővizsgálatára példa a múltból és a jelenből
is [6] a compactin-pravastatin átalakítás esete. A compactin egy penicilliumok fermentá-
ciójával előállítható vegyület, amelyből szelektív hidroxilezéssel pravastatint lehet előállítani,
amely a máj koleszterinszintézisét szelektíven gátolja, ezzel elősegítve a kardiovaszkuláris
betegségek gyógyítását. A szűrővizsgálat során különböző szénforrások mellett 500 mg/L
compactint adtak az agaros tápközeghez, és több mint 100 compactin rezisztens mikroba közül
sikerült két jó hidrexilező aktivitásút kiválasztani (Pseudonocardia autotrphica, Streptomyces
griseolus).
6. A szűrővizsgálatokat egyre gyakrabban alkalmazzák olyan metabolitok keresésére, amelyek az
immunrendszerre hatnak (pl.: immunszupresszánsok). Ilyenkor a screening során a fehér-
vérsejtekkel (limfocitákkal) való reakciót vizsgálják.
7. Az agrár-biotechnológia területén rovarkárosító metabolitokra végeznek gyakran szűrővizs-
gálatokat. Ilyenkor a screening rendszer a célrovart vagy annak hernyóját is alkalmazza.
Mikrobiológiai szűrővizsgálatokat alkalmaznak továbbá új gyomirtószerek, haszonnövény-
növekedést serkentő (auxinhatású) metabolitok keresésére is tesztnövény screeningbe iktatásával.
Gyakran van szükség egy ismert hatóanyag jogvédett előállításának alternatívájára is, ilyenkor
szintén szűrővizsgálattal keresnek másik termelő mikroorganizmust.
Végezetül meg kell említeni, hogy a törzsfejlesztés (ld. Törzsfejlesztés) során is alkalmaznak
szűrővizsgálatokat az új mikrobák között a kedvezőbbek kiszűrésére.
Identifikáció
A mikrobiológiai azonosítás (identifikáció) célja az új izolátumok rendszertani meghatározása,
besorolása. Ehhez számos tulajdonságot kell megvizsgálni, amelyekkel az adott faj egyértelműen
meghatározható.
A rendszertan (taxonómia) valamilyen hasonlóságaik alapján rendszerezi az élőlényeket.
A(z ipari) mikrobiológia hőskorában a rendszertani besoroláshoz a külső megfigyelések és (később) a
biokémiai tulajdonságok alapján osztályozták a mikroorganizmusokat. Napjainkban a rendszertan
átalakulás alatt van, mert a modern géntechnikák lehetővé tették a molekuláris (gén) szintű rokonságok
feltérképezését, és ez gyakran eltér a külsőleg megfigyeltektől.
Meg kell jegyezni, hogy az ipari mikrobiológia területén gyakran szükséges ugyan az új
izolátumok azonosítása, ám sok esetben ezeket a költséges és munkaigényes vizsgálatokat nem, vagy
csak részben végzik el iparjogvédelmi célzattal, hogy ne lehessen könnyen lemásolni a technológiát.
A klasszikus rendszertani besoroláshoz a következő vizsgálati szempontok szerint írják le az
izolátumokat:
makroszkopikusan: telep morfológiája, színe, szaga, mérete
mikroszkopikusan: A) vizuálisan: sejt mérete, alakja, mozgásszerve, sejtmagja
B) mikrobiológiai festésekkel: Gram-festés (sejtfal), spórafestés, csillószínezés,
tokfestés stb.
biokémiai reakciók: oxidázpróba (aerotolerancia), továbbá indol-, ureáz-, kénhidrogén-, arginin-,
xilóz-, ammoncitrátpróbák, aerob/anaerob dextróz fogyasztása.
A rendszertannal bővebben foglalkozik e jegyzet a 3. fejezetben.

26 Ipari mikrobiológia
© Németh Áron, BME © www.tankonyvtar.hu
Törzsfejlesztés
A fermentációs iparban kevés természetből izolált „eredeti” mikrobát használnak. Mivel egy
fermentáción alapuló technológia költségét alapvetően befolyásolja a termelő mikrobatörzs
hatékonysága (hozam, produktivitás, végtiter), ezért különösen a régebbi technológiák az eredeti
termelő törzset mára már jelentősen továbbfejlesztették. Noha a felsorolt 3 kulcsparaméter (hozam,
produktivitás végtiter) jelentősen javítható technológiaoptimálással (pH, oldott oxigén, hőmérséklet,
keverés, lépték stb.), de csak bizonyos korlátokig, ám jobb termelő törzzsel optimálás után még sokkal
jobb hatékonyság érhető el.
Mivel a fenotípusos tulajdonságokat a genom (genotípus) határozza meg, ezért ezt kell megvál-
toztatni a fejlesztés érdekében.
Klasszikus törzsfejlesztés
1.2.7. videó: Klasszikus törzsfejlesztés
A klasszikus technika véletlenszerű mutációk segítségével változtatta meg a termelő mikroba
genomját, majd a megváltozott képességű utódok közül szelektálta a jobban teljesítőket (=random
mutagenezis+screening). Ilyen UV-val generált random mutagenezist mutat be az 1.2.7. videó. A
véletlenszerű mutációk generálása, majd a kedvezőbb klónok szelekciója azt eredményezi, hogy a
kívánt cél eléréséhez igen nagyszámú mutánst kell létrehozni, és így igen sok mutáns utód közül kell
kiszűrni a jobban teljesítőket. Ez azt is jelenti, hogy noha a termelési körülmények eltérnek a
laboratóriumitól, mégis kizárólag kis léptékben lehet gazdaságosan törzsfejlesztést véghezvinni. A
laborléptékben jól teljesítő mutánsokat azután fokozatos léptéknöveléssel (pilot plant =”kísérleti (fél)
üzemi” léptéken keresztül) lehet a termelési szintre juttatni.
Általában a törzsfejlesztés olyan ciklikus folyamat, ahol a fent leírt random mutagenezis+screening
kombinációt folyamatosan ismétlik.
Az egyetlen lehetőség a véletlen mutációk „irányítására” a szelekciós nyomás alkalmazása, azaz
olyan körülmények alkalmazása, amely már a célnak megfelelő mikrobák kitenyésztését segíti, a nem
megfelelőkét hátráltatja.
A modern géntechnológiák lehetővé tették, hogy a korábbi véletlenszerű (random) mutációk
helyett célzott genetikai változást lehessen eszközölni, és így a vizsgálandó mutánsok száma is
radikálisan lecsökkent, ami a törzsfejlesztés (költség)hatékonyságát nagyban megnövelte. A célzott
genetikai beavatkozásokhoz ismerni kell a genom és a funkció kapcsolatát, az anyagcsereutakat és
azok szabályozását (gén azaz expresszió szintű szabályozás, vagy fehérje szintű szabályozás
(aktivátor/inhibítor), előre/visszacsatolások)
Noha hatékonysága kisebb, mégis napjainkban is gyakran használt módszer a klasszikus
mutagenezis+screening kombinációja a törzsfejlesztés céljából. A random (véletlenszerű) mutációk
előidézésére kémiai vagy fizikai mutagéneket alkalmaznak. Míg fizikai behatásként UV besugárzást
szokás alkalmazni, a leggyakoribb kémiai ágens a nitrozoguanidin (ami a DNS-t változtatja meg),
illetve bázisanalógok, amelyek hibás DNS-replikációt eredményeznek, valamint a frame shift
mutagének (általában akridin festékek, amelyek eltolják egy vagy néhány bázispárral a leolvasási
kódot).
Bár a random mutagenezis+screening rendkívül egyszerű (és olcsó, ezek miatt használatos még
ma is), komoly kihívást jelent a megfelelő mutagén dózis megtalálása. Túl nagy dózis esetén annyi
mutáció jut egy sejtre, hogy az elpusztul, és a mutációk után nem marad túlélő. Túl alacsony dózis
pedig nagyon kisszámú mutációt eredményez, amelyek között kisebb valószínűséggel található

1. Alapismeretek, ipari mikrobiológiai módszerek 27
© Németh Áron, BME © www.tankonyvtar.hu
kedvező fenotípusú egyed. Az UV sugárzásnál a besugárzási idő és a fény intenzitása a két
optimálható paraméter, amelyek függenek a gazdaszervezettől. A kémiai mutagéneknél az egyik
paraméter a kontaktidő, a másik a mutagén koncentrációja.
Mivel ezek fajtól függetlenül a DNS-t változtatják meg, általában rákkeltőek!
A random mutációt követő szűrővizsgálat is kétféle elven történhet: A) a random módszer minden
izolátumot megvizsgál termelőképességre, míg a B) racionális módszer valamely biokémiai
tulajdonsághoz köti a termelőképességet, és ezt a biokémiai tulajdonságot monitorozza (pl.:
antimetabolit rezisztens törzsek sokkal több metabolitot termelnek, ha túlélnek).
Modern törzsfejlesztés – anyagcsere-mérnökség (metabolic engineering) [21]
A modern törzsfejlesztés során a genetikai eszközök kihasználásával a klasszikus véletlenszerű
(random) genomváltozásokat lehet kiváltani célzott változtatásokra. Ezáltal jelentősen megnő az
esélye a kívánt tulajdonságú mikroba elérésének (nem kell sok véletlen mutációra várni), és jelentősen
lecsökken a fölöslegesen vizsgált (a nem megfelelően mutálódott) minták száma. Ezzel együtt is a
modern törzsfejlesztés a klasszikushoz hasonlóan gyakran cirkuláris (ismétlődő) műveletsorból áll
(1.2.8. ábra)
1.2.8. ábra: A modern törzsfejlesztés vázlata
A fejlesztés a meglévő törzs metabolikus fluxus analízisével kezdődik, majd ennek módosítását
kell megtervezni a célnak megfelelően (melléktermékképző útvonalak kikapcsolása, új útvonal
génjeinek bevitele, kofaktoregyensúly megtartása stb.), majd a szintézis során a terveknek megfelelő
mikrobát lehet létrehozni. Ezt követően ismét analízisre van szükség, és esetleg ennek során további
fejlesztés (tervezés+szintézis) lehetősége merül föl, illetve kiderülhetnek olyan, korábban ismeretlen
szabályozási mechanizmusok, amelyek a megváltoztatott genom következtében megzavarják,
megváltoztatják a mikroba valódi metabolizmusát a tervezettől eltérően.
Azokban az esetekben, amikor heterológ fehérjeexpresszióra van szükség a termelő mikroba
kialakításához (azaz pl.: új terméket kell előállítani egy adott gazdaszervezettel, vagy ki kell
terjeszteni a szubsztrát spektrumát), a metabolic engineering folyamat metabolikus útvonaltervezéssel
kezdődik. Ez tartalmazhat empirikus terveket vagy kivitelezhető in silico analíziseket, azaz a sejtek
részletes metabolizmus modelljét. Ezután az „útvonaltervezés” után a tervezett törzs elkészítése, majd
további optimalizálás következik a metabolic engineering ciklus szerint.
Míg a molekuláris biológia gyors fejlődésével a szintézislépés sok esetben viszonylag triviálissá
vált (még ha az ipari törzsek fejlesztése az alacsony transzformációs hatékonyság miatt fáradtságos is),
addig a gyors és hatékony iparszerű környezetben végezhető fenotípus-meghatározás eszközei az
analízis lépéshez hiányosak. A teljes genomszekvenciák rendelkezésre állása új HighThroughput
System (HTS) analitikai technikákat eredményezett a transzkriptom, proteom, metabolom számára.
Ezek a technikák azonban további finomításra szorulnak, hogy a szabályozókat azonosítani lehessen,
amelyek kulcsszerepet játszanak a másodlagos anyagcsere útvonalak fluxusainak szabályozásában.

28 Ipari mikrobiológia
© Németh Áron, BME © www.tankonyvtar.hu
Mivel ezen a területen gyakran esik szó egy szervezet valamilyen típusú molekuláinak
összességéről, ezért ezekre a készletekre a gén–genom analógiájára „-om” végződéssel (angolban
„omics”) bevezettek további fogalmakat. A gén–genom felől indulva az RNS-készlet következik
transzkriptom elnevezéssel, majd a proteom (fehérjekészlet), illetve az ezek regulátorainak összessége,
az interaktom (kölcsönhatók). Bevezették továbbá a metabolitok összességére a metabolom és a
fluxusok összességére a fluxom elnevezéseket is.
A biológia központi alapelve („central dogma of biology”), hogy az információ a DNS-en
tárolódik, onnan íródik át az RNS-re, amiről pedig a fehérjék szintetizálódnak. Kiderült azonban, hogy
számos olyan szabályozási funkció létezik, amelyek miatt egy-egy génhez nem lehet egyértelműen
funkciót rendelni. Ezért létre jött egy új kutatási terület a funkcionális genomika (functional genomics),
amelynek feladata, hogy egy adott gén által kódolt fehérje funkcióit feltérképezze. Ehhez azonban
szükséges volt a sejten belüli folyamatok analitikájának fejlődésére.
Az anyagcsere-mérnökség legfőbb célja, hogy genetikai manipulációkkal fejlessze a termelő törzs
metabolikus fenotípusát. A kapott fenotípusok ennek ellenére gyakran szuboptimálisak és nem
kielégítőek a genetikai módosítások közvetett hatásai vagy ismeretlen szabályozó kölcsönhatások
miatt. Ezért erősen ajánlott a teljes metabolikus szabályozást figyelembe venni az anyagcsere-
mérnökség során.
A metabolizmus szabályozása vagy enzim, vagy gén szinten fordul elő. Utóbbi globális
szabályozók és/vagy szigmafaktorok hatása alatt áll. A génszabályozások változnak az idővel a
tenyésztési feltételektől vagy a sejt állapotától függően. A klasszikus metabolikus mérnöki
megközelítések egyik hátránya, hogy ezt nem veszik figyelembe, vagy kevés figyelmet tulajdonítanak
ennek.
Bőséges információ áll rendelkezésre a lokális genetikai szabályozásról és a sejtmetabolizmus
biokémiájáról, illetve fiziológiájáról, de meglepően keveset tudunk a teljes anyagcsere átfogó
szabályozásáról. Sőt, noha a HTS technikákban benne van a lehetőség, hogy a transzkriptomok,
proteomok, metabolomok és fluxonomok közötti mehanizmust feltárja, többségük csak pillanatképet
ad egy állapotról, és az a módszer, ami a különböző szintű információkat egységbe foglalná még
ismeretlen. Így még mindig messze vagyunk a szabályozási elvek sejtszintű megértésétől. Bár már
sikerült megalkotni az Escherichia coli teljes sejtmodelljét, még fontosabbá vált az in vivo vizsgálatok
analízise és a posztgenomikai kutatásokhoz a sejtek egész rendszerének együttes kezelése. Ezen cél
eléréséhez fontos az anyagcsere átfogó szabályozásának vizsgálata a különböző szintek (genom,
transzkriptom, proteom stb.) információinak integrált figyelembe vételével
A legfontosabb információ a komplex anyagcsere-szabályozás mechanizmusának megértésében a
centrális anyagcsere fluxusainak eloszlása lehet, mivel ez a gén- és fehérjeexpresszió (vagy
enzimaktivitás) és az intracelluláris metabolitok kifejeződése. A metabolikus fluxus analízise a kulcs-
metabolitok anyagmegmaradási elvén alapszik. A sejten belüli fluxusokat a mért fluxusokból
anyagmérlegek segítségével határozzák meg. A mérhető külső fluxusok száma korlátozott, és a
sztöhiometriai korlátok miatt gyakran alulhatározott algebrai rendszert kapunk. Ezért kofaktor
anyagmérlegeket is néha be kell vonni a sztöhiometriai modellbe, vagy egy optimalizálandó objektív
függvényt kell bevezetni. A centrális anyagcsere anabolikus és katabolikus funkciókat is ellát (felépítő
és lebontó is), hiszen kofaktorokat és építőköveket szolgáltat a makromolekulák szintéziséhez
(anabolizmus), csakúgy, mint az energiatermeléshez (katabolizmus). Az optimálást a katabolizmus
vagy az anabolizmus, vagy mindkettő szempontjából el lehet végezni a sztöhiometriai korlátok
mellett. A fluxusmérleg-analízist széleskörűen használják a steady state (állandósult állapot)
metabolikus fluxusainak becslésére a sejtnövekedés maximalizálásának érdekében. Az E. coli szinte
optimálisan hasznosítja a szénforrást, ami ennek a törzsnek a különleges előnye. Ennek ellenére a
fluxusszámítások pontossága a kofaktorbecslések megbízhatóságától függ, és a megfelelő objektív
függvényválasztástól. Ismeretlen reakciók jelenléte, amely kofaktort generál vagy fogyaszt,
érvénytelenítheti azt a megközelítést, hogy a kofaktor koncentrációja egyensúlyban marad, és a
kiválasztott objektív függvény nem lesz megfelelő, vagy a megbízhatósága csak bizonyos sejt-
állapotokra fog korlátozódni.
Egy alternatív megközelítés az izotóp-nyomkövetés, ami mérhető mennyiségeket határoz meg az
intracelluláris fluxuseloszláshoz kapcsolódóan. Izotóppal jelzett szubsztrátot lehet adni a sejtnek, és a

1. Alapismeretek, ipari mikrobiológiai módszerek 29
© Németh Áron, BME © www.tankonyvtar.hu
jelölt szénatom el fog oszlani az anyagcsere-hálózaton. A végső izotópdúsulás a sejten belüli
metabolitkészletben mérhetővé válik. A biomassza-hidrolizátumokban az aminosavak sokkal nagyobb
feleslegben vannak jelen, mint prekurzoraik a centrális metabolizmusban. Így könnyebb következtetni
a proteinogén aminosavak jelölt mintázatából az intracelluláris metabolitok jelölési mintájára a
prekurzor–aminosav kapcsolat alapján. A nyomjelző kísérletek általában NMR spektroszkópiát
használnak vagy GC-MS technikát. Az NMR-t a C13
helyi dúsulásának meghatározására egydimenziós
H1 NMR-rel végzik, vagy az izotopomerek mennyiségi meghatározására egydimenziós C13 NMR-t,
vagy kétdimenziós C13 H1 COSY NMR-t alkalmaznak. Bár a C13 NMR egy hatékony és vonzó
technika a metabolikusfluxus-analízis számára, nagy mennyiségű mintát igényel, és a módszer nem
képes detektálni a metabolitokat 10-4M-os koncentráció alatt. Másrészt a GC-MS módszer könnyen
analizál 10-7M-os koncentrációban is. Mindezek miatt a GC-MS megfelelő lehet az alacsony
biomassza-koncentrációjú tenyészetek fluxusanalíziséhez is. Pillanatnyilag ezek a nyomkövető
technikák a közvetlen extracelluláris fluxusmérésekkel kombinálva a legmegfelelőbb módszerek az
intracelluláris metabolikus fluxuseloszlások (metabolic flux distribution=MFD) megállapítására,
csupán néhány modellezési feltételezést használva. Emellett a fluxusarány-analízis is használható az
elágazások fluxusainak megállapítására.
Bár alkalmazásaikban a TCA ciklusra korlátozódnak, néhány további analitikai megoldást a
jelölési kísérletekhez számba kell még venni. A becsült fluxusok konfidencia-intervallumát statisztikai
analízissel lehet megadni. A jelölési kísérletek tervezésének optimalizálására is fejlesztettek már ki
módszereket, és így azonosíthatósági analíziseket is bevezettek. Bár a C13 fluxusanalízist széles körben
alkalmazzák az olyan mikrobákra, mint E. coli, Corynebacterium glutamicum, Saccharomyces
cerevisiae, Bacillus subtilis, más élőlényekre is kiterjesztették, mint például cianobaktériumok,
növényi sejtek és emlős sejtek (köztük egér-, agyi és idegsejtek, valamint rákos sejtek).
Az proteogén aminosavak izotopomer eloszlásainak használata esetén egy jelentős korlát, hogy
csak steady-state esetben alkalmazható. Annak érdekében, hogy az analízist az iparilag érdekes
szakaszos tenyésztésekre vagy dinamikus analízisre lehessen kiterjeszteni, az intracelluláris
metabolitokat kell először meghatározni. Ezek mérhetőek CE-TOF-MS, GC/MS-MALDI/TOF, MS,
LC-MS/MS, vagy LC-MS technikákkal2. A fluxus dinamikáját GC-MS rendszerrel egymagában is
lehet mérni 1,3-propándiolgyártás során. A nem stacionárius fluxusok analíziséhez gyakran kinetikai
modelleket is felhasználnak. Mivel az elsődleges metabolitkészlet nagyságrenddekkel kisebb, mint a
proteogén aminosavakból felépített molekulakészlet, az izotópeloszlás hamarabb éri el az állandósult
állapotot. A fluxusváltozás becsülhető a készletméretekkel felírt differenciálegyenletekkel.
Mind a számításos fluxusmérleg-analízis (fluxus balance analysis – FBA), amely mátrix műve-
letek segítségével képes kezelni a több tucat metabolikus reakcióegyenletet, mind pedig a fluxusok
meghatározására bevezetett kísérletes MFD módszerek hasznosak a sejtfiziológia megértésében. FBA
egyszerre tud kezelni több száz metabolikus útvonalat, míg a kísérletes megközelítés hasznos a főbb
metabolikus útvonalak elsődleges meghatározásában. A fluxuseredményeknek összhangban kell
lennie vagy integrálva kell lennie más információkkal, mint például az anyagcserefluxusok irányával,
az enzimek metabolitok általi allosztérikus szabályozásával és transzkripciós szabályozásokkal stb.,
mivel a metabolikus háló funkcionális viselkedése a génexpresszió, fehérjeexpresszió és az
intracelluláris metabolitok koncentrációjának eredménye. A jelentős méréstechnikai fejlődésnek
köszönhetően minden szintű „omics”-nak a globális monitorozása lehetséges. Annak érdekében, hogy
a metabolikus szabályozás tisztázódhasson, nagyon fontos a metabolikus fluxusok és a többi „omics”
integrálása. Ez a systems biology-nak keresztelt, viszonylag új tudományterület feladata. Meg kell
jegyezni, hogy a génexpresszió, fehérjeexpresszió, sejten belüli metabolitkoncentráció közvetlenül
mérhető, míg a metabolikus fluxusok vagy reakciósebességek nem, ezért becsülni kell őket.
A számos megvalósult példa közül az Escherichia coli-n végzett szisztematikus anyagcsere-
mérnökséget mutatjuk be. Az E. coli teljes levegőztetés mellett is termel jelentős mennyiségű ecet-
savat, amely 10-30%-a is lehet a teljes szénfluxusnak. Ennek oka az, hogy nem megfelelően kontrol-
lált a glükóz felvétele, és a túl sok intracelluláris glükózlebontási terméket (azaz piruvátot) a citrátkör
nem tudja elég hatékonyan feldolgozni, ezért vagy előbb acetil-CoA, majd abból ecetsav képződik
(Pta+Ack, azaz PhosphateAcetylTransferase+Acetil-kináz), vagy a piruvátból közvetlenül (pox azaz
2 Balla József: Analitikai kémia, Budapest, BME

30 Ipari mikrobiológia
© Németh Áron, BME © www.tankonyvtar.hu
piruvát-oxidáz) is ecetsavat állít a sejt elő, előbbi esetében ATP is keletkezik. A glükóz elfogyása után
az ecetsavat visszaalakítja a sejt ATP segítségével acetil-CoA-vá, és a citrátkörön és a terminális
oxidáción keresztül energiatermelésre fordítja. Mivel az ecetsavképződés jelentős szubsztráthányadot
elhasznál, illetve citotoxikus metabolitként a sejtnövekedést gátolja, a kiküszöbölése alapvető
fontosságú az E. coli-val végzett rekombináns (heterológ) fehérjeexpresszió és -előállítás során.
Az egyik lehetőség, hogy csökkent aktivitású glükóztranszferrel rendelkező mutánsokat kell
létrehozni a piruvát akkumlációjának elkerülésére, vagy lehet a glükózkoncentrációt rátáplálásos
technika segítségével alacsony értéken tartani, illetve lehet a foszfotranszferáz rendszer (PTS) glükóz-
specifikus enzimének expresszióját csökkenteni egy represszor molekula expresszáltatásával. Ezeken
felül lehetőség van a citrátkör és a glioxilát ciklus intenzifikálására is. Ez utóbbi enzimeit az aceBAK
gén kódolja az izocitrát-dehidrogenázzal együtt. Ez a gén ecetsavon vagy zsírsavakon történő
növekedéskor aktív, glükózon, glicerinen és piruváton represszálva van. A regulon transzkripciós
faktorai közül a fadR mutációjával elérhető, hogy az utóbbi szénforrásokon is aktív legyen az aceBAK
gén. Így tehát a citotoxikus és szubsztrátveszteséget okozó ecetsav képződése kiküszöbölhető az E.
coli-nál (1.2.9. ábra)
1.2.9. ábra: E. coli ecetsav képzésének eliminációja: A) glükóz transzfer deficiens mutáns,
B) aceBAK génekkel megnövelt glioxalát- és citrátkör- aktivitás.

1. Alapismeretek, ipari mikrobiológiai módszerek 31
© Németh Áron, BME © www.tankonyvtar.hu
A tejsav fermentatív előállításának régóta ismert problémája, hogy a jól termelő lactobacillus-
törzsek igen összetett tápanyagforrásokat (pl.: élesztőkivonat) igényelnek, amelyek viszonylag
költségesek, és feleslegük drágítja, illetve nehezíti a downstream folyamatokat.
Ezért E. coli-t is módosítottak úgy, hogy elfogadható tejsavtermelővé váljon. Ehhez a törzs
glükóz- specifikus PTS fehérjéjének, illetve a piruvát-formiát-liáz enzimének, valamint a saját laktát-
dehidrogenázának génjét inaktiválták, és egy lactobacillus eredetű L-LDH (laktát-dehidrogenázt)
expresszáltattak vele. Így az E. coli alkalmassá vált több mint 70 g/L koncentrációban tejsavat
termelni 77%-os hozammal ecetsav, etanol, formiát és D-tejsav nélkül.
1.3. Mikroorganizmusok ipari alkalmazása
A mikroorganizmusok ipari alkalmazásakor a leghatékonyabb termelés érdekében különböző igényeik
(tápkomponens, levegőztetés, pH stb.) figyelembe vétele és kielégítése a biotechnológus feladata.
Részletesen ezekkel a feladatokkal a Biomérnöki műveletek3 foglalkozik, de néhány mikrobiológiai
aspektust e jegyzet keretein belül is célszerűnek véltünk tárgyalni.
Az oxigén szerepe
A mikrobák centrális (központi) anyagcseréje a következő sémával szemléltethető (1.3.1. ábra)
1.3.1. ábra: A mikrobák centrális anyagcseréje
A központi anyagcsereút során a szénforrás lebontása (pl.: cukrok) a glikolízissel kezdődik és a
piroszőlősavig tart. Az ebből képződő acetilcsoport lép be a citrátkörbe, ahol az oxálecetsavval
összekapcsolódik, és a képződött citromsav két lépésben dekarboxileződik, és CO2 kerül kibocsátásra
(ld. 1.1.4. ábra). A citrátkör során a szén-dioxid-távozás mellett redukált koenzimek is keletkeznek
(NADH2), amelyek a terminális oxidációban a légköri oxigén segítségével a respirációnak nevezett
folyamatban regenerálódnak jelentős ATP energia keletkezése közben. Ha a mikroba légköri oxigént
használ, akkor gyakran találkozik peroxigyökökkel, amelyek igen reaktívak, és így kártékonyak a
sejtekre. Ezek közömbösítésére az aerotoleráns és/vagy aerob mikrobák kataláz enzimeket termelnek
(gyakran peroxiszómában), amelyek a káros peroxidgyököket ártalmatlanítják.
A fermentatív anyagcsere esetében azonban nincs lehetőség a légköri oxigén felhasználására a
redukált koenzimek regenerálása céljából így energianyerésre sem, ezért különböző szerves vegyü-
letek (metabolitok) segítségével történik meg az oxidáció és a kisebb mértékű, de nélkülözhetetlen
3 Lásd Sevella Béla: Biomérnöki műveletek és folyamatok. Budapest, Typotex Kiadó, 2011
G
L
I
K
O
L
Í
Z
I
S
Terminális
oxidáció

32 Ipari mikrobiológia
© Németh Áron, BME © www.tankonyvtar.hu
energiatermelés. Ezek a különböző erjedési folyamatok, amelyek megkülönböztetendők az anerob
légzéstől (amikor is az elektrondonor szerves molekula, pl. cukor), de az elektronakceptor sem nem a
légköri oxigén ((légzés)), sem szerves anyag (fermentáció), hanem oxigéntől eltérő szervetlen
molekula).
Oxigénigény szempontjából a mikrobák lehetnek aerobok (életfunkcióhoz szükséges a légköri
oxigén), aerotoleránsok (nincs szükségük oxigénre, de el tudják viselni ((általában kataláz pozitívak)),
fakultatív anaerobok (ha van oxigén, használják, ha nincs, akkor is élnek és szaporodnak), obligát
anaerobok (nincs szükségük oxigénre, és nem tudják elviselni sem (toxikus rájuk nézve).
A jegyzetben tárgyalt anyagcsere utakat foglalja össze a 2. fejezetben részletesen tárgyalt 2.1.2.
Anyagcsere utak összefoglalása című hyperpicture.
Aerobok tenyésztése
Az oxigén egy különleges szubsztrát, mert gázhalmazállapotban kell a rendszerbe juttatni, és a redox-
egyensúlyon keresztül az oxigén mennyisége befolyásolhatja a mikroba anyagcseréjét. Egyes fermen-
tációk a termék-előállításhoz oxigénfelesleget igényelnek (ekkor maximális a termékhozam), mások
pedig különleges oxigénprofilt, esetleg oxigénlimitet. Mindezek miatt az aerob fermentációknál vagy
felületi tenyésztést kell alkalmazni (szilárd fázisú fermentáció, nem igazán lehet az oxigénszintet
szabályozni), vagy a fermentlébe beoldódott oxigén mennyiségét mérni és gyakran szabályozni kell,
amihez a legcélszerűbb az oxigénelektród használata (pl.: Clark-féle elektród4). Az elektród szolgál-
tatta jel alapján több helyen lehet beavatkozni: 1) keverési sebesség növelésével csökken a buborék-
méret és nő az anyagátadási felület, tehát javul az oxigénellátottság; 2) a levegő térfogatáramának
növelésével szintén több oxigén juttatható a fermentlébe; 3) nagyobb nyomású levegő alkalmazásával
is több oxigén vihető be; 4)végső esetben oxigénnel dúsított levegőt is lehet használni. Noha a
hőmérséklettől jelentősen függ a telítési oxigénkoncentráció (amitől pedig az oxigénbeoldódás
hajtóereje függ), az alkalmazott hőmérsékletet általában a mikroba hőmérsékleti optimuma határozza
meg. A magas hőmérsékleten végzett fermentációknál tehát komoly gondot jelent a megfelelő
levegőztetés biztosítása, mert 60-70°C-on már rosszul oldódik az oxigén (pl.: Thermus thermophylus)
Az ipari fermentációknál (pl.: citromsav- vagy fehérjetermelés E. coli-val) gyakran jelent
problémát, hogy a nagy léptékű reaktorok alja és teteje között jelentős hidrosztatikai nyomáskülönbség
van, ami miatt az oxigénbeoldódás is különbözik, tehát inhomogénné válhat a reaktor. Ennek
kiküszöbölésére több helyen vezetnek be levegőt a fermentorba. A fejtérbe vezetett levegővel a habzás
mértéke is visszaszorítható.
Anaerob fermentációk (erjesztések)
A fejezet elején már részletesen bemutattuk az oxigént hasznosító és nem hasznosító mikro-
organizmusokat. Az oxigént nem igénylő mikrobákat (obligát anaerobok, aerotoleránsok, fakultatív
anaerobok) tenyésztési szempontból most egyszerre tárgyaljuk.
Mindegyik fajta tenyésztésekor mentesülünk a levegőztetés okozta problémáktól (oxigénlimit,
magas keverő fordulatszám és nyíróerő, habzás stb.). A szigorúan anaerobok esetében azonban inert
gáz bevezetésére és alapos eloszlatására (diszpergálására) van szükség, pont, mint az oxigén esetében,
ezért ilyenkor gyakran ugyanazt a gázbevezető rendszert használják, mint a levegőztetéskor.
A klasszikus fermentációs ipar a primer metabolitok termelésével alakult ki. Ezek nagyobb,
iparilag is hasznosítható mennyiségben a különböző erjesztések során keletkeznek, amikor a cukor
részben savakká és alkoholokká oxidálódik, részben pedig az eközben keletkezett redukált koenzim
(NADH2) regenerációjakor redukált szerves vegyületekké alakul (pl. piroszőlősavból tejsav képződik
stb.). Ezeket az erjesztéseket tekintjük át az alábbiakban.
1. Etanolos erjedés
A hexózok erjesztésekor élesztők (Saccharomyces cerevisiae) és egyes baktériumok
(Zymomonas mobilis) metabolizmusa során 1:1 arányban etilalkohol és szén-dioxid keletkezik
a piroszőlősavon (glikolízis) keresztül. Az első lépés a hexokináz katalizálta szubsztrát
4 Lásd Sevella Béla: Biomérnöki műveletek és folyamatok. Budapest, Typotex Kiadó, 2011

1. Alapismeretek, ipari mikrobiológiai módszerek 33
© Németh Áron, BME © www.tankonyvtar.hu
foszforilezés, majd a foszfo-frukto kináz által végzett második foszforilezés eredményeként a
6 szénatomos szubsztrát 2 db 3 szénatomos glicerin-aldehid foszfátra bomlik, amelyek a
glikolízisben foszfoenolpiruvát-piroszőlsav-acetaldehid-etanol útvonalon alakulnak át a
termékké.
A szubsztrát felvétele történhet A) támogatott diffúzióval vagy B) aktív transzporttal. Előbbire
azért van szükség, mert noha a sejten kívüli térben található magasabb koncentrációban a
szubsztrát így a spontán diffúzió is a felvételt segíti, de viszonylag nagy molekulájúak a
monoszacharidok, amelyek ezért csak lassan tudnak a sejtmembránon átjutni, ezért hordozó
molekulák segíthetnek. A termékleadás egyszerű diffúzióval történik, így a magas
termékkoncentráció gátolhatja a saját képződését.
A diszacharidok különböző utakon jutnak el a lebontó apparátushoz: a szacharóz esetében az
invertáz enzim a periplazmás térben hasítja két monoszacharidra a szubsztrátot, amelyek
onnan a fenti mechanizmus szerint hasznosulnak; a laktóz először transzportálódik, majd
intracelluláris β-galaktozidáz hidrolizálja; maltóz esetében ugyanez a helyzet, de α-glükozidáz
végzi az első intracelluláris hasítást
Napjaink kutatásainak fókuszában állnak azok a mikroorganizmusok (pl.: Pichia stipitis),
amelyek pentózokból képesek etanol előállításra, mivel a 3. generációs bioetanol-előállítás
hemicellulóz-tartalmú növényi hulladékok felhasználását tűzte ki célul (a keményítő- és
cellulózalapú technológia után).
Az etanolos erjedés melléktermékei lehetnek az acetaldehid (néha akkumlálódik, de etanollá
alakulhat), illetve a glicerin-aldehid-3-foszfát akkumlációjával a glicerin jelenik meg
melléktermékként.
Tapasztalatok szerint az optimális erjesztéshez némi oxigén is szükséges (a biomassza-
növekedéshez, mert teljesen anaerob körülmények mellett erre képtelenek az élesztők). Az
alkoholos erjedésnél két szabályozási mechanizmust kell figyelembe venni: Pasteur-effektus –
az oxigénlimitben műkődő tenyészet anaerob metabolizmusra vált (lassabb szaporodás, ami
oxigént igényel és etanoltermelés); Crabtee-effektus: magas cukorkoncentráció esetén a
biomassza-képződés helyett az aerob fermentáció anaerob anyagcserére helyezi a hangsúlyt,
mert korlátozott a terminális oxidációs kapacitása a mikrobának.
Az etanoltermelés során így szubsztrát- és termékinhibíció is fellép, utóbbi kb. 112-114 g/L
titernél, ami épp a gazdaságosan kinyerhető termékkoncentráció határán mozog.
A termék etanolfelhasználása széles körben ismert, mint például fertőtlenítőszerek, tisztító-
szerek, italok, üzemanyagok, szintézisek stb.
2. Tejsavas erjedés
A tejsav termelődése igen gyakori az élővilágban a mikrobáktól kezdve az emberig bezárólag.
A mikrobák esetében a legjelentősebb tejsavtermelők a tejsavbaktériumok (lactobacillusok,
lactococcusok, pediococcusok, streptococcusok, leuconostoc fajok stb. ), bacillusok, egyes
gombák (pl.: Rhizopus) és élesztők (pl.: Kluyweromyces lactis). A tejsavas erjedés tipikus
példája a terminális oxidáció pótlásának: oxigén hiányában a redukált koenzim (NADH2)
visszaoxidálásához a centrális metabolizmus molekuláját a piroszőlősavat használják a
molekulát a laktát-dehidrogenáz enzim segítségével, így a koenzimből NAD a piroszőlősavból
tejsav keletkezik. A lactobacillusok emellett a saját biomasszájuk felépítésére szerves
nitrogénforrásokat igényelnek, amelyek nitrogéntartalma mellett a széntartalom is
tápanyagként szolgál, az emellett jelen lévő szénhidrátok bontásával pedig a piroszőlősavon
keresztül történő tejsav-előállítás során energiát termelnek, így bizonyos esetekben a cukor
szinte kizárólag tejsavvá alakul (magas ~95-99%-os hozammal).
Míg a szubsztrát általában aktív transzportal jut be (pl.: glükóz, laktóz – permeáz segít-
ségével), a termelt tejsav disszociálatlan formában szabadon ki/be jut, ám ez toxikus a sejtekre
nézve. A laktátion a protonokkal együtt szimport formában transzportálódik.

34 Ipari mikrobiológia
© Németh Áron, BME © www.tankonyvtar.hu
A bejutott szubsztrát metabolizmusa szerint a tejsavtermelőket két csoportra szokás osztani:
homofermentatívak és heterofermentatívak. Előbbiek esetében a hexózok az Embden–
Meyerhof–Parnas (glikolízis) útvonalon keresztül 1:2 mólarányban tejsavat képeznek, míg a
heterofermentatívak előbb dekarboxilezik a hexózt (a szén-dioxid távozását buborékolás jelzi),
majd a képződött pentózból tejsavat (3 szénatom) és valamely 2 szénatomos mellékterméket
képezik, általában ecetsavat vagy alkoholt. Nyilván a fehér biotechnológia számára a
homofermentatívak a kedvezőek a magas hozammal és konverzióval, de a heterofermentítvak
is iparilag jelentősek: az élelmiszeriparban használatosak, ahol a melléktermékek aránya adja
a kívánt aromát, ízt.
A tejsavtermelők általában fakultatív anaerobok, ami ipari szempontból szintén kedvező,
hiszen nincs szükség költséges levegőztetésre, sem inert gázra, a spontán beoldódó oxigén
viszont nem zavarja a mikrobákat, ugyanakkor gázbevezetés híján habzással sem kell
számolni, ettől a reaktor is jobban kihasználható.
A termék tejsav széles körű felhasználással rendelkezik az élelmiszeriparban, gyógyszer- és
higiéniás termékekben, polimer,- bőriparban stb.
Egy speciális eset a Rhizopus oryzae, amely oxigén jelenlétében a cukrokat szén-dioxiddá
oxidálja, és biomasszát képez, anoxikus (kevés oxigén) körülmények között tejsavat termel,
ám teljesen anaerob körülmények között etanol a főtermék. Noha a termékprofil nagyban függ
a morfológiájától, a fenti három eset általános e fonalas gombánál. A tejsavtermelés
szempontjából a pelletes morfológia a legkedvezőbb, amelynek kialakulása viszont igen sok
tényezőtől függ 1.3.2. ábra [7]. Anyagátadási szempontok miatt az optimális pelletek sima
felületűek és 1-2 mm átmérőjűek. Egy ilyan pellet felszínét mutatja be az 1.3.3. video, illetve
különböző morfológiák láthatók az 1.3.4-1.3.7 ábrákon.
Pellet képződés
Törzs
pHSpóra konc.
Spóra koraTápközeg
KeverésPolimerek
Felület aktív
anyagokSzilárd
részecskék
1.3.2. ábra: A pelletképződést befolyásoló tényezők
1.3.3. Pelletes morfológia

1. Alapismeretek, ipari mikrobiológiai módszerek 35
© Németh Áron, BME © www.tankonyvtar.hu
1.3.4. ábra: Bolyhos pellet (1-2mm)
1.3.5. ábra: Óriás (1cm) pellet
és szabálytalan alakú pellet
1.3.6. ábra: Amorf és óriás (2-3 cm) pellet
1.3.7. ábra: Pelletfelszín mikroszkópos képe
(400-szoros nagyítás)
3. Homoacetogén erjedés
Az ecetsavról mint melléktermékről már több esetben említést tettünk, létezik azonban olyan
erjedési folyamat, amely a fermentlébe az ecetsavat bocsátja ki fő termékként, és emellett
szén-dioxidgáz képződik és távozik a rendszerből. Ilyenkor egy hexózból 2 ecetsav és 2 CO2
keletkezik, melléktermékként etanol fordulhat elő. A hexózok a glikolízisen át, a pentózok
pedig a pentóz foszfát cikluson kersztül bomlanak le piroszőlősavvá, amelyből képződik
közvetlenül vagy acetil-foszfáton keresztül az ecetsav. A szubsztrátfelvételre és a
termékleadásra a tejsavhoz hasonlóan aktív transzport és disszociálatlan forma jellemző,
amely utóbbi ez esetben is citotoxikus hatású.
A termelő mikrobák gyakran termofilek (pl.: Clostridium thermoaceticum, Acetogenium kiwi),
a képződött ecetsavat pedig jellemzően a vegyipar használja fel (az élelmiszeriparban ezek a
mikrobák nemkívánatosak).
4. Propionsavas erjedés
A propionsav jellemzően heterofermentációs folyamatok terméke, ahol a főkomponens, a pro-
pionsav és mellette az ecetsav, a borostyánkősav és a tejsav is megjelenhet. A termelő anyag-
csere során a hexózok a glikolízisen keresztül eljutnak a piroszőlősavig, majd részben tejsavvá
és ecetsavvá, részben oxálacetáttá alakulnak, amely utóbbi a fumársav-borostyánkősav-
propionilCoA-án keresztül propionsavat képez. Jellemzően mezofil, anaerob mikrobák végzik
ezt a fajta fermentációt, mint például Propionibacterium acidipropionici, vagy P. shermanii.
A propionsav-baktériumok tejsavból is képesek propionsavat erjeszteni.
A megtermelt propionsavat takarmány- és élelmiszer-ipari tartósításra, polimerek és
peszticidek gyártására, valamint antifungális szerként is használják.

36 Ipari mikrobiológia
© Németh Áron, BME © www.tankonyvtar.hu
5. Butanolos erjedés
Butanol fermentációs úton kétféle anyagcserével is keletkezhet: A) hexózok bontásából a
glikolízisben piroszőlősav keletkezik, amelynek acetilcsoportját a koenzim-A (CoA) a
citrátkörbe szokta vinni, ám két acetil-CoA-ból aceton keletkezhet, illetve háromból butiril-
CoA, majd abból butanol, és emellett közvetlenül a piroszőlősavból etanol (=ABE), valamint
a citrátkörben szén-dioxid; B) izopropil-alkohol képződik egy dehidrogenáz-enzim segítsé-
gével az acetonból a butanol és az etanol mellett (=IBE)
A technológia nehézsége, hogy a butanol erősebb sejtméreg még az etanolnál is, ezért csak
igen kis koncentrációban halmozzák fel a termelő mikroorganizmusok (0,5-1,0% ~10 g/L).
Emiatt desztillációs, extrakciós és kihajtásos (strippelés) technológiák szükségesek a feldol-
gozási műveletek során, amelyek viszonylag magas fajlagos költséget eredményeznek.
A legáltalánosabb butanoltermelő mikroba az anaerob mezofil Clostridium acetobutylicum.
Ipari butanolos fermentációk az I. világháború utáni német hadiipar számára termeltek
butanolt, a későbbiekben a kőolajalapú technológiák alacsonyabb költségei miatt a butanol-
fermentációs üzemeket bezárták. Napjainkban azonban a „bioetanol” analógiájára szintén
energiahordozóként éli reneszánszát „biobutanol” elnevezéssel – immár a genetikai ismeretek
kiaknázásával – a butanol fermentációs előállítása.
6. Diolok, triolok, poliolok erjesztéses előállításának áttekintése
A) etilénglikol: toxikussága ellenére a borhamisítás kulcsvegyülete, szintetikusan olcsóbb az
előállítása
B) 1,3-propándiol: (bővebben: 1.1.fejezet 1,3-propándiol előállítás) az enterobakterek,
klosztridiumok, egyes tejsavbaktériumok és egyes citrobakterek fajai képesek anaerob
körülmények között a glicerinből (a belső redox egyensúly megőrzésének érdekében) 1,3-
propándiolt előállítani, miközben a redukált NADH2 koenzimet visszaoxidálják. Az 1,3-
propándiol minden esetben heterofermentatív anyagcserével keletkezik, azonban a
melléktermékek a termelő mikrobára jellemzőek (pl.: 2,3-butándiol a Klebsiellákra, vajsav
a Clostridiumokra stb.). Glükóz kofermentációja gyakran segíti az 1,3 propándiol
képződését (mert több NADH2 keletkezik a glükóz lebontásakor, amit a propándiol útnak
kell regenerálni.)
C) 2,3-butándiol: az enterobakterek jellegzetes terméke, amely 2 piroszőlősavból keletkezik.
Gumigyártásra és metil-etil-keton (MEK) előállítására használják, amely utóbbival az
üzemanyagok oktánszámnövelése érhető el.
D) Glicerin (triol): élesztők glükózból termelhetik akár főtermékként is. A bor erjedésekor is
keletkezik pl.: Saccharomycesek, Pichiák közreműködésével. A glicerin felel a bor tes-
tességért, éppen ezért a bor glicerines feljavítása borhamisításnak minősül. A glicerin
régen a szappanfőzés mellékterméke volt, napjainkban egyes biodízeltechnológiák
(metanollal történő növényolaj átészterezése estén) melléktermékeként keletkezik nagy
mennyiségben némileg szennyezett (metanol, katalizátor, lúg stb.) formában.
E) Xillit: xilán hidrolízisével xilóz nyerhető, amelyből pl. Pichiák segítségével xillit állítható
elő, és édesítőszerként forgalmazható.
F) Szorbit(ol): élesztők (pl.: Zymomonas mobilis) képesek glükózból az etanol mellett
szorbitot (az „ol” végződés az angolból ered, magyarul enélkül is használatos, ez esetben
„szorbit”) előállítani.
Fentiekben tehát különböző anaerob fermentációt, erjesztést soroltunk fel. A levegőztetés
problematikája a szubmerz (folyadék fázisú, szuszpenziós) fermentációk óta áll fent, és egy másik
klasszikus fermentációs technikával – a felületi vagy szilárd fázisú fermentációval – részben
kiküszöbölhető.

1. Alapismeretek, ipari mikrobiológiai módszerek 37
© Németh Áron, BME © www.tankonyvtar.hu
Szubmerz kontra felületi fermentáció
Noha a biomérnöki fermentációs tanulmányok során megszoktuk, hogy a mikrobatenyészet az egy
adott faj vizes szuszpenziója, és ezzel a természetben is találkozhatunk (aludttej, joghurt stb.), egy
adott szubsztrát felszínén történő tenyésztés szintén mindennapos (pl.: penészes kenyér), és ugyanezen
folyamat kiaknázható klasszikus technikaként egyes régiókban (főleg Ázsiában).
A szilárd fázisú fermentáció (Solid State Fermentation – SSF; amely csak rövidítésében azonos az
egyidejű cukrosítás és fermentációval –Simultan Saccharification and Fermentation!) során a
biomasszaszaporítás és termékképzés olyan nedves szubsztráton vagy inert hordozón történik, ahol
csak kötött víz található (=vízaktivitás). A természetben ilyen folyamat a komposztálás, silózás vagy a
talajszemcsék ökoszisztémája.
A felületi tenyésztés előnye a szubmerz technikával szemben, hogy egyszerűbb eszközök és
tápközegek használatosak, kevesebb szennyvizet termel, és néha a feldolgozási-tisztítási műveletek
egyszerűbbek (könnyebb a biomassza-eltávolítás különösen). Ugyanakkor hátránya, hogy nem
minden mikrobával végezhető (pl. baktériumok jelentős része), meglehetősen nehezen kontrollálható,
inhomogén rendszer, amitől lassabb a metabolizmus (nincs mindenütt optimális körülmény), és a
„reakció” ez esetben is a kevés kötött vizes fázisban zajlik. Ezért főleg tradicionális ázsiai (pl.: japán)
ételek előállításakor használják.
Szubsztrát/hordozó oldalról a következő 3 szempontot kell figyelembe venni:
1) Anyaga: lehet mezőgazdasági hulladék (hidrolízis nélkül is hasznosíthatók, szemben a szub-
merz fermentációval), szervetlen, nagy szemcsés őrölt kőzet, szintetikus műanyag (poli-
uretán).
2) A szubsztrát/hordozó karakterisztikáját megadja a porozitás, vízmegkötő képesség, mecha-
nikai tűrés.
3) Szubsztrát/hordozó előkezelést igényel-e, úgymint fizikai előkezelések (aprítás, sterilezés
hővel vagy besugárzással), vagy kémiai módszerek (savas/lúgos hidrolízis, extrakció), vagy e
kettő kombinációja.
Bioreaktor oldalról a következő megoldások használatosak:
1) Forgódobos bioreaktor citromsav-előállításhoz Aspergillus niger fonalas gombával (1.3.8.
ábra) [8]
1.3.8. ábra: Forgódobos bioreaktor

38 Ipari mikrobiológia
© Németh Áron, BME © www.tankonyvtar.hu
2) Tálcás reaktor (Koii, 1.3.9. ábra) [9]
1.3.9. ábra: Koji tálcás reaktor
A fenti reaktor előnye, hogy a reaktorban a páratartalmat mérni és szabályozni lehet, a
tenyészeteket tartó tálcásmodulok pedig alulról csővezeték segítségével fűthetőek/hűthetőek.
3) Töltött ágyas bioreaktor
A töltött ágyas reaktoroknál egy oszlopszerű reaktorba durva apríték (darabos) szubsztrátot helyeznek
el, amelyet bepermeteznek a termelő mikrobával, és alulról enyhén levegőztetik.
4) Fluidágyas reaktor
Nagyon hasonló a töltött ágyas reaktorhoz, de ebben az esetben könnyű a hordozó/szubsztrát, és
magasabb a levegő térfogatárama, így a szilárd szubsztrát lebeg. Az intenzívebb levegőztetés miatt
könnyebben kiszárad a tenyészet.
5) Toronyreaktor
Az elve hasonlít egy rektifikáló kolonnára: fentről engedik be a szubsztrátot, amely halad lefelé, és
eközben érintkezik az alulról bevezetett (szembeáramú) levegővel.
6) Alagúttípusú reaktor
Lassú futószalag felett növényházszerűen fóliából készült „reaktor” komposztálási célra.

1. Alapismeretek, ipari mikrobiológiai módszerek 39
© Németh Áron, BME © www.tankonyvtar.hu
A fenti reaktorokat mind azzal a céllal fejlesztették ki, hogy a lehető legegyenletesebben szolgálják ki
a tenyészet igényeit, és ugyanezen célból legyen beavatkozási lehetőség, azaz bizonyos paramétereket
szabályozni lehessen. Szabályozni lenne érdemes a páratartalmat, oxigénkoncentrációt, pH-t,
hőmérsékletet.
Az alábbiakban néhány példát sorolunk fel ipari szilárdfázisú fermentációra.
A) Élelmiszerek előállítása: Penicillium camember, Aspergillus sojae
B) Enzimek előállítása: α-amilázok, proteázok Aspergillus oryzae-vel, cellulázok, pektinázok
Aspergilus niger.
C) Növényvédőszerek: bioinszekticid – Beauveria bassiana (ld. később), biopeszticid –
Fusariumok- ellen antagonista gomba (Trichoderma harzianum) termelése SSF technikával.
D) Csiperke (Agaricus bisporus) és laska (Pleurotus ostreatus) gombák termesztése.
E) Komposztálás kevert tenyészettel, szabadban. Fontos a levegő arány és a páratartalom.
Önmelegedő folyamat. Alkalmas technika a szennyvíziszapok (veszélyes hulladék)
kezelésére.

40 Ipari mikrobiológia
© Németh Áron, BME © www.tankonyvtar.hu
2. AZ IPARI MIKROBIOLÓGIA BIOKÉMIAI ÉS
FIZIOLÓGIAI HÁTTERE
2.1. Szénforrások hasznosulása, különleges metabolizmusok
A mikroorganizmusok anyagcsere-folyamatai rendkívül szerteágazóak ezért célszerű valamilyen
rendszerben tárgyalni őket. A legegyszerűbb megoldás a szénforrások szénatomszáma szerinti
sorbarendezés.
C1-források hasznosulása [19]
Az 1 db szénatomot tartalmazó szubsztrátok száma igen csekély, ám a mikrobák szerepe itt sem
elhanyagolható. Ide sorolhatók a metán (CH4), metil-halogenidek (CH3X, X: Cl, Br, stb.), metanol
(CH3OH), metil-amin (CH3-NH2), hangyasav (HCOOH), metilszulfid ((CH3)2S)) és a szén-dioxid
(CO2). A szén-dioxid-hasznosító mikrobákat metanogéneknek, a többi C1 szubsztrátot hasznosítót
metilotrófnak szokás nevezni. Előbbiek definíció szerint szén-szén kötés nélküli szubsztrátot
használnak, többnyire oxigén segítségével (tehát aerobok), a metanogének jellemzője pedig a
metántermelés.
A felsorolt szubsztrátok hasznosítására képes mikrobákat a természetes szubsztrátok
környezetéből lehet izolálni: metanolt növények felszínéről (metanol=faszesz), metilamin a halak és
növények bomlásakor keletkezik, metilhalogenidet és metilszulfidot pedig a tengeri algák termelnek
nagyobb mennyiségben.
A C1 szubsztrátok hasznosítása olyan előnyökkel jár, mint a csekély verseny (ipari szempontból
ez könnyebb containmentet jelent, azaz csökken a fertőzésveszély), és olcsó kőolajalapú nyersanyagok
jöhetnek számításba (ez mindig relatív).
Metilotrófok a környezetvédelemben
A földi élet szempontjából fontos tevékenység, hogy a metilotrófok az üvegházhatást okozó anaerob
lebontások végtermékeként keletkező metánt (21x több hőt tart vissza, mint a szén-dioxid) szén-
dioxiddá oxidálják.
Másik fontos metilotróf tulajdonság, hogy számos olyan környezetre káros anyagot képesek
átalakítani kevésbé toxikus vagy ártalmatlan vegyületté, amelyek nem igazi szubsztrátjai a mikrobának
(nem kerülnek be a sejtbe és haladnak végig valamelyik anyagcsereúton), így a metilotrófok
talajremediációban is hasznosíthatók szennyezők, illetve hulladékok lebontására.
Metilotrófok a biokatalízisben
Kissé szokatlannak tűnhet a metanol (és társai) mint szubsztrát (ok), ám a XX. Század közepén az
olajárak kedvező szintjén komoly próbálkozások és beruházások történtek, például a brit ICI
vállalatnál kőolaj eredetű metanolon történő SCP (single cell protein) gyártásra. Akkor a metilotróf
mikroba összetételét találták táplálkozástanilag megfelelőnek (emberi és) állati fogyasztára.
A metanolhasznosítás és a „Metanolalapú társadalom” ma reneszánszát éli, és ennek jelentős
magyar vonatkozása is van.
Az 1994-ben kémiai Nobel-díjjal kitüntetett Oláh György kémikus 2005-ben jelentette meg „A
kőolaj és földgáz után: a metanolgazdaság” című értekezését [10].
Ebben 4 érvet sorol fel a metanol mint a gazdaság központi és meghatározó szereplője mellett: 1)
a fosszilis készletek (kőolaj, földgáz, szén) fogytán vannak, 2) a légköri CO2-ből és CH4-ből (üvegház-
hatást okozó gázok) előállítható, 3) egyszerre lehet energia- és nyersanyagforrás, és 4) könnyen
tárolható.
A Nobeldíjat egy különleges reakciómehanizmus felfedezéséért kapta Oláh György, amely
reakcióban „carbocation” keletkezik intermedierként. Ugyanez a köztitermék fordul elő a metánból
történő bázikuskatalizátor melletti oxidáció, illetve szén-dioxidból induló redukció során. Az már

2. Az ipari mikrobiológia biokémiai és fiziológiai háttere 41
© Németh Áron, BME © www.tankonyvtar.hu
ismert volt, hogy metán oxidációjával hangyasav vagy formaldehid keletkezik, de az új reakció-
mechanizmussal e két termék reakcióját is lehetett magyarázni, amely reakció viszont metanolt termel
(2.1.1. ábra).
2.1.1. ábra: Metanol-előállítás szén-dioxidból és metánból
Ezek szerint a metanol a természetben is előfordul, és viszonylag környezet- illetve költségkímélő
módon szintetikusan is előállítható, tehát egy állandóan rendelkezésre álló, olcsó, tiszta szubsztrát. Ez
alapján a jövő biotechnoógiájának egyik kihívása lehet, hogy a metilotrófokat kiaknázhatóvá tegye,
azaz a metabolizmus fő fluxusait a főtermék felé lehessen átirányítani, és ezt a fluxust a
költséghatékonysághoz szükséges szintre lehessen emelni, illetve a melléktermékképzést (tárolás során
képződő metabolitok vagy bomlástermékek, szekretált fehérjék, védekezőrendszerek ((enzimek,
antibiotikumok)) lehessen csökkenteni.
Mindehhez azonban a metilotrófok anyagcseréjét kell jól ismerni.
A metilotrófok a metanolból és a metilaminból is a toxikus intermediert, a formaldehidet képezik,
amelyből szén-dioxidot és energiát termelnek, utóbbit redukált koenzimek (NADH2) formájában,
illetve a formaldehid vagy ribulóz 5-foszfáthoz kapcsolódik és a RuMP ciklusba kerül (ehhez
kapcsolódnak a bioszintetikus utak), vagy szerint képez, és a szerin-cikluson keresztül kapcsolódik a
többi asszimilációs úthoz. A metanogének ugyanezt az elvet követik, de első lépésként
metánmonooxigenáz-enzimmel (pMMO) a metánból metanolt képeznek, ahonnan már azonosak az
anyagcsereútjaik a metilotrófokéval (2.1.2. hyperpicture)
2.1.2. hyperpicture: Az ipari mikrobiológiában fontos anyagcsereutak összefoglalása
C2-metabolizmus
Az egyedüli, cukrokhoz kissé hasonló kétszénatomos molekula a glikolaldehid. A legegyszerűbb
cukorszerű molekula, ezért „dióznak” is lehet nevezni. A pterin szintézisben játszik szerepet (purin-
bázis, GTP-ből), és ezen keresztül a fólátok előállításában, amelyek bioszintézisek kofaktorai. A
szénhidrátok szintézisének első lépcsője, hogy a formóz reakcióban két formaldehidből glikolaldehid
keletkezik, majd újabb formaldehid beépülésével glicerinaldehid, amiből izomerizációval dihidroxi-
aceton, és ez a glicerinaldehiddel ribulózt, izomerizáció után ribózt eredményez (2.1.3. ábra).

42 Ipari mikrobiológia
© Németh Áron, BME © www.tankonyvtar.hu
2.1.3. ábra: Cukorképződés glikoladldehiden keresztül [11]
Ezzel együtt azon tulajdonsága miatt is kutatják, hogy 2 formaldehidből keletkezik egy
glikolaldehid, és ezt a reakciót (a saját képződését) a glikolaldehid katalizálja, tehát ez az első önrepli-
káló mehanizmus.
A spontán ribózképzés és az önreplikációs képesség miatt az élet nyomainak világűrbeli keresése
során az űrkutatás fókuszában áll, továbbá szerin, triptofán, gyógyszer és agrokemikáliák szintézisé-
ben is használatos. Reaktív volta miatt azonban természetben nemigen fordul elő, így klasszikus
szénforrásként nem áll a mikrobák rendelkezésére.
Előállítható szintetikusan és enzimesen etilén-glikolból (pl.: Pichia pastoris alkohol-oxidáz 1
segítségével, vagy Aspergillus japonicus glicerinoxidáz enzim segítségével) is etilén-glikolból.
A kétszénatomos vegyületek közül az etanolt is képesek a mikrobák hasznosítani és ecetsavvá
oxidálni. Míg a már említett crabtree effektus esetén a terminális oxidáció enzimjeinek túlterheltsége
miatt NADH2-felesleg keletkezik – aminek segítségével az alkohol-dehidrogenáz-enzim (ADH)
redukálja az acetaldehidet etanollá –, addig a katabolitrepresszió elmúlásával (azaz a cukor elfogyása
után) az etanolt NADH2 (~energia)- termelés mellett ecetsavvá oxidálja a mikrobák egy része szintén
ADH segítségével.
C3-metabolizmus
A három szénatomos cukrok (triózok) az első olyan molekulák, ahol a cukormolekulának aldehid
(glicerin-aldehid) és keto formája (1,3-dihidroxiaceton) is létezik. Ezek mellett maga a glicerin is (ami
ketocsoport nélküli triol) népszerű három szénatomos szénforrrás. A propionsav-baktériumok pedig a
tejsavat is képesek három szénatomos szubsztrátként hasznosítani.
A glicerinmetabolizmus útvonalait az 1.1. fejezetben már bemutattuk (1.1.8. ábra)
A tejsav mikrobiális előállítását az 1.3. fejezetben már tárgyaltuk.
A tejsav felhasználását a propionsav-baktériumok úgy végzik, hogy első lépésben piroszőlősavvá
oxidálják (NAD+ független laktát-dehidrogenáz (LDH) segítségével), majd az bekerül a citrát-körbe
(oxálecetsavval), és onnan kerül ki propionil-CoA formájában. Általában 3 tejsav molekulából 2
propionsav és 1 ecetsav keletkezik 1 szén-dioxid távozása mellett.

2. Az ipari mikrobiológia biokémiai és fiziológiai háttere 43
© Németh Áron, BME © www.tankonyvtar.hu
A propionsav-baktériumok tejsavfogyasztó tulajdonságát a tejsavtermelő mikrobák keresésekor is
kihasználják a screening vizsgálatoknál a következőképpen: a tápagaron kinövesztik az izolátumokat,
UV-val elölik azokat, s föléjük újabb agarréteget öntenek, amelyben nincs szénforrás, és amelyet
Propionibacterium shermanii-val beoltanak. Ez egyéb metabolizálható szénforrás híján csak azon
izolátumok fölött fog kinőni, amelyek termeltek tejsavat.
C4-metabolizmus
Négy szénatomos cukor- (szerű) molekulák a glicerin (glycerol) analógiájára az eritrittel (másnéven
eritritollal) kezdődnek, amelynek mind a 4 szénatomján 1-1 OH-csoport található (2.1.4. ábra). Jelen-
tősége, hogy belőle Gluconobacter sejtek fermentációjával eritrulózt lehet előállítani a kozmetikai ipar
számára, mert bőrbarnító szerként (a dihidroxiacetonra érzékenyek részére) forgalmazzák. Az eritrulóz
(ketoforma) aldóz formáját eritróznak nevezzük.
2.1.4. ábra: Eritrittől az eritrózig
A négy szénatomos cukrok közé tartozik a treóz is, amely az eritróz izomerje. A két középső OH-
csoport nem transz, hanem cisz helyzetű a treóz esetében.
Az eritróz metabolizmusról keveset lehet tudni (leszámítva, hogy Bertrand már 1900(!)-ban leírta
a Gluconobacter suboxydans tenyészet eritrit-ertiróz átalakítóképességét), de nagy valószínűséggel
foszforilálódik, majd pedig vagy észterképződik belőle, és szedoheptulóz formájában hasznosul [12],
vagy a pentózfoszfát-ciklusba kerül, és energiatermelésre fordítódik [13].
C5-metabolizmus
2.1.5. ábra: Pentóz-foszfát ciklus [14]

44 Ipari mikrobiológia
© Németh Áron, BME © www.tankonyvtar.hu
Az öt szénatomos cukrok javarészt a pentóz-foszfát ciklusban (PFC, angolul: PPP-Penthose phosphate
pathway) hasznosulnak (2.1.5. ábra). Ennek funkciója egyrészt, hogy redukált koenzimet (NADH2)
termeljen, amelyből a terminális oxidáció energiát raktároz ATP formájában. Ezen felül a ciklusban
számos alapvegyület található, amelyek előállítása is a PFC feladata (ribulóz 5-foszfát, szedoheptulóz-
7-foszfát, Fruktóz-6- foszfát, glicerin-aldehid-3-foszfát stb.). Amint az ábráról is látszik, a ciklus egy
oxidatív, és egy nem oxidatív szakaszra bontható. Előbbi feladata a dekarboxilezés, utóbbié a cukrok
felépítése, illetve a lebontó anyagcsere számára a glicerin-aldehid-3-foszfát előállítása.
C6-metabolizmus
A cukorhasznosítás központi útvonala a glikolízis. A hat szénatomszámú cukrok elsősorban ezen
keresztül jutnak el a piroszőlősavig, majd a TCA cikluson keresztül szén-dioxidként a légkörbe és
NADH2-ként a terminális oxidációba kerülnek aerob körülmények esetén. Egyes esetekben (pl.
heterolaktikus erjesztés) egy dekarboxileződés után a pentózfoszfát-ciklusba kerülnek, és azon
keresztül jutnak el a piroszőlősavig.
C7-metabolizmus
A hét szénatomos cukrok kevésbé jellemzőek a mikrobiális anyagcsere-folyamatokban, mint kisebb
szénatomszámú társaik. Leggyakoribb képviselőjük a szedoheptulóz (2.1.6. ábra).
OH OH
O
OH
OH OH
OH
2.1.6. ábra: Szedoheptuloz
Nevezéktanuk is eltér a kisebb cukormolekuláktól: a hét szénatomos szerkezetet általában két
kisebb szénatomszámú cukor nevéből nevezik el (2.1.7. ábra)
Jelentőségük a mikrobiális liposzaharidokban van.
OH OH
OH
OH
OH OH
OHH
D-gluco D-glicero
2.1.7. ábra: D-glicero-D-gluko-heptulóz
Speciális energianyerő metabolizmusok (biometallurgy)
A szénforrások gyakran az energiaforrás szerepét is betöltik, az alábbiakban azonban olyan különleges
mikrobák kerülnek bemutatásra, amelyek szokatlan reakciókkal nyerik a szükséges energiát. Ilyenkor
gyakran különböző fémek redox átalakításának pozitív energiamérlegét aknázzák ki a mikrobák, ezért
iparilag is fontos lehetőséget rejtenek ezek a metabolizmusok a biobányászat (biometallurgia)
területén.
Az érckinyerés és –feldolgozás együttes neve a metallurgia. Ebben tudnak a speciális
anyagcseréjű mikrobák segíteni, ezért nevezik ezt biometallurgiának.
A metallurgiában az ásványokban kötött fémek kinyerésére vagy a klasszikus olvasztást használ-
ják, vagy valamilyen kioldást (=angolul: „leaching”). A leaching során 4-féle eljárás áll rendelkezésre:

2. Az ipari mikrobiológia biokémiai és fiziológiai háttere 45
© Németh Áron, BME © www.tankonyvtar.hu
1) savas kioldás (pl.: Zn), 2) lúgos kioldás (pl.: Al), 3) magas hőmérsékletű kioldás (pl.: S2-), 4)
kioldás komplex képzéssel (Au).
Ilyen kioldási folyamatot képesek a mikrobák is előidézni („bioleaching”), például a kalkopirit
(2.1.8. ábra, legelterjedtebb rézásvány, CuFeS2)-ből a réz kioldására képesek az Acidiphilium-törzsek
(A. multivorum, A. cryptum, A. organovorum) a 2.1.1. egyenlet szerint. Az Acidiphiliumok az alfa-
proteobakterek közé tartozó valódi baktériumok.
2.1.8. ábra: Kalkopirit [15]
OHFe2O0,5H2Fe2
Fe5S2CuFe4CuFeS
2
3
2
2
2
0
23
2
2.1.1. egyenlet
Ezeket a szokatlan – és nemrégiben felfedezett – mikrobákat Peng és mtsi [16] a következőkkel jel-
lemezte: Gram-negatív spórátlan pálcák, amelyek hőmérsékleti optimuma 40°C, pH optimuma 3,5 (!).
A tenyésztésükhöz használható tápközeget a 2.1.1. táblázat mutatja be, amelyen 40 óra alatt lezajlik a
fermentáció.
2.1.1. táblázat: Acidiphiliumok tápközege
Komponens Koncentráció
(g/L)
(NH4)2SO4 3
MgSO4*7H2O 0,5
K2HPO4 0,5
KCl 0,1
Ca(NO3)2 0,01
Szerves anyag 2
Élesztő kivonat 0,01%
A tenyészetet FTIR-rel vizsgálva megállapították, hogy polihidroxibutyrate-ot termel. Jó növeke-
dést tapasztaltak ramnóz, xilóz, mannit, fruktóz, glutaminsav és glicerin szénforrásokon, de nőtt szaha-
rózon, laktózon, hisztidinen, glutaminon és etanolon is.
Tetraciklinnel és ampicillinnel szemben érzékenynek találták ezeket a mikrobákat, míg a sejtfal-
szintézist gátlókkal (penicillinek cefalosporinok) szemben nem.

46 Ipari mikrobiológia
© Németh Áron, BME © www.tankonyvtar.hu
Rendszertanilag a valódi baktériumok (Eubaktériumok)\Alfaproteobakterek\Nitrospira csoportba
tartoznak közeli rokonaikkal a Leptospirillumokkal együtt. Ez utóbbi baktériumok vasoxidálók, ezért a
vaskinyeréshez használják iparszerűen óriási folyamatos üzemű oxidáló tartályokban.
Nagy hasonlóságot mutatnak, de a filogenetika szerint nem közeli rokonok a fentiek a szintén
acidofil vasoxidáló Ferroplasmákkal. Például a Ferroplasma acidophilum egy 35°C-os hőmérsékleti
optimummal rendelkező (mezofil) savkedvelő mikroba 1,7(!) pH optimummal, amely főleg a savas
bányameddőben (salak) található meg, különösen a pirit- (FeS) tartalmúban. Általában azért ezekben a
savas ércmaradványokban tenyészik, mert itt már más mikrobák (pl.: Leptospirillumok és
Acidiphilumok) lesavanyították a pH-t, olyan mértékben, amely az autotróf Ferroplasma számára
optimális. Ilyen körülmények között Ferroplasma már tud energiát nyerni a pirit Fe2+ ionjainak
oxidálásával, amelyhez O2-t használ terminális akceptorként. A folyamat során melléktermékként
kénsav termelődik. Ez tovább savanyítja a tápközeget.
2.2. Primer metabolitok
Az életfolyamatok következtében a tápközegbe kikerülő metabolitokat nevezzük elsődleges (primer)
metabolitoknak. Az életfolyamatokat a hasznosítandó szubsztrátok szerint csoportosítva már
bemutattuk (2.1. fejezet). Általában a különböző erjedések (amelyeket már szintén tárgyaltunk 1.3.
fejezet) termékei tartoznak ide, mint etanol, ecetsav, tejsav, propionsav, 1,3-propándiol, aceton,
butanol, vajsav. Ezek némlyikét élelmiszer-ipari, de főleg vegyipari célokra, illetve bioüzemanyagként
használják. Mindezek azt jelentik, hogy nagy mennyiségben alacsony áron kell ezeket előállítani, ezért
igen hatékony anyagcseréjű termelő mikrobákra van szükség, amely képes jó hozammal jó konverziót
elérni viszonylag gyorsan (=jó produktivitás).
Fiziológiai szempontból általában az energiatermelő folyamatok (NADH2 regenerálás, glikolízis, TCA
ciklus stb.) melléktermékei.
2.3. Szekunder metabolitok
De novo szekundermetabolit-termelés
A primer metabolitokkal szemben a szekunder metabolitok termelése nem a normál életfolyamatok
következménye, hanem a speciális környezeti feltételek által kiváltott reakcióé. A szekunder
metabolitok funkciója ugyanis előnyszerzés biztosítása megromlott életkörülmények (megváltozott
pH, csökkent tápanyag, csökkent élettér) esetén. Ilyen helyzeti előnyök kovácsolásával elősegíthető az
egyed és a faj fennmaradása „nehéz időkben” is.
Társadalmi szempontból a mikrobiális és növényi szekunder metabolitok játszanak fontos
szerepet.
A szekunder metabolitokkal kapcsolatban néhány fogalmat illik tisztázni:
anti- biosz=élet elleni, tehát az antibiotikum minden élőlény elleni szer
inhibítor=gátlószer, ami lassítja, vagy megállítja a növekedést
…sztatikum= leállítja a növekedést (pl.: fungisztatikum)
…cid=pusztító (bakteriocid, fungicid, viricid)
A helyzetielőny-szerzésre különböző stratégiák közül választhat egy adott élőlény:
A) Mozgás (baktériumok, protozoák mozgásszervvel rendelkezhetnek, és vizes közegben képesek
a helyváltoztatásra, növények pedig helyzetváltoztatásra).
B) Terjeszkedő típusú növekedés (pl.. fonalas gombák gyors kolonizációja, mindent beborító
penészfonalak, ehhez szükséges a félnedves szubsztrát).
C) pH megváltoztatása metabolitokkal: savtermelők (pH-t le), ammonifikálók (pH-t fel)
D) Hőmérséklet lokális megváltoztatása a mikrokörnyezetben (pl.: nagy mennyiségű metabolikus
hő termelése, ami megemeli a hőmérsékletet (pl.: komposztban, silóban), és ez kedvez a
termofileknek.
E) Sziderofórok (erős fémkomplexképzők) közegbe bocsátásával az enzimek esszenciális
fémkofaktorainak elszívása.

2. Az ipari mikrobiológia biokémiai és fiziológiai háttere 47
© Németh Áron, BME © www.tankonyvtar.hu
F) Antibiotikumok (szekunder metabolitok) termelése, amelyek a többi élőlényre káros hatással
vannak (sejtfalszintézist, fehérjeszintézist gátolnak stb.).
G) Extracelluláris enzimek termelése több célból is, például a nehezen hozzáférhető szubsztrátok
mobilizálására (amilázok, cellulázok stb.), vagy a konkurencia bontására (lizozim, kitináz stb.)
Az ipari mikrobiológia szempontjából az F) és G) pontok az érdekesek, ám figyelembe kell venni,
hogy a természetben az így kibocsátott anyagok (szekunder metabolitok és extracelluláris enzimek)
csak mikrokörnyezetben kell hogy hassanak, ezért a természetből izolált termelő törzsek csak kis
koncentrációban termelik ezeket az ágenseket, illetve a túl nagy dózis a termelő mikrobát is
károsíthatja.
A legfontosabb szekunder metabolitokat foglalja össze 2.3.1. táblázat.
2.3.1. táblázat: A legfontosabb mikrobiális szekunder metabolitok.
A szekunder metabolitok működését hatásuk szerinti csoportosításban mutatjuk be [17].

48 Ipari mikrobiológia
© Németh Áron, BME © www.tankonyvtar.hu
Antagonizmusok kialakítása
1A) gomba és baktérium küzdelme során a Cephalosporium gramineaum fonalas gomba a
baktérium sejtfalszintézisét gátló antibiotikumot termeli, de csak az őszi búza felszínén (in
vitro Petri csészén nem)
1B) mikoparazitizmus: egyik gomba a másik élősködője. Pl.: az északi fenyő parazitája egy
rozsdagomba (Monocillin nordinii), amelyen élősködik a Cronartium coleosporioides gomba,
és ezt antibiotikum termeléssel éri el. Ezt az antibiotikumot fel lehet használni a fenyő-
kártevő ellen.
1C) bakteriális predáció áll fenn a Mixobaktériumok és áldozataik között, amely utóbbiakat
antibiotikum segítségével ölik el
Olykor előfordul olyan is, hogy saját fajtársai ellen termel antibiotikumot egy mikroba, pl.:
Pseudomonas phenazinium (fenazin típusú antibiotikumot termel), és az antibiotikumot nem
termelő fajtársaikat alaposan túlnőtték.
1D) bakteriális kompetíció: az amőbák általában baktérimokat esznek, de egyes baktériumok (pl.:
Serratia marcescens) piros pigmentanyagot termelnek, ami elöli a protzoát (az albino, azaz
pigmentmentes fehér baktérium nem váltott ki cisztásodást, spóraképzést pusztulást az amőba
részéről)
1E) mikroorganizmus kölcsönhatása magasabb rendű élőlényekkel: gyakran toxintermelés
(növény ellen fitotoxin, ami gyakran más mikrobákra is antagonizmussal hat, mintegy
antibiotikum), amire a növény fitoalexin (~növényi antibiotikum) kiválasztásával válaszol.
1F) mikrobák inszekticid hatására példa a Beauveria bassiana (fonalas gomba) rovarok ellen,
illetve a Bacillus thüringiensis endospórájának ciklikus δ-toxinja a szúnyogok ellen.
A kullancsok is rendelkeznek letalitással járó gombás parzitával. A kullancsok természetes
ellenségeinek felhasználása a kullancspopuláció visszaszorítása érdekében a „Biokontroll”.
Mivel 4 ellenségét ismerjük a kullancsoknak, ők a potenciális jelöltek a biokontroll-funkció
betöltéséhez.
a) sisakos guineai madár kullancsokkal táplálkozik, ám ha sikerülne vele kullancslétszámot
csökkenteni, akkor a madárpopuláció is csökkenne, ettől megint nőhetne a kullancsok
száma, ekkor a madarat újra kellene telepíteni
b) kullancsok rovar parazitája az Ixodiphagus, amely a darázsfélékhez tartozik, és 0,1-5
mm-es nagyságú. Petéit a kulllancsba rakja, amelyek kikelésükkor megölik a kullancsot
c) fonalféreg- (nematóda) parazitával, de ez csak a vérrel magát jól teleszívott nőstény
kullancson hat
d) fonalas gombák parazitizmusa: Metarhizium anisopliae (zöld mészkór), Beauveria
bassiana (fehér mészkór), Lecanicillium psalliotae, Paecilomyces fumoroseus).
A Metharizium anisopliae [18] már rendelezik ipari felhasználással a maláriaszúnyog,
kullancs- és termeszirtások során. Az imperfect gombák aszkomiceta csoportjába
tartozik. 2007 óta ipari lipáztemelő is, mert az indiai biodízelgyártás során
szobahőmérsékleti optimummal rendelkező lipázokra volt szükség. Inszekticid hatásának
kifejtésekor megkapaszkodik a konidiospórája a rovar felszínén, és kutikulumbontó
enzimekkel hifát növeszt a rovar kültakarójába. Amint átér a rovar páncélján, találkozik
annak immunrendszerével, ezért vagy maszkíroznia kell magát, hogy az immunrendszer
ne taszítsa ki, vagy immunszupresszánsok termelésével csökkenti az immunrendszer
hatékonyságát.
A M. anisopliae metabolitjai a destruxinok, amelyek 5 aminosavból és hidroxisavból
felépülő cirkuláris fehérjék. Ez lehetőséget teremt a rekombináns biopeszticid termelésre.
A destruxinok egyébként citotoxikusak, mert Ca2+ transzportot végeznek, és így az
izmokat is blokkolják. Már több mint 30-féle destruxin szerkezetét derítették fel és írták
le.

2. Az ipari mikrobiológia biokémiai és fiziológiai háttere 49
© Németh Áron, BME © www.tankonyvtar.hu
1G) Mikrobák antagonizmusa magasab rendű álatok (és ember) felé: például a fuzáriumos búza
nem alkalmas még takarmányozási célra sem, mert a károsító fuzáriumok mikotoxinokat
termelnek, amelyek mind a haszonállatot, mind az embert veszélyeztetik. A ragadozók is a
mikrobák temelte mikotoxinok miatt esznek általában friss húst, kivéve a dögevők. A poshadt
víz ivásakor a vízben lévő algák termelte toxinok okoznak rosszullétet.
2) Fémionszállító metabolitok
A fémionok (főleg kationok) fiziológiai szerepe meghatározó, mivel az enzimkatalízis
kofaktorai, ezért egyes esszenciális fémionok elszívása a konkurencia elől komoly előnyhöz
juttatja a fémionszállító metabolit termelőjét. A nitrogénfixáló baktériumok esetében például
a nitrogenáz- enzim Mo2+ jelenlétében a legaktívabb, más fémionokkal csökkent aktivitású.
A fémionszállító metabolitok általában ionofórok, azaz olyan transzmembrán ágensek
(többnyire peptidek), amelyek az ionok (elsősorban alkáli, és alkáliföldfém) áramlását a
sejtmembránon keresztül meghatározzák vagy aktív transzporttal, vagy csatornaképzéssel.
A vasszállító faktorok (=sziderofórok) sok esetben antibiotikus hatásúak. A primer és
szekunder metabolitok határán helyezhetők el, mert ugyan nem szükségesek a növekedéshez,
de vashiányos körülmények között stimulálják a növekedést. A mikrobasejteknek kétféle
fémszállító rendszere van: egy kis és egy nagy affinitású. Utóbbihoz tartoznak a sziderofórok
is. Amíg van elegendő fémion jelen, addig a kis affinitású rendszer működik. Ám a fémion
csökkenése (pl.: valamilyen kelátképző megjelenése miatt) aktiválja a sziderofórtermelést.
Számos Streptomyces, Nocardia és Micromonospora faj termel sziderofór faktorokat. A
sziderofórok antibiotikus hatása abban nyilvánul meg, hogy termelőjük kiéhezteti a
konkurenciát vas szempontjából, mivel az nem képes a vas-szideramin komplex felvételére,
és így nem jut vashoz. A sziderofór termelője pedig komplex formában lévő vasat veszi fel.
Ilyen antibiotikum a nocardamin és a desferritriacetilfusigen.
Az ionofórok feladata mikrobákban, hogy segítsenek fenntartani a magas intracelluláris
K+/Na+ arányt is, azaz a koncentráció gradiens miatt kijutott káliumionokat visszapumpálják
a sejtbe, miközben a szintén koncetrációgradiens miatt bejutott nátriumionokat kipumpálják a
közegbe.
Az ionofór antibiotikumok közé tartoznak az enniatinok is, amelyek a destruxinokhoz
hasonlóan ciklikus peptidek és a K+/Na+ transzportban vesznek részt [20]. Piacon lévő
ionofór antibiotikumok a calcimycin, ionomycin, lysocellin és a lasalocid, amelyek a Gram-
pozitív mikrobák ellen hatásosak.
3) Mikroba és növény szimbiózisa szekunder metabolitok segítségével
3A) Tipikus példa a növény-fonalas gomba szimbiózisára az ektomikorhiza, ahol a növényi
tápanyagfelvételt a gyökér körül elhelyezkedő fonalas gomba segíti. Ilyenkor gyakran (pl.:
fenyőféléknél) termelnek a növény védelme céljából antibiotikus hatású szekunder
metabolitokat a baktériumok ellen, csökkentve a betegségveszélyt, és előnyhöz juttatva a
szimbiózist a tápanyagversenyben.
3B) Sziderofórok segítségével baktériumok is élhetnek növényekkel szimbiózisban, ahol a
sziderofórt termelő baktérium segíti a növény növekedését. A pszeudobaktin termelő
Pseudomonas fluorescens vagy Pseudomonas putida például elszívja a vasat a kórokozók
elől egy lineáris hexapeptid (sziderofór) segítségével.
4) Mikroba -nematóda szimbiózisa szekunder metabolit segítségével
Xenorhabdus luminescens baktérium és rokonai a Xenorhabdus fonalférgek endoszim-
bionitái. Ez a szimbiózis együtt támadja meg a rovarokat, amelyeket a baktérium által termelt
antibiotikum pusztít el, és amely antibiotikum a többi baktériumot is távoltartja a rovar-
tetemtől.

50 Ipari mikrobiológia
© Németh Áron, BME © www.tankonyvtar.hu
5) Mikroba-rovar szimbiózist is lehetővé tesznek egyes szekunder metabolitok
Erre példa a rizsen élő barna szöcske, amelyben élő Bacillus polymyxin polymyxin
antibiotikumot termel (ami egy ciklopeptidből és egy hidrofób farokból áll, és a Gram-
negatívak lipopoliszaharid rétegét károsítja), és így védi a szöcskét a rizs flórájától
(pl.:Xanthomonas campestris) .
6) Ferromon szerepű szekunder metabolitok
6A) A Mucorales nemzettségnél hím(+) és nőstény(-) ivarú gaméták találkozásakor a
heterotallikus (+ és -) hifákból történő zigóta kifejlődéséhez trisporinsav szükséges, de ennek
bioszintetkus útvonala részben a +, részben a - ivarú fonalakon van kódolva.
6B) Allomycesek esetében a szirenin nevű szekunder metabolitra a mozgékonyabb hím gaméták a
nőstények felé vándorolnak kemotaxissal.
6C) Egy különleges szimbiózis esetében a rovar (poszméh) nemi ferromonját az emésztőcsator-
nájában élő Aspergillus ochraceus termeli.
7) Morfológiai differenciálódást is befolyásolnak a szekunder metabolitok
Az antibiotikum-termelés gyakran a prespora fázisban történik, a vegetatív forma hanyatló
fázisában. A spórázás és az antibiotikum-termelés közötti kapcsolatot többféleképpen is
bizonyították, például egyes asporogén mikrobák elvesztették antibiotikum-termelő
képességüket, illetve a spóraképzés inhibitora gátolta az antibiotikum-termelést is.
A Bacillus brevis a spórájának külső felszínére Gramicidin S-t (szekunder metabolitot)
választ ki, amely gátolja az elektrontranszport láncot és a kicsírázást. Extracelluláris
proteázzal le tudja hasítani, és így a csírázást megindítani.
Penicillium brevicompactum mikofenolsavat termel a kondidium tartók kifejlődésekor,
aminek hatására a tartó a fény felé fordul.
A másodlagos anyagcsere a primer anyagcsere során keletkező metabolitokból építi fel a
szükséges szekunder metabolitokat. A főbb útvonalak az acetil-CoA-ból keletkező lipidek, illetve az
oxálecetsavból induló szekunder metabolitok, valamint a szintén acetil csoportból induló terpenoidok.
A szekunder metabolitok előállítását számos tényező befolyásolja. Mind szén-, mind
nitrogénforrás tekintetében a nehezebben hozzáférhető, limitált mennyiségű tápanyag kedvez a
másodlagos anyagcseretermékek keletkezésének. Így szénforrásból a különböző poli- és/vagy
oligoszacharidok kedvezőbbek az egyszerű cukroknál. Nitrogénforrásként a fehérjék alkalmazása nem
javasolt, mert a belőlük keletkező aminosavak negatív visszacsatolással hatnak a saját szintézisükre, és
ezen keresztül a szekunder anyagcserére. Foszforforrás tekintetében is a lassúbb növekedés (limitáció)
a kedvező.
Néha szükséges lehet egy-egy kulcsenzim termelődésének indukciója a hatékony másodlagos
anyagcseréhez, mint például Acremonium crysogenum esetében metioninadagolással lehet a
cephalosporin-szintézist fokozni, vagy a Claviceps törzseknél az ergot alkaloidok szintéziséhez
triptofánadagolást alkalmaznak. További tényezők lehetnek a minor komponensek (fémionok) és az
oldottoxigén-koncentráció, amelyek a szubsztrátokhoz hasonlóan hatnak (limitáció a kedvező), illetve
a CO2, amely gátolja például a penicillin képződését, vagy a fény, amelyről csak kevés adat áll
rendelkezésre. A mikrobaszaporodási fázis (trofofázis) és a termék keletkezésének fázisa (idiofázis) a
szekunder anyagcsere sajátosságaként különválik (míg a primer metabolitok termelése általában
növekedéshez kapcsolt a termék képződése), és gyakran az idiofázis alatt alkalmazandó hőmérséklet
alacsonyabb, mint a trofofázisé.
Szteroid- (szekunder metabolit) konverzió
A szteroidok közös vonása a kémiai alapváz (2.3.1. ábra), hogy a másodlagos anyagcsere termékei,
valamint hogy fontos fiziológiai szerepük van az alacsonyabb rendű és a magasabb rendű élőlények-

2. Az ipari mikrobiológia biokémiai és fiziológiai háttere 51
© Németh Áron, BME © www.tankonyvtar.hu
ben, így az emberben is. Ez utóbbi tulajdonságuknak köszönhetően a gyógyszeripar számára régóta
fontos vegyületek, így előállításuk és hasznosításuk számos kutatás tárgya.
2.3.1. ábra: A szteránvázas alapvegyület – a koleszterin– szerkezete
Bioszintézisük során a sejtek acetil-CoA-ból építik fel több lépésben az allilpentenil-pirofoszfátot
és az izopentenil-pirofoszfátot, amelyek addíciójából szkvalenon keresztül épül fel az alapvegyület, a
lanosterin. A szteroidneogenezis során ennek módosításával keletkezik a többi szteránvázas vegyület.
A szteroidok csoportosítása a funkció és a bioszintézis, valamint a termelő élőlény szerint történik
(2.3.2. ábra), így megkülönböztetünk:
2.3.2. ábra: „Szteroidok családfája” [22]
- Állati szteroidokat
- ízeltlábúak szteroidjai
- gerincesek szteroidjai
- hormonok: a kortikoidok olyan hormonok, amelyeket a mellékvesekéreg termel (innen a név
cortex – kéreg). Megkülönböztetjük a sóháztartásban szerepet játszó mineralokortikoidokat és

52 Ipari mikrobiológia
© Németh Áron, BME © www.tankonyvtar.hu
a cukorháztartásban szereplő glükokortikoidokat, valamint a nemi hormonokat (női/férfi nemi
szervek termelik, pl.: ösztrogén, progeszteron, tesztoszteron), illetve az anabolikus
szteroidokat, amelyek androgénreceptor-stimulátorok, és így az izomnövekedést serkentik.
- koleszterin: sejtmembrán-fluiditásért felel
- növényi szteroidok – fitoszterinek: általában a szteroidkonverzió ipari alapanyagai (2.3.3. ábra),
fiziológiai funkciójuk éppúgy a sejtmembrán befolyásolása, mint az állatok esetében a koleszteriné.
- gomba eredetű szteroidok – ergoszterinek
OHOH
OH
Sztigmaszterin Szitoszterin Kampeszterin
2.3.3. ábra: Ipari alapanyagként hasznosított fitoszterinek
Mint az a fentiekből kitűnik, a humánszervezetben nagyon fontos szabályozó szerepet töltenek be
a szteroidok, ezért a gyógyászati célú előállításuk több évtizedes múltra tekint vissza. Ezek kezdetben
kizárólag szintetikus vegyipari lépéseket tartalmaztak, ám a környezetvédelmi előírások
szigorodásával és a sztereokémia fejlődésével a biokémiai lépések egyre nagyobb teret nyertek. Mivel
ilyen vegyületek esetében az optikailag aktív formák több kiralitáscentrummal is rendelkeznek, és a
szteránváz egy egy pontján sztereoszelektív védőcsoportok alkalmazásával, majd eltávolításával
oldható csak meg a biológiailag aktív vegyület előállítása. Ezért célszerűbb a mikrobák enzimeinek
sztereoszelektivitását kihasználva egy-egy specifikus módosítást általuk elvégezetni.
Mivel azonban ezek a vegyületek a másodlagos anyagcsere termékei, ezért nem az akívan
növekedő sejttenyészet végzi a biokonverziót, hanem a felszaporított nyugvó állapotú sejtek indukciót
követően valósítják meg a sztereospecifikus átalakítást. Mivel a szteroid szubsztrát nem a mikroba
szénforrása (legfeljebb energiaforrás), ezért szinte veszteség nélkül (nagy hozammal) alakítható át
termékké. Technológiai nehézséget általában a szteroidok rossz vízoldhatósága jelent, amelyet
gyakran ciklodextrinek alkalmazásával küzdenek le. A ciklodextrinek olyan ciklikus poliszaharid-
molekulák, amelyek magukba zárhatnak apoláris hatóanyagokat (a gyűrű közepébe), és mivel a
ciklodextrinek vízben oldhatóak, így a „beléjük csomagolt” apoláris hatóanyag is oldhatóvá válik. A
ciklodextrin belseje és a folyadékfázis között adszorpciós megoszlás áll fent, azaz ha a csekély
mértékben oldott molekulát a mikrobák a folyadékfázisból elfogyasztják, helyükre a ciklodextrinekből
újabb molekula fog „kioldódni”.
A mikrobiális szteroidkonverziók igen széleskörűen alkalmazhatók, de leggyakrabban a 17-β-
redukció/17-β-oxidáció, illetve ennek valamelyik másik OH-csoporttal együtt történő szinkrón
redukciója/oxidációja (pl.: 3,17-diketoszteroid redukció) fordul elő.
A 17-es atomon történő oxidáció/redukció az élő szervezetekben is igen fontos, mivel a nemi
hormonok egymásba alakulását jelenti. A redox reakciót a 17-hidroxi-szteroiddehidrogenáz-enzim
katalizálja, amelynek aktivitása tehát egyes nemi folyamatok felelőse. Ennek alapján a
gyógyszeriparban is gyakran használják ezeket az enzimes (vagy nyugvósejtes) biokonverziókat a
terápiás hormonkészítmények előállítására. Az első publikáció 1937-ben Saccaromyces cerevisiae-ről
írja le ezt a képességet, azóta azonban számos baktériumról (Pseudomonas, Bacillus, Brevibacterium,
Mycobacterium, Streptomyces fajokról), gombáról (Aspergillus, Mucor, Penicillum, Hansenula,
Pichia, Saccharomyces), algáról (Chlorella, Chlorococcum) és protozoáról (Pentatrichomonas,
Trichomonas) derült ki a 17-OH-SDH aktivitás.

2. Az ipari mikrobiológia biokémiai és fiziológiai háttere 53
© Németh Áron, BME © www.tankonyvtar.hu
2.3.2. gyakori mikrobiális szteroidkonverziók
kísérő
hidroxilezés
17-β-redukció 17-β-oxidáció
C5
C6
C7
C10
C11
C12
C14
C15
Bacillus sp., Botryosphaerica obtusa
Bacillus sp., Botryosphaerica obtusa,
Bacillus sp.
Bacillus sp., Phycomyces
blakesleeanus,
Penicillium crustosum, P.chrysogenum
B. stearothermophilus, B. obtusa,
P. blakesleeanus
Botrytis cinerea, Rhizopus stolonifer
Absidia glauca
Cephalosporium aphidicola, R. stolonifer
A. glauca, B. obtusa
Bacillusok
A. glauca, Aspergillus fumigatus, B. obtusa
Az alábbiakban néhány jellemző ipari szteroidkonverziót mutatunk be.
Ketoredukció Saccharomyces cerevisiae-vel nemi hormonok előállítására
A szitoszterinből előállított androsztén-dionból (AD) ketoredukcióval tesztoszteront lehet előállítani,
noha az élő emberi szervezetben a férfi nemi hormon koleszterinből keletkezik. A ketoredukciót egyes
S. cerevisiae sejtek képesek elvégezni. A tesztoszteronból a C17 metilezésével előbb metil-tesztoszte-
ront (METESZ) lehet előállítani, majd az Arthrobacter simplex baktériummal ismét C1 dehidro-
génezést lehet elvégezni, ami Nerobolt eredményez (roboráló szer), illetve a női nemi hormonok
bioszintézise is koleszterinből és/vagy tesztoszterinen keresztül történik (ösztrogén/progeszteron).
Actinobacter fajok által végzett szteroidkonverziók [24]
Szteroid- (oldallánc) lebontás: Számos Actinobacter faj hasznosítja a szteroidokat, mint egyedüli szén-
és energiaforrást. A lebontási mehanizmus függ a lebontandó szteroid szerkezetétől és a mikroba
fajától. Az esztránokat általában az A gyűrű hasításával bontják le, míg a B gyűrű a leghozzáférhetőbb
része a molekulaváznak az androsztének és a pregnánok esetében. A koleszterinnél és az epesavaknál
a lebomlás kezdődhet az oldalláncon vagy a B gyűrűn. Nocardia fajok és még néhány Streptomyces
ismert A-gyűrű-hasító. Ennek ellenére az Actinobacterek többségét a B gyűrű hasításával jellemzik.
Elsődleges figyelmet kapnak a biotechnológiában azok az Actinobacterek, amelyek a szterinek
oldalláncait bontják, miközben a szteránvázat érintetlenül hagyják. Nocardia restrictus és más
Actinobacterek lehetővé teszik a szélső szubsztituens retroaldol eliminációját a C19-en és ösztrogént
képeznek 3,17-dioxo-C19(CO2OH)-androsztánból. Az ösztrogénhozam csupán 8% volt, ám 19-
hidroxiszitoszterol 3-acetát átalakításával 72%- is elérhető.
Az Actinobacterek (Nocardia restrictus, Proactynomyces erythropolis stb.) a szetrinek kólsavvá
történő alakítását is elvégzik.
A szterin oldalláncának lebontása 14 konszekutív biokémiai reakcióból áll, amelyeket legalább 9
különböző katabolikus enzim katalizál. A főbb megoldandó problémák a 3,17-diketoandrosztanok
oldallánc lebontásával történő előállításakor a következők: 1) a szteroidváz lebomlása, 2) a
reakciótermékek általi oldallánc-oxidáció, 3) szterinek alacsony oldhatósága a vizes közegben. Az 1)
probléma kiküszöbölésére a szubsztrát kémiai módosítása, vagy a 9α-hidroxiláz kémiai inhibíciója,
esetleg 3KSD blokkolt mutánsok használata jelenthet megoldást.
A szitoszterinből az oldallánc lehasításán keresztül Mycobacterium phlei baktériummal állítanak
elő számos üzemben androsztendiont (AD, androst-4-én-3,17-dion), amely több hormonális készít-
mény intermediere. A 70%-os konverziót követően diklóretánnal vagy szelektíven 85%-os metanollal
kiextrahálják a fermentlevet, és az extraktumot bepárolják. Az AD szintézis után acetilént addicio-
náltatnak, és a progeszteron hatású etiszterint nyerik, amelyet tovább is lehet alakítani spironolaktonná
(vízhajtó).
A termelő Mycobacterium phlei nevét a mykolsavnak köszönheti, amely egy hosszú szénláncú
zsírsav a sejtek membránjához kötve. E különleges sejtfalszerkezet miatt nem működik a Gram-festés
sem, illetve különösen ellenállóvá válik a mikroba.

54 Ipari mikrobiológia
© Németh Áron, BME © www.tankonyvtar.hu
A mikrobák életében az oldallánc lebontásnak a fiziológiai szerepe az ATP (energia) generálás
[23]. Ehhez a katabolikus enzimek 4 csoportját használják a mikrobák fel: a zsírsavak β-oxidációs
lebontó enzmeit, az ω-oxidációs enzimeket, a metil-krotonil-CoA karboxiláló rendszert és a propionil-
CoA karboxiláz rendszert. Ezek 9 enzime vesz részt a szteroidok oldallánclebontásában 14 konszeku-
tív reakcióban, amelyek a következőképpen összegezhetők:
1 szitoszterin +3FAD++3NAD++3 coA→ 3 FADH2+ 3 prop-CoA +3 NADH2 +1 AcOH
3 prop-CoA+1 AcOH →18 NADH2+ 7 FADH2 + 1 AD
A keletkezett redukált koenzimek végső oxidációjakor több mint 80 ATP molekula keletkezik 1
oldallánc lebontásakor.
A kapott AD a kiindulási intermediere az etiszterinnek (progeszteron hatású, kémiai úton acetilén
addícióval előállított AD) és a progeszteronoknak
Hidroxilezés: Az Actinobacterok képesek a szteránváz hidroxilezésére különböző pontokon
(1,2,6,7,9,11,12,14,15,16,17). A 9a hidroxilezést a Corynebacterium, Nocardia, Rhodococcus és
Mycobacterium nemzetségek egyes fajai végezhetik. Az Actinobacterok ketoszteroid-9-hidroxilázát
(9-KSH) eddig nem írták le részletesen, valószínűleg az izolálási és tisztítási nehézségei miatt. Egy
ternér 9-KSH komplexet, amely flavoprotein reduktázt és ferredoxint is tartalmaz, izoláltak egy
Nocardia fajból. Az Arthrobacter oxydans 9-KSD-a és 3-KSD-a is plazmidon kódolt, de a nukleotid
szekvenciájuk még nem ismert.
A Rhodococcus 9-KSH-a egy dimer monoxigenáz részt tartalmaz, és a kódoló géneket is
azonosították már.
Az Actinobacterok 11a és 11b hidroxilálóképessége egy ritka tulajdonság. Reichstein’s szubsztrá-
tot hidroxileztek Streptomyces fradiae segítségével hidrokortizonná, de csak csekély hozammal
(1,5%).
Az Actinobacterok azon tulajdonsága, hogy képesek a szteroid vegyületeket a 22,23,24, és 26-os
pozicióban is hidroxilezni intenzív kutatás alatt áll, ami ahhoz is vezetett, hogy feltárják a szteroid-
oldallánc lebomlásának mehanizmusát. Mycobacterium fortuitum szitoszterin átalakítása közben
találtak 22-es atomon történt hidroxilezés 3-oxo-4én származékait. A további kutatások pedig azt
mutatták, hogy a szterinek sorozatos hidroxilezése az oldallánc ezen poziciójában az Actinobacterok
által végzett oldallánclebontás egyes lépései.
A hidrokortizon-előállítás első lépésében Mycobacterium smegmatisszal a szitoszterin
oldalláncát eltávolítják, és a baktérium ugyanazon fermentációban a C9-re OH-csoportot is bevisz
(hidroxilez). Az így kapott 9α-hidroxi-androszténdionból kémiai átalakításokkal állítják elő a
hidrokortizont (2.3.4. ábra). Ezek első lépése a vízelvonás, majd vízaddició (e kettő következtében az
OH csoport átkerül 9-ről a 11-es szénatomra), majd oldallánc-kialakítás következik acetilénnel, végül
annak módosítása.
A fermentációs lépéshez glicerint adnak szénforrás gyanánt, a szitoszterin oldallánchasítása pedig
– az előbbi példához hasonlóan – az energiát szolgáltatja a sejtek számára. Mivel a szitoszterin
kristályos formában van jelen, amelynek felszínéhez a mikrobák odatapadnak, valamint a termék is
kristályos formában keletkezik, látszólag nem történik semmi, pedig a „kristályfermentáció” nyomán
értékes átalakításokra kerül sor a szubsztráton.
A fermentációval előállított 9α-hidroxi-AD feldolgozása az AD-előállítással analóg módon
történik: először teljes extrakcióval kinyerik a maradék szubsztrátot és a keletkezett terméket
(diklórmetánnal), majd szelektív extrakcióban elválasztják a kiindulási és termék szteroidokat
(diizopropil éterrel a terméket oldják, metanollal a szitoszterinmaradékot).
A kinyert 9a-hidroxi-androszténdiont 85%-os foszforsavval főzik víz kilépés érdekében, majd
ismét vizet addicionáltatnak és 11a-hidroxi-androszténdion keletkezik. Erre acetilént addicionáltatnak,
majd az így bevitt oldallánckezdeményt alakítják tovább, amíg a hidrokortizonhoz jutnak.

2. Az ipari mikrobiológia biokémiai és fiziológiai háttere 55
© Németh Áron, BME © www.tankonyvtar.hu
OH
O
OH
OH
O
OH
O
OH
OHOH
OH
OH CH CHOH
O
OH
O OH
Szitoszterin 9-hidroxi-androszténdion
H2O
, 85% H3PO4+H2O
-hidroxi-androszténdion
Kálium+acetilén
11-hidroxi-etiszterol Hidrokortizon
Mycobacterium smegmatis
5-6 nap, 11-12g/L, 70% konverzió, 1-3% inokulumm
2.3.4. ábra: Szitoszterinalapú hidrokortizon- (bio)szintézis
Dehidrogénezés: A C-C kötések dehidrogénezésének képessége egy jellegzetes tulajdonsága az
aktionobaktériumoknak, köztük is az Actinoplanes, Arthrobacter, Corynebacterium, Micromonospora,
Mycobacterium, Nocardia, Nocardioides, Rhodococcus és Streptomyces nemzetségek egyes fajainak.
Általában a C1, C4, C7, C8, C9, és C16 szén és az azt követő szénatom között alakulnak ki a
dehidrogénezés miatt kettős kötések az Actinobacterok aktivitásának köszönhetően.
A 3-ketoszteroidok mikrobiológiai 1(2) dehidrogénezése (amit az 1950-es évek óta használnak) a
prednizolon- és predniszteroidgyártás alapja. Így az Arthrobacter simplexet használják a pregnánok,
androsztánok és szterin, illetve epesavak származékainak előállítására több évtizede.
A hidrokortizon hatékony gyulladáscsökkentő hatóanyag, de kedvezőtlenül hat a sóháztartásra,
ezért a kevesebb mellékhatással rendelkező rokon vegyületeit (pl.: prednizolon) is forgalmazzák. A
prednizolont aerob biokonverzióval az Arthrobacter simplex (más néven Nocardioides simplex vagy
Pimelobacter simplex, propionsav baktérium) állítja elő hidrokortizonból (2.1.13. ábra).
OH
O
OH
O OH
OH
O
OH
O OH
OH
O
OH
O N N
O
O
OH
O OH
O
Hidrokortizon
Arthrobacter simplex
Prednizolon Deperzolon
Triamcinolon-acetenonid(Ftorocort kenõcs)
2.3.5. ábra: Hidrokortizon biokonverziója prednizolonná és származékaivá
Hidrogénezés: A szteránváz kettős kötéseinek hidrogénezését gyakran a dehidrogénezésért felelős
enzimek végzik. Így a Nocardia opaca és N. corallina 3-ketoszteroid-dehidrogenáza (3-KSD)
szteroid- dehidrogénezést és hidrogénezést is végez az 1(2) pozícióban. Ezzel szemben a
Mycobacterium globiforme esetében 3-KSD és 1(2)-dehidrogenázok külön enzimek. Mycobacterium
törzsek, amelyek a szitoszterinből androszténdiont (AD, androszt-4-én-3,17-dion) és androszt-

56 Ipari mikrobiológia
© Németh Áron, BME © www.tankonyvtar.hu
diéndiont (ADD, androszt-1,4-dién-3,17-dion)-t állítanak elő szintén rendelkeznek külön 1(2)
dehidratáló és 1(2) redukáló enzim rendszerrel.
A C4 kettőskötés-redukciójára Streptomyces faecalist használnak.
A pregnánok C1-es kettős kötésének redukciója gyakran a C20 β-redukciójához kapcsolt, amint az
az Arthrobacter simplex és A. globiformis példáján is látszik. Ez utóbbi reakció általában szub-
optimális levegőztetés esetén fordul elő; a reakcióelegy levegőztetése az egyensúlyt a C1-én-20-
ketoszteroid5 felé tolja el.
Epoxilálás: Egyes Nocardia és Corynebacterium fajok képesek a 9(11)-dehydroszteroidokat a 9α,
11α-epoxi-analógjaikká alakítani. Az ilyen átalakításoknál a megfelelő 9α-hidroxi szteroid a
valószínűsített köztitermék.
Alkoholok oxidációja ketonokká vagy aldehidekké: Az Actinobacterok nagyon hatékonyak a 3b-
hidroxilcsoport oxidációjában a megfelelő keton előállítására. Általában széles a szubsztrát-
specifitásuk, és nagy a sztereospecifikusságuk, ami ipari szempontból rendkívül kedvező, mivel szinte
bármilyen szteránvázas szubsztrátot optikailag tiszta termékké képesek alakítani. Így a
Corynebacterium mediolanum a dehidro-epiandrosteront (DHEA) hatékonyan alakítja tesztoszteronná.
A C5-én-3β-hidroxi szteroidok 3β hidroxicsoportjának oxidációját az Actinobacterok a C5->C4 kettős
kötés izomerizációjával kapcsoltan végzik. Nocardia erythropolis esetében megállapították, hogy a
szteroid C5 kettőskötésének hiánya megakadályozta a 4én-3-oxo forma képződését. Ezek az enzimek,
amelyek a 3β-hidroxi-5-én átalakítását 3-oxo-4-énné végzik az oxidázok családjába tartoznak (pl.:
koleszterin oxidáz (CO), EC 1.1.3.6.). Az Actinobacterok CO enzimét jelenleg a koleszterin
kimutatására analitikai célokra használják. Ilyen CO enzimaktivitást számos Actinobacterban találtak,
köztük Nocardia, Rhodococcus, Corynebacterium, Streptomyces esetében. A génmérnöki módszerek
lehetővé tették a hiper- és extracelluláris CO-termelő Actinobacterok létrehozását.
Az egyik leggyakoribb oxidáció a 17β-hidroxi-szteroidok 17-oxo származékainak előállítása
Mycobacterium phlei segítségével (ld. androszténdion előállítás).
Az Actinobacterek további számos hidroxilcsoport oxidációjára képesek (pl.: 3α, 6β, 7α, 11β
stb.), azonban ezek a megfelelő kémiai eljáráshoz képest kevésbé hatékonyak egyelőre.
Szteroidészter-hidrolízis: A szteroidok acetilszármazékainak hidrolizáló képességét a Coryne-
bacterium, Nocardia, Arthrobacter és Streptomyces fajoknál írták le. A hidrolízis itt is kettőskötés
izomerizációhoz kapcsolt: a 3β-acetoxi-C5-én szteroid dezacetilezésekor a 3-hidroxi-C4-én származék
keletkezett.
Ketonok oxidációja észterekké vagy laktonokká: Szteroid jelenlétében a monoxigenáz-enzim oxigént
épített be a C17 és C20 közé progeszteronon Rhodococcus rhodochrous esetében. Az enzim katalizálja
a progeszteron és annak 11α/β-hidroxi származékának oxidatív észterezését, viszont az AD lakto-
nizálását nem. Az enzim tulajdonságai meglehetősen különböztek a gomba eredetű monooxige-
názokétól. A kódoló gént rhodococcusból rekombináns E. coli-ba klónozva nem tapasztalatak akti-
vitást progeszteronon.
Karboniltartalmú vegyületek redukciója: A 3-karbonil csoport redukciója, amelyet az anaerob mikro-
organizmusokra, élesztőkre és egyes fonalas gombákra leírtak, ritka az Actinobacterok között. Ez a
reakció gyakran hidrogénezéshez kötött. Koleszt-4-én-3-on redukálható 5-kolesztán-3-ol-lá Nocardia
corallina segítségével. A szterinek és a pregnan C17-es karbonilcsoportjának Actinobacteres reduk-
ciója az oldallánchasító folyamatokhoz kapcsolt. Így például a Streptomyces levendula által végzett
progeszteron – AD-átlakítás melléktermékeként 1-dehidro-tesztoszteron is képződött.
Az Actinobacterek oldallánclebontó és egyidejűleg 17β-redukciót eredményező képessége
lehetővé teszi az AD intermedier kikerülésével az egylépcsős biotechnológiai tesztoszteron elő-
állítását. Az irodalomban már található olyan publikáció, amely szerint szterinekből egy lépésben kris-
tályos tesztoszteront lehet 98-99%-os tisztasággal előállítani Mycobacterium törzsek segítségével [25].
5 „C1-én” és hasonló kifejezésekkel a C1 szénatom melletti kettős kötést jelöltük.

3. Az ipari mikroorganizmusok és technológiák tárgyalása törzsrendszertanba illesztve 57
© Németh Áron, BME © www.tankonyvtar.hu
3. AZ IPARI MIKROORGANIZMUSOK ÉS TECHNOLÓGIÁK
TÁRGYALÁSA TÖRZSRENDSZERTANBA ILLESZTVE
A jegyzet eddigi fejezeteiben már igen sok mikroorganizmus került megemlítésre, többségükben már
ipari használatban lévők, de mint azt már szintén bemutattuk, számos mikroba rejt kiaknázható
potenciált, amely realizálódhat az adott mikroba tenyésztésével, illetve napjainkban gyakrabban az
adott mikroba génforrásként való felhasználásával. Ezért az ipari mikrobiológia keretein belül mind a
már használatban lévő, mind pedig a kiaknázható potenciált rejtő mikrobákat célszerűnek találjuk
bemutatni, és mivel így már igen nagyszámú törzs kerülhet szóba, célszerű a rendszertani besorolás
szerinti ismertetésük. Ez esetleg olyan összefüggésekre is rávilágít, hogy két különböző technológia
alapjait jelentő mikroba milyen rokonsági kapcsolatban áll egymással, és így az egyes technológiáknál
szerzett tapasztalat segíthet a másik technológia megértésében, fejlesztésében.
3.1. Mikrobiális rendszertan
A tudomány több ezer éves fejlődése során felhalmozott óriási információ halmaz kezelése érdekében
minden szakterületen valamilyen rendezési elv mentén az információkat rendszerbe foglalják. A
biológia, azaz az élővilág tudományterületén az információk klasszikus rendszerezése a megfigyelt
hasonlóságok és az ezek alapján felállított vélt rokonságok alapján történt. Az első ilyen rendszertant
Arisztotelész állította fel az élővilág rendszerezésére több mint 2000 éve a Historia Animalium c.
írásában [26]. A teremtményeket azok hasonlósága alapján csoportosította: állatok vérrel és vér nélkül,
vízi és szárazföldi állatok. Úgy gondolta, hogy a teremtmények hierarchikusan sorbarendezhetők a
legalacsonyabbtól a legfejlettebbig (ami az ember). Az élőlények lényegét fixnek és megváltoztatha-
tatlannak gondolta, és ez a nézete fenn is maradt a következő 2000 évben. Amit viszont a mai napig az
ő nyomán használunk, az a binomiális (kettős) elnevezése az élőlényeknek. Ő még úgy gondolta, hogy
az élőlényeket hasonlóságaik alapján egy „genus”-ba lehet sorolni, ahol az egyes tagokat egy jellemző
saját karakter különbözteti meg (=”difference”), ezért a kettős név Arisztotelésznél „genus,
difference”-ként nézett ki.
Tanait tanítványa, Theophrastus változtatás nélkül kiterjesztette a növényekre, és munkásságával
(növények leírása és nevelése) a botanika alapítójává vált.
A XVI. sz.-ig nem történt érdemi változás a taxonómia területén, míg Andrea Cesalpino olasz
fizikus az általa részletezett növényi leírásokat a növények gyümölcsei és magjai alapján nem kezdte
el rendszerezni, amellyel hatást gyakorolt a következő nagy reformerre Carolus Linnaeus-re (1707–
1778). Linné idejére számos leírás gyűjtemény létezett (pl.: az egyik legnagyobb a svájci fizikus,
Gaspard Bauhin (1560–1620) által írt illusztrált 6000 növény leírását tartalmazó), amelyek
inkoherensek voltak. 1735-ben jelentette meg The system of nature c. művét, amelyben a növényeket
és állatokat a királyságok szintjétől a fajokig besorolta. Ezt a művét többször átdolgozta, mígnem a
világszerte ismert rendszertan létrejött. Például a bálnákat eredetileg a halak közé sorolta, és ahogyan
ő élete végéig javítgatta, frissítgette a rendszertant az újabb és újabb tudományos eredmények alapján,
úgy napjainkig is a rendszertan egy folyamatosan fejlődő tudományág (3.1.1. táblázat).
3.1.1. táblázat: A rendszertan fejlődése
Linnaeus (1735) Haeckel (1866) Chatton (1937) Copeland (1956) Whittaker (1969) Woese (1977) Woese (1990)
"2 ország" "3 ország" "2 birodalom" "4 ország" "5 ország" "6 ország" "3 domén"
Eubaktériumok Baktériumok
Ősbaktériumok Ősbaktériumok
Protiszták Protiszták Protiszták
Gombák Gombák
Növények Növények
Állatok Állatok Állatok Állatok Állatok
NövényekNövények
- Protiszták
Eukarióták
MoneraMonera
Növények
Prokarióták
Eukarióták

58 Ipari mikrobiológia
© Németh Áron, BME © www.tankonyvtar.hu
Ez a fejlődés a XX. században az addig csak módosítgatott rendszertant új alapokra helyezte: míg
a klasszikus rendszertan célja, hogy a valós rokonságokat megállapítsa, és a hasonló fajokat hasonló
rendszertani kategóriákba helyezze, addig a modern rendszertani módszerek rávilágítottak arra a
tényre, hogy a valóságban csak vélt rokoni kapcsolatok vannak egyes esetekben a rendszertanilag
rokonnak tartott fajok között. A modern „rokonság” tesztek természetesen a genetika eszköztárát
használják, és a genetikai állomány azonossága/különbözősége alapján állítják fel a pontos rokonsági
fokot (rokoni távolságot), amelyek alapján filogenetikus fa felállítása válik lehetővé.
A legelterjedtebben használt mikrobaleírások és rendszertan a Bergey I. kiadásában került
publikálásra a ’80-90-es években, majd a modern genetikai alapú II. kiadást megkezdték 2000-ben
(jelenleg is tart). Ennek következtében jelenleg egy átmeneti időszakot élünk, amikor a régi már
elavult, az új pedig még fejlesztés alatt van. A modern kor talán legnagyobb (ám néha megkérdőjelez-
hető hitelességű) információhalmaza az internet, amelyen jelenleg is készül a „Tree of Life” web-
projekt. Ennek lényege, hogy az interneten elérhető tudományos publikációkat összegyűjtse és
felhasználja a mikrobák rendszertanának létrehozásához.
Az új rendszertan hátránya, hogy sokkal lexikálisabb, és kevésbé gyakorlatias: azaz egy adott faj
rendszertani besorolásából a tulajdonságai nem igazán derülnek ki, míg viszont számos további név
igen. Például az Escherichia coli a régi rendszertanban „Gram-negatív\pálca\fakultatív anaerob\
\Escherichia nemzettség” tagjaként került meghatározásra, míg a modern filogenetikus fán
„Baktériumok\Proteobacterek\Gammaproteobacter\Enterobacteriales\Enterobactericeae\Escherichia”
útvonalon található meg. Nyilván az előbbi leírásból több információ volt nyerhető a mikroba felis-
merésével kapcsolatban. Éppen ezért célszerűnek tűnik a klasszikus rendszertan rövid ismertetése is.
Klasszikus rendszertan
A klasszikus mikrobiológiai és biokémiai fenotípizálások segítségével rendszerbe sorolt mikrobák
leíró gyűjteménye a Bergey’s Manual of Systematic Microbiology-ban jelent meg több javított
kiadásban. Ebben a rendszreben a prokariótákat 4 divízióra osztották a sejtfalszerkezetük alapján: I.
Gram-negatív baktériumok (ők egy külső membránréteg miatt nem festhetőek Gram-festéssel), II.
Gram-pozitív baktériumok (amelyeknél a festékkel reagáló mureinháló (poliszaharid) szabadon
hozzáférhető, tehát festhetőek), III. Merev sejtfallal nem rendelkezők (pl.: csavart baktériumok),
illetve IV. Ősbaktériumok (primitívnek látszó, mureint nem (esetleg pszeudomureint) tartalmazó
sejtfallal rendelkező mikrobák) (3.1.1. ábra, lásd még a 62. oldalon).
Filogenetikus rendszertan
A modern – genetikailag támogatott – rendszertan létrehozásához a filogenetikai (phülon: törzs;
geneseis: születés) analízisre van szükség. Ennek a tudományterületnek a célja, hogy az evolúció
rekonstruálásával állapítsa meg a rokoni/leszármazotti viszonyokat az élőlények között. Ehhez
szükséges az elmúlt évtizedek kutatásainak köszönhetően felhalmozódott genomikai és proteomikai
ismeretek rendszerezése, azaz fejlett bioinformatikai infrastruktúra. A filogenetikai analízist 3
egymásra épülő szinten kell elvégezni: 1. szint: genomikai adatokból az azonosságok/különbségek
feltárásával a rokoni kapcsolat és távolság felmérhető; 2. szint: proteomikai adatok segítségével az
egyes fehérjék és változásaik követhetők nyomon, így a rokoni távolság és az evolúciós fa
pontosítható; 3. szint: a különböző algoritmusok különböző evolúciós fákat eredményeznek, amelyek
közül ki kell választani a legvalószínűbbet.
Az egyik sarkalatos pontja a módszernek a megfelelő indikátor génállomány kiválasztása, mivel
ezzel szemben támasztott elvárás, hogy stabil legyen (lehetőleg konzervált, normál életfolyamatok
alatt nem változó), ugyanakkor a nagyobb evolúciós lépést jelentő behatásokra, mutációkra érzékeny
legyen. Ilyen konzervált, de a mutációkat gyűjtő genom található a mitokondriumokban és a
riboszómákban, ezért ezek szekvenciájának meghatározásával lehet a genetikai rokonságokat
feltérképezni.
A másik sarkalatos pontja az analízisnek a megfelelő algoritmus kiválasztása a rokoni távolságok
feltérképezésére. Az egyik leggyakrabban alkalmazott módszer a Neighbour joining (NJ – szomszéd-
összevonó módszer), amelyhez szükséges egy távolságmátrix felírása a vizsgálandó fajok között, majd
ennek transzformált változatában kell megkeresni a legkisebb értékeket (legközelebbi szomszédok),

3. Az ipari mikroorganizmusok és technológiák tárgyalása törzsrendszertanba illesztve 59
© Németh Áron, BME © www.tankonyvtar.hu
megvizsgálni az ő távolságukat a közös ponttól, majd a többi vizsgálandó faj távolságát ugyanettől a
ponttól, azután kezdődik elölről, de az előbb felismert szomszédokat egyként kell kezelni az új
távolsági mátrix felírásakor. Egy másik algoritmus a maximum likelihood (ML – legnagyobb való-
színűség módszere) módszer, amely standard statisztikai megközelítéssel vizsgálja a legvalószínűbb
elágazásokat [30]. Egy harmadik széleskörűen használt módszer a maximum parsimony (MP – legna-
gyobb takarékosság módszere), amely a legkevesebb evolúciós eseményt kereső algoritmus.
A filogenetikus analízis eredményeként rajzolható fel a filogenetikus fa, amelyen az elágazások
távolsága egyenesen arányos az elágazás végpontjain lévő mikrobák genomazonosságával. Egy filo-
genetikus fát mutat a 3.1.2. ábra (lásd még a 63. oldalon).
Az első fontos lépés a mikrobák törzsfejlődésének megértésében a megfelelően objektív krité-
riumok keresése a főbb rendszertani csoportok definiálásához. Jelenleg a baktériumokat mintegy 23
főbb csoportba, azaz törzsbe sorolják, a 16S rRNS fáik alapján. Némely ezen törzsek közül csupán né-
hány fajt tartalmaz (pl.: Thermomicrobia, Chrysiogenetes, Fibrobacteres, Deferribacters), míg mások
– mint például a Proteobaktériumok, Bakteroidák, Cianobaktériumok, Actinobacterok (nagy G+C
tartalmú Gram-pozitívak) és a Firmicutes (kis G+C tartalmú Gram-pozitívak) – nagyon nagyok, és
fajok ezreit számlálják, ideértve az ismert baktériumok több mint 90-95%-át. A kisebb törzsek (pl.:
Fibrobakterek, Chlorobi) valamikor valamelyik nagyobb bakteriális csoport részeként kerültek be-
sorolásra a közös morfológiai/fiziológiai jellemzőiknek megfelelően, de később leválasztották őket a
16S rRNS térkép alapján.
Bakteriális filogentika [27]
A mikrobiológiai rendszertanban a Woese-féle reformok (1990) óta a besorolásokat a következő
csoportokba teszik meg: domén (domén)>kingdom (királyság)>phyla (törzs)>class (osztály) >order
(rend)>family (család)>genus (nemzetség)>species (faj). Az ipari mikrobiológiában használatos
„törzs” kifejezés gyakran nem a fenti rendszertani csoportra utal, hanem arra, hogy egy adott faj sejt-
vonaláról van szó (valamely egyed leszármazottai). Minimális eltérések (pl.: termék hozamban) lehet-
nek egy adott faj sejtvonalai között is. Azoknál a mikrobáknál, amelyeket alaposan tanulmányoztak
már, előfordulnak a fajon belül alfajok (v. fajták vagy variánsok), és azoknak is lehetnek különböző
sejtvonalaik, törzseik.
Azt, hogy milyen tulajdonságok határozzák meg a törzs besorolását alacsonyabb rendű csopor-
tokba, mint például osztályba, rendbe vagy családba, sosem definiálták egyértelműen a tudományos
világban. Éppúgy, mint azt a kérdést, hogy mi alkot egy aldivíziót vagy osztályt egy törzsön belül.
Például, a Proteobakterek törzsét jelenleg 5 aldivízióra (vagy osztályra) osztják, névszerint α, β, γ, δ és
δ proteobakterek. Ezek mindegyike főbb csoportokat alkot a baktériumokon belül, és többségük
világosan megkülönböztethető egymástól, illetve más baktériumdivízióktól, mind filogenetikus
fájukban, mind számos más forrásban. Az alacsonyabbrendű kategóriákba (alkategóriák) való
besorolás objektív feltételei nélkül nem világos, hogy miért pont ezek a főbb bakteriális csoportok
kerültek aldivízióként (vagy osztályként) meghatározásra, míg más alig jellemzett csoportok néhány
ismert fajjal főbb törzsként kerültek leírásra. Így a rendszertani besorolása a különböző bakteriális
csoportoknak jelenleg teljesen önkényes.
Az a kérdés, hogy hogyan lehet a legalacsonyabb rendszertani beosztást a baktériumokon belül
definiálni, kezdetben egy ideális szempontból közelíthető meg. Egy ideális osztályozó rendszerben a
törzs a leszármazás főbb vonalait mutatja be, és a gyökér egy közös bakteriális őst reprezentál. Minden
nagyobb elágazás, amely a törzsből ered, a fa főbb csoportjait (vagy (mikrobiális) törzseit) jeleníti
meg. Egy ideális osztályozó rendszerben ezen elágazások sorrendje a törzsön belül ismert, és a rend-
szer relatív nemzedékeken alapszik. Fontos, hogy ezek a törzsek (vagy elágazások) világosan elkülö-
nüljenek egymástól a fán való helyük alapján. Minden szervezetnek, ami egy elágazást alkot,
rendelkeznie kell olyan tulajdonságokkal, amelyek alapján más ágak szervezeteitől megkülönböz-
tethető. Kisebb ágak és gallyacskák, amelyek a főbb ágak hajtásai, egyre kisebb rendű rendszertani
fokokat reprezentálnak, például kisebb ágak az osztályok, rendek, családok, gallyak a nemzettségek,
fajok. Az azonos kisebb ágakon lévő szervezeteknek több közös tulajdonsága is kell legyen egymás-
sal, mint a fő ág más ágainak szervezeteivel.
Az eddig fő ismertetőjegy alapján meghatározott főbb csoportok száma minimális. Mivel azonban
a fajok különböző elkülönített csoportjai, amelyek ugyanazon ponton ágaznak el még akkor is, ha

60 Ipari mikrobiológia
© Németh Áron, BME © www.tankonyvtar.hu
nincs egyedülálló közös tulajdonságuk, jelenleg provizórikus altörzseit képviselik valamely törzsnek.
Ugyanakkor, amint további ismertetőjegy vagy más információ (ami elkülöníti ezeket az altörzseket,
vagy egyértelművé teszi elágazásuk sorrendjét) elérhetővé válik, ezek az altörzsek egyedi törzsekké
lépnek elő, vagy két elkülönített törzsre osztják az eddigit. A legtöbb főbb törzs jelenleg is valamilyen
csoportspecifikus ismertetőjegy alapján különíthető el. Ezen kritériumok alapján a Proteobaktériumok
legalább 3 fő törzsre oszthatók, mindegyikük jól elkülöníthető a többitől, és az elágazásuk sorrendje is
egyértelmű. A δ-proteobakterek távolról rokonai az α-proteobaktereknek, de ez a kapcsolat még
további finomításra szorul. A β,γ-proteobakterek számos közös ismertetőjeggyel rendelkeznek. Noha
néhány ismertetőjegy a γ-proteobaktereket megkülönbözteti, és későbbi elágazásra utal, a kapcsolat e
két aldivízió (vagy altörzs) között további tanulmányozást igényel. Az ismertető szekvenciákon
alapuló faábra azt is jelzi, hogy a jelenleg fő törzsként jegyzett Bakteroidák (vagy Citofágok,
Flavobakterek Bakteroida csoport), Chlorobi és Fibrobakterek megbízhatóan ugyanazon pontból
ágaznak el, és számos megkülönböztető tulajdonsággal rendelkeznek, amelyek egyedi törzsekké teszik
őket. Így a fentiekben kikötött kritériumoknak megfelelően ezeket a csoportokat egy törzsbe kellene
helyezni, és vagy osztályokat, vagy rendeket kellene alkossanak ebben a fő törzsben. A relatív
elágazásuk ezeknek az alcsoportoknak az FCB törzsön belül szintén jól megállapítható számos
alcsoport-specifikus ismertető szekvencia alapján.
A baktériumok törzsének elágazási sorrendje
A bakteriális filogenetika központi problémája, hogy megértsük, hogy a különböző bakteriális törzsek
hogyan kapcsolódnak egymáshoz, és hogyan ágaznak el a közös őstől. A 16S rRNS vagy más
fehérje/gén alapú filogenetikus családfák nem tudták megoldani ezeket a kritikus aspektusokat, más
jelentős kérdéssel (pl.: fotoszintézis eredete, eukarióta sejt eredete) együtt. Fontos betekintéseket nyúj-
tott ebben a tekintetben a konzervált indelek (=inszerciók és deléciók) felfedezése és analízise számos
univerzálisan elterjedt fehérjében. Ezek a konzervált indelek tulajdonképpen a fővonal ismertető-
jegyei. Az ezekhez vezető genetikai eseményeket általánosan úgy tekintik, mint fontos evolúciós
elágazási pontokat, és ezek eloszlásának mintázata a fajokban értékes információkat szolgáltat a fajok
elágazásának sorrendjéről, és a különböző törzsek közötti kapcsolatról.
Az indel modell alapján felállított bakteriális filogenetikus fa látható a 3.1.3. ábrán (lásd még a 64.
oldalon).
Egy ilyen indelmintázat-példa található az RNS-polimeráz β-alegységén (RpoB). Ez a magja az
RNS polimeráznak, és minden fajban univerzálisan konzerválódott. Az RpoB homológok a különböző
proteobakterekben éppúgy, mint az Aquifex, Klamidiák, Verrucomikrobák, Planctomyceták és BCF
esetében egy nagy inszertet tartalmaznak a fenti fajokra jellemző konzervált régióban, amely inszertet
a többi törzs (Spirohéták, Deinococcus-Thermusok, Chloroflexik, Fusobaktériumok, Actinobacterok,
Thermotogák, Firmicutesek) nem tartalmazza. Az inszert hiánya az archeák (ősbaktériumok) RpoB
homológjaiban egyértelműen arra utal, hogy amely baktériumokból hiányzik, azok az ősibbek. Fajok
közötti eloszlásmintázata alapján ez az inszert valószínűleg az előbbiek közös ősénél keletkezett, így
ezek (proteobakterek, Aquifex, Klamidiák, Verrucomikrobák, Planctomyceszek, és BCF) később
ágaztak le a főágról.
Hasonlóan ehhez egy másik prominens indelt azonosítottak a DNS-giráz B-alegységén. Ebben az
esetben egy konzervált, több mint 150 aminosavból álló inszert található a különböző proteobakterek
és Aquifex, Klamidiák, Verrucomikrobák, valamint Planktomiceszek DNS-giráz B alegységén, de nem
található meg a többi baktériumtörzs – köztük a BCF – tagjai között. Ez az inszert az archeák homológ fe-
hérjéi között sem található meg. Az Rpo és a giráz fajok közötti eloszlás-mintázatainak összehasonlí-
tása alapján megállapítható, hogy a giráz indelje az evolúció későbbi fázisában keletkezett, a Fibro-
bakterek-Chlorobi-Bakteroidák elágazása után. A fajok közötti eloszlás-mintázat alapján ez a két indel
egyértelművé teszi, hogy a Fibrobakter-Chlorobi-Bakteroidák (BCF) csoportok a Spirohéták,
Deinococcus-Thermus, Chloroflexek, Actibobakterek, Thermotogá, és a Firmicutes törzsek után, de a
proteobaktériumok, Aqificales, Klamidiák, Verrucomikrobiák és Planktomiceszek előtt ágaztak el. A
fenti két indelhez hasonlóan számos más fővonalismertető-jegyet fedeztek fel további univerzálisan
elterjedt fehérjékben, amely ismertetők segítik a baktérium- törzsek elágazási sorrendjének megértését.
Az indelek fajok közötti eloszlási mintázata alapján majdnem minden baktériumcsoportot egyértelműen el
lehet különíteni, és lehetséges logikusan kikövetkeztetni a közös őstől való elágazásaik sorrendjét.

3. Az ipari mikroorganizmusok és technológiák tárgyalása törzsrendszertanba illesztve 61
© Németh Áron, BME © www.tankonyvtar.hu
Az indelmodell megbízhatósága és prediktív lehetőségei
A fővonal indeleinek többsége, amelyeken az elágazási mintázatok alapulnak 1997 előtt kerültek
felfedezésre, amikor még a szekvencia adatok igen korlátozottak voltak, és a megszekvenált genomok
száma 10 alatt volt.
Ennek ellenére ezen indelek mindegyike igen specifikus előrejelzéseket tett (vagy tesz) az elő-
fordulásukkal és hiányukkal kapcsolatban a különböző baktériumtörzsek fajaiban. Ha ezek az
elágazási minták megbízhatóak, és ezek az indelek egy ősi vonalban egy adott evolúciós stádiumban
jelentkeztek, akkor minden feljebb fekvő bakteriális törzs fajainak tartalmaznia kell az adott indelt,
míg minden más csoport fajainak, amelyek az inszerciós pont előtt ágaztak el (azaz az inszerciós pont
alatt fekszenek) hiányzik ezen indelje.
Ha a megállapított indeleloszlás a különböző genomokban szorosan követi a modell alapján
elvártat, akkor a megállapított elágazások megbízhatóak. Ha ezek az indelek függetlenül keletkeztek,
vagy az őket tartalmazó gén gyakran került laterális átadásra, akkor az indelek jelenléte vagy hiánya a
különböző fajokban nem követi az elvárt mintázatot. A modellt számos indelen tesztelték különböző
teljesen feltárt genomokban, és megfelelően követték a modellt az eredmények.
A bakteriumtörzsek konzervált indelek alapján származtatott elágazási sorrendje néhány kivétellel
megegyezett a 16S rRNS alapú filogenetikus fákkal, akárcsak számos más fehérje/gén alapú
filogenetikus fákkal. A leggyakrabban publikált fákon, a Thermotoga, Deinococcus-Thermus, Ciano-
baktériumok és a Zöld-nem-kén-baktériumok csoportja korai elágazást mutat, míg mások (mint például
a Proteobacterek és Chlamydiák, valamint a Bakteroides-Chlorobi csoportok) később ágaznak le. Egy
új filogenetikus tanulmány [28] 31 általánosan konzervált fehérje szekvenciájának alapján szintén
határozottan állítja, hogy a Firmicutes a legmélyebb (legelső) elágazási vonal (3.1.3. ábra).
A Firmicutes és Actinobacterek korai elágazását éppúgy, mint a Thermotogákét, Deinococcus-
Thermusokét is alátámasztják a gének sorrendjének elrendeződései a bakteriális genomokban. Lake és
munkatársainak [29] tanulmánya azt is határozottan állítja, hogy a bakteriális fa gyökere nem a Gram-
negatív (vagy diderm) baktériumoknál van. Egy figyelemre méltó különbség a jelenlegi és a 16S rRNS
alapú filogenetikus fák között az Aquifex-ek késői elágazása az újabb tanulmányokban. Ez az anomália
az Aquifex rRNS nagy G+C-tartalma miatt van, ami számos hipertermofil szervezet közös
tulajdonsága, így emiatt az Archeák közé csoportosították, és az rRNS- fán korán elágaznak.
Az indelanalízis alapján levezetett elágazási sorrend a főbb szerkezeti különbségekkel is
összhangban van a baktériumoknál (vagy prokariótáknál). A baktériumokat két csoportra osztják attól
függően, hogy egy membránnal burkoltak (monodermák), vagy két különböző, a periplazmás tér által
elválasztott membránnal (didermák) rendelkeznek. Ez a két csoport durván megfelel a Gram-pozitív és
Gram-negatív csoportosításnak. A konzervált indelek alapján is indokolt ez a szerkezeti elkülönítés, és
azt is kimutatta, hogy a monodermák (Gram-pozitívak) az ősibbek. A levezetett elágazási sorrend a
Deinococcus-Thermus csoportot egy köztes pozicióba helyezi a Gram-pozitívak és negatívak között.
Ez az elhelyezés összhangban van a Deinococcusok egyedülálló szerkezetével, ami egy vékony
peptido-glükán-réteget tartalmaz, és így pozitívan festődnek a Gram-festéssel, de belső és külső
membránnal is körül vannak véve, hasonlóan a legtöbb Gram-negatív baktériumhoz. Ezek a
megállapítások jelzik az evolúciós átmenetet a Gram-pozitívaktól a negatívokig. Így a ritka véletlen
genetikai változások (RGCs: Rare Genetic Changes) alapján felállított bakteriális filogenetika képe jó
egyezést mutat a genotípus és fenotípus között, ami arra utal, hogy megbízható az indelmodell.

62 Ipari mikrobiológia
© Németh Áron, BME © www.tankonyvtar.hu
Coccusok Pálcák
Gram
+1.
Gram -
aerob
2. Gram
- Fak.
anaerob
3.
Gram -
anaerob
4.
Gram +
endospórás
5.
Gram +
spórátlan
6. Gram +
spórátlan
szabálytalan alakú
1. Spirális
és hajtott
bakt.
2.
Spirohéták
3.
Riketsia
félék
4.
Mycoplasma
félék
1.
Lactobacillus
nemzetség
2.
Listeria
nemzetség
1.
Corynebacterium
nemz
2.
Actinomiceták
3.
Streptomiceták
4.
Mycobaktériumok
5.
Nocardia
1.
Enterobacter
család
2.
Nem
enterobacterek
1.
Escherichia
nemzetség
2.
Salmonella
nemzettség
7.
Proteus
nemzettség
1.
Vibrio
nemzettség
2.
Haemophilus
nemzettség
1.
Bacteroides
nemzettsg1. Bacillus
nemzettség
(aerob)
2. Clostridium
nemzettség
(anaerob)
1.
Pseudomonas
nemzettség
1.
Staphylococcus
nemzettség
2.
Micrococcus
nemzettség
3.
Streptococcus
nemzettség2.
Legionella
nemzettség
Gram (-)
baktériumok
(1. Divízió)
Gram (+)
Baktériumok
(2. Divízió)
Merev sejtfal
nélküli baktériumok
(3. Divízió)
Ősbaktériumok
(4.
Divízió)
1.
Coccusok
2.
Pálcák
3.1.1. ábra: Bergey-féle bakteriális rendszertan

3. Az ipari mikroorganizmusok és technológiák tárgyalása törzsrendszertanba illesztve 63
© Németh Áron, BME © www.tankonyvtar.hu
3.1.2. ábra: Filogenetikus fa a 3 domén elmélet alapján [162]

64 Ipari mikrobiológia
© Németh Áron, BME © www.tankonyvtar.hu
3.1.3. ábra: Bakteriális filogenetikus fa az indelmodell alapján [27]

3. Az ipari mikroorganizmusok és technológiák tárgyalása törzsrendszertanba illesztve 65
© Németh Áron, BME © www.tankonyvtar.hu
3.2. Ipari szempontból fontos (ős)baktériumok
A 3.1.3. ábra sorszámozása szerint tárgyaljuk az egyes ágait a bakteriális filogenetikus fának az
alábbiakban. Ennek könnyebb követhetősége érdekében a rendszertant lehet tanulmányozni a 3.2.1.
hyperpicture-ön.
3.2.1. hyperpicture: Rendszertan
1. Archeák
Az ősbaktériumokat széles körben úgy tekintik, mint az élet 3 fő doménjának egyikét, noha eredetük
vita tárgya. Korábban úgy gondolták, hogy csak extrém környezetben élnek, mint extrém forróság
vagy forró és savas, extrém sós vagy nagyon savas, illetve lúgos környezetben, de újabb tanulmányok
bemutatták, hogy széles körűen elterjedtek különböző környezetekben is. Az archeák metanogéneket
is tartalmaznak, amelyek szigorúan anaerob és gyakran termofil környezetben élnek, és amelyek
egyedül képesek arra, hogy a metabolikus energiájukat metanogenezisből redukcióval nyerjék. A 16S
rRNS-fán (3.1.2. ábra) és számos más gén/fehérje szekvenicán alapuló filogenetikus fán az ősbakté-
riumfajok ága határozottan elkülönül minden más szervezettől. Ráadásul, számos további tulajdonság
(mint például az elágazó láncú éterkötésű membrán lipidek, a sejtfalból hiányzó peptidoglükán, sajátos
RNS polimeráz alegység mintázat, módosult bázisok jelenléte a tRNS-ben, egyedi DNS polimeráz) is
azt jelzi, hogy szükséges az archeák külön definiálása. Ennek ellenére ezen tulajdonságok némelyike
nincs meg mindegyik ősbaktériumban, vagy más eukarióta, illetve termofil szervezetben pedig
megtalálható.
Az archeák filogenetikus analízise két főcsoportra (vagy törzsre) osztásukhoz vezetett: Crenarchaeota
és Euryarchaeota. E két csoport fajai – különösen az Euryarchaeotáknál – igen különböznek metabo-
lizmusukban és fiziológiájukban. A metabolikus és fiziológiai, illetve más egyedi tulajdonságaik
alapján az Euryarchaeotáknál 5 funkcionálisan elkülöníthető csoportot határoztak meg: metanogének
(3.2.2. ábra), szulfátredukálók, extrém halofilek, sejtfal nélküliek és extrém termofil kénmetabolizáló
baktériumok. Némely ezen csoportok közül polifiletikus a különböző filogenetikus családfákon. Ezen
különböző csoportoknak unikális gén- vagy fehérjekészlete – ami megkülönböztetné őket minden
mástól – még meghatározásra vár. Az utóbbi években számos archea teljes genomját megszekve-
nálták, és az összehasonlító analízisük értékes információkat szolgáltatott a különböző gének/fehérjék
tekintetében, amelyek az Archeákon belül elkülönítik a csoportokat, valamint a baktériumokkal és
Eukariótákkal való kapcsolatot is meghatározzák.

66 Ipari mikrobiológia
© Németh Áron, BME © www.tankonyvtar.hu
3.2.2. ábra: Methanosarcina metanogén ősbaktérium [178]
Az ősbaktériumok egyik legújabb, 31 jelző fehérje alapján felállított filogenetikus fáját mutatja a
3.2.3. ábra (lásd még a 93. oldalon).
Az ősbaktériumok extrém élőhelyeken is előfordulnak, ezért az enzimeik ezen körülmények
között is aktívak, így noha eddig csak néhány ősbaktérium enzime jutott el a piaci megjelenésig,
hatalmas potenciált rejtenek magukban, ezért intenzíven kutatják őket. A különleges élőhely miatt az
enzimeik stabilak magas hőmérsékleten, extrém alacsony/magas pH-n, nagy nyomáson, esetleg
mindezt magas sótűréssel, oldószer-toleranciával, fémtoleranciával kombinálva [31]. Az extrémofilek
enzimeinek (extremozimok) használatának számos előnye van, például csökkent fertőződésveszély,
jobb anyagátadási sebesség, alacsonyabb viszkozitás, magasabb szubsztrátoldhatóság. Néhány érdekes
tulajdonságot mutat be a 3.2.1. táblázat.
3.2.1. táblázat: Az ősbaktériumok különleges képességei
Tulajdonság Tartomány Faj
nitrátredukáló,
kemolitoautotróf,
termofil
<113°C
Pyrolobus fumarii
metanogén <110°C
<0°C
Methanopyrus kandleri
Methanogenium frigidum
termoacidofil 60-90°C
pH 0-4
aerobok
anaerobok
(kevésbé savas)
Acidianus, Ferriplasma,
Picrophilus, Sulfolobus,
Thermoplasma
Methanogenium frigidum
Thermoproteus, tenax
Methanothermus fervidus
termoaalkalofil 85°C
pH 8,5-9
Thermococcusok
barofil
(nyomáskedvelő)
Paleococcus ferrophilus
Thermococcus barophilus
barofil
(nyomáskedvelő)
20-30 MPa [32] Paleococcus ferrophilus
Thermococcus barophilus
Wolfram mint kofaktor
tg=37min
keményítőtranszport-fehérjék
20 MPa
103°C
Pyrococcus furiosus
A leggyakrabban és legintenzívebben a magasabb hőmérsékleten is stabil és aktív enzimeket
kutatják. Ezidáig nem találtak olyan faktort, amely egyedül érhetné el az enzimek termostabilitását,
ugyanakkor leírtak már néhány jelenséget, amelyek hozzájárulnak az archeák enzimeinek hőstabilitá-
sához: magasabb hidrofóbicitás, és felületi töltöttség, nagyobb számú ionpár és hidrogénkötés,

3. Az ipari mikroorganizmusok és technológiák tárgyalása törzsrendszertanba illesztve 67
© Németh Áron, BME © www.tankonyvtar.hu
csökkent rugalmasság és felületi hurokméret, kisebb felszín/térfogat arány, kevesebb hőérzékeny
aminosav és csonka amino- és karboxiltermináns.
Noha a fenti különleges képességek vagy egy részük a modern genetikai eszközökkel nyilván
kiaknázhatók, egyelőre az extrém körülmények között is stabil aktivitást mutató enzimek forrásaiként
hasznosítják a (fehér)biotechnológia céljaira az ősbaktériumokat. Erre mutatunk be néhány példát az
alábbiakban. [31].
Nagy az ipari érdeklődés a szénhidrát-metabolizmus enzimei – különösen a glikozil hidrolázok
– iránt. Mivel minden keményítőbontó technológiában a szokásos fiziológiai hőmérsékletnél
magasabbat alkalmaznak, a dextrinek, trehalózok, glükóz és fruktóz gyártói profitálhatnak a
termostabil keményítőbontó enzimekből. A Pyrococcus extracelluláris α-amilázai 120°C 4h hőkezelés
után is aktívak (vagy 130°C-os kezelés után is), tehát ideális jelöltek a keményítőipar számára. Az
archeák pullulanázai is 90-100°C között aktívak, és szubsztrát, valamint Ca2+ ionok hiányában is
megőrzik az aktivitásukat. A termoacidofil archeák glükoamilázai 90°C-on és pH=2-n is aktívak
(Thermoplasma acidophilum, Picrophilus torridus és P. oshimae)
A trehalóz (α,α-1,1-glükóz dimer) fontos enzim, antitest, vakcina, hormonstabilzáló szer, elő-
állításában a Sulfolobus-enzimek 75°C-on jól hasznosíthatók.
A ciklodextrinek előállítására a ciklodextrin-glikoziltranszferázokat (CGTáz) használják, amelyek
közül kiemeledő a Thermococcus fajoké a 100°C feletti optimális aktivitással (keményítőből külön-
böző ciklikus dextrinek előállítása), így a keményítőbontás és -ciklizálás egylépcsőben megvaló-
síthatóvá válik.
A cellulózbontó enzimkomplex tagjai (endoglükanáz, exoglükanáz, β-glükozidáz) is fellelhetők
termostabil formában az archeák enzimkészletében. A Pyrococcus furiosus β-glükozidáza 103°C-on is
aktív. A P. horikoshii homológ enzime szerves oldószerben is stabil és aktív, ezért heteroszaharidok
előállítására használható.
Endo-β-1,4-xilanázok segítségével történik a hemicellulóz xilánfrakciójának lebontása. Míg az
archea Pyridictum abyssi xilanáza az egyik legmagasabb hőmérsékleti optimummal (110°C)
rendelkező, a Thermococcus ziligii xilanáza igen specifikus magas hőmérsékleten is (cellulózt nem
bontja, csak a xilánt), ezért a biobleaching számára rejt nagy lehetőségeket.
Hőstabil kitinbontó endo és exo kitinázokat is termelnek az archeák. A Thermococcus
chitonophagus kitináza 120°C 1h kezelést is 50%-os veszteséggel kibír, míg a Pyrococcus furiosus
termoaktív kitinázainak segítségével képes kolloid kitinen, mint egyedüli szénforráson növekedni.
A fehérjebontó enzimek talán a legrégebb óta keresett és piacra dobott biokatalizátorok, mert a
mosószer- és detergensipar igen nagy mennyiségben használja őket. A magas hőmérsékleti optimum
az alkalmazások szempontjából kedvező, mivel a magasabb hőmérsékletű mosás hatékonyabban
tisztít. Ennek elenére az ősbaktériumok számos proteáza közül csak néhányat írtak le eddig
részletesen. Többségük az alkalikus szerin-proteázok családjába tatozik (Thermococcus setteri), de
találtak már tiol proteázt (Thermococcus kodakarensis) és savas protázt, metalloprotázt is Pyrococcus
furiosusban.
A biotechnológia fejlődésével egyre elterjedtebbé vált a PCR technika, ahol a magas hőtűrésű
DNS- polimerázok preferáltak. Az archeák közül ismét a Thermococcus és Pyrococcus törzseket írták
már le hiba nélkül másoló termoaktív DNS-polimeráz-forrásként. Újabban az ősbaktériumok DNS-
polimerázaival egészítik ki a Taq aktivitásokat, mert így csökkenthető a hibás amplifikációk száma.
Az archeák DNS-függő enzimeit a genom szekvenálásokhoz is gyakran alkalmazzák, DNS-ligázaikat
pedig analitikai célokra használják.
A vegyipar számára fontos az optikailag tiszta alkoholok mint építőkövek előállítása. Ehhez
alkohol-dehidrogenázokat (ADH) használnak, amelyek hőstabil homológjai megtalálhatók az
archeákban: Sulfolobus solfataricus (3.2.4. ábra) ADH-a NAD függő és Zn-et is tartalmaz. A
legstabilabbak a pyrococcusok ADH-ai (felezési idő ~150h 80°C-on).

68 Ipari mikrobiológia
© Németh Áron, BME © www.tankonyvtar.hu
3.2.4. ábra: Sulfolobusvírus-fertőzéssel
Szintén az optikailag tiszta vegyületek szintézisében töltenek be fontos szerpet az észterázok.
Némely archea (Aeropyrum pernix, Pyrobaculum calidifontis) eredetű észteráz igen termoaktív, és
még vízzel elegyedő oldószerekben (acetonitril, DMSO) is stabil. A legstabilabb észterázok a
Pyrococcus furiosusból nyerhetők (felezési idő: 50 min 126°C-on).
A vegyipari szintézisek fontos lépései a C-C kötést kialakító reakciók, amelyeket az aldolázok és
a transzketolázok is katalizálnak. Az archeák közül a Sulfolobus solfataricus aldolázát írták már le
piruvát, illetve glicerinaldehid előállítására. A sztereoszelektív Sulfolobus solfataricus amidáza széles
szubsztrát spektrummal rendelkezik, és 95°C-on is még aktív.
Az enzimeiken kívűl az archeák további molekulái is rendelkeznek ipari felhasználással vagy
potenciállal: az ősbaktériumok lipidjei hőstabil liposzómák előállításához alkalmazhatók; Sulfolobus
törzsek antibiotikus hatású fehérjét is termelnek; chaperonjaik a rekombináns fehérjék hajtogatását
segítik. Mivel az ősbaktériumok eltérő kodonrendszert használnak a valódi baktériumoktól, ezért a
génjeik kifejezése bakteriális gazdaszervezetben igen nehézkes, emiatt az utóbbi időben archea-
expressziós rendszereket is létrehoztak Methanococcus és Sulfolobus fajokból.
2. Firmicutes
A csoport neve (=erős bőr) utal a sejtfal jelenlétére, amely a legtöbb nemzetség esetében Gram-pozitív
festésű, de némelyek esetében vékony külső membránnal takart, ezért Gram-negatívan festődik. A
törzs közös vonásaként – szemben a következő Actinobacterek törzzsel – a DNS-ük kis G+C-tartalmát
jelölték ki, ez alapján sorolják ide a nemzettségeket, amelyek között számos ipari jelentőségű akad
(Bacillus, Lactobacillus, Clostridium stb. fajok.) A nemzettségek azonban szinte minden más
tulajdonságban különböznek, mivel vannak pálcikák, kokkuszok, spirálisak, aerobok/anerobok,
endospórásak/spórátlanok is közöttük. Ezek azonban az egyes családokon belül sem eléggé konzervált
tulajdonságok, így nem megfelelőek a besoroláskor, ezért is használják ma már a real-time (valós
idejű) PCR technikát a DNS-alapú osztályozáshoz. A firmicutes rendszertana azonban eme diverzitás
miatt jelentős változásokon ment át és megy át most is. Egyelőre többnyire 3 osztályt különböztetnek
meg a Firmicutes törzsön belül: Bacillusok, Clostridiumok és a Mollicutes [27]. (3.2.2. táblázat, lásd
még a 93. oldalon)
Ipari szempontból a bacilli osztály bacilliales (kataláz pozitívak) rendjének legfontosabb tagja a
bacillusok nemzettsége, amelynek fajai között olyan fontosakat találunk, mint a Bacillus subtilis
(3.2.5. ábra), amelyet szinte a DNS felfedezése (Watson 1953) óta a Gram-pozitív baktériumok

3. Az ipari mikroorganizmusok és technológiák tárgyalása törzsrendszertanba illesztve 69
© Németh Áron, BME © www.tankonyvtar.hu
modellszervezeteként használnak fel, és 1959-ben volt az első sikeres géntranszfer egy Bacillus
subtilis törzsbe [58].
3.2.5. ábra: Bacillus subtilis [178]
A ’90-es évek közepére a biotechnológusok egyik „igáslováról” annyi információ állt rendelke-
zésre (fiziológiai folyamatok ((transzkripciós, transzlációs szabályozások)), tenyésztési és ipari fer-
mentációs tapasztalatok stb.), hogy érdemesnek tűnt belőle expressziós rendszert készíteni, amely
idegen fehérjék (ipari célú, gyógyászati célú, élelmiszercélú stb.) előállítására nagy léptékben alkal-
mas. Az ilyen expressziós rendszerek olyan módosított mikrobák (jelen esetben ez B. subtilis),
amelyek képesek nagy koncentrációban bioaktív formában (helyes elsődleges, másodlagos, harmad-
lagos szerkezettel) fehérjéket a fermentlébe extracellulárisan kiválasztani, miközben eredeti proteáz
aktivitásukat lecsökkentették (és ezáltal a termék lebontás csökkent), persze egyúttal az aminosav-
metabolizmust is szükségszerűen kiegészítették. Így mára valóban a legelőnyösebb és talán a
legelterjedtebb ipari expressziós rendszerré vált a B. subtilis, és olyan fehérjéket állítanak elő vele,
mint a termostabil keményítőbontó enzimek (α-amilázok, glükoamilázok, xilanázok), illetve a humán
interleukin-1 (ILE-1), amely az emebri immunválasz egyik központi citokin6 molekulája [59].
A bacillusok további fajai sem szégyenkezhetnek: A Bacillus coagulans (nevét a tej fehérje kicsa-
pásáról kapta) a tejipari túrógyártás oltóanyaga, illetve jó enzim- (pl.: α- és β-amiláz, xilanáz, celluláz,
alkalikus proteáz, lipáz stb.) és tejsavtermelő. Előbbiek jelentősége a hőstabilitás és különleges pH-
tolerancia miatt kiemelkedő, utóbbi képessége pedig azért jelentős, mert az 50-55°C-on végzett
tejsavfermentációkat nem kell sterilen vezetni, és így jeletős költségmegtakarítás érhető el, ugyan-
akkor hozam és végtiter tekintetében a szokásos (és közeli rokon) Lactobacillus törzsekkel is verseny-
képes [60]. A bacillusok alkalmazásának egyik iskola példája a Bacillus thüringiensis (vagy ékezetek
híjján: B. thueringiensis v. B. thuringiensis), amelynek endospórájának felszínéről endotoxikus
fehérjét izoláltak és klónoztak, mert a biokontroll során a rovarok ellen hatékony bio-irtószer (ipari
szúnyogirtás hatóanyaga). Az 1980-as évektől kezdve alkalmazzák ezt a biopeszticidet, és a ’90-es
évektől kezdve a génmódosított növények egyik nagy csoportját a BT (Bacillus thüringiensis Toxin)-t
kódoló és expresszáltató lepidoptera (lepkefélék) ellen önvédelemmel felszerelt gabonák adják
(Monsanto kukorica stb.) [61]. A Bacillus licheniformis FDA által is elfogadott, GRAS listán szereplő
biztonságos ipari és élelmiszer-ipari mikroorganizmus, amellyel számos heterológ génexpressziót
megvalósítottak, illetve carbohidráz- (főleg α-amiláz) és proteáz- enzimeket állítanak vele elő [62].
Számos patogén mikroba is található a bacillusok között, mint a Bacillus antraxis (Antrax vagy
lépfene) vagy B. cereus. A Bacillialacea mellett két további, főleg egészségügyi vonatkozású család, a
6 ↑ Gilman A, Goodman LS, Hardman JG, Limbird LE. Goodman & Gilman's the pharmacological basis of
therapeutics. New York: McGraw-Hill (2001). ISBN 0-07-135469-7
70 Ipari mikrobiológia
© Németh Áron, BME © www.tankonyvtar.hu
Staphylococcuceae (pl.: S. aureus) és a Listeriaák találhatóak meg (L. monocytogenes, 3.2.6. ábra) a
Bacilliales rendben [64].
3.2.6. ábra: Listeria monocytogenes [178]
A bacillusosztály másik fontos rendje a Lactobacilliales, azaz lactobacillusok. Ide tartoznak az
elsősorban egészségügyi szempontból érdekes Streptococcusok (pl. S. pyogenes (skarlát), S.
pneumoniae, S. bovis), illetve a bélflóra Enterococcusai (pl.: E. faecalis). Mindkét család a tejiparban
játszik szerepet. A S. thermophilus a közeli rokon Lactobacillus delbruecki subsp. bulgaricusszal
együtt joghurtok mikrobiótáját adja, és így élelmiszer-ipari felhasználása jelentős, emellett tejsav-
előállításra is használható [65]. Az Enterococcusok a mediterrán eredetű sajtokban fontos szerepet
játszana egyrészt a sajtok kedvező egészségügyi hatásának kifejtésében, valamint a készítés
technológiájában is [63]. E két kevésbé jelentős család mellett a Lactobacilliceae család tagjai a
Leuconostoc és Lactobacillus (3.2.7.) nemzettségek.
3.2.7. ábra: Lactobacillus acidophilus [178]
Ipari szempontból a Leuconostoc mesenteroides almasav (=2-hidroxi-szukcinát) termelési
potenciálja, valamint dextránszukráz enzimtermelő képessége figyelemre méltó. Utóbbi különösen
divatos a funkcionális élelmiszerek előállítása kapcsán, amely enzim segítségével az

3. Az ipari mikroorganizmusok és technológiák tárgyalása törzsrendszertanba illesztve 71
© Németh Áron, BME © www.tankonyvtar.hu
emésztőenzimeknek ellenálló glükooligoszaharidot szintetizál ez a mikroba [66]. A lactobacillusok
közé tartoznak a Pediococcusok is. Általában ezen tejsavbaktériumok homolaktikus tejsavas
erjesztését használják ki a káposzta savanyításakor. A Pediococcus acidilactici faj a bélflóra tagja, és
részben tejsavtermeléssel, részben pediocinek (pediococcusok által termelt baktericinek) segítségével
antagonizmussal szorítja vissza a fertőzéseket. Alacsony pH-n és magas hőmérsékleten (65°C) is túlél
[71].
Az Oenococcusok (3.2.8. ábra) közé kevés fajt sorolnak, viszont ezek a nevüknek megfelelően a
bor(alkohol)készítés (oenology – enológia) „munkalovai”. A borászati jelentőségük az ízek kialakítá-
sában jelentkezik (pl.: Oenococcus oeni), mivel a keményebb malonsavas (almasavas) ízt (ami a must-
ból ered) alakítják a puhább, teltebb, vajasabb tejsavas ízzé (malo-laktikus fermentáció). E vegyületek
aránya határozza meg a bor ízét.
3.2.8. ábra: Oenococcus oeni [178]
A Lactococcusok a tejgazdaságokban játszanak szerpet a tejtermékek és sajtok előállításánál. A
Lactococcus lactis sp. lactis (3.2.9. ábra) és a L. lactis sp. cremoris oltókultúra szerepét tölti be [72].
Általában homolaktikusak, és fő feladatuk a tej gyors lesavanyítása, ami megakadályozza a
megromlást okozó mikrobák elszaporodását.
3.2.9. ábra: Lactococcus lactis [178]

72 Ipari mikrobiológia
© Németh Áron, BME © www.tankonyvtar.hu
A Firmicutes 2. nagyosztálya a Clostridium-félék osztálya, amely fajokban és nemzettségekben is
igen gazdag [148].
Ipari jelentősége lehet a Sarcina ventriculinak, amely a mikrobiális cellulóztermelés egyik
képviselője. A Clostridium nemzetségből ipari jelentősége lehet a C. thermocellumnak, mivel
cellulózalapon etanolt fermentál, már iparilag alkalmazzák a C. botulinum-ot (3.2.10. ábra), amelynek
endotoxinja idegméreg, de kis koncentrációban botox néven forgalmazzák plasztikai sebészeti céllal.
3.2.10. ábra: Clostridium botulinum
Végül a C. acetobutlicum, amellyel óriási méretekben német technológiával aceton-butanol-etanol
(ABE) femetnációkat végeztek. A Clostridiumok osztályában szinte minden anyagcserére és életfor-
mára találunk példát: vajsavas erjesztő a Butyrivibrio, ecetsavtermelő az Acetobacterium, nitrogén-
fixáló az Anaerococcus, propionsav termelő a Propionispira, borostyánkősav- (szukcinát) termelő a
Succiniclasticum, szulfátredukáló Desulfosporosinus, dehalogenidező (halogenidekkel szubsztituált
szennyezések bontása) Desulfitobacteriumok.
A Firmicutes 3. csoportja a Mollicutes, ahol ipari potenciál alig található, viszont az eddig ismert
legkisebb baktériumokat (Mycoplasma genitalium) idesorolták.
3. Actinobacterek
Az Actinobacterek törzsét Gram-pozitív baktériumok alkotják, amelyeknek nagy a G+C-tartalma
(>55%). Ez a csoport 39 családot és 130 genust tartalmaz, így ez a legnagyobb törzs a baktériumokon
belül. Igen széles morfológiai tartománnyal rendelkeznek a coccusoktól (pl.: Micrococcus) a pálcika-
gömbökön (pl.: Arthrobacter) keresztül, a töredezett hifán át (pl.: Nocardia) az állandó és erősen
differenciált, elágazó micéliumig (pl.: Streptomycesek) bezárólag. Számos Actinobacter spóraképző,
amely spórák a mozgó zoospóráktól a különleges propagulákig terjednek. Élettanilag is nagyon
különböznek, amint az kitűnik számos extracelluláris enzimeik és több ezer metabolitjuk termeléséből
(beleértve az antibiotikumokat is). Az Actinobacterek (különösen a Streptomyces nemzetség) a fő
antibiotikum-termelők a gyógyszeriparban. Ennek ellenére néhány Actinobacter fontos humán- és
állatpatogén. Például a Mycobacterium tuberculosis tuberkulózist okoz, Corynebacterium diphteriae
pedig diftériát, a Propionobacterium acne az akne (bőrhiba) okozója. Az Actinobacter fajok
széleskörűen elterjedtek mind a szárazföldi, mind pedig a vizes ökoszisztémákban, és a bioszanyagok
(cellulóz, kitin) lebontásában és visszaforgatásában (szénkörforgalom) fontos szerepet játszanak. Noha
az Actinobacterek egy elkülönített csoportot képeznek a 16S rRNS-fán, nincs más megbízható
biokémiai vagy molekuláris tulajdonságuk, ami saját jellegzetessége lenne a csoport tagjainak. Ennek

3. Az ipari mikroorganizmusok és technológiák tárgyalása törzsrendszertanba illesztve 73
© Németh Áron, BME © www.tankonyvtar.hu
ellenére az újabb genomszekvencia-analízisek számos konzervált közös és egyedülálló indelt találtak,
amelyek vagy minden Actinobacterra jellemzőek, vagy csak néhány alcsoportjukra, de fontos
információkat szolgáltattak a rendszertanuk, leszármazásuk és egyedülálló biokémiai és élettani
tulajdonságaik megértéséhez.
Az Actinobacterek csoportja igen gazdag fajszámban (3.2.3. ábra). A legnépesebb rend az
actinomycetes rendje (ezen kívül még az Actinobacterek osztályába tartoznak a Rubrocacterek,
Sphaerobacterek, Cariobacterek, Acidimicrobiae-k, és a Bifidobacterek), amelynek tagjai mind
humánegészségügyi, mind ipari szempontból jelentősek.
Az actinomycesek rendje további, igen különböző alrendeket tartalmaz: Micrococcineae,
Corynebacterineae, Propionibacterineae, Pseudonocardineae, Frankinae, Micromonosporineae,
Streptomycineae, Catenulisporinea, Actynomycetaceae és Glycomycetaceae.
A Micrococcusok (Micrococcineae) a természetben széleskörűen megtalálhatók (levegőben, talajban,
porban, vízben), Gram-pozitív gömb alakú baktériumok, általában tertádként fordulnak elő. Erős
sejtfallal rendelkeznek, amely a biomasszatömeg 50%-át is kiteheti. Genomjuk G+C-tartalma nagy, és
plazmidot is gyakran hordoznak, amelyek hasznos tulajdonságokat kódolnak. A M. luteus sárga, a M.
roseus rózsaszín telepeket képez malátakivonatos agaron. Előbbi a toxikus szerves anyagokon (pl.:
piridin) riboflavintermelésre képes. Egyes mikrokokkuszok között csak 50%-os a genomazonosság,
ezért valószínűleg újra kell őket osztályozni a 16S rRNS alapján. Többségük mezofil, de pszichrofil is
akad köztük (M. antarcticus). Habár nem spóraképző, mégis nagyon túlélő fajta (több 10 000 évet is
kibír). Általában szaprofiták vagy kommenzalisták, de opportunista patogének is előfordulnak,
amelyek főleg a legyengült immunrendszerű szervezeteket támadják. Nehéz igazolni a
patogenitásukat, mivel a normál emberi bőrflóra tagjai.
Ipari felhasználásuk többnyire a detoxifikálásra korlátozódik, mivel számos toxikus anyag
lebontására képesek, egyes törzsekkel azonban hosszú szénláncú (C21-C34) alifás szénhidrogént
állítanak elő.
A M. antarcticus faj hidegkedvelő, így 30°C-os hőmérsékleti optimummal rendelkező alfa amliázt
izoláltak belőle, amely 12°C-on is aktív. M.flavust eleveniszapos szennyvíztisztítóból izolálták, és
aerob, heterotrófként írták le. Glicerint, trehalózt és dextrint hasznosít szénforrásként, de glükózt,
fruktózt, laktózt, arabinózt, xilitet, inozitot, ramnózt nem. A M. luteus sárga, obligált aerob, nem
patogén baktérium, amely általában egysejtűként (csoportosulás nélküli formában) található meg.
Koaguláz negatív, bacitracinérzékeny mikroba. A glutaminázenzime ipari szempontból érdekes
(sótűrő) [35] A M. lylae a haltermékek megromlásáért felel, lehet humán patogén is. A M.
mucilaginosusról (3.2.12. ábra) 1981-ben kiderült, hogy a Staphylococcusokhoz jobban hasonlít, ezért
Staphylococcus salvarius néven szerepelt a kis G+C-tartalmú Gram-pozitív baktériumok között, majd
2000-től a Stomatococcus mucialginosus, később a Rothiák közé került, és a száj mikroflórájának
úttörőjeként a plakkok és így a fogszuvasodás kialakulását segíti.

74 Ipari mikrobiológia
© Németh Áron, BME © www.tankonyvtar.hu
3.2.12. ábra: Microccus mucilaginosus [178]
A Micrococcus roseus RJ6 törzset szteroidkonverzióban lehet hasznosítani, mivel egy
plazmidjának termékei a koleszterin -> ADD és AD átalakítást tudják elvégezni [33]
A M. glutamicus törzsnek többféle ipari felhasználása van, a diazo festék eredetű szennyezések
lebontásától [35] a lizin termelésen át [36] a koimmobilizált glutaminsav termeléséig [37]. Utóbbi
potenciált és a termelőtörzseket napjainkban is kutatják és fejlesztik [38].
A mikrokokkuszok egyik iparban is használatos közeli rokonai az Arthrobacter család tagjai.
Jellemzően talajlakók, aerobok, és az exponenciális fázis során pálcikák, a hanyatló fázisban pedig
kokkuszok formájában jelennek meg (reverzió: pálcikák osztódása kokkuszokká). Jól tűrik a
kiszáradást és táplálékhiányt is. Az Arthrobacter simplex szteroidkonverziós képességeit már
tárgyaltuk (2.1. fejezet: Actinobacter-ek által végzett szteroid konverziók), azonban az elmúlt 100
évben többször is új besorolást és nevet kapott (Arthrobacter simplex, Corynebacterium simplex,
Pimelobacter simplex). Jelenleg a szintén Actinomiceta Propionibaktériumok Nocardioides családjába
sorolják Pimelobacter simplex néven, tehát nem a mikrokkuszok Arthrobacter családjába. Az
Arthrobacterek egyik kiemelkedő tagja az A. chlorophenolicus (3.2.13. ábra), amely extrém lebontó-
képességei révén (piridon, klórofenol, Cr(VI) redukciója Cr(III)-má stb.) bioremediációkban
használatos.
3.2.13. ábra: Arthrobacter chlorophenolicus [178]
Az A. mobilis savas ureázát fermentált üdítők karbamidtartalmának, amely a karcinogén
etilkarbamát prekurzora, csökkentésére lehet használni (pl.: japán rizspálinka, azaz szaké esetében)
[44]. Az A. oxydans (újabban A. nicotinovorans [45], neve élőhelyére utal: dohány növényen
nikotinlebontó aktivitás) is iparilag fontos szteroidkonverziós képességekkel rendelkezik, mint azt már
szintén tárgyaltuk (2.1. fejezet: Actinobacterek által végzett szteroidkonverziók).

3. Az ipari mikroorganizmusok és technológiák tárgyalása törzsrendszertanba illesztve 75
© Németh Áron, BME © www.tankonyvtar.hu
A mikrokokkuszok másik érdekes és élelmiszeriparban használt családja a brevibaktériumoké,
amelyek a lábszagért is felelnek (B. linens), és egyes sajtok (Limburger, Port-du-Salut, pálpusztai)
erjesztéséért is. Ezek a mikrobák ugyanis képesek az izzadt lábon az elhalt hámsejtek lebontására
metántiol keletkezése közben, és a szintén jelen lévő propionsav-baktériumok termelte
ecetsavval+propionsavval együtt alakul ki a kellemtlen szag (erre még izovaleriánsav-termeléssel a
Staphylococcus epidermis is rásegít [164]). A sajtaroma kialakításkor pedig a triptofánbontó
képességük játszik fontos szerepet [46]. A pálpusztai sajt felszínére (aerob) kenik az oltóflórát, amely
rózsaszínes (=rúzsflóra), nyálkás bevonatot képez, és igen erős szagot áraszt. Ez a sajt egy viszonylag
fiatal (kicsit több mint 100 éves) hungarikum [47].
A Corynebacteriumok (Corynebacteriaceae) Gram-pozitív, pálca alakú, spórátlan, kataláz pozitív,
aerob vagy fakultatív anaerob, csillótlan, egyenes vagy enyhén hajlott baktériumok, széleskörűen
elterjedtek a természetben, többségük ártalmatlan. Vannak köztük iparban is alkalmazottak (C.
glutamicum) és patogének, mint például a diftéria kórokozója (C. diphteriae (3.2.14. ábra), amely
patogenitását (azaz diftériatoxin-termelő képességét) egy baktriofágtól kölcsönzi). Különös
ismertetőjük, hogy polifoszfát formájában inclusion-bodyban energiát tárolnak (=metakromatikus
granula), amelyeket a metilén-kék festék a szokásos kékfestés helyett rózsaszínűre fest. Az alakjuk
változó is lehet a körülményektől függően (=pleomorf).
3.2.14. ábra: Corynebacterium diphteriae
A Corynebacteriumok sejtfaluk alapján is jól elkülöníthetőek. Mezo-diaminopimelinsavat
tartalmaz a murein hálójuk számos arabinogalaktán ismétlődéssel, csakúgy mint corinemikolsavval
(=mikolsav 22-26 szénatommal), amelyeket diszaharid kapcsol össze (mAGP – mikolil-AG-
peptidoglikán).
Tenyésztésük viszonylag lassú, még kiegészített tápközegen is, és valamennyi törzs biotint (H-
vitamin) igényel. Némelyik törzs tiamint (B1 vitamin) és PABA-t (B10 vitamin) is igényel. Viszonylag
kicsi, szürkés telepeket képeznek, többnyire átlátszóak, de opálos a közepük. Löffler tápközegen
(pepton, glükóz, NaCl, marhahúskivonat, lóvérszérummal (víz helyett) feltöltve) sárgás, fehérbe hajló
színűek a telepek.
Ipari hasznosításuk kiterjed az aminosavak, nukleotidok és más táplálkozási faktorok előállítására,
valamint szénhidrogének lebontására, sajtérlelésre és enzimtermelésre. Egyes törzsek antibiotikus
hatású bakteriocint termelnek. A C. glutamicum a nevéből is láthatóan a glutaminsavgyártásra
alkalmas, és az anyagcsereutak módosításával lizin és treonin aminosavakat is elő lehet vele állítani.
Szintén ezzel a törzzsel sikerült az emberi epidermális növekedési faktort (EGF) aktív formában
előállítani, ami megnyitotta az utat további humánfehérje-expresszáltatásához C. glutamicummal. A
fehérjeexpressziót vagy az általános kiválasztó úton (Sec –> durva felületű endoplazmotkus retikulum

76 Ipari mikrobiológia
© Németh Áron, BME © www.tankonyvtar.hu
-> Golgi), vagy az iker-aginin (Tat –> aktív formában membránon keresztül) transzlokációs úton lehet
elérni.
Egyes Corynebacteriumok lipofilek, azaz zsírban szeretnek szaporodni. A nemlipofileket fermentatí-
vakra és nemfermentatívakra szokás osztani.
Fermentative Corynebacterium-ok:
Corynebacterium diphtheriae group
Corynebacterium xerosis and Corynebacterium striatum
Corynebacterium minutissimum
Corynebacterium amycolatum
Corynebacterium glucuronolyticum
Corynebacterium argentoratense
Corynebacterium matruchotii
Nem fermentative Corynebacterium-ok:
Corynebacterium afermentans subsp. afermentans
Corynebacterium auris
Corynebacterium pseudodiphtheriticum
Lipofilek:
Corynebacterium jeikeium
Corynebacterium urealyticum
Corynebacterium afermentans subsp. lipophilum
Corynebacterium accolens
Corynebacterium macginleyi
CDC coryneform groups F-1 and G
A Corynebaktériumok legközelebbi rokonai a Mycobacteriumok (Mycobacteriaceae), ideértve a
szteroidkonverzióban használt M. phlei és a tuberkulózis kórokozóját (M. tuberculosis), valamint a
lepra kórokozóját (M. leprae) is. Egy részük lassan szaporodik (főleg a kórokozók: marhatuberculózis
(M. bovis), humán tbc (M. tuberculosis 3.2.15. ábra), lepra M. leprae)), mások pedig gyorsan (M.
phlei, M. smegmatis). A mikobaktériumok se nem Gram-pozitívak, se nem negatívak, hanem a Gram-
festésre furcsán reagálnak, mivel a festékek savas/alkoholos lemosásának ellenállnak (acid-fast). Ezen
képességüket és még további néhányat (pl.: antibiotikumrezisztenciák egy része, csak a makrolid
antibiotikumok hatnak) a különleges mikolsavas sejtfaluknak köszönhetik.
3.2.15. ábra: Mycobacterium tuberculosis
A Mycobacteriumok közeli rokonai a Nocardiák. A normál szájflórában is előfordulnak, de
nokardiózisos tüdőgyulladást (N. asteroides, N. brasiliensis) is okozhatnak. A N. lactamdurans ipari
fermentációban termeli a sertéstenyésztés antibiotikumát, az efrotomicint [50]. A nokardiák bioaktív
komponensek előállításáról is ismertek, így a nocardicin [55] (β-laktám vázas antibiotikum, N. uni-
formis termeli), tubelactomicin A [56] (makrolid antibiotikum mikrobaktériumok ellen (=rokonok(!)),
N. vinacea termeli) és a brasilicardin A [57] (triciklusos terpenoid immunszupresszáns, N. brasilensis
termeli) ipari előállításában részt vesznek [51]. A N. farcinica és N. globerula iparilag értékes
amidázok forrása, amelyeket többnyire rekombinánsan állítanak elő, és az akril-amid előállításánál
használnak [52].

3. Az ipari mikroorganizmusok és technológiák tárgyalása törzsrendszertanba illesztve 77
© Németh Áron, BME © www.tankonyvtar.hu
A nokardiák legközelebbi rokonai a Rhodococcusok. Néhányuk patogén, de többségük jóindulatú,
és szinte mindenhol megtalálható, mint például talajban, vízben és eukaróta sejtekben. 2006-ban
készült el a genom megszekveneálása. 9,7Mb hosszúnak és 67% G+C-tartalmúnak találták. A
Rhodococcus törzsek alkalmazása fontos, a széles körű vegyületlebontó képességük és a bioaktív
szteroid-, akrilamid-, akrilsavtermelésük miatt, illetve a fosszilis üzemanyagok biológiai kénmente-
sítése végett. Ezt a nagyfokú genetikai és katabolikus változatosságot nemcsak a nagy kromoszómális
DNS-ének, hanem 3 nagy, lineáris plazmidjának is köszönheti. A Rhodococcusok kísérleti szem-
pontból is kedvező törzsek gyors növekedési sebességüknek köszönhetően. Ennek ellenére még
nincsenek tökéletesen jellemezve, leírva.
Egy másik fontos alkalmazásuk a biokonverziók területén van, amelyek biológiai rendszereket
használnak olcsó nyersanyagok értékesebb vegyületekké alakításában. A Rhodococcusok (3.2.16.
ábra) ezen felhasználása az olyan veszélyes környezetszennyező anyagok lebontásának képességéből
született, mint amilyen a toluol, naftalin és herbicidek. Általában az aromás vegyületek hasznosí-
tásakor az első lépésben az aromás gyűrűt oxigenálják, és két alkoholcsoport keletkezik a gyűrűn.
Ezután belső/külső diolmechanizmus szerint a gyűrű felhasad, és további metabolizmus alá kerül.
Mivel ezen folyamatok kémiája igen sztereospecifikus, a keletkezett diolok kiralitása tervezhető. Míg
a kémiai reakciók kiralitásának kontrollálása a szintetkus kémikusok számára jelentős kihívás, bioló-
giai folyamatokat lehet használni a királis molekulák előállítására, ha a közvetlen kémiai szintézis
megvalósíthatatlan vagy nem hatékony. Erre példa a Rhodococcusok felhasználása az indene elő-
állításában, ami a Crixivan nevű AIDS-gyógyszer prekurzora, egy protáz inhibítor 2 királis centrum-
mal a komplex molekulához szükséges 5-ből.
3.2.16. ábra: Rhodococcus species
Kevéssé jelentős, közeli rokonai a Nocardiáknak és Rhodococcusoknak a Tsukamurellák [53].
Ezek talajban előforduló Actinomyceták, amelyek immunszupresszált páciensekben tüdőgyulladást
okoznak. Különlegességük, hogy a sejtfalban található mikolsavaik extrém hosszú szénlánccal (~60
szénatom) rendelkeznek. A Tsukamurella paurometabola iparilag megfermentált sejtjeinek liofilezett
formáját fitonematódák írtására használják a mezőgazdaságban biokontroll során [54].
Az Actinomyceták következő csoportja a Propionibacteriumok (Propionibacteriaceae), amelyek
nem mozgó, spórátlan, anaerob pálcikák, és gyakran kórokozók (pl.: Propionibacterium acne (3.2.17.
ábra), bőrpórusokban szaporodik el, eltömve azokat, és aknét létrehozva.).

78 Ipari mikrobiológia
© Németh Áron, BME © www.tankonyvtar.hu
3.2.17. ábra: Propionibacterium acne
Nevüket a különleges propionsav-termelő anyagcseréjükről kapták, mint azt már korábban
tárgyaltuk (1.3. fejezet, erjedések). A propionsav-baktériumok kereskedelmi érdeklődésre tartanak
számot a svájci típusú sajtok íz- és lyukképzése kapcsán, továbbá egyre elterjedtebb az a nézet, hogy a
propionsav-baktériumok probiotikumok, egyrészt az általuk termelt propionsav, B12-vitamin,
fólacinok, bakterocinek miatt, másrészt a bélflóra tejsvabaktériumai növekedésének stimulálása végett
[39]. Ilyen faj a P. freundenreichii subsp. shermanii. Éppen a sajtgyártás során merült fel először,
hogy a sajtmátrix kialakulásakor először tejsavas erjedés megy végbe, amelyet az érlelési szakaszban
propionsavas erjedés követ, ezért a propionsav-baktériumok alkalmasak a tejsavasan leerjedt fermentlé
továbbhasznosítására. Ipari szempontból azonban a propionsav-baktériumok (PAB) tenyésztése
jelenleg körülményes, mert lassan növekvő törzsek, amelyek az esetleges bakteriális fertőzésekkel
nem tudnak versenyezni. Egyik megoldás lehet a PAB-ok törzsfejlesztéssel történő „felgyorsítása”,
egy másik pedig az immobilizált PAB-ok alkalmazása.
A Propionibacterium acidipropionici propionsav-termelésre használható 37°C-os optimális
hőmérséklet és anaerob körülmények valamint 6-os pH mellett, míg B12-termelésre is alkalmas 40°C-
os hőmérsékleti optimumammal és 0,5 vvm levegőztetés mellett 6,5-ös pH-n [40].
Ugyanez a baktérium korábban (XX. sz. közepe) Propionibacterium pentosaceum néven szerepel
a szakirodalomban viszonylag nagy terjedelemben, és a baktérium eritritátalakító képességére [41]
valamint nitrát-nitrit átalakítására [42] fókuszálva P. pentosaceum képes a borostyánkősavat is
propionsavvá alakítani [43], amivel megteremti az átjárás lehetőségét a C4 platformból a C3-ba. A
propionibacteriumok egyik iparilag is számottevő csoportja a Nocardioidák családja és azon belül a
Pimelobacter simplex (korábban Arthrobacter simplex: széles körű szteroidkonverziós képességek).
Az actinomyceták következő családja a Pseudonocardia-k (Pseudonocardiacea), amelyek sejtfala
nem tartalmaz mikolsavat. Többynyire lebontóképességüket írták le, így ezek is bioremmediációra
lakalmas fajok, mint például a szénhidrogénbontó Pseudonocardia hydrocarbonoxydans (olykor
Amylocota hydrocarbonoxydans néven szerepel). A P. sulfidoxydans egyes proteobakterekkel együtt a
bioszféra kénkörforgásában játszik szerepet, mivel a kéntartalmú aminosavak (mikrobiális)
lebontásakor dimetil-szulfid és dimetil-diszulfid keletkezik, amelyeket néhány proteobakter és
pszeudonokardia képes csak szulfáttá alakítani. Ezt a tulajdonságát szagmentesítő biofilmek
alkalmazásakor is kiaknázzák [48]. A P. halophobica szintén többszöri átnevezés után (Amylocota
autotrophica) került mai filogenetikus helyére, és korábbi nevén antibiotikumkonverziós irodalmak
lelhetők fel [49]. Az előbbi Pseudonokardiak viszonylag újonnan felfedezett közeli rokona az 1,4-
dioxánnal szennyezett ipari szennyvíziszapból izolált dioxánbontó P. dioxanivorans. Tetrahidrofurán
és egyéb éterszármazékok lebontására is képes.
Az Actinomyceták egyik legismerteb képviselői a Streptomycesek, amelyek másodlagos
metobiltjaikról híresek. A Streptomyces griseus (3.2.18. ábra) tenyészetében fedezték fel (1943) az
aminoglikozid típusú sztreptomicin antibiotikumot.

3. Az ipari mikroorganizmusok és technológiák tárgyalása törzsrendszertanba illesztve 79
© Németh Áron, BME © www.tankonyvtar.hu
3.2.18. ábra: Streptomyces griseus
A neomycin, klóramfnikol, streptomicinek mellett az utóbbi években új szerepet kaptak a
Streptomycesek: heterológ génepresszióra egyre gyakabban használják a Streptomyceseket, mivel az
eukarióta fehérjék E. colival történő kifejeztetése nem volt elég hatékony. Másodlagos metabolitjaik
között számos antifungális szer is található: nisztatin (Streptomyces noursei), amphotericin B (S.
nodosus). A fajokban gazdag nemzetség olyan antibiotikum termelésével rendelezik, mint a klór-
amfenikol (S. venezuelae), tetraciklin (S. rimosus), Vankomicin (S. orientalis). A penicillinrezisztens
mikrobák β-laktamáz enzimeinek gátlásával újra antibiotikum-érzékennyé tehetők. Ehhez klavulán-
savat szoktak a penicillinekkel együtt formulázni, amely kiegészítést a S. clavuligerus faj termel [161].
4. Thermotoga
A Thermotogák termofil vagy hipertermofil, leginkább 80°C (legmagasabb optimum a baktériumok
között) körül, semleges pH-n növekvő mikrobák. Általában pálca alakú sejtek, külső membránnal
(=”toga”) is borítva. Enzimeik a magas hőmérsékleten való aktivitásukról ismertek. Már a ’90-es
években ipari szemszögből foglalkoztak a cellulolítikus és xilánbontó enzimeikkel.
A sótűrésük nagyon változó, míg egyesek extrém magas sókoncentrációt is elviselnek, mások
csak alacsony sókoncentráció mellett növekednek, így olyan változatos élőhelyeken találhatók meg,
mint például a sekélyesek vagy a mélytengeri rendszerek és kontinentális olajmezők. Elsőként a T.
maritimát izolálták egy vulkánból, majd egy olasz sekélytengeri üledékből, amely geotermikusan
fűtve volt. A következő faj a T. neopolitana volt, amelyet egy Nápoly közeli tenger alatti tengeri hő-
forrásból izoláltak. A legújabb Thermotoga törzs 68°C-os melegvízben 6,8-as pH-n nő Németország-
ban. Ezek az aerob Gram-negatív mikroorganizmusok általában spórátlanok, és számos szénhidrátot
metabolizálnak, például glükózt, saccharózt, keményítőt, cellulózt és xilánt. Az L-alanin- termelésük
hasonló az ősbaktériumok közé tartozó Thermococcusokéval, ami azt mutatja, hogy a cukorból L-
alanin-előállítás egy korábbi ősi metabolizmus jellegzetessége.
3.2.19. ábra: Petrotoga sp.

80 Ipari mikrobiológia
© Németh Áron, BME © www.tankonyvtar.hu
Egy hasonló faj az anaerob Petrotoga (3.2.19. ábra) olajtartályokból izolálható, ami felveti a kérdést
mind a mély felszín alatti túlélésükről, mind pedig az őshonosságukról ezen a különleges ökoszisz-
témában.
5. Fusobacteriumok
Anaerob, heterotróf, fonalas szervezetek, noha a humán felsőlégúti (torok, garat) szervek normál
flórájához is tartothatnak, számos fertőzés kórokozói (Fusobacterium necrophorum), ezért minden
Fusobaktert fertőző ágensként kell kezelni. Ennek ellenére bacteriocineket [73], és β-laktamázt [74] is
termeltethetnek velük.
3.2.20. ábra: Fusobacterium novum [178]
6.-7. Deinococcus-Thermus
Két rendet (Deinococcales és Thermales) magába foglaló (ezért kettős nevű), kevés fajt tömörítő
baktériumtörzs, ahová zömében extrémofilek tartoznak. A Deinococcus radioduranst (3.2.21. ábra)
poliextrémofillnek is nevezik, mivel – nevéből adódóan – jól tűri a sugárzásokat, és ezen felül még
hidegtűrő, savtűrő, jól viseli a kiszáradást és a vákuumot is (azaz „űrutazásra” is képes). Az ellenálló
képességét annak köszönheti, hogy a genomját több példányban is fenntartja, illetve hatékony DNS-
javító rendszerrel rendelkezik. [165]
3.2.21. ábra: Deinococcus radiodurans [178]
A Thermus thermophilus japán hőforrásból izolált 80°C-os növekedési optimummal és intenzív
oxigénigénnyel rendelkezik, noha az oxigén oldhatósága e magas hőmérsékleten meglehetősen kicsi.
Ugyanakkor hőstabil enzimek forrás-mikoroorganizmusaként nagy jelentőséggel bír (pl.: rTth-DNS
polimeráz [67], α-glükozidáz [75]). A teljes genomját megszekvenálták, így lehetővé téve a
termofilexpressziós rendszer kialakítását

3. Az ipari mikroorganizmusok és technológiák tárgyalása törzsrendszertanba illesztve 81
© Németh Áron, BME © www.tankonyvtar.hu
8. Chloroflexi
Zöld, nem kénbaktériumok csoportja, általában fonalas morfológiával. A fényt használják
energiaforrásként a zöld pigmentjeik segítségével (nem kloroplaszt!), siklással („gliding”) mozognak.
A fényhasznosító rendszerük a magasabb rendű élőlényeknél megszokott II-típusú fotoszintetikus
reakcióközpontot használja bakterioklorofillal, a Chlorobi fajaihoz hasonlóan. Míg a magasabb rendű
fotoszintetizáló élőlények vizet használnak elektron- donorként, addig a Chloroflex-ek H2S-t (mint a
Chlorobi-k) vagy H2-t. A szokásos Calvin-ciklus helyett pedig egy sajátos, 3-hidroxi-propionát ciklust
használ a szerves anyagok beépítésére. Szemben a szigorúan anaerob Chlorobi fajokkal, képesek aerob
heterotróf anyagcserére vagy anerob fotoheterotróf, illetve anaerob fotoautortóf anyagcserére is [78].
Egy típusfaj a Chloroflexus auranticus (3.2.22. ábra), amely termofil baktérium sötétben, oxigén
jelenlétében is szaporodik, és narancsszínű. Fény mellett fotoszintetizál, és zöld színű telepeket alkot
[76]. Ipari potenciált speciális amilázai jelentenek, amelyek a keményítőt nem maltózra vagy glükózra,
hanem maltotriózra és maltotetraózra (3, illetve 4 tagú oligoszaharidokra) bontja, amelyeknek újabban
nagy jelentősége van az élelmiszeripar, gyógyszeripar és a finomvegyipar számára [77]. Ugyanezen
mikroba különleges 3-hidroxi-piruvát- termelő képessége szintén ipari potenciált rejt, mivel a 3-
hidroxipiruvát és származékai egy fontos platformot alkotnak, amelyek előállításához új megközelítést
és új enzimeket szolgáltathat a Chloroflexus auranticus [79].
3.2.22. ábra: Chloroflexus auranticus [178]
A Chloroflexek másik rendje a Herpetosiphonales, amelynek egyik típus faja a Herpetosiphon
aurantiacus [80], ám ezek iparilag még jelentéktelenek.
9. Cyanobacterek
Fotoszintetikus baktériumok és a kloroplasztok csoportja. A mikroalgák jelentős része is közéjük
tartozik. Poshadt állóvizek lakója, amely vizek az általuk termelt cianotoxinok miatt mérgezőek.
Nevüket kékeszöld színüknek köszönhetik. A cianobaktériumok oxigéntermelő fotoszintetikus képes-
sége tette lehetővé a korábbi redukáló légkör oxidatívvá tételét, ami drámaian megváltoztatta az élet-
formákat a Földön, és megnövelte a biodiverzitást, noha ez veszélyeztette az oxigénintoleráns fajok
fennmaradását. Az endoszimbionita elmélet szerint a növényi és algakloroplasztok endoszimbionita
cianobaktériumokból származtathatók.
Biotechnológiai potenciáljukat jelzi, hogy az egysejtű Synechocystis cianobaktérium volt a 3.
prokarióta, amelynek teljes genomját feltérképezték. Ez továbbra is fontos modellszervezet maradt. A
genomméretük tekintetében igen nagy szórás érzékelhető: Prochlorococcus 1,7Mb, Nostoc
punctiforme 9Mb, Calothrix 12-15Mb, ami már az élesztőével azonos nagyságú.
Felhasználásuk szempontjából némelyeket élelmiszerként forgalmaznak (pl.: Aphanizomenon
flos-aquae), ugynakkor újabban a zöldenergia-termelésben játszhatnak szerepet a napfény közvetlen
elektromossággá alakításával. A makroalgákhoz (ld. később) hasonlóan intenzíven kutatják a
cianobakter-alapú alternatív üzemanyag előállításának (dízel, benzin, kerozin) lehetőségét [81].
Leggyakoribb felhasználásnak az Arthrospira (3.2.23. ábra) nemzettség fajai örvendenek, amelyeket
részletesebben az Algák fejezetben tárgyalunk (ezek elterjedt neve a Spirulina).

82 Ipari mikrobiológia
© Németh Áron, BME © www.tankonyvtar.hu
3.2.23. ábra: Arthrospira (Spirulina) sp. [178]
10. Spirohaeták
Általában csavarodott vagy szabálytalan alakú baktériumok, amelyek többsége kórokozó (pl.: Lyme-
kór -kullancsok által is terjesztett), amelyet a Borrelia burgdorferi okoz, vagy a Treponema pallidum,
amely a szifilisz kórozója). Szaharolítikus [82] és etanoltermelőkre [83] is találunk példát, de ipari
felhasználásuk nincs. A Spirohaeta thermophila fermentatív anyagcserével rendelkezik, és a glükózt
tejsavvá, etanollá, szén-dioxiddá, illetve hidrogénné alakítja [84].
11., 12,. 13. Chlorobik, Fibrobacterek, Bacteroidesek
Egyes források szerint [27] a korábbi CFB (Cytophaga, Flavobacter, Bacteroides) csoportot
átszervezték Fibrobacterek, Chlorobik, Bacteroidesek számára. A Fibrobacterek többnyire a
bendőben találhtaók meg, ezért ipari szempontból cellulázaik figyelemre méltóak. A Fibrobacter
succinogenes jó hozammal (Y=83%, bár alacsony produktivitással J=14 mg/lh) képes a cellulózból
borostyánkősavat (platformalkotó!) előállítani.
A Chlorobi törzs (amelyet zöld kénbaktériumoknak is szokás nevezni) obligát fotolitotrófokat
foglal magába, amelyek szigorúan anaerob fotoszintézist végeznek, azaz elektrondonorként H2S-ot
használnak. Általában anoxikus vizes közegben találhatók meg, ahová a napfény bejuthat. A
fotoszintetkius antennamolekuláikat flexibilesen tudják változtatni, aminek következtében változtatják
az elnyelhető fényhullámhosszt, és így olyan rétegekben is életképesek, ahol más fotoszintetizáló
szervezet számára nincs vagy nem megfelelő a beérkező fény.
3.2.24. ábra: Bacteroides biacutis
A CFB patogén, kommenzalista és szabadon élő mikrobák diverz csoportja. A Bacteroides
fragilis toxi termeléséről ismert, de aminok előállítására is használható, valamint a bélbaktériumok
között is előfordul [69]. A normál humán mikrobióta mintegy 20%-át a Bacteroides-ek teszik ki
(3.2.24. ábra). A bélbaktériumok között különleges szerepük lehet, mivel igen jó cukorlebontó
képességgel rendelkeznek. Az emberi emésztőrendszer felső szakaszán (száj, nyelőcső, gyomor és a

3. Az ipari mikroorganizmusok és technológiák tárgyalása törzsrendszertanba illesztve 83
© Németh Áron, BME © www.tankonyvtar.hu
vékonybél) a cukrok lebomlanak és felszívódnak, így csak a poliszaharidok (pl.: növényi rostok)
szolgálnak a vastagbélben honos mikrobióta számára szén- és energiaforrásul. Az ilyen növényi rostok
fogyasztása csökkentheti a vastagbélrák kialakulásának kockázatát. A különböző fajok szubsztrát-
specifitása jelentős különbségeket mutatnak, így a belőlük nyerhető degradációs enzimek is eltérőek.
A B. fragilis keményítőszármazékok (amilóz és amilopektin) bontásában jeleskedik, a B. ovatusok
között pedig xilán, dextrán, pektin, guargumi és alginát bontók is találhatók.
14., 15., 16. Planctomyceta-k, Verrucomicrobiumok, Chlamydiák
A Planctomicetákkal kapcsolatban az első felfedezés egy magyar nevéhez fűződik (Gimesi Nándor)
1924-ből [70]. Az általa a Lágymányosi-tóban felfedezett csillag alakú élőlényeket (3.2.25. ábra)
planktonikus gombának osztályozta, csak később derült ki, hogy az egyik legfejlettebb prokariótáról
van szó. P. gimesii képes cukrokat fermentálni (erjeszteni). Taxonómiai jellmezőjük a peptidoglükán-
mentes fehérjékkel borított sejtfal, a sarjadzó szaporodás, DNS-t borító membrán, különleges
morfológia (nyeles sejtek, többsejtűek, a nyéllel hajlamosak megtapadni) és részleges
kompartmentizáció [85]. A Verrucomicrobium és Klamidiák fajai általában eukarióták parazitái.
3.2.25. ábra: Planctomyces bekefii
17. Aquificae
Az Aquificae fajok szigorúan hipertermofilek, és jelenleg úgy gondolják, hogy ezek alkotják a
baktériumok közötti első elágazást. Az elágazási pont helye a filogenetikus fán változó, és nem
tisztázott. Ezt a törzset csak mostanában fedezték fel a 16rRNS-ében tapasztalt eltérés alapján.
Azonban ezen kívül nincs más ismert molekula vagy tulajdonság, amely megkülönböztetné őket más
baktériumoktól.
Kemolito-autotrófok, azaz CO2-t fixálnak, és szervetlen anyagok oxidálásával nyerik energiájukat.
A törzs neve is innen ered: hidrogén gáz oxidációjával (akár minimális O2 segítségével) vizet
termelnek (Aquifex=vízjelző). Magas hőmérsékleti optimumuk miatt mint genetikai (és proteomikai)
potenciált kutatják. Általában 2-6 mikronos hosszúságúak (3.2.26. ábra), nem spóraképző Gram-
negatív mikrobák. Az Aquifex pyrophilus anaerob módon is képes növekedni, miközben oxigén helyett
nitrogént redukál.
3.2.26. ábra: Aquifex aeolicus

84 Ipari mikrobiológia
© Németh Áron, BME © www.tankonyvtar.hu
18., 19., 20., 21., 22. Proteobacterek
A „Proteobakter” gyűjtőnevet újabban a korábban használatos bíbor baktérium helyett vezették be,
mivel a nagy diverzitású taxonok között sok nem „bíbor” színű és néhány fotoszintetizáló is található
[86]. A név maga a görög Proteus nevéből származik, jelezvén a sokféle idesorolt fajt. A 16S rRNS-
analízisek alapján a proteobakterek több száz nemzettségét 5 (egyes források szeint 6) osztályba
sorolták, amelyeket a görög ábécé betűivel jelölnek (α, β, γ, δ, ε (δ)). Az rRNS-analízisek szerinti
elágazási sorrend más, mint az ábécé szerinti sorrend, e jegyzetben az eddigi koncepcióhoz híven az
elágazási sorrend szerint tárgyaljuk a proteobaktereket.
Delta-, epszilon-proteobacterek
A delta-proteobacterek 3 nagy csoportra oszthatóak: a szulfát/kén redukáló Desulfovibriokra, a
predátor fajta Bdellovibriokra és a gombaszerű termőtesteket és spórákat képező Myxobacterekre. A
Desulfovibrio fajok a szulfát, nitrit, nitrát, CrVI, UVI redukálóképességük miatt eddig is a
(környezetvédelmi) kutatások középpontjában álltak (pl.: Desulfovibrio desulfuricans, D. vulgaris)
[144]. Az utóbbi évtizedben olyan különleges fajt (és szintén deltaproteobacter rokonait) fedezték fel,
mint a Desulfobacter magneticus, amely magnetotaktikus, és magnetoszómájába magnetit ásványt
(FeII és FeIII oxidot) választ ki (biomineralizáció) [143].
A Bdellovibrio fajok általában más Gram-negatív baktériumok predátorai, és azok periplazmás
terében élnek (pl.: Bdellovibrio bacteriovorus). E különleges baktériumok 160 μm/s sebességgel (saját
hossz 100x-a szekundumonként) nekiütköznek az áldozatnak, és kilyukasztják annak külső
membránját, majd a lyukon át beköltöznek a periplazmás térbe, befoltozzák a lyukat, és várnak. (látens
állapot). Ezt követően hidrolítikus enzimeikkel feloldják a gazdaszervezetet, és fonalas formában
benövik annak belsejét („bdelloplaszt”). Az életciklus végén a fonalak egyedi sejtekké esnek szét. Egy
E. coli sejtből, kb 3-6 Bdellovibrio sejt keletkezik, de a nagyobb áldozatokból akár 80 is keletkezhet
[145]. Biotechnológiai potenciált a litikus enzimeik jelenthetnek, de egyelőre ezek alkalmazása nem
ismert.
A Myxococcus tagjai a legnagyobb genomúak, és tápanyaghiány esetén termőtestet (3.2.27. ábra)
hoznak létre, amely myxospórákat termel. Ezek rendkívűl ellenállóak. Aktív mozgásra („siklás”)
képesek: a temőtestet átrendezik, ezért „nyálkabaktérium”-nak is szokás nevezni őket [146].
3.2.27. ábra: Myxococcus xanthus [178]
A Myxococcus xanthust használják modellszervezetként a Myxococcusok változatos, bioaktív,
másodlagos anyagcseretemékeinek vizsgálatára. Ezek sajátos új szerkezetű antitumor (pl.: epothilon,

3. Az ipari mikroorganizmusok és technológiák tárgyalása törzsrendszertanba illesztve 85
© Németh Áron, BME © www.tankonyvtar.hu
hatása, mint a paclitaxelé), antibiotikus (pl.: orangicin), illetve sziderofór (Myxochellin) hatású
anyagok [147], amelyek komoly (gyógyszer)ipari lehetőségeket rejtenek.
Az epszilon-proteobacterek között a Campylobactereket és a Heliobactereket találjuk. Mindkét
nemzettség emésztőrendszeri megbetegedéseket okoz, a Heliobacterekről az utóbbi években derült ki,
hogy a gyomorfekély kórokozója (Heliobacter pylori, 3.2.28. ábra), amely fekély után 6x nagyobb a
gyomorrák kialakulásának esélye [142]. Ipari alkalmazás vagy annak lehetősége egyiknél sem ismert.
3.2.28. ábra: Heliobacter pylori [178]
Alfa-proteobacterek [87]
A 16S rRNS-alapú filogenetikus fán az alfa-proteobactereket 5 „cluster”-re osztják. Az első a
Caulobakter-féléket tartalmazza három további alcsoportba osztva. Ezek közül ipari potenciállal,
illetve felhasználással rendelkezik a Brevundimonas diminuta, amely kis mérete miatt vízszűrők
hatékonyságának tesztorganizmusa, valamint restrikciós endonukleázok (PdiI, PdmI) forrása, korábbi
neve pedig Pseudomonas diminuta [88]. A Brevundimonas vesicularis pedig a biodegradálható
műanyagok alapanyagának a poli-hidroxi-alkanoátoknak előállítására alkalmazható akár fűrészpor-
alapú fermentációban is [89]. A másik alcsoportba tartoznak a Caulobacterek. A Caulobacter
crescentust (típus törzs) expressziós rendszerként használják, mivel kisméretű fehérjéket poszt-
transzlációs glikozilezés nélkül extracellulárisan termel [90]. Felhasználható haszonállatok vakcinálá-
sára is, mivel egy különleges „S-layer kit” segítségével a sejtek felszínén jelennek meg a klónozott
gének géntermékei, amelyek antigének is lehetnek [91]. Egyes tengeri Cauolbacterek erős antibioti-
kumot (tiotropocint) termelnek, amely egyszerre gátolja a fitoplankton, algák és baktériumok növe-
kedését [92].
A Caulobakterek 2. csoportjába a Rhodobakter-félék tartoznak, idesorolva a Paracoccus nemzet-
séget is, amelynek tagjai (pl.: Paracoccus panthotrophus) felhasználhatók poli-hidroxi-butirát (PHB,
bioműanyag) előállítására [94], astaxantin (karotinoid) előálítására (P. haeundaensis) [95]. A
Paracoccus denitrificans (korábban Micrococcus denitrificans) denitrifikáló (nitrátredukáló)
baktérium, tehát fontos szerepe van a nitrogén-körforgalomban, és egyben a mitokondrium őse, azaz
egy P. denitrificans endoszimbiózisával jöhetett létre az eukarióták mitokondriuma [96]. A Rhodo-
bacterek alkalmazásának lehetőségére példa a Rhodobacter sphaeroides heterológ génexpressziós
rendszere. Emellett széles körű anyagcsere- lehetőségekkel rendelkezik: noha legjobban anaerob
fotoszintézis mellett szaporodik, képes fermentatív heterotróf anyagcserére (aerob, illetve anaerob
légzéssel is), illetve nitrogénfixálásra is. A Rhodobacter capsulatus PHA- és PHB-termelő képességét
széleskörűen kutatják és fejlesztik [97]. Ugyanaezen faj a fotoszintetikus biohidrogén termelésében is
jeleskedik [98]. Szintén biotechnológiai potenciált jelent a R. capsulatus esetében a bacteriocin-
termelés is [99]. Közeli rokonként ide sorolják a Pseudorhodobactereket, Amaricoccusokat, Antarcto-
bactereket, Leisingerákat, Octadecabactereket, is, ám ezek ipari alkalmazása jelenleg nem ismert.

86 Ipari mikrobiológia
© Németh Áron, BME © www.tankonyvtar.hu
A szintén idesorolt Ketogulonicigenium vulgare (3.2.29. ábra) szorbóz dehidrogenázának
irodalma viszonylag kiterjedt, különösen, ha a korábbi nevén (Gluconobacter oxydans DSMz4025)
keressük. Így arra is fényderül, hogy az egyik legrégebbi (bio)technológia a C-vitamin- gyártás során
használatos, amelynek kulcslépése a szorbit-szorbóz átalakítás. Ezt éppen G.oxydans mikrobával
végzik, amelyet 2001-ben soroltak át új taxonómiai helyére [100].
3.2.29. ábra: Ketogulonicigenium vulgare
A Rhodobacter-félék közé tartoznak a Hyphomonasok, amelyek extracelluláris poliszaharidokat
termelnek, és ezekkel mint ragasztóval a biofilmek képződésében játszanak szerepet (pl.: Hyphomonas
neptunium) [93].
Az alfaproteobacterek teljes filogenetikus fája ,számos itt fel nem sorolt fajjal, hozzáférhető a
http://ijs.sgmjournals.org/cgi/data/55/5/1907/DC1/1 címen.
Felhasználási potenciáljuk miatt azonban meg kell még említeni a nitrogén-körforgalomban
fontos szerepet játszó alfaproteobaktereket. Az Azospirillium fajok a mezőgazdaságban kerülnek
szándékosan (farmerek által), illetve természetes úton felhasználásra, mivel ezek megkötik a légköri
nitrogént, és az abból képzett ammóniát glutamin formájában a szimbionita (haszon)növény
gyökerének átadják, amiért cserébe az életfolyamataikhoz és a nitrogénfixációhoz szükséges
energiatermelésükhöz malonsavat (almasav) kapnak a növénytől (heterotróf szimbiózis). Ezáltal
csökkenthető a felhasznált műtrágya mennyisége.
A Nitrobacterek szintén az alfaproteobakterek közé tartoznak, és a nitrifikációért felelnek, azaz az
ammóniából előbb nitritet, majd nitrátot állítanak elő energianyerés céljából szén-dioxid fixáció
közben (kemoautotróf anyagcsere). A hüvelyes növények esetében ez az egymásra utaltság
endoszimbiózis formájában mégszorosabb, és ebben az esetben a Nitrobacterek legközelebbi rokonai
a Rhizobiumok végzik a nitrogénmegkötést.
A Rhizobiumok között található meg újabban (2001) a nagy biotechnológiai múlttal rendelkező
Agrobacterium tumefaciens [170] (3.2.30. ábra), amely növény patogénként a növényi géntranszfer
eszközévé vált.
3.2.30. ábra: Agrobacterium tumefaciens répaszöveten [178]
A Roseobacterek a tengeri mikrobióta leggyakoribb tagjai, amelyek igen változatos másodlagos
anyagcseretermékekkel rendelkeznek ezért intenzív (antibiotikum) kutatások tárgyát képezik [101], de
találhatóak köztük denitrifikálók is (Roseobacter denitrificans) [102].
Végül meg kell említeni, hogy az alfaproteobacterek között olyan patogéneket is találunk, mint a
Rikettsiák és Brucellák. Előbbiek az ízeltlábúak kórokozói, amelyek akár tünetmentes vektorok is

3. Az ipari mikroorganizmusok és technológiák tárgyalása törzsrendszertanba illesztve 87
© Németh Áron, BME © www.tankonyvtar.hu
lehetnek, és csípésük során vagy székletükkel kerülnek át a paraziták az emberbe, ahol a hajszálereket
fertőzik meg, és trombózist, szervi működészavart eredményeznek láz és kiütéses tünetek mellett
[103]. Mivel obligált opportunisták, önmagukban nem tenyészhetők mesterséges környezetben
(fermentorban), csak szöveteken. A Brucellózis abortív betegség, amely a „szarvasláb” nevű beteg-
séget okozza például teheneken, ahonnan tejjel vagy sérült bőrfelületen kerülhet át az emberbe, és
súlyos lázas megbetegedést okoz.
A bioetanol-gyártás bakteriális jelöltje a Zymomonas mobilis is alfa-proteobcter. Különlegessége,
hogy sejtfalában hopaninsavat tartalmaz, amely az eukarióta szterinekhez hasonlít. Ennek
köszönhetően jól tolerálja a magas (13%) etanolkoncentrációt is. Emellett nagyobb etanolhozam és
kisebb biomassza-képződés tapasztalható a Z. mobilis etanolos fermentációjában [160].
Beta-proteobacterek
A Hydrogenophilaceae család tagjai közé tartoznak a Thiobacillus nemzetség fajai, amelyeket
biokontrollra lehet felhasználni (pl.: T.denitrificans), mivel kénporral történő együttes kiszórásuk után
a krumpliföldön a kenet kénsavvá oxidálja, melyet a krumplibogarak lárvái nem viselnek el [104]. A
Thiobacillus ferrooxidans a piriteloxidálást segíti Acidophilumokkal együtt aerob környezetben
(respiráló mikrobák). Néhány Thiobacillust 2000-ben átneveztek Acidithiobacillus, Halothiobacillus
és Thermithiobacillus néven a gammaproteobakterek közé sorolták. A Thiobacillusok igazi ipari
jelentőségét a szulfit ásványokból történő fémfelszabadítás (=”bioleachig”) adja, amelyre példa az
autotróf kemolitotróf Thiobacillus ferrooxidans [105]. A Thiobacillus denitrificans különleges
metabolikus repertoárral rendelkezik: szervetlen kén- vegyületek oxidációjával vagy aerob FeII-
oxidációval kapcsolja össze a denitrifikációt (NO3->NH4+). Fakultatívan választ az aerob respiráció
és a denitrifikáció között, és mezofill, illetve neutrofill hőmérsékleti és pH-tartományokban szapo-
rodik szemben a termo- és acidofil kénoxidáló baktériumokkal. Az a különleges mehanizmus, ahogy
az oldhatatlan szilárd ásványt hasznosítani képes még pontosan nem ismert [106].
A Thiobacillusok közvetlen rokonai a Hydrogenophilus nemzetség tagjai, amelyek különleges-
sége, hogy energiaszükségletüket hidrogén oxidációjával nyerik [171].
A következő csoportot az aerob, fonalas Sphaerotilusok alkotják, típusfajuk a Sphaerotilus
natans, ipari alkalmazásuk nem ismert.
Őket követik a Neisseriák, amelyek tipikus kórokozók: Neissria gonorrhea (nemi betegség), N.
meningitidis (agyhártyagyulladás) [107]. A Spirillumok felhasználásáról nem áll rendelkezésre
információ, viszont a Burkholderia nemzettség tagjai alkalmazhatók [108] a mezőgazdaságban
biokontroll célokra, rhizobiális adalékként, illetve biodegradációban, valamint xilanáztermelő is
található közöttük [109]. A nagyobb felhasználási kör jobban megérthető, ha figyelembe vesszük,
hogy szinte valamennyi Burkholderia faj a Pseudomonasok újrabesorolásakor került leírásra, és
amelyek vizsgálata nagyobb múltra tekint vissza, így alkalmazásuk is szélesebb körű. Példa erre a
Burkholderia cepacia (3.2.31. ábra) és B. gladioli, amelyek rendre a Pseudomonas cepacia és a P.
gladioli gamma-proteobacterekhez történő átsorolásakor kerültek először leírásra Yabouchi és
munkatársai (1992) munkássága nyomán. Mindkét törzs lipázforrásként használatos [110].
3.2.31. ábra: Burkholderia cepacia [178]

88 Ipari mikrobiológia
© Németh Áron, BME © www.tankonyvtar.hu
A Burkholderia rendbe tartoznak az Alcaligenaceae család fajai is, amelyek között a Bordatella
pertusis, vagyis a szamárköhögés kórokozója is megtalálható. Ennek biotechnológiai jelentősége a
gyermekkori vakcina (DiPaTe, korábban DiPerTe) előállításában és alkalmazásában van. A korábbi
DiPerTe vakcina gyakran váltott ki lázas oltátási mellékreakciókat, mióta azonban a pertusis inaktivált
sejtjei helyett antigéneket alkalmaznak (acelluláris azaz sejtmentes készítmény), kevesebb a
mellékreakció. Egy másik különleges nemzetség az Alcaligenes, amelyeket ipari aminosav előállí-
tására használnak, illetve az A. eutrophust PHB-termelésre is használják.
Korábban szintén a Pseudomonas családba sorolták a Zoogloea fajokat, majd elkülönítették a
többi obligát aerob, Gram-negatív spórátlan pálcikától (=Pseudomonasok), mert sajátos zselatinszerű
mátrixot képeznek [111]. Ennek megfelelően biotechnológiai alkalmazásra szinte kizárólag a
„zoogloea gum”, vagyis a különleges extracelluláris poliszaharid előállítása esetén használják e fajo-
kat, köztük leggyakrabban a Zoogloea ramigerat.
A Ferribacterium limneticum szerves savak oxidációjával redukálja a Fe3+-t Fe2+-vé [172]. Ennek
közeli rokona számos nitrogénfixáló nemzetség (Azoarcus, Azovibrio, Azospira stb.), amelyek a
denitrifikációval együtt számos szennyezőanyag lebontását katalizálják (pl.: aromás szennyezők)
[173]. Aerob vagy anaerob heterotróf, nem-szulfát-redukáló, mérsékelten termofildenitrifikáló
baktériumok az olajmezők Petrobacter succinatimandens baktériumai, amelyek borostyánkősavon is
szaporodnak [174]. Különleges fermentatív mikroba a Propionivibrio limicola), amely sem
szénhidrátokon, sem szerves savakon, de mégcsak szerves szennyező anyagokon sem képes nőni,
viszont kinik-savon és sikimik-savon propionsavat, szén-dioxidot ecetsavat termel [175]. A
Rhodocyclus nemzetség tagkai anoxikus fototrófok, és a nevüket a sejtek gyűrűs alakjáról kapták,
aminek következtében a sejt két pólusa közel kerül egymáshoz [176, 177].
Gamma-proteobacterek
Fajokban meglehetősen gazdag proteobacter-csoport, amely számos súlyos betegség (pl.: kolera)
kórokozója mellett néhány biotechnológiailag jelentős mikrobát (egyes Pseudomonas-okat,
Enterobacter-eket) is magába foglal.
A Beggiatoak nagyméretű (200 μm!) fehér fonalas szervezetek, amelyek a néhány ”szintetikus”
szervezet közé tartoznak. Ezek képesek a légköri szén-dioxidból és a tengervízből szervetlen
vegyületek energiájának felhasználásával szénhidrátokat szintetizálni (autotrófok). Noha kataláz
negatívak, légköri oxigén felhasználásával heterotróf anyagcserére is képesek, mivel az oxigén okozta
nem kívánatos oxidációs folyamatokat kén segítségével közömbösíteni tudják [112]. Tengeri szennye-
ződések eltávolításakor megjelennek (indikátor), és nagy méretük miatt szabad szemmel is láthatóan
megjelölik a szennyezést.
A másik autotróf gamma-proteobacter csoport a bíborkén-baktériumok csoportja. Fotoszintézi-
sükhöz nem vizet használnak, mint a legtöbb fotoszintetizáló szervezet (cianobaktériumok, algák,
növények), hanem kénhidrogént, így oxigén helyett kenet szabadítanak fel [113]. Két családra
oszthatóak aszerint, hogy a kenet extracellulárisan (Ectothiorhodospiraceae) vagy intracellulárisan
(Chromaticeae) választják ki. A Chromaticae család legjobban leírt tagja a Thiocapsa roseopersicina,
amely által termelt hidrogenázenzim biotechnológiai (biogáz-termelési) potenciállal rendelkezik,
mivel a Thiocapsa alacsony növekedési hőmérsékleti optimuma (20-25°C, északi-tengeri izolátum(!))
ellenére a hidrogenáza hőstabil (>80°C), proteázoknak ellenáll, és jól tolerálja az oxigént (a Thiocapsa
roseopercicina fakultatív anaerob) [114].
A gamma-proteobacterek speciális alkalmazással bíró nemzettsége az Alcanivorax (pl.: A.
borkumensis, 3.2.32. ábra), amely tengeri olajszennyezések eltávolításánál hasznosítható, amennyiben
kén és nitrogén is rendelkezésére áll [115].

3. Az ipari mikroorganizmusok és technológiák tárgyalása törzsrendszertanba illesztve 89
© Németh Áron, BME © www.tankonyvtar.hu
3.2.32. ábra: Alcanivorax borkumensis
Hasonló képességekkel rendelkeznek a Marinobacterek. A M. hydrocarbonoclasticus (korábban
Pseudomonas nautica) azonban még extrém halotoleráns is [117]
Hidegtűrő gamma-proteobacterek a Glaciecolak (pl.: G. polaris, G. agarilytica), amelyek a
sarkvidéki jeges, oligotróp élőhelyeken is megélnek [116].
Nagy biotechnológiai potenciált rejt a Microbulbifer (újabban Saccharophagus) degradans
enzimkészlete, amely cellulóz, alginát és kitinbontásra is alkalmas enzimeket foglal magába [118].
Ezek segítségével a xilóz- és cellulózalapú etanolgyártás jelöltje.
A gamma-proteobacterek között olyan kórokozók és rokonaik is megtalálhatóak, mint a
Legionellak (pl.: L. pneumophila), Francisellak, Vibriok. A Legionellák halálos légúti fertőzést
okoznak, és a szovjet időkben 100% mortalitásúra mutáltatott fajt is kifejlesztettek belőle (=biológiai
fegyver) [119]. A Francisella tularensis a tularemia kórokozója, amely vadnyulakról kerülhet
kullancsfertőzés vagy vadászat közben az emberekre, és fekélyes lázzal jár, és akár
agyhártyagyulladásba is torkollhat. Az USA-ban biológiai fegyverként is rendszeresítették. Többször
átnevezték, előbb Bacterium tularensis, majd Pasturealla tularensisként került besorolásra [120, 121].
A patogén Vibriók által okozott egyik legsúlyosabb fertőzés a kolera (Vibrio cholerae), amely
szennyezett vízzel terjed. Ugyanakkor a Vibriók fermentálnak (erjesztenek) is, illetve tetrodotoxint
termelnek (Vibrio alginolyticus), amely egy neurtoxikus (nátriumcsatorna szelektív blokkolójaként
blokkolja az akciós potenciál kialakulását) ősi tengeri alkaloid, és a szimbiotikus baktériumok ezzel
nyújtanak védelmet a gazdaszervezeteiknek (hal) a ragadozók ellen, amiért cserébe bőséges táplálékot
kapnak. [122].
A Vibriók különleges rokonai a Photobacteriumok, amelyek biolumineszkálnak (csakúgy, mint a
szintén közeli rokon Lucibacteriumok), azaz fényt bocsátanak ki, és szintén szimbionták, továbbá
pszichrofilek (hidegkedvelők) és piezophilek (másnéven barofilek vagy nyomáskedvelők) [123]. A
Vibrio fisheri további különlegessége, hogy csak egy adott sejtkoncentráció elérése után bocsátanak ki
a sejtek fényt, amely koncentrációt a tengerben szabadon élve nem tudnak elérni, csak a halak
endoszimbiózisában. Ehhez a szervezett válaszreakcióhoz sejt-sejt közötti kommunikáció szükséges
[124].
3.2.33. ábra: Escherichia coli

90 Ipari mikrobiológia
© Németh Áron, BME © www.tankonyvtar.hu
Ipari szempontból talán a legjelentősebb gamma-proteobacter csoport az Enterobactereké.
Köztük olyan fajok találhatóak, mint a legtöbbet vizsgált és iparszerűen is alkalamzott heterológ
expressziós rendszer, az Esherichia coli, amelynek patogén fajtái is léteznek (3.2.33. ábra).
A glicerin – 1,3-propándiol-átalakítás enzimeit is főleg az Enterobacteriaceae között (pl.:
Enterobacter, Citrobacter Klebsiella stb.) találhatjuk meg.
Ide sorolták a mezőgazdasági jelentősséggel bíró Erwina amylovora fajt is, amely az almafélék
tűzelhalásos betegségét okozza.
A Proteus nemzettség tagjai igen változatos formában jelennek meg, ennek köszönhetik nevüket
is. Általában talajlakók és humán patogének (Proteus vulgaris, P. mirabilis, P. penneri) [125].
Talán a legismertebb humánpatogén enterobakterek a Salmonellák, közülük is a Salmonella
enterica sp enterica alfajai, amelyek az ételmérgezések jellegzetes kórokozói: typhi (hastífusz),
choleraesuis (szepszis) enteriditis (heveny gyomorrontás)) [126, 127].
A Salmonellák közeli rokonai az általában húgyuti fertőzést okozó Serratiák (pl.: Serratia
marcescens). Emellett azonban biotechnológiai potenciált is rejtenek, mivel egy értékes
gyógyszerhatóanyagot, a prodigiosint lehet velük hatékonyan ipari hulladékon előállítani. A
prodigiosin egy antifungicid, immunszupresszív és antiproliferatív pigment- molekula [128].
A Shigellak szintén közeli, főemlősöket fertőző rokonai a fenti enterobaktereknek [129]. A
normál bélflóra ecetsav és hangyasav termelésével szorítja vissza a shigellózis (S. flexnerii)
kialakulásának lehetőségeit.
Végül egy szintén komoly, járványos megbetegedés, a pestis kórokozója (Yersinia pestis) és
rokonai is az enterobakterek közé sorolandók. Újabban antibiotikum (v. baktericid) termelését is
bizonyították olyan kórokozók ellen, mint pl.: Pseudomonas aeruginosa vagy Klebsiella pneumoniae,
vagy éppen Staphylococcus aureus [130].
A gamma-proteobacterek másik fontos csoportja a Pseodomonasoké. A típusnemzettség
(Pseudomonas 3.2.34. ábra) fajai a mikrobiológia hőskora óta ismertek, és számos kórokozó tartozik
közéjük (P. putida, P. aeruginosa, P. fluorescens), de olyan hasznos nemzettségek is, mint az
Azotobacterek, Rhizobacterek, Cellvibriók.
3.2.34. ábra: Pseudomonas sp.
Az Azotobacter vinelandii különleges aerob nitrogénfixáló metabolizmussal rendelkezik,
amelynek nitrogenázenzimei eltérnek a szokásos (pl.: Azospirillium) molibdenát központú mehaniz-
mustól. Életformájában is más a megszokott nitrogénfixálókhoz képest: nem növényi szimbionita,
hanem szabadon élő [131]. Több fajuk is (A. vinelandii, A. chroccocum) alginát biopolimert termel, a
barna algákhoz hasonlóan [132].
A Rhizobacterek a Rhizobiumokhoz hasonlóan a hüvelyes növények gyökérszöveteinek endo-
szimbionitája, segítve a növény növekedését megfelelő nitrogéntartalmú tápanyagok ellátása révén (N-
fixálók). Ugyanakkor ezeket a baktériumokat szokás Plant-Growth-Promoting-Rhizobacteriának
(PGPR-növényi növekedést serkentő rhizobaktereknek) nevezni, ahol a pontos támogató mehanizmus

3. Az ipari mikroorganizmusok és technológiák tárgyalása törzsrendszertanba illesztve 91
© Németh Áron, BME © www.tankonyvtar.hu
még nem teljesen ismert, de valószínűleg sziderofórok, fitohormonok segítségével és az ásványianyag-
felvétellel segítik a növény növekedését [133].
A Cellvibrio nemzettség fajai a nevükből is láthatóan cellulózbontók. Köztük a legintenzívebben
vizsgált faj a Cellvibrio japonicus, amelyet japán talajmintából izoláltak 1952-ben. Ennek megfelelően
különböző – a megszokottaktól valamiben eltérő – enzimeiket vizsgálják (arabináz [134], xilanáz
[135] stb.), ezek jelentős alkalmazási potenciállal rendelkeznek a bioipar számára.
A gamma-proteobacterek következő csoportja a Pasteurella-félék, ahová a humánpatogén
Haemophilus influenzae és a házi baromfit fertőző Pasteurella multocida került többek között
besorolásra. Utóbbi biotechnológiai jelentősége, hogy az állattenyésztők számára vakcinaként
inaktivált Pasteurella-sejteket vagy azok antigénjeit forgalmazzák. Hasonló a helyzet az
Actinobacillus pleuropneumoniae esetében, amely a sertésállományt fertőzi, és véres habzó száj/orr,
valamint magas láz mellett az állatok elhullását okozza [136].
A C1-metabolizmusnál már bemutattunk olyan mikrobákat, amelyek a metanol, metán
hasznosítására képesek. A gamma-proteobacterek ezen csoportja a Methylococcus-félék. A csoport
valamennyi nemzetsége metilotróf, metánhasznosító. A névadó nemzettség, amely a metilotróf
kutatások középpontjában áll néhány archeával együtt, a termofil Methylococcus capsulatus [137].
Iparilag több évtizede használt mikrobák a gamma-proteobacterek következő csoportjába sorolt
Xanthomonas-félék, köztük a Xanthomonas campestris, amellyel a széleskörűen (főleg azonban
élelmiszer-ipari célra) hasznosított xantán gumi fermentációs előállítását valósítják meg [141].
A jelen jegyzetben tárgyalt utolsó gamma-proteobacter csoport a Succinivibrio-félék csoportja.
Az idesorolt Anaerobiospirillum succiniproducens okozhat megbetegedéseket (bakterémia) [138], de a
platformalkotó borostyánkősav előállítására is felhasználható [139]. A Succinimonas amylolytica
szintén nagy mennyiségű borostyánkősav előállítására képes, közvetlenül keményítőből kiindulva
[140]. A névadó Succinivibrio dextrinosolvens segítségével heterofementatív tulajdonságának
köszönhetően glükózalapon állíthatók elő borostyánkősav- és tejsav- stb. metabolitok.
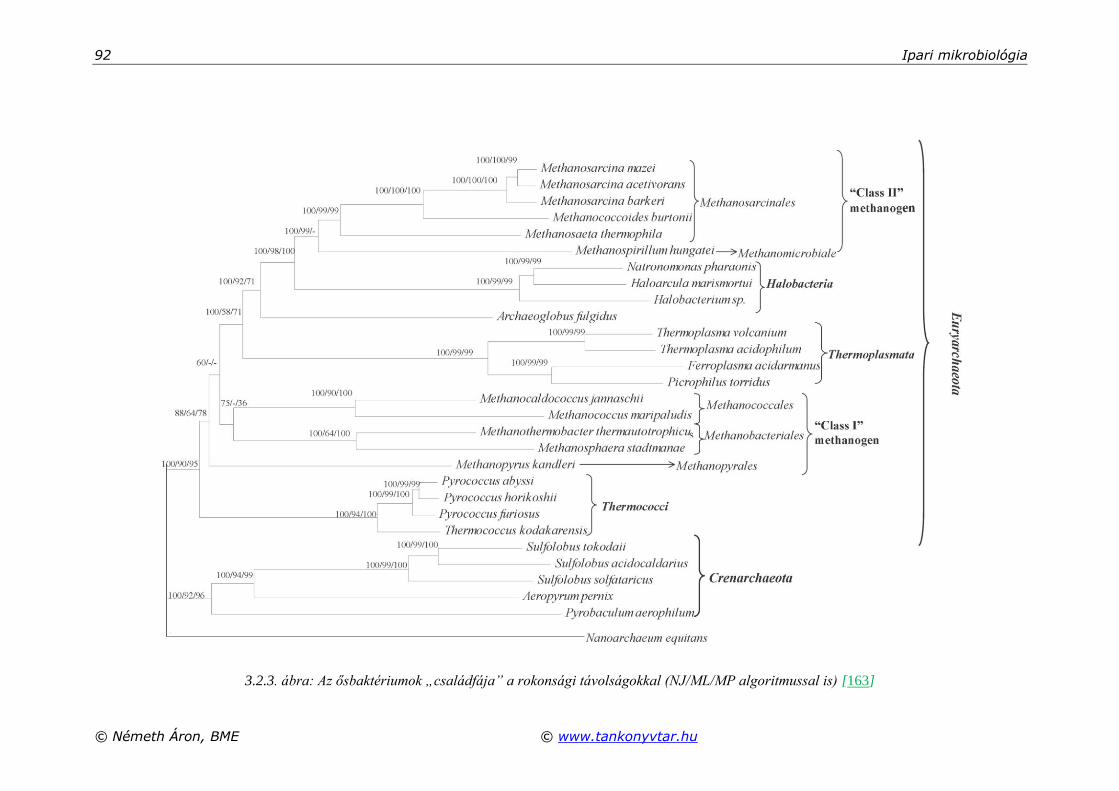
92 Ipari mikrobiológia
© Németh Áron, BME © www.tankonyvtar.hu
3.2.3. ábra: Az ősbaktériumok „családfája” a rokonsági távolságokkal (NJ/ML/MP algoritmussal is) [163]

3. Az ipari mikroorganizmusok és technológiák tárgyalása törzsrendszertanba illesztve 93
© Németh Áron, BME © www.tankonyvtar.hu
3.2.2. táblázat: Bacillusok osztálya
Baktériumok
doménje
Firmicutes
törzs
(alacsony
G+C tart.)
Bacillusok
osztálya
Bacilliales
rendje
(kataláz
pozitív)
Bacillusok
családja
B.subtilis,
B.licheniformis,
B.thüringiensis,
B.coagulans
B. antraxis
Staphylococcusok
családja
S. aureus
Listériák családja Listeria
monocytogenes
Lactobacilliales
rend
(kataláz
negatív)
Lactobacillusok
családja
Lactobacillus
nemzetség
L. casei
L. rhamnosus
L. delbrueckii
L. helveticus
L. coryneformis
Pediococcus
nemzetség
Pediococccus
acidilactici
Leuconostoc
nemzetsége
L.
mesenteroides
Strpetococcus
nemzetsége
S. bovis,
S. pyogenes,
S. thermophilus
Enterococcus
nemzetsége
E. faecalis
Clostridiumok
osztálya
Mollicutes
osztály
94 Ipari mikrobiológia
© Németh Áron, BME © www.tankonyvtar.hu
3.2.11. ábra: Az Actinobacterek családfája (Az NCBI taxonomia adatfile alapján generálva)

3. Az ipari mikroorganizmusok és technológiák tárgyalása törzsrendszertanba illesztve 95
© Németh Áron, BME © www.tankonyvtar.hu
3.3. Ipari szempontból fontos gombák
A gombák rendszertana a szaporodási formák alapján sorolja be a nemzetségeket, fajokat, ezért az
alábbiakban röviden összefoglaljuk a gombák szaporodási folyamatait [149].
3.3.1. animáció: Gombák életciklusai
A gombák ivartalan szaporodása történhet sarjadzással, hifatöréssel, és ivartalan szaporító
képletek (spórák) segítségével, azaz spórázással. Az életciklusuk folyamán ivaros szaporodásra is
képesek, amely során egymástól időben jelentősen elválasztva valósul meg a plazmogámia, kario-
gámia és a meiózis (3.3.1. animáció), így a haploid dikarion és a diploid állapot is stabilan megtalálha-
tó forma. Mindhárom esetben van lehetőség ivartalan esetenként spórás szaporodásra, amely spórák
fenotípusa meglehetősen hasonló, ezért ugyanazon faj különböző állapotú formáit (ivarosan, v.
ivartalanul szaporodó) külön-külön is besorolták a rendszertanba. Ennek következményeként a
korábban elkülönített ivaros (perfect) és ivartalan (imperfect) szaporodású fajok között azonosságokra
derült fény, azaz egy adott fajnak létezik perfect (más néven teleomorph) és imperfect (anamorph)
formája is (legfeljebb még nem ismerjük). Például az anamorph Brettanomyces anomalus teleomorph
formája a Dekkara anomala [150].
A gombák morfológiája is igen változatos, ugyanazon faj különböző állapotai morfológiájukban
is eltérnek: a Saccharomyces cerevisiae pékélesztő egysejtes formában szaporodik szubmerz
körülmények mellett, ám a körülmények megváltozása esetén (szilárd tápközeg, kevés N-forrás)
fonalas növekedésre vált [151].
Mindezeknek köszönhetően a gombák rendszertana morfológiai megfigyeléseken alapult és nem
rokonsági távolságokon, ám az utóbbi évtizedek teljes genomszekvenáló programjainak köszönhetően
fényderült a DNS-alapú rokonságokra, és ezek alapján új rendszertan felállítása kezdődött meg 2007-
től, amely munka a mai napig tart.
Ezek alapján a még nem általánosan elfogadott, de legújabb rendszertan a következők szerint
alakul [153] (3.3.2. ábra). A 3.3.2. ábrán pirossal kiemeltük az iparilag fontos gombákat tömörítő
csoportokat, amelyek a következőkben kerülnek tárgyalásra.

96 Ipari mikrobiológia
© Németh Áron, BME © www.tankonyvtar.hu
3.3.2. ábra: A legújabb gombarendszertan

3. Az ipari mikroorganizmusok és technológiák tárgyalása törzsrendszertanba illesztve 97
© Németh Áron, BME © www.tankonyvtar.hu
Basidomycotak
A basidomycota-k az ascomycotákkal együtt alkotják a Dikarya alkirályságot, amelynek neve a stabil
dikarion állapotra utal, amelyben e két törzs tagjai általában előfordulnak. A basidomycoták
legismertebb formái a kalapos gombák, amelyek felépítését mutatja a 3.3.3. ábra.
3.3.3. ábra: Bazidomiceták felépítése [154]
Nevüket onnan kapták, hogy a termőtestté tömörülő hifák a kalap alján elhelyezkedő lemezeken
bazidiumokat növesztenek, amelyekről rendszerint 4-4 spóra fűződik le.
A kalapos gombák általában az élelmiszeriparban kerülnek piacra, a biotechnológiában ritkábban
hasznosítják őket. Az Agaricomycotina csoportjukba tartozik azonban a Ganoderma lucidum (pecsét-
viaszgomba), amely számos alkalmazással és nagy lehetőségekkel rendelkezik, mivel immunerősítő,
vérnyomáscsökkentő, koleszterincsökkentő hatása van, továbbá javallott vesegyulladás, asztma, gyo-
morbántalmak esetén is. Újabban az AIDS és a rák ellenei küzdelemben is sikerrel alkalmazzák. A széles
körű alkalmazását a mintegy 150 féle triterpén és 50 poliszaharid molekulájának köszönheti [155].
A pecsétviaszgombához hasonlóan a Shii Take gomba (Lentinus v. Lentinula edodes) is az
Agaricomycotak közé tartozik, és az ázsiai hagyományok szerint fogyasztása igen kedvező, mivel anti-
tumor szaharidokat tartalmaz nagy mennyiségben [156].
Az Agaricus brasiliensis fenti rokonaihoz hasonló egészségvédő hatással rendelkezik, és szokásos
biotechnológiai módszerekkel tenyészthető.
További példákat foglal össze a 3.3.1. táblázat a kalapos gombák fermentációs (szubmerz,
bioreaktoros) felhasználására. Amint az a táblázatból is látható, a kalapos gombák fermentációjával
előállított termékek az alábbiak szerint csoportosíthatók.
1) poliszaharidok
Elsősorban az USA-ban, Japánban és Oroszországban nyerik ki vagy a termőtestből, vagy tenyésztett
micélimból, vagy a tenyésztett micélium fermentlevéből. Az így előállított poliszaharidok antitumor-
hatással rendelkeznek, azaz kemotrerápiával kombinálva akadályozzák a rák előrehaladását, és
csökkentik az áttétek kialakulásának esélyét. Nem közvetlen hatást fejtenek ki, hanem a T-sejtes
immunválaszt stimulálják ezek a poliszaharidok. Szerkezetüket tekintve általában β(1-3) kapcsolódású
glükánok, 1-6 elágazással, ami szükséges az antitumorális hatás kiváltásához. A nagy molekula-
tömegűek hatékonyabbak a kisebbeknél. Kereskedelmi forgalomban kapható a lentinán (Lentinus
edodes), PoliSzaharidKrestin (PSK) (Ganoderma lucidum) és a schizophyllan (Schizophyllum
commune).

98 Ipari mikrobiológia
© Németh Áron, BME © www.tankonyvtar.hu
2) terpenoidok
A terpenoidok izoprénszármazékok. Nevüket a terpénről kapták, amely tulajdonképpen izoprén dimer.
A(z) (bio) izoprén új perspektvákat kapott az utóbbi évtizedben, mivel a hagyományos
kőolajkrakkolás mellett rekombináns technikával biológiailag is előállítható polimerizációs és
üzemanyagcélra is [158]! Mivel biológiailag előállítható, megújuló nyersanyagokból is lehet gyártani,
és platformvegyület, tehát származékait egyszerű, konvencionális módszerekkel lehet szintén
előállítani. Ha megfelelő technológiával olcsón elő lehet állítani, üzemanyagcélra is kiváló lehet, mert
magasabb energiatartalommal bír mind a dízelolajnál, mind a benzinnél. A polimer- és üzemanyagcélú
felhasználásán kívül is nagy volumenű alkalmazásnak örvend a gumiipar elasztomerjei, illetve a
ragasztók körében.
Az izoprénszármazék terpenoidokat a szénlánc hossza és az azzal arányos fizikai tulajdonságaik
alapján csoportosítják: illékony mono és szeszkviterpének (sesquiterpenoids, C10-15, esszenciális
olajok), kevésbé illékony triterpének (C20), nem illékony terpenoidok és szterolok (C30), illetve
karotenoid pigmentek (C40). A pecsétviaszgomba (Ganoderma lucidum) által termelt terpéneket
szintén ebben a csoportosításban tárgyaljuk. A ganodermasav A, B, C1, és C2 molekulái az emberi
májsejtek növekedését gátolják, illetve antioxidánsként védik az eritrocitákat és a májsejteket. A
termőtest többi triterpénjével együtt (ganoderiol B, ganodermanontriol) Anti-HIV1 és Anti-HIV1-
proteáz aktivitásúak (előbbi a burokfehérje kötődésének gátlását, utóbbi pedig a virion fehérje
proteolitikus aktiválásának gátlását jelenti). A diterpének között a ciatánokat – amelyek nevüket a
Cyathus helenae madártollról izolált parazita gombáról kapták – számos gomba termőtestjéből
kimutatták, a sphaeroane típusúak pedig algákban is előfordulnak. Idetartozik még a Drosophyla
subatratból előállított antimikrobiális hatású tintinnadiol. A diterépnekre általában is jellemző, hogy
antimikrobiális hatásúak, idegsejt-növekedési faktort indukálnak (NGF), és opioid receptor agonsiták,
aminek a fájdalomcsillapításban van szerepe. A szeszkviterpenoidok és monoterpenoidok szintén NGF
induktorok, inszekticidek és nematicidek.
3) ergoszterol
Az ergoszterol a D-vitamin prekurzora, UV hatására kortizon és progeszteron keletkezik belőle. A
gombáknál a funkciója, hogy a sejtmembrán részeként segítse az integritást és a membrán fehérjék
aktivitását, illetve a sejt energiaháztartását. A kereskedelmi forgalomba kerülő termékeket élesztőből
vagy hulladékmicélium extrakciójával állítják elő.
4) ceramid
Kozmetikumokban nedvesítő ágensként alkalmazzák, de étrend-kiegészítőnek is kitűnő (élelmiszer-
ipar), mivel immunerősítő, rákmegelőző hatást is tulajdonítanak neki.
A bazidomiceták (magasabb rendű gombák) fermentációjának célja, hogy a termőtestekben
megtalálható értékes anyagokat hatékonyan állítsák elő. Ehhez általában talajtenyészeteket alkalmaz-
nak, ami azonban 60-70 napig is eltart, és kevéssé kontrollált. A szubmerz fermentáció fonalas
gombák esetén (kalapos!) a megnövekedő viszkozitás miatt nehézségekbe ütközik, mert oxigénátadási
problémák és magasabb nyíróerő lép fel.
A Ganoderma lucidum esetén 50 g/L alatti (szubsztrát inhibíció miatt) laktózzal vagy szaharózzal
végzik a szekundermetabolit- (termék)előállítást, amely szénforrások azonban a növekedésnek nem
kedveznek. Eközben az oldott oxigén szintjét 20-30% körül célszerű tartani, és így a végső sejt száraz-
anyagtartalom 22 g/L, az elérhető EPS(extracelluláris-poliszaharid) 0,87 g/L, IPS (intracelluláris-
poliszaharid) 2,5 g/L, amely mellett mintegy 300 mg/L GA (ganodermasav) keletkezik. Az optimális
Kla értékét 781/h-ban állapították meg. A 10% körüli oxigénszint az aggregátumokban oxigénlimithez
vezet, ami kisebb mértékű biomassza-képződést, ám magasabb EPS-, IPS-, GA- koncentrációkat
eredményez gyenge produktivitással. Már 20% oxigénszintnél mindez megfordul. A nyíróerő hatását
0,5-1 m/s csúcssebesség tartományban írták le, és megállapították, hogy a pelletméret gyorsabban nő
az alacsonyabb keverés esetén, és ez az IPS-képződésnek kedvez, míg a magasabb keverősebesség
hatására a végső sejt-szárazanyagtartalom (tehát a biomasszahozam is) lecsökent (14->10 g/L-re).
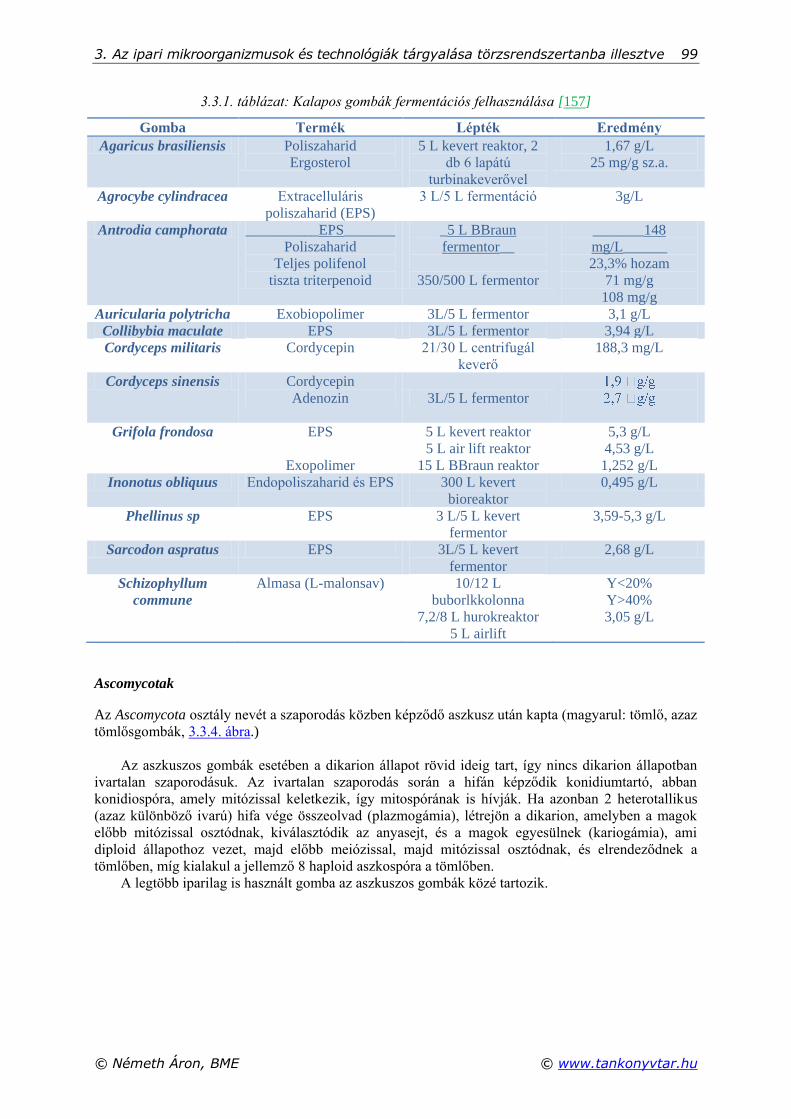
3. Az ipari mikroorganizmusok és technológiák tárgyalása törzsrendszertanba illesztve 99
© Németh Áron, BME © www.tankonyvtar.hu
3.3.1. táblázat: Kalapos gombák fermentációs felhasználása [157]
Gomba Termék Lépték Eredmény
Agaricus brasiliensis Poliszaharid
Ergosterol
5 L kevert reaktor, 2
db 6 lapátú
turbinakeverővel
1,67 g/L
25 mg/g sz.a.
Agrocybe cylindracea Extracelluláris
poliszaharid (EPS)
3 L/5 L fermentáció 3g/L
Antrodia camphorata __________EPS_______
Poliszaharid
Teljes polifenol
tiszta triterpenoid
_5 L BBraun
fermentor__
350/500 L fermentor
_______148
mg/L______
23,3% hozam
71 mg/g
108 mg/g
Auricularia polytricha Exobiopolimer 3L/5 L fermentor 3,1 g/L
Collibybia maculate EPS 3L/5 L fermentor 3,94 g/L
Cordyceps militaris Cordycepin 21/30 L centrifugál
keverő
188,3 mg/L
Cordyceps sinensis Cordycepin
Adenozin 3L/5 L fermentor
Grifola frondosa EPS
Exopolimer
5 L kevert reaktor
5 L air lift reaktor
15 L BBraun reaktor
5,3 g/L
4,53 g/L
1,252 g/L
Inonotus obliquus Endopoliszaharid és EPS 300 L kevert
bioreaktor
0,495 g/L
Phellinus sp EPS 3 L/5 L kevert
fermentor
3,59-5,3 g/L
Sarcodon aspratus EPS 3L/5 L kevert
fermentor
2,68 g/L
Schizophyllum
commune
Almasa (L-malonsav) 10/12 L
buborlkkolonna
7,2/8 L hurokreaktor
5 L airlift
Y<20%
Y>40%
3,05 g/L
Ascomycotak
Az Ascomycota osztály nevét a szaporodás közben képződő aszkusz után kapta (magyarul: tömlő, azaz
tömlősgombák, 3.3.4. ábra.)
Az aszkuszos gombák esetében a dikarion állapot rövid ideig tart, így nincs dikarion állapotban
ivartalan szaporodásuk. Az ivartalan szaporodás során a hifán képződik konidiumtartó, abban
konidiospóra, amely mitózissal keletkezik, így mitospórának is hívják. Ha azonban 2 heterotallikus
(azaz különböző ivarú) hifa vége összeolvad (plazmogámia), létrejön a dikarion, amelyben a magok
előbb mitózissal osztódnak, kiválasztódik az anyasejt, és a magok egyesülnek (kariogámia), ami
diploid állapothoz vezet, majd előbb meiózissal, majd mitózissal osztódnak, és elrendeződnek a
tömlőben, míg kialakul a jellemző 8 haploid aszkospóra a tömlőben.
A legtöbb iparilag is használt gomba az aszkuszos gombák közé tartozik.

100 Ipari mikrobiológia
© Németh Áron, BME © www.tankonyvtar.hu
3.3.4. ábra: Aszkuszos gombák spóraképzése [159]
Az Aspergillusokat (pl.: A. niger) és Penicilliumokat (pl.: P. notatum) széleskörűen alkalmazzák
évtizedek óta. Az előbbieket a citromsavgyártásra az utóbbiakat antibiotikum-termelésre használják.
A Pichiák különleges lebontó enzimekkel rendelkeznek (P. stipitis: xilanázok), és a P. pastoris
még metilotróf is, valamint kiváló gazdaszervezet heterológ fehérjék kifejeztetésre (expressziós
rendszer).
Itt található a kenyérsütéshez és az alkoholgyártáshoz használatos pékélesztő (Saccharomyces
cerevisiae, 3.3.5. videó). Az idesorolt candida fajok általában megtalálhatók a normál humán flórában,
de opportunista patogénekis.
3.3.5. videó: Élesztőtenyészet
Zygomycotak
A zygomiceták esetében csak a haploid alak képes ivartalanul spórákkal szaporodni (konidiospórák).
A 3.3.2. ábrán látható, hogy a rendszertani csoportjuk ipari alkalmazással bíró tagjai a Mucor és
Rhizopus nemzetségek. A 3.3.6. videomikroszkópos felvételen mutatja be a Rhizopus oryzae
micéliumát, konidiumait és konidiospóráit.
3.3.6. videó: Fonalas gomba sporangium

3. Az ipari mikroorganizmusok és technológiák tárgyalása törzsrendszertanba illesztve 101
© Németh Áron, BME © www.tankonyvtar.hu
3.4. Iparilag jelentős algák
Az alga mint gyűjtőnév nem fed rendszertani kategóriát. A prokarióták közül a cianobaktériumok
tartoznak a mikroalgák közé, illetve az eukarióták között mikro- és makroalgákat is találunk. Előbbiek
lebegnek a vízben, utóbbiak többméteresre is megnővén szilárd aljzathoz rögzülnek.
Algasajátosságok közé tartozik a fotoszintetikus képesség, valamint a fotoszintetikus és segéd-
pigmentek. Közös tulajdonságuk továbbá az egysejtű életforma, de különböző szerveződésű szinten
állnak az egyes algák (fonalas, telepes stb.). A különböző pigmentek közös proplasztiszból fejlődnek
ki a környezeti igényeknek megfelelően (algasejt genomja szabályozza), ugyanakkor saját DNS-ük is
van. Megkülönböztetnek kloroplasziszt, amiloplasztot, elaioplasztot (olaj), etioplasztot és kromo-
plasztot.
Rendszerezésük szintén sokáig morfológiai alapon történt (fotoszintetikus pigmentek összetétele,
tartaléktápanyagok, sejtfal szerkezete, ostorok szerkezete és elrendeződése, szemfolt), napjainkban
azonban itt is a DNS-alapú újrarendszerezés zajlik.
Ipari alkalmazásra a cianobacterek közül (prokarióta algák) a már említett Spirullina (pl.:
Spirullina maxima) fajok alkalmasak, részben funkcionális élelmiszerként, részben a biodízel-
technológia nyersanyagának, a lipideknek („növényolaj”) termelőjeként.
3.4.1. ábra: Vörös alga [178]
Az eukarióta algák iparilag is alkalmazott nemzetsége a Chlorella (pl.: Chlorella vulgaris). A
Glaucophyták, amelyek iparilag kevéssé jelentősek, a bizonyítékai a kloroplaszt endoszimbiózissal
magyarázott eredetének, mivel bennük a kloroplaszt még a fagocitózisos membránnal is határolt, így
jelentőségük nem ipari, hanem evolúciótörténeti.
A Rhodopyták (vörös algák, 3.4.1. ábra), már rendelkeznek ipari felhasználással is, például az
agar-agart belőlük (Gellidium amansi) nyerik ki. Karragének gyártására használják a Chondrus
crispust, ír moszatot. A klasszikus zöld algák a felszín közelében szeretnek lebegni, ezért különböző
felületnagyobbító képletet alakítottak ki, amitől igen változatos morfológia jellemzi őket. A
Chlorophyta szintén zöld moszatokat takar, amelyek tömlősgombákkal zuzmót képeznek.
Különleges jelenség a „görögdinnyehó”, amikor enyhén rózsaszínű foltok jelenek meg a hóban a
lábnyomokban. A havon élni (Chlamydomonas nivalis) csak különleges lipidekből álló membránnal
lehet, amely folyékonyabb hidegben, mint melegben. A rózsaszínű pigmenteket az UV sugárzás
kiszűrésére termelik.
Astaxantin termelésére lehetne használni a Haematococcus pluvialis fajt, amely szaporodási
fázisban zöld színű, astaxantin termelésekor pedig vöröses.
A Botriococcus braunii olaj temelése miatt bioüzemanyag-gyártásra lesz felhasználható. Ezt
manapság kétféle rendszerben lehet megoldani: vagy nyitott medencékben, kiszolgáltatva az
időjárásnak, vagy zárt fotobioreaktorokban.
Jelenleg még kevés alkalmazással rendelkenek az igen fejlett csillárkamoszatok (Charophyta). A
sárgás és barnás mosztaok (Ascophyllum nodosum) viszont alginát előállításra használhatók. Végül az
Euglenidek azaz ostoros moszatok (Euglenia viridis) a vízvirágzás leggyakoribb okozói.

102 Ipari mikrobiológia
© Németh Áron, BME © www.tankonyvtar.hu
3.5. Vegyes tenyészetek alkalmazása
A bioszféra minden részén társulások formájában élnek a mikroorganizmusok, és az eddig tárgyalt
alkalmazásokhoz a mikrobilógusok/biotechnológusok erőfeszítések árán jutnak el a színtenyészethez.
Egyes speciális felhasználások esetén – általában olyankor, amikor már van egy specializálódott
vegyes tenyészet – célszerűbb a vegyes mikrobapopulációk használata. Ilyen esetre példa a talaj-
remediáció, bioenergia és zöldkémia egyes területei, szennyvízkezelés, illetve a különböző élelmiszer-
ipari termékek.
A bioremediáció során ex situ vagy in situ távolítják el a talajt ért szennyezéseket azok
lebontásával vagy kevésbé toxikus formába alakítással. A talajok rendelkeznek (a helyi mikrobiótának
köszönhetően) öntisztulással, ám ez igen lassú folyamat, amelyet optimális körülmények megterem-
tésével, esetleg hatékonyabb mikroba (vagy más élőlény) kijuttatásával fel lehet gyorsítani.
A bioremediációra Pseudomonas, Acinetobacter, Citrobacter, Flavobacterium vagy Chromo-
bacter törzseket, illetve ezek keverékét szokás alkalmazni, miközben biztosítják a megfelelő
levegőztetést, N- és P-forrásokat. Cr(VI) detoxifikációjára, redukciójára, akkumulációjára képes az
Arthrobacter oxydans. Különösen jelentős az urán foszfor komplex formájában történő megkötése,
amelyet különböző Citrobacter fajok (C. frendii, C. koseri) képesek elvégezni. Az Acinetobacterek az
aromás szénhidrogének lebontására képesek.
A bioremediációk során gyakran alkalmaznak kometabolizmust. Így a sokáig bonthatatlannak hitt
anyagok is biodegradálhatóak. Ilyenkor általában nem tudják a mikrobák energiatermelésre használni
a bontandó xenobiotkumot (környezetidegen anyag), ezért energiahordozó szubsztrátot kell a
szennyezett területre kijuttatni.
A zöldenergia/vegyipar területén MCB (mixed culture biotechnology) alkalmazására is van
lehetőség, mivel olyan előnyökkel rendelkeznek, mint sterilizálás nélküli termelés, jó alkalmazkodó-
képesség, vegyes szubsztrátok hasznosítása és folyamatosítható eljárás.
A biogáz termelés olyan többlépcsős folyamat, ahol célszerű lehet az MCB-alkalmazás, mivel az
egyes lépcsőket (szubsztrát hidrolízise primer metabolitokig, acetogenezis (H2+CO2ecetsav), illetve
metanogenezis (H2+CO2=>metán (biogáz) <=ecetsav)) más-más mikroba tudja ugyanazon
körülmények mellett leghatékonyabban véghez vinni. A kulcslépés a metán (biogáz) kibocsátása,
amely metanogenesisben ősbaktériumok is részt vehetnek, mint például a Methanococcus jannashi és
Methanosarcina acetivorans.
Az eleveniszapos szennyvíztisztítás során szintén inhomogén kevert kultúra végzi a szennyezések
eltávolítását. Ezek között található habképző Nocardia sp és Microthrix parvicella is. Technológiai
szempontból kényes a populáció egyensúlya, mert ha a fonalas mikrobák szaporodnak el, amelyek
nyálkát is képeznek, akkor megváltoznak az ülepedési tulajdonságok, és kimosódhatnak a mikrobák.
Ostorosok, amőbák is részt vesznek a tisztításban, valamint zsákmánybaktériumok elfogyasztásában.
Fémszennyeződések eltávolítása esetén Klebsiell,a Pseudomonas, Zooglea és Penicillium
nemzetségek alkalmasak, vagy felületi adszorpcióval, vagy citoplazmás felvétellel, olykor illékony
formába alakítással, vagy komplexált kicsapással.
Az élelmiszeriparban szintén gyakran lehet vegyes mikrobatenyészetekkel találkozni. Ezek
jelenléte lehet előnyös (fermentált élelmiszerek), de akár hátrányos (élelmiszerek romlása) is. Utóbbi
oka, hogy az emberi fogyasztásra szánt élelmiszerek a mikrobák számára is ideális tápközegek, ezért
az élelmiszerek közvetítő szerepet játszhatnak a patogén mikrobák terjesztésében is. Egyes mikrobák
jelenléte az élelmiszereken toxintermelésük miatt fokozott veszélyforrás. A romlást okozó mikrobák
jelenléte anyagcseretermékeik folytán jellegzetes szag- és ízanyagokat eredményez.
Ugyanakkor szintén bizonyos (kellemes) szag- és íz anyagok elérése érdekében fogyasztunk
fermentált élelmiszereket is. Ilyen eljárások a savanyítások (káposzta, aludtej stb.), alkoholos
erjedések (szeszes italok), egyes páclevek alkalmazása (szalámi, sonka), illetve érlelési eljárások,
például sajtok esetén.
Az ALKOHOLOS ERJEDÉSEK egyik legrégebben kiaknázott folyamata a borok készítése.
Ezeket az alkoholos erjedéseket a szőlőbogyóról a mustba kerülő vadélesztők keveréke okozza, köztük
apiculatus élesztők (Kloeckera apiculata, Hanseniaspora sp., Brettanomyces sp.), virágélesztők
(Candida sp., Pichia sp.) és nyálkaélesztők (Debaromyces sp., Rhodotorula sp.). A vadélesztők aránya
és aktivitása évről évre változó lehet, így nem lehetne mindig ugyanolyan kiváló minőséget előállítani,

3. Az ipari mikroorganizmusok és technológiák tárgyalása törzsrendszertanba illesztve 103
© Németh Áron, BME © www.tankonyvtar.hu
ezért a nagyobb márkák a vadélesztőket visszaszorítják (kénessav, szeparálás, esetleg pasztőrözéssel),
és saját oltókultúrát (gyakran színtenyészetet) alkalmaznak, amelyek között a Saccharomyces fajok
dominálnak. Különleges folyamat az aszúsodás a boroknál, amelyet a nemes rothasztó Botrytis cinerea
végez úgy, hogy a bogyó behálózásával jelentős vízveszteséget okoz. Érdekesség, hogy a
nemkívánatos szürkerothadást is ugyanez a mikroba okozza.
A húsipari PÁCLEVEK alkalmazásakor általában nitrátos páclevet alkalmaznak (NaCl+KNO3),
amelyekben egyrészt kedvező tejsavbaktériumok (Lactobacillusok), másrészt nitrátbontó mikrobák
szaporodtak el. A minőségi követelményeknek köszönhetően a napjainkban működő nagyüzemek
starter kultúrákat használnak a spontán folyamatok helyett.
A SAVANYÍTÁS-ok egyik fő élelmiszer-ipari területe a tejtermékek gyártása. A frissen fejt
tejben ideális esetben 103-104 db/ml élőcsíra található, amelyek között sem patogének, sem enterális
mikrobák nem lehetnek. A normál tejflóra (Micrococcusok, Lactobacillusok, Streptococcusok) a
fejéskor az állat kültakarójáról és a levegőből kerülhet a tejbe, mivel a mirigyállományban steril tej
képződik. A savanyításért a tejsavbaktériumok felelnek (ld. Firmicutes, C3-metabolizmus) Az
aludtejet a Streptococcus lactis (v. újabban Lactococcus) alvasztja meg, a S. lactis spp aromaticust
pedig acetoin és más ízanyagok termelése miatt vajkutlúrákban alkalmazzák. A S. cremoris a tejföl
jellegzetes tejsavbaktériuma. Magasabb hőmérsékletű érleléseknél (sajtok, joghurtok) S. thermophilus
törzset használnak kevert kultúrában (pl.: sajt: Lactobacillus lactis vagy joghurtok esetében
Lactobacillus bulgaricusszal). A tejtermékek mikrobiotáját foglalja össze a 3.5.1. táblázat.
3.5.1. táblázat: Tejtermékek mikrobiotája
Megnevezés Vajkultúra Joghurtkultúra Kefírkultúra
Mikrobák
Str. lactis
Str. cremoris
Str. diacetilactis
Leuc. citrovorum
Str. thermophilus
Lb. bulgaricus
Str. lactis
Str. cremoris
Lb. casei
Lb. caucasicus
Saccharomyces fragilis
Torula kefyr
Tenyésztési T 20-25 oC 42-45 oC 18-22 oC
Alvadási idő 12-18 óra 2-3 óra 14-20 óra
Kultúra savfoka 36-38 SHo 40-45 SHo 38-40 SHo
Felhasználási
területaludttej, tejföl joghurt kefír
3.6. A vakcinagyártás mikrobiológiája
A kórokozókkal való találkozás túlnyomórészt a gasztroenterális és a légzőszervi szöveteket érinti,
amelyek egészséges élőlényekben átlagos fertőző csíraszám mellet képesek megvédeni a szervezetet
egy fertőzés kialakulásától. Ha azonban valamilyen legyengült állapot vagy sérülés következtében a
szervezet más, belső részeibe kerülnek mikrobák, azaz az első védelmi vonal, a kültakaró mögé, akkor
a második védelmi vonal a vérárammal mindenhol jelen lévő immunrendszer veszi fel a küzdelmet a
kórokozóval szemben. Az immunrendszer egyik részét öröklik az egyedek (veleszületett), a másik
részét pedig az átesett fertőzések során szerzik be (adaptív immunrendszer). Az előbbit makrofágok,
granulociták, „natural-killer” (NK) sejtek és B-sejtes limfociták, utóbit T és B-sejtes limfociták
alkotják. Az immunválasz alapja, hogy a bekerült kórokozó egy speciális, csak rá jellemző molekuláját

104 Ipari mikrobiológia
© Németh Áron, BME © www.tankonyvtar.hu
(antigének: sejtfelszíni fehérje, liposzaharid stb.) érzékelik az immunrendszer fehérvérsejtjei, és
aspecifikus módon megpróbálják likvidáni, vagy legalább izolálni a fertőzést, amíg a specifikus
válasszal elkészül a szervezet. A specifikus választ antitesttermelés formájában valósítja meg az
immunrendszer.
A kórokozók elleni védelem lehet passzív, ha kész antitesteket juttatnak be az oltóanyaggal, illetve
aktív, ha az antitesteket hordozó ágenst (elölt, elgyengített, inaktivált sejt vagy csak az antigén fehérje)
juttatják be az élőszervezetbe. Az utóbbi megoldás hosszú távon hatékonyabb, mert az érzékelt
antigénre a szervezet saját maga tanulja meg az antitest előállítását, így bármikor újra elő tudja állítani,
és egy esetleges újrafertőződést gyorsan kiküszöbölni. Az immunrendszer két részét a 3.6.1. táblázat
hasonlítja össze.
3.6.1. táblázat Az immunrendszer felépítése
Emlős immunrendszere
Veleszületett Adaptív (szerzett)
Funkció
A védelem első vonala,
főleg: patogén érzékelés
(Antigén feldolgozása,
prezentálása, az adaptív válasz
indukciója)
mellesleg: patogén eltávolítása
A védelem második vonala
főleg: patogén eltávolítása,
antigén eltávolítása a
veleszületett rendszer
segítségével
mellesleg: patogén felismerés
evolúciós háttér: ősi, konzervatív fiatalabb
antigénspecifitás kevéssé specifikus nagyon specifikus
előzetes aktiválás nem szükséges szükséges
reakció azonnali késleltetett
memória nincs van
A magyarországi kötelező védőoltásokat a 3.6.2. táblázat foglalja össze.
3.6.2. táblázat: A magyarországi kötelező védőoltások
Beadandó
kor
vakcina
neve
betegség kórokozó baktérium
/vírus
0-6 hét BCG gümőkór, tuberkulózis Bacillus Calmette-Guérin
Mycobacterium tuberculosis baktérium
2 hónap
DTPa
+Hib
+IPV
torokgyík(diftéria),
merevgörcs (tetanusz),
szamárköhögés (pertusis)
+Haemophilus influenzae
+inaktivált poliomyelitis
vírus
Corynebacterium diphteriae
Clostridium tetani
Bordatella pertusis
H. influenzae
polio vírus
bakt.
bakt.
bakt.
bakt.
vírus
3 hónap
DTPa
+Hib
+IPV
torokgyík(diftéria),
merevgörcs (tetanusz),
szamárköhögés (pertusis)
+Haemophilus influenzae
+inaktivált poliomyelitis
vírus
Corynebacterium diphteriae
Clostridium tetani
Bordatella pertusis
H. influenzae
polio vírus
bakt.
bakt.
bakt.
bakt.
vírus
4 hónap
DTPa
+Hib
+IPV
torokgyík(diftéria),
merevgörcs (tetanusz),
szamárköhögés (pertusis)
+Haemophilus influenzae
+inaktivált poliomyelitis
vírus
Corynebacterium diphteriae
Clostridium tetani
Bordatella pertusis
H. influenzae
polio vírus
bakt.
bakt.
bakt.
bakt.
vírus

3. Az ipari mikroorganizmusok és technológiák tárgyalása törzsrendszertanba illesztve 105
© Németh Áron, BME © www.tankonyvtar.hu
15 hónap MMR I.
kanyaró (morbili)
mumpsz (paroitis)
rózsahimlő (rubeola)
Morbili virus (Paramyxo)
Mumps vírus
Measles virus (Paramyxo)
vírus
vírus
vírus
18 hónap
DTPa
+Hib
+IPV
torokgyík(diftéria),
merevgörcs (tetanusz),
szamárköhögés (pertusis)
+Haemophilus influenzae
+inaktivált poliomyelitis
vírus
Corynebacterium diphteriae
Clostridium tetani
Bordatella pertusis
H. influenzae
polio vírus
bakt.
bakt.
bakt.
bakt.
vírus
6 év DTPa
+IPV
torokgyík(diftéria),
merevgörcs (tetanusz),
szamárköhögés (pertusis)
+inaktivált poliomyelitis
vírus
Corynebacterium diphteriae
Clostridium tetani
Bordatella pertusis
polio vírus
bakt.
bakt.
bakt.
vírus
11 év DT torokgyík(diftéria),
merevgörcs (tetanusz),
Corynebacterium diphteriae
Clostridium tetani
bakt.
bakt.
11 év MMR
revakcináció
kanyaró (morbili)
mumpsz (paroitis)
rózsahimlő (rubeola)
Morbili virus (Paramyxo)
Mumps vírus
Measles virus (Paramyxo)
vírus
vírus
vírus
14 év Hepatitis B májgyulladás Hepatitis B virus vírus
Az aktív immunizáláskor alkalmazott vakcinák készítésekor nagyon fontos, hogy a kórokozó
megfelelő molekulája kerüljön kiválasztásra és alkalmazásra. Ugyanazon faj különböző geográfiai
előfordulásai is hordozhatnak különböző antigén molekulákat, ezért különböző szerotípusokba szokás
sorolni a különböző antigénekkel rendelkező, de azonos fajhoz tartozó kórokozókat.
A szerotípus-meghatározás azonban nem olyan hierarchikus, mint a taxonómia, mert például
aVibrio cholerae estében 139 különböző szerotípusú sejtvonal közül csak 2 termelt enterotoxint (ezek
az igazán problémás kórokozók). Ugyanakkor az Actinobacillus pleuropneumoniae esetében 15
szerotípust találtak, amelyek 4-féle toxint termelnek különböző elosztásban. A 15-ös szerotípus
lipopoliszaharidjának O antigénje közös a 3-as és 8-as szerotípuséval, tehát részleges kereszt-
védettséget lehet elérni. Mivel az Actinobacillus pleuropneumoniae toxin (Apx) esetében a IV-es
toxint valamennyi szerotípus termelte, ennek vakcinákban történő alkalmazás indokolt. Megvizsgálva,
hogy milyen védettséget biztosít az Apx IV vakcina, azt tapasztalták, hogy 100%-os védettséget
eredményezett (a baktérium használata 80% körüli védettséget okozott), ugyanakkor a garatmandula
kolonizációját nem tudta megakadályozni, ami továbbfertőzés lehetőségét rejti magában. Ha a toxin
helyett kapszuláris (tok) poliszaharidok kerültek kipróbálásra, csak 70%-os védettség alakult ki, ami a
lipopoliszaharidok esetében tovább csökkent. Ezek alapján belátható, hogy milyen körültekintően kell
kiválasztani a vakcináláshoz megfelelő stratégiát.
Az oltóanyagokat szokás csoportosítani a hatóanyaguk szerint. Így léteznek holt anyagot
tartalmazók (elölt sejt/vírus, inaktivált toxin, transzgenikus/szintetikus antigén), élő, azaz attenuált
(passzált vagy genetikailag deléciós mutáns), élő vektoros (élő vektor pl.: normál flóra tagjai, úgymint
Lactobacillus sp termeli a szükséges antigént a védendő szervezetben), immunkomplex (antitesthez
kötött kórokozó, gyors kolonizáció akadályozva).
Az oltóanyagok lehetnek ún. subunit vakcinák, amelyekben a kórokozó antigéntartalmú részét
használják csupán fel. Általában egyre gyakoribb a transzgénikus subunit oltóanyagok alkalmazása.
Konjugált oltóanyagokat készítenek akkor, ha az antigén nagyon gyenge immunválasz kiváltására
képes csupán. Ilyenkor erős választ kiváltó komponensekhez kötik (pl.:Streptococcus pneumoniae
szerotípus specifikus poliszaharidokat diftériatoxinhoz kötve alkalmazzák).
A vakcinagyártás egyik nagy kihívása a kórokozók előállítása, a másik pedig azok megbízható
inaktiválása. Ez utóbbira szokás kemikáliákat (pl.: formaldehid) vagy fiziko-kémiai kezeléseket (γ-
sugár, β-sugár), esetleg molekuláris biológiai módszereket alkalmazni (pl.: indukálható fág gének
bevitelével lízist idéznek elő).

106 Ipari mikrobiológia
© Németh Áron, BME © www.tankonyvtar.hu
4. A MIKROORGANIZMUSOK IPARI
FELHASZNÁLÁSÁNAK ETIKAI ÉS KOCKÁZATI
KÉRDÉSEI
A biológiai ágensek használatának kockázata két nagy területre osztható: 1) patogén mikrobákkal való
munkavégzés (elsősorban egészségügyi és egyes kutatólaboratóriumok esetében), 2) rekombináns
élőlények használatával kapcsolatos kockázatok. Előbbi esetben közvetlen egészségkárosító
kockázatról beszélünk, míg a transzgenikus szervezetek (GMO) alkalmazásakor a közvetett
(ökológiai) kockázat dominál. A kockázatok felmérése és minimálisra csökkentése mellett mindkét
terület számos etikai kérdést is felvet (fertőzött humán minták kezelése, megbízható GMO termelő
farmerek stb.).
Általánosan elfogadott az az elv, hogy „minden mikroorganizmust potenciális patogénként kell
kezelni”. Ennek oka az, hogy még a legártalmatlanabb ehető mikrobák is a vérbe kerülve szepszist
okozhatnak. A mikrobákat az Egészségügyi Világszervezet (WHO) 4 Biosafety Class-be (BsC Level
1-4) sorolta, és a besorolást külön-külön felállították a vírusokra, gombákra és a baktériumokra is. Az
1-es szintű mikrobák közé az élelmiszer-ipari mikrobák és a normálflóra mikrobái tartoznak
(Lactobacillu-sok, influenzavakcina), a 2-es szintű mikrobák (Clostridium tetani, Candida albicans,
Hepatitis) már patogének, de hatékony védekezési eszköz van ellenük, illetve nem terjednek
járványszerűen; a 3-as jelzésű mikrobák (Bacillus anthracis, Paracoccoidioides brasilensis, HIV) már
komoly megbetegedést okozhatnak, és terjedni is képesek, de a járvány hatékonyan megelőzhető. A 4.
szintre sorolt mikrobák (Myoplasma mycoides, Ebola vírus) a legveszélyesebbek, mivel
gyógyíthatatlan betegséget okoznak, és járványszerűen terjednek. A besorolásból rögtön kitűnik, hogy
időről időre felül kell vizsgálni a besorolást, mivel a tudományos ismertek bővülésével előfordulhat,
hogy a korábban kezelhetetlennek mondott betegség ellen időközben kidolgoztak hatékony
védekezést. Tipikus példa erre a HIV vírus, amely ma már nem a legveszélyesebb 4., hanem a 3.
osztályba tartozik.
A mikrobák kockázatával párhuzamosan a velük dolgozó laboratórimokat is hasonló elvek alapján
osztályokba sorolják. A 4.1.1. táblázatban a javasolt és előírt kockázatcsökkentő feltételeket foglaltuk
össze.
4.1.1. táblázat: A különböző BsC szintre sorolt laboratóriumokkal szembeni követelmények
BsC szint: 1. 2. 3. 4.
labor izolálása - - ~ +
hermetikus
lezárás lehetősége - - + +
szellőztetés be - ~ + +
szellőztetés az
épület
rendszerével
- ~ + -
szellőztetés
elkülönítve - ~ + +
kimenő HEPA-
szűrő - - ~ +
duplaajtós bejárat
(zsilip) - - + +
légzárzuhany - - - +
előtér, öltöző - - + -
előtér zuhannyal - - ~ -
elfolyó anyagok
kezelése - - ~ +

4. A mikroorganizmusok ipari felhasználásának etikai és kockázati kérdései 107
© Németh Áron, BME © www.tankonyvtar.hu
A laboratóriumok második védelmi vonalként működnek, feladatuk a környezet megóvása a
kockázatot jelentő élőlények kijutásától. Az első védelmi vonal a steril fülke vagy a biztonsági fülke,
mivel ezek vannak legközelebb a veszélyforráshoz. Előbbi feladata, hogy a steril munkavégzést
lehetővé tegye, azaz a munkaanyag befertőződését megakadályozza. A biztonsági fülke már nemcsak
az anyagot védi, hanem főleg a felhasználót. A 2. védelmi vonalhoz hasonlóan a lamináris fülkéket is
osztályokba sorolják, amely besorolás nagyjából korrelál a mikrobákkal: I.o. fülkében csak BsC1
mikrobával szabad dolgozni, a II.o. (főleg II/A2-től felfelé) fülke már alkalmas a BsC2 és 3. mikrobák
kezelésére, a III.o.-ba már csak biztonsági fülkék vannak, amelyek BsC3 és 4 mikrobák kezelésére
alkalmasak.
Az ipari mikrobiológia szempontjából a patogén mikrobákra vonatkozó előírások kevésbé
mértékadónak tűnhetnek, mivel a vakcinagyártáson kívül ritkán (vagy egyáltalán nem) használnak
virulens patogén mikrobákat termelési célra. Ugyanakkor az egyre nagyobb méretekben megvalósuló
transzgenikus mikrobát alkalmazó technológiák esetére nagyon hasonló előírások vonatkoznak számos
megfontolás miatt.
A génmódosítás terjedésével a GMO (génmódosított organizmusok) élőlények száma és
mennyisége dinamikusan nő. Igaz ez a haszonnövényekre is, ahol a GMO növényekkel szemben talán
a legnagyobb a társadalmi ellenállás, annak ellenére, hogy a korábbi nemesítési eljárások szintén új
génállományú haszonnövényeket eredményeztek, a pontos genetikai változások ismerete nélkül. Ezzel
szemben a transzgenikus növények létrehozásakor pontosan ismert génátvitelt valósítanak meg egy
adott kedvező tulajdonság megjelenésének érdekében. Meg kell azonban jegyezni azt is, hogy míg a
nemesítés során csak fajon belüli (esetleg rokon fajok közötti) visszakeresztezés (=nemesítés)
történhetett, a modern technológia egészen távoli élőlények közötti géntranszfert is lehetővé tesz (pl.:
hidegtolerancia gén az északi-tengeri lepényhalból búzába), amely miatt a szokatlan gének
kölcsönhatásának eredményeképpen akár váratlan (esetleg káros) folyamatok is létrejöhetnek.
Fenti megfontolásokon felül a GMO élőlények gyakran nagyobb fittséggel rendelkeznek épp a
génmódosítás következtében, amely fittség evolúciós előnyt jelenthet, és így kikerülve a természetbe a
természetes (honos) populációkat visszaszoríthatják7.
Ezen jelenségek váltották ki a világ számos országában azt a törekedést, hogy nemzetközi
egyezményt hozzanak létre a bioszféra diverzitásának megőrzésére (Cartagenai jegyzőkönyv,
2000.01.29), illetve ennek érdekében a transzgénikus szervezetek előállításáról, piacra kerüléséről,
határokon átívelő mozgásáról kölcsönös tájékoztatás megvalósítására.
Hasonló okokból az amerikai Élelmiszer és Gyógyszerügyi Hatóság (FDA) bevezette a GRAS-
(„Gegnerally Recognised As Save”) listát, amelyre csak olyan élelmiszer-ipari és gyógyszeripari
adalékok, hatóanyagok kerülhettek föl, amelyeket: vagy 1) 1958.01.01. óta használnak (tehát kellően
régóta, ahhoz, hogy legyen elegendően nagy fogyasztói létszám), vagy 2) tudományos kísérletekkel
igazolták az ártalmatlanságát, vagy 3) a szakértők egyetértenek a biztonságosságában, általánosan
biztonságosnak ismert.
A mikrobiológiai eredetű adalékok GRAS-listájának egy részét mutatja a 4.1.2. táblázat.
Összefoglalva megállapítható, hogy a mikroorganizmusok ipari alkalmazásakor a lehetséges
kockázatok mérlegelése után a vonatkozó biztonsági előírások betartásával kell eljárni.
7 Szerencsére a piacorientált társadalom túlnyomórészt olyan génmódosításokat végzett eddig, amelyek a termék
minőségét és/vagy hozamát növelték. Ez azonban a termelő szervezet számára plusz teher, azaz inkább evolúciós
hátrány, semmint előny.

108 Ipari mikrobiológia
© Németh Áron, BME © www.tankonyvtar.hu
4.1.2. táblázat: Mikrobiológiai eredetű élelmiszer-adalékok GRAS-listája
§172.155 Streptomyces natalensis és Streptomyces chattanoogensis eredetű Natamycin
§172.325 Saccharomyces cerevisiae eredetű pékélesztő-fehérje
§172.590 Saccharomyces cerevisiae, Saccharomyces fragilis, Candida utilis eredetű élesztő-malátakivonat
§172.620
A Rodophycea osztály Gigartinaceae és Soliericeae családjainak alábbi tagjaiból előállított
karragén (hidrokolloid): Chondrus crispus, Chondrus ocellatus, Eucheuma cottonii, Eucheuma
spinosum, Gigartina acicularis, Gigartina pistillata, Gigartina radula, Gigartina stellata
§172.655 Furcelleran, a Furcellaria fastigiata finomított hidrokolloidja
§172.695 Xanthomonas campestriseredetű xantán gumi
§172.725 Fusarium moniliforme fermetnációjából származó Gibberellin sav
§172.896 száraz élesztő: Saccharomyces cerevisiae, Saccharomyces fragilis; és szárított torula élesztő,
Candida utilis
§172.898 Saccharomyces cerevisiae eredetű pékélesztő glikán
§173.110 Rhizopus niveus eredetű amiloglikozidáz a gélesített keményítő cukorrá történő hidrolíziséhez
§173.120 Aspergillus niger eredetű karbohidrázok és cellulázok kagylók, rákok feldolgozásához
§173.130 Rhizopus oryzae eredetű szénhidrázok a keményítőalapú dextróz előállításához
§173.135 Micrococcus lysodeikticus eredetű kataláz a sajtgyártáshoz
§173.140 Mucor miehei var. Cooney et Emerson eredetű lipázok mint sajtok, zsírok, olajok és tejtermékek
ízfokozói
§173.145 Morteirella vinaceae var. raffinoseutilizer alfa-galaktozidáza a cukorrépa-alapú
szaharózgyártáshoz
§173.150
Mikrobiális tejalvasztó enzimek a sajtgyártáshoz (Endothia parasitica, Bacillus cereus, Mucor
pusillus Lindt illetve módosított Mucor miehei és Aspergillus oryzae, amelyek Rhizomucor miehei
var Cooney et Emerson eredetű aszparaginsav-protáz génjét tartalmazzák
§173.160 Candida guilliermondii mint fermentatív citromsav-előállító mikroba
§173.165 Candida lipolytica mint fermentatív citromsav-előállító mikroba
§173.280 oldószeres extrakciós módszer az Aspergillus niger fermetnlevéből történő citromsav kinyerésére

5. Kitekintés 109
© Németh Áron, BME © www.tankonyvtar.hu
5. KITEKINTÉS
Talán nem szükséges tovább bizonygatni, hogy a biotechnológia egy interdiszciplináris, szerteágazó,
intenzíven kutatott tudományterületből napjainkra a mindennapokat is meghatározó fogalommá vált.
Azt is bemutattuk e jegyzet elején, hogy a biotechnológia szerteágazó területeinek közös pontja a
mikroorganizmus, így az ipari mikrobiológia szorosan összefügg a biotechnológiával. E két terület
jövőjéről is csak egymásra utaltságuk mellett lehet gondolkodni, azaz az ipari mikrobiológia jövője a
biotechnológiai területek és irányok által determinált, ugyanakkor a biotechnológia lehetőségeit a
mikroorganizmusok világa határozza meg. Közös jövőjük még mindig két szálon fut, hiszen a
természetben megtalálható mikrobák és azok különleges tulajdonságainak felkutatása még évtizedekig
eltarthat, figyelembe véve az ismert és a vélt mikrobák számát a Földön, ugyanakkor a rekombináns
termelő törzsek nyújtotta flexibilis és kényelmes lehetőségek nyilván egyre nagyobb teret fognak
hódítani.
Azt gondolhatnánk, hogy a „biotechnológiai forradalom” végét fogja jelenteni, ha a természetben
található összes tulajdonságot és enzimet (azaz az összes létező mikrobát és azok genomját)
feltérképezzük, és a kapott információkészletből gazdálkodva azok végtelen kombinációjával véges
számú tulajdonságból szinte végtelen kombinációjú rekombináns fajt lehet létrehozni, amelyekkel
minden fontos vegyipari eljárás kiválthatóvá válna. Ugyanakkor a protein engineering több évtizedes
gyakorlata jelzi, hogy van lehetőség meglévő tulajdonságok átalakítására, és új tulajdonságok (új
enzimek) létrehozására, ma még főleg meglévő peptidek teljes átszerkesztésével, de idővel talán de
novo előállításával is.
Az a kérdés, hogy a fenti nagy lehetőségekből mi valósul meg a jövőben, aligha válaszolható
meg, hiszen javában tart egy olyan biotechnológiai húzóágazat keresése, amely fenn tudná tartani az
eddigi fejlődést, és további fejlesztéseket generálni. Ilyen potenciális húzóágazat lehet a gyógyszer-
gyártás, a bioüzemanyagok előállítása, a fehér biotechnológia, hogy néhány példát említsünk.
A biotechnológia szkeptikusai napjainkban még könnyen találnak fogást a fejlesztési
törekvéseken, hiszen a kőolajalapú energiaszektor és a vegyipar energia- és anyagigényeinek megújuló
alapokon történő kiváltásához az egész Föld felszínének megművelése is kevés lenne. Az is biztos
azonban, hogy a kőolajkészletek majdani elfogyása után szükség lesz az alternatív lehetőségekre mind
az energia-, mind a nyersanyag-felhasználás területén. Szerencsés esetben az energiaigényt más
további alternatív utakon is ki lehet elégíteni (szél, víz, nap, geotermikus stb.), tehát „csak” a nyers-
anyagigényre kell más biomassza-alapú megoldást találni.
A jövő érdekes kérdése, hogy a fényhasznosító tengeri algák az eddig ki nem használt tengerek
felszínén mennyi biomassza-alapú nyersanyagot, illetve üzemanyagot tudnak majd előállítani. Az
algákkal kapcsolatban is hallani olyan kritikákat, hogy nem elég hatékonyak, ezért a Föld összes
tengere is kevés a jelenlegi és a növekvő energiaigények algákkal történő kielégítéséhez.
Ma még csak remélhetjük, hogy a korlátlannak látszó lehetőségek kiaknázásával sikerül olyan
fejlesztéseket realizálni, amelyekkel elérhetőek az emberiség nagy volumenű igényeinek
kielégítéséhez szükséges termelésihatékonyság-növelés, és a szkeptikusok bánatára hozzájárul majd a
biotechnológia – és a vele szorosan összekapcsolódó ipari mikrobiológia – az emberiség energia- és
nyersanyag-problémáinak megoldásához.

110 Ipari mikrobiológia
© Németh Áron, BME © www.tankonyvtar.hu
6. SZÓSZEDET
acid-fast „saválló”
aerob Oxigénigényes faj.
anaerob Levegőkizárást igénylő anyagcsere vagy faj.
anaerob légzés
Olyan respirációs folyamat, ahol a terminális
oxidációban a végső elektron akceptor nem a
légköri oxigén, hanem valamilyen más szervetlen
molekula (pl.: szulfát)
anaplerotikus
=feltöltő, egyes reakcióutak ciklikusan működnek,
és ha a ciklus tagjai a termékvonal felé mozdultak,
akkor pótolni kell őket a ciklus számára.
antimetabolit rezisztencia
Az antimetabolit egy olyan molekula, amely egy
adott metabolitot specifikusan megköt, így az nem
áll rendelkezésre a sejt számára. Amely sejtek erre
rezisztensek, feltehetően több metabolitot
termelnek, mint amennyi antimetabolit a
rendszerben van, tehát lehetnek jobb termelők.
Arisztotelész
Ókori görög filozófus (Kr. e. 384–322),
kiemelkedő tudós, több tudomány és az európai
kultúra atyja, az első „rendszertan” megalkotója.
Bakteriocin
Baktériumok által termelt fehérjeszerű (peptid)
toxin, amely hasonló baktériumok növekedését
gátolja
biobányászat
A bányászat célja a fémek kinyerése ásványaikból.
Egyes kőzetek fémtartalma, illetve keverék volta
nem teszi lehetővé a hatékony fémkinyerést, ám
mikrobiális folyamatok következtében egyes
fémek kioldhatók, és a híg oldatból kinyerhetők
gazdaságosan.
biokontroll
Agrárkifejezés, amely a kártevők szabályozását
(csökkentését) jelenti természetes ellenségük
(valamilyen más biológiai rendszer) segítségével.
citokin
A citokin név polipeptid regulátorok nagy és
vegyes családját takarja, amelyet a szervezetben
különböző embriológiai eredetű sejtek termelnek
(nem csak az immunrendszer sejtjei).
COSY-NMR
=Correlation spectroscopy, molekula- szerkezet
felderítésére használatos két- dimenziós NMR
spektroszkópia. Egy két- dimenziós NMR-mérés
egy sorozat egydimenziósból áll, amelyek rádió
frekvenciás pulzusok sorozatából állnak
késleltetési periódussal a pulzusok között. Ezeket
az egydimenziós sorozatokat az időzítés, a
frekvencia és az intenzitás különbözteti meg
egymástól. Némely késleltetés alatt, az atommag

6. Szószedet 111
© Németh Áron, BME © www.tankonyvtar.hu
spinek foroghatnak egy adott ideig, amit evolúciós
időnek hívnak. Az utolsó pulzus után a magok a
frekvenciát meghatározzák. Az evolúciós idő
léptetésével kétdimenziós adatok generálhatók az
egydimenziós adat- sorozatokból.
crabtree effektus
Egyes mikrobák (főleg élesztők) estében jellemző,
hogy a túl magas cukor (C/E-forrás) mennyisége a
korlátozott oxidációs kapacitású sejtekben
megfelelő levegőztetés ellenére is anaerob
anyagcserére vált ki.
destruxinok
Ciklikus gomba (pl.: Metharisium anisopliae)
eredetű toxinok, amelyek depsipeptidekből állnak
(peptid kötés helyett helyenként észter kötés van),
és a gerinctelenekben elnyomják az immunválaszt.
diszaharidok
Két monomercukor-egységből felépülő
cukormolekulák, pl.: laktóz (1db glükóz+ 1db
galaktóz), maltóz, cellobióz, szaharóz stb.
diszproporció egyidejű oxidációsszám-növekedés és -csökkenés
egy adott reakció következményeként.
diverzitás =sokféleség
ektomikoriza Fonalas gombák növényi gyökerekkel alkotott
szimbiózisa.
előre/vissza csatolás
szabályozási mehanizmus, amikor egy hálózat
előbbi elemei valamelyik későbbi tagra hatnak
(=előre), például egy metabolit gátolja az őt
tovább alakító enzimek egyikét. A visszacsatolás
esetén az előbbi elem egy még korábbira van (pl.:
gátló) hatással.
enniatinok
Olyan ionofór antibiotikumok, amelyeket fonalas
gombák (pl.: Fusariumok) termelnek, és
depsipeptidből (peptid kötés helyett nelyenként
észter kötés van) állnak.
epidermális növekedési faktor
Az emberi sejtek növekedését, proliferációját,
differenciálódását segítő 53 aminosavból álló
oligopeptid.
expressziószintű szabályozás
Egy adott reakcióútvonalat vagy funkciót
katalizáló enzim(ek) génjének ki/be kapcsolásával
történő szabályozás
expressziós rendszer
Az expressziós rendszerek olyan módosított
mikrobák, amelyek képesek nagy koncentrációban
bioaktív formában (helyes elsődleges,
másodlagos, harmadlagos szerkezettel) idegen
fehérjéket a fermentlébe extracellulárisan
kifejezni, miközben eredeti proteáz aktivitásukat
lecsökkentették (és ezáltal a termék lebontás
csökkent).
extrém termofil Extra magas hőmérsékletet kedvelő.
Extrémofilek Extrém körülmények (pH, hőmérséklet,
sókoncentráció, nyomás) mellett növekvő

112 Ipari mikrobiológia
© Németh Áron, BME © www.tankonyvtar.hu
mikroorganizmusok.
Extrémozim
Extrém körülmények (pH, hőmérséklet,
sókoncentráció, nyomás) mellett aktív enzimek,
fehérjék.
fakultatív anaerob Az anaerob/aerob körülményeket kb. egyformán
szeretik.
fehér biotechnológia
Vagy másnéven "ipari biotechnológia" a
biotechnológia azon ága, amely valamely
vegyipari technológiát képes bioipari eljárással
(enzimes, mikrobiális) technológia szinten
kiváltani.
fehérjeszintű szabályozás
Fehérjék kölcsönhatása révén történő szabályozása
egy reakcióútnak (pl.: egyik fehérje a másik
kofaktora vagy inhibítora, esetleg aktivátora stb.)
Fenotipizálás Fenotípus (a genetikai információ alapján
megjelenő tulajdonság) megállapítása.
fermentatív anyagcsere
Olyan metabolizmus, amelyben az elektron- donor
és az elektronakceptor-molekula is szerves
molekula (pl.: cukorból alkohol erjesztése).
fermentatív faj Erjedést végző faj
filogenetika
Rokonságon alapuló leszármazással foglalkozó
tudomány, amelynek modern formája genetikai
eszközök (génazonosság, mutációs mintázat
alapján) segítségével deríti fel a rokoni
kapcsolatokat.
fitoalexinek Növények által (baktériumok ellen) termelt
antibiotikus hatású molekulák.
fitonematódák növényi fonálférgek, élősködők
fitotoxin Növények ellen termelt mikrobiális méreg.
fittség
A transzgénikus élőlények rátermetségének
jelzésére használatos fogalom, amely az új
tulajdonság evolúciós előnyét méri, és így a
kockázatelemzés első lépése.
formózreakció
A cukorszintézis első lépése, amelyben 2
formaldehidből formóz jön létre, majd erre újabb
formaldehidet addicionáltatva glicerin-aldehid
keletkezik.
foszfotranszferáz rendszer (PTS)
A szubsztrát szintű foszforilációt végző
enzimrendszer, amely foszforcsoportot
addicionáltat a szubsztrátmolekulára (aktiválja azt)
a fszfoenol-piruvátról. Ennek következtében válik
lehetővé a szubsztrát transzportja sejt belsejébe.
GC, HPLC Analitikai (kromatográfiás) módszerek.
halotoleráns
=sótűrő, általában a fiziológiás sókoncentrációnál
magasabb, ozmotikusan veszélyes
sókoncentrációkat elviselők gyűjtőneve

6. Szószedet 113
© Németh Áron, BME © www.tankonyvtar.hu
heterofermentatív
Egy erjesztés vagy fermentáció akkor
heterofermentatív, ha több fajta termék keletkezik
(heterogén termékprofil).
hexózok Hat szénatomból álló cukorvegyületek
(szaharidok), például glükóz, fruktóz, galaktóz stb.
hifa Fonalas gombák többsejtű, differenciálatlan
sejtlánca (fonala).
homofermentatív
Egy erjesztés vagy ferementáció akkor
homofermentatív, ha egyfajta termék keletkezik
(homogén termékprofil).
hozam
hatásfok jellegű mennyiség, amely azt mondja
meg, hogy a szubsztrát mekkora hányada került be
a biomasszába (biomassza-hozam), vagy a
termékbe (termékhozam). Mértékegysége % vagy
g/g. Ha a szubsztrát- fogyás helyett a bemért
szubsztrát mennyiségével számolunk, a hozamhoz
hasonló, azonban értékileg eltérő mennyiséget, a
kihozatalt vagy kitermelést kapjuk.
idiofázis
A mikrobiálistermék-előállítás hanyatló
sejtnövekedéssel, és növekedéshez nem kapcsolt
termékképzéssel jellemezhető szakasza.
Inclusion body
A prokarióták zárvány testje, ahová a
használhatatlan (pl.: heterogén fehérje), illetve a
későbbi használathoz elraktározott anyagokat a
sejtek kiválasztják.
Indel =in(szert)+del(éció), azaz inszercióval, vagy
delécióval létrejövő mutáció.
inszekticid =rovarölő
ionofór
Általában zsíroldható molekula, amelynek
funkciója az ionok szállítása a sejtmembránon
keresztül.
izotopomer Olyan izomerek, amelyek az izotóp atom helyében
különböznek egymástól.
katabolit represszió
Biológiai szabályozási mehanizmus, amely
lehetővé teszi az anyagcsereenzimek szintézisének
gátlásával, hogy mindig a legkedvezőbb
szubsztrátot fogyasszák el előbb a
mikroorganizmusok. (A másika(ka)t hasznosító
enzimek szintézise gátolt.)
kemolito-autotrófok
Szénforrásként légköri CO2-t használnak, energia
forrásként szervetlen vegyületek oxidációját
aknázzák ki.
kenet „kenet” a kén (mint anyag) tárgyragozott (régies)
formája, cserélhető „ként”-re
kofermentáció "=együtt fermentáció", két mikroba vagy két
szubsztrát egyidejű alkalmazása egy tenyészetben.
kólsav Vízoldhatatlan szteránvázas epesav, sói a kólátok.

114 Ipari mikrobiológia
© Németh Áron, BME © www.tankonyvtar.hu
kommenzalista
olyan kapcsolat más (gazda) élőszervezettel,
amely kapcsolat a gazda számára semleges, a
kommenzalista számára pedig előnyös (pl.: a
bőrflóra egyes tagjai)
kompetitív receptor antagonista Olyan molekula, amely kapcsolódni képes a
receptorhoz, de nem aktiválja azt.
leaky mutáns
Olyan mutáns, amely csökkent növekedésű, de
nem hiánymutáns. Ennek oka valamely
fiziológiailag igényelt enzimes reakció csökkent
aktivitása.
magnetotaktikus Mágneses teret magnetit ásványok füzérének
segítségével érzékelni képes mikroba.
makrolid antibiotikum
14, 15, vagy 16 tagú makrolid laktongyűrűt
tartalmazó hatóanyagok, jellemzően fehérje
szintézist gátló antibiotikumok.
malo-laktikus fermentáció
A tejsavbaktériumok által végzett olyan erjedés,
amely során az erősen savas almasav (=malonsav)
az enyhébb tejsavvá és széndioxiddá alakul.
Maximum likelihood (ML) módszer
A filogenetikus analízis egyik módszere, amely
standard statisztikai módszerekkel keresi a
legvalószínűbb leszármazási variációt.
Maximum parsimony (MP) módszer
A filogenetikus analízis egyik módszere, amely a
legkevesebb evolúciós variációt keresi, és fogadja
el.
metabolit Anyagcsere következtében keletkező molekula.
metanogének A szén-dioxidot (nem fotoszintézis útján)
hasznosító mikrobákat nevezik így.
metilotrófok A szén-dioxidon kívüli összes egy szénatomú
szénforrást hasznosító mikrobákat nevezik így.
mezofill Közepes (általában hőmérsékleti) tartományt
kedvelő élőlény.
mezofill Hőmérsékleti optimuma 15–40°C esik.
MIC =MinimalInhibitoryConcentration az a dózis, ami
már hatékony inhibíciót okoz.
microarray technika
Nagyszámú kísérletek végzésére alkalmas
módszer, amely egy furatokkal ellátott lemez
gyors leolvasásán és kiértékelésén (mikroplate
reader) alapul.
mikolsav
A mycobacteriumok sejtfalában található hosszú
szénláncú zsírsav, tehát nem a miko=gomba
elnevezésből származik, noha az actinomyceták
közé sorolt Mycobacterium fonalakat képes
növeszteni.
mutagének Mutációt előidéző hatások, ágensek (lehetnek
kémiaiak (vegyszerek), fizikaiak (pl.: sugárzások).
Neigbhour-joined (NJ) módszer A filogenetikus analízis egyik módszere, amely

6. Szószedet 115
© Németh Áron, BME © www.tankonyvtar.hu
genetikaitávolság-mátrix alapján az egymáshoz
legközelebbi rokonokat keresi meg, majd a közös
elágazási pontjukat helyezi el a többi, távolabbi
rokon között.
objektív függvény
Az optimálások során egy "legjobb" értéket
keresünk. Az a függvény, amelynek értékei között
keressük a legjobbat (optimálisat), az objektív
függvény.
obligált anaerob Szigorúan anaerob körülmények mellett növekvő.
oligotróp Tápanyagszegény élőhely
opportunista patogén
Olyan életforma, amely átlagos
gazdaszervezetekben nem okoz megbetedegedést,
csak annak legyengülése esetén.
Pasteur-effektus
Olyan mikrobiális jelenség, amely során az
elégtelen levegőztetés miatt a mikroba anaerob
anyagcserére vált oxigén jelenlétében is.
pentózok Öt szénatomból álló cukorvegyületek
(szaharidok), például xilóz, arabinóz stb.
peszticid =„kártevőírtó szer”
platformalkotó vegyület (platform kemikália)
Olyan vegyületekre használjuk, amelyeknek egy
sor piacképes, értékes származékát konvencionális
vegyipari folyamatokkal elő lehet állítani.
poliextrémofil
Nemcsak egy paraméter extrém értékeit, hannem
többét is elviseli. Pl.: nemcsak acidofil, hanem
emellett halofil, termofil is egy adott élőlény.
polifilatikus Nem egy közös őstől, hanem több őstől
származtatható törzs, faj.
porozitás
A porozitás vagy hézagtérfogat az üreges szilárd
anyagok jellemzője, amely megadja a hézag
térfogatok arányát az össztérfogathoz képest (pl.:
porszemcsék közötti légtérfogat a porhalom
térfogatához képest, vagy porszemcsében a
hézagtérfogat a porszemcse térfogatához képest.)
Pregnánok
Szteroidok egy csoportja, amelyek a C1-en és C21-
en szubsztituáltak. Pregnánokra (telített pregnán
származékok) és pregnadiénekre (telítettlen
pregnánszármazékok) oszthatók , utóbbiak kettős
kötést is tartalmaznak. Előbbiekhez tartoznak a
kortizon, hidrokortizon, progeszteron.
primer metabolit
a mikrobák normál életfolyamatainak
következtében keletkező, általában szerves
molekula, amely tehát általában növekedéshez
kapcsolt termékképzés.
prodigiosin
Pyrrolylpyrromethane vázú vöröspigment-
molekula, antifungicid, antiproliferatív,
immunszupresszáns, Serratiák termelik.

116 Ipari mikrobiológia
© Németh Áron, BME © www.tankonyvtar.hu
produktivitás
Az adott eljárás termelékenységét méri. A
volumetrikus produktivitás (g/l/h) megadja, hogy
térfogategységenként (L) és időegységenként (h)
hány g termék (termékproduktivitás), illetve hány
g biomassza (biomassza produktivitás) keletkezik.
propagula Szaporítóképlet, elsősorban a növényeknél
használják.
proteinogén aminosav Fehérjefelépítő aminosav.
Proteus Homéroszi hős, aki Posseidon fókáit nevelte, és a
végtelen átalakulás képességét kapta ajándékba.
random mutagenezis+screening
Törzsfejlesztési módszer, amely a hatékonyabb
törzs „készítése” érdekében mesterséges
véletlenszerű mutáltatást végez a szülői
generáción (random mutagenezis), majd a
leánysejteket szűrővizsgálattal minősíti, és
kiválogatja (screening).
Reichstein szubsztrát = 17,21-dihidroxi-progeszteron
rendszertan (taxonómia)
Az élőlények rendszerezésével foglalkozó
tudományág. Újabban nem a fenotípusos
hasonlóság, hanem a genotípusos
azonosság/különbözőség alapján (az evolúció
rekonstruálásával) állapít meg rokoni
kapcsolatokat fajok között, és helyezi el őket
egymástól függő távolságok alapján.
replika módszer
„Petri-csésze-másolás”: a másolandó petrit, fejjel
lefelé ráfordítják egy bársonnyal borított fabakra,
finoman megnyomják, majd leveszik, és egy
másik, steril agarlemezt tesznek fel. Így pont oda
és pont ugyanolyan mikroba kerül , mint ahogy a
master petrin volt.
respiráció
Légzés, amely során a megelőző anyagcsere-
utakon felhalmozott redukált koenzim (pl.:
NADH2) regenerálódik, miközben egy szervetlen
molekula (aerob esetben a légköri oxigén)
redukálódik.
retroaldol elimináció Az aldol (C-C kötést kialakító) reakció visszafelé,
azaz C-C kötést bontó reakció.
reverzió Pálcikák osztódása kokkuszokká
(Micrococcusok).
RGCs „Rare Genetic Changes” – ritka genetikai
változások.
SCP
Single Cell Protein, azaz Egysejtfehérje. Humán
és állati táplálékkiegészítő, a fehérjeszegény
táplálékok feljavítására.
szaprofita Elhalt szerves anyagot lebontó életforma
szekunder metabolit Kedvezőtlen körülmények esetén a mikrobák a
konkurencia elnyomásával probálnak előnyre szert
tenni, és ehhez a másodlagos (szekunder)

6. Szószedet 117
© Németh Áron, BME © www.tankonyvtar.hu
anyagcsere-termékeket állítják elő, és bocsátják ki
a fermentlébe. Anyagcsere szempontjából a primer
metabolitokból épülnek fel a szekunder
metabolitok is.
szelekciós nyomás
A tenyésztési körülmények olyan megválasztása,
amely mellett csak a kívánt célmikroba nő ki, a
többi mikroba visszaszorítás alatt van, viszont a
célmikrobák kinöesztése maximálisan elősegített.
szigmafaktor
Genetikai generális szabályozó fehérje, amely a
prokarióták transzkripcióját képes elindítani vagy
megakadályozni.
szubsztrát/termék inhibíció Nagy termék/szubsztrát koncentráció okozta
lassulás a növekedésben.
TCA TriCarboxilic Acid, vagy citrá kör. (ld. 1.1.4. ábra,
2.1.2. hyperpicture)
termofil Hőmérsékleti optimuma >55°C
termotoleráns Alacsonyabb hőmérsékleti optimum, de képes
magasabb hőmérsékleten is nőni.
tetrád Egyes gömb alakú baktériumok (coccusok) 4-
esével összeállnak és tetrádot alkotnak.
trofofázis A mikrobiálistermék-előállítás sejtszaporodással,
növekedéssel jellemezhető tenyésztési szakasza.
ubiquiter Széles körű előfordulás.
vegetatív sejt Aktív metabolizmussal és életfolyamatokkal
rendelkező sejt.
végtiter A folyamat végére elért koncentráció.

118 Ipari mikrobiológia
© Németh Áron, BME © www.tankonyvtar.hu
7. ÖNELLENŐRZŐ KÉRDÉSEK AZ IPARI
MIKROBIOLÓGIA CÍMŰ JEGYZETHEZ
(A vastaggal szedett kérdések esszék, a kérdés alatt javasolt szempontok segítik a válaszolást.
A tesztkérdéseknél mindig 1 helyes válasz található)
1. fejezet
1. Mi a kapcsolat a biotechnológia és az ipari mikrobiológia között?
A) Az alkalmazott színkódrendszer
B) A technológia, mivel mindkettő e fogalomra fókuszál
C) A két területen alkalmazható műveletek halmaza.
D) A mikroorganizmus, amelynek ipari alkalmazásaival az ipari mikrobiológia foglalkozik.
2. Mely tudományok határterülete az ipari mikrobiológia?
A) Biokémia, mikrobiológia, mérnöki tudományok
B) Biokémia, mikrobiológia, biológia
C) Fiziológia, mikrobiológia, kémia
D) Anatómia, mérnöki tudományok, kémia
3. Milyen szerepe van a biofinomítóknak az ipari mikrobiológiában?
A) Az ipari mikrobiológia a biológia finomított ága, ezért az maga a biofinomító
B) A biofinomító olyan létesítmény, ahol finomkémiai kutatások mellett mikrobiológiai
fejlesztéseket végeznek
C) A biofinomítókat a fehér biotechnológiában alkalmazzák, általában mikrobiális fermen-
tációkon alapulnak (=ipari mikrobiológia)
D) E két kifejezés egymás anagrammája
4. Mutassa be a biofinomítók 3 generációját!
A) Ókori (borászatok), középkori (sörfőzés), újkori (pálinkafőzés)
B) 1-fix paraméterű (nyersanyag, technológia, termék); 2-változó termékpaletta (többi fix); 3-
változtatható alapanyag, technológia, termékpaletta
C) 1-bioetanol-üzem; 2-biodízel-üzem; 3-Biogáz-üzem
D) 1-szénkörforgalomra neutrális; 2-nitrogén körforgalomra neutrális; 3-foszfor körforgalom-
ra neutrális
5. Részletesen mutasson be egy sikeres példát az ipari mikrobiológia történetéből!
(termék, termelő törzs (izolálása), klasszikus és modern törzsfejlesztés)
6. Miért kellett leaky mutánsokat fejleszteni a lizintermeléshez?
A) Hogy „kicsurogjon” a termék lizin.
B) Hogy „áteresszen” a kiütött gén terméke (enzim), és minimális aktivitással megakadá-
lyozza a letalitást.
C) Hogy „áteresszen” a kiütött gén terméke (enzim), és minimális aktivitással elősegítse a
letalitást.
D) Mert a melléktermékek „elfolyatásával” javul a termékhozam.

7. Önellenőrző kérdések az Ipari mikrobiológia című jegyzethez 119
© Németh Áron, BME © www.tankonyvtar.hu
7. Milyen géntechnológiai törzsfejlesztés vezetett a fermentációs az 1,3-propándiol előállításhoz?
A) Rekombináns glicerin előállítása fermentációval.
B) A glicerin- és propándiol-termelő útvonalak egy gazdaszervezetbe integrálása.
C) A propándiol-termeléshez szükséges gének glicerintermelőkbe klónozása.
D) A glicerintermeléshez szükséges gének propándiol-termelőkbe klónozása.
8. Mutassa be és magyarázza el az izolálási művelet lépéseit!
(Például antibiotikum-termelők esetében az élőhelytől a profitig vezető utat!)
9. Hasonlítsa össze a sugárgombák, fonalasgombák és az élesztő(gombá)k izolálását
(Élőhely, azaz izolálás forrása, mintavétel, előkészítés, szelektív adalékolás, inkubálás szem-
pontjából.)
10. Mi az a screening, és milyen szempontokat kell figyelembe venni a végzésekor?
A) A képernyőn „screen” való megjelenítése a videomikroszkópnak, amely során a felbontást
kell szem előtt tartani.
B) Szűrővizsgálat, amely során minden potenciális pozitív választ (keresett tulajdonságú), de
egyetlen negatívválaszt sem szabad figyelembe venni.
C) Szűrővizsgálat, amely során a szűrők integritását vizsgáljuk.
D) Olyan mikrobiológiai művelet, amellyel a használhatatlan mikrobákat kiválasztjuk és
megsemmisítjük.
11. Ismertessen 3 olyan screeninget, ahol tisztulási zóna az indikáció!
A) Inzulintermelők, citromsavtermelők, antibiotikum-termelők
B) koleszterintermelők, lipáztermelők, proteáztermelők
C) DNS-polimeráz-termelők, tejsav-termelők, propionsavtermelők
D) celluláztermelők, tejsavtermelők, antibiotikum-termelők
12. Ismertesse a HighThroughputSystem működését egy kitalált példán keresztül!
(Scale down, mikrotenyésztés és –mérés elve, indikációs lehetőségek)
13. Mutasson be 3 ipari példát sikeres szűrővizsgálatra!
A) Hialuronsav-, epesav-, húgysav-előállítása
B) 7-aminocephalosporánsav, compactin-pravastatin biokonverzió, β-laktamáz-inhibítor
előállítása
C) Kapronsav-, linolénsav-, aszkorbinsav-előállítás
D) eritróz-, glicerin-, oktanolgyártása
14. Hasonlítsa össze a klasszikus és modern identifikációt!
(vizsgálat elve, alapja, leírások pontossága, felállított rendszertan diszkrepanciái)
15. Magyarázza el a törzsfejlesztés lényegét és lépeseit!
(Miért kell és mit kell megváltoztatni, illetve hogyan kell kivitelezni? Klasszikus és modern
módszerek.)
16. Milyen alapanyagokból érdemes Magyarországon tejsavat előállítani? Mi a különbség
köztük fermentációs (mikrobiológiai) műveleti és technológiai szempontból?
(A nyersanyag szénhidrátja és a szénhidrát-hidrolízisek kapcsolata a fermentációval.)

120 Ipari mikrobiológia
© Németh Áron, BME © www.tankonyvtar.hu
17. Milyen kísérleteket célszerű elvégezni tápközeg-optimálás során? Mutassa be a tejsav
példáján!
A) Törzskiválasztás → nyersanyagelőkészítés-vizsgálat → tápközeg-otpimálás →
fermentációs paraméterek optimálása→léptéknövelés
B) Törzskiválasztás → nyersanyagelőkészítés-vizsgálat → tápközeg-otpimálás → →
folyamattervezés szimulációval → fermentációs paraméterek optimálása→léptéknövelés
C) Törzskiválasztás → nyersanyagelőkészítés-vizsgálat → fermentációs paraméterek
optimálása→léptéknövelés
D) Törzskiválasztás → nyersanyagelőkészítés-vizsgálat → tápközeg-otpimálás →
fermentációs paraméterek optimálása→folyamattervezés szimulációval
18. Mi a biokémiai funkciója az elsődleges anyagcseretermékek előállításának anaerob mikrobák
tenyésztése során?
A) NADH2-termelés a glikolízis során.
B) A terminális oxidáció hiányában redukált koenzim regenerálása.
C) CO2-kibocsátás a citrátkör segítségével.
D) Pentózok lebontása a pentóz-foszfát ciklus segítségével.
19. Melyik szubsztrát kedvezőbb az etanolos erjedéseknél: hexózok v. pentózok? Miért?
A) A hexózok, mert esetükben 1:2 arányban képződik az etanol és a CO2.
B) A pentózok, mert esetükben 1:2 arányban képződik az etanol és a CO2.
C) A hexózok, mert esetükben 1:1 arányban képződik az etanol és a CO2.
D) A pentózok, mert esetükben 1:1 arányban képződik az etanol és a CO2.
20. Mi a Pasteur- és a Crabtree-effektus?
A) sok oxigén és kevés cukor esetén anaerob anyagcserére vált az élesztő
B) kevés oxigén és sok cukor esetén anaerob anyagcserére vált az élesztő
C) sok oxigén és kevés cukor esetén aerob anyagcserére vált az élesztő
D) kevés oxigén és sok cukor esetén aerob anyagcserére vált az élesztő
21. Mitől függ, hogy a tejsavas erjedés során a termelő mikroba melyik enantiomert termeli?
A) A felhasznált szubsztrát szénatomszámától.
B) A felhasznált szubsztrát kiralitásától
C) A laktát dehidrogenáz sztereospecifitásától
D) A Lactobacillus sztereospecifitásától
22. Milyen morfológiákkal tenyészthető a Rhizopus oryzae, és melyik mit termel?
A) egysejtű(tejsav), fonalas(alkohol), pelletes(biomassza)
B) egysejtű(alkohol), fonalas(biomassza), pelletes(tejsav)
C) telepes(alkohol), micéliális(tejsav), álszövetes(biomassza)
D) telepes(biomassza), micéliális(alkohol), álszövetes(tejsav)
23. Mely metabolitok a propionsavas erjedés melléktermékei?
A) Almasav, csersav, tejsav
B) Ecetsav, borostyánkősav, tejsav
C) Huminsav, ecetsav, széndioxid
D) Hangyasav, zsírsav, borostyánkősav

7. Önellenőrző kérdések az Ipari mikrobiológia című jegyzethez 121
© Németh Áron, BME © www.tankonyvtar.hu
24. Soroljon fel 3 nehézséget a butanoloserjedés-alapú technológiák kapcsán!
A) Kis szénatomszám, erősen savas karakter, nem illékony termék.
B) Nehezen szűrhető termék, nagy molsúly, alacsony hozam.
C) A termék erős sejtméreg, kis koncentrációban keletkezik, heterofermentatív eljárás.
D) Lassan szaporodó, patogén mikrobák termelik, kis produktivitással.
25. Mutasson be 2 poliol-előállítást mikrobiológiai/fiziológiai/biokémiai szempontból!
A) propándiol előállítása Clostridiummal, xilit(ol)-előállítás Gluconobacterrel
B) szorbit-előállítás Z.mobilis-szal arabinózból, és glicerin(glicerol)-előállítás S.cerevisiae-
vel glükózból.
C) Xilit(ol)-előállítás Pichiával xilózból, és szorbit előállítása Z. mobilisszal glükózból.
D) Xilit(ol)-előállítás Z. mobilisszal xilózból, és szorbit előállítása Pichiával glükózból.
26. Hasonlítsa össze a felületi és szumberz fermentációs eljárást!
(előnyök; hátrányok; alkalmazható szubsztrátok, mikrobák és reaktorok; szabályozható
paraméterek)
2. fejezet
27. Írjon példát C1-szubsztrátra!
A) metanol
B) mentol
C) metil-acetát
D) metil-akrilát
28. Ismertesse a metanolgazdaság működését! Mi lehet a biotechnológia és ipari
mikrobiológia szerepe benne?
(szintetikus lehetőségek, szénkörforgás, metabolikus lehetőségek)
29. Jellemezze a 2 leggyakoribb C2 szubsztrátot!
A) Ecetsav: oxidálhatóenergia-nyeréskor; etil-karbonát: gyakori ásvány.
B) Szénsav: vízben oldható; etilbenzoát: „sejtüzemanyag”.
C) Glikolaldehid: önreplikálódó, etanol: ecetsavvá oxidálható energia nyerésekor.
D) Glikolaktid: tejsav-előállítás; etanol: „sejtüzemanyag”.
30. Hogyan tudják egyes mikrobák a tejsavat szubsztrátként hasznosítani?
A) Piroszőlősavon keresztül propándiollá alakítják
B) Piroszőlősavon keresztül propionsavvá alakítják.
C) Foszfoenol-piruváton keresztül tejcukorrá redukálják.
D) Foszfoenol-piruváton keresztül ATP-vé alakítják.
31. Melyik mikroba rendelkezik széles specifitással a cukoroxidáció területén? (Példa!)
A) Sorbobacter suboxydans (pl.: szorbit-szorbóz átalakítás)
B) Gluconobacter suboxydans (pl.: szorbit-szorbóz átalakítás)
C) Gluconobacter suboxydans (pl.: glükóz-szorbóz átalakítás)
D) Corynebacterium pentosum (pentóz-tejsav átalakítás)
32. Részletesen mutasson be egy példát a bioleaching területéről!
(ásvány, mikroba, energianyerés sztöhiometriája)

122 Ipari mikrobiológia
© Németh Áron, BME © www.tankonyvtar.hu
33. Mit jelent a fungisztatikum és a bakteriocid fogalom? Mely termékcsoportba tartoznak?
A) gombaölő, baktérium növekedést gátló, primer metabolitok
B) gomba növekedést gátló, baktériumölő, primer metabolitok
C) gombaölő, baktérium növekedést gátló, szekunder metabolitok
D) gombanövekedést gátló, baktériumölő, szekunder metabolitok
34. Röviden mutasson be 2 másodlagos anyagcsereterméket az antibiotikus hatás nélküliek közül!
A) Ciklosporin A: ciklikus spóraölő; Aflatoxin: gombaméreg
B) Ciklosporin A: imunszupresszáns; Aflatoxin: gomba termelte méreg
C) Ergot alkaloidok: mellékvesehormonok, Klavulánsav: antibiotikum-rezisztencia
D) Ergot alkaloidok: nemihormonok, Klavulánsav: telítetlen zsírsav
35. Mit takar az enniatin és a destruxin?
A) mikrobiális „ennivalót” és „lebontó”-szert
B) növényi „ennivalót” és „lebontó”-szert
C) ionofór antibiotikumot és ionfór gomba metabolitot.
D) kromofór antibiotikumot és kromofór gomba metabolitot.
36. Mutassa be részletesen a kullancsok ellen használható (mikro)biológiai védekezési
eljárást!
(Kullancsok ellenségei, mikroba hatásmechanizmusa, )
37. Sorolja fel a másodlagos anyagcseretermékek funkcióit, egyet részletesen is ismertessen!
(pl.: termelő mikroba kapcsolata a másik élőlénnyel)
38. Ismertesse a másodlagos anyagcseretermék előállításnak kedvező paramétereket!
A) Lassú keverés, szubsztrát-(Zn, Fe, Mg) inhibíció
B) szubsztrát-(Zn, Fe, Mg) limitáció, magasabb hőmérséklet
C) szubsztrát-(C, N, O) limitáció, magasabb hőmérséklet
D) szubsztrát-(C, N, O) inhibíció, magasabb hőmérséklet
39. Definiálja a következő fogalmakat: Metabolic Engineering, Metabolic Fluxus Analysis,
Metabolic Controll Analysis, Metabolic Network Analysis, Systems Biology, Functional
genomics,
(Mi a hasonló fogalmak közötti különbség? Az adja meg a definíciót.)
40. Miért volt szükség a penicillintermelés fejlesztésekor MCA és MNA-ra? Mik azok?
A) MCA: Metabolic Coefficient Analyisis, MNA: Microbial Network Anal.; Mikroba
átirányítása Penicillin V. felé.
B) MCA: Metabolic Controll Analyisis, MNA: Metabol.Network Anal.; fluxusok átirányítása
Penicillin V. felé.
C) MCA: Microbial Controll Analyisis, MNA: Metabol.Neutral Anal.; Mikroba átirányítása a
Penicillin V. felé.
D) MCA: Mezophilic Controll Analyisis, MNA: Metabol.Network Anal.; fluxusok
átirányítása Penicillin V. felé.

7. Önellenőrző kérdések az Ipari mikrobiológia című jegyzethez 123
© Németh Áron, BME © www.tankonyvtar.hu
41. Írja fel a sztöhiometriai mátrixot és a sebességvektort egy olyan „reakcióhálóra”, ahol A-ból
keletkezik B (1. reakció), B-ből keletkezik C (2. r.), és C-ből keletkezik A (3. reakció). Milyen
vektort lehet ezekkel kiszámolni?
A) vektoraességekreakcioseb,
110
011
101
3
2
1
r
r
r
r
v
v
v
vT
C
B
A
B) vektoraességekreakcioseb,
110
011
101
3
2
1
C
B
A
r
r
r
r
v
v
v
vT
C) vektorasebességekképzodesi,
101
110
011
3
2
1
c
B
A
r
r
r
r
v
v
v
vT
D) vektorasebességekképzodesi,
110
011
101
3
2
1
C
B
A
r
r
r
r
v
v
v
vT
42. Rajzoljon fel a koleszterint! Mutassa meg a mikrobiálisan leggyakrabban módosított pontokat!
A) B)
C) D)
43. Ismertesse a β-redukciót a szteroidok konverziójánál!
(Mely pozícióban gyakori, a reakció élettani jelentősége, példa konverziókra)
44. Miért van szükség ciklodextrinre a szteroidkonverziók során?
A) Hogy az alkaloidok szintézise nagyobb hozammal történjen.
B) Hogy ciklikus dextrinek keletkezzenek a maradék cukorból a feldolgozást könnyítendő.
C) Hogy ciklizálja a dextrineket a könnyebb feldolgozás érdekében.
D) Hogy oldatba vigye a vízben rosszul oldódó szteroidszubsztrátot
45. Soroljon fel 2 Mycobacteriumot, amelyek a szteroidkonverziókban részt vesznek!
A) M. phlei, M. smegmatis
B) M. cholesterus, M. tuberculosis
C) M. pelli, M. smaragdus
D) M. bovis, M. leprae

124 Ipari mikrobiológia
© Németh Áron, BME © www.tankonyvtar.hu
46. Soroljon fel 3 alapanyagot a szteroidkonverziók végzéséhez!
A) Prednizolon, kortizon, progeszteron
B) Sztigmaszetrin, szitoszterin, kampeszterin
C) Ösztron, koleszterin, tesztoszteron
D) METESZ, fitoszterin, kampesztrin
3. fejezet
47. Ismertesse a klasszikus mikrobiális rendszertan legutolsó csoportosításait!
(Hogyan nevezik az egyes csoportokat, melyik redszertan hány csoportra oszt stb.)
48. Miért nem lehet az ősbaktériumokat a Gram-festés szerint egyértelműen besorolni?
A) Mert nincs mureinhálójuk, mégis pozitívan festődnek.
B) Mert van mureinhálójuk, de negatívan festődnek.
C) Mert pszeudomurein-hálójuk van, amely néha pozitívan festődik.
D) Mert külső membrán takarja el a pszeudomurein-hálót, ezért mindig negatívan festődnek.
49. Sorolja fel a klasszikus rendszertani besorolás szempontjait!
A) Makro(sejt)morfológia, mikro(telep)morfológia, szag, szín, csillók száma
B) Makro(telep)morfológia, mikro(sejt)morfológia, szag, szín, biokémiai sajátságok
C) Makro(telep)morfológia, mikro(sejt)morfológia, szag, szín, biokémiai sajátságok
D) Makro(telep)morfológia, mikro(sejt)morfológia, szag, szín, fizikokémiai sajátságok
50. Mutassa be a modern filogenetikai kutatások 3 szintjét!
A) Fajszintű, nemzetségszintű, rendszintű
B) Törzsszintű, osztályszintű, fajszintű
C) Lokális, regionális, kontinentális adatelemzések
D) Genomikai adatelemzés; proteomikai adatelemzés; evolúciós fák összevetése
51. Mutasson be 3 baktériumot a Proteobacterek közül!
(Lehet azonos osztályból (α-ε) vagy különbözőből is.)
52. Melyik rendszertani csoportba (törzsbe) tartoznak a következő, iparilag is használt fontos
fajok: Corynebacterium sp., Clostridium sp., Aspergillus sp. és Rhizopus sp.?
Rendre:
A) Actinomyces; Firmicutes, Ascomyceta, Zygomyceta
B) Archea; Firmicutes, Microsporida, Zygomyceta
C) Actinomyces; Fibrobacter, Microsporida, Saccharomicotina
D) Archea; Fibrobacter, Ascomyceta, Sacharomycotina
53. Ismertesse a gombák szaporodási folyamatait!
(Ivaros, ivartalan, örökítőanyag kópia szám szerint)
54. Mutasson be egy bazidomiceták közé tartozó, iparilag is jelentős gombát!
A) Lentinus edodes: shi take gomba, terpének, immunerősítők előállítása
B) Aspergillus niger: aszkorbinsav előállítása, pelletes morfológia
C) Pichia stipitis: egysejtű eukarióta, C5-hasznosító
D) Ganoderma lucidum: pecsétviaszgomba, terpének termelésére

7. Önellenőrző kérdések az Ipari mikrobiológia című jegyzethez 125
© Németh Áron, BME © www.tankonyvtar.hu
55. Mutasson be 2 ipari terméket, amelyet „kalapos gombák fermentációjával” is elő lehet állítani!
A) ergoszterin (D-vitamin prekurzora); ceramid (kozmetikumokhoz)
B) streptomycin (antibiotikum); lizin (aminosav)
C) kloramfenikol (antibiotikum); ganodermasav (terpenoid)
D) ciklosporin (immunszupresszáns); Anttraciklin (antitumor ágens)
56. Ismertesse a bioizoprént a bazidomicetákkal kapcsolatban!
(izoprén platform, biológiai előálítás, származékai és azok termelői)
57. Mik azok a terpenoidok?
A) Aromás benzolszármazékok
B) Aromás naftalinszármazékok
C) Heteroatomos ciklikus vegyületek
D) Izoprén dimer származékai
58. Hasonlítsa össze az aszkomiceták, bazidomiceták és zygomiceták szaporodási ciklusait!
(Ivaros és ivartalan reprodukció, génállomány száma, az egyes állapotok megléte/hiánya
alapján.)
59. Milyen tulajdonságok alapján sorolták az algákat be a rendszertanba?
A) színük és levélnagyságuk szerint
B) leveleik mintázata alapján
C) méret (morfológia) és tartaléktápanyagaik alapján
D) Biokémiai tulajdonságaik alapján (fermentál, respirál stb.)
60. Soroljon fel 3 különleges plasztiszt az algák sejtfelépítése kapcsán!
A) glükoplaszt, kromoplaszt, protoplaszt
B) amiloplaszt, elaioplaszt, kromoplaszt
C) celluplaszt, protoplaszt, amiloplaszt
D) elaioplaszt, celluplaszt, protoplaszt
61. Hol fordulnak elő a kékmoszatok?
A) mélytengerekben
B) hőforrásokban, lúgos-sós vizekben
C) tengeri üledékben
D) talajvízben
62. Sorolja fel, mire használják a vörös moszatokat!
A) festékiparban (vízfesték)
B) indikátor növénynek (környzetvédelem)
C) agar-agar, karragén, (élelmiszer-előállítás)
D) szennyvíztisztítás (környezetvédelem)
63. Ismertesse a zöld moszatok csoportját!
(Hol találhatók, mire és hogyan használhatók stb.)
64. Hogyan alkalmazhatóak az algák a megújuló energiaforrások terén?
(Mit termelnek, zárt/nyitott rendszerek stb.)

126 Ipari mikrobiológia
© Németh Áron, BME © www.tankonyvtar.hu
65. Sorolja fel, hogy a biotechnológia mely területein használnak mikrobakonzorciumokat!
A) Környezetvédelem, zöld vegyipar, élelmiszeripar
B) Környezetvédelem, gyógyszergyártás, fehér biotechnológia
C) Élelmiszeripar, gyógyszergyártás, műanyagipar
D) Élelmiszeripar, biobányászat, műanyagipar
66. Mutasson be egy remediációban használatos mikrobát!
A) Corynebacterium diphteriae (diftéria megkötése)
B) Arthrobacter simplex (egyszerű szennyezések megkötése)
C) Citrobacter freundi (urán megkötése)
D) Leuconostoc mesenteroides (PLA lebontása)
67. Milyen előnyökkel jár a kevert tenyészetek alkalmazása?
A) sterilezés nélküli technológia, vegyes szubsztráthasznosítás, alkalmazkodó képesség
B) kevesebb reaktor, gyorsabb biokonverzió, nagyobb produktivitás
C) homogén rendszer, nagy hozam, tiszta termék
D) sterilezés nélküli, optikailag aktív technológia
68. Ismertesse, hogy milyen anyagcseréjű mikrobák építik fel a szennyvíziszap-alapú biogáz-
előállító mikrobapopulációt!
(szerves savak→ecetsav→metán→stb.))
69. Soroljon fel 2 kártékony és 2 hasznos mikrobát a szőlő mikroflórájából!
A) Cephalosporangium sp., Botrytis cinerea; Aspergillus niger, Oenococcus sp.
B) Penicillium crysogenum, Anaerobacter; Schizosaccahromyces pombe
C) Penicillium expansum, Acetobacter sp.; Saccharomyces cerevisiae, Debaromyces sp.
D) Citrobacter freundii, Clostridium botulinum; Lactococcus lactis, Bacillus fragilis
70. Miért használnak a húsiparban kevert tenyészeteket?
A) tartósítás, ízesítés
B) savanyítás, pácolás, erjedések
C) sózás, füstölt íz
D) aroma, savanyítás
71. Hogyan működik az emberi immunrendszer első védelmi vonala?
A) immunválasz megtanulása, antigének azonosítása
B) patogének likvidálása, védelmi hormonok indukálása
C) fehévérsejtek folyamatos „járőrözése”
D) patogén érzékelése és eltávolítása, vagy antigén-prezentálás, adaptív válasz előidézése
72. Hogyan működik az emberi immunrendszer második védelmi vonala?
A) patogén eltávolítása
B) az első védelmi vonal felülbírálása
C) az első védelmi vonal értesítése
D) vörösvértestek indukciója
73. Ismertesse a vakcinák szempontjából fontos antigéneket (virális, bakteriális)!
(pl.: fehérje- és más molekulatípusok funkciók alapján)

7. Önellenőrző kérdések az Ipari mikrobiológia című jegyzethez 127
© Németh Áron, BME © www.tankonyvtar.hu
74. Mi az a szerotípus, és mi a jelentősége az oltóanyagok szempontjából?
A) Egy törzs felszíni Ag szerinti besorolása, immunológiailag különböznek.
B) Egy törzs felszíni membrán szerinti besorolása, immunológiailag különböznek.
C) Egy törzs felszíni Ag szerinti besorolása, immunológiailag azonosak.
D) Egy törzs felszíni membrán szerinti besorolása, immunológiailag azonosak.
75. Hogyan csoportosíthatóak az oltóanyagok a bennük lévő „hatóanyag” alapján?
A) Bakteriális, virális stb.
B) Lipid, protein, szaharid
C) Aktív, passzív hatóanyag
D) Holt anyag, attenuált, immunkomplex, élő vektoros
76. Röviden ismertessen egy inaktiválási eljárást oltóanyag készítése során!
A) besugárzás, (foly. kromatográfia), növekedési teszt
B) vegyszer hozzáadása, mintavételekkel csíraszám-ellenőrzés, vegyszer-eltávolítás (foly.
krom.), növekedési teszt
C) sterilezés, vegyszerezés, növekedési teszt
D) kisózás, renaturálás, szűrés, növekedési teszt
4. fejezet
77. Mi az elsődleges védelmi vonal a biológiai biztonságtechnika területén?
A) steril fülke
B) laboratórium
C) gumikesztyű+munkaruha
D) védőszemüveg
78. Mi a másodlagos védelmi vonal a biológiai biztonságtechnika területén?
A) steril fülke
B) laboratórium
C) gumikesztyű+munkaruha
D) védőszemüveg
79. Milyen osztályokba soroljuk a WHO ajánlásának megfelelően a mikrobákat?
A) A-D-ig felhasználásuk szerint
B) A-D-ig előfordulásuk szerint
C) 1-4-ig előállításuk szerint
D) 1-4-ig veszélyességük szerint
80. Mi az a Cartagenai jegyzőkönyv?
A) Cartagenai konferencia absztrakt könyve
B) Az aláírók csökentik a CO2-kibocsátásukat
C) Az aláírók kölcsönösen információt cserélnek a GMO-k áramlásáról.
D) Az aláírók növelik a környezetvédelmi kiadásaikat
E)
81. Ismertesse a „biológiai containment” fogalmát és lehetőségeit!
(GMOk kockázata és programozhatósága)

128 Ipari mikrobiológia
© Németh Áron, BME © www.tankonyvtar.hu
Tesztmegoldó kulcs
1.D 2.A 3.C 4.B 6.B 7.B 10.B 11.D 13.B 17.A
18.B 19.C 20.B 21.C 22.B 23.B 24.C 25.D 27.A 29.C
30.B 31.B 33.D 34.B 35.C 38.C 40.B 41.C 42.D 44.D
45.A 46.B 48.C 49.B 50.D 52.A 54.D 55.A 57.D 59.C
60.B 61.B 62.C 65.A 66.C 67.A 69.C 70.B 71.D 72.A
74.A 75.D 76.B 77.A 78.B 79.D 80.C

8. Hivatkozások 129
© Németh Áron, BME © www.tankonyvtar.hu
8. HIVATKOZÁSOK
[1] M.Koffas és G. Stephanopoulus (review, 2005)
[2] W. Leuchtenberger Appl Microbiol Biotechnol (2005) 69: 1–8
[3] S. Udaka J Bacteriol. 1960 May; 79(5): 754–755
[4] Microbial production of 1,3-propanediol: Recent developments and emerging opportunities
Biotechnology Advances 27 (2009) 895–913
[5] Biebl H, Menzel K, Zeng A-P, Deckwer W-D. Microbial production of 1,3-propanediol. Applied
Microbiology and Biotechnology 1999,52:289-297.
[6] Chao-Chen et. al. Screening of Compactin Resistant microorganism capable of converting compactin to
pravastatin Current Microbiology 2006,53:108-112
[7] Vásárhelyi Miklós: Tejsavtermelés fonalas gombával, Diplomamunka, BME, 2010.
[8] PRADO, F. C. et al. Citric acid production by solid-state fermentation on a semi-pilot scale using
different percentages of treated cassava bagasse. Braz. J. Chem. Eng. [online]. 2005, vol.22, n.4, pp. 547-
555. ISSN 0104-6632.
[9] Lüth et al (2003) USP 6,620,614 B1
[10] http://www.mta.hu/index.php?id=634&no_cache=1&backPid=390&tt_news=5151&cHash=71cfcfb769
[11] Wikipedia the online ecyclopedia: http://en.wikipedia.org/wiki/File:FormoseReaction.png
[12] D-ERYTHROSE METABOLISM IN A STRAIN OF ALCALIGENES FAECALIS J.Bact. 1956,71(6),
649-654 http://jb.asm.org/content/vol71/issue6/index.dtl
[13] http://www.genome.jp/kegg/pathway/map/map00030.png
[14] Wikipedia the online ecyclopedia:
http://upload.wikimedia.org/wikipedia/en/archive/5/5e/20061120074006!Ppp.svg
[15] Wikipedia the online ecyclopedia: http://en.wikipedia.org/wiki/File:Min_chalcopyrite.jpg
[16] Peng et al Trans. Nonferrous.Met.Soc.China (2008)18, 1443-1449
http://www.ysxbcn.com/upfile/soft/200916/23-p1443.pdf
[17] Arnold L. Demain,Aiqi Fang: The Natural Functions of Secondary Metabolites, (2000) Advances in
Biochemical Engineering/Biotechnology,Vol. 69,
http://www.springerlink.com/content/eand70p6w3ntnhwn/
[18] Nature Reviews Microbiology 5, 377-383 (May 2007)
http://www.nature.com/nrmicro/journal/v5/n5/abs/nrmicro1638.html
[19] Greg Crowther: Alcohol consumption as a way of life: the metabolism of methylotrophic bacteria
Metabolic modelers' group meeting, November 2004, Division of Allergy and Infectious Diseases within
the Department of Medicine at the University of Washington.
http://faculty.washington.edu/crowther/Research/methylo_intro_Nov2004.ppt
[20] Martyn Ford: Mode of Action of Destruxin, Centre for Molecular Design, University of Portsmouth,
http://www.port.ac.uk/research/cmd/slideshows/filetodownload,43809,en.ppt
[21] Kazuyuki Shimizu: Toward systematic metabolic engineering based on the analysis of metabolic
regulation by the integration of different levels of information, review, Biochemical Engineering Journal
46 (2009) 235–251
http://www.sciencedirect.com/science?_ob=ArticleURL&_udi=B6V5N-4WKK1T6-
1&_user=303407&_coverDate=11%2F01%2F2009&_rdoc=1&_fmt=high&_orig=search&_origin=searc
h&_sort=d&_docanchor=&view=c&_acct=C000016619&_version=1&_urlVersion=0&_userid=303407
&md5=ce39df0bd5e33986e88c26bde68cee6b&searchtype=a
[22] Wikipedia – The free encyclopedia – Steroids. http://en.wikipedia.org/wiki/Steroid#Steroidogenesis
[23] Szentirmai A. Microbial physiology of sidechain degradation of sterols, Journal of Industrial
Microbiology & Biotechnology Volume 6, Number 2, 101-115, DOI: 10.1007/BF01576429 (1990)
[24] M. V. Donova: Transformation of Steroids by Actinobacteria: A Review, Applied Biochemistry and
Microbiology, 2007, Vol. 43, No. 1, pp. 1–14. http://www.springerlink.com/content/j75176573642n780/
[25] Egorova, O., Nikolayeva, V., and Donova, M., J. Steroid Biochem. Mol. Biol., 2002, vol. 81, no. 3, pp.
273–279.

130 Ipari mikrobiológia
© Németh Áron, BME © www.tankonyvtar.hu
[26] http://davesgarden.com/guides/articles/view/2051/
[27] Bacterial (Prokaryotic) Phylogeny Webpage (March 2007).
http://www.bacterialphylogeny.com/index.html
[28] Ciccarelli, F. D., Doerks, T., von Mering, C., Creevey, C. J., Snel, B., and Bork, P. (2006). Toward
automatic reconstruction of a highly resolved tree of life. Science 311, 1283-1287.
[29] Lake, J. A., Herbold, C. W., Rivera, M. C., Servin, J. A., and Skophammer, R. G. (2007). Rooting the tree
of life using nonubiquitous genes. Mol.Biol.Evol.24, 130-136.
[30] Computational phylogenetics, Wikipedia, the free encyclopedia
http://en.wikipedia.org/wiki/Computational_phylogenetics 2010. október 26.
[31] Ksenia Egorova and Garabed Antranikian: Industrial relevance of thermophilic Archaea, Current Opinion
in Microbiology 2005, 8:649–655
[32] COSTANZO BERTOLDO AND GARABED ANTRANIKIAN: The Order Thermococcales,
Prokaryotes (2006) 3:69–81, Chapter 5. DOI: 10.1007/0-387-30743-5_5
http://www.springerlink.com/content/v20wq8765k62167x/fulltext.pdf
[33] N. Dogra and G.N. Qazi: Biotransformation of cholesterol by Micrococcus species mediated by plasmid
DNA, World Journal of Microbiology & Biotechnology 15: 411±415, 1999.
http://www.springerlink.com/content/u5546k47448h4p36/
[34] Kazuaki Yoshimune, Yasuo Shirakihara, Mamoru Wakayama, Isao Yumoto: Crystal structure of salt-
tolerant glutaminase from Micrococcus luteus K-3 in the presence and absence of its product l-
glutamate and its activator Tris, FEBS Journal, Volume 277, Issue 3, pages 738–748, February 2010
http://onlinelibrary.wiley.com/doi/10.1111/j.1742-4658.2009.07523.x/abstract
[35] R.G. Saratale, G.D. Saratale, J.S. Chang, and S.P. Govindwar, Ecofriendly degradation of sulfonated
diazo dye C.I. Reactive Green 19A using Micrococcus glutamicus NCIM-2168, Bioresource Technology
Volume 100, Issue 17, September 2009, Pages 3897-3905
http://www.sciencedirect.com/science?_ob=ArticleURL&_udi=B6V24-4W386XT-
3&_user=10&_coverDate=09%2F30%2F2009&_rdoc=1&_fmt=high&_orig=search&_origin=search&_s
ort=d&_docanchor=&view=c&_searchStrId=1521854297&_rerunOrigin=google&_acct=C000050221&_
version=1&_urlVersion=0&_userid=10&md5=f333166c8a0e4b58df8db08291149fef&searchtype=a
[36] MEENA LAKSHMAN and M.R. RAGHAVENDRA RAO: Excretion of lysine by Micrococcus
glutamicus J. Biosci., Vol. 3 Number 1, March 1981, pp. 51-55
http://www.ias.ac.in/jarch/jbiosci/3/51-55.pdf
[37] I. Sunitha, K. Jagannadha Rao and C. Ayyanna: Coimmobilized whole cells of Pseudomonas reptilivora
and Micrococcus glutamicus in calcium alginate gel for the production of L-glutamic acid. Bioprocess
and Biosystems Engineering, Volume 18, Number 1, 55-58, DOI: 10.1007/PL00008974 (1997)
http://www.springerlink.com/content/b32gujlh4pj7l2rl/
[38] GANGULY S. ; BANIK A. K. ; Induced mutation and selection of High Yielding Strain of
Micrococcus glutamicus for glutamic acid production, Journal of the Indian Chemical Society,
2010, vol. 87, no6, pp. 717-721
http://cat.inist.fr/?aModele=afficheN&cpsidt=22991226
[39] N. Gardner, C.P. Champagne: Production of Propionibacterium shermanii biomass and vitamin B12
on spent media, Journal of Applied Microbiology, Volume 99, Issue 5, pages 1236–1245, November
2005
http://onlinelibrary.wiley.com/doi/10.1111/j.1365-2672.2005.02696.x/full
[40] A. Quesada-Chanto, A. S. Afschar and F. Wagner: Optimization of a Propionibacterium acidipropionici
continuous culture utilizing sucrose, Applied Microbiology and Biotechnology Volume 42, Number 1, 16-
21, (1994) DOI: 10.1007/BF00170217
http://www.springerlink.com/content/jj4124461174p833/
[41] Edward J. Wawszkiewicz: Erythritol metabolism by Propionibacterium pentosaceum. Role of L-
erythrulose 1-phosphate, Biochemistry, 1968, 7 (2), pp 683–687,
DOI: 10.1021/bi00842a025, http://pubs.acs.org/doi/abs/10.1021/bi00842a025
[42] Maria L. W. Gent-Ruijters, Francisca A. Meijere, Wijtske Vries and A. H. Stouthamer: Lactate
metabolism in Propionibacterium pentosaceum growing with nitrate or oxygen as hydrogen acceptor,
Antonie van Leeuwenhoek Volume 42, Number 3, 217-228, (1976) DOI: 10.1007/BF00394118
http://www.springerlink.com/content/lk3j875h85v44244/

8. Hivatkozások 131
© Németh Áron, BME © www.tankonyvtar.hu
[43] E. A. DELWICHE', E. F. PHARES, AND S. F. CARSON: SUCCINIC ACID DECARBOXYLATION
SYSTEM IN PROPIONIBACTERIUM PENTOSACEUM AND VEILLONELLA GAZOGENES, J.
Bacteriol..(1956); 71: 503-625 http://jb.asm.org/cgi/reprint/71/5/598.pdf
[44] Miyagawa K, Sumida M, Nakao M, Harada M, Yamamoto H, Kusumi T, Yoshizawa K, Amachi T,
Nakayama T.: Purification, characterization, and application of an acid urease from Arthrobacter
mobilis, J Biotechnol. 1999 Feb 19;68(2-3):227-36. http://www.ncbi.nlm.nih.gov/pubmed/10194859
[45] Yukiko Kodama*, Hiroshi Yamamoto, Norihide Amano and Teruo Amachi: Reclassification of Two
Strains of Arthrobacter oxydans and Proposal of Arthrobacter nicotinovorans sp. nov., Int J Syst
Bacteriol 42 (1992), 234-239; DOI 10.1099/00207713-42-2-234
http://ijs.sgmjournals.org/cgi/content/abstract/42/2/234
[46] Ummadi, M.; Weimer, B. C., Tryptophan Catabolism in Brevibacterium linens as a Potential Cheese
Flavor Adjunct. Journal of Dairy Science 2001, 84, (8), 1773-1782.
http://www.dfrc.wisc.edu/DFRCWebPDFs/2001-Ummadi-JDS-84-1773.pdf
[47] Wikepedia a szabad enciklopédia http://hu.wikipedia.org/wiki/P%C3%A1lpusztai_sajt
[48] Reichert, K.; Lipski, A.; Pradella, S.; Stackebrandt, E.; Altendorf, K., Pseudonocardia asaccharolytica sp.
nov. and Pseudonocardia sulfidoxydans sp. nov., two new dimethyl disulfide-degrading actinomycetes
and emended description of the genus Pseudonocardia Int J Syst Bacteriol 1998, 48, (2), 441-449. doi:
10.1099/00207713-48-2-441. http://ijs.sgmjournals.org/cgi/content/abstract/48/2/441
[49] KEIKO NAKAGAWA, KAZUO SATO, YOSHIHISA TSUKAMOTO and AKIO TORIKATA:
MICROBIAL CONVERSION OF MILBEMYCINS: 28-HYDROXYLATION OF MILBEMYCINS BY
Amycolata autotrophica, The Journal of Antibiotics Vol.46 , No.3(1993)pp.518-519
http://www.journalarchive.jst.go.jp/english/jnlabstract_en.php?cdjournal=antibiotics1968&cdvol=46&noi
ssue=3&startpage=518
[50] Cover, W. H.; Kirpekar, A. C.; George, H.; Stieber, R. W., Calcium inhibition of efrotomycin production
by Nocardia lactamdurans Journal of Industrial Microbiology and Biotechnology, (1991) Vol. 7, pp 41-
44.
[51] Yamamura, H.; Tamura, T.; Sakiyama, Y.; Harayama, S., Nocardia amamiensis sp. nov., isolated from a
sugar-cane field in Japan, Int J Syst Evol Microbiol 2007, 57, (7), 1599-1602. doi: 10.1099/ijs.0.64829-0.
http://ijs.sgmjournals.org/cgi/reprint/57/7/1599
[52] Xu M, Yu HM, Tan TW, Zhu YQ, Shen ZY: Molecular cloning of an amidase gene from Nocardia sp.
and its expressionin Escherichia coli Sheng Wu Gong Cheng Xue Bao. (2006) Jul; 22(4):682-5. article in
chineese http://www.ncbi.nlm.nih.gov/pubmed/16894910
[53] COLLINS, M. D.; SMIDA, J.; DORSCH, M.; STACKEBRANDT, E., Tsukamurella gen. nov. Harboring
Corynebacterium paurometabolum and Rhodococcus aurantiacus. Int J Syst Bacteriol 1988, 38, (4), 385-
391. doi: 10.1099/00207713-38-4-385 http://ijs.sgmjournals.org/cgi/reprint/38/4/385
[54] Hernández, A.; Weekers, F. d.; Mena, J. s.; Borroto, C.; Thonart, P., Freeze-drying of the biocontrol agent
Tsukamurella paurometabola C-924 Predicted stability of formulated powders. Industrial Biotechnology
2006, 2, (3), 209-212. doi:10.1089/ind.2006.2.209.
http://www.liebertonline.com/doi/pdf/10.1089/ind.2006.2.209
[55] Wikipedia 2010. november 23. http://en.wikipedia.org/wiki/Nocardicin_A
[56] Kinoshita, N.; Homma, Y.; Igarashi, M.; Ikeno, S.; Hori, M.; Hamada, M., Nocardia vinacea sp. nov.
Actinomycetologica 2001, 15, (1), 1-5.
[57] Shigemori, H.; Komaki, H.; Yazawa, K.; Mikami, Y.; Nemoto, A.; Tanaka, Y.; Sasaki, T.; In, Y.; Ishida,
T.; Kobayashi, J. i., Brasilicardin A. A Novel Tricyclic Metabolite with Potent Immunosuppressive
Activity from Actinomycete Nocardia brasiliensis. The Journal of Organic Chemistry 1998, 63, (20),
6900-6904.
[58] Colin R. Harwood: Bacillus subtilis and its relatives: molecular biological and industrial workhorses,
Trends in Biotechnology, Volume 10, 1992, Pages 247-256
[59] Sui Lam Wong: Advances in the use of Bacillus subtilis for the expression and secretion of heterologous
proteins, Current Opinion in Biotechnology, Volume 6, Issue 5, 1995, Pages 517-522
http://www.sciencedirect.com/science?_ob=MImg&_imagekey=B6VRV-45765YH-4R-
1&_cdi=6244&_user=303407&_pii=0958166995800859&_origin=search&_zone=rslt_list_item&_cover
Date=12%2F31%2F1995&_sk=999939994&wchp=dGLbVzz-
zSkzS&md5=849ab35125b5f838beb157d724845d6f&ie=/sdarticle.pdf

132 Ipari mikrobiológia
© Németh Áron, BME © www.tankonyvtar.hu
[60] T. Payot, Z. Chemaly and M. Fick Lactic acid production by Bacillus coagulans—kinetic studies and
optimization of culture medium for batch and continuous fermentations , Enzyme and Microbial
Technology, Volume 24, Issues 3-4, 3 February 1999, Pages 191-199[61]
[61] Vaughan A. Hilder and Donald Boulter: Genetic engineering of crop plants for insect resistance – a
critical review (1999), Crop Protection, 18, 177-191
http://www.cib.espol.edu.ec/Digipath/D_Papers/42684.pdf
[62] Boer, A. S.; Priest, F.; Diderichsen, B. r., On the industrial use of Bacillus licheniformis: a review. In
Applied Microbiology and Biotechnology, ed.; 'Ed.'^'Eds.' Springer Berlin / Heidelberg: 1994; 'Vol.' 40,
p^pp 595-598. http://www.springerlink.com/content/n6t3p7112042v146/fulltext.pdf
[63] ENTerococci Industrial Platform http://imol.vub.ac.be/IMDO/projects/ENTIP.html
[64] Wikipedia, the online encyclopedia: http://en.wikipedia.org/wiki/Bacillus
[65] Garbayo, I.; VÃlchez, C.; Vega, J.; Nava-Saucedo, J.; Barbotin, J., Influence of immobilization
parameters on growth and lactic acid production by Streptococcus thermophilus and Lactobacillus
bulgaricus co-immobilized in calcium alginate gel beads. In Biotechnology Letters, ed.; 'Ed.'^'Eds.'
Springer Netherlands: 2004; 'Vol.' 26, p^pp 1825-1827.
http://www.springerlink.com/content/kj81u15234j1g452/fulltext.pdf
[66] Application of Leuconostoc Mesenteroides Strains Isolated from Chinese Food Sources to Produce
Oligosaccharides to Be Used as Prebiotics (2009) web article, (2011. január 4.) http://www.latest-
science-articles.com/Basic_Sciences/Application-of-Leuconostoc-Mesenteroides-Strains-Isolated-
from-Chinese-Food-Sour-763.html
[67] Henne A. et al.: The genome sequence of the extreme termofil Thermus thermophilus, Nat. Biotechnol.
22 (5), 547-553, 2004
[68] Nature Biotechnology (2004), 22(5), p.547 http://www.nature.com/nbt/journal/v22/n5/full/nbt956.html
[69] A. A. SALYERS,* J. R. VERCELLOTTI, S. E. H. WEST, AND T. D. WILKINS: Fermentation of
Mucin and Plant Polysaccharides by Strains of Bacteroides from the Human Colon, APPLIED AND
ENVIRONMENTAL MICROBIOLOGY, (1977), Vol. 33, No. 2, p. 319-322,
[70] Zs. Langó: Who has first observed Planctomycetes, (2005), Acta Microbiologica et Immunologica
Hungarica, 52(1), p.73-84 http://www.akademiai.com/content/qu476r5626227075/
[71] Wikipedia, the onlyne Encyclopedia: http://en.wikipedia.org/wiki/Pediococcus_acidilactici (2010.12.31)
[72] Wikipedia, the onlyne Encyclopedia: http://en.wikipedia.org/wiki/Lactococcus (2010.12.31)
[73] A.A.E Oliveira, L.M. Farias, J.R. Nicoli, J.E. Costa and M.A.R. Carvalho: Bacteriocin production by
Fusebacterium isolates recovered from the oral cavity of human subjects with and without periodontal
disease and of marmosets Res. Microbiol. (1998), 149, 585-594
http://www.sciencedirect.com/science?_ob=ArticleURL&_udi=B6VN3-3XXDWSF-
6&_user=303407&_coverDate=09%2F30%2F1998&_rdoc=1&_fmt=high&_orig=search&_origin=searc
h&_sort=d&_docanchor=&view=c&_acct=C000016619&_version=1&_urlVersion=0&_userid=303407
&md5=8c40601af3e32d2b2165cdecca539853&searchtype=a
[74] Kononen, E.; Kanervo, A.; Salminen, K.; Jousimies-Somer, H., beta -Lactamase Production and
Antimicrobial Susceptibility of Oral Heterogeneous Fusobacterium nucleatum Populations in Young
Children. Antimicrob. Agents Chemother. 1999, 43, (5), 1270-1273.
http://aac.asm.org/cgi/reprint/43/5/1270
[75] Anna Zdzieb_o, Jo´ zef Synowiecki : New source of the thermostable a-glucosidase suitable for single
step starch processing Food Chemistry 79 (2002) 485–491
[76] Wikipedia, the online Encyclopedia http://en.wikipedia.org/wiki/Chloroflexus (2010.12.31)
[77] Khanok Ratanakhanokchai,† Jun Kaneko, Yoshiyuki Kamio, and Kazuo Izaki: Purification and Properties
of a Maltotetraose- and Maltotriose-Producing Amylase from Chloroflexus aurantiacus, Appl Environ
Microbiol. 1992; 58(8): 2490–2494
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC195809/pdf/aem00049-0158.pdf
[78] M T Madigan and T D Brock: Photosynthetic sulfide oxidation by Chloroflexus aurantiacus, a
filamentous, photosynthetic, gliding bacterium. J Bacteriol. 1975 May; 122(2): 782–784.
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC246117/?tool=pmcentrez
[79] Hugler, M.; Menendez, C.; Schagger, H.; Fuchs, G., Malonyl-Coenzyme A Reductase from Chloroflexus
aurantiacus, a Key Enzyme of the 3-Hydroxypropionate Cycle for Autotrophic CO2 Fixation,
DOI: 10.1128/JB.184.9.2404-2410.2002. J. Bacteriol. 2002, 184, (9), 2404-2410.
http://jb.asm.org/cgi/content/short/184/9/2404

8. Hivatkozások 133
© Németh Áron, BME © www.tankonyvtar.hu
[80] J G Holt and R A Lewin: Herpetosiphon aurantiacus gen. et sp. n., a new filamentous gliding organism.
J Bacteriol. 1968; 95(6): 2407–2408.
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC315177/pdf/jbacter00588-0459.pdf
[81] Wikipedia, the online Encyclopedia: http://en.wikipedia.org/wiki/Cyanobacteria
[82] Prokaryotes (2006), Chapter 4.1, 7:195-210: Free living saccharolytic Spirohetes: The genus Spirohaeta
http://resources.metapress.com/pdf-preview.axd?code=g50hph505208ltu1&size=largest
[83] Ghasem D Najafpour, Lim Jit Kang: Evaluation and isolation of ethanol producer strain SMP-6, The
Proceedings of RSCE and 16th SOMChE, 28-30 Oct 2002 Malaysia
http://www.andrew.cmu.edu/user/jitkangl/Fermentation%20of%20Ethanol/Fermentation%20of%20Ethan
ol.htm
[84] Peter H. Janssen and Hugh W. Morgan: Glucose Catabolism by Spirochaeta thermophila RI 19.B1,
JOURNAL OF BACTERIOLOGY,(1992), Vol. 174, No. 8 p. 2449-2453
http://jb.asm.org/cgi/reprint/174/8/2449.pdf
[85] The Prokaryotes: Handbook of Microorganism (2006) 7:757-793 Chapter 8.1.: The order
Planctomycetales including the genera Planctomyces, Pirellula, Gemmata, and Isosphaera, and the
candidatus genera Brocadia, Kuenenia, and Scalindua
http://books.google.hu/books?id=L-
cCqW4IVj0C&pg=PA757&dq=The+prokaryotes+chapter+8.1&hl=hu&ei=7RAjTdDhJsqZOqjbyPsI&sa
=X&oi=book_result&ct=result&resnum=2&ved=0CC0Q6AEwAQ#v=onepage&q=The%20prokaryotes
%20chapter%208.1&f=false
[86] Stackebrandt, E.; Murray, R. G. E.; Truper, H. G., Proteobacteria classis nov., a Name for the
Phylogenetic Taxon That Includes the "Purple Bacteria and Their Relatives" Int J Syst Bacteriol 1988, 38,
(3), 321-325. DOI: 10.1099/00207713-38-3-321. http://ijs.sgmjournals.org/cgi/reprint/38/3/321
[87] Lee, K.-B.; Liu, C.-T.; Anzai, Y.; Kim, H.; Aono, T.; Oyaizu, H., The hierarchical system of the
'Alphaproteobacteria': description of Hyphomonadaceae fam. nov., Xanthobacteraceae fam. nov. and
Erythrobacteraceae fam. nov. Int J Syst Evol Microbiol 2005, 55, (5), 1907-1919. DOI:
10.1099/ijs.0.63663-0. http://ijs.sgmjournals.org/cgi/reprint/55/5/1907
[88] Labrat.com: http://www.thelabrat.com/restriction/sources/Brevundimonasdiminuta.shtml
[89] Silva, J. A.; Tobella, L. M.; Becerra, J.; Godoy, F.; Martínez, M. A., Biosynthesis of poly-[beta]-
hydroxyalkanoate by Brevundimonas vesicularis LMG P-23615 and Sphingopyxis macrogoltabida LMG
17324 using acid-hydrolyzed sawdust as carbon source. Journal of Bioscience and Bioengineering 2007,
103, (6), 542-546. http://www.sciencedirect.com/science?_ob=ArticleURL&_udi=B6VSD-4P5RTMP-
7&_user=10&_coverDate=06%2F30%2F2007&_rdoc=1&_fmt=high&_orig=search&_origin=search&_s
ort=d&_docanchor=&view=c&_searchStrId=1598408746&_rerunOrigin=google&_acct=C000050221&_
version=1&_urlVersion=0&_userid=10&md5=c09ab5dd7a8153a8d705eccf4490a012&searchtype=a
[90] Production of heterologous polypeptides from freshwater caulobacter, United States Patent 6861245
http://www.freepatentsonline.com/6861245.html
[91] CAULOBACTER S-LAYER KITS: For Recombinant Protein Secretion AND Peptide/Protein Display
http://www.flintbox.com/public/project/1487/
[92] Yasuhiro et al.:Production of thiotropocin by a marine bacterium, Caulobacter sp. and its antimicroalgal
activities. Journal of Marine Biotechnology, (1997); 'Vol.' 5, pp 225-229.
http://www.springerlink.com/content/0hja15c5dgc3ew1l/
[93] Microbe wiki: http://microbewiki.kenyon.edu/index.php/Hyphomonas_neptunium
[94] van Aalst-van Leeuwen, M. A.; Pot, M. A.; van Loosdrecht, M. C. M.; Heijnen, J. J., Kinetic modeling of
poly(β-hydroxybutyrate) production and consumption by Paracoccus pantotrophus under dynamic
substrate supply. Biotechnology and Bioengineering 1997, 55, (5), 773-782.
http://onlinelibrary.wiley.com/doi/10.1002/(SICI)1097-0290(19970905)55:5%3C773::AID-
BIT7%3E3.0.CO;2-8/abstract
[95] Young Tae Kim, Keunho Ji and Tae Hyug Jeong: Improved production of astaxanthin of the Paracoccus
haeundaensis by mutagenesis FASEB J. April 2010 24 (Meeting Abstract Supplement) 682.3
http://www.fasebj.org/cgi/content/meeting_abstract/24/1_MeetingAbstracts/682.3
[96] Wikipedia, the online Encyclopedia: http://en.wikipedia.org/wiki/Paracoccus_denitrificans
[97] Kranz, R.; Gabbert, K.; Locke, T.; Madigan, M., Polyhydroxyalkanoate production in Rhodobacter
capsulatus: genes, mutants, expression, and physiology. Appl. Environ. Microbiol. 1997, 63, (8), 3003-
3009. http://aem.asm.org/cgi/content/short/63/8/3003

134 Ipari mikrobiológia
© Németh Áron, BME © www.tankonyvtar.hu
[98] Uyar, B.; Schumacher, M.; Gebicki, J.; Modigell, M., Photoproduction of hydrogen by Rhodobacter
capsulatus from thermophilic fermentation effluent. In Bioprocess and Biosystems Engineering, ed.;
'Ed.'^'Eds.' Springer Berlin / Heidelberg: 2009; 'Vol.' 32, p^pp 603-606.
http://www.springerlink.com/content/r628k6535t8kw72g/
[99] Sang Seo Lib et al: Bacteriocin from Purple Nonsulfur Phototrophic Bacteria, Rhodobacter
capsulatus, Journal of Bacteriology and Virology 2009. Vol. 39, No. 4 p.269 – 276, DOI
10.4167/jbv.2009.39.4.269
http://synapse.koreamed.org/Synapse/Data/PDFData/0079JBV/jbv-39-269.pdf
[100] Taro Miyazaki,* Teruhide Sugisawa,‡ and Tatsuo Hoshino§ : Pyrroloquinoline Quinone-Dependent
Dehydrogenases from Ketogulonicigenium vulgare Catalyze the Direct Conversion of l-Sorbosone to l-
Ascorbic Acid† Appl Environ Microbiol. 2006; 72(2): 1487–1495. doi: 10.1128/AEM.72.2.1487-
1495.2006.
[101] Galata 3 : „Roseobacter bacteria – the ocean’s stars” project, Dánia
http://130.226.56.246/uk/Menu/Science/Roseobacter+bacteria+%E2%80%93+the+ocean%E2%80%99s+
stars
[102] Roseobase, Genomic resource for marine Roseobacters: http://www.roseobase.org/roseo/och114.html
[103] Sulinet, Digitális tudástár: http://sdt.sulinet.hu/Player/Default.aspx?g=954cccbf-ed5b-47dd-bb73-
28761eae299b&cid=ad91047e-4823-4393-b137-5f3cb2b945e9
[104] Wikipedia, the online encyclopedia: http://en.wikipedia.org/wiki/Hydrogenophilaceae
[105] Wikipedia, the online encyclopedia: http://en.wikipedia.org/wiki/Acidithiobacillus
[106] Joint Genom Institute – Genom portal: http://genome.jgi-psf.org/thide/thide.home.html
[107] Wikipedia, the online encyclopedia: http://de.wikipedia.org/wiki/Neisseria#Neisseria_meningitidis
[108] Wikipedia, the online encyclopedia: http://en.wikipedia.org/wiki/Burkholderia
[109] Mohana, S.; Shah, A.; Divecha, J.; Madamwar, D., Xylanase production by Burkholderia sp. DMAX
strain under solid state fermentation using distillery spent wash. Bioresource Technology 2008, 99, (16),
7553-7564.
[110] Anke Baselin: Optimization of lipase production in Burkholderia glumae, PhD-disszertáció (2005), Ruhr
Universitaet (Bochum).http://deposit.ddb.de/cgi-
bin/dokserv?idn=978412117&dok_var=d1&dok_ext=pdf&filename=978412117.pdf
[111] Dugan, P.; Stoner, D.; Pickrum, H., The Genus Zoogloea. In The Prokaryotes, ed.; Dworkin, M.; Falkow,
S.; Rosenberg, E.; Schleifer, K.-H.; Stackebrandt, E., Springer New York: 2006; 'Vol.' p^pp 960-970.
http://www.springerlink.com/content/m202714852322t01/
[112] MicrobeWiki: http://microbewiki.kenyon.edu/index.php/Beggiatoa
[113] Wikipedia, the online Ecyclopedia: http://en.wikipedia.org/wiki/Purple_sulfur_bacteria
[114] Fodor Barna: Hidrogenáz érés a Thiocapsa roseopersicina baktériumban, PhD disszertáció (2003),
Szegedi Tudományegyetem, http://doktori.bibl.u-szeged.hu/37/3/tz_hu937.pdf
[115] Wikipedia, the online encyclopedia: http://en.wikipedia.org/wiki/Alcanivorax
[116] Van Trappen, S.; Tan, T.-L.; Yang, J.; Mergaert, J.; Swings, J., Glaciecola polaris sp. nov., a novel
budding and prosthecate bacterium from the Arctic Ocean, and emended description of the genus
Glaciecola Int J Syst Evol Microbiol 2004, 54, (5), 1765-1771. DOI: 10.1099/ijs.0.63123-0.
http://ijs.sgmjournals.org/cgi/citmgr?gca=ijs;54/5/1765
[117] Gauthier, M. J.; Lafay, B.; Christen, R.; Fernandez, L.; Acquaviva, M.; Bonin, P.; Bertrand, J.-C.,
Marinobacter hydrocarbonoclasticus gen. nov., sp. nov., a New, Extremely Halotolerant, Hydrocarbon-
Degrading Marine Bacterium, Int J Syst Bacteriol 1992, 42, (4), 568-576. DOI: 10.1099/00207713-42-4-
568
http://ijs.sgmjournals.org/cgi/reprint/42/4/568
[118] Joint Genome Institute: http://genome.jgi-psf.org/micde/micde.home.html
[119] Assyrian International News Agency (2008): http://www.aina.org/news/20081201063837.htm
[120] Wikipedia, the online encyclopedia: http://en.wikipedia.org/wiki/Francisella
[121] Wikipedia, the online Encyclopedia: http://hu.wikipedia.org/wiki/Tular%C3%A9mia
[122] Wikipedia, the online encyclopedia: http://en.wikipedia.org/wiki/Tetrodotoxin
[123] Wikipedia, the online encyclopedia: http://en.wikipedia.org/wiki/Photobacterium

8. Hivatkozások 135
© Németh Áron, BME © www.tankonyvtar.hu
[124] Carnegie Melon University (Pittsburgh), „Molecular Biology of Prokaryotes” course:
http://www.bio.cmu.edu/courses/03441/TermPapers/97TermPapers/lux/communication.html
[125] Phage Therapy Center (The Republic of Georgia):
http://www.phagetherapycenter.com/pii/PatientServlet?command=static_proteus
[126] Wikipedia, the online encyclopedia: http://en.wikipedia.org/wiki/Salmonella_enterica
[127] Hazipatika.com: http://www.hazipatika.com/services/betegseglexikon/view/Salmonellozis?id=236
[128] Casullo de Araújo, H. W.; Fukushima, K.; Takaki, G. M. C., Prodigiosin Production by Serratia
marcescens UCP 1549 Using Renewable-Resources as a Low Cost Substrate. Molecules 2010, 15, (10),
6931-6940. http://www.mdpi.com/1420-3049/15/10/6931/pdf
[129] Baross Rezső: A hangyasav és a propionsav hatása, Országos mezőgazdasági szakfolyóirat - X. évfolyam
- 2006/09 ISSN 2061-552 http://www.agronaplo.hu/szakfolyoirat/2006/09/pr/2369
[130] Amjad Khalil, B., Z. Musa Abu and A. Mohammad, 2006. Production of antimicrobial agents from
thermophilic Yersinia sp.1 and Aeromonas hydrophila isolated from hot spring in Jordan valley.
Biotechnology, 5: 252-256. http://scialert.net/abstract/?doi=biotech.2006.252.256
[131] John Innes Center (Norwich, UK): http://www.jic.ac.uk/SCIENCE/molmicro/Azot.html
[132] A. Khanafari; A. Akhavan Sepahei: Alginate biopolymer production by Azotobacter chroococcum from
whey degradation, Int. J. Environ. Sci. Tech., 4 (4): 427-432, 2007
[133] Wikipedia, the online encyclopedia: http://en.wikipedia.org/wiki/Rhizobacter
[134] Nurizzo, D.; Turkenburg, J. P.; Charnock, S. J.; Roberts, S. M.; Dodson, E. J.; McKie, V. A.; Taylor, E.
J.; Gilbert, H. J.; Davies, G. J., Cellvibrio japonicus [alpha]-L-arabinanase 43A has a novel five-blade
[beta]-propeller fold. 2002, 9, (9), 665-668. http://www.nature.com/nsmb/journal/v9/n9/full/nsb835.html
[135] Pell, G.; Szabo, L. r. n.; Charnock, S. J.; Xie, H.; Gloster, T. M.; Davies, G. J.; Gilbert, H. J., Structural
and Biochemical Analysis of Cellvibrio japonicus Xylanase 10C Journal of Biological Chemistry 2004
279 (12 ), 11777-11788 DOI: 10.1074/jbc.M311947200
http://www.jbc.org/content/279/12/11777.full.pdf+html
[136] Tenyészsertés-állomány mentesítése Actinobacillus pleuropneumoniae fertőzéstől, Magyar
Állatorvosok lapja 2010(7) (2010.08.17.) http://www.magyarallatorvosoklapja.hu/node/79
[137] MicrobeWiki: http://microbewiki.kenyon.edu/index.php/Methylococcus_capsulatus
[138] McNell, M. M.; Martone, W. J.; Dowell, V. R., Bacteremia with Anaerobiospirillum succiniciproducens
Review of Infectious Diseases 1987 9 (4 ), 737-742 DOI: 10.1093/clinids/9.4.737
http://cid.oxfordjournals.org/content/9/4/737.full.pdf+html
[139] Nghiem, N.; Davison, B.; Suttle, B.; Richardson, G., Production of succinic acid by anaerobiospirillum
succiniciproducens. In Applied Biochemistry and Biotechnology, ed.; 'Ed.'^'Eds.' Humana Press Inc.:
1997; 'Vol.' 63-65, p^pp 565-576. http://www.springerlink.com/content/f86w142j30883u64/
[140] M. P. Bryant, Nola Small, Cecelia Bouma, and Hilda Chu: BACTEROIDES RUMINICOLA N. SP. AND
SUCCINIMONAS AMYLOLYTICA THE NEW GENUS AND SPECIES, J Bacteriol. 1958 July; 76(1):
15–23. http://www.ncbi.nlm.nih.gov/pmc/articles/PMC290147/pdf/jbacter00504-0033.pdf
[141] Wikipedia, the online encyclopedia: http://en.wikipedia.org/wiki/Xanthomonas_campestris
[142] The Merck Manuals: http://www.merckmanuals.com/professional/sec02/ch013/ch013b.html
[143] Sakaguchi, T.; Arakaki, A.; Matsunaga, T., Desulfovibrio magneticus sp. nov., a novel sulfate-reducing
bacterium that produces intracellular single-domain-sized magnetite particles. Int J Syst Evol Microbiol
2002, 52, (1), 215-221. http://ijs.sgmjournals.org/cgi/reprint/52/1/215
[144] MicrobeWiki: http://microbewiki.kenyon.edu/index.php/Desulfovibrio
[145] Wikipedia, the online encyclopedia: http://en.wikipedia.org/wiki/Bdellovibrio
[146] Wikipedia, the online encyclopedia: http://en.wikipedia.org/wiki/Myxobacteria
[147] Peter Meiser: Myxococcus xanthus – a myxobacterial model strain as multiproducer of secondary
metabolites, PhD disszertáció (2008), niversitaet des Saarlandes. http://scidok.sulb.uni-
saarland.de/volltexte/2008/1683/pdf/Dissertation_PeterMeiser_Internetversion.pdf
[148] Wikipedia, the online encyclopedia: http://en.wikipedia.org/wiki/Clostridiaceae
[149] Sveiczer Ákos: Mikrobiológia – II. Részletes Mikrobiológia jegyzet, BME 2005
http://bmekornyesz.hu/sqlatm/index.php?a=dl&f=silo/4.%20f%C3%A9l%C3%A9v/Mikrobiol%C3%B3g
ia/elojegyzet2.pdf

136 Ipari mikrobiológia
© Németh Áron, BME © www.tankonyvtar.hu
[150] Smith MT, van Grinsven AM. Dekkera anomala sp. nov., the teleomorph of Brettanomyces anomalus,
recovered from spoiled soft drinks. Antonie Van Leeuwenhoek 1984,50:143-148
http://www.springerlink.com/content/q62764570q553565/
[151] Susanne Prinz,1 Iliana Avila-Campillo,1 Christine Aldridge,1 Ajitha Srinivasan,1 Krassen Dimitrov,1
Andrew F. Siegel,1,2 and Timothy Galitski1,3: Control of Yeast Filamentous-Form Growth by Modules in
an Integrated Molecular Network, Genome Res. 2004, 14(3): 380–390.
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC353223/pdf/0140380.pdf
[152] Wikipedia, the online Encyclopedia: http://en.wikipedia.org/wiki/Fungus#Taxonomy
[153] Hibbett DS, et al. (2007). "A higher level phylogenetic classification of the Fungi". Mycological Research
111 (5): 509–47
http://www.clarku.edu/faculty/dhibbett/AFTOL/documents/AFTOL%20class%20mss%2023,%2024/AFT
OL%20CLASS%20MS%20resub.pdf
[154] Wikipedia, the online encyclopedia: http://en.wikipedia.org/wiki/Basidiomycota
[155] Maszlavér Petra: A PECSÉTVIASZ GOMBA, GANODERMA LUCIDUM (CURT.: FR.) KARST
HAZAI TERMESZTEHETŐSÉGÉNEK LEHETŐSÉGEI, PhD disszertáció, Budapesti Corvinus
Egyetem (2008) http://phd.lib.uni-corvinus.hu/319/
[156] M. Zhang, S.W. Cui, P.C.K. Cheung and Q. Wang: Antitumor polysaccharides from mushrooms: a
review on their isolation process, structural characteristics and antitumor activity, Trends in Food Science
& Technology 18 (2007) 4-19
[157] Ya-Jie Tang*, Li-Wen Zhu, Hong-Mei Li and Dong-Sheng Li: Submerged Culture of Mushrooms in
Bioreactors – Challenges, Current State-of-the-Art, and Future Prospects, Food Technol. Biotechnol. 45
(3) 221–229 (2007) http://www.ftb.com.hr/45/45-221.pdf
[158] World Congress on Industrial Biotechnology and Bioprocessing, 2009.07.22, Montreál, Kanada
[159] BIODIDAC -A bank of digital resources for teaching biology:
http://biodidac.bio.uottawa.ca/thumbnails/filedet.htm?File_name=ASCO005B&File_type=GIF
[160] Wikipedia, the online encyclopedia:
http://www.albertschatzphd.com/?cat=articles&subcat=streptomycin&itemnum=001
[161] Wikipedia, the online encyclopedia: http://en.wikipedia.org/wiki/Streptomyces#Biotechnology
[162] Wikipedia, the online encyclopedia: http://commons.wikimedia.org/wiki/File:Phylogenetic_tree_hu.svg
[163] Beile Gao and Radhey S Gupta: Phylogenomic analysis of proteins that are distinctive of Archaea and its
main subgroups and the origin of methanogenesis, BMC Genomics 2007, 8:86, pp1-23,
http://www.biomedcentral.com/1471-2164/8/86
[164] Discovery Health Webpage:
http://health.howstuffworks.com/wellness/men/how-to-get-rid-of-foot-odor1.htm
[165] Wikipedia, the online Encyclopedia: http://en.wikipedia.org/wiki/Deinococcus_radiodurans
[166] Michael R. Tansey and Thomas D. Brock: The Upper Temperature Limit for Eukaryotic Organisms, Proc
Natl Acad Sci U S A. 1972 September; 69(9): 2426–2428.
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC426956/
[167] Singh, S.; Madlala, A. M.; Prior, B. A., Thermomyces lanuginosus: properties of strains and their
hemicellulases. FEMS Microbiology Reviews 2003, 27, (1), 3-16
http://www.sciencedirect.com/science?_ob=ArticleURL&_udi=B6T37-481FWG3-
1&_user=303407&_coverDate=04%2F30%2F2003&_rdoc=1&_fmt=high&_orig=search&_origin=searc
h&_sort=d&_docanchor=&view=c&_searchStrId=1619039986&_rerunOrigin=google&_acct=C0000166
19&_version=1&_urlVersion=0&_userid=303407&md5=0002c9fee56e661909a8dfdc0572ce0b&searcht
ype=a
[168] Joint Genom Institute – Genom portal: http://genome.jgi-psf.org/Spoth1/Spoth1.home.html
[169] Rezessy Sz. J.: Thermomyces lanuginosus -galaktozidáz enzim előállítása és jellemzése, PhD
disszertáció (2003), Budapesti Corvinus Egyetem http://phd.lib.uni-corvinus.hu/478/1/de_1612.pdf
[170] Young, J.; Kuykendall, L.; Martinez-Romero, E.; Kerr, A.; Sawada, H., A revision of Rhizobium Frank
1889, with an emended description of the genus, and the inclusion of all species of Agrobacterium Conn
1942 and Allorhizobium undicola de Lajudie et al. 1998 as new combinations: Rhizobium radiobacter, R.
rhizogenes, R. rubi, R. undicola and R. vitis. Int J Syst Evol Microbiol 2001, 51, (1), 89-103.
http://ijs.sgmjournals.org/cgi/reprint/51/1/89?maxtoshow=&hits=10&RESULTFORMAT=&author1=Yo
ung&fulltext=A+revision+of+Rhizobium+Frank+1889&andorexactfulltext=and&searchid=1&FIRSTIN
DEX=0&sortspec=relevance&resourcetype=HWCIT

8. Hivatkozások 137
© Németh Áron, BME © www.tankonyvtar.hu
[171] Wikipedia, the online encyclopedia: http://en.wikipedia.org/wiki/Thiobacillusű
[172] Cummings, D. E.; Caccavo Jr., F.; Spring, S.; Rosenzweig, R. F., Ferribacterium limneticum, gen. nov.,
sp. nov., an Fe(III)-reducing microorganism isolated from mining-impacted freshwater lake sediments. In
Archives of Microbiology, ed.; 'Ed.'^'Eds.' Springer Berlin / Heidelberg: 1999; 'Vol.' 171, p^pp 183-188.
http://resources.metapress.com/pdf-preview.axd?code=7hb3yc6qjwmd6h29&size=largest
[173] Reinhold-Hurek, B.; Hurek, T., The Genera Azoarcus, Azovibrio, Azospira and Azonexus. In The
Prokaryotes, ed.; Dworkin, M.; Falkow, S.; Rosenberg, E.; Schleifer, K.-H.; Stackebrandt, E., 'Ed.'^'Eds.'
Springer New York: 2006; 'Vol.' p^pp 873-891. http://www.springerlink.com/content/l3g777m801tl215j/
[174] Salinas, M. B.; Fardeau, M.-L.; Cayol, J.-L.; Casalot, L.; Patel, B. K. C.; Thomas, P.; Garcia, J.-L.;
Ollivier, B., Petrobacter succinatimandens gen. nov., sp. nov., a moderately thermophilic, nitrate-reducing
bacterium isolated from an Australian oil well, Int J Syst Evol Microbiol 2004, 54, (3), 645-649.DOI:
10.1099/ijs.0.02732-0. http://ijs.sgmjournals.org/cgi/reprint/54/3/645
[175] Brune, A.; Ludwig, W.; Schink, B., Propionivibrio limicola sp. nov., a fermentative bacterium specialized
in the degradation of hydroaromatic compounds, reclassification of Propionibacter pelophilus as
Propionivibrio pelophilus comb. nov. and amended description of the genus Propionivibrio. Int J Syst
Evol Microbiol 2002, 52, (2), 441-444. http://ijs.sgmjournals.org/cgi/reprint/52/2/441
[176] Wikipedia, the online enyclopedia: http://en.wikipedia.org/wiki/Rhodocyclaceae
[177] PFENNIG, N., Rhodocyclus purpureus gen. nov. and sp. nov., a Ring-Shaped, Vitamin B12-Requiring
Member of the Family Rhodospirillaceae, Int J Syst Bacteriol 1978, 28, (2), 283-288. DOI:
10.1099/00207713-28-2-283., http://ijs.sgmjournals.org/cgi/reprint/28/2/283
[178] A rendszertani (III.) fejezetben bemutatott mikroszkópos képek mindegyike a Wikipedia
(http://en.wikipedia.org/wiki/Main_Page) szabad felhasználású képei.

138 Ipari mikrobiológia
© Németh Áron, BME © www.tankonyvtar.hu
ÁBRÁK, ANIMÁCIÓK, VIDEÓK, TÁBLÁZATOK JEGYZÉKE
Ábrák, animációk, videók
1.1.1. ábra: Az ipari mikrobiológia helye a tudományban ................................................................. 7 1.1.2. ábra: A lizin képlete ................................................................................................................. 7 1.1.3. animáció: A bioautográf módszer ............................................................................................ 8 1.1.4. ábra: Glutaminsav- és lizinmetabolizmus ................................................................................ 9 1.1.5. ábra: Az 1,3-propándiol-előállítás alakulása az utóbbi két évtizedben .................................. 10 1.1.6. ábra: A propándiol okozta különleges szerkezetű PTT polimer (PET: polietilén-tereftalát,
PTT: poli-trimetilén-tereftalát, PBT: polibutilén-tereftalát, ET: etilén, PR: propilén, BU: butilén,
TFS: tereftálsav) ............................................................................................................................... 10 1.1.7. animáció: 1,3-PD különleges szerkezete ................................................................................ 10 1.1.8. ábra: az 1,3-propándiol-előállító technológiák ....................................................................... 11 1.1.9. ábra: Az anaerob glicerin metabolizmus ................................................................................ 12 1.1.10. ábra: A glicerinmetabolizmus enzimei: GDH, GDHt, PDOR, (DHA: dihidroxiaceton,
HPA: hidroxipropionaldehid, PD: 1,3-propándiol) .......................................................................... 13 1.1.11. hyperpicture: A rekombináns PD fermentációs technológia ................................................ 13 1.2.1. ábra: Izolálás szilárd mintáról ................................................................................................ 15 1.2.1. videó: Izolálás......................................................................................................................... 15 1.2.2. ábra: Anaerob tenyésztőrendszerek ........................................................................................ 17 1.2.3. videó: Törzsfenntartás aktív formában ................................................................................... 20 1.2.4. ábra: Ferdeagaros, anaerob és szúrt tenyészetek, Petri-csészék és oltókacsok ....................... 21 1.2.5. ábra: Microplate reader – HTS szűrővizsgálathoz ................................................................. 22 1.2.6. ábra: Antibiotikum-termelők szűrővizsgálati tesztje .............................................................. 24 1.2.7. videó: Klasszikus törzsfejlesztés ............................................................................................ 26 1.2.8. ábra: A modern törzsfejlesztés vázlata ................................................................................... 27 1.2.9. ábra: E. coli ecetsav képzésének eliminációja: A) glükóz transzfer deficiens mutáns,
B) aceBAK génekkel megnövelt glioxalát- és citrátkör- aktivitás. .................................................. 30 1.3.1. ábra: A mikrobák centrális anyagcseréje ................................................................................ 31 1.3.2. ábra: A pelletképződést befolyásoló tényezők ....................................................................... 34 1.3.3. Pelletes morfológia ................................................................................................................. 34 1.3.4. ábra: Bolyhos pellet (1-2mm) ................................................................................................. 35 1.3.5. ábra: Óriás (1cm) pellet és szabálytalan alakú pellet ............................................................ 35 1.3.6. ábra: Amorf és óriás (2-3 cm) pellet ....................................................................................... 35 1.3.7. ábra: Pelletfelszín mikroszkópos képe (400-szoros nagyítás) ............................................... 35 1.3.8. ábra: Forgódobos bioreaktor .................................................................................................. 37 1.3.9. ábra: Koji tálcás reaktor ......................................................................................................... 38 2.1.1. ábra: Metanol-előállítás szén-dioxidból és metánból ............................................................. 41 2.1.2. hyperpicture: Az ipari mikrobiológiában fontos anyagcsereutak összefoglalása ................... 41 2.1.3. ábra: Cukorképződés glikoladldehiden keresztül .................................................................. 42 2.1.4. ábra: Eritrittől az eritrózig ...................................................................................................... 43 2.1.5. ábra: Pentóz-foszfát ciklus .................................................................................................... 43 2.1.6. ábra: Szedoheptuloz ............................................................................................................... 44 2.1.7. ábra: D-glicero-D-gluko-heptulóz .......................................................................................... 44 2.1.8. ábra: Kalkopirit ...................................................................................................................... 45 2.3.1. ábra: A szteránvázas alapvegyület – a koleszterin– szerkezete .............................................. 51 2.3.2. ábra: „Szteroidok családfája” ................................................................................................ 51 2.3.3. ábra: Ipari alapanyagként hasznosított fitoszterinek .............................................................. 52 2.3.4. ábra: Szitoszterinalapú hidrokortizon- (bio)szintézis ............................................................. 55 2.3.5. ábra: Hidrokortizon biokonverziója prednizolonná és származékaivá ................................... 55 3.1.1. ábra: Bergey-féle bakteriális rendszertan ............................................................................... 62 3.1.2. ábra: Filogenetikus fa a 3 domén elmélet alapján ................................................................. 63

Ábrák, animációk, videók, táblázatok jegyzéke 139
© Németh Áron, BME © www.tankonyvtar.hu
3.1.3. ábra: Bakteriális filogenetikus fa az indelmodell alapján ...................................................... 64 3.2.1. hyperpicture: Rendszertan ...................................................................................................... 65 3.2.2. ábra: Methanosarcina metanogén ősbaktérium ..................................................................... 66 3.2.4. ábra: Sulfolobusvírus-fertőzéssel ........................................................................................... 68 3.2.5. ábra: Bacillus subtilis ............................................................................................................ 69 3.2.6. ábra: Listeria monocytogenes ................................................................................................ 70 3.2.7. ábra: Lactobacillus acidophilus ............................................................................................. 70 3.2.8. ábra: Oenococcus oeni ........................................................................................................... 71 3.2.9. ábra: Lactococcus lactis ......................................................................................................... 71 3.2.10. ábra: Clostridium botulinum ................................................................................................ 72 3.2.12. ábra: Microccus mucilaginosus ........................................................................................... 74 3.2.13. ábra: Arthrobacter chlorophenolicus ................................................................................... 74 3.2.14. ábra: Corynebacterium diphteriae ........................................................................................ 75 3.2.15. ábra: Mycobacterium tuberculosis ....................................................................................... 76 3.2.16. ábra: Rhodococcus species ................................................................................................... 77 3.2.17. ábra: Propionibacterium acne ............................................................................................... 78 3.2.18. ábra: Streptomyces griseus ................................................................................................... 79 3.2.19. ábra: Petrotoga sp. ................................................................................................................ 79 3.2.20. ábra: Fusobacterium novum ................................................................................................ 80 3.2.21. ábra: Deinococcus radiodurans ............................................................................................ 80 3.2.22. ábra: Chloroflexus auranticus .............................................................................................. 81 3.2.23. ábra: Arthrospira (Spirulina) sp. .......................................................................................... 82 3.2.24. ábra: Bacteroides biacutis ..................................................................................................... 82 3.2.25. ábra: Planctomyces bekefii ................................................................................................... 83 3.2.26. ábra: Aquifex aeolicus .......................................................................................................... 83 3.2.27. ábra: Myxococcus xanthus .................................................................................................. 84 3.2.28. ábra: Heliobacter pylori ....................................................................................................... 85 3.2.29. ábra: Ketogulonicigenium vulgare ....................................................................................... 86 3.2.30. ábra: Agrobacterium tumefaciens répaszöveten .................................................................. 86 3.2.31. ábra: Burkholderia cepacia .................................................................................................. 87 3.2.32. ábra: Alcanivorax borkumensis ............................................................................................ 89 3.2.33. ábra: Escherichia coli ........................................................................................................... 89 3.2.34. ábra: Pseudomonas sp. ......................................................................................................... 90 3.2.3. ábra: Az ősbaktériumok „családfája” a rokonsági távolságokkal
(NJ/ML/MP algoritmussal is) .......................................................................................................... 92 3.2.11. ábra: Az Actinobacterek családfája (Az NCBI taxonomia adatfile alapján generálva) ....... 94 3.3.1. animáció: Gombák életciklusai .............................................................................................. 95 3.3.2. ábra: A legújabb gombarendszertan ....................................................................................... 96 3.3.3. ábra: Bazidomiceták felépítése .............................................................................................. 97 3.3.4. ábra: Aszkuszos gombák spóraképzése ............................................................................... 100 3.3.5. videó: Élesztőtenyészet ........................................................................................................ 100 3.3.6. videó: Fonalas gomba sporangium ....................................................................................... 100 3.4.1. ábra: Vörös alga ................................................................................................................... 101
Táblázatok
1.1.1. táblázat: Színkódrendszer a Biotechnológiai területek jelölésére ............................................ 5 1.2.1. táblázat: Fonalas gombák ipari felhasználása ......................................................................... 18 1.2.2. táblázat: Termofil mikroorganizmusok lelőhelyei ................................................................. 20 2.1.1. táblázat: Acidiphiliumok tápközege ....................................................................................... 45 2.3.1. táblázat: A legfontosabb mikrobiális szekunder metabolitok. ................................................ 47 2.3.2. gyakori mikrobiális szteroidkonverziók ................................................................................. 53 3.1.1. táblázat: A rendszertan fejlődése ............................................................................................ 57 3.2.1. táblázat: Az ősbaktériumok különleges képességei................................................................ 66 3.2.2. táblázat: Bacillusok osztálya .................................................................................................. 93

140 Ipari mikrobiológia
© Németh Áron, BME © www.tankonyvtar.hu
3.3.1. táblázat: Kalapos gombák fermentációs felhasználása .......................................................... 99 3.5.1. táblázat: Tejtermékek mikrobiotája ...................................................................................... 103 3.6.1. táblázat Az immunrendszer felépítése .................................................................................. 104 3.6.2. táblázat: A magyarországi kötelező védőoltások ................................................................. 104 4.1.1. táblázat: A különböző BsC szintre sorolt laboratóriumokkal szembeni követelmények ..... 106 4.1.2. táblázat: Mikrobiológiai eredetű élelmiszer-adalékok GRAS-listája ................................... 108





























