Embed Size (px)
Citation preview

Investigating Bacterial Pathogen-host Interaction by Using Scanning Electron Microscopes and Transmission Electron Microscopes
Lan Hu M.D., Ph.D. CFSAN, FDA, Laurel, MD 20708 Corresponding author: Lan Hu, M.D., Ph.D.; Email: [email protected]
Electron microscopes have been widely used to investigate cell structures under high resolution. A scanning electron microscope (SEM) provides tridimensional images of a cell surface, while a transmission electron microscope (TEM) is commonly employed to observe the internal structure changes of host cells after bacterial adherence and invasion. The SEM and TEM can reveal bacterial location, binding patterns, and host cell structure changes/damages under high resolution. The normal intestinal epithelial cells constitute a physical barrier to prevent microorganisms from entering the bloodstream. Intracellular bacterial pathogens including Campylobacter jejuni have specific tissue adherent and invasive capabilities, and can breach the barrier, and cause human gastroenteritis and other diseases. Our EM study demonstrates unique bacterial adherence, invasion, early endocytosis, and transcellular transcytosis in human intestinal cells. Although numerous molecular biological methods have been developed, electron microscopes are still valuable tools to study the bacterial pathogen-host interaction. This chapter will present an overview of the uses of the SEM and TEM to investigate the interaction of Campylobacter jejuni and human intestinal cells, and also introduce our specimen preparation procedures for the SEM and TEM observation. Our study demonstrates that the combination of scanning electron microscopy with transmission electron microscopy can provide precise and detailed information for understanding bacterial pathogen-host interaction.
Key words: scanning electron microscope, transmission electron microscope, bacteria-host interaction, Campylobacter jejuni, foodborne bacteria, invasion
Introduction
An electron microscope (EM) uses electrons instead of light to create an image of a specimen. The resolution of a microscopic image is determined by the wavelength of the illumination source that is being used to probe the specimen. Because the wavelength of electrons accelerated in an EM is much smaller than that of visible light, an electron microscope has much greater resolution than a light microscope [1,2]. Electron microscopy can reveal the detailed structure of biological and non-biological specimens such as bacteria, viruses, chlamydia, mycoplasmas, cells, tissues, clinical samples, metal materials, and others [3-6]. The modern EM creates images that can be visualized by a computer or detected by a digital camera. A transmission electron microscope (TEM) uses a high voltage electron beam to create an image, and offers structural and composition information with high resolution. It can image a specimen up to 1 micrometer (μm) thickness, and can achieve better than 0.5 angstrom (Å) resolution and about 10,000,000 times magnification. A scanning electron microscope (SEM) is the EM analog of a stereo light microscope, which uses a focused beam of electrons and is capable of producing images of the surface structure of a specimen. It can provide the details of the surface structures of cells and whole organisms that are not possible to be observed by a TEM. It can also be used for particle counting and size determination. Although numerous molecular biological methods have been developed, the electron microscopy is still in the forefront of both clinical diagnoses and microbiological studies. In clinical laboratories, the electron microscopy is particularly valuable in the surveillance of emerging diseases and potential bioterroristic viruses, chlamydia, mycoplasmas, and new or unculturable bacteria. In the microbiological studies, the electron microscopy has demonstrated how bacterial pathogens adhere to and invade host cells, and how bacteria replicate and cause damages in host cells. The electron microscopy provides key information on the cell function and cell damages [4, 5]. This chapter will present an overview of using the SEM and TEM to investigate bacteria-host interaction, focusing on our results and specimen preparation procedures for SEM and TEM in Campylobacter jejuni infected human intestinal Caco-2 cells.
The advantages and disadvantages of EM in microbiological research
The SEM and TEM are the basic types of EM, and are extensively employed in microbiological research and clinical laboratories due to their multiple advantages [1,2,4,5]. A SEM has several advantages over a light microscope. (1) It has a large depth of field, which allows to observe a large area of a specimen at the same time, and to create a tridimensional image. (2) It has a high resolution of about 2 nanometer (nm). (3) It uses electromagnets rather than lenses, so the degree of magnification can be easily controlled.
Microscopy and imaging science: practical approaches to applied research and education (A. Méndez-Vilas, Ed.)
195
___________________________________________________________________________________________

(4) Scanning electron microscopy is one of the rapid front line tests in disease outbreaks or identification of a new pathogenic agent. (5) Both liquid and tissue samples can be analyzed and no prior information is required. The disadvantages for the SEM are: (1) SEM utilizes vacuum conditions and water would vaporize in the vacuum, so all water must be removed from the samples. (2) SEM should use conductive specimens, and non-conductive materials have to be treated by covering the specimen with a thin layer of conductive materials such as gold, palladium alloy, carbon, osmium, etc. (3) The images are black and white. A TEM is a powerful electron microscope with many advantages. (1) It has much higher resolution than SEM. The image resolutions for TEM are around 1-2Å with a maximum potential magnification of approximate 0.2 nanometer. (2) High energy electrons (80-200 keV) are transmitted through electron transparent samples (~100 nm thick). (3) The TEM images are high quality and detailed. The disadvantages of TEM include: (1) The samples are limited to those that are electron transparent, ultrathin, dried, and solid samples. (2) The images are black and white. (3) Although serial ultrathin sections have been proven to be a useful technique for EM, the sample preparation for TEM may be laborious and time consuming, and requires specially trained technicians.
Investigating bacterial pathogen-cell interaction using electron microscopy
Understanding the complex pathogenic bacteria-host interaction requires investigation at the level of single cells/tissues and single infection events. For this purpose, the electron microscopy is employed to examine bacterial infection in cultured cells or animal models. The TEM and SEM are particularly useful in studying morphologic changes of cell and tissue structures before and after bacterial infection. Intestinal epithelial cells linked by tight junction forms a physical barrier that protects against invasion of nonpathogenic bacteria residing in the intestinal lumen. Salmonella, Shigella, enteropathogenic and enterohemorrhagic Escherichia coli (EPEC and ETEC) are foodborne bacterial pathogens. The pathogens have specific tissue-adhering, and/or -invading capabilities and can physically breach the intestinal mucosal barrier [7]. Bacterial diseases of the gastrointestinal tracts result from a complex set of interaction of the offending bacteria and the gastrointestinal cells. These enteric pathogens typically exploit host cell machinery, establishing a unique association with human intestinal epithelial cells that result in disease initiation, develop, and spread. The bacterial adherence rapidly initiates host signal transduction events including the secretion of effector proteins via type 3 secretion system 1 (T3SS1) into the intestinal cells that lead to cytoskeletal rearrangements of the cells and bacterial colonization (for EPEC and ETEC) or internalization (for Salmonella and Shigella). After the bacteria invade the enteric cells, the T3SS2 secrets and regulates effector proteins for bacterial survival and replication. Understanding the mechanisms by which bacteria induce infectious diseases is important for controlling and preventing the diseases. Cultured intestinal cells are commonly used as a simple model for investigating the colonization and invasion because they are more easily controlled than whole animal models. Electron microscopy is a perfect method to examine the binding and invasive abilityes of pathogenic bacteria in the host cells [8,9]. Electron microscopy combined with immune labelling technique and mutation methods plays a critical role in the investigation of the molecular basis and function of T3SS. Electron microscopy revealed supramolecular structures spanning the inner and outer membranes of flagellated and nonflagellated strains of the bacteria. The needle structure of T3SS is composed of a 7-8-nm-wide and 60-nm-long needle emanating from the bacterial surface as well as a shorter cylinder in 20-30 nm in diameter located in the bacterial membrane [10]. The T3SS is a key virulence factor of the bacteria. Campylobacter jejuni is a leading cause of foodborne gastroenteritis, and also a cause of a post-infectious paralyzing neuropathy Guillain-Barre syndrome. Campylobacter-mediated pathogenesis involved gut adherence and invasion of human intestinal cells. In contrast to most entereopathogens such as Salmonella, Shigella, EPEC, and ETEC, C. jejuni lacks traditional virulence factors like T3SS. The molecular mechanism of C. jejuni intestinal inflammation is still not understand very well. After ingestion with contaminated food or water, the bacterium travels deep down into the colon where they colonize, replicate, breach epithelial barrier, and results in acute inflammation [11,12]. The epithelial cells and immune cells within the intestinal mucosa such as dendritic cells, neutrophils, and macrophages induce interleukin and chemokine production, recruit inflammatory cells, result in host cell death, and cause gastroenteritis [12]. C. jejnui strain 81-176 has been reported that it exhibited relative high invasion capacity in human cultured epithelial cells, caused bloody colitis, and invaded the intestinal cells via a unique mechanism that was microtubule dependent [13]. The SEM and TEM are perfect techniques to study Campylobacter’s adherence, invasion, and transcytosis events using differentiated human intestinal cells.
Microscopy and imaging science: practical approaches to applied research and education (A. Méndez-Vilas, Ed.)
196
___________________________________________________________________________________________

Sample preparation procedures for scanning and transmission electron microscopy
Growth of bacteria.
C. jejuni was grown in Mueller-Hinton biphasic medium and on Mueller-Hinton agar (Difco) at 37°C under a Campylobacter microaerophilic atmosphere containing 10% O2, 5% CO2, and 85% N2.
Cell lines, media, and culture conditions.
Immortalized human colonic cancer cells (Caco-2), obtained from the American Type Culture Collection (ATCC), were maintained in liquid nitrogen and cultured in minimal essential media with 20% heat-inactivated fetal calf serum (FCS) (Gibco), 2 mM l-glutamine, and 0.1 mM nonessential amino acids, as recommended by the ATCC [14].
Adherence and invasion assays.
(1) Cell preparation. Coco-2 cells at a concentration of 105/ml were added into a Transwell clear polyester membrane filter (3 um pore size, 6.5 mm diameter, Millipore Corp.), which was inserted into each well of a 24-well plate, and incubated in a 5% CO2-95% air atmosphere to grow for 7-15 days. The cell culture medium was replaced daily.
(2) Caco-2 cells differentiated significantly and formed polarized monolayers that had intercellular tight junctions and defined apical and basolateral surfaces after 7 to 15 days in culture. The integrity of tight junction was monitored by measuring monolayer transepithelial electrical resistance (TER) with a dual-voltage ohmmeter (Millicel-ERS, Millipore) and brush border organization was verified by electron microscopy [15].
(3) Adding bacteria. Bacteria suspended in minimal essential medium at the multiplicity of infection (MOI) of 20 or 100 were added to each well. Infected monolayers were incubated for 10 to 120 min at 37°C in a 5% CO2-95% air atmosphere to allow adhesion and invasion to occur.
(4) Washing. The infected monolayers were washed three times with Earle's balanced salt solution [14].
Specimen preparation procedure for scanning electron microscopy.
(1) Rinsing. The monolayers infected with C. jejuni for varies times were washed with phosphate-buffered saline (PBS).
(2) Chemical fixation. The monolayers were immersed for 2 h at 4°C in primary fixative (pH 7.2) consisting of 4% paraformaldehyde, 1% glutaraldehyde, and 0.1 M sodium cacodylate.
(3) Rinsing. The specimens were rinsed three times with 0.1 M cacodylate buffer (pH 7.2). (4) Postfixation. The specimens were postfixed for 1.5 h in 1% osmium tetroxide in 0.1 M cacodylate buffered
solution, then rinsed three times with 0.1 M cacodylate buffer. (5) Dehydration. The specimens were dehydrated gradually using 30%, 50%, 70%, and 95% ethanol. Then they were
put into absolute ethanol for 10 min, and the step was repeated two more times. (6) Dryness. Each specimen was immersed in liquid carbon dioxide and dried using a Denton DCP-1 critical point
dryer. (7) Coating. Each specimen was mounted on a stub and coated with gold-palladium using a Denton vacuum
evaporator. (8) Observation. The specimens were observed by using a Hitachi S4700 field emission scanning electron microscope
[16].
Specimen preparation procedure for transmission electron microscopy.
(1) Rinsing. The cells grown on Transwell membrane filters infected with C. jejuni 81-176 for varies times were washed with PBS.
(2) Chemical fixation. The infected cells are fixed in 3% glutaraldehyde in 0.1 M sodium cacodylate buffer (pH 7.2) for 1 h at 4°C.
(3) Rinsing. The specimens were rinsed three times with 0.1 M cacodylate buffer. (4) Postfixation. The specimens were resuspended in fixative of 3% glutaraldehyde in 0.1 M sodium cacodylate
buffered solution for 1 h. (5) Dehydration. The specimens were dehydrated in graded ethanol series. (6) Embedding. The specimens were embedded in polymer resins to stabilize them sufficiently to allow ultrathin
section. (7) Section. Thin sections were cut on an ultramicrotome. Each epoxy resin block was trimmed with a diamond/glass
knife and the targeted monolayer was selected using a light microscope by viewing semithin sections stained with toluidine blue. The ideal block was then cut into ultrathin sections.
(8) Coating. The ultrathin sections are collected on 3 mm copper (mesh) grids and stained with uranyl acetate and lead citrate.
Microscopy and imaging science: practical approaches to applied research and education (A. Méndez-Vilas, Ed.)
197
___________________________________________________________________________________________

(9) Observation. The specimens were observed with a Philips 400 HM TEM operating at an acceleration voltage of 80 kV [16,17,18].
Investigating C. jejuni adherence, invasion, and transcytosis in differentiated intestinal cells by scanning and transmission electron microscopy
The results of intestinal biopsies of patients, experimental animal models, and human cultured cell assays have demonstrated that C. jejuni is able to bind to and invade enterocytes, suggesting that gut adherence, invasion, and mucosal transcytosis are important steps in Campylobacter pathogenesis. To obtain more detailed information about the events and cell structure changes associated with Campylobacter binding and entry, the differentiated Caco-2 cells were infected with C. jejuni 81-176 at the MOI of 20 or 100 from 10 min to 120 min. Both SEM and TEM were used to examine the early Campylobacter-human enteric cell interaction. Adherence. Scanning microscopy was used to examine the binding of Campylobacter to the surface of differentiated human intestinal cells. Human Caco-2 cells that were grown on Transwell membrane filters for 7-15 days formed confluent differentiated monolayers, which hold dense apical microvilli and tight junctions. C. jejuni was confirmed to bind to Caco-2 cells after 10 min infection by the images of the electron microscope. The SEM revealed a striking interaction between the lateral surface of C. jejuni flagella and microvillus tips of Caco-2 cells 10 min postinfection (Fig. 1). The TEM shows contacts of the bacterial outer membranes and the sides or tips of the host cell microvilli (Fig. 2B). The interaction suggests that there are specific bindings between the sides of flagella and components of the rounded tips of cell microvilli [14].
Fig. 1. Scanning and transmission SEM (A) or TEM (B) images of C. jejuni adherence to Caco-2 monolayers 10 min postinfection. (A) Bar = 1.5 μm, (B) Bar = 0.2 μm (Adapted from Hu et al., 2008. “Copyright © American Society for Microbiology [Infect Immun. 2008, 76:5294-304. doi: 10.1128/IAI.01408-07]”). Invasion. Campylobacter interacts with the intestinal mucosal surface, triggers special host signal transduction events that induce host cytoskeletal rearrangements and result in bacterial uptake. The image of ultrathin section observed by the TEM (Fig. 2A) shows one bacterium interacting end-first with a host cell apical surface with its polar flagella 10 min postinfection at the MOI of 100. Fig. 2B shows that one bacterium has presumably just been internalized by endocytosis (arrow), at least two bacteria have been internalized in endosomes (arrow heads), and several bacteria appear to bound to host cell microvilli (asterisks) 10 min postinfection. The images confirm that C. jejuni internalized into the host cells at early times [14]. Translocation. Bacterial translocation across the intestinal mucosal barrier, and cause further dissemination of the bacteria. The pathways used for bacteria to traverse the intestinal barrier include: (1) bacteria pass through enterocytes or M cells (transcellular route). (2) bacteria pass between adjacent epithelial cells (paracellular route). Polarized Caco-2 cells provides a nice model to study the pathways. C. jejuni can penetrate from the apical surface to the basolateral surface of polarized Caco-2 cells without disrupting transepithelial electrical resistance [19].
A B
Microscopy and imaging science: practical approaches to applied research and education (A. Méndez-Vilas, Ed.)
198
___________________________________________________________________________________________
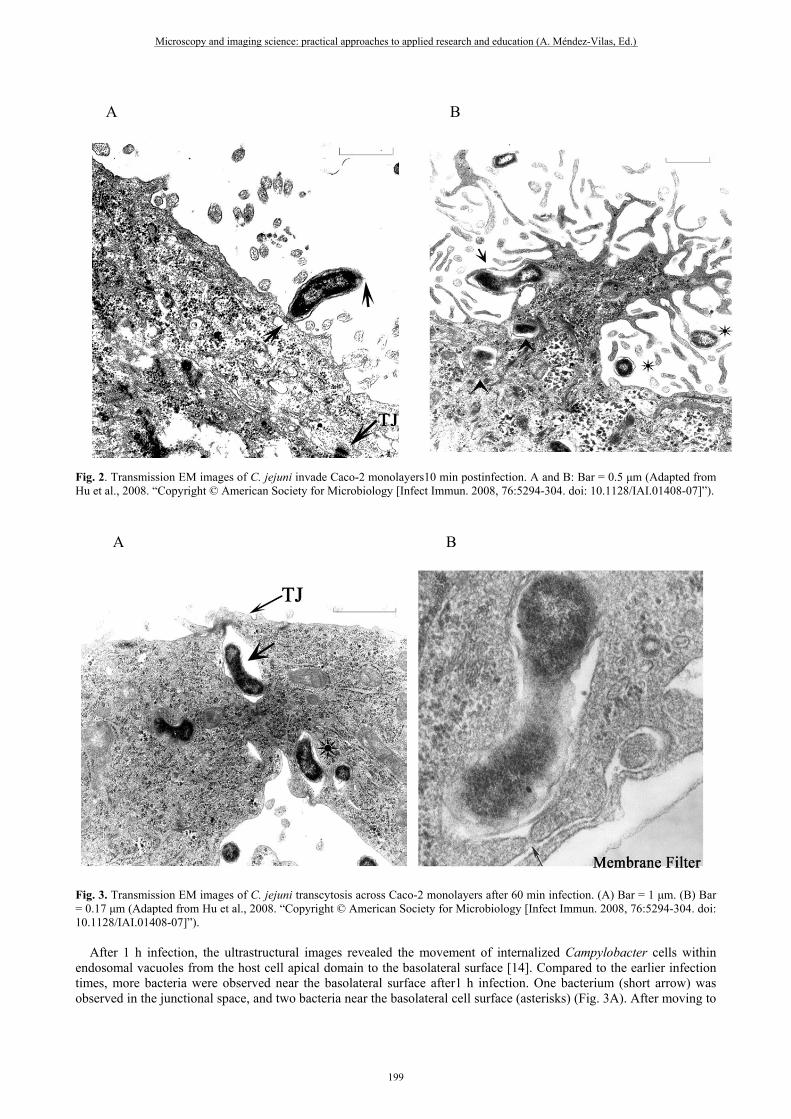
Fig. 2. Transmission EM images of C. jejuni invade Caco-2 monolayers10 min postinfection. A and B: Bar = 0.5 μm (Adapted from Hu et al., 2008. “Copyright © American Society for Microbiology [Infect Immun. 2008, 76:5294-304. doi: 10.1128/IAI.01408-07]”).
Fig. 3. Transmission EM images of C. jejuni transcytosis across Caco-2 monolayers after 60 min infection. (A) Bar = 1 μm. (B) Bar = 0.17 μm (Adapted from Hu et al., 2008. “Copyright © American Society for Microbiology [Infect Immun. 2008, 76:5294-304. doi: 10.1128/IAI.01408-07]”). After 1 h infection, the ultrastructural images revealed the movement of internalized Campylobacter cells within endosomal vacuoles from the host cell apical domain to the basolateral surface [14]. Compared to the earlier infection times, more bacteria were observed near the basolateral surface after1 h infection. One bacterium (short arrow) was observed in the junctional space, and two bacteria near the basolateral cell surface (asterisks) (Fig. 3A). After moving to
B A
B A
Microscopy and imaging science: practical approaches to applied research and education (A. Méndez-Vilas, Ed.)
199
___________________________________________________________________________________________

the basolateral surface, bacteria are released by an apparent exocytosis event at the basolateral surface. The figure shows that Campylobacter may pass between cells by paracellular route without host cell endocytosis. Fig. 3B shows one bacteria near the basolateral surface and is in the process of being released from the cell 60 min postinfection [14]. It appears that Campylobacter may cross polarized epithelial cells via transcellular route. Our EM studies plus kinetic analysis of Campylobacter-host interaction have demonstrated unique host cell adherence, early endocytosis-specific structures, and transcellular and paracellular routes of C. jejuni entering human intestinal cells [14].
New techniques and developments in electron microscopy
Electron microscopes are important diagnostic and research tools in diagnosing and researching infectious diseases and cancers at ultrastructural levels. The electron microscopy often combines with other methods such as light microscopy, immunofluorescence technique, mutation methods, and other methods. New types of EM have been developed to get higher quality images for exploring the bacterial pathogen-host interaction. Scanning transmission electron microscope (STEM). It combines the principles of TEM and SEM. A STEM requires very thin samples like TEM, and scans a very finely focused beam of electrons across the sample in a raster pattern like a SEM. Its primary advantage over conventional SEM imaging is the improvement in spatial resolution [20]. Cryo-electron microscopy (CryoEM). It has increasingly replaced the traditional methods of sample preparation for EM, and is able to obtain the images from fully frozen-hydrated specimens in a near-native state. The technique allows the TEM to view molecular structure of proteins and large molecules [2]. MicroED. The term MicroED refers various electron microscopic imaging modalities when applied to samples embedded in vitreous ice [21]. Liquid cell electron microscope. A liquid cell EM allows to view the liquid samples with powerful capabilities of an EM and to observe biological structures under high resolution without the need of drying and freezing [22]. Although numerous molecular biological methods have been developed, electron microscopy is still a valuable tool to study the bacterial pathogen-host interaction. Our study demonstrates that the combination of scanning electron microscopy with transmission electron microscopy can provide high quality and detailed images to reveal bacterial location, binding patterns, invasion and exocytosis events, as well as the host cell structure changes/damages. New technique developments in the EM would provide more detailed and precise information for exploring the bacterial pathogen-host interaction.
References [1] Woods AE, Stirling JW. Electron microscopy. In: Bancroft JD, Gamble M, editors. Theory and Practice of Histological
Techniques, 6th edition. Churchill Livingstone; 2008. P. 601-40. [2] Pilhofer M, Ladinsky MS, McDowall AW, Jensen GJ. Bacterial TEM: new insights from cryo-microscopy. Methods Cell Biol.
2010; 96:21-45. [3] Fong CK. Electron microscopy for the rapid detection and identification of viruses from clinical specimens. Yale J. Biol. Med.
1989; 62:115–30. [4] Goldsmith CS, Miller SE. Modern uses of electron microscopy for detection of viruses. Clin. Microbiol. Rev. 2009. 22:552–563. [5] Hazelton PR, Gelderblom HR. Electron microscopy for rapid diagnosis of infectious agents in emergent situations. Emerg. Infect.
Dis. 2003; 9:294–303. [6] Carabeo RA, Grieshaber SS, Fischer E, Hackstadt T. Chlamydia trachomatis induces remodelling of the actin cytoskeleton during
attachment and entry into HeLa cells. Infect Immun. 2002; 70:3793–803. [7] Lencer WI. Microbial invasion across the intestinal epithelial barrier. Pediatr Res. 2001. 49:4-5. [8] Galán JE, Collmer A. Type III secretion machines: bacterial devices for protein delivery into host cells. Science. 1999;
284(5418):1322-8. Review. [9] Malt LM, Perrett CA, Humphrey S, Jepson MA. Applications of microscopy in Salmonella research. Methods Mol Biol. 2015;
1225:165-98. [10] Kubori T, Matsushima Y, Nakamura D, Uralil J, Lara-Tejero M, Sukhan A, Galan JE, Aizawa S-I. Supramolecular structure of
the Salmonella typhimurium type III protein secretion system. Science. 1998; 280:602–5. [11] Black RE, Levine MM, Clements ML, Hughes TP, Blaser MJ. Experimental Campylobacter jejuni infection in humans. J.
Infect. Dis. 1988; 157:472–9. [12] Hu L, Kopecko DJ. Interactions of Campylobacter with eukaryotic cells: gut luminal colonization and mucosal invasion
mechanisms, In: Nachamkin I, Blaser MJ, editors. Campylobacter, 2nd ed. Washington, DC: ASM Press; 2008; P. 191-215. [13] Hu L, Kopecko DJ. Campylobacter jejuni 81-176 associates with microtubules and dynein during invasion of human intestinal
cells. Infect Immun. 1999; 67:4171-82.
Microscopy and imaging science: practical approaches to applied research and education (A. Méndez-Vilas, Ed.)
200
___________________________________________________________________________________________

[14] Hu L, Tall, BD, Curtis SK, Kopecko DJ. Enhanced microscopic definition of Campylobacter jejuni 81-176 adherence to, invasion into, translocation across, and exocytosis from polarized human intestinal Caco-2 Cells. Infect. Immun. 2008; 76, 5294-304.
[15] Beatty WL, Méresse S, Gounon P, Davoust J, Mounier J, Sansonetti PJ, Gorvel JP. Trafficking of Shigella lipopolysaccharide in polarized intestinal epithelial cells. J Cell Biol. 1999; 145:689-98.
[16] Oelschlaeger TA, Tall BD. Uptake pathways of clinical isolates of Proteus mirabilis into human epithelial cells. Microb Pathog. 1996; 20:1–16.
[17] Tall BD, Gray RT, Shah DB. Bacterial adherence and biofilm behavior of Vibrio vulnificus. In: Bailey G W, Rieder C L, editors. Proceedings of the Annual Meeting of the Microscopy Society of America. San Francisco, Calif: San Francisco Press, Inc.; 1993. P. 378–9.
[18] Huang XZ, Tall B, Schwan WR, Kopecko DJ. Physical limitations on Salmonella typhi entry into cultured human intestinal epithelial cells. Infect. Immun. 1998. 66:2928-2937.
[19] Boehm M, Hoy B, Rohde M, Tegtmeyer N, Baek KT, Oyarzabal OA, Brondsted L, Wessler S, Backert S. Rapid paracellular transmigration of Campylobacter jejuni across polarized epithelial cells without affecting TER: role of proteolytic-active HtrA cleaving E-cadherin but not fibronectin. Gut Pathog. 2012; 4:3.
[20] Nellist PD. Scanning Transmission Electron Microscopy. Editors. Hawkes PW, Spence JCH. Science of Microscopy. 2007, P. 65-132.
[21] Nannenga BL, Gonen T. MicroED opens a new era for biological structure determination. Curr Opin Struct Biol. 2016; 40:128-135.
[22] Ross EM. Opportunities and challenges in liquid cell electron microscopy. Science. 2015; 350(6267):aaa9886.
Microscopy and imaging science: practical approaches to applied research and education (A. Méndez-Vilas, Ed.)
201
___________________________________________________________________________________________





























