Embed Size (px)
Citation preview

Intragenomic Distribution and Stability of Transposable Elements inEuchromatin and Heterochromatin of Drosophila melanogaster:Elementswith Inverted Repeats Bari 1, hobo,and pogo
Carmen Di Franco,2 Alessandro Terrinoni,1 Patrizio Dimitri, 2 Nikolaj Junakovic1
1 Centro per lo Studio degli Acidi Nucleici C.N.R.2 Dipartimento di Genetica e Biologia Molecolare Universita` ‘‘La Sapienza’’ P. le A. Moro 5, 00185 Roma, Italy
Received: 21 August 1996 / Accepted: 24 March 1997
Abstract. The elements of theBari 1, hobo,andpogotransposon families that are located in euchromatin, het-erochromatin, and on the Y chromosome have been iden-tified, and their stability has been assessed by Southernblot analysis. The fraction of heterochromatic elementsappears to be distinctive of all transposon families tested,except for Bari 1. Evidence for instability of heterochro-matic elements is described. The analysis of unstableelements in differentDrosophilastocks suggests that thehost genome contributes to the stability/instability oftransposon families.
Key words: Drosophila melanogaster— Euchroma-tin — Heterochromatin — Intragenomic parasites —Transposable elements
Introduction
It has been hypothesized that transposable elements(TEs) are essentially intragenomic parasites that estab-lish little functional relationship with the host genome.Their mutagenic potential would be, by and large, neutralor detrimental to the host, while their ubiquity wouldmainly rest on their ability to replicate autonomously (fora review, see Charlesworth et al. 1994). In another in-terpretation, the mutagenic potential of TEs is believed to
be important in evolution by giving rise to adaptive ge-netic variability (for a review, see MacDonald 1995).
A testable criterion that has been put forward to dis-tinguish between these interpretations is whether ele-ments are detected that preferentially interact with spe-cific genomic sites or regions suggesting a functionalinteraction with the host genome (Orgel and Crick 1980).Another parameter that appears relevant to the relation-ship between TEs and the host is evidence for autono-mous mobility of the elements of different transposonfamilies (Di Franco et al. 1992, Nuzhdin and McKay1995). This autonomy could be related to structural dif-ferences that would make some transposon families moreactive than others. Alternatively, there could be host fac-tors that regulate the activity of specific transposon fami-lies or groups of families. Evidence for the former inter-pretation would stress the autonomy of TEs versus thehost and add to the selfish DNA hypothesis. Evidence forthe latter would show that at least one functional inter-action between the two has evolved suggesting that otherinteractions, still to be worked out, are possible.
This report is aimed at detecting stable or preferentialinteractions between transposable elements and hetero-chromatin, euchromatin, or specific genomic sites inDrosophila melanogaster.DNA samples extracted from100 adult males, females, and 100 pairs of larval salivaryglands of 12Drosophila stocks have been analyzed bythe Southern blot analysis. Individual flies have alsobeen tested. By the comparison between the stocks andthe analysis of intrastock polymorphisms, stability hasbeen assessed. Probes homologous to transposon fami-Correspondence to:N. Junakovic
J Mol Evol (1997) 45:247–252
© Springer-Verlag New York Inc. 1997

lies have been sequentially hybridized to the same filters.In the accompanying article (Terrinoni et al. 1997), therationale of this experimental approach and the resultsobtained with five non-LTR retrotransposon families aredescribed. Here, the elements with inverted repeats of thetransposon familiesBari 1, hobo,andpogoare studied.
The results show that the fraction of heterochromaticelements is distinctive of all transposon families tested,exceptBari 1. Elements of most families appear to avoidthe Y chromosome. Evidence for instability of hetero-chromatic elements is described. The analysis of inter-and intrastock polymorphisms shows that there are notransposon families inherently more unstable than othersand suggests a role of the host in regulating the stability/instability of transposon families.
Materials and Methods
Drosophila Stocks.Four reactive and four inducer strains definedunder the I-R dysgenic system and fourcn bwlines that originate fromthe same laboratory population have been analyzed. The phenotypicmarkers are described in the accompanying article (Terrinoni et al.1997).
In situ Hybridization.Preparation of polytene chromosome fromsalivary glands and fluorescent in situ hybridization (FISH) were car-ried out as described by Gatti et al. (1994).
Southern Blot Analysis.DNA extraction, digestion withHind IIIrestriction enzyme, agarose gel electrophoresis, transfer, and hybrid-ization were performed as previously described (Di Franco et al. 1989,1995).
Probes. Bari 1:Internal 1.1 KbHind III-Sma1 fragment.hobo:Internal EcoRI-EcoRIfragment 864 bp long.pogo: 1.6 Kb Hind III-EcoRI internal fragment.
Results
In Figure 1, the autoradiographs revealed byBari 1,hobo, and pogo probes in 12Drosophila stocks areshown. Three lanes per stock, bearing DNA samples ex-tracted from males, females, and larval salivary glands,respectively, are compared.
Bari 1 Elements
Location. The number of elements located inpolytenized regions of salivary glands ranges from 1 (M5stock) to 23 (Ch stock). The prominent band, 1.7 Kblong, that is strongly reduced in intensity in the ‘‘salivaryglands’’ lanes is the monomer of the heterochromaticcluster (Caizzi et al. 1992). The number of bands due toother elements located in underreplicated regions variesfrom 1 to 4 per stock.
Stability.Most stocks give rise to heterogeneous pat-terns including heterochromatic elements. The similarityof the patterns detected inseF8 and cn strains is dis-cussed below. A band is detected in all samples andwhich is also present in the ‘‘salivary glands’’ lane; thisis, most likely, the fixed element at the position 91Fpreviously detected by in situ hybridizations (Caggese etal. 1995).
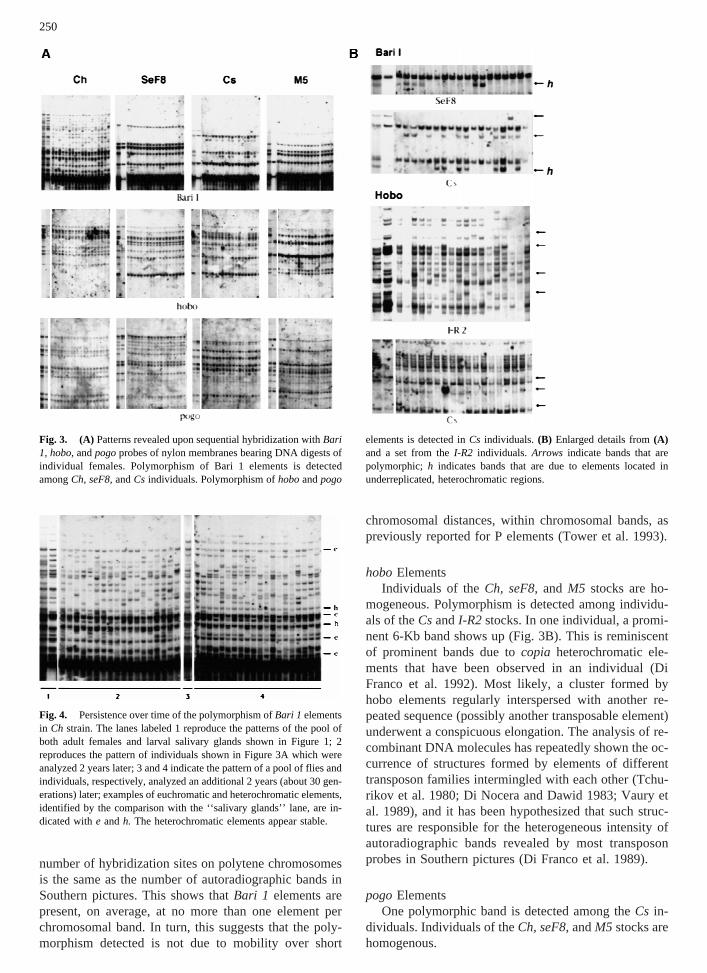
Fig. 1. From left to right: Three lanes per stock bear aliquots ofHind III digests of DNA extracted from 100 males, 100 females, and100 pairs of larval salivary glands (50 males and 50 females). TheSouthern pictures were obtained by sequentially reprobing the samefilters. Bari 1: The bar labelled 1.7 Kb indicates a band that is promi-nent in the lanes from ‘‘adults’’ and strongly reduced in the ‘‘salivaryglands’’ lanes. This is due toHind III monomers of the heterochromaticcluster (Caizzi et al. 1992). Thearrow on the left-hand side of the threesets indicates the element fixed at 91F (Caggese et al. 1995).Arrowsonthe right-hand side of the fourcn bwlines indicate bands that are sharedby two lines (bands 1 and 4), three lines (2), or all lines (3 and 5),suggesting a common origin. Noteworthy is the similarity of the Bari 1patterns between the stocksseF8andcn (see Discussion).hobo: Thenumber and intensity of most bands detected in the ‘‘adults’’ lanes arestrongly reduced in the ‘‘salivary glands’’ lanes in all stocks, exceptI-R2, which is presumably the only (H) stock. In theM5 strain, a bandis detected which is present in the ‘‘males’’ and ‘‘salivary glands’’lanes (arrow). This is due to an element located in a Y chromosomeregion which is polytenized in the salivary glands chromosomes. Thepatterns are heterogeneous among most stocks, except theseF8andcnstrains and thecn bw lines 3 and 4 which appear identical.pogo: Thearrows indicate the band that is shared by all stocks, exceptM5; this ispresumably the fixed site previously described (Tudor et al. 1992).
248
hoboElements
With respect to hobo,Drosophilastrains can be of the E(empty) or H (hobo) type. In E strains, sequences ho-mologous tohobo (termed non-hobos) give rise to het-erogeneous Southern patterns amongDrosophilastocks.They are located in underreplicated chromocentric re-gions as no hybridization signal is observed by the in situhybridization to polytene chromosomes. In H strains, inaddition to heterochromatic elements,hobosare detect-able on chromosomal arms of polytene chromosomes(Boussy and Daniels 1991; Streck et al. 1986; Blackmanand Gelbart 1989).
Location.Most bands detected in the ‘‘adults’’ lanesare absent or markedly reduced in intensity in the ‘‘sali-vary glands’’ lanes of all stocks exceptI-R2. Hence,most stock are of the E type. TheI-R2 stock is probablyan H strain because most bands detected in the ‘‘adults’’lanes do have their counterpart in the ‘‘salivary glands’’lanes.
Stability. The heterogeneity of the heterochromaticelements (non-hobos) in most stocks is confirmatory ofprevious observations (Boussy and Daniels 1991;Boussy and Periquet 1993). The patterns detected incnand seF8 strains and thecn bw lines 3 and 4 appearidentical. This is discussed below.
pogoElements
Location.A large proportion of the bands detected inthe ‘‘adults’’ lanes are present in the ‘‘salivary glands’’lanes. Hence,pogoelements are mostly located in eithereuchromatin or heterochromatic regions that arepolytenized in the salivary glands chromosomes.
Stability. A band is detected that exhibits the samemobility in 11 out of the 12 stocks tested; this is probablydue to the fixed element previously described (Tudor etal. 1992). All other bands, including those that are due toheterochromatic elements, give rise to patterns that areheterogenous among the stocks.
The Fraction of Elements in Heterochromatin
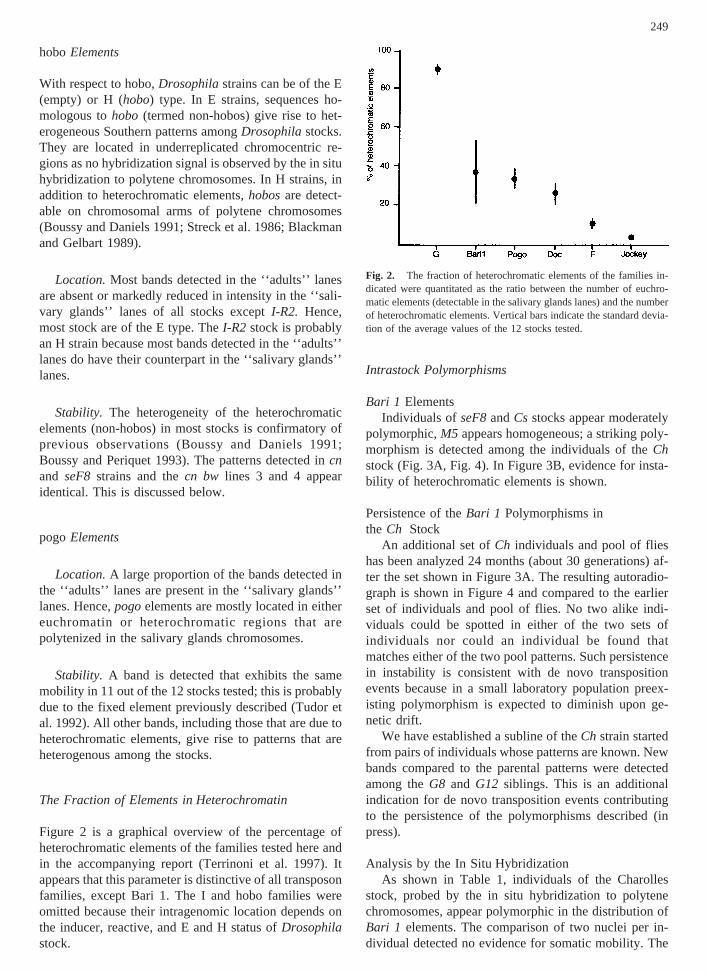
Figure 2 is a graphical overview of the percentage ofheterochromatic elements of the families tested here andin the accompanying report (Terrinoni et al. 1997). Itappears that this parameter is distinctive of all transposonfamilies, except Bari 1. The I and hobo families wereomitted because their intragenomic location depends onthe inducer, reactive, and E and H status ofDrosophilastock.
Intrastock Polymorphisms
Bari 1 ElementsIndividuals ofseF8andCs stocks appear moderately
polymorphic,M5 appears homogeneous; a striking poly-morphism is detected among the individuals of theChstock (Fig. 3A, Fig. 4). In Figure 3B, evidence for insta-bility of heterochromatic elements is shown.
Persistence of theBari 1 Polymorphisms inthe Ch Stock
An additional set ofCh individuals and pool of flieshas been analyzed 24 months (about 30 generations) af-ter the set shown in Figure 3A. The resulting autoradio-graph is shown in Figure 4 and compared to the earlierset of individuals and pool of flies. No two alike indi-viduals could be spotted in either of the two sets ofindividuals nor could an individual be found thatmatches either of the two pool patterns. Such persistencein instability is consistent with de novo transpositionevents because in a small laboratory population preex-isting polymorphism is expected to diminish upon ge-netic drift.
We have established a subline of theCh strain startedfrom pairs of individuals whose patterns are known. Newbands compared to the parental patterns were detectedamong theG8 and G12 siblings. This is an additionalindication for de novo transposition events contributingto the persistence of the polymorphisms described (inpress).
Analysis by the In Situ HybridizationAs shown in Table 1, individuals of the Charolles
stock, probed by the in situ hybridization to polytenechromosomes, appear polymorphic in the distribution ofBari 1 elements. The comparison of two nuclei per in-dividual detected no evidence for somatic mobility. The
Fig. 2. The fraction of heterochromatic elements of the families in-dicated were quantitated as the ratio between the number of euchro-matic elements (detectable in the salivary glands lanes) and the numberof heterochromatic elements. Vertical bars indicate the standard devia-tion of the average values of the 12 stocks tested.
249
number of hybridization sites on polytene chromosomesis the same as the number of autoradiographic bands inSouthern pictures. This shows thatBari 1 elements arepresent, on average, at no more than one element perchromosomal band. In turn, this suggests that the poly-morphism detected is not due to mobility over short
chromosomal distances, within chromosomal bands, aspreviously reported for P elements (Tower et al. 1993).
hoboElementsIndividuals of theCh, seF8,and M5 stocks are ho-
mogeneous. Polymorphism is detected among individu-als of theCsandI-R2 stocks. In one individual, a promi-nent 6-Kb band shows up (Fig. 3B). This is reminiscentof prominent bands due tocopia heterochromatic ele-ments that have been observed in an individual (DiFranco et al. 1992). Most likely, a cluster formed byhobo elements regularly interspersed with another re-peated sequence (possibly another transposable element)underwent a conspicuous elongation. The analysis of re-combinant DNA molecules has repeatedly shown the oc-currence of structures formed by elements of differenttransposon families intermingled with each other (Tchu-rikov et al. 1980; Di Nocera and Dawid 1983; Vaury etal. 1989), and it has been hypothesized that such struc-tures are responsible for the heterogeneous intensity ofautoradiographic bands revealed by most transposonprobes in Southern pictures (Di Franco et al. 1989).
pogoElementsOne polymorphic band is detected among theCs in-
dividuals. Individuals of theCh, seF8,andM5 stocks arehomogenous.
Fig. 3. (A) Patterns revealed upon sequential hybridization withBari1, hobo,andpogoprobes of nylon membranes bearing DNA digests ofindividual females. Polymorphism of Bari 1 elements is detectedamongCh, seF8,andCs individuals. Polymorphism ofhoboandpogo
elements is detected inCs individuals. (B) Enlarged details from(A)and a set from theI-R2 individuals. Arrows indicate bands that arepolymorphic; h indicates bands that are due to elements located inunderreplicated, heterochromatic regions.
Fig. 4. Persistence over time of the polymorphism ofBari 1 elementsin Ch strain. The lanes labeled 1 reproduce the patterns of the pool ofboth adult females and larval salivary glands shown in Figure 1; 2reproduces the pattern of individuals shown in Figure 3A which wereanalyzed 2 years later; 3 and 4 indicate the pattern of a pool of flies andindividuals, respectively, analyzed an additional 2 years (about 30 gen-erations) later; examples of euchromatic and heterochromatic elements,identified by the comparison with the ‘‘salivary glands’’ lane, are in-dicated withe andh. The heterochromatic elements appear stable.
250

Discussion
We have asked whether instances of preferential locationbetween the elements of theBari 1, hobo, and pogofamilies and genomic sites or regions are detectable inDrosophila melanogaster.The stability of elements lo-cated in different genomic regions has been assessed bythe comparison between numerousDrosophila stocksand the analysis of individual flies. The patterns wererevealed by reprobing the same filters that have beentested with non-LTR retrotransposons described in theaccompanying report (Terrinoni et al. 1997).
The Fraction of Heterochromatic Elements IsDistinctive of Most Transposon Families
Under the hypothesis that accumulation of TEs in het-erochromatin is due to the deleterious effects of mostinsertions (for a review see Charlesworth et al. 1994), thefinding that the fraction of heterochromatic elements isdistinctive of most transposon families suggests thattransposon families are not equally deleterious. Thiscould be the case if there were transposon families in-trinsically more unstable than others which would bemore deleterious because of the higher rate of de novoinsertions. As discussed below, there is evidence sug-gesting that there are no transposon families more un-stable than others.
Another possibility is that the fraction of heterochro-matic elements per family could be correlated with theinvasion time ofD. melanogaster:the older the invasion,the higher the fraction of heterochromatic elements. Theobservation that bothpogo (this report) andjockeyele-ments (accompanying report, Terrinoni et al. 1997) ap-pear to be preferentially located in euchromatin and re-stricted toD. melanogasterand sibling species (Tudor etal. 1992; Mizroki and Mazo 1990) is consistent with thisinterpretation; however, the variability of the heterochro-matic fraction of Bari 1 elements suggests that otherforces contribute to the euchromatic/heterochromaticdistribution of transposon families.
There Are No Transposon Families That Are MoreUnstable Than Others
a) Evidence From Inter-Stock Polymorphisms:In Fig-ure 1, theBari 1 andhobopatterns inseF8andcnstrains
appear identical or similar. This is rather surprising be-cause so far allDrosophila strains and even lines oflaboratory stocks maintained separate for periods of timerevealed totally heterogeneous patterns (Junakovic et al.1984; see also the heterogeneity inpogopatterns amongcn bw lines). With respect tohobo,which are located inunderreplicated heterochromatic regions, this identitycould be due to inactive elements. Upon contamination,different strains believed to be unrelated can turn homo-geneous. None of the two interpretations appears satis-factory because 1) thehobo patterns are heterogenousamong the other stocks showing that they are potentiallyactive and 2) the heterogeneity of thepogopatterns pro-vides an internal control for diversity betweenseF8andcn strains. In addition,jockey, F and Doc elements,probed on the same filter, give rise to heterogeneouspatterns betweenseF8andcn (Terrinoni et al. 1997).
Another possible interpretation of the homogeneitybetweenseF8andcn strains is thatBari 1 andhoboarefamilies more stable thanpogo. In other words, therewould be a hierarchy of stability/instability betweenthese families. If so, then elements of theBari 1 andhobofamilies are expected to be invariably more stable thanpogo in other Drosophila strains. TheBari 1 and pogoare heterogeneous incn bw lines 3 and 4, whereashobopatterns appear identical. This is consistent with the hy-pothesis ofhobobeing the most stable of the three fami-lies. However, in thecn bwlines 1 and 2,hoboandpogopatterns are totally heterogeneous, whereasBari 1 aresimilar. This is not consistent with the hypothesis of ahierarchy of stability between these families.
Evidence from Intrastock Polymorphisms:Individualsof the Ch strain that are highly polymorphic in the dis-tribution of Bari 1 elements are homogeneous in thedistribution ofhobo, pogo,and 11 additional transposons(Terrinoni et al. 1997 and unpublished). Under the hy-pothesis of a hierarchy of stability between transposonfamilies, this suggests thatBari 1 family is the mostunstable of the families tested. However,Bari 1 elementsappear homogeneous among the individuals of theM5stock which are polymorphic in the distribution ofI,jockey and F elements. This is illustrated in Table 2which shows that elements of all transposon familiestested in this and the accompanying report are polymor-phic in at least one stock. In all stocks tested, there is atleast one transposon family that gives rise to homoge-neous patterns. There is no evidence that one transposonfamily is more unstable than others suggesting that in-stability is correlated to the genomic background. In turn,this suggests as well that there are no transposon familiesmore deleterious than others. On the other hand, there arefamilies that are more abundant in heterochromatin thanothers (see above). Hence, there appears to be no corre-lation between accumulation of TEs in heterochromatinand deleterious effects in euchromatin.
Table 1. Intrastock polymorphism ofBari 1 elements in Charollesstock as determined by FISH technique to larval polytene chromo-somes
Individuals
X-Chromosome sites
2F 5F 13D 16F 19D
1 — — + + +
2 — + + — +3 — + + + +4 — + + + —5 + — — + +
251

Acknowledgments. This work was supported by the Progetto Final-izzato Ingegneria Genetica C.N.R. and the Foundation Cenci-Bolognetti, Institut Pasteur. We thank M. Tudor for critical reading ofthe manuscript, F. Spirito, and G. Modiano for stimulating discussionsand I. Zhimulev for help in interpreting the in situ pictures.
References
Blackman RK, Gelbart WM (1989) Mobile DNA. In: Berg DE, HoweMM (eds) American Society for Microbiology, Washington DC, pp523–527
Boussy IA, Daniels SB (1991) hobo transposable elements inDro-sophila melanogasterandD. simulans.Genet Res Camb 58:27–34
Boussy IA, Periquet G (1993) The transposable element hobo inDro-sophila melanogasterand related species. In: McDonald J (ed)Transposable elements and evolution. Kluwer Academic Publisher,Dordrecht, pp 192–200
Caizzi R, Caggese C, Pimpinelli S (1992) Bari 1, a new transposon-likefamily in Drosophila melanogasterwith unique heterochromaticorganization. Genetics 133:335–345
Caggese C, Pimpinelli S, Barsanti P, Caizzi R (1995) The distributionof the transposable element Bari 1 in theDrosophila melanogasterandDrosophila simulansgenomes. Genetica 96:269–283
Charlesworth B, Sniegowski P, Stephan W (1994) The evolutionarydynamics of repetitive DNA in eukaryotes. Nature 371:215–220
Di Franco C, Pisano C, Dimitri P, Gigliotti S, Junakovic N (1989)Genomic distribution of copia-like transposable elements in so-matic tissues and during development ofDrosophila melanogaster.Chromosoma 98:402–410
Di Franco C, Galuppi D, Junakovic N (1992) Genomic distribution oftransposable elements among individuals of an inbredDrosophilaline. Genetica 86:1–11
Di Franco C, Terrinoni A, Galuppi D, Junakovic (1995) DNA extrac-tion from singleDrosophila flies.DIS 76:172–174
Di Nocera PP, Dawid IB (1983) Interdigitated arrangement of twooligo(A)-terminated DNA sequences inDrosophila.Nucleic AcidsRes 11:5475–5482
Gatti M, Bonaccorsi S, and Pimpinelli S (1994) Looking atDrosophilamitotic chromosomes. Methods Cell Biol 44:371–391
Junakovic N, Caneva R, Ballario P (1984) Genomic distribution ofcopia-like elements in laboratory stocks ofDrosophila melanogas-ter. Chromosoma 90:378–382
McDonald JF (1995) Transposable elements: possible catalysts of or-ganismic evolution. Trends Ecol Evol 10(3)(105):123–126
Mizrokhi LJ, Mazo AM (1990) Evidence for horizontal transmission ofthe mobile element jockey between distantDrosophila species.Proc Natl Acad Sci USA 87:9216–9220
Nuzhdin SV, Mackay TFC (1995) The genomic rate of transposableelements movement inDrosophila melanogaster.Mol Biol Evol12:180–181
Orgel LE, Crick FHC (1980) Selfish DNA: the ultimate parasite. Na-ture 284:601–608
Streck RD, MacGaffey JE, Beckendorf SK (1986) The structure ofhobo transposable elements and their insertion sites. EMBO J 5:3615–3623
Tchurikov NA, Zelentsova ES, Georgiev GP (1980) Clusters contain-ing different mobile dispersed genes in the genome ofDrosophilamelanogaster.Nucleic Acids Res 8:429–439
Terrinoni A, Di Franco C, Dimitri P, Junakovic N (1997) Intragenomicdistribution and stability of transposable elements in euchromatinand heterochromatin ofDrosophila melanogaster.I: non-LTR ret-rotransposon. J Mol Evol (in press)
Tower J, Karpen GH, Craig N, Spradling AC (1993) Preferential trans-position of Drosophila P elements to nearby chromosomal sites.Genetics 133:347–359
Tudor M, Lobocka M, Goodell M, Pettitt J, O’Hare K (1992) The pogotransposable element family ofDrosophila melanogaster.Mol GenGenet 232:126–134
Vaury C, Bucheton A, Pe´lisson A (1989) Theb-heterochromatic se-quences flanking the I elements are themselves defective transpos-able elements. Chromosoma 98:215–224
Table 2. Overview of intrastock polymorphisms detected here and inthe accompanying report (Terrinoni et al. 1997)
Ch seF8 Cs M5 I-R2
I 58 h h p p p
I 38 h h p p pG h h h h pjockey h h p p —F h h p p —Doc h p p h —Bari 1 p p p h —pogo h h p h —hobo h h p h p
Upon reprobing the same filters, homogenous (h) and polymorphic (p)patterns are revealed. The number of bands that give rise to the poly-morphisms range from 1 (jockey inM5 stock) to over 20 (Bari 1 in Chstock). Note that (bold letters)Bari 1 elements are the only ones to bepolymorphic in theCh stock, whereas the same elements are stable inthe M5 stock which is polymorphic in the distribution of the elementsof other families
252





























