Embed Size (px)
Citation preview

Interspecific and Extraspecific Pregnancies inEquids: Anything GoesW. R. Allen and R. V. Short
From the University of Cambridge, Department of Clin-ical Veterinary Medicine, Cambridge, England (Allen)and the University of Melbourne, Royal Women's Hos-pital, Carlton, Victoria, Australia (Short). W. R. Allen Isnow at the TBA Equine Fertility Unit, Mertoun Pad-docks, Woodditton Road, Newmarket, Suffolk CB8 9BH,England. This paper was delivered at a symposium en-titled "Interspecles Hybrids In Mammals" In associa-tion with the New Zealand Genetlcal Society and Aus-tralasian Gene Mapping Workshop In Dunedln, NewZealand, from November 30-December 1, 1995.
Journal of Heredity 1997^8:384-392; 0022-1503/97/*5.00
Equids possess the unusual ability to Interbreed freely among the phenotypicaliyand karyotypically diverse member species of the genus to produce viable, butusually Infertile, offspring. The mule (female horse x male donkey) was humanity'sfirst successful attempt at genetic engineering and Its clear expression of bothparental phenotypes has contributed much to our understanding of genetic Inher-itance over the centuries. Even more surprising, mares and donkeys have beenshown to be capable of carrying to term a range of true, xenogenelc extraspeciespregnancies created by embryo transfer, including Przewalski's horse (Equus prez-walskll; In = 66)-ln-horse, (£. caballus; In = 64), and Grant's zebra (E. burchelll;2n = 44)-in-horse pregnancies. Fetal genotypes has a marked influence on placen-tal development in equids, expeclally on the width and general development of theannulate chorionic girdle, progenitor tissue of the gonadotrophin (eCG)-secretingendometrlal cups. However, transfer of intact and bisected deml-mule embryos (E.mulus; In = 63) to Jenny donkeys (£. aslnus; 2n = 62) showed convincingly thatmaternal uterine environment, probably mediated by Intrauterlne growth factor pro-duction, can exert an overriding Influence on chorionic girdle development and itsInvasion of the matemai endometrlum. Transfer of donkey embryos (2n = 62) tohorse mares (2n = 64) results In the development of an exceptionally small cho-rionic girdle that completely falls to invade the endometrlum to form endometrlalcups. Around 70% of these donkey-ln-horse preganancies are aborted betweendays 80 and 85 of gestation in conjuctlon with delayed and abnormal placentalattachment combined with a vigorous maternal cell-mediated reaction against thexenogeneic donkey trophoblast. This model of pregnancy loss shows strong evi-dence of Immune memory and the rate of fetal death Is reduced by immunizationof the surrogate mare against donkey lymphocytes. The findings suggest an im-portant role for the invasive trophoblast cells of the equine placenta in initiatingand driving attachment and interdigitatlon of the non-Invasive placenta for fetalsustenance, and in modulating materno-fetal Immunologlcal Interaction to enablesurvival of the antlgenetlcally foreign fetus in the uterus.
The genus Equus is unusual for the abilityof its phenotypicaliy and karyotypicallydiverse member species to interbreedfreely to produce viable, although usuallyinfertile, offspring. From the "AncientHorse of Mongolia" {Equus przewalskii; 2n= 66), through the many and disparatebreeds of domestic horse (E. caballus; 2n= 64), to the European ass or domesticdonkey (E. asinus; 2n = 62), through themany Asiatic and African species of wildass (e.g., E. hemionus; 2n = 54), to thevery beautiful and finely striped Grevyi'szebra of Somalia and Northern Kenya (£.greuyii; In = 46), the range of subspeciesof common zebra in Central and EasternAfrica (E. burchelli; In = 44) and finally tothe now rare Mountain Zebra of Southwest
Africa (£". zebra; 2n = 32), the consider-able range of hybrid foals born during thepast 150 years makes it reasonable to sup-pose that any equine species will conceiveto any other equine species if the femaleis inseminated with fertile semen at thecorrect time in her ovulatory cycle (Ewart1898; Gray 1971; Short 1975b; Tegetmeierand Sutherland 1895).
Equally unusual, and of considerable in-terest scientifically, has been the more re-cent discovery that at least the horse,donkey, and mule (E. mulus mulus; 2n =63) within the equine family will accept,gestate, carry to term, give birth to andrear successfully truly xenogeneic extras-pecific foals created by the use of thetechnique of between-species embryo
384

transfer (Allen 1982b). In this article wereview some of the developmental, endo-crinological, and immunological difficul-ties associated with the establishment ofpregnancy and the survival of the xeno-geneic fetus to term in mares and otherequids when carrying interspecific hybridor transferred extraspecific foals.
Interspecies Pregnancies
The Mule and HlnnyThe mule (female horse X male donkey;2n = 63) and the reciprocal cross, the nin-ny (female donkey x male horse; 2n = 63)are by far the most common equine hy-brids, simply because the horse and don-key are the only two species of equids tohave been domesticated in significantnumbers. Many millions of mules havebeen produced over the past 5,000 yearsfor their mixture of the physical and men-tal attributes of the two parental species.But in addition to its practical usefulness,the mule has intrigued and puzzled biolo-gists since the time of Aristotle. As de-scribed by Short (1975a), the clear and ap-parently equal mixture in the mule of thephenotypic characteristics of its two par-ents ran counter to Aristotle's "seed andsoil" view of reproduction, in which themale donated the essential seed for thenew offspring while the female merely pro-vided the soil in which the male seedcould grow. Centuries later, Spenser(1638-1686) and De Graaf (1641-1673)both used the mule as an example in theirindependent and revolutionary proposalsthat the females of all mammalian speciesalso contribute "seed" to the generationof new individuals (see Jocelyn and Setch-ell 1972). In more recent times, the infer-tility of mules and hinnies has similarlyaroused the interest of geneticists. Wod-sedalek (1916) was the first to reason thatspermatozoa are not produced in the tes-tes of male mules because of an incom-patibility between the paternal and mater-nal sets of chromosomes leading to ablock in meiosis. Fifty years later, Taylorand Short (1973) demonstrated that thesame chromosomal incompatibility leadsto partial meiotic arrest in female mulesand hinnies, which results in a severelydepleted stock of oocytes at birth.
It is Interesting that extensive data onthe sex ratio of mules at birth showthere is marked excess of females (56 fe-male:44 male; Craft 1938), a dramaticconfirmation of Haldane's law, whichstates that "in the case of interspecifichybrids the heterogametic sex is the one
Figure 1. (a) F, Interspecles foal produced by mating Its Welsh Pony domestic horse mother (E caballur, 2n =64) to a Przewalsld's horse stallion (E. przeioalskii; 2n = 66). (b) Two half-slb Interspecles F, foals produced bymating their F, Interspecles Przewalskl's horse X domestic horse mothers to a domestic horse pony stallion. Thefilly foal on the left In the photograph had a karyotype of 2n = 65 and was very Przewalskl's horsellke In appear-ance, whereas the colt foal on the right had a karyotype of 2n = 64 and, apart from a prominent dorsal stripe,was Indistinguishable from a normal pony foal.
most likely to be absent, rare or sterile"(Haldane 1922). The most plausible ex-planation for this phenomenon wouldappear to lie in the high mutation rate ofgenes on the Y chromosome, and es-pecially of the testis-determining gene,SRY (Tucker and Lundrigan 1993; Whit-field et al. 1993). Perhaps the horse anddonkey SRY have diverged sufficientlyfrom one another during the course oftheir independent evolution to the pointwhere donkey SRY (and other Y-linkedgenes concerned with spermatogenesis)may now be unable to induce normaltesticular development in some male hy-
brids. It would follow that the deficiencyof males could be explained by the pres-ence of a corresponding number of XYfemales in the hybrid offspring.
We still lack conclusive proof as towhether there is any consistent pheno-typic difference between mules and hin-nies, although in the light of our recentunderstanding about the exclusively ma-ternal inheritance of mitochondrial DNAand the role of genetic imprinting (Bar-ton et al. 1985), such differences be-tween the reciprocal hybrids might beexpected.
The interspecies mating of a horse mare
Allen and Short • Equine Hybrids 3 8 5

to a Jack donkey to produce a mule is asfertile, or even more so in terms of con-ception rates, as the straightforward intra-species mating between either of the pa-rental species. However, in the experienceof the authors, and as is common knowl-edge in countries like Spain, Portugal,Greece and India, where equine hybridsare still bred and used in large numbers,the reciprocal cross between the femaledonkey and the male horse is very muchless fertile. For example, during a 7 yearperiod in the authors' laboratory in Cam-bridge, England, and at the Equine Immu-nogenetics Center in Ithaca, New York, atotal of 159 attempts were made in suc-cessive breeding seasons to establish nin-ny pregnancies in 51 normal fertile femaledonkeys, using 6 pony stallions of knownhigh fertility with a mixture of natural mat-ing and artificial insemination. Only 18conceptions (14%) occurred and 11 ofthese were in the same 3 female donkeysmated or inseminated in successive years(Allen et al. 1993b).
This striking disparity in fertilizationrates between reciprocal hybrid matingsin equids has parallels in other species.For example, high fertilization rates areachieved when inseminating female goatswith ram semen, but many fewer concep-tions occur when reciprocally inseminat-ing ewes with goat semen (Hancock andMcGovern 1964; Hancock et al. 1968). Sim-ilarly, hybrid matings between the hare(Lepus americanus) and the rabbit (Oryc-tolagus cuniculus) produce high rates offertilization when rabbits are inseminatedwith hare semen, whereas less than 10%of hare oocytes are fertilized by rabbit se-men (Chang et al. 1964). The mechanismsresponsible for these large differences re-main unknown, although it is tempting tosuppose that the phenomenon of geneticimprinting (Barton et al. 1985) may play arole.
Other Equine HybridsThe wide range of equine hybrids pro-duced over the years has been well cata-loged by Gray (1971), King (1965), andShort (1975b). These have included suchunlikely combinations as the progeny pro-duced by the mating of a domestic donkeyto Somali, African, and Asiatic wild asses,and to Burchell's, Grant's, and Mountainzebra. Similarly, domestic horses havebeen mated to all the main zebra species,including Burchell's, Chapman's, Grant's,and Hartman's. In all the cases of success-ful birth in the latter group, the female ofthe partnership was a horse, in a similar
manner to the production of mules. In anyinstances where reciprocal crosses wereattempted by mating a male horse to fe-male wild asses or zebra, conceptionsfailed to occur (Gray 1971).
From these findings, and from theabove-mentioned relative infertility of thefemale donkey x male horse mating toproduce hinnies, it is tempting to specu-late that fertilization may occur readily inany cross where the sire has a lower dip-loid chromosome complement than thedam. Conversely, conception may be moredifficult in those crosses in which the sirehas a higher diploid number than the dam.
Fertility of Equine HybridsAlthough male mules and hinnies cansometimes produce a few spermatozoa,these appear to be morphologically ab-normal, with small heads (Chandley et al.1974). There are no verified accounts ofmale mules or hinnies of proven fertility,in accord with Haldane's law. On the otherhand, a number of unverified reports of in-dividual fertile female mules were made inthe early part of this century, the most no-table of which was an animal owned byTexas A&M College in the 1920s that gavebirth to two foals. One of these, sired bya donkey, was a female, and was typicallymulelike in appearance and was infertile.The other, a colt sired by a horse, wascompletely horselike in appearance andwas fully fertile (Anderson 1939).
During the past decade there has beensomething of an upsurge of fertile femaleequine hybrids reported in the scientificliterature and the majority of these havebeen verified karyotypically. For example,Ryder et al. (1985) confirmed by karyotyp-ing the offspring and parents a foal bornto a female mule mated to a donkey. Ronget al. (1988) confirmed karyotypically botha fertile mule and a fertile hinny in China,and Henry et al. (1995) described a femalemule in Brazil that has given birth to sev-eral foals, some sired by a donkey and oth-ers by a horse. Chandley (1988) discussedthe mechanisms that might function to en-able these odd female mules and hinniesto be fertile. She argued in favor of the af-finity hypothesis proposed originally byMichie (1953), whereby chromosomallybalanced haploid gametes might occasion-ally be produced by the movement of cen-tromeres of similar ancestry to oppositepoles at anaphase I of meiosis. This couldleave a whole set of female parental chro-mosomes from one species remaining inthe oocyte, with subsequent elimination ofthe paternal set to the polar body. Elimi-
nation of the donkey paternal set of chro-mosomes from the mule oocyte wouldleave only the maternal horse set remain-ing so that another mule would be pro-duced if the animal was mated to a don-key. Conversely, a pure horse would resultif the mating were to a horse stallion. Thisis exactly what apparently happened inthe case of the Texas (Anderson 1939), Ne-braska (Ryder et al. 1985), and Brazilmules (Henry et al. 1995). However, whenChandley and her colleagues karyotypedthe fertile mule and hinny in China, theyfound a mixture of horse and donkey chro-mosomes that suggested there had beenrandom inheritance of both maternal andpaternal chromosomes in the oocyte thatwas fertilized (Chandley 1988).
Not all possible equine hybrids may beas infertile as the mule and hinny. Both themale and female offspring (2n = 65) of thecross between Przewalski's horse (E. prze-walskii; 2n = 66) and domestic horse (E.caballus; 2n = 64) are fully fertile (Chan-dley et al. 1975; Koulischer and Frechkop1966; Short et al. 1974). Also of consider-able interest was the finding that the threeprogeny produced by mating an F, malePrzewalski's horse X domestic horse hy-brid (Figure la) to three pony mares werea filly with a karyotype of 2n = 65 that wasvery similar to Przewalski's horse in ap-pearance and two colts, both of which hada karyotype of 2n = 64 and, apart from aprominent dorsal stripe, were indistin-guishable in general appearance from anormal horse foal (Figure lb).
Gray (1971) listed a number of examplesof hybrids between the domestic donkeyand both Burchell's and Chapman's zebrathat were reported to be fertile, althoughnone of these cases were ever verified. Onthe other hand, Gray (1971) also listed ex-amples of hybrids between the domesticdonkey and the Persian wild ass (£ hem-ionus onager) and Somali wild ass (E. asi-nus africanus) and other hybrids betweenthe various species of zebra and the do-mestic horse as being infertile.
Extraspecies Equine PregnanciesIn addition to their readiness to interbreedto produce hybrids, the use of embryotransfer techniques has highlighted a re-markable ability of the member species ofthe genus to accept and carry to term (ornear term) a variety of truly xenogeneicextraspecies pregnancies. Over a 15 yearperiod, in collaboration with the Zoologi-cal Society of London and the Equine Im-munogenetics Center of Cornell University(Ithaca, New York), live offspring have
386 The Journal of Heredity 1997:88(5)
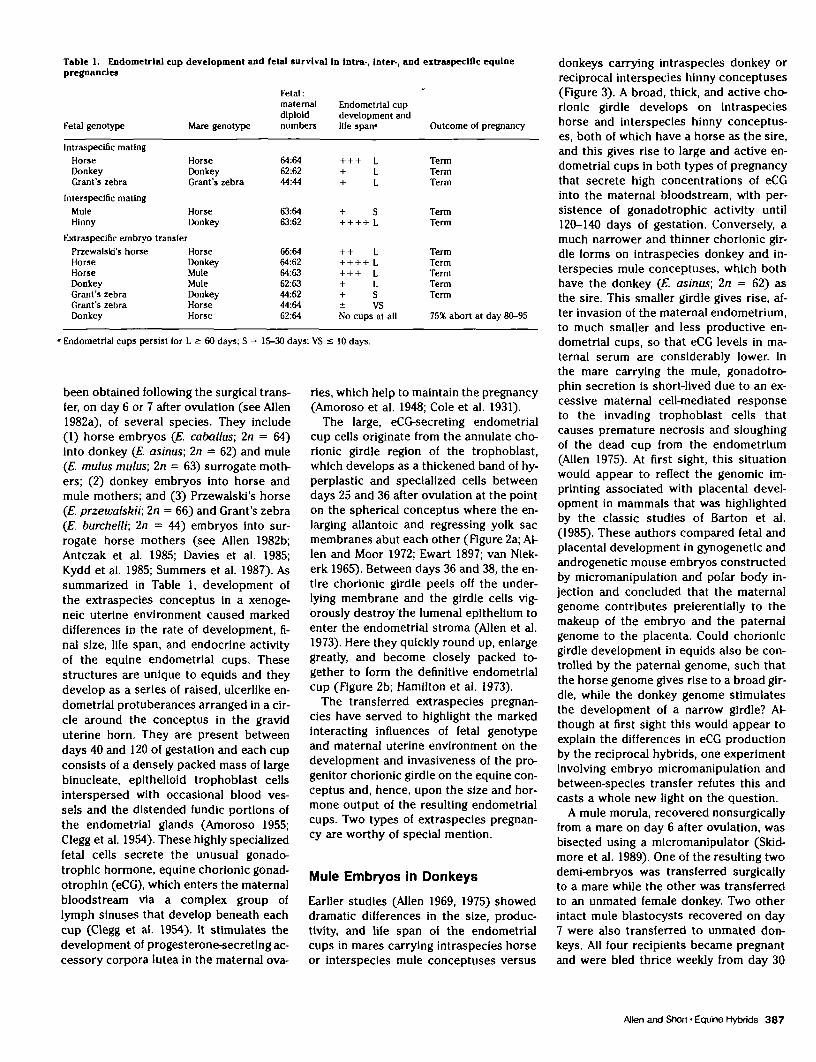
Table 1. Endometrlal cnp development and fetal snrvlval In Intra-, inter-, and extraspeciflc equinepregnancies
Fetal genotype Mare genotype
Fetal:maternal Endometrlal cupdiplold development andnumbers life spair* Outcome ol pregnancy
Intraspecific matingHorseDonkeyGrant's zebra
Interspecific matingMuleHlnny
Extraspecific embryo transferPrzewalsld's horseHorseHorseDonkeyGrant's zebraGrant's zebraDonkey
HorseDonkeyGrant's zebra
HorseDonkey
HorseDonkeyMuleMuleDonkeyHorseHorse
64:6462:6244:44
63:6463:62
66:6464:6264:6362:6344*244:6462:64
+ + + L+ L+ L
+ S+ + + + L
+ + L+ + + + L+ + + L+ L+ S± VSNo cups at all
TermTermTerm
TermTerm
TermTermTermTermTerm
75* abort at day 80-95
• Endometrlal cups persist for L 2: 60 days; S = 15-30 days; VS s 10 days.
been obtained following the surgical trans-fer, on day 6 or 7 after ovulation (see Allen1982a), of several species. They include(1) horse embryos (E. caballus; In = 64)into donkey (E. asinus; 2n = 62) and mule(E. mulus mulus; In = 63) surrogate moth-ers; (2) donkey embryos into horse andmule mothers; and (3) Przewalski's horse(E. przewalskii; In = 66) and Grant's zebra(E. burchelli; 2n = 44) embryos into sur-rogate horse mothers (see Allen 1982b;Antczak et al. 1985; Davies et al. 1985;Kydd et al. 1985; Summers et al. 1987). Assummarized in Table 1, development ofthe extraspecies conceptus in a xenoge-neic uterine environment caused markeddifferences in the rate of development, fi-nal size, life span, and endocrine activityof the equine endometrial cups. Thesestructures are unique to equids and theydevelop as a series of raised, ulcerlike en-dometrial protuberances arranged in a cir-cle around the conceptus in the graviduterine horn. They are present betweendays 40 and 120 of gestation and each cupconsists of a densely packed mass of largeblnucleate, epithelioid trophoblast cellsinterspersed with occasional blood ves-sels and the distended fundic portions ofthe endometrial glands (Amoroso 1955;Clegg et al. 1954). These highly specializedfetal cells secrete the unusual gonado-trophic hormone, equine chorionic gonad-otrophin (eCG), which enters the maternalbloodstream via a complex group oflymph sinuses that develop beneath eachcup (Clegg et al. 1954). It stimulates thedevelopment of progesterone-secreting ac-cessory corpora lutea in the maternal ova-
ries, which help to maintain the pregnancy(Amoroso et al. 1948; Cole et al. 1931).
The large, eCG-secreting endometrialcup cells originate from the annulate cho-rionic girdle region of the trophoblast,which develops as a thickened band of hy-perplastic and specialized cells betweendays 25 and 36 after ovulation at the pointon the spherical conceptus where the en-larging allantoic and regressing yolk sacmembranes abut each other (Figure 2a; Al-len and Moor 1972; Ewart 1897; van Niek-erk 1965). Between days 36 and 38, the en-tire chorionic girdle peels off the under-lying membrane and the girdle cells vig-orously destroy the lumenal epithelium toenter the endometrial stroma (Allen et al.1973). Here they quickly round up, enlargegreatly, and become closely packed to-gether to form the definitive endometrlalcup (Figure 2b; Hamilton et al. 1973).
The transferred extraspecies pregnan-cies have served to highlight the markedInteracting influences of fetal genotypeand maternal uterine environment on thedevelopment and invasiveness of the pro-genitor chorionic girdle on the equine con-ceptus and, hence, upon the size and hor-mone output of the resulting endometrialcups. Two types of extraspecies pregnan-cy are worthy of special mention.
Mule Embryos in Donkeys
Earlier studies (Allen 1969, 1975) showeddramatic differences in the size, produc-tivity, and life span of the endometrialcups in mares carrying intraspecies horseor interspecies mule conceptuses versus
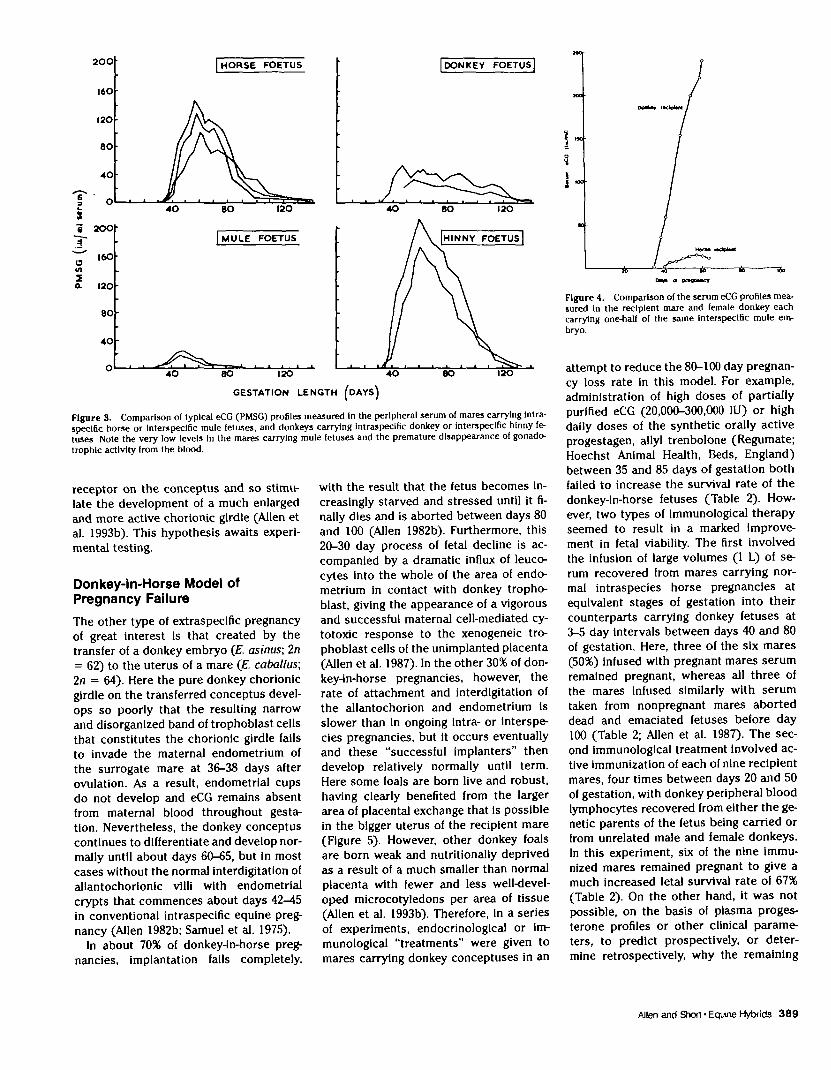
donkeys carrying intraspecies donkey orreciprocal interspecies hinny conceptuses(Figure 3). A broad, thick, and active cho-rionic girdle develops on intraspecieshorse and interspecies hinny conceptus-es, both of which have a horse as the sire,and this gives rise to large and active en-dometrial cups in both types of pregnancythat secrete high concentrations of eCGinto the maternal bloodstream, with per-sistence of gonadotrophic activity until120-140 days of gestation. Conversely, amuch narrower and thinner chorionic gir-dle forms on intraspecies donkey and in-terspecies mule conceptuses, which bothhave the donkey (E. asinus; In = 62) asthe sire. This smaller girdle gives rise, af-ter invasion of the maternal endometrium,to much smaller and less productive en-dometrial cups, so that eCG levels in ma-ternal serum are considerably lower. Inthe mare carrying the mule, gonadotro-phin secretion is short-lived due to an ex-cessive maternal cell-mediated responseto the invading trophoblast cells thatcauses premature necrosis and sloughingof the dead cup from the endometrium(Allen 1975). At first sight, this situationwould appear to reflect the genomic im-printing associated with placental devel-opment in mammals that was highlightedby the classic studies of Barton et al.(1985). These authors compared fetal andplacental development in gynogenetlc andandrogenetic mouse embryos constructedby micromanipulation and polar body in-jection and concluded that the maternalgenome contributes preferentially to themakeup of the embryo and the paternalgenome to the placenta. Could chorionicgirdle development in equids also be con-trolled by the paternal genome, such thatthe horse genome gives rise to a broad gir-dle, while the donkey genome stimulatesthe development of a narrow girdle? Al-though at first sight this would appear toexplain the differences In eCG productionby the reciprocal hybrids, one experimentInvolving embryo micromanipulation andbetween-specles transfer refutes this andcasts a whole new light on the question.
A mule morula, recovered nonsurgicallyfrom a mare on day 6 after ovulation, wasbisected using a micromanipulator (Skid-more et al. 1989). One of the resulting twodemi-embryos was transferred surgicallyto a mare while the other was transferredto an unmated female donkey. Two otherintact mule blastocysts recovered on day7 were also transferred to unmated don-keys. All four recipients became pregnantand were bled thrice weekly from day 30
Allen and Short • Equine Hybrids 3 8 7
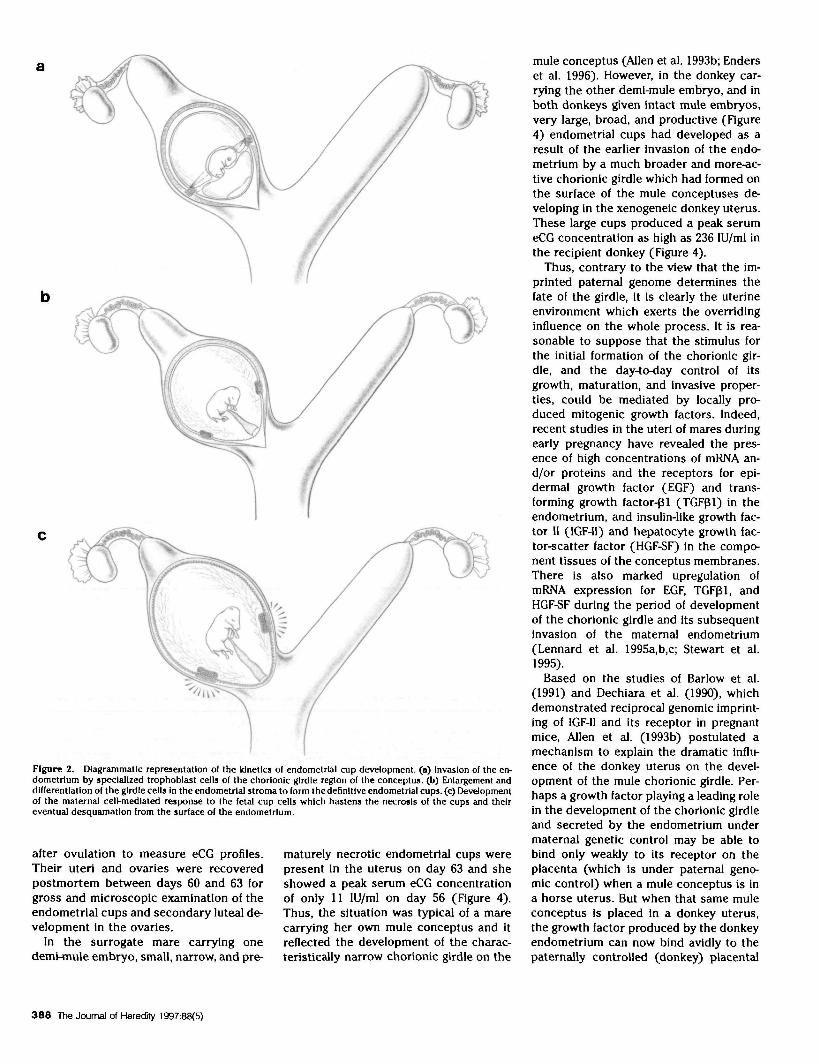
Figure 2. Diagrammatic representation of the Idnetics of endometrtal cup development, (a) Invasion of the en-dometrium by specialized trophoblast ceils of the chorlonlc girdle region of the conceptus. (b) Enlargement anddifferentiation of the girdle cells In the endometrial stroma to form the definitive endometrial cups, (c) Developmentof the maternal cell-mediated response to the fetal cup cells which hastens the necrosis of the cups and theireventual desquamatlon from the surface of the endometrlum.
after ovulation to measure eCG profiles.Their uteri and ovaries were recoveredpostmortem between days 60 and 63 forgross and microscopic examination of theendometrial cups and secondary luteal de-velopment in the ovaries.
In the surrogate mare carrying onedemi-mule embryo, small, narrow, and pre-
maturely necrotlc endometrial cups werepresent in the uterus on day 63 and sheshowed a peak serum eCG concentrationof only 11 IU/ml on day 56 (Figure 4).Thus, the situation was typical of a marecarrying her own mule conceptus and itreflected the development of the charac-teristically narrow chorionic girdle on the
mule conceptus (Allen et al. 1993b; Enderset al. 1996). However, in the donkey car-rying the other demi-mule embryo, and inboth donkeys given intact mule embryos,very large, broad, and productive (Figure4) endometrial cups had developed as aresult of the earlier invasion of the endo-metrium by a much broader and more-ac-tive chorionic girdle which had formed onthe surface of the mule conceptuses de-veloping in the xenogeneic donkey uterus.These large cups produced a peak serumeCG concentration as high as 236 IU/ml inthe recipient donkey (Figure 4).
Thus, contrary to the view that the im-printed paternal genome determines thefate of the girdle, it is clearly the uterineenvironment which exerts the overridinginfluence on the whole process. It is rea-sonable to suppose that the stimulus forthe initial formation of the chorionic gir-dle, and the day-to-day control of itsgrowth, maturation, and invasive proper-ties, could be mediated by locally pro-duced mitogenic growth factors. Indeed,recent studies in the uteri of mares duringearly pregnancy have revealed the pres-ence of high concentrations of mRNA an-d/or proteins and the receptors for epi-dermal growth factor (EGF) and trans-forming growth factor-61 (TGFpi) in theendometrium, and insulin-like growth fac-tor II (IGF-II) and hepatocyte growth fac-tor-scatter factor (HGF-SF) in the compo-nent tissues of the conceptus membranes.There Is also marked upregulation ofmRNA expression for EGF, TGFB1, andHGF-SF during the period of developmentof the chorionic girdle and Its subsequentInvasion of the maternal endometrium(Lennard et al. 1995a,b,c; Stewart et al.1995).
Based on the studies of Barlow et al.(1991) and Dechiara et al. (1990), whichdemonstrated reciprocal genomic imprint-ing of IGF-II and its receptor in pregnantmice, Allen et al. (1993b) postulated amechanism to explain the dramatic influ-ence of the donkey uterus on the devel-opment of the mule chorionic girdle. Per-haps a growth factor playing a leading rolein the development of the chorionic girdleand secreted by the endometrium undermaternal genetic control may be able tobind only weakly to its receptor on theplacenta (which is under paternal geno-mic control) when a mule conceptus is ina horse uterus. But when that same muleconceptus Is placed in a donkey uterus,the growth factor produced by the donkeyendometrium can now bind avidly to thepaternally controlled (donkey) placental
388 The Journal erf Heredity 1997:88(5)
2OO
I6O
ISO
8O
4O
T °si soo
om
0. 120
8O
4O
O
| HORSE FOETUS DONKEY FOETUS
4O BO I2O 12O
MULE FOETUS
4O BO I2O 4O
GESTATION LENGTH (DAYS)
BO I2O
Figure 3. Comparison of typical eCG (PMSG) profiles measured In the peripheral serum of mares carrying Intra-speclfic horse or Interspecific mule fetuses, and donkeys carrying intraspecific donkey or interspecific hlnny fe-tuses Note the very low levels in the mares carrying mule fetuses and the premature disappearance of gonado-trophlc activity from the blood.
receptor on the conceptus and so stimu-late the development of a much enlargedand more active chorionic girdle (Allen etal. 1993b). This hypothesis awaits experi-mental testing.
Donkey-in-Horse Model ofPregnancy Failure
The other type of extraspecific pregnancyof great interest is that created by thetransfer of a donkey embryo (E. asinus; 2n= 62) to the uterus of a mare (£. caballus;In = 64). Here the pure donkey chorionicgirdle on the transferred conceptus devel-ops so poorly that the resulting narrowand disorganized band of trophoblast cellsthat constitutes the chorionic girdle failsto invade the maternal endometrium ofthe surrogate mare at 36-38 days afterovulation. As a result, endometrial cupsdo not develop and eCG remains absentfrom maternal blood throughout gesta-tion. Nevertheless, the donkey conceptuscontinues to differentiate and develop nor-mally until about days 60-65, but in mostcases without the normal interdigitation ofallantochorionic villi with endometrialcrypts that commences about days 42—45in conventional intraspecific equine preg-nancy (Allen 1982b; Samuel et al. 1975).
In about 70% of donkey-in-horse preg-nancies, implantation fails completely,
with the result that the fetus becomes in-creasingly starved and stressed until it fi-nally dies and is aborted between days 80and 100 (Allen 1982b). Furthermore, this20-30 day process of fetal decline is ac-companied by a dramatic influx of leuco-cytes into the whole of the area of endo-metrium In contact with donkey tropho-blast, giving the appearance of a vigorousand successful maternal cell-mediated cy-totoxic response to the xenogeneic tro-phoblast cells of the unimplanted placenta(Allen et al. 1987). In the other 30% of don-key-in-horse pregnancies, however, therate of attachment and interdigitation ofthe allantochorion and endometrium isslower than in ongoing intra- or interspe-cies pregnancies, but it occurs eventuallyand these "successful implanters" thendevelop relatively normally until term.Here some foals are born live and robust,having clearly benefited from the largerarea of placenta! exchange that is possiblein the bigger uterus of the recipient mare(Figure 5). However, other donkey foalsare born weak and nutritionally deprivedas a result of a much smaller than normalplacenta with fewer and less well-devel-oped microcotyledons per area of tissue(Allen et al. 1993b). Therefore, in a seriesof experiments, endocrinological or im-munological "treatments" were given tomares carrying donkey conceptuses in an
Figure 4. Comparison of the serum eCG profiles mea-sured in the recipient mare and female donkey eachcarrying one-hall of the same Interspecific mule em-bryo.
attempt to reduce the 80-100 day pregnan-cy loss rate in this model. For example,administration of high doses of partiallypurified eCG (20,000-300,000 IU) or highdaily doses of the synthetic orally activeprogestagen, allyl trenbolone (Regumate;Hoechst Animal Health, Beds, England)between 35 and 85 days of gestation bothfailed to increase the survival rate of thedonkey-in-horse fetuses (Table 2). How-ever, two types of immunological therapyseemed to result in a marked improve-ment in fetal viability. The first involvedthe infusion of large volumes (1 L) of se-rum recovered from mares carrying nor-mal intraspecies horse pregnancies atequivalent stages of gestation into theircounterparts carrying donkey fetuses at3-5 day intervals between days 40 and 80of gestation. Here, three of the six mares(50%) infused with pregnant mares serumremained pregnant, whereas all three ofthe mares infused similarly with serumtaken from nonpregnant mares aborteddead and emaciated fetuses before day100 (Table 2; Allen et al. 1987). The sec-ond immunological treatment involved ac-tive immunization of each of nine recipientmares, four times between days 20 and 50of gestation, with donkey peripheral bloodlymphocytes recovered from either the ge-netic parents of the fetus being carried orfrom unrelated male and female donkeys.In this experiment, six of the nine immu-nized mares remained pregnant to give amuch increased fetal survival rate of 67%(Table 2). On the other hand, it was notpossible, on the basis of plasma proges-terone profiles or other clinical parame-ters, to predict prospectively, or deter-mine retrospectively, why the remaining
Allen and Short • Equine Hybrids 3 8 9

Figure 5. (a) A recipient horse mare with her transferred extraspeclfic donkey foal, and (b) a female donkeyrecipient with her transferred extraspecific horse foal. Note the great difference In size between the foals comparedto the size of the genetic parents, caused by the donkey developing In a larger-than-normal uterus and the horsefoal developing In the uterus of the smaller donkey mother.
Table 2. The extraapecifle donkey-ln-horse pregnancy model or pregnancy failure: endocrinologies!and Immunologlcal treatments
TreatmentNo. ofmares
No. that abortedbefore day 120
Fetal sur-vival («)
Control 24 19
Endocrinologlcal (from day 36)Allyl trenbolone (progestagen) 4 4eCG (high doses) ? 8
ImmunologlcalInfusion of pregnant mares' serum (day 40-80) 6 3Infusion of nonpregnant mares' serum (day 40-80) (control) 3 3Immunized against donkey lymphocytes (day 20-50) 9 3
21
014
500
67
three immunized mares aborted their don-key conceptuses (Allen et al. 1987).
These inexplicable differences in the re-sponses of individual mares when carry-ing donkey conceptuses beg the questionof whether the whole phenomenon of ear-ly pregnancy failure in this model is me-diated immunologically or has a geneticbasis. Two further findings argue for acombination of these two influences, asexemplified by the high correlation thatexists in the outcome of successive don-key pregnancies established in the samerecipient mares. On the one hand, of 10mares that carried their first donkey preg-nancy successfully, no fewer than 9 car-ried a second donkey conceptus withoutany form of treatment, and 2 of these car-ried a third donkey pregnancy, again with-out treatment. On the face of it, this resultseems to hint strongly of a genetic com-ponent in the recipient mares wherebyone has, in simple terms, "good mothers"and "bad mothers" with regard to the abil-ity to gestate a foreign donkey embryo (Al-len et al. 1987). In contrast, however, all ofseven mares that aborted their first don-key pregnancies aborted second donkeyconceptuses transferred to them in the fol-lowing breeding season; six of these sevenabortions occurred significantly earlier ingestation and both of the third successiveabortions of donkey fetuses carried out intwo of the mares occurred even earlierstill. This pronounced decrease in the in-terval from embryo transfer to abortion insecond and later donkey-in-horse pregnan-cies is reminiscent of immune memory,one of the hallmarks of the immune sys-tem (Allen et aJ. 1987). Clearly, much morework is needed before we will even beginto distinguish between the two phenome-na.
Conclusions
In summary, the genus Equus is of consid-erable interest because of the ability of itsmember species to produce a wide rangeof interspecific hybrids, which appear toobey Haldane's law, and to carry an arrayof phenotypically, genetically, and immu-nologically different interspecific and ex-traspecific pregnancies. This readiness ofthe maternal immune system to acceptsuch "foreignness" in the uterus duringthe course of gestation is all the more sur-prising in view of the unusual injection ofspecialized trophoblast cells that expresshigh levels of major histocompatibilltycomplex (MHC) class I (Donaldson et al.1992), species-related (Allen 1975) and tis-
3 9 0 The Journal of Heredity 1997:88(5)

Figure 6. Genetic drift In the nursery. Three recipient pony mares with their transferred extraspeclfic Przewal-sld's horse (left), donkey (center), and Grant's zebra (right) foals.
sue-speclfic (Oriol et al. 1989) antigensinto the maternal endometrium to providethe endometrial cup reaction (Allen et al.1973). This considerable antigenic chal-lenge to the mother is clearly recognizedand responded to by her, as exemplifiedby her production of high titres of allo-specific lymphocytotoxic antibody (Allen1979; Antczak et al. 1984) and her vigorouscell-mediated reaction against the endo-metrial cups, especially in the interspeciesand extraspecies pregnancies (Allen 1975;Allen et al. 1987).
But perhaps of even greater interest isthe evidence provided by the donkey-in-horse model of extraspecies pregnancy,which points to a definite need for the en-dometrial cup reaction for placentationand fetal survival, not as a result of thehormonal output of the cups, but becauseof the strong antigenic stimulus the invad-ing trophoblast cells give to the maternalorganism. If the annulate chorionic girdledoes not develop properly and fails to in-vade the maternal endometrium aboutdays 36-38, the implantatlon/placental in-terdlgitation reaction is not triggered inthe endometrium and a savage cell-medi-ated attack is mounted against the xeno-geneic placenta and fetus. But if the girdledoes manage to invade the endometrium,the resulting antigenic stimulation of themother induces in her a protective, ratherthan cytotoxic, immunological responsetoward her fetus. The amazing strengthand success of this protective reaction,when elicited correctly, is illustrated bythe considerable genetic diversity of theoffspring of the group of experimentalpony mares shown In Figure 6.
ReferencesAllen WR, 1969. Factors Influencing pregnant mare se-rum gonadotrophln production. Nature 223*4-66.
Allen WR, 1975. The Influence of fetal genotype upon
endometrial cup development and PMSG and proges-tagen production In equlds. J Reprod Fert (suppl 23):405-413.
Allen WR, 1979. Maternal recognition of pregnancy andImmunological Implications of trophoblast-endometrt-um interactions In equlds. In: Maternal recognition ofpregnancy. CIBA Foundation Symposium no 64. Am-sterdam: Excerpta Medlca; 323-352.
Allen WR, 1982a. Embryo transfer In the horse. In.Mammalian egg transfer (Adams CE, ed) Boca Raton,Florida: CRC Press; 135-154.
Allen WR, 1982b. Immunological aspects of the equineendometrial cup reaction and the effect of xenogenelcextraspecies pregnancy In horses and donkey. J ReprodFert (suppl 31)57-94.
Allen WR, Hamilton DW, and Moor RM, 1973. The originof the equine endometrial cups. D. Invasion of the en-dometrium by trophoblast. Anat Rec 177.485-502.
Allen WR, Kydd JH, and Antczak DF, 1993a. Interspeciesand extraspecies equine pregnancies. In. Equine repro-duction (McKlnnon AO and Voss JL, eds). Philadelphia:Lea and Feblger, 536-553.
Allen WR, Kydd JH, Boyle MS, and Antczak DF, 1987.Extra-specific donkey-ln-horse pregnancy as a model ofearly fetal death. J Reprod Fert (suppl 35)197-209.
Allen WR and Moor RM, 1972. The origin of the equineendometrial cups. I. Production of PMSG by fetal tro-phoblast cells. J Reprod Fert 29:313-316.
Allen WR, Skidmore JA, Stewart F, and Antczak DF,1993b. Effects of fetal genotype and uterine environ-ment on placental development In equlds. J ReprodFert 97:55-60.
Amoroso EC, 1955. The endocrinology of pregnancy. BrMedBull 17:117-125.
Amoroso EC, Hancock JL, and Rowlands IW, 1948. Ovar-ian activity In the pregnant mare. Nature (Lond) 161:355-356.
Anderson WS, 1939. Fertile mare mules J Hered 30:549-551.
Antczak DF, Davles CJ, Kydd JH, and Allen WR, 1985.Immunological aspects of pregnancy In mules. EquineVet J (suppl 3):68-72.
Antczak DF, Miller J, and Remlck LH, 1984. Lymphocytealloantlgens of the horse. II. Antibodies to ELA antigensproduced during equine pregnancy. J Reprod Immunol6283-287.
Barlow DF, Stoger R, Herrmann BG, Salto K, and Schwel-fer N, 1991. The mouse Insulin-like growth factor type-2receptor is Imprinted and closely linked to the Tmelocus. Nature 349:84-87.
Barton SC, Suranl MAH, and Norrls ML, 1985. Role ofpaternal and maternal genomes in mouse development.Nature 311374-376.
Chandley AC, 1988. Fertile mules. J R Soc Med 81:2.
Chandley AC, Jones RC, Dott HM, Allen WR, and Short
RV, 1974. Melosis In Interspecific equine hybrids. I. Themale mule (Equus asinus X £ caballus') and hlnny (£caballus X £ asinus). Cytogenet Cell Genet 13:330-341.
Chandley AC, Short RV, and Allen WR, 1975. Cytoge-netlc studies of three equine hybrids. J Reprod Fert(suppl 23):365-370.
Chang MC, Marston JH, and Hunt DM, 1964. Reciprocalfertilization between the domesticated rabbit and thesnowshoe hare with special reference to Inseminationof rabbits with an equal number of hare and rabbitspermatozoa. J Exp Zool 155:437^45.
Clegg MT, Boda JM, and Cole HH, 1954. The endome-trial cups and allantochorionlc pouches In the marewith emphasis on the source of equine gonadotropin.Endocrinology 54:448-463.
Cole HH, Howell CF, and Hart GH, 1931. The changesoccurring In the ovary of the mare during pregnancy.Anat Rec 49:199-209.
Craft WA, 1938. The sex ratio In mules and other hybridmammals. Q Rev Blol 13:19-40.
Davles CJ, Antczak DF, and Allen WR, 1985 Reproduc-tion In mules: embryo transfer using sterile recipients.Equine Vet J (suppl 3):63-67.
Dechlara TM, Efstratiadls A, and Robertson EJ, 1990. Agrowth-deficiency phenotype In heterozygous micecarrying an Insulin-like growth (actor 2 gene disruptedby targeting. Nature 345:78-80.
Donaldson WL, Oriol JG, Plavln A, and Antczak DF,1992. Developmental regulation of class I major histo-compatlbllity complex antigen expression by equinetrophoblast cells. Differentiation 52:69-78.
Enders AC, Meadows SJ, Stewart F and Allen WR, 1996.Failure of endometrial cup development in the donkey-ln-horse model of extraspecific equine pregnancy. JAnat 188:575-589.
Ewart JC, 1897. A critical period In the development ofthe horse. London: Adam and Charles Black.
Ewart JC, 1898. On zebra-ass hybrids; with observa-tions on the relationships of the zebras. Veterinarian71185-202.
Gray AP, 1971. Mammalian hybrids. Slough, England:Commonwealth Agricultural Bureaux
Haldane JBS, 1922. Sex ratio and unisexual sterility Inhybrid animals. J Genet 12:101-109.
Hamilton DW, Allen WR, and Moor RM, 1973. The originof equine endometrial cups. III. Ught and electron mi-croscopic study of fully developed equine endometrialcups. Anat Rec 177:503-518.
Hancock JL and McGovem FT, 1964. The transport ofsheep and goat spermatozoa In the ewe. J Reprod Fert15:283-287.
Hancock JL, McGovern FT, and Stamp JT, 1968. Failureof gestation of goat x sheep hybrids In goats and sheep.J Reprod Fert (suppl 3):29-36.
Henry M, Gastal EL, Plsjhelro LEL, and Guimaraes SEF,1995. Mating patterns and chromosome analysis of amule and her offspring. Biol Reprod monogr ser 1:273—280
Jocelyn HD and Setchell BP (trans), 1972. Regnier deGraaf on the human reproductive organs. J Reprod Fert(suppl 17).
King JM, 1965. Comparative aspects of reproduction inequldae (PhD dissertation). Cambridge: CambridgeUniversity.
Koullscher L and Frechkop S, 1966. Chromosome com-plement: a fertile hybrid between Equus przewalskiiand Equus caballus. Science 151:93-95.
Kydd J, Boyle MS, Allen WR, Shepard A, and SummersPM, 1985. Transfer of exotic equine embryos to do-mestic horse* and donkeys. Equine Vet J (suppl 3):80-84.
Lennard SN, Stewart F, and Allen WR, 1995a. Insulin-likegrowth factor II gene expression In the fetus and pla-centa of the horse during the first half of gestation. JReprod Fert 103:169-179.
Allen and Short • Equine Hybrids 391

Lennard SN, Stewart F, and Allen WR, 1995b. Trans- Samuel CA, Allen WR, and Steven DH, 1975. Ultrastruc- bryos of Przewalskl's horse (Equus przewalskii) andforming growth factor bl (TGFbl) expression In the tural development of the equine placenta J Reprod Grant's zebra (£ burchelli) to domestic mares (£ co-endometrium of the mare during placentatlon. Mol Re- Fert (suppl 23):575-578. ballus). J Reprod Fert 80:13-20.prod Dev 42:131-140. S h o r t ^ ] 9 ? 5 a ^ e v o l u t l o n of ( h e h o r s e j Rgp,.^ T a y l o r M ^ j ihort RV, 1973. Development of the germLennard SN, Stewart F, Allen WR, and Heap RB, 1995c. Fert (suppl 23):l-6. cells in the ovary of the mule and hinny. J Reprod FertGrowth factor production in the equine uterus. Blol Re- S h o r t R v , 9 7 5 b ^ c o n t r l b u t i o n of t h e m u l e t 0 K l . 32:441-445.prod monogr ser l.lbl-1 /u. e n t | f i c t n o u g n t j Reprod Fert (suppl 23)^59-364. Tegetmeier WB and Sutherland CL, 1895. Horses, asses,
Mlchie D, 1953. Affinity: a new genetic phenomenon in S n o r t RV| chandley AC, Jones RC, and Allen WR, 1974 z e b r a S ' m u l e S ^ m U ' e b r e e d l n 8 - London- H o r a c e Cox-the house mouse, evidence from distant crosses. Na- Me|osls In interspecific equine hybrids. U. The Prze- Tucker PK and Lundrigan BU 1993. Rapid evolution ofture 171.26-27. walskl horse/domestic horse hybrid (Equus przewalskii the sex determining locus In Old World mice and rats.Orlol JG, Poleman DF, Antczak DF, and Allen WR, 1989. x £ caballus). Cytogenet Cell Genet 13:465-178. Nature 364:715-717.A monoclonal antibody specific for equine trophoblast. SUdmore JA, Boyle MS, Cran D, and Allen WR, 1989. van Nlekerk CH, 1965. The early diagnosis of pregnan-Equlne Vet J (suppl 8):14-18. Micromanlpulatlon of equine embryos to produce mon- cy, the development of the foetal membranes and nl-Rong R, Chandley AC, Song J, McBeath S, Tan PP, Bai o zy8° t l c t w l n s- E t ' u l n e V e t J ('"PP1 8):126-12S- datlon in the mare. J S Air Vet Med Assoc 36:483-188.Q, and Speed RM, 1988. A fertile mule and hinny In Stewart F, Lennard SN, and Allen WR, 1995. Mecha- Whltfield LS, Lovell-Badge R, and Goodfellow PN, 1993.China. Cytogenet Cell Genet 47:134-139. nlsms controlling formation of the equine chorlonlc glr- Rapid sequence evolution of the mammalian sex-deter-o j r.» r-u 11. ir- D J, »T J D , ui die. Biol Reprod monogr ser 1:487-495. mining gene SRY. Nature 364:713-715.Ryder DA, Chemmick LG, Bowling AT, and Benlrschke K o 55K, 1985. Male mule qualifies as the offspring of a female Summers PM, Shephard AM, Hodges JK, Kydd J, Boyle Wodsedalek JE, 1916. Causes of sterility In the mule,mule and jack donkey. J Hered 76379-381. MS. and Allen WR, 1987. Successful transfer of the em- Blol Bull Mar Biol Lab Woods Hole 30 1-56.
3 9 2 The Journal of Heredity 1997:88(5)





























