Embed Size (px)
Citation preview

Rom J Leg Med 15 (3) 167 – 181 (2007) © 2007 Romanian Society of Legal Medicine
________________________ * Corresponding author, Prof. Univ. Dr. Gheorghe S. Drăgoi, MD, PhD, Member of
Academy of Medical Sciences, e-mail: [email protected], Academic Hospital No. 2, Craiova.
167
Interrelaţii tisulare în evoluţia barierelor hemochoriale. Implicaţii în patologie şi expertiza medico-legală a gravidităţii şi avortului
Gheorghe S. Drăgoi *, Liviu Radu, Mugurel Robert Popescu, Radu Neamţu _____________________________________________________________________ ABSTRACT: Tissue Interrelations in the Evolution of Hemochorial Barriers. Implications in Forensic Expertise of Pregnancy and Abortion: Homo Sapiens Sapiens placenta is of hemochorial type. In pregnancy evolution there is a phenotype transformation of villous structures. Therefor, we shall move from hemobichorial – maternoembrionar type to hemomonochorial – maternofetal type.The authors have noticed the structural differences between the two villous types through the evaluation of tissue interrelations in between hemochorial barriers differentiating the fibrinoid and villous elements of extracellular matrix. The existence of PAS positive and fibrinoid storage are the main factors for accumulating calcar salts in citotrofoblast location.Those observations are important in the medical expertise because they bring arguments upon the changes that come in disturbing maternofetale circulation. Keywords : placenta, placentar villous, hemochorial barrier, trophoblast.
A. Motivaţia, scopul şi obiectivele cercetării
valuarea interrelaţiilor tisulare în etapele genezei, creşterii şi diferenţierii vilozităţilor choriale este fundamentală pentru rigurozitatea diagnosticului
microanatomic de transformare fenotipică a barierelor hemochoriale. Placenta de Homo Sapiens Sapiens este de tip hemochorial. Particularităţile structurale ale placentei sunt determinate de geneza şi evoluţia elementelor sale structurale: chorionul vilozitar (Chorion frondosum) din partea fetală (Pars fetalis) şi decidua bazală sau caduca bazală (Decidua basalis) din porţiunea maternă a placentei (Pars materna). Hotarele placentei sunt realizate de două structuri numite plăci: placa chorială (Lamina corionica) şi placa deciduală sau stratul compact din decidua bazală (Pars basalis). Joncţiunea structurilor trofoblastice cu decidua bazală este realizată la nivelul membranei bazale subdeciduale. Între cele două plăci, chorială şi deciduală există camerele intervilozitare (Spatium intervillosum) în care circulă sângele matern din sistemul vascular uteroplacentar.
E

GS Dragoi Tissue Interrelations in the Evolution of Hemochorial Barriers
168
În patologia generală şi medicolegală, diagnosticul de sarcină se impune înaintea precizării cauzelor de întrerupere a gestaţiei. Placenta considerată ca un întreg dar şi vilozităţile choriale izolate sunt probanţi anatomici ai gestaţiei. Întreruperea spontană a sarcinii poate fi determinată fie de tulburările circulaţiei sanguine, fie de perturbări ale procesului de placentogeneză. Se consideră că diagnosticul de certitudine al sarcinii se bazează pe prezenţa vilozităţilor choriale (Kernbach, 1953; Beliş, 1990). Leziunile structurilor placentei pot fi argumente suplimentare în susţinerea diagnosticului de avort spontan (involuntar) în care unii autori includ expulzia fătului oprit în evoluţie şi reţinut un timp variabil în cavitatea uterină (Lacassagne şi Martin, 1921), iar alţii şi expulzia unui făt neviabil care moare rapid după expulzie (Durigon, 1988). Heterogenitatea structurală şi variabilitatea descrierilor elementelor microanatomice sunt de multe ori bariere greu de depăşit în formularea răspunsurilor la numeroase întrebări:
Există o filiaţie între cito- şi sinciţiotrofoblast în timpul gestaţiei? Ce sunt, de fapt, celulele Langhans? De ce dispare citotofroblastul după primul trimestru de gestaţie? Persistă în vilozităţile hemo-monochoriale sinciţiotrofoblastul şi care este originea sa
în caz afirmativ? În practica histopatologică, prezenţa fibrinoidului placentar ridică probleme legate de
originea şi structura sa. Deşi fibrinoidul a fost semnalat la nivelul placentei în anul 1877 de catre Langhans, totuşi în istoria cunoaşterii sale au fost formulate mai multe întrebări decât răspunsuri: Care este rolul îndeplinit de fibrinoid în homeostazia biosistemului placentar?; Cum intervine el în geneza şi transformarea fenotipică a vilozităţilor placentare?; Este el un factor inductor în interacţiunile epiteliomezenchimale? Depunerea anormală de săruri de calciu în structurile placentei (calcinoza) este relativ frecventă. Fujikura (1963) a observat că 15% din placente au grade variabile de depuneri calcare. Se cunosc puţine date legate de tranzitul ionilor de calciu prin membranele fetoplacentare. Procesul de calcipexie este incomplet cunoscut. Scopul lucrării este înţelegerea transformărilor fenotipice temporospaţiale a vilozităţilor hemochoriale şi de a cunoaşte semnificaţia modificărilor structurilor sistemului placentar ce pot fi implicate în oprirea evoluţiei, în moartea şi reţinerea embrionului sau fătului în cavitatea uterină, urmate de declanşarea avortului patologic, de cunoaşterea contribuţiei fibrinoidului la procesele complexe de geneză şi evoluţie a vilozităţilor placentare, de cunoaşterea stereodistribuţiei sărurilor de calciu în structurile placentare în vederea evaluării riguroase a consecinţelor calcinozei asupra barierelor hemochoriale în cazul expertizelor medicolegale. B. Probleme legate de terminologia utilizată
Abateri în utilizarea unor termeni de anatomie macro- şi microscopică prin necunoaşterea încărcăturii lor semantice generează confuzii în cunoaşterea structurilor funcţionale ale placentei. Din această categorie fac parte termenii de chorion, deciduă, calcinoză, calcificare.
Chorionul (Gr. Khorion = Membrană a fătului) este termenul introdus în embriologie pentru nominalizarea structurilor care se diferenţiază din trofoblast şi care participă la formarea membranei choriale. Din luna a doua de viaţă antepartum trofoblastul ia numele de chorion. Porţiunea funcţională a chorionului, în descrierile de embriologie este chorionul vilos (Chorion frondosum) care împreună cu decidua bazală formează placenta. Preluarea acestui

Romanian Journal of Legal Medicine Vol. XV, nr. 3, 2007
169
termen de către histologi pentru definirea „lamei de ţesut conjuctiv” localizată în profunzimea mucoaselor, seroaselor sau dermului a realizat deturnarea conţinutului semantic al termenului de chorion. Această eroare a rămas adânc înrădăcinată în manualele de histologie, deşi în paralel au fost introduşi termenii de stromă şi de matrix extracelular în tratatele de anatomie microscopică.
Decidua (Lat. Deciduus = care cade; de la Decidere = a cădea) este termenul utilizat pentru descrierea stratului funcţional al endometrului din structura placentei care se va detaşa în momentul delivrenţei. Partea bazală a deciduei (Decidua basalis) realizează legătura strânsă cu corionul vilozitar ca element fetal al placentei.
Calcinoza (Lat. Calx, calcis = var, calciu) este termenul utilizat pentru depunerea anormală de săruri de calciu în parenchimul unui organ. Ea poate fi circumscrisă sau generalizată. În studiul placentei, în mod eronat se foloseşte termenul de calcificare (Lat. Calx, calcis = var, calciu; Lat. Ficare – derivat de la facere = a face). Din opunct de vedere etimologic, termenul de calcificare reprezintă o producere de calciu. Din punct de vedere semantic, sensul termenului de calcificare variază atât în ortologie, cât şi în patologie. Deşi în condiţiile genezei osului, el este folosit pentru nominalizarea unui proces fiziologic în timpul osificării, totuşi în patologie este frecvent utilizat pentru depunerea sărurilor de calciu în ţesuturi sau în organe care nu le conţin în mod normal şi este deetrminată de existenţa unor leziuni degererative inflamatorii sau necrotice. Pentru evitarea confuziilor s-a creat un termen mai general de calcipexie (Lat. Calx, calcis = var, calciu; Gr. Pexis = Fixare) care nominalizează procesul de fixare a calciului la nivelul unei matrice proteice.
C. Material şi metode Studiul s-a realizat pe 1317 placente provenite de la avorturi patologice şi naşteri
premature şi la termen. Placentele au fost prelucrate riguros înainte de fixare şi de includere la parafină. Analiza macroscopică a fost însoţită permanent de macrofotografii ale discului placentar ca un întreg, dar şi a secţiunilor seriate secante, perpendiculare pe faţa fetală (Figura nr. 1). 860 de placente au fost utilizate pentru evaluarea microanatomică a transformărilor fenotipice a vilozităţilor hemochoriale, 227 de placente au fost analizate în scopul stabilirii microtopografiei fibrinoidului placentar şi 230 de placente, pentru analiza locaţiei procesului de calcipexie. Examinarea secţiunilor microanatomice şi achiziţia de imagine s-au realizat cu camera fotodigitală anexată Fotonomicroscopului de Cercetare Nikon Elipse 600.
D. Rezultate
1. Analiza microanatomică a barierelor hemobichoriale din primul trimestru al gestaţiei
Pe secţiunile seriate efectuate prin fragmentele recoltate din placentele de prim trimestru al gestaţiei, am remarcat o homogenitate structurală prin prezenţa „vilozităţilor mezenchimale” secţionate sub unghiuri variabile (Figura nr. 3). Diametrul transvers mediu al acestor vilozităţi este de 178 de microni cu variaţii de amplitudine între 118 – 248 microni. În structura axei centrale am identificat mezenchimul vilozitar străbătut de rare capilare sanguine. Periferia vilozităţilor este înconjurată de un strat de epiteliu trofoblastic (Figura nr. 3) alcătuit din celule Langhans (citotrofoblastul din Terminologia Internaţională ) şi un syncitium (sinciţiotrofoblastul din Terminologia Internaţională). Membrana bazală subtrofoblastică este bine vizibilă pe secţiunile de impregnare argentică după metoda Gömöri (Figura nr. 3 H). La examinarea secţiunilor tratate după metoda Mac Mannus se remarcă cu
GS Dragoi Tissue Interrelations in the Evolution of Hemochorial Barriers
170
uşurinţă prezenţa de glicozaminoglicani în locaţia membranei bazale subtrofoblastice dar şi în zona microvilozităţilor sinciţiotrofoblastului (Figura nr. 3 G).
A
C
D B
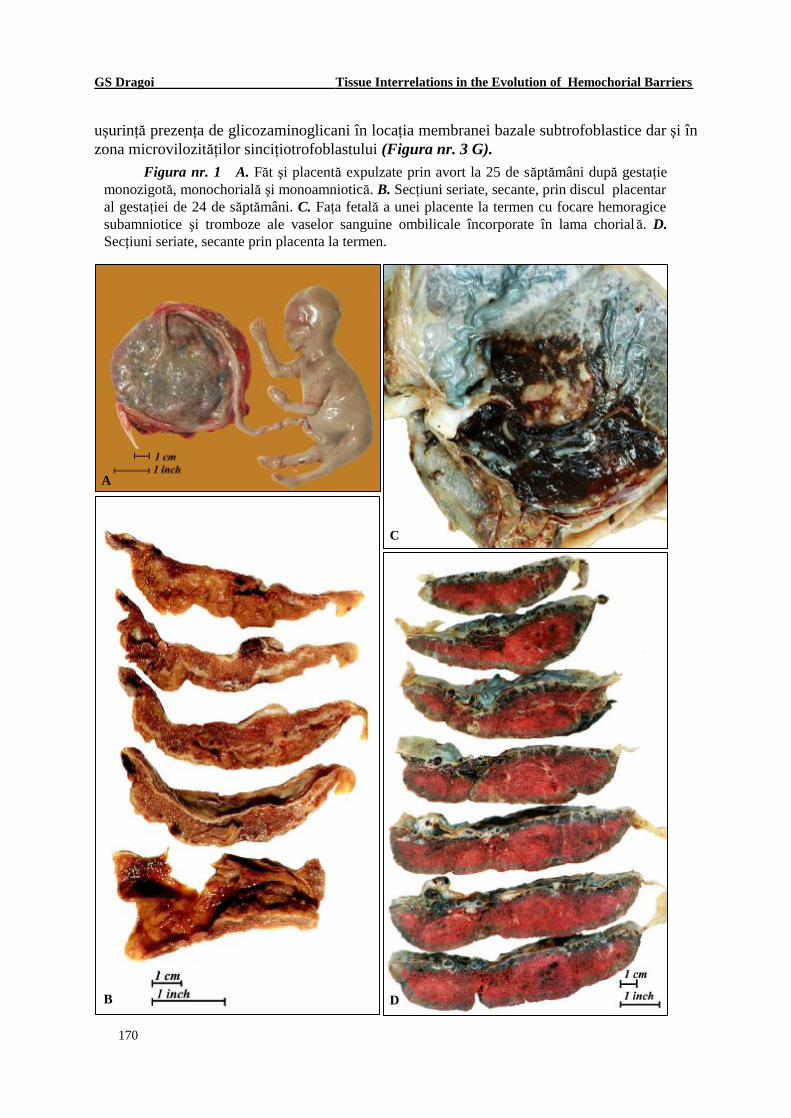
Figura nr. 1 A. Făt şi placentă expulzate prin avort la 25 de săptămâni după gestaţie monozigotă, monochorială şi monoamniotică. B. Secţiuni seriate, secante, prin discul placentar al gestaţiei de 24 de săptămâni. C. Faţa fetală a unei placente la termen cu focare hemoragice subamniotice şi tromboze ale vaselor sanguine ombilicale încorporate în lama chorială. D. Secţiuni seriate, secante prin placenta la termen.

Romanian Journal of Legal Medicine Vol. XV, nr. 3, 2007
171
Granule de săruri calcare au fost identificate, pe secţiunile de impregnare argentică după metoda Kosa, între membrana bazală subtrofoblastică şi celulele Langhans (Figura nr. 3 D). Numeroase vilozităţi sunt în raporturi de contiguitate prin intermediul microvilozităţilor plasmoidale realizând veritabile „sinapse trofoblastice” (Figurile nr. 3 E şi 3 F).
Citotrofoblastul are ataşate capilare sanguine astfel încât membranele bazale subtrofoblastică şi subendotelială capilară devin tangente (Figura nr. 3 F).
A
B C
1
2
3
4 5
6
Epiteliul amniotic
Mezenchimul chorionului
Stratul fibrinoid Langhans
Vas ombilica
Figura nr. 2 Anatomia topografică secţională a placentei la termen: A. Secţiune macroscopică perpendiculară pe suprafaţa fetală a discului placentar şi delimitată de lama chorială (1) şi placa bazală (2). B. Secţiune microanatomică prin placa bazală. C. Secţiune microanatomică prin lama chorială. 1 – Lama chorială; 2 – Placa bazală; 3 – Stratul fibrinoid Rohr; 4 – Stratul fibrinoid uteroplacentar Nitabush; 5 – Membrana bazală subdeciduală; 6 – Decidua bazală (stratul compact). Metode de vizualizare: A. Secţiune macroscopică; B. Impregnare argentică Gömöri; C. Coloraţie Hematoxilină – Eosină. Oc. 7; Ob. 10, x 70 (B, C).
GS Dragoi Tissue Interrelations in the Evolution of Hemochorial Barriers
172
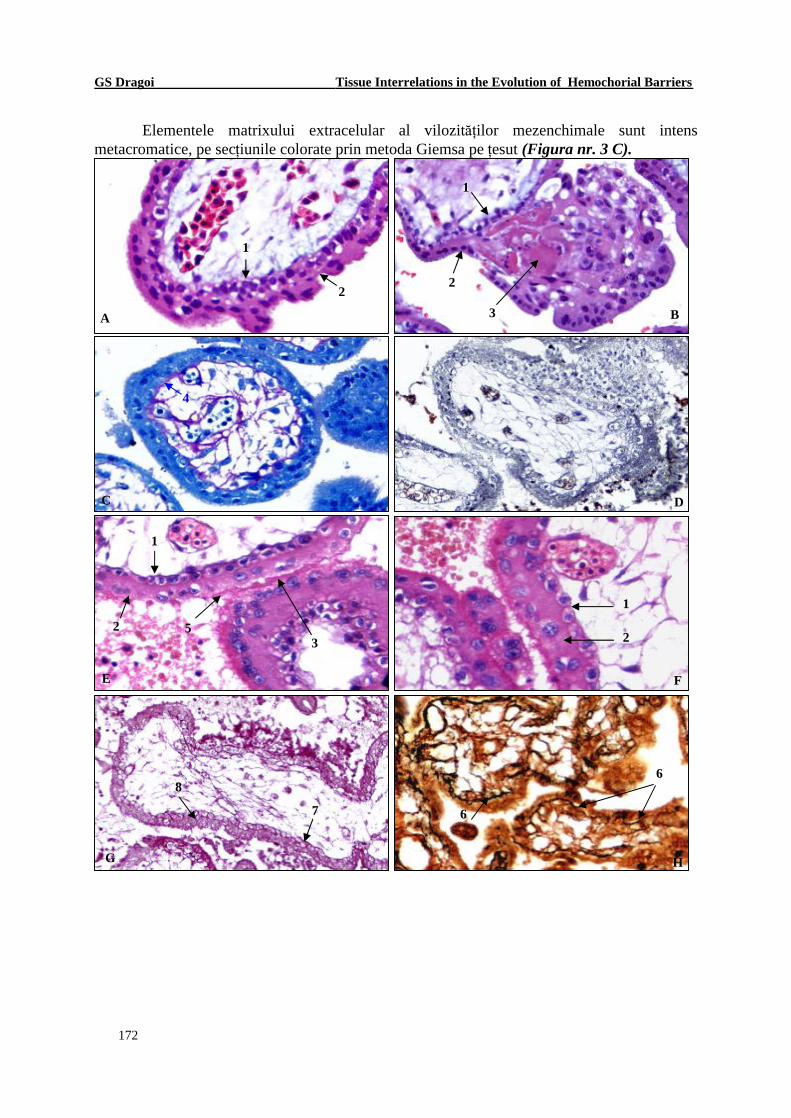
Elementele matrixului extracelular al vilozităţilor mezenchimale sunt intens metacromatice, pe secţiunile colorate prin metoda Giemsa pe ţesut (Figura nr. 3 C).
A B
C D
E F
G H
1
2
1
2
3
4
1
2 5 3
1
2
6
6 7
8

Romanian Journal of Legal Medicine Vol. XV, nr. 3, 2007
173
Figura nr. 3 Particularităţile structurale şi histochimice ale barierei hemo-bichoriale din vilozităţile mezenchimale ale primului trimestru al gesta ţiei. 1. Citotrofoblast. 2. Sinciţiotrofoblast. 3. Substanţă fibrinoidă. 4. Membrană bazală subtrofoblastică – MPZ acid pozitiv. 5. „Sinapsă trofoblastică”. 6. Membrană bazală subtrofoblastică impregnată argentic. 7. Membrană bazală subtrofoblastică PAS pozitivă. 8. Substanţă PAS pozitivă în structura trofoblastului.
Metode de vizualizare : Coloraţia Hematoxilină-Eosină (A; B; E; F); Coloraţia Giemsa pe ţesut (C); Impregnare argentică după metoda Kosa (D), Coloraţie Mac Mannus (G) şi Impregnare argentică Gömöri (H). Oc. 7; Ob. 20 (D; G); 40 (A; B; C; E; F; H); X 140 (D; G); X 280 (A; B; C; E; F; H)
GS Dragoi Tissue Interrelations in the Evolution of Hemochorial Barriers
174
A
B
C
D
1
2
2
3
4 5
6
7
1
9
8
7
6
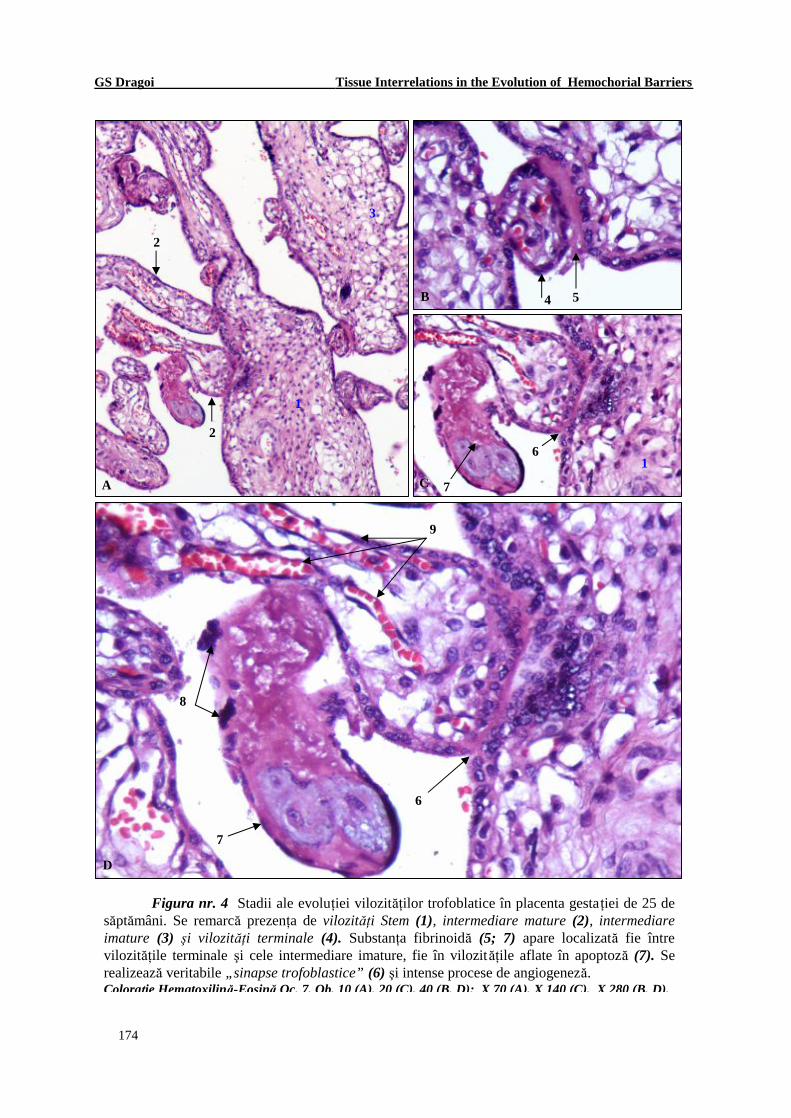
Figura nr. 4 Stadii ale evoluţiei vilozităţilor trofoblatice în placenta gestaţiei de 25 de săptămâni. Se remarcă prezenţa de vilozităţi Stem (1), intermediare mature (2), intermediare imature (3) şi vilozităţi terminale (4). Substanţa fibrinoidă (5; 7) apare localizată fie între vilozităţile terminale şi cele intermediare imature, fie în vilozităţile aflate în apoptoză (7). Se realizează veritabile „sinapse trofoblastice” (6) şi intense procese de angiogeneză. Coloraţie Hematoxilină-Eosină Oc. 7, Ob. 10 (A), 20 (C), 40 (B, D); X 70 (A), X 140 (C), X 280 (B, D).

Romanian Journal of Legal Medicine Vol. XV, nr. 3, 2007
175
Figura nr. 5 Raporturile lamei choriale cu substanţa fibrinoidă, cu celulele Stem trofoblastice şi cu trofoblastul extravilozitar (Celulele X) în placenta la termen.
1. Epiteliul amniosului. 2. Lama chorială marginită de un strat de substanţa fibrinoidă (Langhans). 3. Vilozitate Stem. 4. Conectarea vilozităţii Stem prin substanţa fibrinoidă cu stratul fibrinoid Langhans din structura lamei choriale. 5. Insulă de trofoblast extravilozitar înconjurată de substanţă fibrinoidă. 6. Celule Stem trofoblastice. 7. Vilozităţi terminale în apoptoză.
Coloraţie Hematoxilină-Eosină, Ob. 7; Ob. 4 (A); 10 (B); 20 (C); 40 (D); X 28 (A); X 70 (B); X 140 (C); X 280 (D).
A
B
C
D
1
2
3 4
4
5
6
7
2

GS Dragoi Tissue Interrelations in the Evolution of Hemochorial Barriers
176
A
B
C
D
E
1
2
2
2
2
2
3
4
Figura nr. 6 Raporturile microanatomotopografice ale elementelor din placa bazală a placentei la termen. 1. Decidua basalis. 2. Substanţa fibrinoidă din stratul Nitabush. 3. Vilozitate intermediară matură. 4. Vilozitate terminală în apoptoză.
Coloraţie Hematoxilină-Eosină, Ob. 7; Ob. 4 (A ); 10 (B, D); 20 (C); 40 (E); X 28 (A); X 70 (B, D); X 140 (C); X 280 (E).
Romanian Journal of Legal Medicine Vol. XV, nr. 3, 2007
177
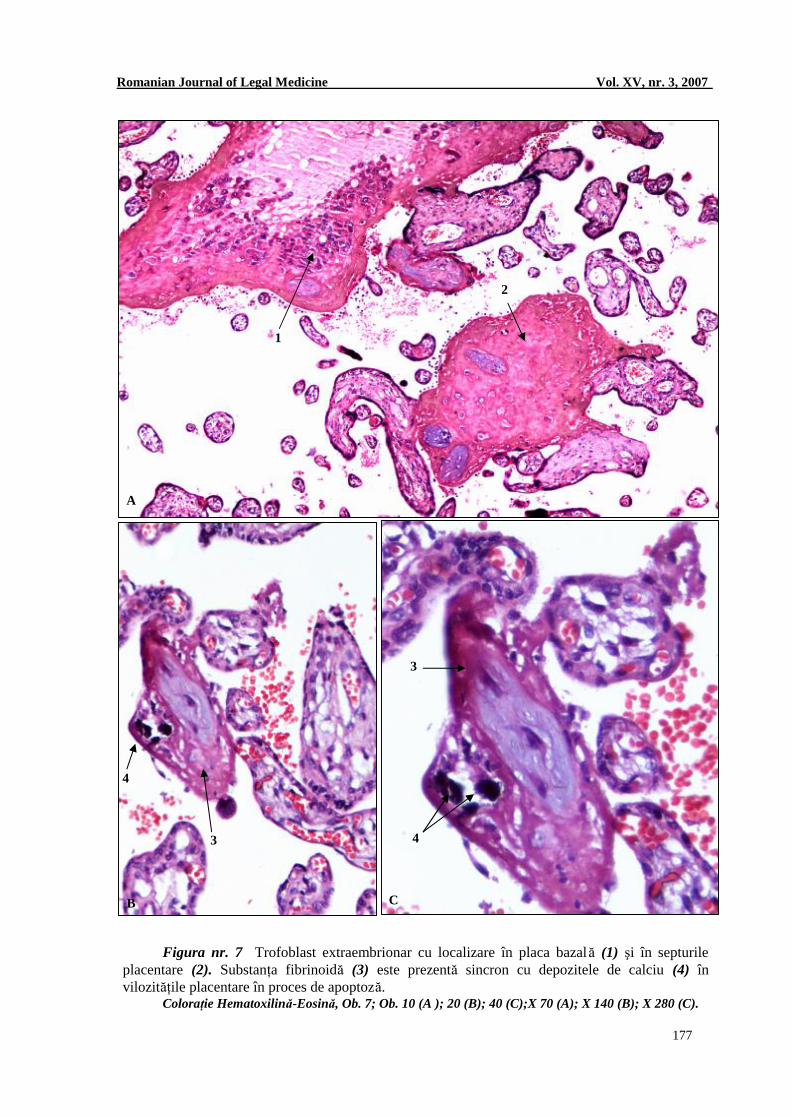
Figura nr. 7 Trofoblast extraembrionar cu localizare în placa bazală (1) şi în septurile placentare (2). Substanţa fibrinoidă (3) este prezentă sincron cu depozitele de calciu (4) în vilozităţile placentare în proces de apoptoză.
Coloraţie Hematoxilină-Eosină, Ob. 7; Ob. 10 (A ); 20 (B); 40 (C);X 70 (A); X 140 (B); X 280 (C).
A
B C
1
2
3
4
4
3

GS Dragoi Tissue Interrelations in the Evolution of Hemochorial Barriers
178
Figura nr. 8 Microanatomia şi histochimia barierei hemo-monochoriale din vilozităţile placentare din al treilea trimestru al gesta ţiei.
Metode de vizualizare: Coloraţia Hematoxilină-Eosină (B; C; D; E); Coloraţia Van Gieson (A); Impregnare argentică după metoda Kosa (I); Impregnare argentică Gömöri (F; G; J) şi Mac Mannus (H). Ob. 7; Ob. 20 (A; D; E; F; H; J); Ob. 40 (B; C; G; I); X 140 (A; D; E; F; H; J); X 280 (B; C; G; I).
A B
C
E
G
I
D
F
H
J

Romanian Journal of Legal Medicine Vol. XV, nr. 3, 2007
179
2. Analiza microanatomică a barierelor hemomonochoriale din al treilea trimestru al gestaţiei
În secţiunile seriate efectuate prin fragmentele recoltate din placentele de al treilea trimestru al gestaţiei, am constatat o heterogenitate structurală prin prezenţa de vilozităţi mezenchimale (~1%), intermediare imature (4%), intermediare mature (25%), Stem (20%) şi terminale (50%) (Figura nr. 4). Vilozităţile terminale au fost identificate cu uşurinţă pe secţiunile seriate colorate cu Hematoxilină-Eosină ca ramuri ale vilozităţilor intermediare mature (Figura nr. 4 B).
Diametrul mediu calculat este de 55 microni cu o amplitudine de variaţie între 38 şi 78 de microni. În stroma laxă se află capilare dilatate cu traiect sinosoidal. Ele determină prin presiunea exercitată asupra trofoblastului, atrofia până la înlocuirea acestuia cu un depozit fibrinoid (Figura nr. 8). Se observă transformarea barierei hemochoriale într-o „membrană vasculofibrinoidă” (Figurile nr. 8A; 8B, 8F).
Examinarea comparativă a imaginilor din secţiunile colorate cu Hematoxilină-Eosină (Figura nr. 3A) şi de impregnare argentică după metoda Kosa permite evaluarea formaţiunilor existente în spaţiul ce a aparţinut sinciţiotrofoblastului.
Pe secţiunile colorate cu Hematoxilină-Eosină apar în acest spaţiu un şirag de structuri ovoidale colorate intens cu Hematoxilină, cunoscută ca un colorant nuclear (Figura nr. 8A). La o primă evaluare, suntem tentaţi de a omologa aceste structuri cu nucleii sincitiotrofoblastului.
Ori, examinarea secţiunilor de impregnare argentică după metoda Kosa, demonstrează prezenţa de conglomerate ovoidale de granule de săruri calcare exact în fosta locaţie a sinciţiotrofoblastului (Figura nr. 8 I) unde am identificat şiragul de structuri hematoxilinopozitive, glicozaminoglicani PAS pozitivi (Figura nr. 8H) şi nu în ultimul rând, depozite fibrinoide (Figura nr. 8B).
În egală măsură, am constatat raporturi de contiguitate între vilozităţile placentare cu realizarea de sinapse trofoblastice (Figurile nr. 4 şi 5), pe de o parte şi existenţa de trofoblast extravilozitar, cu grade variabile de diferenţiere (Figura nr. 7A), pe de altă parte. 3. Analiza distribuţiei spaţiale a substanţei fibrinoide placentare Fibrinoidul a fost identificat perivilozitar, intravilozitar, în zona subiacentă plăcii choriale, pe suprafaţa plăcii bazale, în joncţiunea maternofetală, la nivelul insulelor de celule extravilozitare şi în septurile placentare.
Ne-a atras atenţia în mod deosebit topografia fibrinoidului la nivelul insulelor de celule trofoblastice extravilozitare.
Celulele Langhans prezente în aceste insule suferă un proces de apoptoză şi sunt incluse într-un matrix de tip fibrinoid (Figura nr. 7). Am identificat cu uşurinţă condensări de substanţă fibrinoidă atât la nivelul lamei choriale sub forma stratului fibrinoid Langhans (Figura nr. 5), cât şi la nivelul plăcii bazale sub forma straturilor fibrinoide Rohr şi Nitabush (Figurile nr. 2 şi 6).
Observaţiile noastre coroborate cu datele din literatură ne-au creat convingerea că fibrinoidul placentar nu este o structură patologică, ci face parte din sistemul anatomic al placentei, asigurând adezivitatea uteroplacentară, reglarea circulaţiei intravilozitare, procese de transport maternofetal, protecţie imunologica si nu in ultimul rând o barieră la invazia trofoblastului în structurile uterine.

GS Dragoi Tissue Interrelations in the Evolution of Hemochorial Barriers
180
4. Analiza repartiţiei spaţiale a conglomeratelor de săruri calcare
Am analizat comparativ aceleaşi sectoare din secţiunile colorate prin metodele standard cu cele din secţiunile colorate prin metoda Kosa de vizualizare a sărurilor de calciu. În placentele de prim trimestru de gestaţie am identificat granule de săruri calcare Kosa pozitive, localizate atât sub membrana bazală a citotrofoblastului, precum şi în spaţiile interviloase.
În placentele de ultim trimestru de gestaţie am identificat conglomerate de săruri de calciu pe marginea trofoblastului dar şi în mezenchimul vilozitar cu sau fără depuneri fibrinoide.
În egală măsură, depozitele de săruri calcare sunt prezente şi în insulele de trofoblast intervilozitar. Septurile intervilozitare, placa coriala şi placa bazală conţin în schimb conglomerate comasate de săruri calcare ce conferă structurilor menţionate duritate remarcabilă (Figurile nr. 3 D, 7 B, 7 C, 8 I).
E. Discuţia rezultatelor Structura funcţională a barierelor hemochoriale este incomplet cunoscută. În dinamica
gestaţiei există trei tipuri de locaţii ale schimburilor biunivoce materno-embriofetale: „bariera hemobichorială”, „bariera hemomonochorială” şi „membrana vasculo-fibrinoidă”. În structura „barierei hemo-bichoriale” am remarcat inexistenţa unei graniţe anatomice între citotrofoblast şi sinciţiotrofoblast dar prezenţa de glicozaminoglicani între lama bazală trofoblastică şi suprafaţa sinciţiotrofoblastului precum şi în jurul celulelor Langhans.
În structura „barierei hemo-monochorială”, celulele stratului Langhans dispar prin apoptoză (moarte celulară programată), iar sinciţiotrofoblastul devine atrofic. În alcătuirea „membranei vasculofibrinoide” se diferenţiază în locul sinciţiotrofoblastului un depozit fibrinoid, PAS pozitiv, ce include depozite de săruri calcare („calcinoză placentară”) care pot genera un obstacol în realizarea schimburilor materno-fetale.
Depozitele fibrinoide au fost menţionate în literatură, în special în ariile lipsite de sinciţiotrofoblast. Acesta pare a fi un mecanism de reparare în cadrul etapei de reepitelizare (Nelson, 1990). În stroma adiacentă acestui fenomen, Castelluci (1991) a remarcat prezenţa unei glicoproteine a matrixului extracelular (Tenascina).
Se creează astfel condiţii anatomice pentru perturbarea schimburilor maternofetale şi implicit prin apoptoza structurilor placentare iar în final, ori moartea in utero a fătului, ori la scurt interval de la parturiţie.
Progrese recente s-au înregistrat în cunoşterea transformării fenotipice a trofoblastului. Placenta de Homo Sapiens Sapiens conţine o celulă specializată numită trofoblast care este prima linie diferenţiată în dezvoltarea embrionului şi joacă roluri cheie în nidaţie şi în placentogeneză.
Celula trofoblastică a fost clasificată în citotrofoblast şi sinciţiotrofoblast. În stadiul iniţial al gestaţiei celula Stem trofoblastică orientată către decidua maternă cu care se contopeşte formează syncytium (sinciţiotrofoblastul clasic). În zilele 8 şi 9 de la fecundaţie apar în sinciţiotrofoblast lacune vasculare numite lacune trofoblastice. Stratul de citotrofoblast poate prolifera rapid în aceste spaţii pentru a genera vilozităţile choriale primare. Ulterior, proliferarea citotrofoblastului, creşterea mezenchimului chorionic (de sub stratul de citotrofoblast) şi dezvoltarea vaselor sanguine determină transformarea fenotipică a vilozităţilor primare în vilozităţi secundare care sunt alcătuite din celule mezenchimale

Romanian Journal of Legal Medicine Vol. XV, nr. 3, 2007
181
înconjurate de un strat intern de citotrofoblast şi un strat extern multinucleat de sinciţiotrofoblast (Figurile nr. 3 A , 3 B, 3 C, 3 E, 3 F).
Stratul de sinciţiotrofoblast este incriminat în transportul nutrimentelor, gazelor şi hormonilor - lactogen şi chorionic gonadotrofinic (Benirschke şi Kaufmann, 2001).
Recent, o glicoproteină de membrană numită Syncytin a fost implicată ca mediator al fuziunii celulelor din linia trofoblastică (Mi şi col., 2000). Descoperirea ei ridică vălul necunoaşterii filiaţiei dintre cele două straturi trofoblastice – citotrofoblastul şi sincitiotrofoblastul.
Hibridizarea in situ a demonstrat că syncytin se manifestă specific în stratul de sincitiotrofoblast (Mi şi col., 2000). Aceste studii sugerează că syncytin poate media fuziunea citotrofoblastelor în stratul de sincitiotrofoblast, iar manifestarea syncytin este strâns reglată, în timp şi spaţiu, într-o manieră de menţinere a integrităţii şi funcţiei stratului de sinciţiotrofoblast.
Syncytin are rol în diferenţierea trofoblastului la om şi anume în medierea fuziunii citotrofoblastului şi în sinciţializarea în timpul diferenţierii trofoblastului. Nivelele de syncytin au fost măsurate în placentele normale şi în condiţii variate de gestaţie (Knerr, 2004) a corelat nivelul syncytin mRNA cu vârsta gestaţiei şi greutatea placentei. S-a observat un nivel scăzut al acestui marker la pacientele cu preeclampsie.
F. Concluzii
1. În evoluţia structurilor barierelor hemochoriale intervin elemente ce aparţin fătului
(Trofoblastul) şi mamei (Decidua basalis).
2. Transformarea fenotipică a trofoblastului este dependentă de prezenţa Syncytinei.
3. Fibrinoidul placentar reprezintă o structură histochimică generată de celulele
Langhans şi reprezintă o cauză a derulării temporospaţiale a algoritmului genezei şi evoluţiei
placentei prin procese de regeneză, remodelare, angiogeneză şi transformare fenotipică.
Perturbările în diferenţierea fibrinoidului placentar au ca efecte modificări vasculare, hipoxie
şi implicit suferinţă fetală.
4. Calcinoza placentară apare ca un epifenomen în evoluţia placentei. În etapa
embrionară, placenta conţine granule Kosa pozitive care sunt în limitele ortologiei şi pot fi un
marker pentru tranzitul ionilor de calciu prin bariera hemobichorială. În etapa fetală, a
evoluţiei placentei, am identificat conglomeratele de granule de calciu, în special la nivelul
trofoblastului. Aceasta atrage atentia asupra posibilităţii apariţiei unui obstacol în realizarea
schimburilor gazoase şi nutriţionale prin membranele fetoplacentare.
5. Cunoaşterea limitelor de evoluţie a barierelor hemochoriale reprezintă un factor
primordial în evaluarea structurilor placentare în patologia generală şi medicolegală a gestaţiei
şi avortului.

GS Dragoi Tissue Interrelations in the Evolution of Hemochorial Barriers
182
Bibliografie
1. Ander Z., Bilegan I., Molnar V., Medicina Legală, Ed. Didactică şi Pedagogică, Bucureşti, 1966. 2. Beliş V. (Sub redacţie), Investigaţia microscopică în Medicina Legală. Ed. Academiei Române, Bucureşti,
1993. 3. Beliş V., Îndreptar de practică medicolegală, Ed. Medicală, Bucureşti, 1990. 4. Benirschke K., Kaufmann P., Pathology of the Human Placenta, 4th Ed., pp. 49-56, Springer-Verlag, New
York Inc, New York, 2001. 5. Borke J.L., Calcium pump epitopes placental trophoblast basal plasma membranes. Am. J. Physiol, 257:
C341-C346, 1989. 6. Care A.D., The placental transfer of calcium, J. Dev. Physiol, 15: 253-257, 1991. 7. Castelluci M., Classen-Linke I., Mulhauser J., Kaufmann P., Zardi L., Chiquet-Ehrismann R., The
Human Placenta: A Model for tenascin Expression. Histochemistry 95: 449-458, 1991. 8. Chang C., Chen P.T., Chang G.D., Huang C.J., Chen H., Functional characterization of the Placental
Fusogenic membrane Protein Syncytin, Biology of Reproduction, 71, 2004. 9. Durigon M., Pathologiè médico-legale, Ed. Masson, Paris, 1988. 10. Fisher S.J., Cui T.Y, Zhang L., Grahl K., Guo – Yang Z., Tarpey J., Damsky C.H., Adhesive and
degradative properties of the human placental cytotrophoblast cells in vitro, J. Cell Biol., 109:891-902, 1989.
11. Kernbach M., Medicina judiciară, Ed. Medicală, Bucureşti, 1953. 12. Knerr I., Huppertz B., Weigel C., Dotsch J., Wich C., Schild R.L., Beckmann M.W., Racher W.,
Endogenous retroviral syncytin: compilation of experimental research on syncytin and its possible role in normal and disturbed human placentogenessis. Mol. Hum. Reprod. 2004, 10: 581-588.
13. Knofler M., Vasicek R., Schreiber M., Placenta, 22, Supl. A, S83-S92, 2001. 14. Lacassagne A., Martin E., Precis de Médicine Legale, Ed. Masson et Cie, Paris, 1921. 15. Mi S., Lee X., Li X.P., Veldman G.M., Finnerty H., Racie L., La Vallie E., Tang X.Y., Edouard P., Howes
S., Keith J.C., Mc Cay J.M:, Syncytin is a captive retroviral envelope protein involved in human placental morphogenesis. Nature, 403, 785-789, 2000.
16. Nelson D.M., Crouch E.C., Curran E.M., Farmer D.R., Trofoblast Interaction with Fibrin Matrix. Epittelialization of Perivillous Fibrin Deposits as a Mechanism for Villous Repair in the Human Placenta. Am. J. Pathol. 136: 855-865, 1990.
17. Preda I., Moraru I., Probleme de obstetrică şi ginecologie medicolegală, In Medicina Legală sub redacţia Moraru I., Ed. Medicală, Bucureşti, 1967, p. 644 – 683.
18. Scripcaru G., Astărăstoaie V., Scripcaru C., Medicina Legală, Ed. Didactică şi Pedagogică, Bucureşti, 1993.
19. Scripcaru Gh., Terbancea M., Patologie Medicolegală, Ed. Didactică şi Pedagogică, Bucureşti, 1983. 20. Yu C., Shen K., Lin M., Chen P., Lin C., GCMa Regulates the Syncytin – Mediated Trophoblastic Fusion,
The Journal of Biological Chemistry, vol. 277, No. 51, pp. 50062-50068, 2002.