Embed Size (px)
Citation preview

JOURNAL OF BACTERIOLOGY, May 1982, p. 471 482 Vol. 150, No. 20021-9193/82/050471-12$02.00/0
Interconversion and Uptake of Nucleotides, Nucleosides, andPurine Bases by the Marine Bacterium MB22
MARTINE FORET AND JAN AHLERS*Institut far Biochemie und Molekularbiologie, Fachbereich Biologie, Freie Universitat Berlin, I Berlin 33,
GermanyReceived 17 July 1981/Accepted 12 December 1981
Whole cells and isolated membranes of the marine bacterium MB22 convertednucleotides present in the external medium rapidly into nucleosides and then intobases. Nucleosides and purine bases formed were taken up by distinct transportsystems. We found a high-affinity common transport system for adenine, guanine,and hypoxanthine, with a Km of 40 nM. This system was inhibited noncompeti-tively by purine nucleosides. In addition, two transport systems for nucleosideswere present: one for guanosine with a Km of 0.8 ,uM and another one for inosineand adenosine with a Km of 1.4 ,uM. The nucleoside transport systems exhibitedboth mixed and noncompetitive inhibition by different nucleosides other thanthose translocated; purine and pyrimidine bases had no effect. The transport ofnucleosides and purine bases was inhibited by dinitrophenol or azide, thussuggesting that transport is energy dependent. Inside the cell all of the substrateswere converted mainly into guanosine, xanthine, and uric acid, but also anabolicproducts, such as nucleotides and nucleic acids, could be found.
We recently investigated the kinetic proper-ties of a membrane-bound nucleotidase fromdifferent strains of marine bacteria (1, 17). Toobtain information about the ability of marinebacteria to use the metabolites formed by theaction of this enzyme, we investigated the me-tabolism and transport of nucleosides and purinebases.
Until now such studies have been performedmainly with Escherichia coli (2, 5, 7, 9, 11, 12,18) using either the wild types (2, 11, 18), specialmutants (3, 4, 7-9), or membrane vesicles (5).The results reported have not yet presentedunequivocal proof concerning the mechanism(s)of nucleoside or base transport. Some authorsfavor a group translocation mechanism (5, 18),whereas others implicate an active transportmechanism (3, 4, 8, 9, 11). Munch-Petersen andPihl (8) propose a proton motive force as themain energizer of nucleoside transport. Burton(3) presents evidence for porter systems foradenine, hypoxanthine, and uracil dependent ona proton motive force and facilitated by intracel-lular metabolism of the free bases. Most authorspropose the existence of more than one trans-port system (4, 7, 8, 11). The interpretation ofsuch transport data is often complicated byseveral interconversions of substrates and re-sulting products by enzymes located in the cyto-plasm, at the inner or outer surface of the plasmamembrane, or in the periplasmic space (2, 3, 5,18).The data obtained with E. coli cannot be
readily transfered to other microorganisms. Forinstance, Pickard et al. (13, 14) described twonucleoside deaminases in Micrococcus sodonen-sis. With E. coli, nucleoside metabolism occursmainly by the action of phosphorylases (5).Pickard also found that the entry of nucleosidesinto M. sodonensis was neither saturable, norwas there any competitive inhibition betweenthe nucleosides studied, suggesting the entry byfree diffusion.These results suggest the necessity for a gen-
eral approach to examine the uptake and inter-conversions of nucleotides, nucleosides, andpurine bases by marine bacteria, a group oforganisms exposed to a completely differentenvironment than E. coli. We have alreadyproven that at least one enzyme involved in themetabolism of these substrates, the nucleotid-ase, has quite different properties than the re-spective enzyme of E. coli (1, 17). This enzymeis not located in the periplasmic space, butrather is firmly attached to the membrane.
MATERIALS AND METHODSOrganism. The marine bacterium MB22 was a kind
gift from A. J. De Siervo, University of Maine atOrono.Some of the microbiological characteristics of MB22
were published recently (17).Chemicals. 8-14C-labeled adenine, guanine, hypo-
xanthine, adenosine, guanosine, AMP, and GMP wereobtained from the "Commissariat a l'energie atomi-que" (Gif-sur-Yvette, France), and 'IC-labeled ino-
471
on June 30, 2020 by guesthttp://jb.asm
.org/D
ownloaded from

472 FORET AND AHLERS
sine was from the Radiochemical Centre, Amersham,England.The unlabeled purine and pyrimidine bases were
purchased from Pharma-Waldhof, Dusseldorf, Germa-ny; the purine nucleosides were from Boehringer,Mannheim, Germany. Cytidine and uridine were fromServa, Heidelberg, Germany. Yeast extract was fromDifco, Detroit, Mich. All other reagents were obtainedfrom Merck, Darmstadt, Germany.Membrane filters (pore size 0.45 ,um) were supplied
by Schleicher & Schull, Dassel, Germany, and Poly-gram Cel 400 UV254 thin-layer chromatography sheetswere by Macherey, Nagel and Co., Duren, Germany.
Materials used for autoradiography were KodirexX-ray film (9 by 12 cm), Kodak D 19 developer, andKodak AL 4 X-ray fixer.
Culture conditions. The cells were cultivated at 30°Cin artificial seawater (ASW), pH 7.4 (0.4 M sodiumchloride-0.05 M magnesium sulfate-0.01 M potassiumchloride). To the ASW, 2% peptone from casein,0.005% yeast extract, 0.05% ammonium sulfate, and0.05% Tris were added, and the mixture was shakenon a rotary shaker. Cells were usually harvested in thestationary phase.
Preparation of membranes. The cells were treatedwith lysozyme. After lysis of the spheroplasts byosmotic shock, the plasma membranes were isolatedby centrifugation.
Metabolic conversion studies. Stationary cells (56 mg[wet weight]) were washed twice with ASW and thensuspended in 6.4 ml of ASW. The cell suspension wasincubated for varying periods of time with 1.6 ml of150 FM (25 Ci/mol) purine, nucleoside, or nucleotidesolution, and 1-ml volumes were centrifuged to sepa-rate the medium from the cells. A 10-,ll portion of thesupernatant was applied to polygram Cel 400 UV254thin-layer chromatography sheets against a referencemixture and developed in 1 M ammonium acetate. Thelabeled spots were assayed for radioactivity in aBeckman liquid scintillation counter.A pool extraction of the cells was performed by the
extraction procedure of Nazar et al. (10) and analyzedautoradiographically as described by Seipel and Rei-chert (16).
Kinetic studies. A 0.5-mg amount of cells (wetweight) generally obtained from the stationary phasewas incubated at 25°C for 30 s with 0.03 to 0.2 ,M [8-"C]hypoxanthine, -guanine, -adenine or with 0.5 to 2FM [8-14C]inosine, -guanosine, or -adenosine in 1 mlof ASW and then collected on membrane filter. Thefilters were assayed for radioactivity in a Beckmanscintillation counter. The scintillation cocktail was 100g of naphthalene plus 5 g 2,5-diphenyloxazole per literdioxan.For the study of the different inhibitors, 1 to 10 FM
unlabeled purine and pyrimidine nucleosides or 0.1 to0.5 ILM purine and pyrimidine bases were added to theassay.Measurement of accumulation. A 0.5-mg amount of
cells (wet weight) was incubated at 25°C with 15 ,uM8-14C-labeled nucleoside or with 2 ,M 8-14C-labeledpurine base as described above. Energy poisons (1mM dinitrophenol or 50 mM sodium azide) were addedto the cell suspension 10 min before the labeledcompound or at the time indicated in the figure. Atdifferent times portions were taken and assayed forradioactivity.
Analysis of kinetic data and estimation of error. Thekinetic constants were obtained from double recipro-cal plots and by least-squares fit to the hyperbolicMichaelis-Menten equation (J. Ahlers, A. Arnold, F.R. V. Dohren, and H. W. Peter, Enzymkinetik, 2nded., in press) assuming a constant relative error. Allvalues used for kinetic characterization were averagesof four or more experiments performed in duplicate.
RESULTSInterconversions of nucleotides, nucleosides,
and bases. We incubated the cells with labeledpurine nucleotides, nucleosides, or bases. Atdifferent incubation times, medium and cellswere separated and analyzed by chromatogra-phy and autoradiography.
Analysis of the external medium. Figure 1shows the result of our analysis of the externalmedium, demonstrating the interconversion ofGMP into different metabolites. It can be seenthat GMP immediately disappeared from themedium due to its conversion into guanosine.Guanosine, however, was then converted toguanine and finally to xanthine. The results withother substrates are compiled in Table 1. It canbe seen that all nucleotides were immediatelyconverted to nucleosides. Interestingly, whenAMP was the substrate, an additional deami-nation took place which resulted in the rapidappearance of inosine rather than adenosine. Inaccordance with Fig. 1, all nucleosides were
15
10x
E
5
5 10 15 20 25Time (min)
FIG. 1. Metabolites present in the external mediumafter incubation of marine bacteria with labeled GMPat 25°C. Symbols: 0, GMP; 0, guanosine; O, guanine;U, xanthine. The different metabolites were separatedchromatographically as indicated in the text.
J. BACTERIOL.
on June 30, 2020 by guesthttp://jb.asm
.org/D
ownloaded from

INTERCONVERSION AND UPTAKE OF PURINES 473
TABLE 1. Composition of the external medium'
Substrate Incubation Presence of metabolitesbtime (min) Guanosine Inosine Adenine Guanine Hypoxanthine Xanthine
AMP 1 +++ +++ 010 0 +++ +
GMP 1 +++ ++ ++10 0 +++ +++
Adenosine 1 +++ +++ +10 0 +++ ++
Guanosine 1 + + + + + +10 0 +++ +++
Inosine 1 +++ 010 +++
Adenine 1 + + + + +10 + + ++ ++
Guanine 1 +++ 010 +++++
Hypoxanthine 1 +++ 010 ++ +
aMetabolites found by thin-layer chromatography of the external medium after 1 and 10 min of incubation ofthe marine bacteria at 25°C with labeled nucleotides, nucleosides, and purine bases in ASW (pH 7.4).
b Symbols: 0, only traces; +, small amount; + +, medium amount; +++, large amount.
split to purine bases. When adenosine was usedas substrate, a deamination could again be ob-served. The purine bases were finally convertedto xanthine at a slower rate than the above-mentioned conversion. In the case of adenine,conversion to xanthine was via a deamination tohypoxanthine.
Analysis of the internal pool. Upon examiningthe compounds formed inside the cells afterincubation with several labeled substrates, werecognized a great variety of products (Table 2).After 10 min, only about 25 to 30%o of theradioactivity was found in the insoluble nucleicacid fraction (not shown). Thus, under theseexperimental conditions, the catabolic pathwayseemed to be more important.Table 2 shows that the main products inside
the cells were guanosine, xanthine, and uricacid, which is for the most part independent ofthe substrate offered at the outside. Nearlyevery possible metabolite in the catabolic chainbetween nucleotides and uric acid also ap-peared.Comparison of the products inside and outside
the cell. A comparisoti of Tables 1 and 2 indi-cates that each metabolite found in the mediumalso appeared in the pool extract of the cells.From these results the question arises whetherthe substrates were converted at the outside of
the membrane and then translocated into thecell, or whether the substrates were taken upand the products appeared in the external medi-um after conversion inside the cells. To discrimi-nate between these possibilities, we examinedthe conversions of several substrates by isolatedmembranes. Furthermore, we performed kineticinvestigations on possible transport systems.
Conversions by isolated membranes. Whenstudies on the conversion of several substrateswere performed using isolated membranes rath-er than whole cells, we obtained the resultsshown in Table 3. Isolated membranes were stillable to catalyze both the conversions of nucleo-tides into nucleosides and of nucleosides intopurines. Therefore, both nucleotidase and nu-cleosidase activity must be membrane bound. Adeaminase activity was lacking in these prepara-tions. Possibly, this enzyme had been solubi-lized from the membrane or inactivated by themembrane preparation procedure.
Kinetic investigations. When 8-14C-labeled nu-cleosides or purine bases were added to marinebacteria, a rapid increase in the cell radioactivitycontent was evident. The investigations on met-abolic conversion have shown that the radioac-tivity of the cells represents the sum of theradioactivity incorporated into the various poolsof metabolic products (Tables 1 and 2, Fig. 1).
VOL. 150, 1982
on June 30, 2020 by guesthttp://jb.asm
.org/D
ownloaded from

474 FORET AND AHLERS
+ + + + + + + + + +++ +
+ ++ + ++ ++++ ++ ++ ++ +++ ++ + ++ ++
+ +
+ + + + + +
+ +
+ ++ ++ +
+ +
++ ++ ++++ ++ ++++ ++
+ ++ +
+ + + + + +
+ + ++
- - -S - -
00
o < 0
J. BACTERIOL.
++ ++
+++ +++
+ +
U
4.)
x
4.
4)
0
4)
0
4)
0
4)
04)C)
4)
+ +
0,:
*;.4)i
0
4)~
4)
.-4
0
0
m
F-A
+ +
+ +
-orA
>14
di
~4)
,54)
r3
.o F-0
la4
w, -
e 0
*i;g,D*X
4)o
+ +
+ +
o
I940
99t
994
f
f
I
I
VI
0440
4-
04
1
0400
Pz
on June 30, 2020 by guesthttp://jb.asm
.org/D
ownloaded from

INTERCONVERSION AND UPTAKE OF PURINES 475
TABLE 3. Conversion of nucleotides, nucleosides, and purine bases by isolated membranesaPresence of metabolitesb
SubstrateAdenosine Guanosine Inosine Adenine Guanine Hypoxanthine
AMP + +GMP + +Adenosine + +Guanosine + +Inosine + +Adenine +Guanine +Hypoxanthine +
a The identification of the metabolites present in the medium was performed after incubation of isolatedmembranes of marine bacteria with different nucleotides, nucleosides, and purine bases. The conditions were thesame as in Table 1.
b For explanation of symbols, see Table 1.
To avoid misleading interpretations of the re-sults obtained, we performed kinetic measure-ments under conditions where only a very smallamount of substrate was taken up. Thus steady-state conditions are always maintained and re-verse transport can be neglected. Kinetic inves-tigations with several nucleosides and basesalways resulted in a hyperbolic v versus [S]curve. This indicates a mediated transport ratherthan free diffusion of the compounds.
Effect of the growth phase on transport. My-gind and Munch-Petersen (9) reported that mea-surements on transport of pyrimidine nucleo-sides by E. coli gave very unreproducible resultswhen cells were harvested in the stationaryrather than in the logarithmic phase. To achieveoptimal and reproducible conditions for the ki-netic measurements of transport by marine bac-teria, we performed purine and nucleoside varia-tions using cells harvested at various timesduring growth. We found that the growth phasedid not influence Km or VmX. for purine trans-port, nor the Km of nucleoside uptake. However,Vmax for nucleoside transport was twofold high-er when the cells were harvested in the station-ary rather than in the logarithmic phase (notshown). Therefore, all kinetic measurementswere performed using stationary cells.Nudeoside transport. We measured the uptake
of labeled adenosine, guanosine, and inosineunder steady-state conditions. 8-14C-labeled nu-cleosides were used rather than U-14C com-
pounds to avoid interference by the uptake ofribose possibly produced by an external nucleo-sidase.Table 4 shows that the Vma values for the
nucleosides were similar, whereas the Km forguanosine was considerably lower than the re-
spective constants for adenosine and inosine.For further characterization of this transportsystem, we investigated the influence of unla-
beled nucleosides as well as purine and pyrimi-dine bases on the uptake of nucleosides.
Effect of unlabeled nudeosides. To determinewhether the results shown in Table 4 representone or more transport systems and to evaluatethe specificity of the nucleoside transport sys-tem, we performed kinetic experiments usinglabeled inosine in the presence of several con-centrations of unlabeled adenosine (Fig. 2) orguanosine (Fig. 3). If the purine nucleosideswere transported by the same carrier, we wouldhave expected a competitive inhibition, with Kivalues representing the affinity of the inhibitorsvaried to that transport system. Such a competi-tive inhibition can be seen in Fig. 2, in which theeffect of various concentrations of adenosine oninosine uptake is shown. From the secondaryplot (Fig. 2, inset), we obtained a Ki of 1.4 ,uM(Table 5), which is identical, within the limits oferror, with the Km of 1.5 ,uM for adenosine(Table 4).
In contrast to adenosine, the presence ofguanosine led to a noncompetitive inhibition ofinosine transport (Fig. 3). Therefore, guanosinemust be bound by a different site of the translo-cation system other than the binding site for
TABLE 4. Kinetic constants of the nucleosidetransport systema
Vm.Substrate Km (FLM) (nmol/minper mg of
cells)
Adenosine 1.5 ± 0.4 1.5 + 0.3Inosine 1.4 ± 0.4 1.2 + 0.3Guanosine 0.7 ± 0.2 1.3 ± 0.3
a The data were obtained from reciprocal plots aftermeasurement of the uptake of labeled nucleosidesincubated for 30 s at 25°C with the marine bacteria.
VOL. 150, 1982
on June 30, 2020 by guesthttp://jb.asm
.org/D
ownloaded from

476 FORET AND AHLERS
1:
v}I
U)
o1
_o ?IE
ccy
8
6
3-
) El.
2
0.5 1.0 1.5linosiner1 (FM)1
FIG. 2. 1/v versus 1/[inosine], plotted for various concentrations of adenosine. Experiments were performedat 25°C in ASW (pH 7.4) as described in the text. Symbols: 0, control; 0, 2 ,M adenosine; *, 5 FM adenosine.(Inset) Secondary plot of the apparent Km values obtained from the Lineweaver-Burk plot versus [adenosine].
9
5(Uo EC .
.C
2 4 6 8 1019"nOSine (PM )
1.5 2.0f inosineIJ (PM )
FIG. 3. 1/v versus 1/[inosine], plotted for various concentrations of guanosine. Symbols: V, control; 0, 1 .LMguanosine; 0, 2 I.M guanosine; U, 5 ,uM guanosine; O, 10 ,uM guanosine. The experiments were done under thesame conditions as described in Fig. 2. (Inset) Secondary plot of l/V,,,, versus [guanosine]. The values weretaken from the intercepts of the straight lines of Fig. 3 with the ordinate.
J. BACTERIOL.
on June 30, 2020 by guesthttp://jb.asm
.org/D
ownloaded from

INTERCONVERSION AND UPTAKE OF PURINES 477
TABLE 5. Inhibition of nucleoside transport'Labeled Unlabeled Typeoni Ksubstrate inhibitor Type of ihibitLon(>M)
Inosine Inosine Competitive 1.4Adenosine Competitive 1.4Guanosine Noncompetitive 3.0Cytidine Mixed 5.5Uridine Mixed 6.0
Adenosine Inosine Competitive 1.5Guanosine Noncompetitive 2.5
Guanosine Guanosine Competitive 0.8Inosine Noncompetitive 2.0Adenosine Noncompetitive 1.7Cytidine Mixed 5.0Uridine Mixed 3.0
a The type of inhibition was determined from reciprocal plots (e.g., Fig. 2 to 5). The inhibition constants werederived from the secondary plots (e.g., insets in Fig. 2, 3, and 5). The conditions were the same as in Table 4.
inosine or adenosine. A Ki value of 3 ,Mresulted from the secondary plot (Fig. 3, inset);this was considerably higher than the respectiveKm value (0.7 FM, Table 4). Besides adenosineand inosine, guanosine could also be taken upwith a similar V,,, and with an even higheraffinity (Table 4). The differences in Km and Kiofguanosine in connection with the noncompeti-tive inhibition of guanosine on inosine transport
indicated the presence of another nucleosidetransport system.
Figure 4 shows the effects of several unlabelednucleosides on the uptake of labeled guanosine.The presence of unlabeled guanosine leads to acompetitive inhibition since unlabeled substratecompetes with labeled substrate for the translo-cation site. However, inhibition studies usinginosine as well as adenosine revealed a noncom-
5
41
U)
c c
[guanosinel- (1.iM1lFIG. 4. 1/v versus 1/[guanosine], plotted for 1 FM adenosine, inosine, or guanosine. Symbols: 0, control; *,
adenosine; 0, inosine; V, guanosine. The conditions were the same as in Fig. 2.
VOL. 150, 1982
on June 30, 2020 by guesthttp://jb.asm
.org/D
ownloaded from
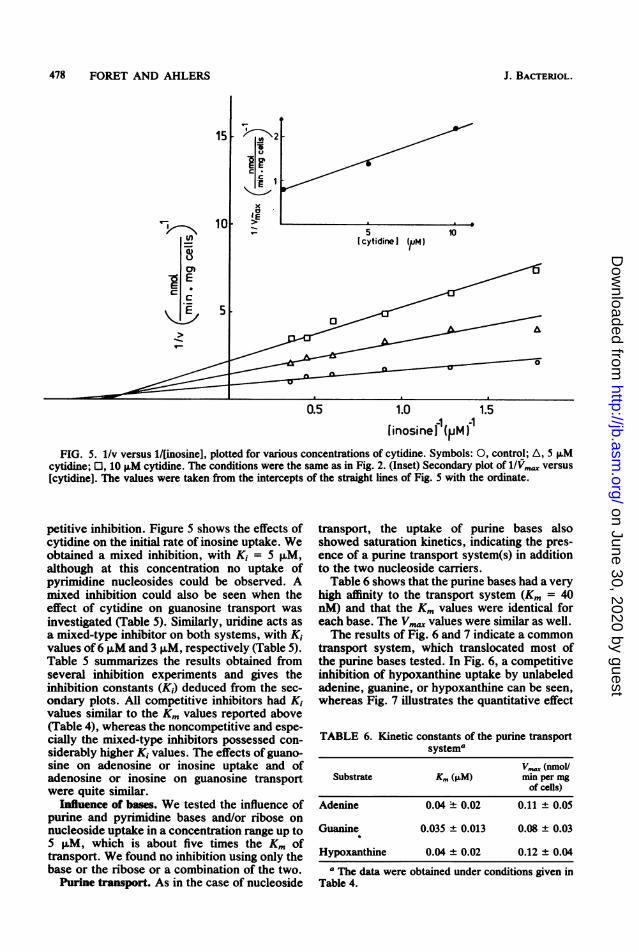
478 FORET AND AHLERS
tn
8c"
XEc c-
2
ic
I
1
xa
-f5
[cytidine I (PM)10
0
0.5 1.0 1.5-1 -1
[inosinei (PM)FIG. 5. 1/v versus 1/[inosine], plotted for various concentrations of cytidine. Symbols: O, control; A, 5 ,uM
cytidine; 0, 10 FM cytidine. The conditions were the same as in Fig. 2. (Inset) Secondary plot of 1/Vmax versus[cytidine]. The values were taken from the intercepts of the straight lines of Fig. 5 with the ordinate.
petitive inhibition. Figure 5 shows the effects ofcytidine on the initial rate of inosine uptake. Weobtained a mixed inhibition, with Ki = 5 ,M,although at this concentration no uptake ofpyrimidine nucleosides could be observed. Amixed inhibition could also be seen when theeffect of cytidine on guanosine transport wasinvestigated (Table 5). Similarly, uridine acts asa mixed-type inhibitor on both systems, with K,values of 6 ,uM and 3 p,M, respectively (Table 5).Table 5 summarizes the results obtained fromseveral inhibition experiments and gives theinhibition constants (K,) deduced from the sec-ondary plots. All competitive inhibitors had K1values similar to the Km values reported above(Table 4), whereas the noncompetitive and espe-cially the mixed-type inhibitors possessed con-siderably higher Ki values. The effects of guano-sine on adenosine or inosine uptake and ofadenosine or inosine on guanosine transportwere quite similar.
Influence of bases. We tested the influence ofpurine and pyrimidine bases and/or ribose onnucleoside uptake in a concentration range up to5 p,M, which is about five times the Km oftransport. We found no inhibition using only thebase or the ribose or a combination of the two.
Purine trnport. As in the case of nucleoside
transport, the uptake of purine bases alsoshowed saturation kinetics, indicating the pres-ence of a purine transport system(s) in additionto the two nucleoside carriers.
Table 6 shows that the purine bases had a veryhigh affinity to the transport system (Km = 40nM) and that the Km values were identical foreach base. The Vm., values were similar as well.The results of Fig. 6 and 7 indicate a common
transport system, which translocated most ofthe purine bases tested. In Fig. 6, a competitiveinhibition of hypoxanthine uptake by unlabeledadenine, guanine, or hypoxanthine can be seen,whereas Fig. 7 illustrates the quantitative effect
TABLE 6. Kinetic constants of the purine transportsystema
V.. (nmol/Substrate Km (>M) min per mg
of cells)Adenine 0.04 4 0.02 0.11 + 0.05
Guanine 0.035 + 0.013 0.08 + 0.03
Hypoxanthine 0.04 + 0.02 0.12 ± 0.04a The data were obtained under conditions given in
Table 4.
J. BACTERIOL.
on June 30, 2020 by guesthttp://jb.asm
.org/D
ownloaded from

INTERCONVERSION AND UPTAKE OF PURINES 479
* ' 30
10 20 1 1 301 hypoxonthine f (pjM)
FIG. 6. 1/v versus 1/[hypoxanthinel, plotted fordifferent concentrations of adenine, guanine, or hypo-xanthine. Symbols: *, control; 0, 0.1 F.M adenine; *,0.15 ,uM guanine; V, 0.2 ,uM hypoxanthine. Theconditions were the same as in Fig. 2.
of adenine on guanine uptake. From the second-ary plot, a !, value can be deduced, which isidentical with the respective Km value of Table6. Xanthine bases, pyrimidine bases, and pyrim-idine nucleosides had no inhibitory effect on thispurine base transport system. However, purinenucleosides led to a strong noncompetitive inhi-bition of purine base transport (Fig. 8) with aninhibition constant of Xi = 60 nM; i.e., they hada greater affinity than the noncompetitive effec-tors for the nucleoside transpdrt systems.Dependence of nudeoside and purine base
transport on energy. The accumuiation of gua-nine (Fig. 9A) and guanosine (Fig. 9B) wasstrongly inhibited by the addition of 1 mMdinitrophenol or 50 mM sodium azide. Inhibitionwas detected regardless of whether the inhibi-tors were added before the addition of the radio-actively labeled compounds or during the courseof accumulation. Similar effects were obtainedwith both energy poisons on all purine bases andnucleosides.
DISCUSSION
In most microorganisms, purine and pyrimi-dine nucleotides may be synthesized from sim-pler metabolites via the de novo pathways, orfrom preformed bases and nucleosides, whenthese precursors are available. Several marinebacteria possess very high nucleotidase activity(1). This enzyme, which has access to substratesin the external medium (1), may be responsiblefor converting external nucleotides into metabo-lites. These can then be transported into thecells and thus can be used, via the salvagepathway, to build up internal nucleotides andnucleic acids.
In contrast to the results of Pickard (13) fornucleoside transport by M. sodonensis, our ki-netic investigations supply evidence for the ab-sence of free diffusion and for the presence of
10[guanine]-1 (pMr1
15
FIG. 7. 1/v versus 1/[guanine], plotted for variousconcentrations of adenine. Symbols: *, control; 0,0.15 ,uM adenine; 0, 0.3 ,uM adenine; 0, 0.5 ,uMadenine. The conditions of experiments were the sameas described in Fig. 2. (Inset) Secondary plot of theapparent Km values obtained from the Lineweaver-Burk plot versus [adenine].
VOL. 150, 1982
on June 30, 2020 by guesthttp://jb.asm
.org/D
ownloaded from

480 FORET AND AHLERS
70
60
IQ
It.)CC
E>
10 20 30 1 -140[hypoxonthinel (P.M)
FIG. 8. 1/v versus 1/[hypoxanthine], plotted for 0.1 ,uM guanosine, adenosine, or inosine. Symbols: V,control; 0, guanosine; 0, adenosine; *, inosine. The conditions were the same as in Fig. 2.
several transport systems involved in the uptakeof purine bases and nucleosides.The catabolism of the different substrates may
be a source of error in the interpretation of theresults of nucleoside uptake. However, sinceinitial rate data were measured, we could ex-clude a significant conversion of nucleosides.Therefore the kinetic constants obtained weremost likely due to nucleoside transport.
Furthermore, unlabeled purines had no effecton these transport processes. The competitiveinhibition of inosine uptake by unlabeled adeno-sine and the noncompetitive inhibition by guano-sine and vice versa indicated the presence ofdifferent transport systems for nucleosides: acommon one for inosine and adenosine andanother one for guanosine. Similar results were
reported by Munch-Petersen and Pihl (8), whofound a special transport system for guanosinein addition to the system available for all othernucleosides. Pyrimidine nucleosides at a con-
centration at which no measurable uptake couldbe observed led to a mixed inhibition of bothtransport processes, indicating the presence ofeffector sites in addition to the translocationsites of both carriers.
Interestingly, the kinetic constants of inhibi-
tion were well above the Km values for uptake ofthese compounds. Thus a regulation of transportoccurred when there was an abundance of othernucleosides other than that which was to betransported.The effector sites in both transport systems
must be relatively specific, since neither purinebases, nor ribose, nor both compounds togethereffect the transport. Such effector sites, althoughquite abundajnt in enzymes, have not been fre-quently reported for purine base or nucleosidetransport by bacteria. However, Peterson et al.(11), in investigating the nucleoside transport inE. coli, gave evidence that the sites for interac-tion of adenosine and uridine, on the cytidinetransport system, are not involved in the trans-port of the inhibiting nucleosides.
Considering the uptake of bases, we foundthat the marine bacteria possessed a commontransport system with very high affinity foradenine, guanine, and hypoxanthine (Table 6),whereas pyrimidine bases and xanthine were nottransported under these conditions. The kineticconstants were identical for the three substrates.The Km was only about 1/40 of the Km fornucleosides, whereas the Vml, was an order ofmagnitude lower. The Km values for the uptake
J. BACTERIOL.
on June 30, 2020 by guesthttp://jb.asm
.org/D
ownloaded from

INTERCONVERSION AND UPTAKE OF PURINES 481
- 0.3
--0.20EC
a 0.1._CE
dl
Os9.980.6C
C
c 0.3 /^
10 20 30 40 50Time (min)
FIG. 9. Accumulation of purine base and nucleo-side in MB22. The accumulation was measured asdescribed in the text. Either 2 FM [8-14C]guanine (A)or 15 FLM [8-14C]guanosine (B) was added at time zero.1 mM dinitrophenol (A) or 50 mM azide (B) was addedeither 10 min before addition of the substrate to thecell suspension or at the time indicated in the figure.Symbols: 0, [8-14C]guanine (control); 0, +1 mMdinitrophenol; +, plus preincubation with 1 mM dini-tropherol; *, [8-14C]guanosine (control); E, +50 mMazide; A, plus preincubation with 50 mM azide.
of purine bases were much lower than thosereported by Hochstadt-Ozer and Stadtman (6)for E. coli, but only slightly below those valuesgiven by Roy-Burman and Visser (15) for E. coli.Whereas pyrimidine bases and pyrimidine nucle-osides had no effect on the uptake of purinebases, there was a noncompetitive inhibition byguanosine, adenosine, or inosine (Fig. 9). There-
fore, we suppose that there was an effector siteat the purine base transport system similar tothat discussed above for the nucleoside translo-cation systems.Thus it is obvious that there are manifold
possibilities for regulation of the utilization ofthe catabolic products of nucleic acids from theenvironment. We found that, after incubation ofmarine bacteria with nucleotides, several nucle-osides and bases appeared very rapidly in theexternal medium. They could be incorporatedinto the cell for use in the salvage pathway bytwo nucleoside transport systems, whose activi-ties were determined by the type and concentra-tion of other nucleosides present, or by a high-affinity purine base transport system which waseffected by nucleosides as well. Inside the cellthe translocated substrates were converted tonucleotides and nucleic acids or were catabo-lized.We assumed that the inhibition of nucleoside
and base transport by higher concentrations ofother nucleosides represented a mechanism tostop uptake under conditions in which the cellhas an abundance of these substances. Thus itwas unnecessary for the cell to take them up.On the other hand, when there were very low
concentrations of nucleosides present in theexternal medium, the uptake of purine bases bytheir high-affinity transport system may havebecome important. Thus these mechanisms pro-vided the cells with the ability to use externalsubstrates under unfavorable conditions.From Fig. 9 it can be calculated that the
marine bacterium MB22 was able to accumulatepurine bases as well as nucleosides from theenvironment. We found a 200-fold increase inradioactivity when purine bases were appliedand a 60- to 90-fold increase in the case ofnucleosides. However, because of several con-versions of the transported compounds insidethe cell (Table 2), the exact ratios could not bedetermined. Yet data from energy poisons suchas dinitrophenol and azide show the dependenceof the transport on energy. Similar results werereported by Munch-Petersen and Pihl (8) as wellas by Mygind and Munch-Petersen (9) for nucle-oside transport in cells of E. coli.
ACKNOWLEDGMENT
We thank U. Reichert and R. Schmidt for valuable discus-sion. This work was supported by a research grant of theDeutsche Forschungsgemeinschaft.
LITERATURE CITED
1. Ahlers, J., T. Guinther, and H. W. Peter. 1978. Phospho-lipid composition of plasma membranes and kinetic prop-erties of membrane-bound nucleotidase from marine bac-teria. Int. J. Biochem. 9:573-578.
2. Bagnara, A. S., and L. R. Finch. 1974. The effects of bases
VOL. 150, 1982
on June 30, 2020 by guesthttp://jb.asm
.org/D
ownloaded from

482 FORET AND AHLERS
and nucleosides on the intracellular contents of nucleo-tides and 5-phosphoribosyl-1-pyrophosphate in Esche-richia coli. Eur. J. Biochem. 41:421-430.
3. Burton, K. 1977. Transport of adenine, hypoxanthine anduracil into Escherichia coli: Biochem. J. 168:195-204.
4. Dodskl, J. 1972. The components of the nucleoside-transporting system in Escherichia coli. Biochim.Biophys. Acta 282:393-400.
5. HMqcstadt4Ozer, J. 1972. The regulation of purine utiliza-tion in bacteria. J. Biol. Chem. 247:2419-2426.
6. Hochsadtdzer, J., and E. R. Stadtman. 1971. The regula-tion of purine utilization in bacteria. J. Biol. Chem.246:5294-5303.
7. Komatsu, Y., and K. Tanaka. 1972. A showdomycin-resistant mutant of Escherichia coli K-12 with alterednucleoside transport character. Biochim. Biophys. Acta288:390-403.
8. Munch-Petese, A., and N. J. PIh. 1980. Stimulatoryeffect of low ATP pools on transport of purine nucleosidesin cells of Escherichia coli. Proc. Natl. Acad. Sci. U.S.A.77:2519-2523.
9. Myglnd, B., and A. Munch-Petersen. 1975. Transport ofpyrimidine nucleosides in cells of Escherichia coli K 12.Eur. J. Biochem. 59:365-372.
10. Nazr, R. N., H. G. Lawford, and J. Tze-Fel Wong. 1970.An improved procedure for extraction and analysis ofcellular nucleotides. J. Anal. Biochem. 35:305-313.
11. Peterson, R. N., J. Bonface, and A. L. Koch. 1967. Energyrequirements, interactions and distinctions in the mecha-nisms for transport of various nucleosides in Escherichiacoli. Biochim. Biophys. Acta 135:771-783.
12. Peteno, R. N., and A. L. Koch. 1966. The relationship ofadenosine and inosine transport in Escherichia coli. Bio-chim. Biophys. Acta 126:129-145.
13. PHkard, M. A. 1980. Passive diffusion of nucleosides intoMicrococcus sodonensis membrane vesicles. Can. J. Bio-chem. 58:457-460.
14. Pckard, M. A., L. Philllppe, and J. N. Campbell. 1974.Metabolism and transport of purine nucleosides by mem-brane preparations of Micrococcus sodonensis. Can. J.Biochem. S2:83-89.
15. Roy-Burman, S., and D. W. Visser. 1975. Transport ofpurines and deoxyadenosine in Escherichia coli. J. Biol.Chem. 250:9270-9275.
16. Sedpel, S., and U. Redchert. 1977. Two-dimensional thinlayer chromatography-autoradiography of intracellularpurine interconversion products. J. Chromatogr. 135:485-488.
17. Sbnonls, M., S. Rade, and J. Ahlen. 1981. Kinetic charac-terization of a membrane-bound nucleotidase from themarine bacterium MB22. Int. J. Biochem. 13:733-738.
18. Yagil, E., and I. R. Beacham. 1975. Uptake of adenosineS'-monophosphate by Escherichia coli. J. Bacteriol.121:401-405.
J. BACTERIOL.
on June 30, 2020 by guesthttp://jb.asm
.org/D
ownloaded from





























