Embed Size (px)
Citation preview



Innova Biotec es una publicación semestral del Instituto de Innovación en Biotecnología e Industria.
Directora Ejecutiva del IIBI: Dra. Bernarda Castillo
Director de la Revista: Dr. Andrés Guerrero.
Consejo Editorial:Dr. Diógenes Aybar, Academia de Ciencias de la República Dominicana / MESCYTDr. Luis Wong Vega.Dra. Bernarda Castillo, IIBIIng. Hector Rosario, IIBI
Correctores:Blasina Fabián Adames, MscLic. Benita Castillo
Diseño y Diagramación:Lic. Wilfredo M. Tavárez Q.Lic. Fausto Fray
Dirección:Instituto de Innovación en Biotecnología e IndustriaAve. Núñez de Cáceres, esq. Oloff PalmeSan Gerónimo, Santo Domingo, D.N.República Dominicana
Teléfonos: (809) 566 8121, Ext.. 2221 o 2222Fax: (809) 227 8810Email: [email protected], Vol. 1, ISSN 2226-2490
Junio 2012

Innova BiotecNo.1, Vol. 1, ISSN 2226-2490

5
Indice.
- Presentación de Innova Biotec, por Bernarda Castillo, Directora Ejecutiva del IIBI...........................................................................................5
Trabajos científicos
- Caracterización molecular del mango banilejo (Mangifera indica L.) por medio de análisis AFLP, por José R. Núñez, Ineko Hodai, Guarina Del Monte y Atharva Veda Rosa......................................................................6
- Respuesta a cinco tratamientos de asepsia en distintos explantes de guanábana (Annona muricata L.), por Adriana Chavarría Lines, Yaneury Doñé Dipré y Luis Wong-Vega......................................................................14
- Harina de Nopal (Opuntia ficus) aplicada en la Elaboración de Productos de Panificación, por Oscar Merette, Elisa Goméz, Carlos Calderón y Lucía Beriguete..............................................................................18
- Bioprospección de plantas endémicas dominicanas: Aislamiento e identificación de compuestos con posible acción anticancer presentes en Agave antillarum, Agave brevipetala y Agave brevispina, por Susana De Jesús, S. Bhave, S. Clinton, K. Riedl y S. Schwartz.....................................................................................................28
- Análisis de la variabilidad genética de dos especies de pecaríes (Género Tayassu) en Colombia, Bolivia, Ecuador y Perú utilizando marcadores microsatélites, por Carlos Fernando Vergara y Manuel Ruiz García....................................................................38
- Instrucciones para Publicación....................................................................................59

66
Presentación por Dra. Bernarda Castillo, Directora Ejecutiva.
El IIBI, institución estatal creada mediante decreto no. 58-05 del poder ejecutivo, tiene como misión conducir investigación científica, asi como transferencia e innovación tecnológica enfocada a la biotecnología en diversas vertientes (vegetal, industrial, biomedica, medio ambiental) para contribuir a mejorar la competitividad de la Nación Dominicana.
En su corta historia, el IIBI ha logrado establecerse como institución de vanguardia que busca la excelencia en su quehacer cotidiano. En 2007 fuimos certificados bajo la norma ISO 9001 (versión 2001) y en 2010 continuamos re-certificados con la versión 2008 de la norma ISO 9001. El año 2009 fue testigo de un hito trascendental en la historia de nuestro país, al lograr nuestros laboratorios la acreditación ISO/IEC 17025:2005 para 4 ensayos y en 2010 incrementamos los ensayos acreditados de 4 a 16, siendo la primera institución en República Dominicana en lograr este objetivo que contribuye a garantizar la calidad de los productos industriales dominicanos dirigidos tanto al mercado nativo como a la exportación; nuestros laboratorios además de servir de apoyo a la investigación, proveen valiosos servicios a la industria nacional.
En lo que se refiere a investigación, centro de nuestra misión, desde el 2005, año en que inician sus operaciones tanto el IIBI como el FONDOCYT (fondo concursable del Ministerio de Educación Superior, Ciencia y Tecnología - MESCYT), cada año hemos accedido a financiamiento para proyectos de investigación. Además del financiamiento provisto por FONDOCYT, nuestra cartera de proyectos (más de 30, concluidos o en ejecución) ha recibido el apoyo de otros fondos como el FONIAF, FIES y de organismos internacionales de cooperación. Las áreas de investigación son muy diversas pues abarcan disciplinas relacionadas con la agricultura, medio ambiente, seguridad alimentaria, biomedicina, productos nutricionales, cuidado personal y tecnología de alimentos.
La introducción precedente es necesaria para dar a conocer el contexto en que se produce el lanzamiento de nuestra revista científica INNOVA BIOTEC que representa un punto notable en la consolidación del accionar científico del IIBI.
En este primer número presentamos a la comunidad científica y académica cinco artículos científicos que revelan la diversidad de resultados trabajados por nuestro personal investigador que van desde una innovadora propuesta de harina de nopal (Opuntia ficus L.) hasta la caracterización molecular del mango banilejo (Mangifera indica L.), producto emblemático de nuestra agricultura con acceso a importantes nichos de exportación. Este primer número incluye además, una pionera investigación en establecimiento in vitro de guanábana (Annona muricata L.), novedosos hallazgos en posibles propiedades anticancerígenas de especies endémicas de Agaves (antillarum, brevipetala y brevispina) y un analisis genético poblacional de dos especies de pecaríes (género Tayassu) que habitan en la región amazónica de varios paises sudamericanos.
Esperamos que esta contribución de nuestros investigadores sea acogida con beneplácito por la comunidad científica y académica, tanto en la República Dominicana como en el Caribe, en Latinoamerica y en el Mundo.

7
Caracterización molecular del mango banilejo (Mangifera indica L.) por medio de análisis AFLPJosé R. Núñez, Ineko Hodai, Guarina Del Monte y Atharva Veda Rosa. Instituto de Innovación en Biotecnología e Industria (IIBI). Centro de Biotecnología Vegetal (CEBIVE). E-mail: [email protected]
Palabras Clave: Mango, caracteres morfológicos, marcadores moleculares, AFLP.
Resumen
Se realizó una caracterización molecular y morfológica de mangos banilejos (Mangifera indica L.), usando el método de análisis AFLP y características agro- morfológicas organolépticas con el objetivo final de uniformizar la variedad. La recolección del material vegetal se hizo en tres huertos comerciales propiedad de miembros del clúster de mango de la República Dominicana localizados en la provincia Peravia. Los análisis fueron conducidos en los laboratorios del Instituto de Innovación en Biotecnología e Industria (IIBI) localizados en La Isabela, Santo Domingo Oeste siguiendo protocolos estándares establecidos para estos procedimientos. Para la caracterización morfológica se tomaron los parámetros: Diámetro del fruto (DF), Longitud del fruto (LF), Peso del fruto (PF), Diámetro de la semilla (DS), Longitud de la semilla (LS), Peso de la semilla (PS), Peso de la cáscara (PC), Peso del mesocarpio (PM) y Grado brix (GB), como % de sólidos solubles. Para los marcadores moleculares de ADN se usó la técnica de Polimorfismo de Longitud de Fragmento Amplificado (Amplified Fragment Length Polymorphism) AFLP, por sus siglas en inglés usando un kit de Invitrogen™ prediseñado, el cual incluye su propio protocolo de trabajo. El análisis de conglomerado de AFLP determinó la formación de cinco grupos entre las 38 muestras analizadas demostrando la gran diferencia entre los mangos banilejos cultivados en la zona de Baní, República Dominicana. En base a los datos obtenidos se seleccionaron cuatro ejemplares para realizarle su perfil genético y recomendarlos como patrones para la obtención de yemas en los programas de multiplicación de la variedad.
Key words: Mango, morphological characters, molecular markers, AFLP.
Abstract
A molecular and morphological characterization was performed on mangoes (Mangifera indica L.) of the banilejo type, using AFLP analysis and agro-morphological and flavor characteristics with the final objective of standardizing the variety. Plant material collection was done in three different commercial orchards owned by members of the Dominican Republic mango cluster. These orchards are located in the Peravia Province, 60 kilometers south of the city of Santo Domingo. The analyses were conducted in the laboratories of the Instituto de Innovación en Biotecnología e Industria (IIBI) locate at La Isabela, following standard protocols established for these procedures. For the morphological characterization the following parameters were considered: Fruit diameter (DF), Fruit length (LF), Fruit weight (PF), Seed length (LS), Seed weight (PS), Peel weight (PC), Pulp weight (PM) and Brix grade (GB), as % of soluble solids. For the DNA molecular markers the technique of Amplified Fragment Length Polymorphism (AFLP) was used using a pre-designed kit from Invitrogen™ which

88
includes its own working protocol. The conglomerate analysis of AFLP determined the formation of five groups within the 38 samples analyzed showing a great variability between these types of mangoes grown around the Baní area of the Dominican Republic. Based on the data obtained, four individual trees were selected to perform a genetic fingerprinting and to recommend them as source buds for plant propagation of the variety.
Introducción
El mango es nativo del sur de Asia especialmente Birmania e India Oriental. En el sureste asiático se conocen 41 especies afines de las cuales pudo haberse obtenido el mango actual por selección y cruzamiento. La India ha sido el centro de selección principal de variedades donde se cultiva esta fruta desde hace cientos de años y de donde se difundió al resto de los trópicos a partir de 1700 cuando los Portugueses lo llevaron a Brasil. Fue introducido a la isla Española por los franceses al final del siglo 18 (Geilfus, 1989).
Existen dos razas de mango, una de la India y la otra de las Filipinas y el sureste asiático. La raza india es intolerante a la humedad, el crecimiento nuevo es brillante y rojizo el cual es susceptible al mildiu y el fruto es monoembriónico de mucho color y forma regular. La raza Filipina tolera el exceso de humedad, el crecimiento nuevo es de color verde pálido o rojizo y resiste al mildiu. Su fruto poliembriónico es verde pálido y de forma alargada y arriñonada (Sampson, JA.1986).
Los marcadores de ADN han permitido la caracterización de genotipos, independiente de las influencias de las condiciones de crecimiento ambientales, de la edad fisiológica de la planta y del tipo de tejido que se analice (Vainstein y Ben-Meir, 1994). El análisis de polimorfismo de longitud de fragmento amplificado (AFLP) por sus siglas en inglés, es un método eficiente de huella digital de ADN basado en una amplificación selectiva usando PCR (reacción en cadena de la polimerasa) de fragmentos de restricción de una digesta total del ADN genómico. La técnica del AFLP fue desarrollada básicamente para revelar las diferencias entre cultivares de especies vegetales y ha sido aplicado en varios cultivos (Corsini y col. 1999).
En la actualidad todos los cultivares de mango han resultado de selecciones al azar o por mejoramiento clásico (Lavi y col. 2004). Lavi y col. (2004) aplicaron AFLP para estudiar la relación genética entre 16 cultivares de patrones de mango y para la construcción de un mapa genético de estos. Los autores encontraron que el promedio de bandas compartidas es de 81% (Lavi y col. 2004). Kashkush y col. (2001) encontraron que seis combinaciones de cebadores (primers) de AFLP produjeron 204 bandas claras y en promedio 34 bandas para cada combinación de 16 cultivares y 7 patrones de mango.
Materiales y Metodos Material vegetal:
Las muestras biológicas se obtuvieron en las fincas del Dr. Mois Haché, del señor, Amable Díaz y del señor Angel Peguero, los tres pertenecientes al Clúster de Mango de la República Dominicana. Estas fincas están localizadas en la Provincia Peravia, distante unos 60 km al suroeste de Santo Domingo.
Las muestras se tomaron en hora de la mañana y luego de colocarse en bolsas plásticas se identificaron apropiadamente y se colocaron en un recipiente herméticamente cerrado conteniendo espuma refrigerante PolarPack® completamente congelada. Esto pudo conservar las muestras en un estado fresco hasta llegar al laboratorio en donde se colocaron en un refrigerador a -21o C. Estas muestras se mantuvieron en óptimas condiciones de frescura hasta que fueron procesadas. Usando unas tijeras filosas se procedió a cortar las hojas en pequeños trocitos. Para la extracción de ADN se usaron 2 gramos de muestra y para la extracción de proteínas se usó un gramo de muestra.

9
Caracterización agro-morfológica:
Se seleccionaron árboles de la finca comercial del Dr. Mois Haché los cuales se designaron con las siglas FMH. Los árboles seleccionados de la finca comercial del señor Amable Díaz fueron designados con las siglas FAD y los seleccionados de la finca comercial del señor Angel Peguero fueron designados con las siglas FAP. Los criterios tomados en consideración para la selección de los árboles fueron basados en las recomendaciones de los productores, las cuales incluían la productividad de los árboles particulares, la uniformidad y tamaño de los frutos y la uniformidad visual de los árboles. De la finca del Dr. Mois Haché se seleccionaron doce (12) árboles individuales; de la finca del señor Amable Díaz se seleccionaron diez (10) árboles; y de la finca del señor Ángel Peguero se seleccionaron cinco (5) árboles. Esto dió un total de veintisiete (27) árboles individuales en el estudio. A cada árbol seleccionado para análisis de AFLP se le tomaron muestras de frutos (cinco por cada árbol) para evaluar la morfología y calidad de los mismos. Los parámetros que se midieron fueron los siguientes:
ParámetroDiámetro del fruto (DF), cmLongitud del fruto (LF), cmPeso del fruto (PF), gmDiámetro de la semilla (DS), cmLongitud de la semilla (LS), cmPeso de la semilla (PS), gmPeso de la cáscara (PC), gmPeso del mesocarpio (PM), gmGrado brix (GB), % sólidos solubles
Caracterización molecular:
La extracción de ADN se llevó a cabo usando un protocolo de D.L. Nickrent (Molecular Methods in Plant Biology, Second Edition, 1996. Department of Plant Biology Southern Illinois University) el cual se modificó y se adaptó para su uso en mango por ser esta una planta muy difícil para la extracción de ADN.
Se molieron 3 gramos de cada muestra en un mortero con nitrógeno líquido hasta obtener un polvo fino y se añadió una cantidad de 25 ml de buffer CTAB previamente calentado en un baño de agua a 95º C, después de lo cual se continuó moliendo por un minuto adicional. Se coló el extracto en un tubo de centrífuga de 50 ml usando un embudo y gasa. A este extracto se le añadieron 50 unidades de proteasa K y 0.5 ml de DTT (Ditiotreitol) (Sigma D0632) y después de mezclar bien se colocó en un baño maría 1 hora con movida ocasional. Se añadió 2/3 volumen de cloroformo:isoamílico (24:1) y se centrifugó a 8,500 rpm por 15 minutos después de agitarse cuidadosamente en una centrífuga de alta capacidad marca Fisher Marathon 21000 (Fisher Scientific, Pittsburg PA, USA). Después de la centrifugación se separó la capa acuosa superior y se colocó en un tubo limpio de 50 ml a la cual se le añadió 2/3 volumen de isopropanol frío y se colocó a -20º C por al menos una hora después de lo cual se centrifugó por 20 minutos a 10,000 rpm para la precipitación del ADN.
El ADN se re-purificó con fenol:cloroformo:isoamílico dos veces dando como resultado un ADN de muy buena calidad. La cuantificación del ADN se realizó a través de un proceso de espectrofotometría con un espectrofotómetro marca BECKMAN COULTER Modelo DU Series 700 UV/Vis de rastreo (Beckman Coulter, Inc. Fullerton, California USA. El proceso de AFLP se llevó a cabo con un kit para tales fines provisto por Invitrogen Corporation (invitrogen™ AFLP® Analysis System I), Carlsbad, CA, siguiendo el protocolo incluído en éste. Los ciclos de PCR se llevaron a cabo en un termociclador marca Genius (Techne Incorporated, New Jersey, 08540-6328, USA).La electroforesis se corrió en una cámara vertical marca FisherBiotech modelo FB-SEQ-3345 (Fisher Scientific, Pittsburg, Pa, USA) y la tinción se realizó con nitrato de plata al 0.2% por 20 minutos después de haberse oxidado
Con el empleo del paquete de estadística del Programa SAS (Sta-tistical Analysis System), se realizó un análisis de estos parámetros
cuyos resultados se muestran más adelante.

1010
con ácido nítrico (1%) por 3 minutos. La preservación de las gelatinas para su lectura se hizo por tratamiento con metanol al 10% por 15 minutos.
Resultados y Discusión
De acuerdo a los datos obtenidos en los análisis fenotípicos, los árboles FMH1, FMH8, FMH10, FMH12, FAD3, FAD4, FAD5, FAD10, FAP2 y FAP3 fueron seleccionados para continuar su evaluación fenotípica además y de los análisis de AFLP. Los promedios de los parámetros analizados para los diez árboles seleccionados se muestran en la Tabla 1. Los árboles FMH1 y FMH10 de la finca del señor Mois Haché fueron preseleccionados por sus cualidades al igual que los árboles FAD4 y FAD10 de la finca del señor Amable Días y el árbol FAP3 de la finca del señor Ángel Peguero. Estos resultados muestran la gran variabilidad morfológica y organoléptica que existe en los cultivares de mangos de los denominados banilejos que se cultivan en la región sur del país, específicamente en la Provincia Peravia de la República Dominicana.
Para los análisis de AFLP se hicieron numerosas pruebas preliminares para determinar la mejor combinación de cebadores. En las pruebas preliminares se realizaron 35 combinaciones de cebadores de las cuales se seleccionaron 4 como las mejores. Estas fueron: EAGC-MCTG, EAGG-MCTG, EACA-MCAT y EACA-MCTA (ENNN se refiere a los cebadores que se adhieren al sitio de restricción de la EcoR I, mientras que N es un nucleótido selectivo que se extiende al fragmento de restricción; MNNN se refiere al sitio de restricción de la Mse I). Con estas cuatro combinaciones se trabajó durante un tiempo para determinar con cual se obtenia mejor. La combinación de cebadores que se usó en este kit fue EACA- MCTA por ser la que mejores resultados dió durante las pruebas preliminares. Con esta combinación se analizaron las 38 muestras y se obtuvieron excelentes y numerosas bandas polimórficas en las gelatinas. Los análisis de AFLP muestran gran diferencia entre las muestras analizadas como se ve en las figuras 1 y 2.
Para evaluar la similitud de las muestras mediante AFLP, se hizo un análisis cualitativo de las bandas esparcidas sobre las gelatinas como muestra la Figura 1. La Figura 2 es una representación diagramática parcial de los patrones de bandas de la Figura 1 donde se muestran las posiciones de las bandas de cada uno de los individuos. Estos patrones de bandas propios de cada muestra fueron expresados en una matriz numérica compuesta por 0 (cero) para la ausencia de banda y 1 (uno) para la presencia de las mismas. Los datos obtenidos con este sistema fueron sometidos a un análisis de similitud usando el paquete estadístico Statistica y el subprograma STA-CLU, especial para hacer análisis de conglomerados, con el método de agrupamiento de distancia euclidiana obteniéndose el dendrograma que aparece en la Figura 3.
En la figura 3 se muestra un dendrograma de las 38 muestras analizadas mediante la técnica de AFLP. Las 38 muestras fueron separadas en cinco grupos distintos con diferentes distancias de ligamiento entre si. El primer grupo está compuesto por cuatro individuos con tres diferentes distancias de los cuales dos individuos (FAD55 y FAD53) son idénticos. En este grupo sólo un individuo (FMH3) no procede de la finca del señor Amable Díaz. El árbol FMH3 y el FAD8 aunque no idénticos entre sí tienen una distancia de ligamiento muy cercana.
El segundo grupo, compuesto por seis individuos de la misma finca, separó cuatro distancias de ligamiento diferentes. Los árboles FAD21 y FAD6 resultaron idénticos entre sí al igual que los FAD9 y FAD5 aunque las diferencias entre los seis integrantes de este grupo no son muchas. De este grupo, el árbol FAD4, con poca relación con los otros miembros del grupo, fue preseleccionado por sus cualidades fenotípicas. El grupo 3 del dendrograma está compuesto por doce individuos con marcada variación en las distancias de ligamiento. De este grupo, los árboles FMH1 y FMH10 fueron preseleccionados. En este grupo se pueden notar siete diferentes subgrupos de distancias de ligamiento con identidad de los individuos FMH5 y FMH4; FMH1 y FAD51; FMH10 y FMH8; y FAD42 y FAD41.

11
El grupo 4 de siete árboles está compuesto por 4 subgrupos de distancias de ligamiento. Los individuos FAD2 y FAP5 son idénticos entre sí al igual que FAD1 y FAP4; y FAD3 y FAP2. En este grupo no se preseleccionó ningún árbol. El último grupo (5) de nueve individuos identificó siete subgrupos de distancias de ligamiento de los cuales se preseleccionaron los árboles FAD10 y FAP3 por sus cualidades fenotípicas. En este grupo sólo los árboles FAD11 y FAP1 resultaron idénticos. Este fue el grupo que resultó con mayores diferencias en distancias de ligamiento.
Los árboles seleccionados fueron localizados en los grupos 2, 3 y 5. FAD4 fue localizado en el grupo 2 siendo este árbol diferente a todos los otros del grupo. El grupo 3 incluyó los árboles seleccionados FMH3 y FMH10 los cuales, aunque en el mismo grupo mostraron una gran distancia genética entre sí. El grupo 5 incluyó los árboles seleccionados FAD10 y FAP3 en el cual el árbol FAP3 no presento ningún parentesco con el resto del grupo y cuya distancia genética con el FAD10 fue muy pronunciada. Estos árboles fueron debidamente identificados, y deberán ser cuidados y acondicionados para que sirvan de suplidores de yemas en programas de propagación de la variedad.
Agradecimiento
Los autores quieren expresar su agradecimiento al Consejo Nacional de Investigaciones Agropecuarias y Forestales (CONIAF) no sólo por el apoyo financiero que brindó a este proyecto, sino por el entusiasmo que siempre mostró en la realización del mismo y por la extensión del plazo otorgado para la finalización del mismo. Va nuestro agradecimiento particular al Profesor Ing. Gabriel Domínguez por su apoyo continuo al proyecto y su comprensión en las razones de atraso en la finalización.
Al Ing. Agron. Rafael Matos por su colaboración en el análisis estadístico de los datos. A todo el personal del IIBI y del CEBIVE que colaboraron para llevar a cabo este proyecto en especial a la Dra. Bernarda Castillo por su decidido apoyo al proyecto y por su pronta respuesta a las solicitudes de compra de materiales y al Ing. Félix Rivas por el continuo apoyo moral.
Al Cluster de Mango por darnos su aval en esta investigación y en particular al Dr. Mois Haché, al señor Angel Peguero y al señor Amable Díaz por permitirnos usar sus fincas en la recolección de muestras y por su apoyo incondicional que nos brindaron.
La realización de esta investigación fue posible gracias al aporte del Fondo Concursable de Investigacion (FONDOCYT) Del Ministerio de Educación Superior, Ciencia y Tecnología de la República Dominicana (MESCYT) quien proveyó parte del financiamiento.
Bibliografía
Corsini, G., A. Manubens, M. Lladser, S. Lobos, and D. Seelenfreund. 1999. AFLP Analysis of the Fruit Fly Ceratitis capitata. Focus Volume 21 Number 3, 72-73.
Geilfus, Frans. 1989. El árbol al servicio del agricultor: manual de agroforestería para el desarrollo rural. Vol. 2: guía de especies. Santo Domingo, DO: Enda.Caribe y CATIE, pp.3.
Gutiérrez Mulas, M., Morela Fuchs P. Lourdes del C. González R. Asia Y, Zambrano. 2004. Caracterización isoenzimática de genotipos de algodón (Gossypium sp.) del banco de germoplasma del CENIAP, Venezuela. PGR Newsletter No.130, pp. 20-24.
Herrero, R., M.J. As¡ns, E.A. Carbonell and L. Navarro. 1996. Genetic diversity in the orange subfamily

1212
Aurantioideae. I. Intraspecies and intragenus variability. Theor. Appl. Genet. 92:599-609.
Jacobs, B.F., C.R. Perth, and S. Guttman. 1984. Genetic relationships in Abies (fir) of eastern United States: an electrophoretic study. Can. J. Bot. 62:609-616. Kashkush, K., F. Jinggui, E. Tomer, J. Hillel, and U. Lavi. 2001. Cultivar identification and genetic map of mango (Mangifera indica). , Vol. 122, No. 1, pp. 129-136(8).
Lavi, U., K. Kashkush, D. Sa'ada, H. Shats, U. Ravid, and E. Tomer. 2004. Mango breeding and the potential of modern biology. Acta Hort. (ISHS) 645:51-59. Nickrent, D.L. Molecular Methods in Plant Biology, Second Edition, 1996. Department of Plant Biology Southern Illinois University.
Patel, D. 1994.Gel Electrophoresis: Essential Data. 1994. John Wiley & Sons. Chichester, UK. Pp. 8-56.Samson, J. A. Tropical Fruits. 2nd ed. Longman Scientific and Technical. 1986. pp. 216-234.
Vainstein, A. and H. Ben-Meir. 1994. DNA Fingerprint Analysis of Roses. J. Amer. Soc. Hort. Sci. 119(5): 1099-1103. Vallejos, C.E. 1983. Enzyme Activity Staining. In: S.D. Tanksley and T.J. Orton (Editors), Isozymes in Plant Genetics and Breeding, Part A. Elsevier Science Publishers B.V., Amsterdam.
Vos, P. and M. Kuiper. 1997. AFLP analysis pp. 115-131: In G. Caetano-Anollés and P.M. Gresshoff, editors. DNA markers: protocol, applications, and overviews. Wiley-Liss, Inc. New York. 1997.
Anexos.
Tabla 1. Promedio de los parámetros analizados en las diez muestras seleccionadas (segunda cosecha). MuestraParámetro FMH1 FMH8 FMH10 FMH12 FAD3 FAD4 FAD5 FAD10 FAP2 FAP3
DF, cm 6.84 5.04 5.84 5.90 5.62 5.56 5.52 5.96 5.42 5.94
LF, cm 8.5 6.72 7.5 7.96 6.96 7.08 6.94 6.96 6.70 7.22
PF, gm 181.1 119.16 176.88 172.04 158.2 155.8 149.54 170.78 148.56 177.24
DS, cm 3.76 2.84 3.22 3.08 3.32 2.92 2.92 3.18 2.78 2.90
LS, cm 6.5 5.24 5.78 5.94 5.52 5.30 5.14 5.20 4.86 5.26
PS, gm 40.62 25.78 36.7 33.66 33.98 28.76 28.22 29.44 27.10 29.34
PC, gm 32.64 20.72 27.34 29.60 26.00 26.10 24.46 26.00 21.58 29.70
PM, gm 122.04 72.02 110.1 106.44 98.86 100.24 96.24 115.10 99.34 117.12
GB, %ss 17.2 18.2 18.4 17.0 19.0 17.0 20.2 18.6 20.4 22.2
PS/PF 0.19 0.21 0.21 0.20 0.21 0.19 0.19 0.17 0.18 0.17
Figura1. Gelatina de electroforesis mostrando las bandas de AFLP.

13
Muestra Posición BandasFAP1 FAP2 FAP3 FAD4 FAD5 FAD6 FMH1 FMH2 FMH3 FMH4 FMH5
▬ ▬ 1
▬ 2
▬ 3
▬ ▬ ▬ ▬ ▬ ▬ ▬ ▬ 4
▬ ▬ ▬ ▬ ▬ ▬ ▬ ▬ 5
▬ ▬ ▬ ▬ ▬ ▬ ▬ 6
▬ ▬ ▬ ▬ ▬ ▬ 7
▬ ▬ ▬ ▬ ▬ ▬ ▬ 8
▬ ▬ ▬ ▬ ▬ 9
▬ ▬ ▬ ▬ ▬ ▬ 10
▬ ▬ ▬ ▬ ▬ ▬ 11
▬ ▬ ▬ ▬ ▬ ▬ 12
▬ ▬ ▬ ▬ ▬ ▬ 13
▬ ▬ ▬ ▬ ▬ ▬ ▬ ▬ ▬ ▬ 14
▬ ▬ ▬ ▬ ▬ ▬ 15
▬ ▬ ▬ ▬ ▬ ▬ ▬ 16
▬ ▬ ▬ ▬ ▬ ▬ ▬ 17
▬ ▬ ▬ ▬ ▬ 18
▬ ▬ ▬ ▬ 19
▬ ▬ ▬ ▬ ▬ ▬ ▬ ▬ 20
▬ ▬ ▬ ▬ ▬ 21
▬ ▬ ▬ ▬ ▬ ▬ 22
▬ ▬ ▬ ▬ ▬ 23
▬ ▬ ▬ ▬ ▬ ▬ ▬ ▬ ▬ 24
▬ ▬ ▬ 25
▬ ▬ ▬ ▬ ▬ ▬ ▬ ▬ ▬ 26
▬ ▬ 27
▬ ▬ ▬ ▬ ▬ ▬ ▬ 28
▬ ▬ ▬ 29
▬ ▬ ▬ ▬ ▬ ▬ 30
▬ ▬ ▬ 31
▬ ▬ ▬ ▬ ▬ ▬ 32
▬ ▬ 33
▬ ▬ ▬ ▬ ▬ ▬ ▬ 34
▬ ▬ ▬ ▬ ▬ ▬ 35
▬ ▬ ▬ ▬ ▬ 36
▬ ▬ ▬ ▬ ▬ 37
▬ ▬ ▬ ▬ ▬ ▬ 38
▬ ▬ ▬ ▬ ▬ ▬ ▬ ▬ ▬ ▬ 39
▬ ▬ ▬ ▬ ▬ ▬ ▬ 40

1414
▬ ▬ ▬ ▬ ▬ ▬ 41
▬ ▬ ▬ ▬ ▬ 42
▬ ▬ ▬ ▬ ▬ ▬ ▬ 43
▬ ▬ ▬ ▬ ▬ 44
▬ ▬ ▬ ▬ 45
▬ ▬ ▬ ▬ 46
▬ ▬ ▬ ▬ ▬ ▬ 47
▬ ▬ ▬ ▬ ▬ ▬ ▬ ▬ 48
▬ ▬ ▬ ▬ ▬ ▬ 49
▬ ▬ ▬ ▬ ▬ ▬ 50
▬ ▬ ▬ ▬ ▬ ▬ 51
▬ ▬ ▬ ▬ ▬ ▬ ▬ ▬ 52
▬ ▬ ▬ ▬ 53
▬ ▬ ▬ ▬ ▬ 54
▬ ▬ ▬ ▬ ▬ 55
56
▬ 57
▬ ▬ ▬ 58
▬ ▬ ▬ ▬ 59
▬ ▬ ▬ ▬ ▬ ▬ ▬ 60
Figura 2. Representación diagramático parcial de los patrones de banda obtenidos en la corrida de electroforesis en gelatina de poliacrilamida de los AFLP. Por cuestión de espacio no se pueden mostrar todas las bandas y todos los individuos.
Figura 3. Dendrograma de las 38 muestra analizadas mediante la técnica de AFLP.

15
Respuesta a cinco tratamientos de asepsia en distintos explantes de guanábana (Annona muricata L.)Yaneury Doñé Dipré, Adriana Chavarría Lines y Luis Wong-Vega
Palabras clave: Annona muricata L., guanábana, asepsia, micropropagación
Resumen
La guanábana (Annona muricata L) es una especie frutal de gran interés en la fabricación de pulpas y jugos. Sus estudios biotecnológicos hasta ahora han sido muy escasos, de ahí la importancia de establecer un método de asepsia efectivo que permita obtener material limpio para transformar genéticamente la especie. En este estudio se probaron cinco diferentes tratamientos de asepsia que fueron adaptados de otros tratamientos de especies cercanas. Los tratamientos más efectivos en cuanto a la viabilidad y el control de la contaminación de los explantes fueron la combinación de los agentes (Agrimicin y Bravo) y el hipoclorito de sodio durante 5 minutos. Sin embargo, el explante que presentó la mayor viabilidad fue el hipocotilo que llegó a formar plantas asépticas.
Keywords: Annona muricata L., soursop, asepsis, micropropagation.
Abstract
The soursop (Annona muricata) is a fruit species of great interest in the manufacture of pulp and juice. their biotechnological studies so far have been very limited, hence the importance of establishing an effective aseptic method to obtain clean material to genetically transform this species. In this study we tested five different species treatments nearby. The most effective treatments in terms of viability and pollution control explants were the combination of agents (agrimicin and bravo) and sodium hypochlorite for 5 minutes. However, the explant showed the highest viability was the hypochlorite that made aseptic plants.”
Introducción
La guanábana (Annona muricata L.) es una especie originaria de América tropical y pertenece a la familia Annonaceae. Este frutal tuvo una amplia expansión en tiempos prehispánicos, sin embargo, no se conoce en estado silvestre. (Ramírez-Villalobos et al, 2002)
Las especies del género Annona se han propagado tradicionalmente por semilla y los métodos convencionales de propagación vegetativa son muy lentos. Debido a que las colecciones de estas plantas en el campo, generalmente están expuestas a plagas, enfermedades, problemas edáficos, climáticos y de espacio, el cultivo in Vitro puede ser utilizado para establecer bancos de germoplasma asépticos y conservar así la diversidad necesaria. (Velásquez et al, 2004) (Rivero et al, 2006).

1616
Además, el cultivo in Vitro ofrece muchas ventajas en la práctica de la multiplicación vegetativa, entre las cuales destaca la propagación de un gran número de especies difíciles de multiplicar por los métodos clásicos. (Velásquez et al, 2004)
Para el caso de Annona muricata L. no se tiene reportado en la literatura un método de asepsia así como la manipulación genética de la especie. De ahí la importancia de establecer un método de asepsia efectivo, ya que es el paso inicial de cualquier estudio de transformación genética.
En este estudio se comparó la efectividad de cinco tratamientos de asepsia adaptados de trabajos similares en otras especies.
Materiales y Métodos
Para cada experimento independiente, se tomaron tres partes de la planta joven de guanábana crecidas en el vivero; la radícula, la plúmula y el hipocotilo. Se ensayaron cinco tratamientos de desinfección para cada tejido joven, los cuales fueron evaluados al cabo de una semana. Los tratamientos para estos tejidos fueron:
1. Inmersión de los tejidos en Bravo al 0.5% y Agrimicin al 0.3% por 30 minutos, seguido de un baño de cloro al 5.25% por 15 minutos, seguido de tres enjuagues con agua destilada (30 segundos cada enjuague) y al final un enjuague en ácido cítrico al 0.5%.
2. Inmersión de los tejidos en Bravo al 0.5% y Agrimicin al 0.3% por 30 minutos, seguido de un baño de cloro al 5.25% por 5 minutos, seguido de tres enjuagues con agua destilada (30 segundos cada enjuague) y al final un enjuague en ácido cítrico al 0.5%.
3. Inmersión de los tejidos en Bravo al 0.5% y Agrimicin al 0.3% por 30 minutos, seguido de un baño de cloro al 1% por 5 minutos, seguido de tres enjuagues con agua destilada (30 segundos cada enjuague) y al final un enjuague en ácido cítrico al 0.5%.
4. Inmersión de los tejidos en Bravo al 1% y Agrimicin al 1% por 30 minutos, seguido de un baño de cloro al 5.25% por 5 minutos, seguido de tres enjuagues con agua destilada (30 segundos cada enjuague) y al final un enjuague en ácido cítrico al 0.5%.
5. Inmersión de los tejidos en Bravo al 0.5% y Agrimicin al 0.3% por 4 horas, seguido de un baño de cloro al 5.25% por 5 minutos, seguido de tres enjuagues con agua destilada (30 segundos cada enjuague) y al final un enjuague en ácido cítrico al 0.5%.
Una vez finalizado el tratamiento de asepsia, los explantes fueron inoculados en medio de cultivo MS. Cada frasco contenía un solo tipo de tejido seccionado en tres partes. La eficacia de cada tratamiento fue evaluada cada día y se contaba como positiva si el explante y el medio de cultivo estaba libre de cualquier patógeno, hongo o bacteria. Para cada tratamiento se inoculaban siete frascos, es decir en cada repetición se tenían siete frascos por cada tratamiento. Se hicieron tres repeticiones de cada ronda de experimentos. Cada ronda de experimentos tenía los cinco tratamientos de asepsia.
17
Resultados
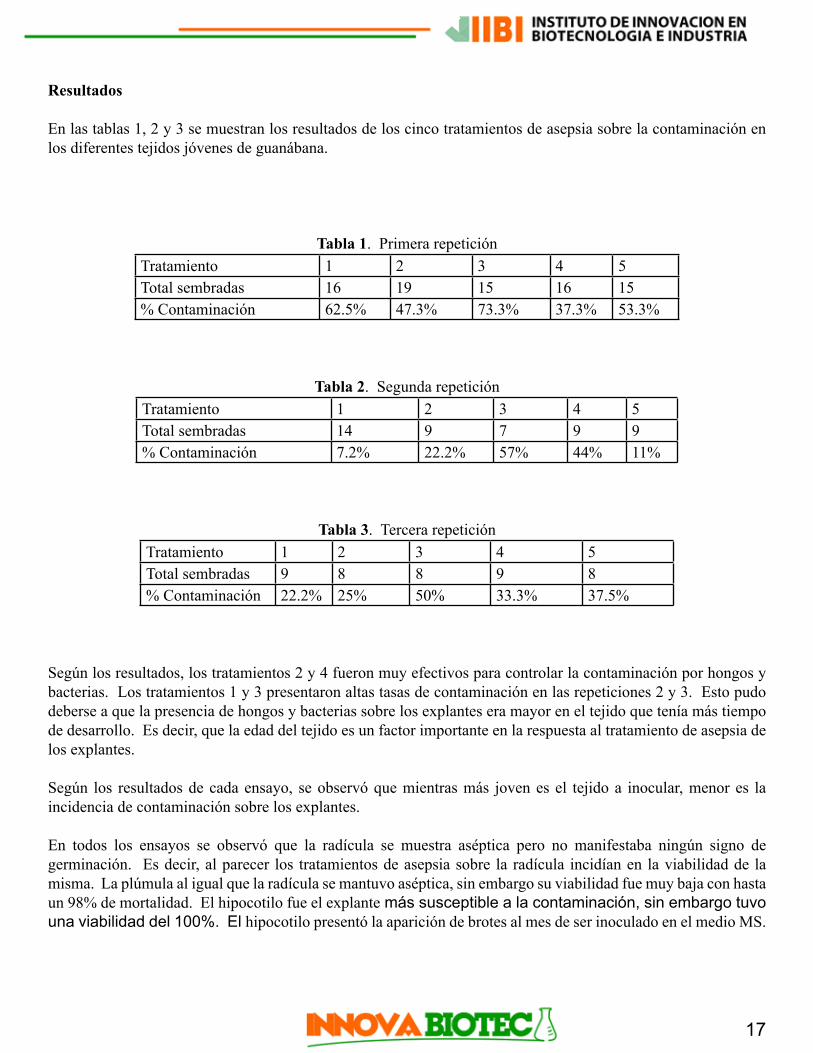
En las tablas 1, 2 y 3 se muestran los resultados de los cinco tratamientos de asepsia sobre la contaminación en los diferentes tejidos jóvenes de guanábana.
Tabla 1. Primera repeticiónTratamiento 1 2 3 4 5Total sembradas 16 19 15 16 15% Contaminación 62.5% 47.3% 73.3% 37.3% 53.3%
Tabla 2. Segunda repeticiónTratamiento 1 2 3 4 5Total sembradas 14 9 7 9 9% Contaminación 7.2% 22.2% 57% 44% 11%
Tabla 3. Tercera repeticiónTratamiento 1 2 3 4 5Total sembradas 9 8 8 9 8% Contaminación 22.2% 25% 50% 33.3% 37.5%
Según los resultados, los tratamientos 2 y 4 fueron muy efectivos para controlar la contaminación por hongos y bacterias. Los tratamientos 1 y 3 presentaron altas tasas de contaminación en las repeticiones 2 y 3. Esto pudo deberse a que la presencia de hongos y bacterias sobre los explantes era mayor en el tejido que tenía más tiempo de desarrollo. Es decir, que la edad del tejido es un factor importante en la respuesta al tratamiento de asepsia de los explantes.
Según los resultados de cada ensayo, se observó que mientras más joven es el tejido a inocular, menor es la incidencia de contaminación sobre los explantes.
En todos los ensayos se observó que la radícula se muestra aséptica pero no manifestaba ningún signo de germinación. Es decir, al parecer los tratamientos de asepsia sobre la radícula incidían en la viabilidad de la misma. La plúmula al igual que la radícula se mantuvo aséptica, sin embargo su viabilidad fue muy baja con hasta un 98% de mortalidad. El hipocotilo fue el explante más susceptible a la contaminación, sin embargo tuvo una viabilidad del 100%. El hipocotilo presentó la aparición de brotes al mes de ser inoculado en el medio MS.

1818
Discusión
Los resultados anteriores sugieren que la combinación de agentes (Bravo y Agrimicin) y el hipoclorito de sodio afectan la viabilidad de la radícula, es decir, aún cuando la misma está aséptica no presenta regeneración. Por otra parte, la plúmula al ser tratada con los agentes (Bravo y Agrimicin) y el hipoclorito de sodio, presenta una alta mortalidad. Estas al cabo de cinco días finalizan por completo.
Las hojas de las plantas germinadas a partir de los hipocotilos asépticos fueron seccionadas e inoculadas en el medio MS con distintas combinaciones de hormonas. Al cabo de dos meses los explantes no presentaron contaminación bacteriana ni fúngica, lo que indicaría que el método de asepsia ha sido efectivo para establecer tejido limpio.
Agradecimientos
La realización de esta investigación fue posible gracias al aporte del Fondo Concursable de Investigacion (FONDOCYT) Del Ministerio de Educación Superior, Ciencia y Tecnología de la República Dominicana (MESCYT) quien proveyó parte del financiamiento.
Bibliografía
Ramírez-Villalobos, M., Urdaneta, A. & S. León de Sierralta. 2002. Establecimiento in Vitro de explantes adultos del guanábano (Annona muricata L.) tratados con hipoclorito de sodio. Rev. Fac. Agron. 19 (1): 48-55.
Rivero, G., Ramírez, M. & S. Sierralta. 2006. Efecto de la planta madre en el establecimiento in Vitro del guanábano. Agronomía Tropical 56(4): 627-631.
Velázquez, M., González, A., Mata, F. León de Sierralta, S., Esparza, D. & M. Ramírez. 2004. Tipo de sombreamiento y tiempo de crecimiento de brotes laterales sobre la viabilidad de explantes de Annona muricata L. Rev. Fac. Agron.-Luz 21(1): 12-18.

19
Harina de Nopal (Opuntia ficus) aplicada en la Elaboración de Productos de Panificación.E. Lucía Berigüete, Oscar Merette, Elisa Gómez, Carlos Calderón
Palabras Claves: Nopal (Opuntia ficus), cladodios, harina, deshidratado, panificación
Resumen
Se realizó una caracterización del nopal por familia, género y subgénero en el jardín botánico, también se determinaron las principales características nutricionales, la ubicación de poblaciones silvestres o posibles cultivares en el país, se estableció el cultivo a nivel experimental en el IIBI del género Opuntia), con la finalidad de disponer de materia prima durante los ensayos de producción de harina en pequeña escala en laboratorios y producción a nivel piloto en la planta del IIBI. La validación y uso de la harina de nopal en la elaboración de productos de panificación se realizó con la colaboración de la Panificadora Moderna ubicada en el ensanche la Fe y la Panadería el Metro ubicada en Villa Mella, República Dominicana. La harina de nopal se utilizó en sustitución de la harina de trigo a diferentes proporciones en la elaboración de productos de panificación de consumo cotidiano que se comercializan en el país: empanada, Pan, Bizcocho, Galletas, Coconete y Pudín.
Para cada producto de panificación se establecieron los diseños y protocolos de elaboración, respetando la formulación de los productos de cada panificadora o panadería y sólo se sustituyeron los porcentajes de la harina de nopal a incorporar en los productos del %: 30, 25 20, 15 y 10., y se realizó una evaluación sensorial utilizando la escala hedónica de cinco puntos para cada producto (Anzaldúa, A. 1994). Los atributos evaluados en el pan utilizando el 10% de harina de Nopal, la apariencia, sabor y color con puntuación de 4 a 5, el menos valorado fue el olor en una escala de 3. En el coconete utilizando un 30%, los atributos evaluados de color, sabor, olor y apariencia fueron calificados de 4 a 5 puntos en la escala hedónica. Este análisis estableció que es factible sustituir un promedio 20 % de harina de nopal en productos de panificación por no haber diferencias significativas en cuanto a los atributos evaluados con y sin nopal. En el análisis químico presentó un incremento nutricional favorable en el contenido de calcio, calorías y grasa. (Laboratorios IIBI), en costo fue establecida una competitividad comercial favorable al uso de harina de nopal.
Keywords: Nopal (Opuntia ficus), pads, flour, powder, baking Abstract. Characterization was performed by cactus family, genus, subgenus in the botanical garden, also identifies the key nutritional characteristics, the location of wild populations or potential cultivars in the country, was established on an experimental crop in the genus Opuntia IIBI ), in order to provide raw material for production testing of flour in small scale laboratory and pilot scale production plant in IIBI. The validation and use of nopal flour in the preparation of bakery products was conducted in collaboration with the Moderna Bakery located in the widening of the Faith and the Metro Bakery located in Villa Mella, Dominican Republic. This flour is used in place of wheat flour in different proportions in the preparation of bakery products that are traded daily consumption in the country: Pie, Bread, Biscuits, Cookies, Coconet, and pudding.

2020
For each bakery product were established protocol design and development, respecting the formulation of the products of each bakery or bakery and only replaced the percentage of nopal flour to incorporate into the products of% 30, 25 20, 15 and 10., and conducted a sensory evaluation using the five-point hedonic scale for each product (Anzaldúa, A. 1994). The attributes evaluated using bread flour 10% Nopal, as the best were the appearance, taste and color score of 4 to 5 I like and love, the smell was the lowest rated on a scale of 3 or I like or dislike. In concómete using a 30% color attributes evaluated taste; smell and appearance were rated from 4 to 5 points hedonic scale. This analysis demonstrated that it is feasible to replace an average 20% of nopal flour in bakery products by no significant difference in terms of attributes evaluated with and without cactus and chemical analysis showed an increase in favorable nutritional content of calcium, Calories and fat. (IIBI Laboratories) in costs was established favorable business competitiveness using the nopal flour.
Introducción
El nopal (familia: Cactáceas, género: opuntia, subgénero: Platypuntia). Es originario de América tropical y subtropical; se encuentra entre las principales plantas que se localizan en las zonas tropicales y subtropicales de África, Asia y América Latina, lugares donde predominan temperaturas altas (FAO 2006), hoy día, se encuentra ampliamente distribuido en todo el mundo en forma silvestre o cultivada, en una gran variedad de condiciones agroclimáticas (Sáenz C., 2006).
El nopal abunda de manera silvestre en la República Dominicana, se produce en todas las zonas semi-áridas y ocupa dimensiones territoriales en las regiones suroeste y nordeste del país. Las analíticas realizadas a la materia prima del nopal (raquetas, botánicamente llamadas cladodios o pencas), aportan importantes cantidades de nutrientes que varían según el estadio del corte o cosecha, clasificando como tierno, semi tierno y lleno. Fué seleccionado el semi-tierno para la elaboración de harina por aportar más nutrientes característicos de la especies. (Laboratorio IIBI, 2006). La harina de nopal o polvo de nopal tiene aplicación en la industria panificadora en la preparación de galletas, panes, bizcochos, pastas, cremas y postres. Su aporte en fibras dietéticas resulta importante, en virtud de que el consumo de fibras de tipo soluble, representa una mejoría significativa de los procesos digestivos, constipación, alimentos dietéticos demandados por los consumidores. (Sáenz C., 2006).
Por Iniciativa del IIBI, el apoyo de la FAO, la participación la Panificadora Moderna y la Panadería el Metro, se procedió a reunir información sobre las principales variedades de nopales en el país, su valor nutricional, alimenticio y usos, por lo que fue seleccionada la variedad con menos espina y fácil manipulación en los procesos de elaboración (opuntia ficus). El proceso de elaboración de harina de nopal fue establecido con un protocolo desarrollado por el IIBI, basado en los parámetros de control de los procesos (humedad, granometría) y las normativas nacionales para la molienda. El porcentaje de harina de nopal de un 20% establecido por su aporte nutricional y disponibilidad de materia prima local combinada con el 80% la harina de trigo mejoró los aportes nutricionales, organolépticos y económicos a las formulaciones de panificación que se realizaron y estudiaron.
Materiales y Métodos
Ubicación y caracterización materia prima.
Este estudio fue realizado en las instalaciones del IIBI, en la planta piloto y laboratorios, la materia prima para la elaboración de la harina (los cladodios frescos) de nopal; para la parte experimental fueron recolectados en la plantación experimental del IIBI. La producción a mediana escala para la formulación del producto, la materia prima se obtuvo del cultivo modelo en el Centro de Biotecnología Vegetal del IIBI.
Obtención de la Harina
El procedimiento para la obtención de la harina de nopal consistió en la recolección de los cladodios semi tierno, de 15 a 20 días del brote, la selección fue sin daño físico ni biológico, el lavado y el retiro de las espinas se realizó con agua potable comercial y la desinfección consistió en sumergir los cladodios en una solución de cloro

21
50ppm., por 10 minutos y luego fueron enjaguados con abundante agua potable. Para el rebanado se realizó en una rebanadora marca SNF de acero inoxidable, a un grosor de 2cm para logar partes uniformes y obtener el mínimo de tiempo en la deshidratación. La deshidratación del nopal fue realizada en un deshidratador marca TSUNG HSING, donado por la Misión Taiwán al IIBI, con 40 bandejas de 72.2 cm2, con un voltaje de 110 VAC, alimentado con gas propano como combustible. El nopal rebanado uniformemente fue colocado en las bandejas y puesto en deshidratador por 3 horas a una temperatura de 65°C., con un control de actividad de agua (Aw) cada 30 minutos, utilizando un medidor de actividad de agua marca ROTRONIC.
El proseo de molienda se realizó en el molino D6A de martillo marca FITZMILL utilizando un tamiz con una abertura de 0.30 mm con perforaciones redondas en acero inoxidable, el cual fue alimentado con nopal seco con AW 0.4 y se procedió a la molienda para obtener la harina de nopal, la cual fue empacada para enviar a los laboratorios del IIBI, las panaderías y almacenada a una temperatura ambiente de 23°c promedio y una humedad relativa menos de 60% tomada con un Hidrómetro marca Fisher Scintific, y con estos controles fue establecida la vida útil en anaquel de la harina de nopal.
Elaboración de los productos de panificación
La formulación de los productos de panificación fue establecida con la participación de los industriales de la harina y maestro de panadería, seleccionados los productos de mayor venta cotidiana en la panadería el Metro y panificadora Moderna: panes, bizcochitos, coconetes, pudín y empanadas. Estos y otros productos fueron revalidados para degustación y exhibición como: galletas dulces y saladas, bizcochos. Estas panaderías están ubicadas en la provincia Santo Domingo y el Distrito Nacional. Respetando la formulación de los productos de cada empresa, le acompañó un técnico del IIBI, especialista en nutrición y tecnología de los alimentos, para la mezcla planteada sustituyendo un % de 10, 15, 20, 25, 30 y 35 de la harina de trigo por la harina de nopal, para establecer sus características organolépticas y nutricional. Empleando el método oficial de la AOAC (1990) y otros autorizados y validados, todos los análisis se realizaron por duplicado en los laboratorios del IIBI. Las determinaciones analísticas al nopal fresco en tres etapas de crecimiento fueron: Tierno con menos de diez días, semi-tierno entre 10 a 30 días y lleno con mas de 30 días después del brote del cladodio; para establecer la cosecha según el mejor aporte nutricional. Formulada una mezcla de un 80% harina de trigo y un 20% harina de nopal (MHTN01), se determinaron los nutrientes de interés y se analizó VS harina de trigo 100% (MHT02). Los productos de panificación formulados con MHTN01 y los mismos productos de panificación formulados con (MHT02), fueron evaluados por la diferencia mayor o menor de: energía, grasas, carbohidratos, azucares totales, proteínas, cenizas, calcio, hierro, zinc, fósforo y humedad.
Para establecer el perfil organoléptico de los productos de panificación, se aplicó una evaluación sensorial comparativa entre los productos de panificación empleando la mezcla MHTN01 y los productos de panificación convencionales. Este estudio fue realizado por un panel de 20 jueces no entrenados del IIBI, cuyas edades estaban entre 22-38 años. Se determinó el grado de preferencia de cada producto utilizando una escala hedónica del 1 al 5, donde 1 es me disgusta mucho y 5 me gusta mucho (ver anexo 2). Los atributos sensoriales evaluados fueron color, olor, sabor, consistencia y apariencia. El costo se estableció para la producción de harina y los productos formulados, a través de los precios de la materia prima al por mayor, energía eléctrica, vida útil de los equipos, carga laboral y otros gastos fijos, para el nopal se cargaron los costos de reelección y transporte. Para la interpretación y discusión de los resultados se realizó un análisis de Varianza a los datos obtenidos durante el proceso de esta investigación.
Resultados y Discusión
Para la caracterización de la materia prima para fines de alimentación y usos industriales, es necesario conocer la composición química y elementos de interés presentes en cada una de las etapas de crecimiento del cladodio clasificado como tierno con menos de 10 día después del brote de la yema y Semi-tierno de 10 a 30 después del brote de la yema y lleno con más de 30 días después del brote de la yema (cultivo experimental IIBI 2006).

2222
Tabla No. 1 – Composición química en 100 gramos de nopal fresco de distintas edades en días después del brote visible del cladodio o penca:
Fuente: Servicios Analíticos SERAN-IIBI
Los datos de la Tabla No-1, muestran que los cladodios de nopal óptimo para cosechar y producir harina de nopal por su mayor aporte de sólidos y carbohidratos, es la etapa de semi-tierno, aunque la proteína es un elemento de interés en los alimentos, la harina presentó una textura desagradable a la vista (presencia de tejido leñoso) cuando se utilizó cladodios llenos. El tierno muestra un aporte importante en minerales de interés en sólidos y carbohidratos es el que menos aporta elementos importantes a la harina.
Proporción de Harina de Nopal
Los productos de panificación mostraron resultados organolépticos favorables hasta un 20% en la sustitución de harina de trigo por harina de Nopal. En las proporciones subsiguientes, 25% y sobre todo en la de 30%, los productos de panificación presentaron cambios organolépticos notorios como son: textura dura, presencia de filamentos, color verdoso y olor a hoja seca.
Composición Química de las Harinas
Tabla No. 2: Composición química de la harina de nopal 100%, la mezcla formulada con 80% harina trigo + 20% harina de Nopal (MHTN) y harina de trigo 100%.
Fuentes: SERAN-IIBI * datos etiqueta harina de trigo comercial, D/N datos no disponibles.La tabla No. 2 presenta datos de las harinas de nopal y trigo, información nutricional importante que nos permite tomar decisión para la formulación de las mezclas para disminuir el contenido de carbohidratos, grasa y aumentar
Compuestos
En 100 g de Nopal FrescoCladodios (Pencas)
Tierno Semi-tierno Lleno
Energía (kcal) 26.30 26.58 19.18Humedad (%) 92.61 90.90 92.48Sólidos solubles totales (%) 7.39 9.10 7.52Carbohidratos totales (%) 3.60 5.64 1.01Proteína (%) 0.95 1.78 3.74Grasas (%) 0.90 0.1 0.02Ceniza 1.94 2.58 2.75Calcio (ppm) 928 784 1113Hierro (ppm) 35.6 8.92 9.51Zinc (ppm) 14.4 5.28 4.13
Compuestos100 g de Harina
Harina de Nopal 100%
80% Harina de Trigo 20% Harina de Nopal
Harina de Trigo 100%
Energía (kcal) 266.3 345 353Grasa gr (%) 0.02 1 1.5Humedad (%) 9.0 11 9.0Carbohidratos gr (%) 58.4 73 77Proteínas gr (%) 8.1 11 9Calcio mg (%) 80.6 144 N/DHierro 40.5 1.29 N/Dzinc 3.8 1 N/DFósforo mg (%) N/D 0.01 N/DAzúcares Totales (%) N/D 0.4 N/D

23
el contenido de proteína y calcio en la mezcla. La harina de nopal presentó un valor importante de hierro que no se expresa en la mezcla. En general el valor nutricional mejoró con la mezcla de la harina de nopal con harina de trigo.
1. Al hacer la comparación del valor nutricional de productos elaborados con la fórmula 80:20 (MHTN01) Vs 100 (MHT02) el aporte mas significativo de nutrientes fue de calcio en la mezcla. Estos aportes se manifiestan en los productos analizados, al igual que es evidente la disminución de las calorías, excepto en el coconete ya que su ingrediente característico es el coco con un alto contenido de grasa.
Resultado de la Evaluación Sensorial
Grafico 1: Promedios de los parámetros Evaluados en el Análisis Sensorial de los Productos de Panificación al 20% Harina de Nopal.
Al analizar el gráfico 1 las puntuaciones obtenidas en la evaluación sensorial de los productos de panificación empleando harina de nopal mezclada con harina de trigo (MHT02) en comparación con los productos de panificación sólo con harina de trigo podemos observar una ligera preferencia por los productos sin harina de nopal, sin embargo los dos productos clasificaron en escala de 4, en los atributos evaluados: apariencia, color, olor, sabor, respectivamente.
Además de la evaluación sensorial se han hecho diversas degustaciones en ferias y eventos, donde más de 2,000 personas han consumido los productos de panificación desarrollados con un 20% harina de nopal, evidenciando la aceptación que tienen estos productos en el público en general.
Análisis de Costos
Para determinar los costos de producción de harina de nopal en la República Dominicana se realizó un balance de Materia (diagrama 1).
Diagrama No. 1: Balance de Materia para obtener 1000 Kg de Harina de Nopal

2424
Para establecer el balance de materia se realizaron más de 10 repeticiones verificando los mismos parámetros para obtener resultados transferibles. El rendimiento de harina a partir del nopal fue de 20%. El balance de energía eléctrica se determinó tomando en consideración la capacidad del equipo, el tiempo de uso, el consumo y costo de kilowatts por hora.
El costo de producción de harina de nopal es aceptable para el mercado y la proporción a utilizar en la elaboración de productos de panificación en relación a los precios del mercado nacional.
Los industriales elaboran diariamente aproximadamente 10 mil sacos de harina de trigo importado con tendencia a elevarse cuando inicia el suministro del desayuno escolar. La harina de nopal con un 20% como complemento a la harina de trigo en la elaboración de productos de panificación, aporta una reducción de costos y un nivel de independencia alimentaria.

25
Conclusión
El tiempo en días para la cosecha de los cladodios óptimos para la producción de harina o polvo es de 10 a 30 días después del brote calificado como semi-tierno, por su aporte de sólidos totales y carbohidratos.
La combinación de la mezcla de un 20% de harina de nopal con 80% de harina de trigo resultó la más manejable tecnológicamente en la elaboración de los productos de panificación, según los parámetros de manipulación de la masa, fermentación, configuración, horneado, características físicas y visuales de los productos elaborados con las diferentes proporciones de harina de nopal. En el análisis sensorial realizado con jueces no entrenados los productos de panificación evaluados (pan, bizcocho, pudín, empanada y coconete) con la fórmula MHTN 80:20, en comparación con estos mismos productos con harina trigo 100%, la puntuación fue muy similar con valores menores en los que contenían nopal, lo que tiene su explicación en que el nopal es un alimentos que no forma parte de la dieta diaria.
Los productos analizados, en los que se utilizó la fórmula MHTN 80:20, presentaron resultados satisfactorios desde el punto de vista de proceso de elaboración y valor nutricional; mostrando diferencias significativas en los componentes de calcio, cuyo parámetro mejoró al mezclar harina de trigo con nopal. Un aspecto interesante a considerar es la disponibilidad de la harina elaborada con esta cactácea producida en el país, la reducción de precio y la posibilidad de generación de empleos y ahorro de devisas.
Para la Trasferencia Tecnológica a una empresa o emprendedor interesado en la producción de harina de nopal o la elaboración de productos de panificación empleando una proporción de harina de nopal es recomendable hacer un estudio de factibilidad que permita establecer la disponibilidad, el fomento del cultivo de nopal. Así como estudios de mercado para establecer un nivel de consumo. Las grandes extensiones de terrenos áridos de baja productividad en el país puede ser una alternativa utilizada para promover el cultivo de nopal como potencial para producir harina local y otros usos en la alimentación; dentro de la política estatal para crear empleos, fomentar la agroindustria, innovar con nuevos productos en el mercado nacional e internacional, enfrentar la crisis alimentaria y contribuir a la seguridad alimentaria de la Isla.
Agradecimiento
Los autores expresan su agradecimiento a todo el personal del IIBI que colaboraron para llevar a cabo este proyecto, en especial a la Dra. Bernarda Castillo por su iniciativa decidida a gestionar los recursos y apoyo incondicional del proyecto, al Ing. Andrés Guerrero por su colaboración en el análisis estadístico e interpretación de los resultados, al Ing. Félix Rivas por su continuo apoyo para la redacción de la propuesta, al Lic. Alejandro Tabar y el personal de Servicios Analísticos por gestionar y apoyar las actividades programadas para las analíticas y trabajo de campo, a la Lic. Gloria Santos y todo el equipo de administración incluyendo el personal de mantenimiento con ellos fue posible procesar y evaluar los productos. A la FAO en nombre del Sr. Antonio Morales, Héctor Mata por apoyar la Industrialización del Nopal en Neyba que sirvió de base a esta investigación, en particular al Ing. Andrés Gómez del Programa Especial para la Seguridad Alimentaria, PESA. Yaquelín Silfa y toda la directiva y membrecía del Bloque de Organizaciones de Neyba BONE.
Al Sr. Francisco Pollok de Panificadora Moderna, la Lic. Nelly Berigüete y Alcides de la Panadería el Metro con todo su personal por permitirnos elaborar los productos en sus instalaciones.
La realización de esta investigación fue posible gracias al aporte del Fondo Concursable de Investigacion (FONDOCYT) Del Ministerio de Educación Superior, Ciencia y Tecnología de la República Dominicana (MESCYT) quien proveyó parte del financiamiento.

2626
Referencias
Anzaldúa A. La Evaluación sensorial de los alimentos en la teoría y la práctica, editorial ACRIBIA, S.A, Zaragoza, España, 1994.
Sáenz C. Utilización Agroindustrial del Nopal. Boletín de Servicios Agrícolas de la FAO, 162, Roma, 2006.Cerezal, P. y Duarte, G. Algunas características de tunas (Opuntia ficus-índica (L.) Miller) cosechadas en el altiplano andino de la 2da Región de Chile. Disponible en línea: http://www.scielo.org.ve/scielo.php
Consejo de Promoción de Nopal y Tuna (2009). Usos y Aplicaciones del Nopal. México: Autor. Disponible en: http://www.cpnt.org.mx/pdf/usosApliNopal.pdf
El Nopal., disponible en línea: http://www.giga.com/mag/Tratado_Nopal.htm. El nopal y sus propiedades., disponible en línea: http://www.alimentacion-sana.com.ar/informaciones/novedades/nopal.htm; Nopal, manjar saludable. Disponible en línea: http://www.alimentacion-sana.com.ar/informaciones/novedades/nopal.htm
Memorias “VII Simposium Nacional: Taller de “Producción y Aprovechamiento del Nopal al Nordeste de México”. Facultad de Agronomía, Universidad Autónoma de Nuevo León., del 24 al 25 de octubre de 2008. Mina, Nuevo León, México. 179 p.
Ríos j. Y Quintana v. (2004) MANEJO GENERAL DEL CULTIVO DEL NOPAL. México. 81 p. Disponible en línea:http://www.sra.gob.mx/internet/informacion_general/programas/fondo_tierras/manuales/Manejo_general_cultivo_Nopal.pdf
Sáenz, C., Sepúlveda E., Pak N., Vallejos X. (2002) Uso de fibra dietética de nopal en la formulación de un polvo para flan. V. 52 n. 4 Caracas.
Sandoval Iturbide Antelma, Navarro Cruz Addí Rhode, Ávila Sosa Sánchez Raúl, Lazcano Hernández Martín, Dávila Márquez Rosa María. Elaboración de un producto de panificación utilizando harina de nopal viejo o pie de cría (Opuntia ficus indica). Roma, 2006
Anexos 1: Fotografías A. Cultivo experimental de nopal (tuna, alquitira) en el IIBI
Plantación de nopal con 3 meses, de cultivada y se está evaluando el tiempo del cladodio desde el brote hasta la cosecha para clasificar tierno, semi- tierno y lleno
Esta es la misma plantación 2 años, después, donde fueron cosechados los cladodios o materia prima para producir la harina de nopal.

27
B. Esta son los parámetros visuales de las harinas y su aspecto al ser mezclada para la elaboración o formulaciones de los productos de panificación.
Harina de Nopal 100% 80% Harina de Trigo mezclada
con 20% Harina de Nopal (MHTN01)
Harina de Trigo 100% Comercial
Anexo 2. Productos de panificación que vende la panadería El Metro, elaborado con la formulación MHTN01 y los productos de panificación con solo harina de trigo, como acostumbran los cliente a comprar a diario.
Empanadas con Mermelda de Nopal
Pan Bizcochitos Coconete Pudin

2828
Formulario utilizado para la evaluación sensorial (escala hedónica de 5 puntos).
Evaluación Sensorial Productos con Harina de Nopal
Fecha:_____________
Generalidades del juez:
Nacionalidad ____________________ Edad _______ Sexo ______
Instrucciones: 1) Indique su juicio de la característica y escriba la escala a la que califica el producto según el atributo en cada muestra.
2) Marque con una (X) si existe diferencia entre las muestras presentadas: SI hay diferencia NO hay diferencia
Comente la Respuesta 2: _________________________________________
Leyenda: “MHTN01” Mezcla de un 20% Harina de Nopal y 80% harina de trigo “MHTC02” Muestra de 100% Harina de Trigo comercial importado.
80% Harina de Trigo 20% Harina de Nopal
Pan Pudín Bizcochitos Coconete Empanadas
Escala CaracterísticasAtributos
Muestras1 Me disgusta mucho MHTN01 MHTC022 Me disgusta Apariencia3 Ni me disgusta, ni me gusta Color4 Me gusta Olor5 Me gusta mucho Sabor

29
Bioprospección de Plantas Endémicas Dominicanas: Aislamiento e Identificación de compuestos con posible acción Anticancer Presentes en Agave Antillarum, Agave Brevipetala y Agave Brevispina.
Susana De Jesús1, S. Bhave2, S. Clinton2, K. Riedl3 y S. Schwartz31 Biotecnología Farmacéutica de Productos Naturales, Instituto de Innovación en Biotecnología e Industria (IIBI), Santo Domingo, RD.2 Department of internal medicine, The Ohio State University, Columbus, OH, USA.3 Department of food science and technology, The Ohio State University, Columbus, OH, USA.
Palabras Claves: Bioprospección, plantas endémicas, aislamiento, identificación, Agave, spp.
Resumen
Las especies del género Agave son plantas tropicales usadas por los nativos de la isla Hispaniola con propósitos medicinales. Estudios recientes en una especie endémica de este género (Agave intermixta) mostraron que ésta posee actividad antiinflamatoria y citotóxica. El propósito de este trabajo fue investigar la potencial actividad anticáncer de diferentes especies del género Agave, e identificar y aislar compuestos con esta actividad. Los extractos crudos de Agave sp y Agave antillarum fueron probados contra 29 microorganismos, y sus fracciones fueron probadas contra el patógeno Cándida albicans. Luego la habilidad de inhibir en crecimiento de la línea celular cancerígena de próstata PC3, fue evaluada con los extractos crudos de A. antillarum, A. brevispina, A. brevispina??, A. sp, y A. espina grande, la fracción de A. antillarum que contenía saponinas, y los extractos hidrolizados. Los extractos crudos de Agave sp y Agave antillarum fueron activo contra Cándida albicans y no fueron activos contra el resto de las bacterias. Las fracciones de los extractos, que contenían saponinas, fueron también activas contra C. albicans. Todos los extractos crudos exhibieron inhibición de crecimiento en una manera dosis-dependiente para PC3. Los extractos hidrolizados de los Agave mostraron ser activos contra PC3, pero la actividad fue más baja que la mostrada por el extracto crudo, excepto por A. antillarum, para el cual su extracto hidrolizado fue más activo. La fracción de A. antillarum que contenía saponinas mostró inhibir el crecimiento de PC3, sin embargo, la actividad fue más baja que la mostrada por el extracto crudo. Esto pone en evidencia que otros compuestos cuentan para la actividad y que las saponinas no son las responsables por toda la actividad observada. El estudio de LC MS mostró la presencia de dioscin, diosgenin, hecogenin, tigogenin, manogenin, y otros iones no especificados en los extractos de los Agave.

3030
Keywords: Bioprospecting, endemic plants, isolation, identification, Agave spp.
Summary
Species of the genus Agave are tropical plants used by the natives of the island of Hispaniola for medicinal purposes. Recent studies in an endemic species of this genus (Agave Intermixt) showed that it possesses antiinflammatory and cytotoxic activity. The purpose of this study was to investigate the potential anticancer activity of different species of the genus Agave, and identify and isolate compounds with this activity. Crude extracts of Agave and Agave sp antillarum were tested against 29 microorganisms, and its fractions were tested against the pathogen Candida albicans. Then the ability to inhibit growth of prostate cancer cell line PC3 was evaluated with extracts of A. antillarum, A. brevispina, A. brevispina?, A. sp, and A. large thorn, the fraction of A. antillarum containing saponins, and hydrolyzed extracts. Crude extracts of Agave and Agave sp antillarum were active against Candida albicans and were not active against other bacteria. The fractions of the extracts containing saponins were also active against C. albicans. All crude extracts showed inhibition of growth in a dose-dependent manner for PC3. The hydrolyzed extracts of Agave shown to be active against PC3, but the activity was lower than that shown by the crude extract, except for A. antillarum, for which the hydrolyzed extract was more active. The fraction of A. antillarum containing saponins was shown to inhibit the growth of PC3, however, activity was lower than that shown by the crude extract. This shows that other compounds for activity and have the saponins are responsible for all activity observed. The study of LC MS showed the presence of dioscin, diosgenin, hecogenin, tigogenin, manogenin, and other unspecified ions in the extracts of Agave.
Introducción
Las especies del género Agave son comúnmente conocidas en la República Dominicana por el nombre popular de ‘Maguey de Bestia’ o ‘Maguey’. En la isla Hipaniola, los usos medicinales de estas especies incluyen: laxativos (raíces), cura de llagas de animales (hojas), astringente (Liogier, 2000), inflamaciones, tumores, infecciones, artritis (Quílez et al., 2004), y remoción de fibroides uterinos (Ososki et al., 2002). Estos usos medicinales ponen en evidencia su potencial como fuente de nuevas moléculas bioactivas. En general, los Agave contienen fibras, que en algunas especies son de valor comercial (aunque las especies nativas de la Hispaniola no son explotadas para este propósito), y saponinas.Las saponinas y sus agliconas son metabolitos secundarios complejos las cuales exhiben un gran número de actividad biológica incluyendo anti-cáncer, activación de p53, apoptosis (Corbiere et al., 2003), antiinflamatoria (García et al., 2000), antifúngica y antiviral, haemolítica (Takechi and Tanaka, 1995), hipocolesterolémica, permeabilizante de membrana, inmunoestimulante (Francis et al., 2002), e ingesta de alimentos y crecimiento en animales (Makkar and Becker, 1996). Muchos estudios muestran que la saponina dioscin, así como su aglicona diosgenin poseen actividad anticáncer, e inducen apoptosis (Corbiere et al., 2003; Cai et al., 2002; Wang et al., 2001). También se ha llegado a la conclusión de que la estructura del espirotenol podría jugar un papel muy significante en la actividad. Trouillas et al., 2005, comparó la relación estructura-función de diferentes saponinas muy relacionadas. Con estos estudios, se llegó a la conclusión de que su actividad fue parcialmente atribuida a la presencia del 5,6 doble enlace, y la hetero azúcar.
El propósito de este estudio fue evaluar la actividad citotóxica de diferentes especies del género Agave, e identificar y aislar compuestos con esta actividad.

31
Materiales y métodos
Material vegetal. Las hojas de Agave sp fueron recolectadas en Altos de Chavón, La Romana, República Dominicana. Las hojas de Agave antillarum Descourt, fueron recolectadas en la provincial de Azua, República Dominicana, y fueron identificadas por el botánico A. Veloz. Las hojas de Agave brevispina Trel. Agave brevispina??, y Agave espina grande fueron recolectadas en El Aguacate, Sierra de Bahoruco, República Dominicana. Estas fueron identificadas por el botánico Dr. J. Salazar, en el Jardín botánico nacional de Santo Domingo. Voucher de los especimenes fueron depositados. Sólo A. antillarum y A. brevispina han sido bien caracterizadas.
Preparación de los extractos. Las hojas frescas de las plantas fueron cortadas en trozos, y los compuestos fueron extraídos con etanol utilizando un soxhlet. Los extractos fueron luego concentrados in vacuou utilizando un rotavapor standard de diosgenin (Sigma Chemical Co; St. Louis, MO).
Ensayo antimicrobiano. Este ensayo fue realizado en el laboratorio de microbiología del IIBI. Para esto utilizamos 29 microorganismos patógenos: Bacillus cereus (2 strains), Bacillus subtillis, Staphylococcus aureus (4 strains), Proteus mirabilis (2 strains), Pseudomonas aeruginosa (2 strains), Enterococcus faecalis (2 strains), Enterococcus faecium, Leuconostoc mesenteriae, Salmonella arizona, Salmonela dervi, Salmonella sp (2 strains), Salmonella abaetetuba, Escherichia coli (3 strains), Citrobacter freundii, Klebsiella pneumoniae, Enterobacter cloacae, Listeria monocytogenes, Proteus vulgaris, y Candida albicans, usando el método agar difusión. Brevemente, un disco de papel filtro de 0.5mm fue impregnado con el extracto, y se dejó secar. Una vez seco el extracto, el disco se colocó en la caja petri que contenía medio sólido y el microorganismo. Los platos fueron incubados a 37 ºC y observados 24 h más tarde.
Fraccionamiento de los extractos crudos. Después de realizar el ensayo antimicrobiano, el fraccionamiento de los extractos se realizó para localizar la polaridad de los compuestos activos. Para esto fueron utilizados, una columna pyrex y sílica gel, y un sistema de solvente en gradiente desde 95% hexano: 5% acetona a 100% acetona. La proporción extracto: sílica gel fue 1:10 respectivamente. Se obtuvieron seis fracciones.
Hidrólisis de los extractos. La hidrólisis de los extractos fue realizada usando una versión modificada de la propuesta por Taylor et al., 2000. Se tomaron 10 ml de los extractos crudos de los Agave fueron evaporados hasta que quedaron 4 ml. Luego fueron añadidos, 3.2 ml de una mezcla de 70% 2-propanol con 30% H2SO4 1M. La mezcla se dejó a 100 °C por 2 horas. Después de las 2 horas, se dejó refrescar y fueron añadidos 4.8 ml de H2O. Las agliconas fueron extraídas con 3.2 ml de MTBE, tres veces (para cada extracción, la mezcla se centrifugó por 10 min a 10,000 rpm), y luego se neutralizado con 2.4 ml de NaOH 1M, dos veces. Los compuestos extraídos con MTBE fueron luego lavados con 2.4 ml de H2O, y secados por MgSO4 usando lana de vidrio. Luego, los extractos fueron secados bajo N2 comprimido, y almacenado a -20 °C hasta análisis.
Para la hidrólisis de la fracción conteniendo saponinas, se hizo una modificación. Se mezcló 15 ml de la fracción, con una solución de H2SO4 2.88 M en 2-propanol. La mezcla se calentó por 2 horas a 100 °C. Después de este tiempo y cuando la mezcla se refrescó, se le añadieron 37.5 ml de H2O y se extrajo la mezcla con 25 ml de MTBE, tres veces. Los compuestos extraídos con MTBE fueron luego neutralizados con 12.5 ml de NaOH 1M, dos veces, y luego lavados con 12.5 ml of H2O, secados con MgSO4 usando lana de vidrio, y finalmente secados bajo N2 comprimido.
Crecimiento In Vitro de células cancerígenas de próstata (PC3). Las células humanas cancerígenas de próstata (PC3), fueron mantenidas como cultivo monocapa en RPMI suplementado con 10% de suero bovino fetal, 2% L-glutamina, y 0.2% de una mezcla de penicilina/estreptomicina, en una atmósfera saturada de humedad 95%

3232
aire/5% CO2 a 37°C. Los estudios fueron conducidos con 6.5 x 104 células por platos de 10 cm2. Las células fueron tratadas con los extractos crudos, fracción de extracto crudo conteniendo saponinas, y los extractos parcialmente hidrolizados. Para disolver los extractos hidrolizados, se utilizó DMSO (para una concentración final de ≤ 0.1%), y fueron encubados por 48 horas. En los ensayos con los extractos hidrolizados, se utilizó DMSO como control. El conteo de células fue hecho utilizando un hemocitómetro. Todos los ensayos fueron realizados en duplicado.
Análisis en HPLC. La separación cromática de los extractos de los Agave, fue realizada usando una columna de fase reversa Varian Inertsil ODS-3, 250 x 4.6 mm, 5μ, C18, en un HPLC Waters 2695. Los extractos hidrolizados secos fueron reconstituidos con volúmenes apropiados de metanol: H2O (9:1) respectivamente. Muestras de 50μl fueron inyectadas con un método socrático (90% methanol: 10% H2O con 0.1% de ácido fórmico), con un flujo de 1 ml/min por 40 minutos, y la temperatura de la columna a 30 °C. Los picos fueron detectados a 202 nm usando un detector Waters 2996 con arreglo de foto-diodo.
Análisis LC MS. Para este experimento, fue empleado un método socrático compuesto de 0.1% de acido fórmico en H2O (A), y 0.1% de acido fórmico en metanol (B). El método fue corrido con 90% B en una columna de fase reversa Varian Inertsil ODS-3, 250 x 4.6 mm, 5μ, C18, por 40 minutos a un flujo de 1 ml/min. Aproximadamente un décimo del eluyente del HPLC fue enviado a la fuente electro-spray de un espectrómetro de masa QToF Premier (Micromass, Beverly, MA), vía un “flow splitter”. El voltaje capilar se mantuvo a 3.2 kV, y el cono de muestra estuvo a 35V. La temperatura del “source block” fue 100°C, y el gas de disolución fue enviado a 400 L/h y 400°C. El espectrómetro de masa fue calibrado, para exactitud de masa, con formato de sodio, y la prueba “lockspray” conteniendo leucine enkephalin como “lockmass acquiring” cada 10 segundo durante la corrida analítica. Un estándar de diosgenin fue utilizado para anticipar la presencia de este en los extractos.
Resultados
Los extractos crudos de Agave sp y Agave antillarum fueron activos contra Candida albicans produciendo halos de inhibición de 2.7 y 2.38 cm respectivamente (Tabla 1). Estos extractos no mostraron actividad en el resto de las bacterias. Las fracciones de estos extractos, que contenían saponinas, fueron también activas contra C. albicans (Fig. 1 y Tabla 1). Estos resultados confirman la propiedad antifúngica de las saponinas previamente reportada (Kumar et al., 2006; Sautour et al.,2007; Xie et al., 2006).
Todos los extractos crudos exhibieron inhibición de crecimiento en un formato dosis-dependiente para las células PC3 (Fig. 2). Los extractos hidrolizados mostraron ser activos contra PC3, pero la actividad fue menor que la mostrada por los extractos crudos, exepto para A. antillarum (Fig. 3). La fracción de A. antillarum que contenía las saponinas mostró inhibición del crecimiento de PC3, sin embargo, la actividad fue menor que la mostrada por el extracto crudo (Fig.4). Esto pone en evidencia que otros compuestos cuentan para la actividad y que las saponinas no son las responsables de toda la actividad observada. Los picos intensos del MS fueron fácilmente fraccionados en el espectrómetro de masa para liberar iones consistentes con muchos sapogenoles previamente identificados incluyendo dioscin (577 m/z), diosgenin (415 m/z), hecogenin (431 m/z), togogenin (417 m/z), manogenin (447 m/z), y otros iones con (433 y 413) m/z (Fig. 6). Los espectros UV de todos los picos mostraron su lambda máxima alrededor de 210 nm típico de saponinas.
Los extractos crudos de Agave antillarum, Agave brevispina, y Agave sp, mostraron ser muy activos contra PC3. Los datos obtenidos muestran que el ID50 para estos extractos fue más alto que los propuestos por el “Protocols of the National Cancer Institute of USA” (Geran et al., 1972) para extractos crudos de plantas (< 30μg/ml). Estos resultados preliminares ponen en evidencia que las plantas ofrecen una gran fuente de compuestos que pueden conllevar al desarrollo de nuevos medicamentos para enfermedades con opciones de tratamientos limitados.

33
Fig. 1. Actividad mostrada por los extractos crudos (A y B), y fracciones de los extractos crudos (C y D), contra Cándida albicans. Este ensayo fue realizado usando el método Agar Difusión, donde un disco de 0.5 mm fue impregnado con el extracto. Cuando el extracto se había secado, este fue puesto en una caja Petri que contenía el medio sólido inoculado con el microorganismo. Los platos fueron incubados a 37 ºC y observados 24 h más tarde. Los extractos crudos fueron probados contra 29 microorganismos y las fracciones contra C. albicans.
Tabla 1 Efecto de los extractos crudos de Agave antillarum y Agave sp contra Candida albicans. Halo de inhibición medido en cm.
Extracto C. albicansAgave antillarum Halo (cm)Crudo 2.38Fracción 1 *Fracción 2 *Fracción 3 1.1Fracción 4 1.5Fracción 5 2.2Fracción 6 *Agave sp Halo (cm)crudo 2.7Fracción 1 *Fracción 2 *Fracción 3 *Fracción 4 *Fracción 5 2Fracción 6 1.6*= no inhibition
Dose-dependent inhibition for PC3Experiment 4 (48 h)
0
20
40
60
80
100
120
0 200 400 600 800 1000 1200
Crude extract (ug/ml)
Cel
l nu
mb
er (
% c
on
tro
l)
Aa
Ab
Ab??
Asp
Aeg
Fig. 2. Inhibición dosis-dependiente In Vitro del número de células para PC3 por los extractos crudos de Agave antillarum (Aa), Agave brevispina (Ab), Agave brevispina?? (Ab??), Agave sp (Asp), y Agave espina grande (Aeg). Las células fueron mantenidas
como monocapas y tratadas por 48 h, y el número de células fue determinado por conteo directo.

3434
Dose-dependent inhibition for PC3Experiment 5
0
20
40
60
80
100
120
140
0 100 200 300 400 500 600
Hydrolized crude extract (ug/ml)
Cell n
um
ber
(% c
on
tro
l)
Ab??
Asp
Aeg
Dose-dependent inhibition for PC3Experiments 3 (48 h)
0
20
40
60
80
100
120
140
0 100 200 300 400 500 600
Hydrolized crude extract (ug/ml)
Ce
ll n
um
be
r (%
co
ntr
ol)
Aa
Ab
Fig. 3. Inhibición dosis-dependiente In Vitro del número de células para PC3 por los extractos hidrolizados de Agave brevispina?? (Ab??), Agave sp (Asp), Agave espina grande (Aeg) (A), Agave antillarum (Aa) y Agave brevispina (Ab) (B). Las células fueron mantenidas como monocapas y tratadas por 48 h, y el número de células fue determinado por conteo directo.
Dose-dependent inhibition for PC3
0
20
40
60
80
100
120
0 500 1000 1500 2000 2500
Saponin fraction (ug/ml)
Cel
l num
ber (
% c
ontr
ol)
Saponin fraction (SF)
Dose-dependent inhibition for PC3
0
20
40
60
80
100
120
140
160
180
0 100 200 300 400 500 600
Hydrolized saponin fraction (ug/ml)
Cel
l num
ber (
% c
ontr
ol)
Sapogenins of AaSf
Fig. 4. Inhibición dosis-dependiente In Vitro del número de células para PC3 por la fracción de Agave antillarum (Aa) conteniendo saponinas (A), y la fracción hidrolizada de Agave antillarum (Sf) que contenía las saponinas (B). Las células fueron mantenidas como monocapas y tratadas por 48 h, y el número de células fue determinado por conteo directo.
A B
A B

35
Fig 5. Análisis de HPLC de los extractos crudos hidrolizados de Agave antillarum (Aa), Agave brevispina (Ab), Agave sp (Asp), y la fracción hidrolizada de A. antillarum que contenía saponinas (H sf de Aa). La separación se llevó a cavo utilizando una columna de fase reversa Varian Inertsil ODS-3, 250 x 4.6 mm, 5μ, C18, en un HPLC Waters 2695. Las muestras fueron inyectadas con un método isocrático (90% metanol: 10% H2O con 0.1% de ácido fórmico), con un flujo de 1 ml/min por 40 minutos, y la temperatura de la columna a 30 °C. Los picos fueron detectados a 202 nm usando un detector Waters 2996 con arreglo de foto-diodo. Esta separación muestra un pico a los 5 minutos que pertenece a dioscin.

3636
Fig. 6. Compuestos identificados en los extractos crudos de Agave antillarum y Agave sp. Dioscin (A), una saponina que ha mostrado poseer efectos citotóxicos, e induce apoptosis. Diosgenin (B), la aglicona de dioscin, ha mostrado inducir apoptosis, inhibir la proliferación de diferentes líneas tumorales, aumentar la activación de p53, y poseer actividad antiinflamatoria. Hecogenin (C) y tigogenin (D), son agliconas saturadas que han mostrado poseer una moderada actividad citotóxica.

37
Referencias
Borris, R. P. (1996). Natural products research: perspectives from a major pharmaceutical company. Journal of Ethnopharmacology 51: 29-38.
Cai, J., M. Liu, Z. Wang, and Y. Ju. (2002). Apoptosis Iduced by dioscin in Hela cells. Biol. Pharm. Bull. 25 (2): 193-196.
Corbiere, C., B. Liagre, A. Bianchi, K. Bordji, M. Dauça, P. Netter, and J-L Beneytout. (2003). Different contribution of apoptosis to the antiproliferative effects of diosgenin and other plant steroids, hecogenin and tigogenin, and human 1547 osteosarcoma cells. International Journal of Oncology 22: 899-905.
Finlay, G.J., B.C. Baguley and W. R. Wilson. (1984). A semi-automated microculture method for investigating growth inhibitory effects of cytotoxic compounds on exponentially growing carcinoma cells. Anal. Biochem., 139: 272-277.
Francis G, Kerem Z, Harinder MPS, Becker K. (2002). The biological action of saponins in animal systems: a review. Br J Nutr 88:587–605.
García, M. D., A. M. Quilez, M. T. Saenz, M. E. Martinez-Dominguez, y R. de la Puerta. (2000). Anti-inflammatory activity of Agave intermixta Trel. and Cissus sicyoides L., species used in the Caribbean traditional medicine. Journal of Ethnopharmacology 71: 395-400.
Geran, P. I., N. H. Greenberg, N. M. McDonald, A. M. Schumacher, and B. J. Abbot. (1972). Protocols for screening chemicals agents and natural products against animal tumors and other biological systems. Cancer Chemother. Rep 3:17.
Gupta, M. P. (1995). Plantas medicinales iberoamericanas. Editoral presencia Ltda, Santafe de Bogota, Colombia, xxiii, p 617.
Hostettmann, K., y J.-L. Wolfender. (1997). The search for biologically active secondary metabolites. Pestic. Sci 51: 471-482.
Kumar, R., A. K. Chaturvedi, P. K. Shukla, and V. Lakshmi. (2006). Antifungal activity in triterpene glycosides from the sea cucumber Actinopyga lecanora. Bioorg. Med. Chem. Lett. (Dec 21).
Liogier, H. A. (2000). Diccionario botánico de nombres vulgares de la Española. Segunda edicción. Editora Corripio, c. por a. República Dominicana, p 290.
Loizzo, M. R., R. Tundis, G. A. Statti, F. Menichini, and P. J. Houghton. (2005). In-vitro antiproliferativr effects on human tumor cell lines of extracts and jacaranone from Senecio leucanthemifolius Poiret. Journal of pharmacy and pharmacology 57: 897-901.
Makkar HPS, Becker K. (1996). Effect of Quillaja saponins on in vitro rumen fermentation. In: Waller GR, Yamasaki Y (eds) Saponinins used in food and agriculture. Plenum, New York, pp 377–386.
Ososki, A. L., P. Lohr, M. Reiff, M. J. Balick, F. Kronenberg, A. Fung-Berman, y B. O’Connor. (2002). Ethnobotanical

3838
literature survey of medicinal plants in the Dominican Republic used for women’s health conditions. Journal of Ethnopharmacology 79: 285-298.
Phillipson, J. D. (2001). Phytochemistry and medicinal plants. Phytochemistry 56: 237-243.
Quílez, A. M., M. T. Sáenz, M. D. Garcia, y R. de la Puerta. (2004). Phytochemical analisis and anti-allergic study of Agave intermixta Trel. and Cissus sicyoides L. Journal of Pharmacy and Pharmacology 56: 1185-1189.
Quinn, M. J., A. d’Onofrio, B. Møller, R. Black, C. Martinez-Garcia, H. Møller, M. Rahu, C. Robertson, L. J. Schouten, C. La Vecchia & P. Boyle. (2003). Cancer mortality trends in the EU and acceding countries up to 2015. Annals of Oncology, 14: 1148- 1152.
Saenz, M. T., M. D. Garcia, A. Quilez, y M. C. Ahumada. (2000). Cytotoxic activity of Agave intermixta L. (Agavaceae) and Cissus sicyoides L. (Vitaceae). Phytotherapy Research 14: 552-554.
Sautour, M., T. Miyamoto, and M. A. Lacaille-Dubois. (2007). Steroidal saponins from Asparagus actifolius. Phytochemistry (Apr 19).
Slater, T.F., B. Sawyer and U. Strauli. (1963). Studies on succinate-tetrazolium reductase systems. III Points of coupling of four different tetrazolium salts. Biochem. Biophys. Acta.77: 383-393.
Takechi M, Tanaka Y. (1995). Haemolytic and time course differences between steroid and triterpenoid saponins. Planta Med 61:76–77.
Taylor, W. G., J. L. Elder, P. R. Chang, and K. W. Richards. (2000). Microdetermination of diosgenin from Fenugreek (Trigonella foenum-graecum) seeds. J. Agric. Food Chem. 48: 5206-5210.
Trouillas, P., C. Corbiere, B. Liagre, J-L Duroux, and J-L Beneytout. (2005). Structure-function relationship for saponin effects on cell cycle arrest and apoptosis in the himan 1547 osteosarcoma cells: a molecular modeling approach of natural molecules structurally related to diosgenin. Bioorganic & Medicinal Chemistry 13: 1141-1149.
Wang, Z., J. Zhou, Y. Ju, H. Zhang, M. Liu, and X. Li. (2001). Effects of two saponins extracted from the Polygonatum zanlanscianense pamp on the human leukemia (HL-60) cells. Biol. Pharm. Bull. 24(2): 159-162.
Watts, M.E., I.J. Roberts, and M. Woodcock. A comparison of colorimetric and clonogenic assays for hypoxic-specific toxins with hamster and human cells. Int. J. Radiation Oncology Biol. Phys.16: 939-942
Xie, B. B., H. Y. Liu, W. Ni, C. X. Chen, Y. Lü, L. Wu, and Q. T. Zheng. (2006). Five new steroidal compounds from Ypsilandra thibetica. Chem. Biodivers. 3(11): 1211-8.
Agredecimientos
La realización de esta investigación fue posible gracias al aporte del Fondo Concursable de Investigacion (FONDOCYT) Del Ministerio de Educación Superior, Ciencia y Tecnología de la República Dominicana (MESCYT) quien proveyó parte del financiamiento.

39
Análisis de la Variabilidad Genética de Dos Especies de Pecaríes (Tayassu) en Colombia, Bolivia, Ecuador y Perú Utilizando Marcadores Microsatélites.Carlos Fernando Vergara Castillo, Manuel Ruiz García
Palabras clave: Genética de poblaciones, Marcadores moleculares, Microsatelites, Pecarí de collar, Pecarí de labio blanco, Heterogeneidad genética, acervo genético.
Resumen
En el presente trabajo se aportaron pruebas feacientes de la importancia y el poder que los análisis genéticos poblacionales en conjunción con procedimientos moleculares que involucren metodologías con marcadores microsatélites, tienen en la obtención de conocimiento acerca de la historia natural de dos especies de pecarí. Los procedimientos moleculares se llevaron a cabo analizando 75 muestras de pecarí de collar y 71 muestras de pecarí de labio blanco, pertenecientes a los países de Colombia, Perú, Bolivia y Ecuador, utilizando marcadores tetranucleótidos para cerdo doméstico (Sus scrofa).
Los análisis moleculares ofrecieron un importante poder de discriminación para asignar a los animales estudiados en sus respectivas localidades geográficas. La heterogeneidad genética encontrada en ambas especies estudiadas permitió considerar a las poblaciones de T. pecari y T. tajacu procedentes de los diferentes países de origen como poblaciones con un grado de diferenciación genética apreciable.
Introducción
Sudamérica, Centroamérica, las tierras bajas de México y las Antillas pueden ser consideradas como una sola región zoogeográfica, conocida como región neotropical. La fauna se caracteriza por la variedad y carencia de afinidad con la fauna de otros continentes, incluso con la del norte de Estados Unidos y la meseta mexicana. En ella se han desarrollado familias de mamíferos que no existen en ninguna otra región del mundo.
La biodiversidad entendida como capital biológico para el desarrollo de los países sustenta un fino equilibrio, entre las áreas de producción y ecología del cual depende la supervivencia de muchas especies sometidas a explotación. La pérdida de dicha diversidad constituye una de las crisis más apremiantes del mundo aumentando la preocupación sobre el estado de los recursos biológicos (www.redlist.org, 2002). Actualmente se realizan esfuerzos conjuntos para el estudio de la diversidad biológica que incluye recursos genéticos, especies y ecosistemas. Su conocimiento no sólo busca la conservación y uso sostenible, sino la distribución justa y equitativa que resulta del uso de todos los recursos genéticos, (Krieger y Ochoa, 2002).

4040
Colombia ocupa un lugar clave y estratégico formando un puente entre Mesoamérica y Suramérica incluyendo los ríos Amazonas y Orinoco. Tiene el máximo de hábitats desde desiertos y pluvioselvas, de los llanos y sabanas subtropicales a las cimas mas elevadas de las montañas. Debido a esto, Colombia por su gran extensión y al variado relieve, presenta ambientes desde el desértico hasta los pluviales en una serie ecológica húmeda y desde los ecuatoriales y tropicales hasta los de superparamo en el gradiente altitudinal de nuestras cordilleras, lo anterior unido a su ubicación geográfica da como resultado que su flora y fauna sean muy variadas y complejas, (Estrada, 1980).
El suborden de mamíferos Suiformes, contiene tres familias: Hippopotamidae (hipopótamos), Tayassuidae (pecaríes) y Suidae (verdaderos cerdos). Antes de su introducción al nuevo mundo por los humanos, las especies de la familia Suidae eran pertenecientes exclusivamente a un taxón del viejo mundo, como lo son los hipopótamos (Theimer y Keim, 1998).
El Pecarí o Saíno, nombre común de ciertas especies de cerdos pequeños salvajes similares al jabalí, constituyen la familia de los Tayasuidos, incluida en el orden de los Artiodáctilos. Existen tres especies de pecaríes: El pecarí de collar se clasifica como Tayassu tajacu, el pecarí de labio blanco como Tayassu pecarí; por último, la tercera especie Catagonus wagneri. Viven desde el norte y centro de América hasta Argentina. Son animales de costumbres nocturnas y de dieta omnívora (Theimer y Keim, 1998). Con el descubrimiento de un tercer pecarí en 1975, otros autores como Weyzel (1975), Wetzel (1977) citado por Theimer y Keim, (1998), argumentaron que el pecarí de labio blanco y el pecarí de collar eran lo suficientemente similares entre sí, comparados con el nuevo pecarí descubierto, por esto ubicaron al pecarí de collar y al de labio blanco en un solo género llamado Tayassu. El reciente pecarí descubierto, fue relacionado inmediatamente con fósiles pertenecientes a Sudamérica y fue clasificado perteneciente al género Catagonus.
El pecarí de collar, también llamado tsainú, tiene, como su nombre indica, una banda o collar de pelo blanco alrededor del cuello y su área de distribución es muy amplia. Vive en grupos pequeños de entre dos y cincuenta individuos y se alimenta de insectos, raíces, frutas y reptiles. El tipo de hábitat que ocupa es muy variado; en algunos lugares se sabe que viven en cavidades naturales o en madrigueras construidas por otros animales. El pecarí de labio blanco es más grande que la especie anterior; su distribución es más restringida, habita en la selva tropical y vive en piaras de hasta cien individuos. La tercera especie se diferencia de las anteriores en que tiene las patas, las orejas y el hocico más grande; fue descrita en un principio a partir de un resto fósil y se pensaba que estaba extinta desde hace once mil años, pero se descubrió un individuo vivo en Argentina y se registró por primera vez en la literatura científica en el año de 1975.
El pecarí de labio blanco, prefiere los hábitats húmedos en los bajiales, mientras que el pecarí de collar prefiere los hábitats más secos en las tierras altas. Ambas especies están bien adaptadas a los bosques de tierra firme, mientras que el pecarí de labio blanco puede usar los bosques de várzea mejor que el pecarí de collar debido a sus largos desplazamientos. Estas diferencias en el hábitat, está reflejada en la dieta de los pecaríes en las estaciones inundables y no inundables (Bodmer, 1997).
El cariotipo del pecarí de labio blanco consta de un número diploide de 26 cromosomas. Análisis de bandas G, de los trece pares de cromosomas de esta especie sugieren que tres de estos cromosomas evolucionaron por eventos de fusión Robertsoniana de pares de cromosomas ancestrales Giannoni y Ferrari, (1976). Con respecto al cromosoma 6, se concluyó que fue el resultado de un evento de fusión entre un par de cromosomas ancestrales similares a los cromosomas 9 y 14 de Tayassu tajacu.

41
El cromosoma 8, de cromosomas ancestrales similares al 8 y al 10 en Tayassu tajacu y el cromosoma 2 de cromosomas ancestrales similares a los cromosomas 2 y 3 del cerdo doméstico (Sus scrofa), (Giannoni y Ferrari, 1976). Hibridizaciones en cautiverio entre Tayassu pecarí y Tayassu tajacu, han sido reportadas en tres ocasiones en el Zoológico de Londres (Zuckerman, 1953) y el zoológico de Manaus (Sowls, 1984). No se conocen reportes de hibridización de estas especies en estado natural.
La importancia de este estudio con este género en particular radica, en que los pecaríes son recursos naturales significativos, principalmente en la Amazonia y en todos los lugares en que habitan en general. Su papel ecológico es muy importante, debido a que es la principal fuente de alimento para varias especies de carnívoros, además contribuyen de alguna manera a la dispersión de semillas a pesar de que se les considera pobres en dicha actividad, si los comparamos con las aves o los murciélagos. Debido a que consumen frutos que se encuentran en el suelo, han desarrollado estrategias de alimentarse tanto de la pulpa como de las semillas de dichos frutos, esto los convierte en buenos depredadores de semillas. Por otro lado, son la fuente más importante de carne, tanto para subsistencia de los habitantes de las regiones en las cuales habita, como para su venta en los mercados urbanos. Adicionalmente, las pieles de pecarí son legalmente exportadas y vendidas desde sus lugares de origen, siendo un factor importante para la economía de las diferentes regiones en donde se encuentra (Bodmer et al., 1997).
Ante el desconocimiento de muchos aspectos que tienen que ver con estos organismos y ante la notable falta de estudios moleculares que caracteriza a este género en particular, se pretende ampliar con análisis de microsatélites, aspectos hasta hoy en día desconocidos, que definen la forma como están estructuradas genéticamente estas poblaciones, complementándose así a los estudios de carácter ecológico que se han adelantado hasta el momento. De esta manera, se contribuye al conocimiento de estas especies, para aquellos estudios con ánimos conservacionistas, tratándose de organismos que ofrecen potencial económico y comercial. Además, se ofrece mayor conocimiento acerca de la historia evolutiva del género, analizando las estructuras poblacionales actuales y las diferencias a nivel molecular que ofrecen las poblaciones gracias a su diversa distribución en Sudamérica.
Este estudio pretende determinar mediante el análisis de microsatélites, la estructura genética y reproductiva de las diferentes poblaciones de pecaríes, correspondientes a Colombia, Ecuador, Bolivia y Perú; así como la variabilidad genética existente entre las dos especies principales en estudio: Tayassu pecarí y Tayassu tajacu, además de determinar dicha variabilidad genética entre organismos de una misma especie dentro de un mismo grupo y también de diferentes grupos que habitan distintas regiones de Sudamérica.
En términos generales, la variabilidad genética se caracteriza por la presencia de dos o mas alelos en una población, independientemente de sus efectos fenotípicos, por eso se habla frecuentemente de locus polimórficos. El término polimórfico usualmente se refiere a aquellos alelos poco comunes que tienen una frecuencia mayor a 0.01. El polimorfismo es causado por diferencias alélicas en un solo locus (Futuyma, 1997).
Dobzhansky en su libro “Genetics and the Origin of Species”, consideró la variabilidad, como un carácter a nivel de especie, relacionado con la supervivencia de la misma. También afirmó, que una acumulación de variabilidad genética, actúa como un mecanismo que combate la extinción. Especies que concentran adaptaciones muy estrechamente, son favorecidas por la selección natural en un momento dado (Lloid y Gould, 1993).

4242
Los microsatélites del DNA, son una clase especial de repeticiones en tándem, que consiste en fragmentos de DNA de aproximadamente 2 a 6 pb por repetición, los cuales se repiten hasta mas de 100 veces (Tautz et al. 1993, citado por Zhivotovsky y Feldman, 1995). Los microsatélites son altamente polimórficos, presentando en algunos casos docenas de alelos por cada locus y tasas de mutación mas altas de 10-3 (Jeffreys et al. 1988; Kelley et al. 1991, citado por Zhivotovsky y Feldman, 1995) o 10-4 (Levinson et al. 1987; Henderson et al. 1992, citado por Zhivotovsky y Feldman, 1995). Por esta razón, su uso es bastante apropiado en taxonomía molecular, evolución y genética de poblaciones (Bowcock et al. 1994, citado por Zhivotovsky y Feldman, 1995). En poblaciones y en genética evolutiva, los microsatélites son una herramienta muy poderosa porque, además de la frecuencia de los alelos, el número de repeticiones de un alelo puede ser visto como características cuantitativas. Esta aproximación es útil para la genética cuantitativa y de hecho puede ser utilizada para estimar el momento central en el número de repeticiones en las poblaciones, para evaluar distancias genéticas entre poblaciones y sus correspondientes estadísticos F.
Materiales y Métodos
Para este estudio se emplearon técnicas moleculares conjuntamente con estudios morfológicos tomados de análisis craneométricos practicados a un total de 72 cráneos pertenecientes a las dos especies. Este análisis consistió en tomar una serie de medidas a cada uno de los cráneos con el objetivo de establecer diferencias a nivel sistemático y taxonómico, además, esta técnica también permite establecer diferencias entre individuos que tienen lugares de procedencia distintos.
Para los estudios moleculares, se analizaron 75 muestras de pecarí de collar y 71 muestras de pecarí de labio blanco, correspondientes a sangre, pelo, dientes, músculo, cartílago y piel, provienen principalmente de los siguientes países: Colombia, Perú, Ecuador, Brasil y Bolivia.
Las muestras de sangre fueron obtenidas de animales en campo y corresponden a 5ml de sangre preservada en EDTA disódico. Las muestras de músculo obtenidas en campo, fueron inmediatamente preservadas en alcohol para su posterior transporte al laboratorio. Las demás muestras fueron obtenidas también de animales en cautiverio, pieles de colección, animales muertos por cazadores, especímenes encontrados en mercados urbanos etc.
La preparación de las muestras de pieles, dientes y tejidos blandos consistió en lavar cuidadosamente los tejidos con el objetivo de extraer la mayor cantidad posible de contaminantes que puedan interferir con el resultado final del procedimiento de extracción, tales como microorganismos, químicos, etc.El DNA genómico se aisló utilizando principalmente el método orgánico de extracción de fenol-cloroformo con algunas modificaciones dependiendo del tipo de tejido.
La muestra fue mezclada con buffer de lisis ( 50 mM Tris-HCl pH 7.5; 1mM EDTA, 100 mM NaCl, 1% SDS) con proteinasa K (10 mg/ml) e incubada a 55 °C de un día para otro. Posteriormente fue centrifugada y el sobrenadante será extraído tres veces con fenol-cloroformo-isoamilico (25:24:1) y dos con cloroformo-isoamilico (24:1). La fase acuosa fue precipitada con etanol absoluto y 10% de 3 M acetato de sodio y centrifugada. Después el DNA precipitado fue lavado tres veces con 70% etanol y finalmente fue resuspendido en agua o en buffer TE.
Los productos de DNA extraídos en cada muestra fueron analizados en geles de agarosa al 0.8% y visualizados con bromuro de etidio para evaluar su cantidad y calidad de acuerdo a la metodología

43
citada por Sambrook y Russell (2001). De acuerdo con la cantidad de DNA presente en cada muestra, se realizaron las respectivas diluciones para obtener la concentración de trabajo adecuada.
Para las muestras de pelo se utilizó el protocolo de extracción con resina Chelex al 10%, con algunas modificaciones del método de Walsh (1991); para esto únicamente se emplearon pelos que contenían bulbo o raíz, cortando una longitud aproximada de 1 cm desde el bulbo.
Posteriormente, al obtener el DNA se procedió a amplificar los fragmentos de interés por medio de la técnica llamada PCR (Reacción en Cadena de la Polimerasa); en la cual, en primer lugar se hicieron pruebas con primers de cerdo doméstico (Sus scrofa) ya que no se contaba con los específicos para las especies de pecarí. Se realizó una estandarización del protocolo de PCR y se comprobó que dichos primers funcionaban correctamente como se describe a continuación.
Las reacciones de PCR se llevaron a cabo de la siguiente manera:
El volumen final de las reacciones de PCR que tienen que ver con muestras obtenidas de sangre, diente, músculo y cartílago por medio de los protocolos de extracción con fenol-cloroformo, corresponde a 25 μl, utilizando las siguientes cantidades:
14.5 μl de agua, 2.5 μl, de MgCl 2 3mM, 2.5 μl de buffer 10x, 0.5 μl de dNTPs, 1 μl de cada marcador, 2 μl de DNA de cada muestra y 1 μl de Taq polimerasa. Los marcadores utilizados en principio (Tabla 1), fueron SW951, SW857, IGF1 y SW240, originalmente diseñados para cerdo doméstico (Sus scrofa).
Tabla 1. Marcadores STR pertenecientes a cerdo doméstico utilizados y estandarizados para las dos especies de pecarí.
Las reacciones de PCR fueron llevadas a cabo en un termociclador Geneamp System 9600 termocycler de Perkin Elmer. Las temperaturas que se emplearon fuerón:
1 ciclo a 95ºC durante 5 minutos, 30 ciclos de 1 minuto a 94ºC, 1 minuto a 55ºC y 1 minuto a 72ºC, luego 1 ciclo de 5 minutos a 72ºC y luego la temperatura final a 4ºC.
Los productos de PCR se conservarón en nevera a una temperatura de 4ºC hasta su utilización en los geles de poliacrilamida. Luego, los productos amplificados de PCR fueron corridos en geles denaturantes de poliacrilamida al 6% en una cámara vertical Hoefer SQ3. La tinción de los geles se realizó con nitrato de plata, la cual permite visualizar las bandas de DNA con una resolución de hasta 1 pb de diferencia. Este método permite analizar bandas hasta de una sola base nucleotídica de diferencia.

4444
Análisis Genético Poblacionales:
Alelos y polimorfismo
En primer lugar, se calcularon las frecuencias alélicas de cada uno de los loci estudiados, además del número promedio de alelos por locus presentes en las diferentes poblaciones analizadas; posteriormente se observó la distribución de estas frecuencias alélicas y ver si se sobrelapan o no; también se calculó la diversidad genética en las diferentes poblaciones de pecaríes utilizando el índice de heterocigocidad esperada de Nei, (1978).
Las estimaciones del número de alelos, fueron normalizadas debido al tamaño diferente de las muestras representativas para cada población perteneciente a cada país. Esto se hizo utilizando aproximaciones bootstrap y jackknife (Ruiz-García, 2001).
Equilibrio Hardy-Weinberg y desequilibrio gamético.
Se calculó el equilibrio Hardy-Weinberg para determinar el exceso o déficit de heterocigotos y/u homocigotos dentro de las poblaciones observadas pertenecientes a los países de interés y en la población total en general. Para esto se utilizaron diferentes estrategias. Los estadísticos F de Weir y Cockerhan (W-C F) y los estadísticos F de Robertson y Hill (R-H F), fueron utilizados para calcular los excesos de homocigotos o de heterocigotos en las poblaciones de pecaríes. Para medir las probabilidades exactas de estos estadísticos, se utilizó el método de las cadenas de Markov con 10000 repeticiones y 500 batches y 10000 Interacciones por Batch, siguiendo a Raymond y Rousset (1955) con el programa Genepop 3.1.
El equilibrio Hardy-Weinberg se analizó por locus, por población y simultáneamente por loci y población utilizando el método de Fisher. El desequilibrio gamético fue calculado mediante los procedimientos de cadenas de Markov y el método de Fisher, utilizando los mismos parámetros que se tuvieron en cuenta en el cálculo del equilibrio Hardy-Weinberg.
Heterogeneidad genética y flujo genético
Para calcular la heterogeneidad genética entre las diferentes poblaciones de las dos especies de pecarí, se utilizó el método de probabilidades exactas con cadenas de Markov utilizando 1000 dememorizaciones, 50 batches y 1000 interacciones por batch. Se utilizó también el método de Fisher como un test global simultáneo, considerando todos los loci analizados.
Los estadísticos F de Wright fueron utilizados por el procedimiento de Michalakis y Excoffier (1996). La significancia de estos estadísticos F fue medida utilizando 1000 permutaciones Jackknife y 1000 permutaciones bootstrap utilizando el programa FSTAT. Además, las desviaciones estándar con intervalos de confianza del 95% y el 99% de los estadísticos FIT y FST fueron llevadas a cabo utilizando Jackknife sobre las poblaciones y sobre los loci, así como bootstrap sobre los loci (Ruiz-García, 2001).Para efectos del estadístico FST el flujo genético indirecto teórico fue estimado (Nm) ¬ para las diferentes poblaciones de pecarí, aplicado un modelo de islas infinitas (Ruiz-García 1993, 1998; Ruiz-García y Álvarez 2000) y un modelo n-dimensional de islas (Takahata 1983; Crow y Aoki 1984). Posteriormente el flujo genético fue calculado utilizando el modelo de alelos privados (Slatkin, 1985; Barton y Slatkin, 1986).

45
Asignación poblacional
Las muestras de pecaríes con lugar de procedencia geográfica conocida fueron procesadas con el programa GENECLASS de Cornuet y col. (1999), el cual es muy útil para asignar individuos a poblaciones concretas.
Se aplicaron métodos de distancias genéticas y métodos de verosimilitud. Para los métodos de distancias genéticas, los cuales asignan individuos a la población más cercana a la cual pertenecen calculando la distancia genética entre los individuos y la población analizada, se utilizaron las distancias genéticas de Nei (1978), Cavalli-Sforza y Edwards (1967), DAS (shared allele distance o distancia de alelos compartidos; Chakraborty y Jin, 1993) y la distancia δμ2 (Goldstein y col., 1995).
En cuanto a los métodos de verosimilitud, se utilizó el método Bayesiano similar al propuesto por Rannala y Mountain (1997) y el método de frecuencias propuesto por Paetkau et al., (1995), así como los métodos estadísticos “leave one out” y “as is”.
Cuellos de botella
Otro de los análisis genético-poblacionales que se realizaron, consistió en detectar si las poblaciones de pecaríes en estudio han pasado por cuellos de botella recientes. Se entiende por la palabra reciente, cuando las especies han pasado a través de un cuello de botella hace 2Ne-4Ne generaciones, siendo Ne el número efectivo de animales en cada población y considerándose también el tiempo que dura cada generación en este organismo en particular. Las poblaciones que han experimentado cuellos de botella recientes, simultáneamente sufren una pérdida en el número de alelos y en los niveles esperados de heterocigocidad. Calculando estos valores se puede determinar si en estas poblaciones ocurrió un evento de estas características (Cornuet y Luikart, 1996; Luikart y Cornuet, 1998).
Para detectar si existieron recientes cuellos de botella en las poblaciones estudiadas, se utilizaron dos métodos: El primero de ellos, basado en la teoría que afirma que las poblaciones que recientemente han sufrido cuellos de botella, simultáneamente decrece el número de alelos y los niveles esperados de heterocigocidad. Sin embargo, el número de alelos (ko) se reduce más rápidamente que los valores de heterocigocidad esperada (Cornuet y Luikart, 1996; Luikart y Cornuet, 1998). Sin embargo el valor de heterocigocidad esperada, calculado a través del número de alelos (Heq) es más bajo que el obtenido en la heterocigocidad esperada (He). Este exceso de heterocigocidad esperada en contraste con aquel obtenido a través del número de alelos, es claramente demostrable a través del modelo de alelos infinitos y no tan claramente a través del modelo mutacional step-wise (Otha y Kimura, 1973). En una población que haya presentado cuellos de botella, una gran fracción de los loci analizados presentarán un marcado exceso en la heterocigocidad esperada. Para calcular esta probabilidad, se utilizaron cuatro pruebas: Una prueba de significancia, una prueba de diferencias estandarizadas, una prueba “signed rank” o prueba de signo de Wilcoxon y gráficos que describan la forma de la distribución de las frecuencias alélicas. Una población que no haya pasado por un cuello de botella reciente, presentará una distribución en forma de L como lo que se esperaría en una población estable en equilibrio genético deriva-mutación, por otro lado, una población que haya presentado un cuello de botella reciente presentará una distribución “mode-shift”.

4646
La prueba de Wilcoxon es quizás la herramienta más poderosa cuando se trabaja con un número escaso de loci microsatélites como es el caso de este estudio. Todos los análisis descritos anteriormente se llevaron a cabo en el programa BOTTLENECK de Piry y col., (1999).
Números efectivos
Nielsen (1997), mediante modelos matemáticos propuestos por Otha y Kimura (1973) y Wehrhahn (1975), calculó la probabilidad de que un alelo seleccionado al azar, posee m repeticiones más que otro alelos también seleccionado al azar, aplicando la teoría de la coalescencia para obtener las funciones de verosimilitud de θ para muestras de un tamaño determinado. El tiempo de coalescencia de dos alelos determinados presenta una distribución exponencial con una media igual a 1 y el número condicional de mutaciones de cada linaje sigue una distribución de Poisson con una media de θc / 2.
En la siguiente expresión, la probabilidad q(ω) de la muestra es determinada por la adición de todos los posibles estados alélicos previos por la probabilidad de estar en un estado determinado y por la probabilidad de transición de este estado al estado actual. Teniendo en cuenta el modelo mutacional escogido (Un solo paso) esta probabilidad se puede calcular de la siguiente manera:
q(ω) = (θ / (n + θ – 1)) Σ (ni + 1/(n)) ½ q (ω + εi - εj ) + (n - 1)/ (n + θ - 1) Σ (nj – 1)/ (n – 1) q (ω - εj ), donde εi es un vector unitario el cual agrega valores iguales a 1 a la entrada de i en ω. Este procedimiento es fácil de desarrollar si tenemos la capacidad de obtener los valores de (θ/(n + θ – 1)), el cual, es la probabilidad de que el último evento justo antes del presente, sea una mutación considerando de que previamente ha ocurrido un evento mutacional o un evento coalescente, por eso, si tenemos (n – 1) / (n + θ – 1) el cual es la probabilidad de que ocurra un evento de coalescencia considerando que haya ocurrido un evento de mutación o de coalescencia previamente; también (ni – 1) / (n – 1), el cual es la probabilidad de que una mutación ocurra en un alelo i dado que ya ha ocurrido una mutación; y (nj– 1) / (n – 1), el cual es la probabilidad de que dos alelos que provienen del estado j sean coalescentes, dado que haya ocurrido un evento coalescente anteriormente. La probabilidad de que una mutación ocurra de un estado i hacia un estado j dado que ocurra una mutación en el estado i, es ½ (Ruiz-García y col., en prensa).
También se analizaron si cada uno de los microsatélites para cada una de las especies de pecarí estaban afectados por diferentes tasas de mutación. Para esto se comenzó con la hipótesis θ1= θ2= θ (Valores de θ para dos microsatélites diferentes) y se le aplicó una prueba de verosimilitud con la expresión –2log (L1(θ)L2(θ))/(L(θ1, θ2)), la cual sigue una distribución χ2 con un grado de libertad. Una probabilidad más baja de α = 0.05 indicaría que los microsatélites presentan diferentes tasas de mutación.
También se midió si el modelo de mutación paso a paso se ajusta significativamente mejor que el de un solo paso en cada una de las especies de pecarí. Teniendo en cuenta la expresión –2log (L(θ, p =0) / L(θ, p)) la cual en muestras grandes sigue una distribución de χ2 con un grado de libertad bajo la hipótesis nula de que p = 0, una probabilidad más baja de α = 0.05 indicaría que el porcentaje por mutaciones según el modelo paso a paso es significativamente diferente que el modelo de un solo paso y en este caso este último modelo es rechazado. (Ruiz-García y col., en prensa).

47
3- RESULTADOS
Alelos y polimorfismos
El loci con mayor número de alelos fue sw857 con un total de 32 alelos en el pecarí de collar y 18 alelos en el pecarí de labio blanco en la población colombiana. Este último en la población peruana alcanzó un total de 12 alelos y en la población boliviana un total de 8 alelos. IGF1 presentó un total de 10 alelos en Colombia para Tayassu tajacu y 9 alelos para Tayassu pecarí. Mientras que sw951 y sw240 fueron los loci que menor número de alelos presentaron en las diferentes poblaciones, con un mínimo de 2.En los resultados de los niveles de heterocigocidad genética con el método de Nei y el número de alelos encontrados por locus y por población para cada una de las especies (Datos no presentados), se observa que la diversidad genética fue elevada con valores de 1 para IGF1 en las poblaciones de Ecuador, Perú y Bolivia; valores de 0.944 y 0.833 para sw857 en la población colombiana y boliviana respectivamente y valores de 0.5 para sw951 en la población de Ecuador.
Equilibrio Hardy-Weinberg y Desequilibrio gamético
Los resultados del equilibrio HW para las dos especies de pecaríes en estudio (Datos no presentados) se observa que para Tayassu tajacu, ninguno de los marcadores mostró valores de p significativos por países lo cual nos lleva a concluir que las poblaciones de esta especie en los países estudiados para los marcadores analizados sí se encuentran en equilibrio Hardy Weinberg. Esto significa que al tomar estas poblaciones en el seno de los países, las diferencias genéticas entre ellos no fueron muy grandes. Por el contrario, en T. pecarí analizando la población total para esta especie, todos los valores a excepción de los marcadores sw857 y sw240 en la población boliviana y de sw951 en la población colombiana, presentaron valores significativamente positivos para los dos estadísticos F analizados, (Weir & Cockerham, 1984 y Robertson & Hill, 1984), con valores que se encontraban entre los rangos de 0.244 - 0.443 y 0.127 -0.405, respectivamente.
Teniendo en cuenta los análisis de desequilibrio gamético, vemos que se presentan dos casos de desequilibrio en la población de Colombia de Tayassu tajacu para los marcadores IGF1 con sw951 y para T. pecarí en los marcadores IGF1 y sw857 lo cual nos indica posibles eventos que se estén dando que afectan la evolución de estas dos especies o tal vez fusiones o arreglos cromosómicos que hayan ocurrido en el pasado que nos hagan pensar que estos locus microsatélites se encuentran en el mismo cromosoma o ligados de alguna manera en ambas especies.
Heterogeneidad genética y flujo genético.
Para Tayassu tajacu, dos de los marcadores estudiados mostraron un grado de diferenciación genética significativo para las poblaciones en estudio: (IGF1, p= 0.01568 y sw857, p= 0.01588). El marcador sw951 (p= 0.73858) no mostró diferencias significativas dado que para este locus los alelos encontrados en las poblaciones en estudio fueron pocos y resultaron similares. Analizando el grado de diferenciación genética para cada par de poblaciones, utilizando el método de cadenas de Markov, tenemos que para el locus IGF1 se presenta un grado de diferenciación genética significativo para el pecarí de collar en las poblaciones de Ecuador con respecto a las poblaciones de Bolivia y Perú (p=0.04728; p= 0.04647, respectivamente). También se observa un grado significativo de diferenciación entre la población de Perú y Colombia (p= 0.01902). Para el marcador sw857, la

4848
población de Ecuador muestra un marcado grado de diferenciación con las poblaciones de Perú, Bolivia y Colombia (p= 0.02271, p= 0.03154 y p= 0.00931, respectivamente). Por el contrario, el marcador sw951 no mostró un grado de diferenciación genética significativo entre las 4 poblaciones estudiadas (Goudet, 1996).
Cada prueba se realizó para cada uno de los marcadores microsatélites analizados, así como de manera conjunta para todos ellos. En el caso de las pruebas para Fis y Fit, para Tayassu tajacu todos los valores fueron altamente significativos a excepción del marcador sw951; para los marcadores IGF1 y sw857 se obtuvieron valores de 0.00020 para ambos casos respectivamente. Al realizar el test de manera global, los valores dieron de igual manera significativos lo que corrobora los resultados del estadístico F de W&C.
Los resultados del estadístico Rst (Rousset 1996) para Tayassu tajacu, fueron estimados para la población global y no fueron significativamente diferentes de los valores calculados para Fst de W &C, por lo tanto se podría llegar a conclusiones similares a las planteadas anteriormente.
Los resultados del programa STRUCTURE (Pritchard, Donnelly y Stephens 2000) se utilizaron para analizar la posibilidad que se encontraran, para las muestras globales de ambas especies, entre 1 y 15 acervos genéticos diferentes.
Para ambas especies de pecarí, el número de acervos genéticos más óptimos fue de 4, para k= 4, valor en el cual se obtuvieron los valores medios del Ln de verosimilitud más elevados. Eso significa que en ambas especies de pecarí existe una estructura genética consolidada y que, al menos, existen 4 poblaciones genéticamente bien diferenciadas en el rango geográfico estudiado.
Asignación poblacional
Los resultados con los métodos de distancias genéticas y de máxima verosimilitud utilizados para la asignación poblacional de individuos correspondientes a la especie Tayassu pecarí y Tayassu tajacu se aplicaron con un total de 71 individuos de procedencias geográficas conocidas, los cuales se subdividieron en 36 ejemplares correspondientes a Bolivia, 9 individuos procedentes de Perú y 26 procedentes de Colombia.
Para los cálculos de distancias genéticas y de máxima verosimilitud utilizando, el método “as is” clasificó de una manera más correcta a los ejemplares en sus correspondientes lugares de procedencia que con el método “leave one out”.
La mejor clasificación de individuos se obtuvo con el método de la distancia de Nei DA y el procedimiento “as is” con un total de 85.92% de los pecaríes correctamente clasificados en su correspondiente país (61/71), seguido por el método Bayesiano con un total de 83.10% de individuos correctamente asignados (59/71); el método DAS de igual manera que el anterior con un total de 83.10% individuos correctamente asignados (59/71). Los porcentajes de clasificación más bajos fueron obtenidos por el método de frecuencias utilizando el procedimiento “as is” con un 39.44% de individuos correctamente asignados (28/71) y utilizando el procedimiento “leave one out”, la distancia estándar de Nei dió un porcentaje del 59.15. Por lo tanto, para el número limitado de marcadores moleculares empleados, los primeros métodos parecen los más poderosos para discriminar individuos. Porcentajes entre un 83 y

49
un 86% de asignación correcta muestra que existen diferencias genéticas considerables entre esas poblaciones de pecaríes.
El mayor porcentaje de asignación lo produjo el método de distancia mínima de Nei con un 68% de individuos correctamente asignados (51/75), seguido por el método bayesiano con un 64% de asignaciones correctas (48/75). Los porcentajes de clasificación más bajos se dieron con el método “leave one out”, específicamente con la distancia de Goldstein con un 5.33% (4/75) lo cual podría considerarse anormalmente bajo.
Cuellos de botella
En el pecarí de labio blanco (T. pecarí) para la población de Bolivia, bajo el modelo mutacional I.A.M no se detectaron diferencias significativas entre las heterocigocidades observadas y esperadas para los marcadores analizados, lo cual concuerda con la afirmación de que esta población no ha sufrido cuellos de botella recientes. Bajo el modelo mutacional step-wise únicamente se detectó un aumento en la heterocigocidad observada de 0.799 y 0.695 en los loci sw857 y sw951 respectivamente, respecto a la heterocigocidad calculada con el número de alelos presentes y que fue de 0.586 y 0.410, presentando diferencias significativas con p= 0.0036 y p= 0.0114, respectivamente. Dado que en este caso los valores de heterocigocidad calculada a partir de las frecuencias alélicas son menores que las heterocigocidades calculadas a partir del número de alelos, corresponde nuevamente con la hipótesis de que dicha población no ha pasado por cuellos de botella. Con las otras pruebas aplicadas como la de diferencias estandarizadas, la prueba de Wilcoxon, el test del Signo y un descriptor gráfico de la forma de la distribución de las frecuencias alélicas, se confirma la hipótesis de que no hubo cuellos de botella en esta población de pecaríes de labio blanco. Sin embargo en la prueba de diferencias estandarizadas, se observo bajo SMM un valor significativamente negativo (T2= -4.212; p= 0.00001) lo cual sugiere que dicha población está experimentando expansión, flujo genético entre acervos diferentes o se encuentra bajo el efecto Wahlund de subdivisión. Teniendo en cuenta lo anterior, se aportan más evidencias de que en la población en cuestión no se detectan cuellos de botella porque es posible que otros eventos poblacionales se superpongan a la acción de los cuellos de botella. La prueba grafica mode-shift, corrobora la inexistencia de cuellos de botella al ofrecer una distribución normal en forma de L, lo cual permite concluir que no existe evidencia que soporte eventos de cuellos de botella recientes en la población de pecarí de labio blanco de Bolivia.
Teniendo en cuenta la prueba de significancia (sign test) y la prueba mode-shift, se confirma que dicha población no han ocurrido eventos de cuello de botella dado que la primera no muestra valores significativos y la segunda ofrece una curva normal en forma de L. Sin embargo, en la prueba de diferencias estandarizadas se obtuvo que bajo el modelo I.A.M un valor positivamente significativo lo cual induce a pensar que la población colombiana existió; alguna vez estuvo sometida a un evento de cuello de botella.
Números efectivos
Los números efectivos de las poblaciones de pecarí correspondientes a Tayassu tajacu y Tayassu pecarí pertenecientes a Colombia y Bolivia fueron calculados utilizando la expresión Ɵ = 4Ne U, usando dos tipos de tasas de mutación correspondientes a 5.6 x 10 –4 y 7 x 10 –5 respectivamente,

5050
con el objetivo de evaluar cuál de estos dos rangos se ajusta mejor a los tamaños demográficos esperados en estas especies.
Para la poblaciones colombianas de T pecarí, los valores de Ne para el marcador IGF1 con tasas de mutación de 5.6 x 10 –4, oscilaron entre 7915 y 11853 individuos, para el marcador sw951 los valores de Ne oscilaron entre 163 y 176 individuos y para el marcador sw857 entre 13046 y 40825 individuos. Los valores promedio en este caso fueron de 16301 pecaríes para el modelo uni-step y 8358 pecaríes para el modelo multi-step. Para tasas de mutación de 7 x 10 –5, los valores de Ne para todos los marcadores aumentaron considerablemente, en el primer caso para el marcador IGF1 oscilaron entre 63324 y 94820 individuos; para el marcador sw951 dichos valores se comportaron entre 1306 y 1406 individuos y para el marcador sw857 los valores fueron significativamente mayores encontrándose entre los 104368 y 326599 individuos. Presentando valores promedio de 130410 pecaríes para el modelo uni-step y 66864 pecaríes para el modelo multi-step.
Para Tayassu tajacu en poblaciones colombianas, trabajando con una tasa de mutación de 5.6 x 10 –4, los valores de Ne oscilaron para el marcador IGF1 entre 5179 y 8253 reproductores; para el marcador sw951 los valores como en los casos precedentes fueron menores, comprendidos ente 159 y 176 y para el marcador sw857 oscilaron entre 34532 y 36783 individuos. Los promedios de Ne, en este caso fueron 14040 pecaríes para el modelo uni-step y 14320 pecaríes para el modelo multi-step. Como se esperaba, trabajando con una tasa de mutación de 7 x 10 –5 los valores de Ne aumentaron como en el caso anterior, resultando para IGF1 rangos entre los 41428 y los 66026 individuos; para sw951 valores entre 1269 y 1408 reproductores y para sw857 oscilaron entre los 276254 y los 294268 individuos. Y los promedios fueron de 112321 pecaríes para el modelo uni-step y 114562 pecaríes para el modelo multi-step. Se puede observar que para los dos modelos mutacionales, las estimas arrojadas por los marcadores dieron bastante parecidas.
Discusión
Datos moleculares
Diversidad genética.
En cuanto a los 4 marcadores utilizados en pecarí de labio blanco y los 3 utilizados en pecarí de collar, el 100% de estos fueron polimórficos. Los niveles de diversidad genética en las poblaciones de Tayassu tajacu fueron elevados para las poblaciones en estudio especialmente para el marcador IGF1 con valores entre 0.885 y 1 y para el marcador sw857 con valores entre 0.75 y 0.944. Para T. pecarí los valores resultaron un poco menores pero se mantienen en un nivel de elevada diversidad genética con rangos entre 0.585 y 0.951 para los dos marcadores mencionados anteriormente.
Resulta evidente entonces que la riqueza genética de estas especies es bastante considerable tanto en las poblaciones de Colombia y Bolivia como en las poblaciones de Perú y Ecuador que presentan valores un poco menores pero se mantienen en niveles elevados. Teniendo en cuenta que estas especies son objeto de caza excesiva en todas las localidades estudiadas ya que por su carne y la comercialización de su piel son objetivo primordial para los habitantes de esas regiones, podemos considerar que los niveles de diversidad genética se mantienen elevados, al menos para los marcadores estudiados. Además, la continua destrucción de su hábitat natural ha fragmentado

51
las poblaciones hasta el punto de haberse producido su desaparición de grandes áreas donde se distribuía originalmente (Bodmer y Sowls, 1998). Dado esto, se podría establecer que estos niveles de diversidad genética altos son producto de que estas especies han tenido tradicionalmente elevada variabilidad genética y tasas reproductivas igualmente altas, lo cual permiten que las poblaciones se recuperen rápidamente a estos eventos de sobrecaza y de destrucción de hábitat. Por otro lado, es probable que estos niveles de diversidad genética detectados en la actualidad no sean producto de las actividades de explotación recientes a las que están sometidas estas especies, sino que pueden ser el resultado de una estructura genética ancestral característica de cada una de estas especies (Ruiz et al. 2003).
Alelos y polimorfismos
Con respecto al número de alelos encontrados para cada especie en particular tenemos que el marcador en el cual se encontró un mayor número de alelos fue sw857 con un total de 32 alelos registrados en la población colombiana para Tayassu tajacu y 25 para Tayassu pecarí, seguido por 10 alelos para el marcador IGF1 en las dos especies y para el marcador sw951 un total de 6 alelos para Tayassu pecarí y 2 para Tayassu tajacu siendo este el marcador que menor número de alelos fueron encontrados. Asimismo la gran variabilidad alélica presentada por estos marcadores, por sw857 en particular, en estas especies los convierten en una herramienta fundamental para discriminar individuos entre si y para efectuar asignamientos de individuos de localidades geográficas desconocidas dado su alto grado de polimorfismo. Algunas poblaciones como la de Ecuador, Perú y Bolivia para el caso de Tayassu tajacu, presentan un número de alelos reportados para todos los marcadores relativamente bajo en comparación con la población de Colombia. Esto es debido, posiblemente, a que el número de muestras obtenidas para esta especie fue relativamente pequeño para dichas poblaciones, lo cual puede ser un limitante para este tipo de análisis. Caso contrario para T pecarí en el cual la cantidad de muestras obtenidas para cada una de estas poblaciones fue más elevada. Para fines conservacionistas o de zoocría, el alto grado de polimorfismo que presentan estos marcadores puede ser una herramienta muy útil cuando se planteen planes de cruzamiento en cautiverio, ya que por la facilidad con la que se pueden realizar asignamientos poblacionales, el distinguir los diferentes acervos genéticos de los cuales provienen los individuos en estudio, permite de acuerdo a los objetivos que se planteen, evaluar que tipo de cruzamiento sería el más adecuado.
Equilibrio Hardy-Weinberg
Con respecto al equilibrio Hardy Weinberg, se detectó un claro exceso de homocigotos en las poblaciones analizadas de Tayassu tajacu y Tayassu pecarí, a nivel global con excepción del marcador IGF1 en todas las poblaciones y sw857 en la población de Perú. Tanto en las poblaciones de Tayassu tajacu como en Tayassu pecarí en términos globales no se detectó equilibrio HW aunque algunos marcadores en el caso de Tayassu pecarí analizados individualmente presentaban una tendencia hacia el equilibrio. Este exceso de homocigotos contrasta con los resultados obtenidos por Campero (1999) en pecaríes de labio blanco en Costa Rica, el cual detectó un exceso de heterocigotos en las poblaciones estudiadas y un grado de diferenciación genética bajo entre sus grupos analizados. Este exceso de homocigotos reportado en este estudio puede ser debido a que se esté asumiendo que cada una de las poblaciones pertenecientes a cada país representan una población total y en realidad existan subpoblaciones dentro de cada una de ellas (efecto Wahlund), (Ruiz y Payan 2002). La existencia de

5252
3 subespecies para Tayassu tajacu (T.t. angulatus, T.t. patira y T.t. tajacu) y de 5 subespecies para Tayassu pecari (T. p. ringens, T.p. spiradens, T.p. equatoris, T.p. pecarí y T.p. albirostris) las cuales se encuentran bien distribuidas a lo largo del área de estudio, podría explicar de manera más correcta que el exceso de homocigotos que se dio en los datos analizados se deba a un efecto de subdivisión poblacional o efecto Wahlund. Otra posible explicación a este exceso de homocigotos puede ser por efecto de cuellos de botella recientes, pero esta hipótesis puede ser descartada dado que no se encontró evidencia en las poblaciones estudiadas en las múltiples pruebas aplicadas de que hayan estado sometidas a eventos de este tipo dado que los niveles de diversidad genética fueron bastante altos en las poblaciones de pecarí estudiadas. Se descarta en primera instancia los efectos de la endogamia y los efectos de deriva genética, debido a que si estos eventos tuvieran repercusión en las poblaciones analizadas los niveles de diversidad genética se encontrarían en un nivel inferior menor. El efecto de la selección a favor de los homocigotos podría descartarse también, dado que la probabilidad de que este evento afecte a los dos marcadores de igual manera es demasiado remota.
Desequilibrio gamético
Como se mencionó anteriormente, se presentó evidencia de que ocurría cierto nivel de desequilibrio gamético en la población colombiana de Tayassu tajacu para los marcadores IGF1 y sw951 lo cual nos indica posibles eventos que se estén dando que afectan la evolución de estas dos especies. La deriva genética y la consanguinidad son usualmente los principales factores que causan desequilibrio gamético (Ruíz-García y Payan, 2002), pero como se mencionó anteriormente no existen evidencias en este estudio que soporten esta hipótesis para ninguna de las poblaciones de pecarí analizadas. Sin embargo, el flujo genético entre poblaciones con acervos genéticos bien diferenciados o la selección epistática son los responsables en algunos casos de la ocurrencia de desequilibrio gamético en las poblaciones (Hedrick 1985, Ruiz-García, 2000). Para este estudio como se mencionará más adelante, se reporta evidencia de flujo genético y los acervos de las diferentes poblaciones estudiadas se diferencian consistentemente entre si, lo cual podría significar una explicación para el desequilibrio gamético presentado por los marcadores anteriormente mencionados. Pero, la inexistencia de un exceso de heterocigotos también es un inconveniente para esta hipótesis. Puede ser simplemente que esos marcadores estén parcialmente ligados en un mismo cromosoma.
Heterogeneidad genética.
Los análisis de heterogeneidad genética en ambas especies estudiadas, arrojaron resultados suficientemente altos que permitieron considerar a las poblaciones de Tayassu pecarí y Tayassu tajacu procedentes de los diferentes países de origen como poblaciones con un grado de diferenciación genética apreciable. Estos valores para Tayassu tajacu oscilaron entre 0.073 y 0.095 para Gst. Para el estadístico Fst resultó un valor de 0.05 para esta misma especie y para T. pecarí 0.079 y para el estadístico Rst, 0.046 para Tayassu tajacu, siendo todos ellos significativos. Esto es, aunque en este estudio se trabajó con un número pequeño de microsatelites debido a la inexistencia de marcadores específicos para las especies de pecarí, este número pequeño es suficiente, dado su alto grado de polimorfismo, para poder diferenciar los acervos genéticos de los cuales provienen los individuos.
Flujo genético
Las estimaciones de flujo genético (Nm) entre las diversas poblaciones pone en evidencia un número mayor de migrantes para la especie Tayassu tajacu (8.48 individuos / generación) que para Tayassu

53
pecarí (2.20 individuos / generación). Este número de migrantes es bastante alto lo que induce a pensar que existe una conexión genética entre las poblaciones estudiadas aún cuando la heterogeneidad genética es significativa. Los resultados obtenidos en este estudio fueron menores que los obtenidos para pecaríes del parque nacional Corcovado en Costa Rica utilizando marcadores enzimáticos y microsatélites (Campero 1999), el cual para T pecarí obtuvo un valor Nm= 3.93 individuos / generación. En concordancia con lo anterior, en cuanto a morfología y probablemente en cuanto a etología, las poblaciones de pecarí correspondientes a los dos estudios son similares, pero es probable que la distribución de dichas poblaciones en Costa Rica, dado que se trata de un espacio considerablemente más pequeño y que el grado de interacción de las manadas es posiblemente más alto, si lo comparamos con la distribución de las poblaciones en los cuatro países que involucra este estudio, los niveles de flujo genético van a resultar lógicamente más altos que los observados aquí. Es evidente la gran diferencia existente entre las cantidades de flujo genético que presentan las dos especies de pecarí, este mayor grado de conexión entre las poblaciones de pecarí de collar puede estar ligado a que su distribución en Suramérica es mucho más uniforme y cubre una mayor área que la del pecari de labio blanco (figura 3), el cual se reporta como inexistente en gran parte de Venezuela y una buena porción de los llanos orientales y parte de la zona norte de la amazonia colombiana (Grubb y Groves 1998). Esta mayor área de cubrimiento por parte del pecarí de collar le permite mantener más fácilmente la conectividad entre las poblaciones de Perú, Ecuador y Colombia o entre las poblaciones bolivianas y las peruanas. Igualmente parece claro que la capacidad de flujo génico es más elevada para el pecarí de collar que para el pecarí de labio blanco o bien, el tiempo de divergencia de las poblaciones de la primera especie son ostentablemente menores que en la segunda.
Asignación poblacional
Los procedimientos de asignación de individuos mostraron que los métodos que mejor resultaron para el pecarí de collar fueron los de Distancia mínima de Nei, el método Bayesiano y el método de frecuencias, todos utilizando la metodología “as is”. Para el pecarí de labio blanco, los mejores métodos para realizar este análisis fueron los de Nei (DA), el método bayesiano y el método de distancia de alelos compartidos (DAS) utilizando de igual manera la metodología “as is” dando porcentajes de más del 80% de asignaciones correctas y más del 60% de asignaciones correctas en el caso del pecarí de collar, los porcentajes en esta última especie dieron un poco menores pero de igual manera significativos. Estos porcentajes resultaron menores debido a que a diferencia de los análisis realizados en pecarí de labio blanco, se trabajaron en este caso 4 poblaciones en lugar de tres y el número de individuos por población fue mucho menor que en el caso anterior. Por el contrario las metodologías que arrojaron los menores porcentajes fueron los métodos de Frecuencias en el caso del pecarí de labio blanco y la distancia Ɗ2 de Goldstein en el caso del pecarí de collar, con porcentajes de 39.44% y 5.33% respectivamente.
Estos análisis aportaron información relevante sobre la dinámica poblacional de esta especie, dado que se ratifica la existencia de cuatro acervos genéticos diferentes tanto para el pecarí de labio blanco como para el pecarí de collar correspondientes a Colombia, Perú, Bolivia y Ecuador cuando se aplicó el programa STRUCTURE. En la mayoría de los casos, los animales que no fueron asignados correctamente como por ejemplo, animales colombianos asignados a Perú o animales peruanos asignados a Bolivia, etc. coinciden con que la procedencia de dichos animales es de regiones que circundan las zonas limítrofes entre los diferentes países por lo tanto es muy probable que debido a los niveles de flujo genético existentes reportados aquí, exista un intercambio genético que pueda ser detectado por estos análisis. En los casos en que la asignación definitivamente no corresponda,

5454
generalmente esto puede ser debido a que esos animales no tuvieron resultados completos para todos los microsatélites analizados así que en estos casos, estas asignaciones pierden suficiente validez como para ser tenidas en cuenta. También es posible que el número muestral representativo para algunos países no sea suficiente. Cualquiera que fuera el caso, es claro que apoyándose en otros análisis además de este, existen puntos de corte entre los diferentes acervos genéticos tanto para el pecarí de labio blanco como para el pecarí de collar. Estos resultados corroboran nuevamente la gran utilidad que tienen los marcadores microsatélites para discriminar poblaciones diferenciadas en función de las áreas a las cuales pertenecen.
Como se mencionó anteriormente, hay que tener en cuenta el número de subespecies morfológicamente definidas que se encuentran en cada país. En particular para el caso de Colombia, en la especie Tayassu tajacu, se encuentra ampliamente distribuida la subespecie T.t. patira y únicamente en la parte sur de la amazonia se encuentra T. t tajacu. El mismo caso ocurre en Perú y Bolivia en el cual aparecen reportadas las dos subespecies mencionadas anteriormente y a diferencia de los anteriores en Ecuador únicamente está reportada T.t. patira. Por otro lado, T pecarí para el caso de Colombia, se encuentran reportadas 4 subespecies (T.t. ringens, T.t. spiradens, T.t.equatoris y T.t.pecari), para Ecuador únicamente T.t. equatoris, para Perú aparecen T.t. equatoris y T.t. albirostris y para Bolivia únicamente T.t. albirostris (Bodmer y Sowls, 1998, March 1998).
Aunque esta metodología resulta muy útil para detectar subespecies dentro de una población dada, en este caso los niveles de flujo genético son bastante elevados, así que no necesariamente los resultados aquí obtenidos demuestran eso. Sin embargo, las estimas altas de flujo genético global basadas en estadísticos de heterogeneidad genética podrían enmascarar una posible limitación de conexión entre las poblaciones (Ruiz-García y col., 2003).
Cuellos de botella
Los resultados aquí obtenidos fueron bastante parecidos para las dos especies en estudio. La mayoría de estos resultados ofrecidos por los marcadores fueron negativos para los dos modelos mutacionales tratados, el modelo de alelos infinitos y el modelo “step-wise”. Esto nos permite concluir que no ocurrieron eventos de cuello de botella recientes en ninguna de las especies analizadas.
Para el caso de ambas especies, los valores de heterocigocidad calculada a partir del número de alelos son mayores que las heterocigocidades calculadas a partir de las frecuencias alélicas, corresponde nuevamente con la hipótesis de que dicha población no ha pasado por cuellos de botella. Con las otras pruebas aplicadas como la de diferencias estandarizadas, la prueba de Wilcoxon, Sign test y la prueba gráfica de la distribución de las frecuencias alélicas, se confirma la hipótesis de que no hubo cuellos de botella en estas poblaciones.
Ciertos eventos evolutivos pueden afectar de buena manera los resultados obtenidos en el anterior análisis. Una de las condiciones que propone este análisis es que las poblaciones deben encontrarse en equilibrio mutación – deriva y de acuerdo con los análisis anteriormente aplicados los resultados plantean que las poblaciones en estudio no lo están, como se discutió anteriormente. Es posible que se deba al efecto Wahlund (subdivisión poblacional). No obstante, los análisis que se aplicaron para detectar diferenciación poblacional así como los resultados obtenidos por el programa STRUCTURE, diferencian muy bien los distintos acervos genéticos por lo tanto reducen los efectos que pudiera tener el efecto Wahlund en los datos aquí obtenidos.

55
Números efectivos
Para esto, se utilizaron dos tasas de mutación diferentes, 5.6 x 10-4 que es la tasa de mutación típica para los mamíferos y 7 x 10-5 que es la tasa de mutación típica para cerdos. Además, fueron calculadas teniendo en cuenta los dos modelos mutacionales (multi-step y uni-step) que pueden afectar a los microsatélites. Cabe anotar que no existen censos poblacionales para estas especies en la actualidad, por lo que los valores obtenidos aquí significan un primer acercamiento al número de individuos reproductores existentes en promedio a lo largo de la historia de la especie. Los números efectivos que mejor ofrecen tamaños reales para las poblaciones actuales son los obtenidos con la tasa de mutación de 7 x 10-5. Por lo tanto, se dará énfasis a estos resultados. Para Tayassu tajacu correspondientes a la población de Bolivia, los valores de Ne oscilaron para todos los marcadores entre 5.003 y 9.527 pecaríes para el modelo multi-step y entre 15.428 y 19.140 pecaríes para el modelo uni-step. Los cálculos para esta especie no se realizaron en los demás países debido al menor número de muestras que se obtuvieron.
En las poblaciones de Tayassu pecarí de Bolivia los valores Ne oscilaron entre 63.324 y 326.599 pecaríes para el modelo uni-step y para el modelo multi-step, valores Ne entre 94.820 y 104.368 pecaríes. Para Colombia con el modelo uni-step, los números Ne se dieron entre 41428 y 294268 pecaríes y para el modelo multi-step se dieron entre 66026 y 276254 pecaríes. Los dos modelos mutacionales ofrecen valores que pueden acercarse a los tamaños efectivos reales de las poblaciones actuales. Si esto es así, y aunque los niveles de diversidad genética son elevados lo cual puede evidenciar que las poblaciones de pecarí se recuperan rápidamente de las presiones de sobrecaza y destrucción de hábitat, esto no significa que no se vaya a llegar a un punto en el que estas presiones antrópicas superen los niveles de recuperación que poseen estas especies. Por lo tanto no se deben descuidar los programas de conservación para estas especies. Los datos obtenidos por el marcador sw951 en las dos especies y sw240 en T. pecarí fueron descartados debido a que se considera que los valores de Ne ofrecidos por estos marcadores difieren considerablemente de los promedios normales que dieron en los demás marcadores. Esto puede ser debido a dos razones. La primera de ellas es que exista alguna constricción mutacional en esos microsatélites. La segunda es que existen fenómenos de selección purificante actuando directamente o en áreas ligadas a ellos o simplemente estos marcadores se encuentren en áreas de baja recombinación (Ruiz-García et al. 2003).
Conclusiones
Las poblaciones de Tayassu pecarí y Tayassu tajacu pertenecientes a Colombia, Perú, Bolivia y Ecuador no se encuentran en equilibrio Hardy-Weinberg. En concordancia con lo anterior, se detectó un claro exceso de homocigotos para todas las poblaciones anteriormente mencionadas. Esto, puede ser debido a que se esté asumiendo que cada una de las poblaciones pertenecientes a cada país representan una población total y en realidad existan subpoblaciones dentro de cada una de ellas (efecto Wahlund).La diversidad genética de estas especies es bastante considerable en todas las poblaciones analizadas. Estos niveles de diversidad genética altos son producto de que estas especies han tenido tradicionalmente elevada variabilidad genética y tasas reproductivas igualmente altas, lo cual permite que las poblaciones se recuperen rápidamente de eventos de sobrecaza y de destrucción de hábitat. También es probable, que los niveles de diversidad genética detectados en la actualidad no sean producto de las actividades de explotación reciente a las que están sometidas estas especies, sino que

5656
pueden ser el resultado de una estructura genética ancestral característica de cada una de ellas.
Se detectaron niveles de desequilibrio gamético en la población colombiana de Tayassu tajacu para los marcadores IGF1 y sw951 y para Tayassu pecarí en los marcadores IGF1 y sw857. Una causa que podría haber producido este resultado es la existencia de flujo genético entre acervos genéticos significativamente diferenciados para los microsatélites estudiados. Pero, la inexistencia de un exceso de heterocigotos resulta un inconveniente para esta hipótesis. Puede ser simplemente que esos marcadores estén parcialmente ligados en un mismo cromosoma debido a reareglos cromosómicos que se hayan dado en los Tayassuidae respecto a los Suidae.
La heterogeneidad genética encontrada entre las poblaciones de cada una de las especies estudiadas permitió considerar a las poblaciones de Tayassu pecarí y Tayassu tajacu procedentes de los diferentes países de origen como poblaciones con un grado de diferenciación genética apreciable.
El nivel de flujo genético entre las diferentes poblaciones estudiadas fue bastante alto, lo que induce a pensar que existe una conexión genética entre dichas poblaciones aún cuando la heterogeneidad genética es significativa. Los valores de Nm = 2 en Tayassu pecarí y Nm = 8 en Tayassu tajacu, deja en evidencia que los acervos genéticos de la primera especie son mucho más diferenciados que los de la segunda.
Se ratificó, mediante técnicas de máxima verosimilitud, la existencia de cuatro acervos genéticos diferentes tanto para el pecarí de labio blanco como para el pecarí de collar correspondientes a las poblaciones de Colombia, Perú, Bolivia y Ecuador. Los porcentajes más altos de asignación poblacional para T. pecarí, corroboran el mayor grado de diferenciación que tienen los acervos genéticos detectados para esta especie.
No se detectó evidencia de eventos de cuellos de botella recientes en las poblaciones estudiadas para ninguna de las dos especies de pecarí.
Los promedios de los números efectivos (Ne) para T. pecarí son menores que para T. tajacu, esto ratifica nuevamente el mayor grado de diferenciación existente entre los acervos de la primera especie, dado que, a mayor número de individuos reproduciéndose en una población, mayor será la probabilidad de que haya conexión genética entre los acervos y como consecuencia el grado de diferenciación genética será menor.
Los análisis moleculares, aún con un número limitado de marcadores, ofrecieron un mayor poder de diferenciación entre los individuos estudiados que los métodos craneométricos. Estos, en general, fueron incapaces de discriminar de manera correcta a los animales estudiados en sus respectivas localidades geográficas. La asignación de especímenes de acuerdo a la localidad geográfica a la cual pertenecen utilizando métodos que involucran análisis craneométricos, depende más de las propiedades matemáticas intrínsecas que posee cada método utilizado en este estudio, que de las posibles correlaciones reales de origen biológico que exhiban las diferentes especies en el medio natural.

57
Bibliogrfía
Barton , N., Slatkin M. 1986. A quasi-equilibrium theory of the distribution of rare alleles in a subdivided population. Heredity 56: 409-416.
Bodmer R., Aquino R., Puertas P., Reyes C., Fang T., y Gottdenker N. 1997. Manejo y uso sustentable de pecaríes en la Amazonía peruana. Ocasional paper No 18 de la Comision de Supervivencia de Especies. Quito. Ecuador.
Bodmer R, Sowls L. 1998. Plan de acción y evaluación de la condición actual de los pecaríes. Museo Nacional de Historia Natural. La Paz- Bolivia.
Campero H. 1999. Variación y estructura genética dentro y entre grupos de chanchos de monte (Tayassu pecari) en el Parque Nacional Corcovado, Costa Rica. Tesis Mag. Sc. Universidad Nacional. Heredia (Costa Rica), 78p.
Cornuet J. and Luikart G. Description and evaluation of two test for detecting recent bottlenecks. 1996. Genetics. 144: 2001-2014.
Goldstein D., Linares L., Cavalli-Sforza and Feldman M. An evaluation of genetic distances for use with microsatellite loci. 1995. Genetics. 139: 463-471.
Grubb P, Groves P. 1998. Los Dicotylidae del Neotropico (Tayassu y Catagonus) Plan de acción para su conservación y evaluación de su condición actual. . Museo Nacional de Historia Natural. La Paz- Bolivia.
Luikart G. and Cornuet J. Empirical evaluation of a test for identifying recently bottlenecks. Populations from allele frequency data. 1998. Conservation Biology. 12: 228-237.
Estrada J.V. 1980. Las especies menos protegidas de nuestra fauna. Museo de Historia Natural, Universidad de Caldas. Informe.
Fradrich, H. 1968. Swine and peccaries. Pp.76-108, in Grzimek’s animal life encyclopedia (B. Grzimek ed.). Van Nostrand Reinhold Company, New York, 13:1-566.
Giannoni, M. A. Y I. Ferrary, 1976. Estudo biometrico do cariotipo da especie Tayassa albirostris Illinger 1815. Ciencia e Cultura, 28:432-435.
Griffiths, R. C., and S. Tavaré. 1994. Simulating probability-distributions in the coalescent. Theoretical Population Biology 46: 131-159.Hall. E. R. The mammals of North America. Second edition. John Wiley and Sons, New York 2:601-1181 + 90pp.
Hedrick P. W. 1985. Coat variants in cats. Gametic disequilibrium between unlinked loci. Journal of heredity 76: 127-131.

5858
Idiaquez, 1978. Mamíferos de mi tierra. López y Compañía. Tegucigalpa Honduras, 427pp.
Kendall, M., Stuart, A. 1977. The advanced theory of statistics. Volume I. Macmillan, New York.
Krieger, K. y D. Ochoa. (eds). 2002. Estado de la investigación en la Biodiversidad y la cooperación Científica y Técnica en Colombia. Instituto de Investigación de Recursos Biológicos Alexander von Humboldt. Informe.
March I. 1998. Plan de acción y evaluación de la condición actual de los pecaries. Museo Nacional de Historia Natural. La Paz- Bolivia.
Méndez, E. 1970. Los principales mamíferos silvestres de Panama. Zool. Lab. Conmemorativo Gorgas, Ciudad de Panama, Panama, 283pp.
Ohta, T., and M. Kimura. 1973. A model of mutation appropiate to estimate the number of electrophoretically detectable alleles in a finite population. Genetical Research 22: 201-204.
Paetkau, D., Shields, G., Strobeck,. C. 1998. Gene flow beteen insular, coastal and interior populations of brown bears in Alaska. Molecular Ecology, 7: 1283-1292.Perry R. 1970. The world of the jaguar. Taplinger press, New York. 168pp.
Piry, S., Luikart, G., Cornuet, M. 1999. BOTTLENECK: A computer program for detecting recent reductions of the effective population size using allele frecuency data. Journal of Heredity 90: 502-503.
Reich, D. E. and Goldstein D. B. Genetic evidence for a Paleolithic human population expansion in Africa. 1998. Proceedings of the National Academy of Sciences of the United States of America. 95: 8119-8123.
Ruiz-Garcia M. 1997. Genetic relationship among some new cat populations sampled in Europe. Spatial autocorrelation analysis. Journal of Genetics. 76: 1-24.
Ruiz-Garcia M. 1998. Genetic structure of different cat population (Felis catus) in Spain, Italy and Argentina at microgeographical level. Acta Theriologica 43:39-64.
Ruiz-Garcia M. 2000. Genetic microstructure in two Spanish cat populations. II: Gametic disequilibrium and spatial autocorrelation. Genes and Genetics systems 75:281-292.
Ruiz-Garcia, M. 2003. Molecular population genetic analysis of the spectated bear (Tremarctos ornatus) in the northern Andean area. Hereditas 138: 81-93.
Ruiz-Garcia, M. y Payan-Garrido E. 2002. Molecular and morphological evolution of the jaguar (Pantera onca) in Colombia using microsatellite markers and craneometric data. (en prensa).
Ruiz-Garcia M, Orozco P, Payan E, Castellanos A. 2003. Genética Molecular de Poblaciones aplicada al estudio de dos grandes carnívoros Neotropicales (Tremarctos ornatus Cuvier, 1825-Oso Andino y

59
Panthera onca Linné, 1758-Jaguar): lecciones de conservación. Bol. R. Esp. Hist. Nat. (Sec Biol.), 98 (1-4), 135-158.
Ruiz-Garcia M., Escobar, P., Mudry, M., Ascunce, M., Gutierrez, G. Microsatellite DNA analyses of four Alouatta species (Atelidae, Primates): Evolutionary Microsatellite dynamics. (en prensa).
Sambrook, J., and Russell, D.W. 2001. Molecular cloning. A Laboratory Manual. Third edition. Cold Spring Harbord Laboratory press.
Slatkin, M. 1993. Isolation by distance in equilibrium and non-equilibrium populations. Evolution 47: 264-279.
Sokal R. And Oden N. Spatial autocorrelation in biology methodology. 1978. Biol. J. Linn. Soc. 10:199-228.
Sowls, L. K. 1978. Collared pecari. Pp. 191-205, in big game of North America: ecology and management. (J. L. Schmidt and D. C. Gilbert ed.). Stackpole Books, Harrysburg, Pennsylvania, 494pp.
Sowls, L. K. 1984. The peccaries. Univ. Arizona press, Tucson, 251pp.
Theimer T. and Keim P. Phylogenetic Relationship of Peccaries based on Mitochondrial Citochrome b DNA Secuences. 1998. Journal of mammalogy. 79(2): 566-572.
Theimer T. and Keim P. Geographic Patterns of Mitochondrial-DNA Variation in Collared Peccaries. 1994. Journal of mammalogy. 75(1): 121-128.
Weber, J. L., and C. Wong. 1993. Mutation of human short tandem repeats. Human Molecular Genetics 2: 1123-1128.
Wehrhahn, C. 1975. The evolution of selectively similar electrophoretically detectable alleles in finite natural populations. Genetics 80: 375-394.
Wetzel, R. M. y J. W. Lovett. 1974. A collection of mammals from the Chaco of Paraguay. Univ. Connecticut Ocass. Papers. Biol. Sci. ser. 2:203-216.
Zhivotosky, L. A., and M. W. Feldman. 1995. Microsatellite variability and genetic distances. Proc. Natl. Acad. Sci. USA 92: 11549-11552.
Zuckermann, S. 1953. The breeding season of mammals in captivity. Proc. Zool. Soc. London, 122:827-950.www.redlist.org

6060
Revista semestral Innova BiotecInstrucciones para Publicación
1. Definición
La revista “INNOVA BIOTEC” es una publicación periódica semestral (dos números regulares anualmente) publicada por la Dirección Ejecutiva del Instituto de Innovación en Biotecnología e Industria (IIBI).
“INNOVA BIOTEC” va dirigida a estimular editorialmente la producción científica y tecnológica nacional, así como a difundir a niveles extranacionales los resultados del trabajo de investigación teórica y experimental de nuestra comunidad cientíica.
Esta publicación periódica es interdisciplinaria, pero va dirigida preferentemente a trabajos experimentales dentro de los dominios temáticos de las Ciencias Naturales, Exactas, Agronómicas, Médicas, de la Tierra y del Ambiente, de la Ingeniería y de la Tecnología. De manera especial se presentarán artículos sobre biotecnología, disciplina que es parte fundamental de la misión del IIBI. Así mismo, se publicarán artículos de discusión y ensayos sobre temas generales de la ciencia, específicamente, sobre política y gestión científica, historia de la ciencia, filosofía de la ciencia, bioética, etc.
Está abierta al personal docente e investigativo del IIBI, pero acepta gustosamente colaboraciones de investigadores provenientes de otros Centros de Educación Superior, Entidades Gubernamentales o Institutos de Investigación nacionales o extranjeros.
2. Política Editorial
2.1 Todos los artículos serán responsabilidad exclusiva de los autores. Con el fin de prevenir el fraude o el plagio, el IIBI podrá verificar datos sobre la trayectoria científica de los autores y, sobre ésta base, decidir sobre su publicación definitiva.
2.2 Existirán dos tipos de artículos, según su origen: colaboraciones regulares enviadas por los autores y colaboraciones solicitadas por el Editor. En el primer caso, los trabajos serán enviados a revisión por parte de evaluadores externos, libremente seleccionados por el Editor, quienes de manera imparcial y anónima aprobarán, recomendarán cambios o rechazarán la publicación del trabajo remitido. En el segundo caso, será prerrogativa final del Editor publicar o no la colaboración solicitada.

61
2. Los trabajos a ser considerados para publicación deberán ser absolutamente inéditos. Los mismos deberán ser enviados por triplicado, vía correo ordinario, a la siguiente dirección:
Revista Innova Biotec c/o Dirección Ejecutiva Instituto de Innovación en Biotecnología e Industria Ave. Núñez de Cáceres, esq. Olof Palme, Sector San Gerónimo, Santo Domingo, D.N. República Dominicana
2.4 Los artículos enviados serán publicados Ad Honorem y la revista “Innova Biotec” retribuirá su aporte otorgando gratuitamente a cada colaborador un número determinado de copias de la publicación.
2.5 El IIBI conservará los derechos de autor sobre el material impreso en la edición de la que se trate. El autor mantendrá sus derechos de autor sobre ediciones sucesivas.
2.6 No se devolverán colaboraciones no solicitadas por el Editor.
3. Especificaciones Técnicas para las Colaboraciones.
3.1 La revista publicará tres tipos de colaboraciones:
3.1.1 Ensayos cortos y monografías.
Que versarán sobre revisiones bibliográficas divulgativas o artículos de reflexión y opinión sobre algún tema prioritario, relacionado a las Ciencias anteriormente definidas. - Se redactarán bajo la perspectiva de dirigirse a un público general. - Su extensión será de un máximo de 15 cuartillas, escritas a doble espacio. - Podrán incluír ilustraciones o gráficos, pero en un número estrictamente mínimo, nunca superior a una o dos, en cada caso.
3.1.2 Comunicaciones cortas.
Que versarán sobre resultados experimentales derivados de investigaciones, generalmente en progreso y que, por su novedad e importancia, deben ser publicados con celeridad. - Se redactarán dirigiéndose a una audiencia especializada. - Su extensión máxima será de 10 cuartillas, escritas a doble espacio.- Podrán incluír ilustraciones o gráficos, pero en un número mínimo, nunca superior a cinco, en total.

6262
3.1.3 Artículos científicos.
Que versarán sobre resultados originales derivados de investigaciones experimentales, en los dominios de las ciencias básicas o aplicadas. - Se redactarán dirigiéndose a una audiencia especializada. - Su extensión máxima será de 20 cuartillas, escritas a doble espacio.- Podrán incluír ilustraciones o gráficos, en un número nunca superior a diez, en total.
3.2 Todos los trabajos deberán tener un título claramente identificado. Adjunto al título irá el nombre del o los autores, así como su filiación institucional y su dirección postal. El primer autor listado será considerado como Autor Principal y a él será dirigida cualquier comunicación referente al trabajo. Toda ésta información deberá concentrarse en una misma página, exclusiva para éstos fines.
3.3 En el caso de los artículos científicos:
- deberán usarse las unidades del Sistema Internacional de medidas, obligatoriamente.- deberá incluírse siempre un resumen (abstract) escrito en español, con su respectiva traducción al inglés. La extensión de cada una de éstas versiones del resumen no podrá ser superior a media cuartilla, escrita a doble espacio, respectivamente.- deberán incluírse cinco palabras claves en español (y sus respectivas traducciones al inglés).- deberán contener las siguientes secciones: introducción, materiales y métodos, resultados, discusión y bibliografía. - las referencias bibliográficas deberán listarse según el sistema Harvard. Dichas referencias deberán ser preferiblemente recientes, generadas dentro de los últimos diez años anteriores a la fecha de publicación.
3.4 Las ilustraciones (dibujos, esquemas, fotografías) y gráficos (cuadros, tablas) deberán acompañar las copias del texto, presentándose en el formato final, listos para ser reproducidos. Deberán estar acompañados de sus respectivos títulos y estar numerados en secuencia, según el texto del trabajo. Los gráficos no deberán repetir la información de las ilustraciones.
3.5 Los textos enviados para ser considerados para publicación, podrán ser remitidos en un medio digital como CD. Pueden ser creados usando LATEX o con procesadores de texto como Open Office, Microsoft Word o Word Perfect.





























