Embed Size (px)
Citation preview

Inhibitory Effects of Brazilin on the VascularSmooth Muscle Cell Proliferation and
Migration Induced by PDGF-BB
Jing Guo,* Li Li,* Yu-Jie Wu,† Yu Yan,† Xiao-Na Xu,† Shou-Bao Wang,*
Tian-Yi Yuan,† Lian-Hua Fang† and Guan-Hua Du*
*State Key Laboratory of Bioactive Substances
and Functions of Natural Medicines
†Beijing Key Laboratory of Drug Targets Identification
and Drug Screening, Institute of Materia Medica
Chinese Academy of Medical Sciences
and Peking Union Medical College
Beijing 100050, China
Abstract: Abnormal vascular smooth muscle cell (VSMC) proliferation and migration con-tribute to the pathogenesis of vascular diseases including atherosclerosis and restenosis.Brazilin isolated from the heartwood of Caesalpinia sappan L. has been reported to exhibitvarious biological activities, such as anti-platelet aggregation, anti-inflammation, vasor-elaxation and pro-apoptosis. However, the functional effects of Brazilin on VSMCs remainunexplored. The present study investigated the potential effects of Brazilin on platelet-derived growth factor (PDGF)-BB induced VSMC proliferation and migration as well as theunderlying mechanism of action. VSMC proliferation and migration were measured byCrystal Violet Staining, wound-healing and Boyden chamber assays, respectively. Cell cyclewas analyzed by flow cytometry. Enzymatic action of matrix metalloproteinase-9 (MMP-9)was carried out by gelatin zymography. Expression of adhesion molecules, cell cycleregulatory proteins, the phosphorylated levels of PDGF receptor β (PDGF-RβÞ, Src, extra-cellular signal regulated kinase (ERK) and Akt were tested by immunoblotting. The presentstudy demonstrated that pretreatment with Brazilin dose-dependently inhibited PDGF-BBstimulated VSMC proliferation and migration, which were associated with a cell-cycle arrestat G0/G1 phase, a reduction in the adhesion molecule expression and MMP-9 activation inVSMCs. Furthermore, the increase in PDGF-Rβ, Src, ERK1/2 and Akt phosphorylationinduced by PDGF-BB were suppressed by Brazilin. These findings indicate that Brazilin
Correspondence to: Dr. Lian-Hua Fang and Dr. Guan-Hua Du, Institute of Materia Medica, Chinese Academy ofMedical Sciences and Peking Union Medical College, 1 Xiannongtan Street, Beijing 100050, China. Tel: (þ86)10-6316-5313; Fax: (þ86) 10-6316-5184, E-mail address: [email protected] (Lian-Hua Fang); [email protected]
(Guan-Hua Du).
The American Journal of Chinese Medicine, Vol. 41, No. 6, 1283–1296© 2013 World Scientific Publishing Company
Institute for Advanced Research in Asian Science and MedicineDOI: 10.1142/S0192415X13500869
1283
Am
. J. C
hin.
Med
. 201
3.41
:128
3-12
96. D
ownl
oade
d fr
om w
ww
.wor
ldsc
ient
ific
.com
by N
AT
ION
AL
CH
UN
G H
SIN
G U
NIV
ER
SIT
Y o
n 03
/28/
14. F
or p
erso
nal u
se o
nly.

inhibits PDGF-BB induced VSMC proliferation and migration, and the inhibitory effects ofBrazilin may be associated with the blockade of PDGF-Rβ - ERK1/2 and Akt signalingpathways. In conclusion, the present study implicates that Brazilin may be useful as an anti-proliferative agent for the treatment of vascular diseases.
Keywords: Brazilin; Vascular Smooth Muscle Cell; Proliferation; Migration; PDGF.
Introduction
The abnormal proliferation and migration of vascular smooth muscle cells (VSMCs) arekey events in the development of vascular diseases including atherosclerosis and restenosis(Dzau et al., 2002). Although these processes are triggered by multiple cytokines andgrowth factors, one of the principal regulators of chemoattraction and mitogenesis forVSMCs is platelet-derived growth factor (PDGF)-BB, the expression of which is increasedin atherosclerotic lesions (Bornfeldt et al., 1995; Levitzki, 2005; Huang et al., 2007).Binding of PDGF-BB to the PDGF recetpor β (PDGF-Rβ) induces receptor dimerizationand autophosphorylation, which allows binding and activation of cytoplasmic SH2-domaincontaining signal transduction molecules, such as PLC�, Src, Grb-2, and phosphatidyl-inositol 3 kinase (PI3K). These signal transduction molecules initiate a number of differentsignaling pathways in cells that lead to actin reorganization, cell growth, migration anddifferentiation. PI3K/Akt and mitogen-activated protein kinase (MAPK)/ERK are the twomajor PDGF signaling pathways (Dong et al., 2010; Moon et al., 2012), and the activationof them is required for mitogenic signaling through a number of tyrosine kinases. Recently,some studies have shown that the nonreceptor tyrosine kinase Src family plays animportant role in PDGF-BB-induced signaling events leading to cell proliferation,migration, and gene expression via interactions with multiple signaling pathways (Wal-tenberger et al., 1999; Yamboliev et al., 2001; Tsang et al., 2002; Veracini et al., 2006;Reddy et al., 2009).
Brazilin [7,11b-dihydrobenz (b) indeno[1,2-d]pyran-3,6a,9,10(6H)-tetrol] (Fig. 1) iso-lated from the heartwood of Caesalpinia sappan L. has been used as a beverage and foodcolorant in several Asian countries (Kim et al., 2012) as well as a natural red pigmentfor histological staining (Augulis and Sepinwall, 1969). Previous studies have demon-strated various biological activities of Brazilin, such as hypoglycemic activity (Moon et al.,1993), anti-hepatotoxicity (Moon et al., 1992), anti-platelet aggregation (Chang et al.,2013), induction of immunological tolerance (Mok et al., 1998), anti-inflammation (Baeet al., 2005), vasorelaxation (Sasaki et al., 2010), anti-allergic asthma properties (Lee et al.,2012), etc. Recent studies revealed the function of Brazilin on melanin synthesis sup-pression (Mitani et al., 2013) and pro- apoptosis in certain cells (Kim et al., 2012; Leeet al., 2013). However, little is known about the possible effect of Brazilin on VSMCproliferation and migration. The present study was conducted to further investigate theeffect of Brazilin on PDGF-BB induced VSMC proliferation and migration. The PDGF-related signaling in VSMCs was also considered.
1284 J. GUO et al.
Am
. J. C
hin.
Med
. 201
3.41
:128
3-12
96. D
ownl
oade
d fr
om w
ww
.wor
ldsc
ient
ific
.com
by N
AT
ION
AL
CH
UN
G H
SIN
G U
NIV
ER
SIT
Y o
n 03
/28/
14. F
or p
erso
nal u
se o
nly.

Materials and Methods
Antibodies and Major Reagents
Anti-phospho-PDGF-Rβ, anti-PDGF-Rβ, anti-phospho-ERK1/2, anti-ERK1/2, anti-phospho-Akt, anti-Akt, anti-cyclin D1, anti-cyclin E and anti-CDK2 antibodies were purchased fromCell Signaling Technology, Inc. (Beverly, MA, USA). Anti-intercellular adhesion mol-ecule 1 (ICAM-1) and anti-vascular cell adhesion molecule 1 (VCAM-1) antibodies wereobtained from Santa Cruz Biotechnology (Santa Cruz, CA, USA). PDGF-BB was obtainedfrom R&D System (Minneapolis, MN, USA). Brazilin was purchased from PureoneBiotechnology (Shanghai, China).
Cell Culture
Rat aortic VSMCs were isolated by enzymatic dispersion as described previously (Leeet al., 2011). Cells were cultured in DMEM, supplemented with 10% FBS to subcon-fluence and synchronized by serum deprivation for 24 h. Synchronized cells were treatedwith different concentrations of Brazilin for 2 h and then stimulated with PDGF-BB (20 ng/mL) for indicated time periods.
Cell Viability Assay
VSMCs were seeded into 96-well culture plates. When cells reached 70% confluence, themedium was replaced with serum-free medium. After 24 h incubation the cells wereexposed to different concentrations of Brazilin for an additional 24 or 48 h (Lee et al.,2011). Cell viability was measured using MTT assay.
Figure 1. Chemical structure of Brazilin.
BRAZILIN INHIBITS VSMC PROLIFERATION AND MIGRATION 1285
Am
. J. C
hin.
Med
. 201
3.41
:128
3-12
96. D
ownl
oade
d fr
om w
ww
.wor
ldsc
ient
ific
.com
by N
AT
ION
AL
CH
UN
G H
SIN
G U
NIV
ER
SIT
Y o
n 03
/28/
14. F
or p
erso
nal u
se o
nly.

Cell Proliferation Assay
VSMCs were seeded into 96-well plates (3� 103 cells per well) and grown in DMEMcontaining 10% FBS for 24 h. The cells were then synchronized by serum deprivationfor 24 h and pretreated with various concentrations of Brazilin for 2 h before stimulationwith or without PDGF-BB. Cell proliferation was measured by Crystal Violet Staining(Rothmeier et al., 2009).
In Vitro Wound-Healing Assay
VSMCs were grown to confluence in 24-well plates, starved in serium-free DMEMmedium for 24 h, and then injured with a single scratch using a sterile pipette tip. Cellswere then incubated in the absence or presence of PDGF-BB (20 ng/mL) or Brazilin (1, 3,10, 30�M). The wounds were photographed at 0 and 20 h after scratch.
Modified Boyden Chamber Migration Study
Cell migration was also performed using a modified Boyden chamber model (Transwell,8.0�m pore size, Corning, NY) as previously described (Lo et al., 2012). Briefly, cellswere trypsinized, washed with phosphate-buffered saline (PBS) and suspended in serum-free DMEM medium. A total of 100�L of cell suspension (2� 104 cells) was added tothe top of chamber. The lower chambers were filled with 500 �L DMEM supplementedwith PDGF-BB (20 ng/mL) or vehicle in the presence or absence of Brazilin. Thechambers were incubated at 37�C for 8 h to allow for cell migration. The cells from theupper surface of the filters were gently removed while the cells on the lower surface ofthe filters were fixed with 4% paraformaldehyde and stained with 0.1% crystal violet.Migrant cells were quantified by blind counting under a microscope (Olympus IX71,Tokyo, Japan). Eight random areas of cells per membrane were counted and averaged.Experiments were performed at least three times.
Cell-Cycle Progression Analysis
VSMC cell cycle distribution was analyzed using flow cytometry as previously described(Dong et al., 2010; Sun et al., 2012). The percentage of cells in different phases of thecell cycle was analyzed using Cell-FIT software (Becton Dickinson Instruments).
Zymography
Matrix metalloproteinase-9 (MMP-9) activity in the medium was analyzed by non-reducing SDS-PAGE in 10% gels containing 0.1% gelatin as previously described (Donget al., 2010).
1286 J. GUO et al.
Am
. J. C
hin.
Med
. 201
3.41
:128
3-12
96. D
ownl
oade
d fr
om w
ww
.wor
ldsc
ient
ific
.com
by N
AT
ION
AL
CH
UN
G H
SIN
G U
NIV
ER
SIT
Y o
n 03
/28/
14. F
or p
erso
nal u
se o
nly.
Immunoblotting Assay
Immunoblotting was performed as previously described (Fang et al., 2004; Lee et al.,2011; Moon et al., 2012). VSMCs were stimulated with 20 ng/mL PDGF-BB for 1 min forPDGF-Rβ, 5 mins for Src (Reddy et al., 2009) and ERK1/2, and 15 mins for Akt. VSMCswere stimulated with 20 ng/mL PDGF-BB for 24 h for the expression of cyclin D, cyclin E,CDK2, p27 and the adhesion molecule (ICAM-1 and VCAM-1). The bands were quan-tified by Quantityone software (Bio-Rad, Richmond, CA, USA) and normalized toGAPDH as an internal control.
Statistical Analysis
Results are expressed as mean� S:E:M. A one-way analysis of variance (ANOVA) andDunnett’s post-test were used for multiple comparisons. p � 0:05 was considered asstatistically significant.
Results
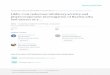
Brazilin Inhibits PDGF-BB-Stimulated VSMC Proliferation
As shown in Fig. 2A, treatment with PDGF-BB for 24 h and 48 h significantly promotedVSMC proliferation. Pretreatment with Brazilin in the concentration range from 1 to30�M for 2 h inhibited PDGF-BB induced VSMC proliferation in a concentrationdependent manner. When quiescent cells were treated with Brazilin (1–30�M) for 12–48 h in the absence of PDGF-BB, no significant difference was observed in the viability
(A) (B)
Figure 2. Brazilin inhibits VSMC proliferation induced by PDGF-BB. Synchronized VSMCs were pretreated withvarious concentrations of Brazilin for 2 h, and then stimulated with (for proliferation assay) or without (for cell
viability assay) 20 ng/mL PDGF-BB for indicated time periods. (A) VSMC proliferation was detected by crystalviolet staining. (B) Viability of VSMCs was determined by MTT assay. Data are expressed as mean� S:E:M:
ðn ¼ 4Þ. ##p < 0:01, vs. untreated control; *p < 0:05, **p < 0:01, vs. PDGF-BB control.
BRAZILIN INHIBITS VSMC PROLIFERATION AND MIGRATION 1287
Am
. J. C
hin.
Med
. 201
3.41
:128
3-12
96. D
ownl
oade
d fr
om w
ww
.wor
ldsc
ient
ific
.com
by N
AT
ION
AL
CH
UN
G H
SIN
G U
NIV
ER
SIT
Y o
n 03
/28/
14. F
or p
erso
nal u
se o
nly.

of VSMCs as compared with the untreated cells. These results suggest that Brazilin hasno cytotoxicity on VSMCs at the concentrations tested (Fig. 2B) and the anti-proliferative effect of Brazilin on VSMCs was not due to cytotoxicity.
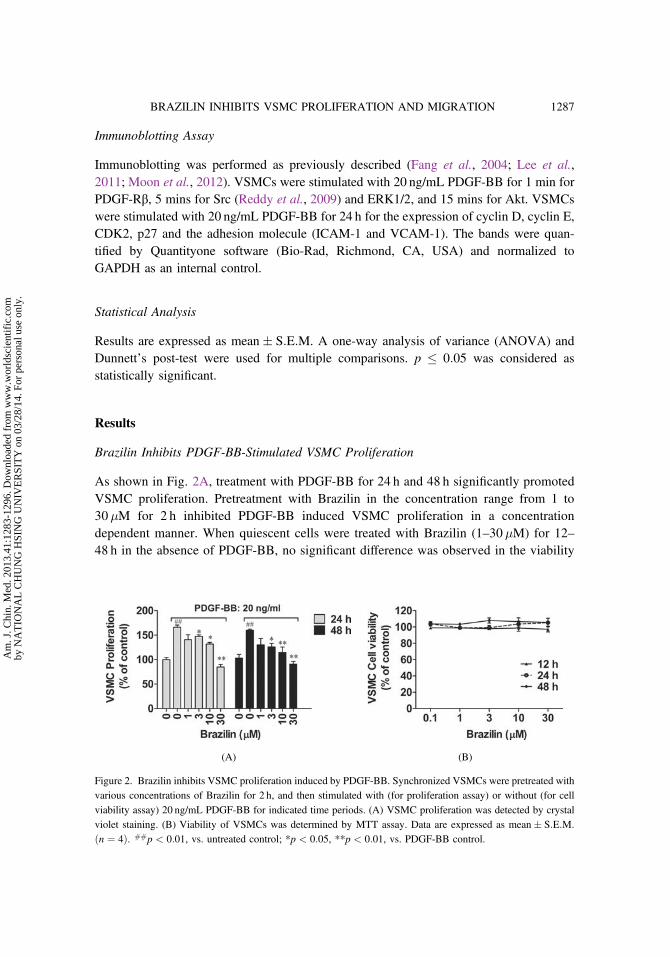
Brazilin Inhibits PDGF-Stimulated VSMC Migration
To address the effect of Brazilin on VSMC migration, which represents a cru-cial process in the pathogenesis of neointima formation, a wound-healing assaywas performed. As shown in Fig. 3A, PDGF-BB (20 ng/mL) induced VSMCs migratedsooner and distinctly narrowed the wound distance after 20 h treatment. Brazilin (1–30�M) concentration-dependently attenuated PDGF-BB induced VSMC migration.
The modified Boyden chamber experiment was then carried out to confirm theinhibitory effect of Brazilin on VSMC migration. As indicated in Figs. 3B and 3C,treatment with PDGF-BB for 8 h markedly increased the number of VSMCs thatmigrated through the Transwell chamber. However, pretreatment with Brazilin (3, 10and 30�M) for 2 h prior to PDGF-BB stimulation significantly reduced the number ofmigrated cells.
Furthermore, the levels of migration of regulatory proteins, including ICAM-1and VCAM-1 were detected by Western blot. As shown in Fig. 3D, Brazilin pretreatmentinhibited the upregulation of ICAM-1 and VCAM-1 induced by PDGF-BB in aconcentration-dependent manner. Zymography assay showed that Brazilin decreasedMMP-9 activity induced by PDGF-BB. These findings suggest that Brazilin inhibits themigration of VSMCs induced by PDGF-BB by suppressing the expression of migration-related proteins in these cells.
Brazilin Induces a Cell Cycle Arrest in PDGF-Stimulated VSMCs
To explore the mechanism of the inhibitory effect of Brazilin on PDGF-induced VSMCproliferation, the effects of the Brazilin on cell cycle progression were analyzed. Serum-deprivation of VSMCs for 24 h resulted in an approximately 88% synchronization of thecell cycle in G0/G1 phase. PDGF-BB stimulated cells in S phase increased from 6% to24%, and concomitantly decreased cells in G0/G1 phase from 88% to 61%. In contrast,Brazilin-treated cells showed a concentration-dependent accumulation of cells in G0/G1phase. Pretreatment with Brazilin at the concentrations of 3, 10 and 30 �M significantlyincreased cells in G0/G1 phase to approximately 68% (p < 0:05), 71% (p < 0:05), and81% (p < 0:01) (Figs. 4A and 4B). These findings indicate that G0/G1 cell cycle arrestinduced by Brazilin treatment contributes to its inhibitory effect on PDGF-stimulatedVSMC proliferation.
Cell cycle progression is strictly controlled by the positive and negative regulators thatact at checkpoints throughout the cell cycle. Then the protein levels of G0/G1 phaseregulatory proteins were detected, and as shown in Figs. 4C–4G, treatment with Brazilin
1288 J. GUO et al.
Am
. J. C
hin.
Med
. 201
3.41
:128
3-12
96. D
ownl
oade
d fr
om w
ww
.wor
ldsc
ient
ific
.com
by N
AT
ION
AL
CH
UN
G H
SIN
G U
NIV
ER
SIT
Y o
n 03
/28/
14. F
or p
erso
nal u
se o
nly.
(A)
(B)
(C) (D)
Figure 3. Brazilin inhibits VSMC migration and the associated adhesion molecule expression induced by PDGF-BB. (A) VSMCs migrated from wound edge following culturing with or without PDGF-BB in the presence orabsence of Brazilin for 20 h. The upper and lower pictures represent the images of before and 20 h after PDGF-BBstimulation, respectively. The yellow frame in the picture represents denuded area. These pictures are repre-
sentative of at least three independent experiments. (B), (C) Representative images and number of migratedVSMCs in Transwell assay after treatment with or without PDGF-BB in the presence or absence of Brazilin for8 h. These pictures are representative of at least three independent experiments. Data are expressed as
mean� S:E:M. ##p < 0:01, vs. untreated control; **p < 0:01, vs. PDGF-BB control. (D) Protein levels ofICAM-1 and VCAM-1 in cell lysate and enzymatic activity of MMP-9 in conditioned medium. VSMCs werestimulated with PDGF-BB for 24 h following Brazilin pretreatment at indicated concentration, then the enzymatic
activity of MMP-9 in the conditioned medium were measured by gelatin zymography assay. The protein levels ofICAM-1 and VCAM-1 in cell lysate was determined by Western blot. GAPDH was used as an internal control.WB, Western-blot; Zym, Zymograph.
BRAZILIN INHIBITS VSMC PROLIFERATION AND MIGRATION 1289
Am
. J. C
hin.
Med
. 201
3.41
:128
3-12
96. D
ownl
oade
d fr
om w
ww
.wor
ldsc
ient
ific
.com
by N
AT
ION
AL
CH
UN
G H
SIN
G U
NIV
ER
SIT
Y o
n 03
/28/
14. F
or p
erso
nal u
se o
nly.

(1–30�M) concentration-dependently decreased the PDGF-BB induced expression ofcyclin E and CDK2. p27, an important negative regulator of CDK2/cyclin E, was upre-gulated by Brazilin in PDGF-BB stimulated VSMCs. The protein level of cyclin D was notaffected by Brazilin.
(A)
(B) (C)
(D) (E)
Figure 4. Brazilin induces a cell cycle arrest in PDGF-BB stimulated VSMCs. Cells were exposed to various
concentrations of Brazilin for 24 h in serium-free medium, and then stimulated with or without PDGF-BB foranother 24 h. (A), (B) Individual nuclear DNA content was as reflected by fluorescence intensity of incorporatedpropidium iodide. (n ¼ 3). Bra ¼ Brazilin. Cell cycle regulatory proteins CyclinD (D), Cyclin E (E), CDK2 (F)
and p27 (G) were detected by Western blot. Representative graphs of Western blots (C). GAPDH was used as aninternal control. The relative level of these proteins is expressed as fold increase compared to that of the untreatedcontrol, which was given an arbitrary value of 1 (C). Data are expressed as mean� S:E:M: ðn ¼ 4Þ. #p < 0:05 vs.
untreated control; *p < 0:05, **p < 0:01vs. PDGF-BB control.
1290 J. GUO et al.
Am
. J. C
hin.
Med
. 201
3.41
:128
3-12
96. D
ownl
oade
d fr
om w
ww
.wor
ldsc
ient
ific
.com
by N
AT
ION
AL
CH
UN
G H
SIN
G U
NIV
ER
SIT
Y o
n 03
/28/
14. F
or p
erso
nal u
se o
nly.

(F) (G)
Fig. 4. (Continued )
(A) (B)
(C) (D)
Figure 5. Brazilin inhibits PDGF-Rβ – ERK1/2 and Akt signaling cascades activated by PDGF-BB in VSMCs.Synchronized VSMCs were pre-incubated with Brazilin at different concentrations for 2 h and then stimulated with20 ng/mL PDGF-BB for 1min for PDGF-Rβ, 5min for Src and ERK1/2, and 15min for Akt. Representative
graphs of data from three independent experiments are presented (top panel). The phosphorylation levels ofPDGF-Rβ (A), Src (B), Akt (C) and ERK1/2 (D) are expressed as fold increase compared to that of untreatedcontrol (bottom panel). Data are shown as the mean� S:E:M: from three independent experiments. ##p < 0:01,
vs. untreated control; *p < 0:05, **p < 0:01, vs. PDGF-BB control.
BRAZILIN INHIBITS VSMC PROLIFERATION AND MIGRATION 1291
Am
. J. C
hin.
Med
. 201
3.41
:128
3-12
96. D
ownl
oade
d fr
om w
ww
.wor
ldsc
ient
ific
.com
by N
AT
ION
AL
CH
UN
G H
SIN
G U
NIV
ER
SIT
Y o
n 03
/28/
14. F
or p
erso
nal u
se o
nly.

Brazilin Inhibits the Increase in PDGF-Rβ, Src, Akt, ERK1/2 PhosphorylationInduced by PDGF-BB in VSMCs
The signaling pathways that might be involved in VSMC proliferation and migration werethen measured. Results showed that Brazilin markedly inhibited PDGF-BB induced-PDGF-Rβ activation (Fig. 5A) in a concentration-dependent manner in VSMCs. Thesimilar blockade of the downstream kinases activation including Src, Akt and ERK1/2were also observed (Figs. 5B–5D).
Discussion
VSMC proliferation and migration are crucial events involved in the pathophysiology ofvascular diseases including atherosclerosis and restenosis (Louis and Zahradka, 2010).PDGF-BB is one of the most potent mitogens and chemoattractants for VSMCs and playsthe central role in provoking restenosis. We now have clearly demonstrated that Brazilininhibits PDGF-BB induced VSMC proliferation and migration without influencing VSMCviability.
During the formation of neointima, the processes of VSMC proliferation and migrationare interrelated. Cell adhesion molecules ICAM-1, VCAM-1 (Dong et al., 2010; Koonet al., 2011) and MMP-9 have been reported to be major regulators in VSMC migrationand neointima formation (Cho and Reidy, 2002; Li et al., 2011; Karki et al., 2013). Thepresent study demonstrated that Brazilin inhibits PDGF-induced VSMC migration as wellas VSMC proliferation. The inhibitory effects may at least partially be due to the sup-pression of the increased expression of migration-related proteins induced by PDGF.
Cell cycle progression is tightly regulated through a complex network of positive andnegative regulatory molecules, including cyclins, cyclin-dependent kinases (CDKs) andcyclin-dependent kinase inhibitor (CKIs). Cyclin D, cyclin E, CDK2, CDK4 and CDK6 areknown to play an important positive role in the G0/G1 transition; p27 is a negativeregulator of CDK2/cyclin E and can block cell cycle in G0/G1 phase (Liang et al., 2008;Dong et al., 2010). A previous study reported that Brazilin attenuated the expression ofcyclin D1, cyclin B1, and cyclin E, as well as activated p21 and p27 in U266 cells (Kimet al., 2012). Here, we showed that Brazilin pretreatment leads to a down-regulation ofcyclin E/CDK2 expression and an up-regulation of p27 in PDGF-induced VSMCs. Themodulation of both the key positive and negative regulators of G1/S transduction indicatesthat multifaceted effects on multiple target molecules by Brazilin contribute to the anti-proliferative effects of it on PDGF-stimulated VSMCs.
PDGF-BB binds to the extracellular domain of the PDGF receptor and subsequentlyincreases the receptor’s kinase activity, which in turn recruits and phosphorylates multipleintracellular substrates such as Src family kinase, phosphatidylinositol 3-kinase (PI3K),Ras-GAP, etc. (Kazlauskas, 1994). The phosphorylated PDGF receptor and/or its sub-strates then associate with intracellular signaling enzymes. Inhibitors of PDGF signalinghave successfully conferred resistance to the pathological processes associated with pro-liferative vascular diseases such as atherosclerosis and restenosis. Directly targeting PDGF
1292 J. GUO et al.
Am
. J. C
hin.
Med
. 201
3.41
:128
3-12
96. D
ownl
oade
d fr
om w
ww
.wor
ldsc
ient
ific
.com
by N
AT
ION
AL
CH
UN
G H
SIN
G U
NIV
ER
SIT
Y o
n 03
/28/
14. F
or p
erso
nal u
se o
nly.

with an anti-PDGF antibody (Englesbe et al., 2004) or using PDGF-R inhibitors such asimatinib mesilate can attenuate neointimal development (Makiyama et al., 2008; Masudaet al., 2011). MAPK/ERK1/2 and PI3K/Akt are two major PDGF signaling pathwaysthat contribute to VSMC proliferation and migration (Dong et al., 2010; Moon et al.,2012). PDGF-BB potently activates MAPK/ERK by triggering Ras-Raf activation inVSMCs. Either an ERK1/2 inhibitor (Pyles et al., 1997) or the gene transfer of an ERK1/2dominant-negative mutant (Izumi et al., 2001) can inhibit VSMC proliferation and preventneointimal formation in balloon-injured arteries. PI3K/Akt signaling is triggered by PDGF-BB and is involved in PDGF-BB induced migration and proliferation in VSMCs (Li et al.,2001; Kim and Yun, 2007). Moreover, the nonreceptor tyrosine kinase Src family kinasesmay act as upstream kinases of Akt and MAPK/ERK signaling cascades (Bromann et al.,2004; Lee et al., 2007). Numerous studies demonstrated that Src family kinases play anessential role in PDGF-BB induced VSMC cell progression and migration (Kazlauskas,1994; Waltenberger et al., 1999; Yamboliev et al., 2001; Bromann et al., 2004; Veraciniet al., 2006). In the current study, phosphorylation of PDGF-Rβ, Src, ERK1/2 and Aktkinases were induced by PDGF-BB. VSMCs pretreated with Brazilin inhibited the phos-phorylation of them in a concentration-dependent manner. These results indicate thatinhibiting PDGF-BB induced activation of PDGF-Rβ and its downstream signaling path-way may have contributed to the inhibition of VSMC proliferation and migration exertedby Brazilin.
In conclusion, the present study provides the first scientific evidence that Brazilin has aninhibitory effect on PDGF-BB induced VSMC proliferation and migration. Suppression ofthe increased expression in migration-related proteins induced by PDGF, G0/G1 phase cellcycle arrest, and the suppression of PDGF-Rβ signaling cascades may lead to the inhibition.In addition, the down-regulation of cyclin E-CDK2 expression and up-regulation of p27expression may result in G0/G1 cell cycle arrest. This study provides a novel insight into thebiological activities of Brazilin on VSMC proliferation and migration underlying neointimaformation, which suggests a potential therapeutic application of Brazilin in the treatment ofvascular diseases though further study it still needs to be confirmed in vivo.
Acknowledgments
This study was supported by the national scientific and technological major special project“significant creation of new drugs” (2013ZX09103001-008, 2012ZX09103101-078) andspecial foundation on scientific and technological basic work (No. 2007FY130100).
References
Augulis, V. and J. Sepinwall. Brazilin-toluidine blue O and hematoxylin-darrow red methods forbrain and spinal cord. Stain Technol. 44: 131–137, 1969.
Bae, I.K., H.Y. Min, A.R. Han, E.K. Seo and S.K. Lee. Suppression of lipopolysaccharide-inducedexpression of inducible nitric oxide synthase by brazilin in RAW 264.7 macrophage cells. Eur.J. Pharmacol. 513: 237–242, 2005.
BRAZILIN INHIBITS VSMC PROLIFERATION AND MIGRATION 1293
Am
. J. C
hin.
Med
. 201
3.41
:128
3-12
96. D
ownl
oade
d fr
om w
ww
.wor
ldsc
ient
ific
.com
by N
AT
ION
AL
CH
UN
G H
SIN
G U
NIV
ER
SIT
Y o
n 03
/28/
14. F
or p
erso
nal u
se o
nly.

Bornfeldt, K.E., E.W. Raines, L.M. Graves, M.P. Skinner, E.G. Krebs and R. Ross. Platelet-derivedgrowth factor. Distinct signal transduction pathways associated with migration versus pro-liferation. Ann. NY Acad. Sci. 766: 416–430, 1995.
Bromann, P.A., H. Korkaya and S.A. Courtneidge. The interplay between Src family kinases andreceptor tyrosine kinases. Oncogene 23: 7957–7968, 2004.
Chang, Y., S.K. Huang, W.J. Lu, C.L. Chung, W.L. Chen, S.H. Lu, K.H. Lin and J.R. Sheu. Brazilinisolated from Caesalpinia sappan L. acts as a novel collagen receptor agonist in humanplatelets. J. Biomed. Sci. 20: 4, 2013.
Cho, A. and M.A. Reidy. Matrix metalloproteinase-9 is necessary for the regulation of smooth musclecell replication and migration after arterial injury. Circ. Res. 91: 845–851, 2002.
Dong, L.H., J.K. Wen, S.B. Miao, Z. Jia, H.J. Hu, R.H. Sun, Y. Wu and M. Han. Baicalin inhibitsPDGF-BB-stimulated vascular smooth muscle cell proliferation through suppressingPDGFRbeta-ERK signaling and increase in p27 accumulation and prevents injury-inducedneointimal hyperplasia. Cell Res. 20: 1252–1262, 2010.
Dzau, V.J., R.C. Braun-Dullaeus and D.G. Sedding. Vascular proliferation and atherosclerosis: newperspectives and therapeutic strategies. Nat. Med. 8: 1249–1256, 2002.
Englesbe, M.J., S.M. Hawkins, P.C. Hsieh, G. Daum, R.D. Kenagy and A.W. Clowes. Concomitantblockade of platelet-derived growth factor receptors alpha and beta induces intimal atrophy inbaboon PTFE grafts. J. Vasc. Surg. 39: 440–446, 2004.
Fang, L.H., Y.H. Zhang, J.J. Ma, G.H. Du, B.S. Ku, H.Y. Yao, Y.P. Yun and T.J. Kim. Inhibitoryeffects of tetrandrine on the serum- and platelet-derived growth factor-BB-induced prolifer-ation of rat aortic smooth muscle cells through inhibition of cell cycle progression, DNAsynthesis, ERK1/2 activation and c-fos expression. Atherosclerosis 174: 215–223, 2004.
Huang, M., J.B. Duhadaway, G.C. Prendergast and L.D. Laury-Kleintop. RhoB regulates PDGFR-beta trafficking and signaling in vascular smooth muscle cells. Arterioscler. Thromb. Vasc.Biol. 27: 2597–2605, 2007.
Izumi, Y., S. Kim, M. Namba, H. Yasumoto, H. Miyazaki, M. Hoshiga, Y. Kaneda, R. Morishita, Y.Zhan and H. Iwao. Gene transfer of dominant-negative mutants of extracellular signal-regulated kinase and c-Jun NH2-terminal kinase prevents neointimal formation in balloon-injured rat artery. Circ. Res. 88: 1120–1126, 2001.
Karki, R., E.R. Jeon and D.W. Kim. Nelumbo nucifera leaf extract inhibits neointimal hyperplasiathrough modulation of smooth muscle cell proliferation and migration. Nutrition 29: 268–275,2013.
Kazlauskas, A. Receptor tyrosine kinases and their targets. Curr. Opin. Genet. Dev. 4: 5–14, 1994.Kim, B., S.H. Kim, S.J. Jeong, E.J. Sohn, J.H. Jung, M.H. Lee and S.H. Kim. Brazilin induces
apoptosis and G2/M arrest via inactivation of histone deacetylase in multiple myeloma U266cells. J. Agric. Food Chem. 60: 9882–9889, 2012.
Kim, T.J. and Y.P. Yun. Antiproliferative activity of NQ304, a synthetic 1,4-naphthoquinone, ismediated via the suppressions of the PI3K/Akt and ERK1/2 signaling pathways in PDGF-BB-stimulated vascular smooth muscle cells. Vascul. Pharmacol. 46: 43–51, 2007.
Koon, C.M., K.S. Woo, P.C. Leung and K.P. Fung. Salviae Miltiorrhizae Radix and PuerariaeLobatae Radix herbal formula mediates anti-atherosclerosis by modulating key atherogenicevents both in vascular smooth muscle cells and endothelial cells. J. Ethnopharmacol. 138:175–183, 2011.
Lee, C.C., C.N. Wang, J.J. Kang, J.W. Liao, B.L. Chiang, H.C. Chen, C.M. Hu, C.D. Lin, S.H. Huangand Y.T. Lai. Antiallergic asthma properties of brazilin through inhibition of TH2 responses inT cells and in a murine model of asthma. J. Agric. Food Chem. 60: 9405–9414, 2012.
Lee, C.K., H.M. Lee, H.J. Kim, H.J. Park, K.J. Won, H.Y. Roh, W.S. Choi, B.H. Jeon, T.K. Park andB. Kim. Syk contributes to PDGF-BB-mediated migration of rat aortic smooth muscle cells viaMAPK pathways. Cardiovasc. Res. 74: 159–168, 2007.
1294 J. GUO et al.
Am
. J. C
hin.
Med
. 201
3.41
:128
3-12
96. D
ownl
oade
d fr
om w
ww
.wor
ldsc
ient
ific
.com
by N
AT
ION
AL
CH
UN
G H
SIN
G U
NIV
ER
SIT
Y o
n 03
/28/
14. F
or p
erso
nal u
se o
nly.

Lee, D.Y., M.K. Lee, G.S. Kim, H.J. Noh and M.H. Lee. Brazilin inhibits growth and inducesapoptosis in human glioblastoma cells. Molecules 18: 2449–2457, 2013.
Lee, J.J., W.Y. Zhang, H. Yi, Y. Kim, I.S. Kim, G.N. Shen, G.Y. Song and C.S. Myung. Anti-proliferative actions of 2-decylamino-5,8-dimethoxy-1,4-naphthoquinone in vascular smoothmuscle cells. Biochem. Biophys. Res. Commun. 411: 213–218, 2011.
Levitzki, A. PDGF receptor kinase inhibitors for the treatment of restenosis. Cardiovasc. Res. 65:581–586, 2005.
Li, L., H.N. Zhang, H.Z. Chen, P. Gao, L.H. Zhu, H.L. Li, X. Lv, Q.J. Zhang, R. Zhang, Z. Wang,Z.G. She, R. Zhang, Y.S. Wei, G.H. Du, D.P. Liu and C.C. Liang. SIRT1 acts as a modulatorof neointima formation following vascular injury in mice. Circ. Res. 108: 1180–1189, 2011.
Li, X.A., C. Bianchi and F.W. Sellke. Rat aortic smooth muscle cell density affects activation of MAPkinase and Akt by menadione and PDGF homodimer BB. J. Surg. Res. 100: 197–204, 2001.
Liang, K.W., S.C. Yin, C.T. Ting, S.J. Lin, C.M. Hsueh, C.Y. Chen and S.L. Hsu. Berberine inhibitsplatelet-derived growth factor-induced growth and migration partly through an AMPK-dependent pathway in vascular smooth muscle cells. Eur. J. Pharmacol. 590: 343–354, 2008.
Lo, H.M., Y.J. Tsai, W.Y. Du, C.J. Tsou and W.B. Wu. A naturally occurring carotenoid, lutein,reduces PDGF and H2O2 signaling and compromised migration in cultured vascular smoothmuscle cells. J. Biomed. Sci. 19: 18, 2012.
Louis, S.F. and P. Zahradka. Vascular smooth muscle cell motility: from migration to invasion. Exp.Clin. Cardiol. 15: e75–85, 2010.
Makiyama, Y., K. Toba, K. Kato, S. Hirono, T. Ozawa, T. Saigawa, S. Minagawa, M. Isoda,F. Asami, N. Ikarashi, M. Oda, M. Moriyama, M. Higashimura, T. Kitajima, K. Otaki andY. Aizawa. Imatinib mesilate inhibits neointimal hyperplasia via growth inhibition of vascularsmooth muscle cells in a rat model of balloon injury. Tohoku J. Exp. Med. 215: 299–306,2008.
Masuda, S., K. Nakano, K. Funakoshi, G. Zhao, W. Meng, S. Kimura, T. Matoba, M. Miyagawa,E. Iwata, K. Sunagawa and K. Egashira. Imatinib mesylate-incorporated nanoparticle-elutingstent attenuates in-stent neointimal formation in porcine coronary arteries. J. Atheroscler.Thromb. 18: 1043–1053, 2011.
Mitani, K., F. Takano, T. Kawabata, A.E. Allam, M. Ota, T. Takahashi, N. Yahagi, C. Sakurada,S. Fushiya and T. Ohta. Suppression of melanin synthesis by the phenolic constituents ofsappanwood (Caesalpinia sappan). Planta Med. 79: 37–44, 2013.
Mok, M.S., S.D. Jeon, K.M. Yang, D.S. So and C.K. Moon. Effects of Brazilin on induction ofimmunological tolerance by sheep red blood cells in C57BL/6 female mice. Arch. Pharm. Res.21: 769–773, 1998.
Moon, C.K., K.S. Park, S.G. Kim, H.S. Won and J.H. Chung. Brazilin protects cultured rat hep-atocytes from BrCCl3-induced toxicity. Drug Chem. Toxicol. 15: 81–91, 1992.
Moon, C.K., S.H. Lee, M.O. Lee and S.G. Kim. Effects of Brazilin on glucose oxidation, lipogenesisand therein involved enzymes in adipose tissues from diabetic KK-mice. Life Sci. 53: 1291–1297, 1993.
Moon, C.Y., C.R. Ku, Y.H. Cho and E.J. Lee. Protocatechuic aldehyde inhibits migration andproliferation of vascular smooth muscle cells and intravascular thrombosis. Biochem. Biophys.Res. Commun. 423: 116–121, 2012.
Pyles, J.M., K.L. March, M. Franklin, K. Mehdi, R.L. Wilensky and L.P. Adam. Activation of MAPkinase in vivo follows balloon overstretch injury of porcine coronary and carotid arteries. Circ.Res. 81: 904–910, 1997.
Reddy, M.A., S. Sahar, L.M. Villeneuve, L. Lanting and R. Natarajan. Role of Src tyrosine kinase inthe atherogenic effects of the 12/15-lipoxygenase pathway in vascular smooth muscle cells.Arterioscler. Thromb. Vasc. Biol. 29: 387–393, 2009.
BRAZILIN INHIBITS VSMC PROLIFERATION AND MIGRATION 1295
Am
. J. C
hin.
Med
. 201
3.41
:128
3-12
96. D
ownl
oade
d fr
om w
ww
.wor
ldsc
ient
ific
.com
by N
AT
ION
AL
CH
UN
G H
SIN
G U
NIV
ER
SIT
Y o
n 03
/28/
14. F
or p
erso
nal u
se o
nly.

Rothmeier, A.S., I. Ischenko, J. Joore, D. Garczarczyk, R. Furst, C.J. Bruns, A.M. Vollmar andS. Zahler. Investigation of the marine compound spongistatin 1 links the inhibition ofPKCalpha translocation to nonmitotic effects of tubulin antagonism in angiogenesis. FASEB J.23: 1127–1137, 2009.
Sasaki, Y., M. Suzuki, T. Matsumoto, T. Hosokawa, T. Kobayashi, K. Kamata and S. Nagumo.Vasorelaxant activity of Sappan Lignum constituents and extracts on rat aorta and mesentericartery. Biol. Pharm. Bull. 33: 1555–1560, 2010.
Sun, L., R. Zhao, L. Zhang, T. Zhang, W. Xin, X. Lan, C. Huang and G. Du. Salvianolic acid Ainhibits PDGF-BB induced vascular smooth muscle cell migration and proliferation while doesnot constrain endothelial cell proliferation and nitric oxide biosynthesis. Molecules 17: 3333–3347, 2012.
Tsang, F., H.H. Choo, G.S. Dawe and W.S. Wong. Inhibitors of the tyrosine kinase signaling cascadeattenuated thrombin-induced guinea pig airway smooth muscle cell proliferation. Biochem.Biophys. Res. Commun. 293: 72–78, 2002.
Veracini, L., M. Franco, A. Boureux, V. Simon, S. Roche and C. Benistant. Two distinct pools of Srcfamily tyrosine kinases regulate PDGF-induced DNA synthesis and actin dorsal ruffles. J. Cell.Sci. 119: 2921–2934, 2006.
Waltenberger, J., A. Uecker, J. Kroll, H. Frank, U. Mayr, J.D. Bjorge, D. Fujita, A. Gazit,V. Hombach, A. Levitzki and F.D. Bohmer. A dual inhibitor of platelet-derived growth factorbeta-receptor and Src kinase activity potently interferes with motogenic and mitogenicresponses to PDGF in vascular smooth muscle cells. A novel candidate for prevention ofvascular remodeling. Circ. Res. 85: 12–22, 1999.
Yamboliev, I.A., J. Chen and W.T. Gerthoffer. PI 3-kinases and Src kinases regulate spreading andmigration of cultured VSMCs. Am. J. Physiol. Cell Physiol. 281: C709–718, 2001.
1296 J. GUO et al.
Am
. J. C
hin.
Med
. 201
3.41
:128
3-12
96. D
ownl
oade
d fr
om w
ww
.wor
ldsc
ient
ific
.com
by N
AT
ION
AL
CH
UN
G H
SIN
G U
NIV
ER
SIT
Y o
n 03
/28/
14. F
or p
erso
nal u
se o
nly.