Embed Size (px)
Citation preview

Vol. 41, No. 2JOURNAL OF VIROLOGY, Feb. 1982, p. 692-7020022-538X/82/020692-11$02.00/0
Inhibition of Sindbis Virus Maturation After Treatment ofInfected Cells with Trypsin
RAQUEL H. ADAMS AND DENNIS T. BROWN*Cell Research Institute and Department of Microbiology, University of Texas at Austin, Austin, Texas 78712
Received 21 May 1981/Accepted 14 September 1981
Brief treatment of Sindbis virus-infected BHK-21 or Vero cells with lowconcentrations of trypsin irreversibly blocked further production of progenyvirions after removal of the enzyme. The inhibitory effects of the trypsintreatment could only be demonstrated in cells in which virus infection wasestablished; optimal inhibition occurred at ca. 3 h postinfection. Production ofvirus structural proteins PE2, El, and C occurred at normal levels in inhibitedcells. PE2 and El were also transported to the cell plasma membrane duringinhibition; however, PE2 was not cleaved to E2, and little capsid protein becamemembrane associated relative to control cells. Although trypsin treatment had noeffect on Sindbis protein synthesis, the production of both 26S and 42S RNA wasgreatly reduced. Similar trypsin treatment of BHK cells infected with vesicularstomatitis virus had no detectable effect on the course of virus infection.
Sindbis virus, the alphavirus prototype, iscomposed of three virus-coded structural pro-teins (28, 30). One of these proteins, the capsidprotein (C), associates with the virus RNA toform a protein-nucleic acid complex, the nucleo-capsid, which has icosahedral symmetry (38).The two remaining proteins (E1 and E2) areassociated with a membrane bilayer which en-closes the nucleocapsid (5, 35). The virus mem-brane proteins are glycosylated by host cellenzymes and are thus hybrid structures derivedfrom both host and virus origin (21). The virusparticle is itself a hybrid aggregate, since thelipids which form the virus membrane are ac-quired in the final budding stages of assemblyfrom preexisting host cell membranes (12, 22).A wealth of experimentation has been con-
ducted to investigate the biochemical eventsassociated with the production and maturationof Sindbis virus proteins (3, 28, 35). Thesestudies, together with numerous studies involv-ing other model enveloped viruses, have led tothe conclusion that the events involved in proc-essing virus membrane proteins are similar, ifnot identical, to those involved in processinghost cell membrane proteins. The three Sindbisvirus structural proteins are derived from asingle precursor polyprotein (29, 31). This pre-cursor is processed by proteolysis into the cap-sid protein, one of the envelope proteins (E1),and a precursor to the second envelope protein(PE2). It has been demonstrated that PE2 and E1are integrated into membranes of the roughendoplasmic reticulum (9, 40) and are then trans-ported to the smooth membranes and eventuallyto the plasma membrane before the final budding
events (2, 4, 9, 25). Modification of the proteinsby glycosylation (7, 21, 34) and lipidation (32,33) takes place while these proteins are en routeto the cell surface. The capsid protein combineswith virus RNA to form the nucleocapsid in thecytoplasm of the host cell. Nucleocapsid struc-tures then become associated with the portion ofthe PE2 polypeptide which is exposed to thecytoplasm after PE2 has reached the plasmamembrane (9). At this point, the conversion ofPE2 to E2 takes place, and envelopment of thecapsid in the virus-modified cell plasma mem-brane is initiated. The process of envelopment isintimately associated with the conversion of PE2to E2, and, in every instance in which thisconversion is blocked, virus envelopment doesnot take place (2, 5, 9, 14, 15, 25, 36).Many host functions are involved in the pro-
duction and maturation of Sindbis virus compo-nents. The extensive exploitation of the hostprotein synthetic and processing apparatus hasmade viruses such as Sindbis excellent modelsfor the study of normal cell processes. In spite ofthe many obvious functional roles the host cellplays in Sindbis development, it is well knownthat cell viability is not necessary for the produc-tion of large amounts of virus. Pretreatment ofcells with high concentrations of actinomycin D(an inhibitor of DNA-dependent RNA polymer-ase) (26) or enucleation (17, 19) of cells does notdestroy the ability of vertebrate cells to produceprogeny virus. That vertebrate cell viability isnot essential is emphasized by the fact thatalphavirus infection of these cells results in anarrest of host cell macromolecular synthesis afew hours after infection (39). These observa-
692
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 21
Oct
ober
202
1 by
188
.168
.178
.115
.

SINDBIS VIRUS MATURATION 693
tions suggest that, in the vertebrate cell, rela-tively stable host functions established beforeinfection are utilized by infecting virions forreplication.
Invertebrate cells respond quite differently toSindbis virus infection. Host cell macromolecu-lar synthesis and growth are not arrested duringinfection; rather, virus production coexists withhost functions, resulting in a true persistentinfection (13, 24). In striking contrast to infectedvertebrate cells, we have found that Sindbisvirus production in invertebrate cells is sensitiveto actinomycin D (26). In the invertebrate sys-tem, the synthesis of viral structural proteins,including E2, continues in the presence of acti-nomycin D, although the assembly of the Sind-bis virus components into mature virus is inhib-ited. We interpreted these data as suggestingthat, in contrast to vertebrate cells, invertebratecells require the continued synthesis of an unsta-ble function, possibly of host origin, for theassembly of virus components into mature viri-ons.We have continued in our attempts to estab-
lish the existence of host functions essential tothe later stages of alphavirus maturation in ver-tebrate cells by seeking methods of alteringpossible host functions after the cell has lost theability to replace them. In this report, we de-scribe experiments examining the effects of tryp-sin treatment of vertebrate cells on their abilityto produce Sindbis virus. The proteolytic en-zyme trypsin was chosen for this study since it isknown that this agent has no assayable effect oncell viability. The enzymatic activity of trypsincan also be completely inhibited by soybeantrypsin inhibitor (10), thereby allowing studieson the effects of limited proteolysis in the ab-sence of continued enzymatic activity.
MATERIALS AND METHODS
Cells, virus, and media. BHK-21 cells were grown inEagle minimal essential medium (MEM; 8) supple-mented with 10% fetal calf serum (GIBCO, GrandIsland, N.Y.), 2 mM glutamine, and 10% tryptosephosphate broth, as described previously (9). Heat-resistant Sindbis virus, which served as wild type, andvesicular stomatitis virus (VSV), Indiana strain, wereprovided by E. Pfefferkorn (Dartmouth Medical Col-lege, Hanover, N.H.). Virus stocks were prepared andtitrated in BHK cells as previously described (23).
Infection of cells and radioactive labeling procedures.To minimize host cell protein and RNA synthesis,cells were pretreated for 90 min at 37°C with EagleMEM containing 4 ,ug of actinomycin D per ml andmaintained in the drug throughout infection. Themonolayers were infected with 100 PFU of wild-type(heat-resistant Sindbis virus) per cell, as has beendescribed (23). Experiments with VSV were carriedout by infecting BHK cells with 50 PFU per cell. Allother procedures involving VSV were as describedwith Sindbis, except where noted. Infected cells were
incubated at 37°C for 2 h in Eagle MEM supplementedwith 1% fetal calf serum and starved for methionine inmethionine-free Eagle MEM as described previously(9). Monolayers were labeled as indicated in the textwith 40 ,uCi of [35S]methionine (1,100 Ci/mmol; Amer-sham Corp., Arlington Heights, and G. D. Searle andCo., Chicago, Ill.) per ml in MEM. In pulse-chaseexperiments, infected monolayers were labeled with100 p.Ci of [35S]methionine per ml for 10 min, washed,and chased in warm media containing 50 ,ug of cyclo-heximide (Sigma Chemical Co., St. Louis, Mo.) per mland 3 mM methionine. Cells and chase media wereharvested by washing once with phosphate-bufferedsaline deficient in calcium and magnesium (PBS-D).Cells were separated from the supernatants by centrif-ugation at 1,000 rpm for 5 min and washed once withPBS-D.
Purification of labeled virus. The presence of radio-active virus in the chase medium was assayed withdensity gradient centrifugation in 15 to 35% (wt/wt)linear potassium tartrate gradients, according to rou-tine procedure (9).
Trypsinization procedures. Trypsin stocks weremade by dissolving 3 x crystallized trypsin (Worthing-ton Diagnostics, Freehold, N.J.) to a concentration of0.4 mg/ml in phosphate-buffered saline. Trypsin solu-tions were sterilized by ultrafiltration and stored at-20°C. The amount of soybean trypsin inhibitor (Wor-thington Diagnostics) necessary to inhibit the action oftrypsin was determined spectrophotometrically withthe benzoyl-L-arginine ethyl ester assay (10). Theconcentration of trypsin necessary to demonstrate theinhibitory effect on Sindbis virus maturation was de-termined as follows. Monolayers of uninfected cellswere treated with 0.5-log increment dilutions of en-zyme and monitored for morphological changes withphase-contrast microscopy. Identical dilutions of tryp-sin were used to treat Sindbis virus-infected cells at 3 hafter infection. These cells were then labeled at 37°Cwith 25 ,uCi of [35S]methionine per ml for 2 h. Theinhibitory effect of trypsin on virus production wasassayed by determining the incorporation of the labelinto physical and infectious virus particles. A trypsinconcentration of 50 ,ug/ml was found to be optimalsince this concentration produced minimal morpholog-ical changes in the cell monolayers while completelyblocking incorporation of label into mature virus andreducing yields of infectious virions (see below). Tryp-sin concentrations of 50 to 400 ,ug/ml were found to beinhibitory to Sindbis virus production without inhibit-ing the production of virus protein (see below).
In the experiments described below, the infectedmonolayers were washed once with warm PBS-D andtreated with warm trypsin (25°C) at a concentration of50 pLg/ml in PBS-D for 5 min. Trypsin activity wasarrested by the addition of soybean trypsin inhibitor(at a concentration of 50 ,ug/ml in PBS-D) in twice thevolume required to neutralize the trypsin. The cellswere washed from the mixture by dilution in phos-phate-buffered saline and centrifugation at low speed.The cells were suspended in complete Eagle mediumand labeled in suspension. Three hours postinfectionwas chosen as the time to treat the cells with trypsin inmost of the experiments described below, as this wasthe earliest time at which the maximum inhibitoryeffect was exhibited (see Fig. 2 and Discussion).Control experiments demonstrated that these concen-
VOL. 41, 1982
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 21
Oct
ober
202
1 by
188
.168
.178
.115
.

694 ADAMS AND BROWN
trations of trypsin or soybean trypsin inhibitor or amixture of both would not reduce the infectivity (asdetermined by plaque assay) of a standard preparationof Sindbis virus (Fig. 1).
Polyacrylamide gel electrophoresis. Analysis of 3S-labeled proteins of Sindbis virus- and VSV-infectedcells was carried out by electrophoresis of the proteinsthrough sodium dodecyl sulfate containing 11% poly-acrylamide gels by the method of Laemmli (18). Cellsor virus was prepared in sample buffer and electro-phoresed as described previously (9, 36). Fluorogra-phy of the gels was performed by the method ofBonner and Laskey (1). Gels were dried and exposedto Kodak X-Omat RP X-ray film.Membrane fractionation. Cells were pretreated with
actinomycin D and infected as described above. Cellmonolayers were treated with trypsin at 3 h postinfec-tion, pulse-labeled for 10 min, and chased for theindicated times at 37°C, as detailed in Results. Mem-brane fractionation was by the protocol of Erwin andBrown (9), omitting the PBS-D washes before homog-enization. Cells were placed directly into E-T buffer
16
14 -
12-
I 10 20FRACTION NO.
FIG. 1. Production of Sindbis virus particles bytrypsin-treated and untreated BHK-21 cells. Sindbisvirus-infected BHK-21 cells were treated with trypsin([1) or mock treated (0) as described in the text. Aftertreatment, the cells were labeled with [35S]methionine,and virus released into the media was concentrated oncontinuous density gradients as described in the text.A, Infected cells treated with soybean trypsin inhibitoralone. The supernatant of unirnfected cells, which hadbeen treated with trypsin followed by inhibitor asdescribed in the text, was added to an infected mono-layer for 5 min at 3 h postinfection (0). Inhibition ofvirus production could be detected if an infectedmonolayer was treated (as described in the text) withthe inhibitor-inactivated trypsin-containing wash de-rived from another culture of infected cells (A) (ascompared with 0, above).
(0.001 M Tris-0.01 M EDTA, pH 8.0) after centrifuga-tion. Membrane fractions were pooled and dialyzedagainst two changes of water with 1 ,ug of phenyl-methyl sulfonyl fluoride per ml. Dialyzed fractionswere frozen, lyophylized, and boiled in sample bufferbefore electrophoresis.
Trichloroacetic acid precipitation of protein andRNA. Identical monolayers of cells were treated with 4,ug of actinomycin D per ml 90 min before and through-out infection. Cells were infected with Sindbis virusand treated with trypsin, followed by inhibitor or PBS-D (mock treated) at 3 h postinfection for 5 min asdescribed above. The cells were labeled directly in thetrypsin-soybean trypsin inhibitor for 3 h with either 50,uCi of [35S]methionine or 100 puCi of [3H]uridine (45Ci/mmol; Amersham Corp. and G. D. Searle) per ml.Cell monolayers were disrupted by using 1% sodiumdodecyl sulfate, and the resulting DNA mass wasdisrupted by repeated passage through a 25-gaugeneedle. Protein and RNA synthesized after trypsintreatment were determined by assaying total amountsof label incorporated into cold 15% trichloroaceticacid-insoluble material.
Extraction of 42S and 26S RNA. Cultures containingequivalent numbers of cells were treated with actino-mycin D 90 min before and throughout infection withSindbis virus. Infected cells were incubated at 37°C for3 h in fresh media, using the trypsinization protocoloutlined above. Monolayers were labeled with 75 uCiof [3H]uridine per ml in MEM plus 10% fetal calfserum for 3 h at 37°C. Equal numbers of trypsin- ormock-treated infected monolayers were swollen for 5min in cold Tris-EDTA buffer (10 mM Tris-1 mMEDTA, pH 7.4) containing 0.3% diethyl pyrocarbonate(Sigma Chemical Co.) and 50 ,ug of heparin per ml(Sigma). Samples were disrupted in the cold, using astainless-steel Dounce homogenizer (Kontes Co.,Vineland, N.J.). Cell homogenates were immediatelymade 1% with respect to sodium dodecyl sulfate andphenol extracted by using the procedure of Penman(20). The RNA precipitates generated were pelleted bycentrifugation at 15,000 rpm for 15 min at 0°C. PelletedRNA was suspended in gradient buffer (0.01 M NaCl-1mM EDTA-0.01 M Tris-0.2% sodium dodecyl sulfate-1M formamide-0.3% diethyl pyrocarbonate, pH 7.4)and loaded onto a 17-ml, 15 to 30% sucrose gradient (inthe same gradient buffer). The samples were run for 24h at 26,500 rpm at 23°C in a Sorvall AH627 rotor.Gradients were fractionated as 300-,d samples directlyinto scintillation vials, and radioactivity was deter-mined as previously described (9). Viruses harvestedafter the labeling period were run on a 10 to 35%potassium tartrate gradient. Gradients were fractionat-ed and counted as described above.
Electron microscopy. BHK-21 cells were infected asdescribed and incubated at 37°C. Cells were trypsin-ized at 3 h postinfection, as described above, or mocktreated, suspended in fresh medium, and incubated foran additional 5 h. Samples were washed with buffer(0.1 M K2HPO4-0.12 M NaCl, pH 7.2) and fixed with2.5% glutaraldehyde plus 1.5% formaldehyde in 0.1 Mphosphate buffer, pH 7.2, for 2 h at room temperature.The cells were then washed twice for 30 min in 0.1 Mphosphate buffer (pH 7.2) and postfixed with 1%osmium tetroxide in the same buffer for 2 h at roomtemperature. This treatment was followed by two 30-min washes in phosphate buffer. The samples were
J. VIROL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 21
Oct
ober
202
1 by
188
.168
.178
.115
.

SINDBIS VIRUS MATURATION 695
dehydrated in a graded ethanol series, after whichthey were imbedded in Spurr low-viscosity epoxyresin (37). Ultrathin sections were mounted on un-coated, 200-mesh copper grids and stained in saturatedethanolic uranyl acetate and lead acetate. All prepara-tions were examined and photographed with a 100CXelectron microscope (JEOL USA, Medford, Mass.).
RESULTSVirus production in trypsin-treated Sindbis- or
VSV-infected BHK-21 cells. The effects of tryp-sin treatment on the ability of VSV- and Sindbisvirus-infected BHK-21 cells to produce progenyvirus are presented in Table 1 and Fig. 1. WhenSindbis virus-infected cells were treated withtrypsin (as described in Materials and Methods)at 3 h postinfection, a striking reduction in theyields of virus produced by the cells relative tonontreated controls was observed. This reduc-tion could be measured as either infectious virus(Table 1) or as physical particles (Fig. 1). In asimilar experiment with VSV-infected BHK-21cells, trypsin treatment did not result in a reduc-tion in virus production (Table 1). The observedreduction of Sindbis virus yields by trypsintreatment of cells was more dramatic in cellswhich were infected in the presence of actino-mycin D (Table 1). This result was due, in part,to the increase in yields obtained in control cellsin the presence of this agent (26).Soybean trypsin inhibitor alone or trypsin
mixed with inhibitor before its addition to theinfected cells could not produce the inhibitoryeffect (Fig. 1). These experiments, as well as thecontrols described in Materials and Methods,show that the reduction in virus yields does notresult from the inactivation of mature virions byresidual trypsin activity.
TABLE 1. Effect of trypsin treatment on productionof infectious virus by Sindbis or VSV-infected BHK-
21 cells
Virus yield(PFU per
Type of cells cell) 10 hafter
infection
Sindbis virus-infected 1,400Trypsin-treated, Sindbis virus- 300
infectedaActinomycin D-treated, Sindbis virus- 2,300
infectedbActinomycin D-treated, trypsin-treated, 100
Sindbis virus-infectedVSV-infected 1,200VSV-infected, trypsin-treated 1,600
a Cells treated with trypsin at 3 h after infection asdescribed in the text.
b Cells treated with actinomycin D (4 ,ug/ml) 90 minbefore infection and throughout infection.
These data suggest that treatment of Sindbisvirus-infected cells, but not VSV-infected cells,with trypsin resulted in the loss of the cell'scompetence to produce either infectious orphysical virus particles. This could be achievedby two possible mechanisms: (i) the treatment ofcells with trypsin may result in the destructionby proteolysis of some function essential tovirus production through either a reorganizationof the cell surface or by direct proteolytic attack,or (ii) trypsin treatment might result in therelease from the infected cell of some agentcapable of suppressing virus production. Thefollowing experiment was done in an attempt todistinguish between these possibilities. BHK-21cells infected with Sindbis virus for 3 h weretreated with trypsin, followed by trypsin inhibi-tor as described in Materials and Methods. Atthe conclusion of the trypsin treatment protocol,the infected cells were removed from the tryp-sin-soybean inhibitor-containing mixture. Thecell-free reaction mixture (total, 5 ml) was thenapplied to an identically infected and incubated(non-trypsin treated) monolayer for a period of 5min. This monolayer was washed and labeledwith [35S]methionine as described above, andradioactive virus was purified as described inMaterials and Methods. Surprisingly, this treat-ment consistently resulted in a 50 to 60% reduc-tion in the yield of labeled virus particles (Fig.1). A control experiment was conducted by thesame protocol, except the initial trypsinizationwas performed on a mock-infected BHK mono-layer, and produced a yield of progeny virusparticles equal to that of an untreated control(Fig. 1).The results of this experiment suggest that
treatment of a Sindbis virus-infected BHKmonolayer with trypsin results in the release ofsome factor(s) capable of suppressing virus pro-duction. This factor(s) cannot be produced bysimilar treatment of uninfected cells. The natureof this inhibiting activity is currently under in-vestigation.The experiments described above and shown
in Fig. 1 and Table 1 were repeated, substitutingVero cells for BHK-21 cells. The results ob-tained were similar to those obtained with BHKcells (data not shown), indicating that this phe-nomenon is not restricted to one particular hostcell type.
Effect on virus yields of treated BHK-21 cellswith trypsin at various times before and afterinfection with Sindbis virus. The experimentsdescribed above were carried out with proteoly-sis of infected cells at 3 h postinfection, a timewhen virus infection is well established. Theresults suggest that the Sindbis virus-inhibitingactivity could only be generated by the trypsini-zation of infected cells. To determine how this
VOL. 41, 1982
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 21
Oct
ober
202
1 by
188
.168
.178
.115
.

696 ADAMS AND BROWN
activity was related to the cexperiment was carried outexposed to trypsin, as descious times relative to the tireffect of trypsin on virus prowas determined by assayingtious virus released from thafter the trypsin treatment, itrypsinization was postinfeiinfection, in those cases whewas before infection (Fig. 2if a monolayer was treatebefore or immediately beforion virus yield was observed.between infection and tr)creases, the yield of virus detory effects of the trypsin Imaximum at 2.5 to 3 h presults suggest that, whencarried out early in infectiorthe inhibitory effect on virduced.
Protein synthesis in Sirtrypsin-treated cells. The ftwas done to determine tUtreatment on virus proteiners of BHK cells were treatD for a period of 90 minSindbis virus (the effect ofonstrated in actinomycin D1]). The monolayers were
10101
0.
-l 0
HOURS
FIG. 2. Yields of Sindbis vitreated with trypsin at variousinfection. Identical monolayerwith trypsin as described in tlvirus infection (arrow). The Narrow is the yield of virus obttrypsin treated immediately aftduction was assayed 10 h afterpoints to the right of the arrow)(data point to the left of the ar
,ycle of infection, an trypsin at 3 h postinfection (Fig. 2). Subsequentin which cells were to trypsin treatment, the cells were labeled for 2ribed above, at vari- h with [35S]methionine, washed, and placed inne of infection. The normal medium containing 10 times normal con-)duction in each case centrations of nonradioactive methionine for 20the amount of infec- h at 37°C. Identical monolayers were treatedie monolayer at 10 h with buffer containing no trypsin or with bufferin those cases where containing only soybean trypsin inhibitor. At thection, or 10 h after end of the incubation period, the medium from thewre trypsin treatment monolayers was collected, layered onto continu-). It was found that, ous sucrose gradients, and centrifuged as de-d with trypsin 1 h scribed in Materials and Methods. Untreatede infection, no effect cells or cells treated with only soybean trypsinAs the time interval inhibitor produced large amounts of radioactiveypsin treatment in- progeny virus, as was expected (Fig. 1). Noecreases. The inhibi- significant amount of label could be detected intreatment were at a the virus region of the gradient prepared withbostinfection. These media from cells treated with trypsin (Fig. 1).trypsin treatment is The monolayers themselves were dissolved ini or before infection, electrophoresis buffer. Equivalent trypsin-treat-us production is re- ed and untreated monolayers were found to
contain similar amounts of trichloroacetic acid-idbis virus-infected, precipitable methionine (Table 2), indicating that)llowing experiment trypsin treatment did not affect the ability ofie effect of trypsin infected cells to produce virus protein. Electro-synthesis. Monolay- phoresis of the treated and untreated cell digests:ed with actinomycin reveal that, although both monolayers producedi and infected with virus precursor proteins PE2, E1, and C, cellstrypsin can be dem- treated with trypsin accumulated PE2 and lacked-treated cells [Table E2 (Fig. 3A). These data suggest that, afterthen treated with trypsin treatment, virus protein synthesis con-
tinues normally, but that cleavage of PE2 to E2 isblocked and that this block may prevent virusenvelopment.
Kinetics of virus maturation in trypsin-treatedcells. To determine at what point after trypsintreatment virus maturation was arrested, cellswere pulse-labeled for 10 min with [35S]methio-nine (40 p,Ci/ml) immediately after and at 30 and90 min after treatment with trypsin. After thepulse, the cells were chased for 3 h more inmedia containing cycloheximide (50 Fg/ml) toarrest further protein synthesis. At the end ofthe chase period, the medium was collected fromeach experiment and subjected to density gradi-ent centrifugation as described in Materials andMethods. Label banding in the region of virusdensity is shown in Fig. 4. A 10-min pulse,
| * | followed by a 3-h chase in untreated (control)2 3 4 5 cells, resulted in significant amounts of label
chased into virions. A small amount of labelirus from BHK-21 cells could be detected in the region of the gradienttimes before and after corresponding to virus when cells were pulseds of cells were treated immediately after trypsin treatment. A de-he text before or after creased amount of label was recovered as virusalned wndicedllbywere from cells labeled 30 min after trypsin treatment,er infection. Virus pro- and no label could be detected in virus when thetrypsin treatment (data pulse was carried out at 90 min post-trypsinand 10 h after infection treatment. Polyacrylamide gel electrophoresis ofTOW). the label recovered in virus at 0 and 30 min post-
J. VIROL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 21
Oct
ober
202
1 by
188
.168
.178
.115
.
SINDBIS VIRUS MATURATION 697
TABLE 2. Protein and RNA synthesis in Sindbis virus-infected BHK-21 cells after treatment with trypsinaAmt (cpm x 1o-4) of synthesis in following
Expt type of cells:Control Trypsin-treated
Sindbis virus-infected cells ([35S]methionine incorporated 39.5 40.6into trichloroacetic acid-insoluble material)b
Sindbis virus-infected cells ([3H]uridine incorporated 12.0 1.6into trichloroacetic acid-insoluble material)c
Uninfected cells ([3H]uridine incorporated into 40.0 36.0trichloroacetic acid-insoluble material)c
a Identical monolayers of cells were treated with actinomycin D (4 p.g/ml) 90 min before and throughoutinfection. Cells were treated or mock treated with trypsin (as described in the text) at 3 h postinfection.
b Cells were labeled with [35S]methionine (50 ,XCi/ml) in methionine-free medium from 30 min after thetreatment described in footnote a until 6 h postinfection.
c Cells were labeled with tritiated uridine (100 XCi/ml) as described in footnote b.
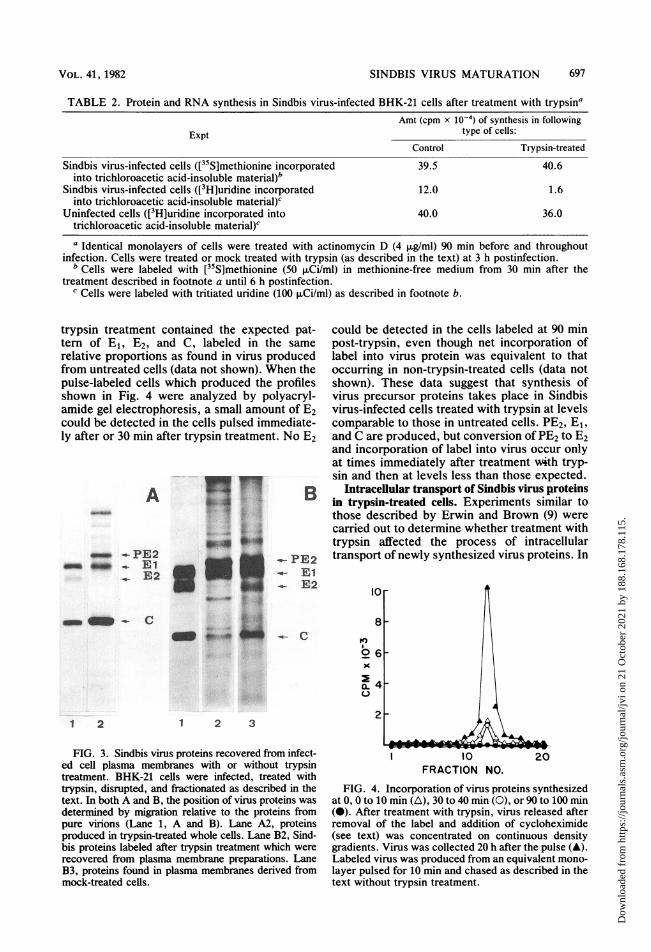
trypsin treatment contained the expected pat-tern of E1, E2, and C, labeled in the samerelative proportions as found in virus producedfrom untreated cells (data not shown). When thepulse-labeled cells which produced the profilesshown in Fig. 4 were analyzed by polyacryl-amide gel electrophoresis, a small amount of E2could be detected in the cells pulsed immediate-ly after or 30 min after trypsin treatment. No E2
A
- -PE2- - _ E1
_ E2
mm- C
1 2
lw .. p
~~o i~~~~~~~-, -~~~~~~..4
.-- PE2_- E1
E2
o
a-K
1 2 3
FIG. 3. Sindbis virus proteins recovered from infect-ed cell plasma membranes with or without trypsintreatment. BHK-21 cells were infected, treated withtrypsin, disrupted, and fractionated as described in thetext. In both A and B, the position of virus proteins wasdetermined by migration relative to the proteins frompure virions (Lane 1, A and B). Lane A2, proteinsproduced in trypsin-treated whole cells. Lane B2, Sind-bis proteins labeled after trypsin treatment which wererecovered from plasma membrane preparations. LaneB3, proteins found in plasma membranes derived frommock-treated cells.
could be detected in the cells labeled at 90 minpost-trypsin, even though net incorporation oflabel into virus protein was equivalent to thatoccurring in non-trypsin-treated cells (data notshown). These data suggest that synthesis ofvirus precursor proteins takes place in Sindbisvirus-infected cells treated with trypsin at levelscomparable to those in untreated cells. PE2, E1,and C are produced, but conversion of PE2 to E2and incorporation of label into virus occur onlyat times immediately after treatment with tryp-sin and then at levels less than those expected.
Intracellular transport of Sindbis virus proteinsin trypsin-treated cells. Experiments similar tothose described by Erwin and Brown (9) werecarried out to determine whether treatment withtrypsin affected the process of intracellulartransport of newly synthesized virus proteins. In
I0FRACTION NO.
FIG. 4. Incorporation of virus proteins synthesizedat 0, 0 to 10 min (A), 30 to 40 min (0), or 90 to 100 min(0). After treatment with trypsin, virus released afterremoval of the label and addition of cycloheximide(see text) was concentrated on continuous densitygradients. Virus was collected 20 h after the pulse (A).Labeled virus was produced from an equivalent mono-layer pulsed for 10 min and chased as described in thetext without trypsin treatment.
VOL. 41, 1982
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 21
Oct
ober
202
1 by
188
.168
.178
.115
.

698 ADAMS AND BROWN
these experiments, it was found that migrationof virus membrane proteins PE2 and E1 fromrough membranes to smooth and plasma mem-branes was not affected by treatment with tryp-sin and that similar amounts of virus label weretransported to the plasma membrane fraction inthe presence and absence of trypsin treatment(data not shown). Polyacrylamide gel electro-phoresis of the plasma membrane fraction re-vealed the presence of PE2 and E1 in thissubcellular fraction (Fig. 3B) in amounts equiva-lent to those in similarly labeled non-trypsintreated cells. No polypeptide E2 could be detect-ed in this fraction. Surprisingly, the plasmamembranes isolated from trypsin-treated cellscontained very little capsid protein relative tonontreated cells (Fig. 3B and reference 9, Fig.4). This result suggests that, although capsidprotein is produced in trypsin-treated cells, thiscomponent failed to bind to modified plasmamembrane, as was the case in nontreated cells.Electron microscopy of Sindbis virus-infected
trypsin-treated cells. The observation that capsidprotein synthesized in trypsin-treated cells didnot become plasma membrane associated sug-gested that maturation of Sindbis virus fromtrypsin-treated cells might be prevented by afailure to assemble capsid protein into nucleo-capsid structures. Sindbis virus-infected cellsprepared for electron microscopy 8 h postinfec-tion and 5 h after treatment or mock treatmentwith trypsin are shown in Fig. 5. Infected cellsnot treated with trypsin contained large numbersof nucleocapsid structures in their cell cyto-plasm (Fig. 5A). Cells treated with trypsin con-tained very few distinguishable nucleocapsidstructures (Fig. 5B). Although some virus parti-cles could be seen associated with the surface oftrypsin-treated cells (Fig. 5B [arrows]), none ofthese could be identified as budding virus whenexamined at high magnification. These particlesare most likely virus stuck to the cell surfacefrom the inoculum or from particles producedbefore trypsin treatment. This observation sup-ported the contention that nucleocapsid assem-bly is inhibited in trypsin-treated cells. Becausesynthesis of capsid protein occurs at approxi-mately normal levels in trypsin-treated cells, thelack of nucleocapsid assembly could result froma failure to produce virus progeny RNA, theother component required for virus nucleocap-sid assembly.RNA synthesis in Sindbis virus-infected tryp-
sin-treated cells. Total RNA synthesis in Sindbisvirus-infected trypsin-treated and mock-treatedcells was determined by assaying the amount of[3H]uridine incorporated into cellular trichloro-acetic acid-insoluble material over a 3-h periodafter treatment (Table 2). It was found that,although virus proteins accumulated at normal
levels, RNA synthesis was reduced to 13% ofnormal levels. This observation could not beattributed to a lack of [3H]uridine uptake bytrypsin-treated, Sindbis virus-infected BHKcells. Control experiments (Table 2) showed nodifferences in the total amount of [3H]uridineuptake by trypsin-treated cells as compared withmock-treated cells.
It has been demonstrated that, in Sindbisvirus-infected cells, two forms of plus-strandRNA are generated from a common 42S-strandtemplate (16). One of these species is the 26SRNA which codes for the virus structural pro-teins. The other is 42S RNA which is incorporat-ed into progeny virions and may also code fornonstructural proteins responsible for virus rep-lication (16). We determined the relativeamounts of the RNA species synthesized intrypsin-treated and nontreated cell cultures.Identical cultures of BHK-21 cells were treatedwith actinomycin D and infected as described inMaterials and Methods. At 3 h postinfection, themonolayers were treated or mock treated withtrypsin as described in Materials and Methods.After treatment, the cells were labeled with[3H]uridine (75 p,Ci/ml) for 3 h at 37°C. TotalRNA was extracted from the cells as describedin Materials and Methods, and the RNA wassedimented in 15 to 30% sucrose gradients. RNAspecies were found to sediment in peaks corre-sponding to those of 42S and 26S RNA species.The total amount of label in each of these peakswas determined. The medium of the labeled cellcultures was subjected to isopycnic centrifuga-tion on linear potassium tartrate gradients, andthe amount of label banding at virus density (42SRNA) was mathematically added to the cellular42S RNA component to give a value for total42S RNA synthesized by the trypsin-treated andnontreated cells. This was done to account forthe label lost from cells due to virus maturation(Table 3). The yields of both 26S and 42S RNAswere reduced by approximately equal amountsin trypsin-treated cells, indicating that trypsintreatment reduced total virus RNA synthesisand not the synthesis of one particular species ofRNA.
DISCUSSIONThe data presented above demonstrate that
Sindbis virus-infected BHK-21 or Vero cellslose their ability to efficiently produce progenyvirions when treated briefly with trypsin at 3 hpostinfection. Trypsin treatment was found tohave no effect on the production of VSV fromBHK-21 cells. Trypsin is also used to enhanceinfectious paramyxovirus production in certaincell lines (27). This latter experiment, as well ascontrol experiments described above, indicates
J. VIROL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 21
Oct
ober
202
1 by
188
.168
.178
.115
.

SINDBIS VIRUS MATURATION
'C. a'C-~~~~~~~~~~~~~~~~~~ira
41~
.4..~~~~~~~~~~~~~~~~0
t~ ~ ~ ;k..r-k'
A. "
4 4i ~~,") 4
ir.'t
I .$
rirw ''tt-l - rl
,'iI...4b, \
¶ -zar '',t;Q< ,,'a 2 -
I
--
ki - t -
FIG. 5. Electron micrographs of trypsin- and mock-treated Sindbis virus-infected BHK-21 cells. Identicalmonolayers of Sindbis virus-infected BHK-21 cells were treated with trypsin or mock treated as described in thetext at 3 h postinfection. At 8 h postinfection, the cells were collected and prepared for electron microscopy asdescribed in the text. Cells not treated with trypsin (A) were found to contain many nucleocapsids (someindicated by arrows). Virus was also found budding from the plasma membrane (insert). Trypsin-treated cells (B)contained few identifiable capsid structures, although some mature virus could be seen at the cell surface(arrows). Magnification: A, x15,000 (insert, x154,000); B, x11,000.
VOL. 41, 1982 699
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 21
Oct
ober
202
1 by
188
.168
.178
.115
.

700 ADAMS AND BROWN
TABLE 3. Incorporation of [3H]uridine into 26S and42S RNA in Sindbis virus-infected, trypsin-treated,
and nontreated cellsa
Total cpm (x 1o-5) incor-Type of cells porated into:
42S RNA 26S RNA
Trypsin-treated 1.6 8.5Nontreated 10.3 49.7
Ratio (treated/nontreated) 0.15 0.17
a Samples were prepared as described in text.
the nontoxic nature of trypsin treatment andfurther shows that the effects of trypsin were notattained by inactivation of virus after matura-tion.The synthesis of Sindbis virus-specified poly-
peptides continued at normal levels in trypsin-inhibited cells. Polypeptides PE2, E1, and Ccould be readily identified by polyacrylamide gelelectrophoresis, and virus glycoproteins PE2and E1 were transported to cell plasma mem-branes in the inhibited cells. Trypsin-inhibitedcells differed from control cells in that PE2 wasnot converted to E2 during inhibition, and C wasnot associated with plasma membrane in thequantities seen in control cells. Electron micros-copy of inhibited and control cells revealed veryfew detectable nucleocapsid structures in inhib-ited cells relative to control cells. The reducedlevel of assembly of nucleocapsids in the pres-ence of normal levels of capsid protein is proba-bly due to the significantly reduced levels of 26Sand 42S progeny RNA synthesis occurring in theinhibited cell cultures. Thus, the failure to pro-duce mature virions in inhibited cells seems tohave resulted from a greatly reduced assemblyof nucleocapsids, which was in turn due to a90% reduction in progeny RNA synthesis. Theobservation that 26S RNA synthesis can begreatly reduced by trypsin treatment early ininfection without affecting the net synthesis ofvirus structural proteins implies that, at 3 hpostinfection, sufficient stable messenger hasaccumulated to sustain normal levels of virusproduction.These observations expand our knowledge of
alphavirus assembly in that they show that Cdoes not attach to modified plasma membranesunless incorporated with progeny RNA into anucleocapsid structure. They further suggestthat conversion of PE2 to E2 may be dependentupon the attachment of nucleocapsid to thecytoplasmic carboxy terminus of the PE2 poly-peptide, although we have previously demon-strated that this attachment in and of itself is notsufficient to induce the conversion of PE2 to E2(4, 25, 36). The primary effects of trypsin inhibi-
tion on Sindbis maturation seem to be at thelevel of RNA synthesis. How this inhibitionoccurs is not clear. The experiments presentedherein show that the infection of cells by Sindbisvirus must be well established (3 h postinfection)for trypsin treatment to have its maximum in-hibitory effect. Surprisingly, the medium of tryp-sin-treated Sindbis virus-infected cells containssomething which blocks Sindbis virus produc-tion when this medium is applied to other Sind-bis virus-infected cultures. A similar mediumderived from trypsin-treated uninfected cells didnot have this effect. These results suggest thattrypsin treatment of Sindbis virus-infected cellsresults in the release into the media of someagent capable of inhibiting progeny RNA syn-thesis. Transmembranal effects of an externallyacting agent, anti-measles virus serum, on cyto-plasmic processing of virus structural proteinshas been demonstrated by Fuginami and Old-stone (11). It is not clear as to whether thesetrypsin-produced components (or trypsin itself)produce their inhibitory effect transmembranally(that is, by acting on the outer surface of the cellto alter some internal fraction) or if the compo-nent finds its way to some internal site ofactivity. Characterization of the inhibitory agentreleased from Sindbis virus-infected trypsin-treated cells, however, is now in progress.
It is interesting that the maximum inhibitoryeffect of trypsin on progeny virus production isattained when trypsin treatment takes place atapproximately the time at which Bruton andKennedy (6) found that negative-strand RNAsynthesis is being terminated. It is conceivablethat trypsin treatment released from the cell anagent(s) which inactivates those negative-strand-containing replicating complexes respon-sible for the production of plus-strand 26S and42S RNA. This data would suggest that, onceinactivated, these complexes cannot be returnedto an active plus-strand-producing state. Whentrypsin is applied at times before 3 h postinfec-tion, virus-infected cells are still producing nega-tive-strand RNA which can be used to replacesome of the inactivated plus-strand synthesizingcomplexes. Once negative-strand RNA synthe-sis is turned off (ca. 3 h postinfection), plus-strand RNA synthesis stops for lack of function-al negative-strand-containing templates. Thecomplete insensitivity of VSV-infected cells tosimilar treatment with trypsin may derive frombasic differences in the process of replication ofthis negative-strand virus. This and other possi-ble explanations for the results presented abovewill be pursued in future studies.The above observations suggest that trypsin
treatment of Sindbis virus-infected cells maypresent a unique opportunity to study eventsrelated to virus protein processing in wild-type-
J. VIROL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 21
Oct
ober
202
1 by
188
.168
.178
.115
.

VOL. 41, 1982
infected cells in the absence of virus maturationand E2 production. These cells also provide asource of plasma membrane modified by wild-type virus glycoproteins without associated nu-cleocapsids, and such components may prove tobe important sources of material for studies ofnucleocapsid-membrane interactions. The syn-thesis of normal levels of capsid protein in theabsence of 42S RNA synthesis may also providea source of non-RNA-associated capsid protein.
ACKNOWLEDGMENTS
This research was supported by Public Health Service grantAI-14710 and Public Health Service training grant TGCA-09182, from the National Institutes of Health and by grant F-717 from the Robert A. Welch Foundation. Raquel Adams is aFellow of the Compton-Danforth Foundation and was sup-ported for part of this research by the National Institutes ofHealth.We thank Bob Slocum, who assisted with the electron
microscopy and was supported by Public Health Servicetraining grant TGCA-09182.
LITERATURE CITED
1. Bonner, W. M., and R. A. Laskey. 1974. A film detectionmethod for tritium-labeled proteins and nucleic acids inpolyacrylamide gels. Eur. J. Biochem. 46:83-88.
2. Brache, M., and M. J. Schlesinger. 1976. Defects in RNAtemperature sensitive mutants of Sindbis virus and evi-dence for a complex of PE2-E, viral glycoproteins. Vi-rology 74:441-449.
3. Brown, D. T. 1980. The assembly of alphaviruses, p. 473-501. In R. W. Schlesinger (ed.), The togaviruses. Academ-ic Press, Inc., New York.
4. Brown, D. T., and J. F. Smith. 1975. Morphology of BHK-21 cells infected with Sindbis virus temperature-sensitivemutants in complementation groups D and E. J. Virol.15:1262-1266.
5. Brown, D. T., M. R. F. Waite, and E. R. Pfefferkorn. 1972.Morphology and morphogenesis of Sindbis virus as seenwith freeze-etching techniques. J. Virol. 10:524-536.
6. Bruton, C. J., and S. I. T. Kennedy. 1975. Semliki Forestvirus intracellular RNA: properties of the multistrandedRNA species and kinetics of the positive and negativestrand synthesis. J. Gen. Virol. 28:111-127.
7. Burke, D., and K. Keegstra. 1979. Carbohydrate structureof Sindbis virus glycoprotein E2 from virus grown inhamster and chicken cells. J. Virol. 29:546-554.
8. Eagle, H. 1959. Amino acid metabolism in mammalian cellcultures. Science 130:432-437.
9. Erwin, C., and D. T. Brown. 1980. Intracellular distribu-tion of Sindbis virus membrane proteins in BHK-21 cellsinfected with wild-type virus and maturation-defectivemutants. J. Virol. 36:775-786.
10. Fratalli, V. 1969. Soybean inhibitors. III. Properties of alow molecular weight soybean protease inhibitor. J. Biol.Chem. 244:274.
11. Fuginami, R. S., and M. B. A. Oldstone. 1979. Antiviralantibody reaction on the plasma membrane alters virusexpression inside the cell. Nature (London) 279:529-530.
12. Hirschberg, C. B., and P. W. Robbins. 1972. The glycolip-ids and phospholipids of Sindbis virus and their relation tothe lipids of the host cell plasma membrane. Virology61:602-608.
13. Igarashi, A., R. Koo, and V. Stollar. 1977. Evolution andproperties of Aedes albopictus cell cultures persistentlyinfected with Sindbis virus. Virology 82:69-83.
14. Johnson, D. C., and M. J. Schlesinger. 1980. Vesicularstomatitis and Sindbis virus glycoprotein transport to thecell surface is inhibited by ionophores. Virology 103:407-424.
15. Johnson, D. C., M. J. Schlesinger, and E. L. Elson. 1981.
SINDBIS VIRUS MATURATION 701
Fluorescence photobleaching recovery measurements re-veal differences in envelopment of Sindbis and vesicularstomatitis viruses. Cell 23:423-431.
16. Kennedy, S. I. T. 1980. Synthesis of alphavirus RNA, p.351-368. In R. W. Schlesinger (ed.), The togaviruses.Academic Press, Inc., New York.
17. Kos, K. A., B. A. Osborn, and R. A. Goldsby. 1975.Inhibition of group B arbovirus antigen production andreplication in cells enucleated wtih cytochalasin B. J.Virol. 15:913-917.
18. Laemmli, V. K. 1970. Cleavage of structural proteinsduring the assembly of the head of bacteriophage T4.Nature (London) 227:680685.
19. Ortin, J., and E. Vinuela. 1977. Requirement of cellnucleus for African swine fever virus replication in Verocells. J. Virol. 21:902-905.
20. Penman, S. 1966. RNA metabolism in the Hela cellnucleus. J. Mol. Biol. 17:117-130.
21. Pesonin, M., and 0. Renkonin. 1976. Sequence and ano-meric configuration of monosaccharides in type A glyco-peptides of Semliki Forest Virus. Biochim. Biophys. Acta455:510-525.
22. Pfefferkorn, E. R., and R. L. Clifford. 1964. The origin ofthe proteins of Sindbis virus. Virology 23:217-223.
23. Renz, D., and D. T. Brown. 1976. Characteristics ofSindbis virus temperature-sensitive mutants in culturedBHK-21 and Aedes albopictus (mosquito) cells. J. Virol.19:775-781.
24. Riedel, B., and D. T. Brown. 1977. The role of extracellu-lar virus in the maintenance of the persistent infectioninduced in Aedes albopictus (mosquito) cells by Sindbisvirus. J. Virol. 23:554-561.
25. Scheefers, H., U. Scheefers-Borchel, J. Edwards, and D. T.Brown. Distribution of virus structural proteins and pro-tein-protein interaction in plasma membrane of babyhamster kidney cells infected with Sindbis or vesicularstomatitis virus. Proc. Natl. Acad. Sci. U.S.A. 77:7277-7281.
26. Scheefers-Borchel, U., H. Scheefers, J. Edwards, and D. T.Brown. 1981. Sindbis virus maturation in cultured mosqui-to cells is sensitive to actinomycin D. Virology 110:292-301.
27. Scheid, A., and P. W. Choppin. 1974. Identification of thebiological activities of paramyxovirus proteins. Activationof cell from hemolysis and infectivity by proteolyticcleavage of an inactive precursor protein of Sindbis virus.Virology 57:475-490.
28. Schlesinger, M. J., and L. Kaariainen. 1980. Translationand processing of alphavirus proteins, p. 371-389. In R.W. Schlesinger (ed.), The togaviruses. Academic Press,Inc., New York.
29. Schlesinger, M. J., and S. Schlesinger. 1973. Large-molec-ular-weight precursors of Sindbis virus proteins. J. Virol.11:1013-1016.
30. Schlesinger, M. J., S. Schlesinger, and B. W. Burge. 1972.Identification of a second glycoprotein in Sindbis virus.Virology 47:539-541.
31. Schlesinger, S., and M. J. Schlesinger. 1972. Formation ofSindbis virus proteins: identification of a precursor forone of the envelope proteins. J. Virol. 10:925-932.
32. Schmidt, M. F. G., and M. J. Schlesinger. 1979. Fatty acidbinding to vesicular stomatitis virus glycoproteins: a newtype of post-translational modification of the viral glyco-protein. Cell 17:813-819.
33. Schmidt, M. F. G., and M. J. Schlesinger. 1980. Relationof fatty acid attachment to the translation and maturationof vesicular stomatitis and Sindbis virus membrane glyco-proteins. J. Biol. Chem. 255:3334-3339.
34. Sefton, B. M., and B. W. Burge. 1973. Biosynthesis of theSindbis virus carbohydrates. J. Virol. 12:1366-1374.
35. Simons, K., Garoff, H., and A. Helenius. 1980. Alphavirusproteins, p. 317-331. In R. W. Schlesinger (ed.), Thetogaviruses. Academic Press, Inc., New York.
36. Smith, J. F., and D. T. Brown. 1977. Envelopment ofSindbis virus: synthesis and organization of proteins in
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 21
Oct
ober
202
1 by
188
.168
.178
.115
.

702 ADAMS AND BROWN
cells infected with wild-type and maturation-defectivemutants. J. Virol. 22:662-678.
37. Spurr, A. R. 1969. A low viscosity epoxy resin embeddingmedium for electron microscopy. J. Ultrastruct. Res.26:31-43.
38. von Bonsdorff, C.-H., and S. C. Harrison. 1978. Hexago-nal glycoprotein arrays from Sindbis virus membranes. J.Virol. 28:578-583.
J. VIROL.
39. Wengler, G. 1980. Effects of alphavirus on host cellmacromolecular synthesis, p. 459-471. In R. W. Schle-singer (ed.), The togaviruses. Academic Press, Inc., NewYork.
40. Wirth, D. F., H. F. Lodish, and P. W. Robbins. 1979.Requirements for the insertion of the Sindbis envelopeglycoproteins into the endoplasmic reticulum membrane.J. Cell Biol. 81:154-162.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 21
Oct
ober
202
1 by
188
.168
.178
.115
.





























