Embed Size (px)
Citation preview

Cancer Therapy: Preclinical
Inhibition of the EGFR/STAT3/CEBPD AxisReverses Cisplatin Cross-resistance withPaclitaxel in the Urothelial Carcinoma of theUrinary BladderWei-Jan Wang1, Chien-Feng Li2,3,4,5, Yu-Yi Chu6, Yu-Hui Wang6, Tzyh-Chyuan Hour7,Chia-Jui Yen8,Wen-Chang Chang9,10, and Ju-Ming Wang6,9,10,11
Abstract
Purpose: Cisplatin (CDDP) is frequently used in combinationchemotherapy with paclitaxel for treating urothelial carcinoma ofthe urinary bladder (UCUB). CDDP cross-resistance has beensuggested to develop with paclitaxel, thus hindering successfulUCUB treatment. Therefore, elucidating the mechanisms under-lying CDDP-induced anticancer drug resistance is imperative andmay provide an insight in developing novel therapeutic strategy.
Experimental Design: Loss-of-function assays were performedto elucidate the role of the EGFR and STAT3 in CDDP-inducedCCAAT/enhancer-binding protein delta (CEBPD) expression inUCUB cells. Reporter and in vivo DNA-binding assays wereemployed to determine whether CEBPD directly regulates ATPbinding cassette subfamily Bmember 1 (ABCB1) andATPbindingcassette subfamily C member 2 (ABCC2) activation. Finally, axenograft animal assay was used to examine the abilities of
gefitinib and S3I-201 (a STAT3 inhibitor) to reverse CDDP andpaclitaxel sensitivity.
Results: CEBPD expression was maintained in postoperativechemotherapy patients, and this expression was induced byCDDP even in CDDP-resistant UCUB cells. Upon CDDP treat-ment, CEBPD activated ABCB1 and ABCC2. Furthermore, theEGFR/STAT3 pathway contributed to CDDP-induced CEBPDexpression in UCUB cells. Gefitinib and S3I-201 treatment sig-nificantly reduced the expression of CEBPD and enhanced thesensitivity of CDDP-resistant UCUB cells to CDDP and paclitaxel.
Conclusions:Our results revealed the risk of CEBPD activationin CDDP-resistant UCUB cells and suggested a therapeutic strat-egy for patients with UCUB or UCUB resisted to CDDPand paclitaxel by combination with either gefitinib or S3I-201.Clin Cancer Res; 23(2); 503–13. �2016 AACR.
IntroductionUrothelial carcinoma of the urinary bladder (UCUB) is the
fourth and tenth most commonmalignancy in men and women,
respectively. At present, cisplatin (CDDP) is the most commonagent for treating locally advanced and metastatic UCUB. How-ever, the intrinsic and acquired CDDP resistance of UCUB is amajor clinical problem, which eventually results in treatmentfailure (1). In addition, paclitaxel, a microtubule dissociationinhibitor, is awidely accepted combination agent for use inCDDPin UCUB therapy. However, CDDP-resistant tumors (includingUCUB) are not completely responsive to salvage combinationchemotherapy involving paclitaxel (2). Therefore, novel thera-peutic combinations or treatments are required to reverse CDDP-induced paclitaxel cross-resistance.
Approximately 50% of bladder tumors exhibit strong immu-nohistochemical staining for EGFR, which is also associated withmuscle invasion and poor tumor differentiation (3, 4). In addi-tion, a subgroupofmuscle-invasive bladder carcinomadisplayinga basal-like phenotype is sensitive to EGFR inhibitor erlotinib (5).However, the underlying mechanisms remained unknown. Gefi-tinib, also known as ZD1839, selective EGFR tyrosine kinaseantagonist that inhibits tumor cell growth (6), attenuates UCUBcell proliferation and enhances apoptosis (7). However, thedetailed mechanisms underlying the effects of EGFR in UCUBcells drug resistance remain largely unknown.
Under normal physiologic conditions, the expression levels oftranscription factor CEBPD are relatively low, but the levels can beupregulated by various extracellular stimuli (8). CEBPD is alsoresponsive to several anticancer drugs, including vitamin D3 (9)and 1-(2-hydroxy-5-methylphenyl)-3-phenyl-1,3-propanedione(HMDB; ref. 10). Several studies have suggested that the
1Institute of Basic Medical Science, National Cheng Kung University, Tainan,Taiwan R.O.C. 2Department of Pathology, Chi-Mei Medical Center, Tainan,Taiwan R.O.C. 3Department of Biotechnology, Southern Taiwan University ofScience and Technology, Tainan, Taiwan R.O.C. 4National Institute of CancerResearch, National Health Research Institutes, Tainan, Taiwan R.O.C. 5GraduateInstitute of Medicine, College of Medicine, Kaohsiung Medical University, Kaoh-siung, Taiwan R.O.C. 6Institute of Bioinformatics and Biosignal Transduction,National Cheng Kung University, Tainan, Taiwan R.O.C. 7Department of Bio-chemistry, Kaohsiung Medical University, Kaohsiung, Taiwan R.O.C. 8Division ofHematology/Oncology, Department of Internal Medicine, National Cheng KungUniversity Hospital, National Cheng Kung University, Tainan, Taiwan R.O.C.9Center of Infectious Disease and Signaling Research, National Cheng KungUniversity, Tainan, Taiwan R.O.C. 10Graduate Institute of Medical Sciences,College of Medicine, Taipei Medical University, Taipei, Taiwan R.O.C. 11Center ofMolecular Inflammation, National Cheng Kung University, Tainan, Taiwan R.O.C.
Note: Supplementary data for this article are available at Clinical CancerResearch Online (http://clincancerres.aacrjournals.org/).
C.F. Li and J.M. Wang contributed equally and share last authorship.
CorrespondingAuthor: Ju-MingWang, Institute of Bioinformatics andBiosignalTransduction, College of Bioscience and Biotechnology, National Cheng KungUniversity, 1 Ta-Hsueh Road, Tainan 701, Taiwan. Phone: 886-6275-7575, ext.31067; Fax: 886-6208-3663; E-mail: [email protected]
doi: 10.1158/1078-0432.CCR-15-1169
�2016 American Association for Cancer Research.
ClinicalCancerResearch
www.aacrjournals.org 503
on April 8, 2021. © 2017 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from
Published OnlineFirst July 19, 2016; DOI: 10.1158/1078-0432.CCR-15-1169

p38/CREB (cAMP responsive element binding protein) and JAK/STAT3 pathways play crucial roles in CEBPD transcriptionalactivation (10, 11). Previous studies have also suggested thatCEBPD plays a vital role in inflammation and inflammatorydisease processes (12, 13). However, unlike the inflammatoryeffectorsNF-kBand STAT3 that are consistently activated in cancercells, CEBPD inactivation has been observed in several types ofcancer, including cervical and hepatocellular carcinoma (10, 14),breast (15), prostate cancer (9), and leukemia (16). In addition toacting as a tumor suppressor, several recent reports have suggestedthat CEBPD plays an oncogenic role under certain conditions(17, 18). Furthermore, CEBPD attenuation can sensitize CDDP-induced cell death in CDDP-resistant UCUB cells (19). However,the details remain largely unclear, and the improvement inCEBPDactivation followingCDDP andpaclitaxel cross-resistanceremains uninvestigated.
Multidrug resistance (MDR) transporter proteins are mostknown for their contribution to chemoresistance through thecellular efflux of anticancer drugs. Several members of the ABCtransporter family can induce MDR. To date, tumor tissue studieshave consistently revealed that in most cancer cells, the majormechanismunderlyingMDR involves ABCB1, ABCC1, or ABCG2.ABCC2 endows tumor cells with resistance to various anticancerdrugs, including CDDP (20). Furthermore, the ABC transporterfamily proteins have well-known specific chemotherapy sub-strates: ABCC2 effluxes CDDP andmitoxantrone, whereas ABCC1effluxes mitoxantrone and paclitaxel (21). Currently, most evi-dence for additional roles is correlative, and definitive studies arerequired to confirm the causality.
In this study, we showed that CDDP-resistant UCUB cells alsoexhibited considerable paclitaxel resistance, CEBPD responded toCDDP treatment, and CEBPD activation contributed to ABCB1and ABCC2 expression in UCUB cells. Furthermore, CDDP-induced EGFR phosphorylation and gefitinib markedly inhibitedCEBPD levels in UCUB cells. We observed that STAT3 activationwas EGF responsive and sustained in CDDP-resistant NTUB1(NTUB1/P) cells. Moreover, CEBPD expression correlated withpEGFR and pSTAT3 levels in specimens from patients with UCUBreceiving postoperative chemotherapy. The ABC transportersABCB1 and ABCC2 were upregulated by CDDP-induced CEBPD
through transcriptional regulation and contributed to CDDPresistance and CDDP-induced paclitaxel cross-resistance. Finally,CDDP administered in combination with either gefitinib orS3I-201 significantly inhibited viability of CDDP-resistant UCUBcells and tumor growth. Overall, our results suggest that gefitiniband S3I-201 are useful adjuvants for treating CDDP-resistantUCUB, particularly for patients who are administered these drugsin combination with CDDP or paclitaxel.
Materials and MethodsPatients and tumor specimens
This study was approved by the Institutional Review Board(approval number IRB10302015) of the Chi Mei Medical Center(Tainan City, Taiwan). All samples were obtained from the ChiMei Medical Center BioBank and were previously collectedaccording to the official ethical guidelines. We retrieved UCUBcases (1996–2004) from the aforementioned medical centerarchives for immunohistochemical and survival analysis, asdescribed previously (22). To evaluate the significance of CEBPDexpression with respect to a CDDP-based chemotherapyresponse, we enrolled 79 UCUB cases with pT2 to pT4 tumorsor with nodal involvement in which CDDP-based postoperativeadjuvant chemotherapy was applied. To closely validate thesignificant findings, 60 independent cases were collected fromLiou Ying Campus of Chi Mei Healthcare system. The clinico-pathologic evaluation criteria were essentially identical to thoseused in our previous study (23). An expert pathologist (C.-F. Li)reevaluated hematoxylin and eosin–stained sections from eachcase.
Cell lines and culture conditionsJ82, TSGH8301, and TCCSUP cells were maintained in
DMEM supplemented with 10% FBS, 100 U/mL penicillin, and100 mg/mL streptomycin. The J82 and TCCSUP cells werepurchased from ATCC. Human NTUB1 (24) and the TSGH8301(25) cell lines were obtained from and authenticated byT.-C. Hour (Kaohsiung Medical University, Kaohsiung, Taiwan).The NTUB1 and nasopharyngeal cancer HONE1 cells weremaintained in RPMI1640 media supplemented with 10% FBS,100 U/mL penicillin, and 100 mg/mL streptomycin. StableCDDP-resistant NTUB1/P and HONE1 (HONE1/R) clones weremaintained in the aforementioned media with 10 mmol/LCDDP (Sigma).
Cell viability and cell death assaysCell survival was measured using the Cell Counting Kit-8
(Sigma). Experimental cells were treated with either CDDP(20 mmol/L; BioVision) or paclitaxel (1 mmol/L; BioVision) for24 hours. For the combination treatment, the cells were treatedwith CDDP or paclitaxel plus either gefitinib (5 mmol/L; BioVi-sion) or S3I-201 (10 mmol/L; BioVision) for 24 hours. Thepercentages of cell viability and death were determined for eachtreatment through comparisons with the untreated controlgroup. For cell death assays, the cells were plated and culturedin regular media as previously mentioned for 16 hours andthen treated with CDDP alone or in combination with eithergefitinib or S3I-201 for another 24 hours. The cells were stainedwith propidium iodide (Sigma) and analyzed through flowcytometry.
Translational Relevance
Despite extensive investigation, urothelial carcinoma of theurinary bladder (UCUB) remains a relatively understudieddisease, and nonsurgical treatments have undergone littledevelopment in recent decades. We demonstrated thatEGFR-induced CEBPD promotes cisplatin (CDDP) and pac-litaxel cross-resistance in CDDP-resistant UCUB by elevatingATP binding cassette subfamily Bmember 1 (ABCB1) andATPbinding cassette subfamily C member 2 (ABCC2) levels,respectively. In addition, our results validated suppression ofCEBPDby gefitinib or S3I-201 to enhance the apoptotic effectsin UCUB and CDDP-resistant UCUB cells in vitro and in vivo.The study also describes the first preclinical evaluation ofS3I-201 in UCUB and supports that CEBPD inhibition canreduce drug resistance and promote therapeutic efficiency inCDDP-treated UCUB.
Wang et al.
Clin Cancer Res; 23(2) January 15, 2017 Clinical Cancer Research504
on April 8, 2021. © 2017 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from
Published OnlineFirst July 19, 2016; DOI: 10.1158/1078-0432.CCR-15-1169

Chromatin immunoprecipitation–PCR assayThe chromatin immunoprecipitation (ChIP) assay was per-
formed as described previously (13). In brief, the experimentalcells were treated with 1% formaldehyde for 15 minutes, and thecross-linked chromatin was prepared and sonicated to an averagesize of 500 bp. The DNA fragments were immunoprecipitatedusing either specific antibodies recognizing CEBPD or controlrabbit immunoglobulin G (IgG) at 4 �C for 12–16 hours. Fol-lowing cross-link reversal, quantitative real-time PCR assays wereperformed using the precipitated DNA and primers correspond-ing to specific target gene regions. The primers were as follows:
CEBPD-A (A), 50-GCCAAGTCCTGGTTTTGATT-30; CEBPD (AS),50 -GCCCCCTCTCAGTTCCTC-30; CEBPD-B (A), 50-GGTTTCAC-CATGTTGACCAG-30; CEBPD-B (AS), 50-AGAATGGGCTTTGT-CATGTG-30; ABCB1-A (A), 50-CTCTGGTACTGGGATAAACACT-TGTA-30; ABCB1-A (AS), 50-TGATTGCAAACTTCTAGTCAAGACA-30; ABCB1-B (A), 50-CCTGGACCCAGCTTTCACTA-30; and ABCB1-B (AS), 50-AGCACAAATTGAAGGAAGGAG-30. The amplifiedproducts were resolved using agarose gel electrophoresis andconfirmed using sequencing.
Lentivirus knockdown assaysViruses were produced in Phoenix cells by cotransfecting various
short hairpin RNA (shRNA) expression vectors with pMD2.G andpsPAX2. The lentiviral knockdownexpression vectorswereobtainedfrom the National RNAi Core Facility (Genomic Research Center ofthe Institute of Molecular Biology, Academia Sinica, Taiwan). Afterthe viral infection efficiencywas determined, lentiviruses containingshb-galactosidase (shLacZ) or shCEBPD were used for infectingNTUB1/P cells for 72 hours at a multiplicity of infection. For alllentiviral experiments, media containing uninfected viruses wereremoved before further analysis. The shRNA sequences in thelentiviral expression vectors were as follows: shLacZ, 50-CCGGTGTTCGCATTATCCGAACCATCTCGAGATGGTTCGGATAA-TGCGAACATTTTTG-30; shCEBPD, 50-CCGGGCCGACCTCTTC-AACAGCAATC TCGAGATTGCTGTTGAAGAGGTCGGCTTTTT-30;shABCB1, 50-CCGGGCTGCTTTCCTGCTGATCTATCTCGAGATA-GATCAGCAGGAAAGCAGCTTTTTG-30 and shABCC2, 50-CCGG-CCTGGTGGATAGCAACAATATCTCGAGATATTGTTGCTATCCAC-CAGGTTTTTG-30.
MDR pump activity assaysThe experimental cells were plated overnight in a growth
medium at a concentration of 60,000 cells/well/90 mL in 96-wellplates. The next day, the cells were treated with CDDP andgefitinib or S3I-201 for 24 hours. Furthermore, the cells weremixed with 100 mL/well of MDR dye-loading solution (Abcam)and incubated for 4 hours at room temperature. Sample fluores-cence intensity was then measured at 525 nm by using a fluores-cence reader (Thermo Scientific).
Immunohistochemical stainingTissue sections (4 mm)were cut from paraffin-embedded tissue
blocks, placed onto precoated slides, deparaffinized, and rehy-drated. Their antigens were retrieved, and endogenous peroxidasewas blocked, as described previously (23). Moreover, the slideswere incubated for 1 hours with primary antibodies for CEBPD(1:200; ab65081; Abcam), pEGFR (1:25; Tyrosine 1086–specific,Zymed), pSTAT3 (1:25; Tyrosine 705-specific, Santa Cruz Bio-technology), and ABCB1 (1:100, C-19, Santa Cruz Biotechnolo-gy). We then detected the primary antibodies using the DAKO
ChemMate EnVision kit (K5001; DAKO). To ensure immunos-taining quality, a sample prepared without the primary antibodyserved as the negative control.
IHC interpretation and scoringAn expert pathologist (C.-F. Li), blinded to clinical and follow-
up data, evaluated the IHC results. Immunoreactivity was eval-uatedusing both the percentage and intensity of positively stainedtumors, illustrating representative staining patterns (pEGFR,membranous and cytoplasmic; pSTAT3, nuclear; CEBPD, nuclear;ABCB1, cytoplasmic) to calculate the H-score according to thefollowing equation: H-score ¼ SPi(i þ1), where i is the intensityof the stained tumor cells (0–3), andPi is thepercentage of stainedtumor cells for each intensity (0%–100%). This formula generatesa score between 100 and 400; 100 and 400 indicate that 100% ofthe tumor cells were negative (0) and strongly stained (3þ),respectively (23).
Animal studiesMale 6- to 8-week-old NOD/SCID mice were purchased from
the Laboratory Animal Center of the National Cheng KungUniversity (Tainan, Taiwan). NTUB1/P cells (1 � 107) in 100 mLof PBS were subcutaneously inoculated into the right flanks of themice. Once macroscopic tumors (100–125 mm3) formed, theanimals (n¼4per group)were randomly separated into 4 groups.The groups were then administered intraperitoneal treatments asfollows: the control group received the vehicle (PBS), the CDDPgroup received CDDP (5 mg/kg/2 days) for 3 weeks, the CDDPplus gefitinib group received CDDP (5 mg/kg/2 days) and gefi-tinib (10 mg/kg/2 days) for 3 weeks, and the CDDP plus S3I-201group received CDDP (5mg/kg/2 days) and S3I-201 (10mg/kg/2days) for 3 weeks. Animal weights and tumor dimensions weremeasured every 3 days by calipers, and tumor volumes wereestimated from the tumor length and width using the formula[L�W2]/2, where L andW are the length and width, respectively.
Statistical analysisStatistical analysis was performed using SPSS V.14.0 software
(SPSS Inc.). The median IHC H-scores for CEBPD, pEGFR,pSTAT3, and ABCB1 were used as cutoffs for separating thecases into high and low expression groups. Furthermore, a c2
test was used to assess the associations among CEBPD, pEGFR,pSTAT3, and ABCB1 expression. The endpoint analyzed wasmetastasis-free survival (MeFS), which was calculated from thedates of curative surgery and development of distal spreading.Patients lost to follow-up were censored on the final follow-update. We plotted survival curves by using the Kaplan–Meiermethod and evaluated predictive differences between thegroups by using the log-rank test. For all analyses, two-sidedtests of significance were conducted, with P < 0.05 consideredsignificant. The statistical significance of the variations betweenthe mean values was estimated using the SigmaPlot softwarepackage and independent Student t test for unequal variances.The data were expressed as the mean � SEM, and P < 0.05 wasconsidered statistically significant.
ResultsSignificance of CEBPD immunoreactivity in UCUB
UCUB cells exhibited nuclear CEBPD. After dichotomizing thetumors into low and high CEBPD expression groups, we found
Targeting CEBPD Prevents Drug Resistance in UCUB
www.aacrjournals.org Clin Cancer Res; 23(2) January 15, 2017 505
on April 8, 2021. © 2017 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from
Published OnlineFirst July 19, 2016; DOI: 10.1158/1078-0432.CCR-15-1169

that increasedCEBPDexpressionwas significantly associatedwithnodal involvement (P ¼ 0.013; Supplementary Table S1). Nota-bly, UCUB patients with high CEBPD levels had significantlyworse MeFS (P ¼ 0.0077; Supplementary Fig. S1A), suggesting apotential role of CEBPD in UCUB progression following postop-erative adjuvant chemotherapy. Of note, the aforementionedfindings remained significant in our independent cohort(Supplementary Fig. S1B)
CEBPD expression is increased in CDDP-resistant UCUB celllines and contributes to CDDP resistance and CDDP-inducedpaclitaxel cross-resistance
To investigate the involvement of CEBPD in the development ofCDDP resistance, we evaluated the association of CEBPD expres-sion and CDDP sensitivity in UCUB cell lines, including J82,TSGH8301, TCCSUP, and NTUB1. CEBPD abundance was corre-lated with UCUB cell resistance to CDDP-induced cell death
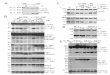
A
B C
ED
CTL
1.5
1
0.5
Cel
l via
bili
ty(v
s. c
on
tro
l)
0J82
J82
TSG
TSGH
TCC
TCC
CEBPD
1.5NTU
shLacZ8
1.5
1.0
0.5
0
1.5
CEBPD
a-Tubulin
1.0
0.5
0
6
4
2
0
shCD2
NTP
PTX
Cel
l via
bili
ty (
vs. c
on
tro
l)
Cel
l via
bili
ty (
vs. c
on
tro
l)
Rel
ativ
e fl
uo
resc
ence
un
its
(x10
3 , 4
85/5
20 n
m)
1
0.5
0
a-Tubulin
NTU
NTU
CDDP - - - - -+ + + + +
- + - +
CDDPPTXCDDP
shC
D2
shC
D1
shLa
cZ
shLac
Zsh
CD1sh
CD2
CTL
- + ++-+ + +-
shC
D2
shC
D1
shLa
cZ
+ + +-
NTP
NTP
CDDP 10 mmol/LCDDP 20 mmol/LCDDP 30 mmol/L
J82 TSG TCC NTP
CEBPD
a-Tubulin
NTU
Figure 1.
CEBPD expression is responsive to CDDPand is associated with CDDP andpaclitaxel (PTX) cross-resistance. A,CDDP affects cell viability and CEBPDexpression in various UCUB cells. UCUBcell lines were treated with variousconcentrations of CDDP for 24 hours.CEBPD expression levels were evaluatedusing Western blotting with theindicated antibodies, and cell viabilitywas examined using the CCK8 assay.TSGH8301 (TSG), TCCSUP (TCC), NTUB1(NTU), and CDDP-resistant NTUB1/P(NTP) cells. B, CEBPD expression isresponsive to CDDP treatment andsustained in all UCUB cells. A Westernblot analysis was conducted with lysatesharvested from CDDP-treated all UCUBcell lines for 24 hours. Western blotanalysis of CDDP-treated UCUB celllysates. C, NTP cells are insensitive topaclitaxel treatment. NTU and NTP cellswere treated with or without paclitaxeland assessed for cell viability by usingthe CCK8 assay after 24 hours ofincubation.D, CEBPD attenuation in NTPcells sensitizes them to CDDP andpaclitaxel. NTP cells were pretreatedwith lentiviruses containing eithershLacZ or shCEBPD (shCD1 and shCD2).After 48 hours of incubation,experimental cells were treated withCDDP or paclitaxel for 24 hours and thenassessed for cell viability by using theCCK8 assay. E, CEBPD affects the ABCtransporter–mediated pump activity inNTP cells. NTP cells were pretreatedwith lentiviruses containing eithershLacZ or shCEBPD (shCD2). The cellswere then treated with or without CDDP.After 24 hours, an MDR dye-loadingsolution was incubated in each well for 4hours, and the resulting fluorescenceintensity was detected using an ELISAreader. In this figure, ��� and �� denote asignificant difference with P < 0.001 and<0.05, respectively. CTL, control.
Wang et al.
Clin Cancer Res; 23(2) January 15, 2017 Clinical Cancer Research506
on April 8, 2021. © 2017 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from
Published OnlineFirst July 19, 2016; DOI: 10.1158/1078-0432.CCR-15-1169

(Fig. 1A). Our previous studies suggested that CEBPD is responsiveto anticancer drugs (26, 27). We further assessed whether CDDPaffected CEBPD expression in UCUB cells. In addition to theincreased basal CEBPD levels being observed in CDDP-resistantNTUB1/P cells, further CEBPD inductionwas observed after CDDPtreatment in NTUB1/P cells as well as other UCUB cells (Fig. 1B).These results suggested that not only CEBPD expression is associ-ated with CDDP resistance in UCUB cells but also that CDDP-induced CEBPD expression leads to drug resistance of UCUB cells.
Cancer cells evade death and exhibit increased drug efflux thatcontributes to drug resistance. As previously mentioned, CDDPtreatment could lead to paclitaxel cross-resistance; however, theunderlying mechanisms in UCUB cells remain unclear. Weassessed whether CDDP-resistant NTUB1/P cells were alsopaclitaxel insensitive. NTUB1/P cells exhibited attenuatedpaclitaxel sensitivity compared with NTUB1 cells (Fig. 1C). Fur-thermore, we examined the involvement of CEBPD in CDDPresistance and CDDP-induced paclitaxel resistance in our system.The loss-of-function assays using shLacZ and shCEBPD wereperformed to verify the role of CEBPD in CDDP resistance andCDDP-induced paclitaxel resistance. The results revealed that theattenuation of CEBPD levels by shCEBPD led to increased CDDPsensitivity and CDDP-induced paclitaxel resistance in NTUB1/Pcells (Fig. 1D).We also assessedwhether CEBPD contributes to anincreased drug efflux. In NTUB1/P cells, the loss of CEBPDenhanced cellular fluorescence intensity in the MDR pump activ-ity assay (Fig. 1E), suggesting that CEBPD contributes to theincrease in drug efflux.
EGFR inhibitor significantly enhances CDDP sensitivity indrug-resistant UCUB cells by reducing the CEBPD activity
A recent study reported the involvement of EGFR in drug-resistant UCUB (28); however, the mechanisms of EGFR-medi-ated drug resistance in UCUB remain unclear. We observed thatCDDP inducedEGFRexpression andactivity inNTUB1/P cells butnot in NTUB1 cells (Fig. 2A). We then examined the EGFR loss offunction by treating UCUB cells with acquired resistance with anEGFR inhibitor. This assay revealed that gefitinib treatment alonesignificantly induced cell death inNTUB1/P cells; however, it onlymarginally affected NTUB1 cells (Fig. 2B). Furthermore, we ana-lyzed the anticancer effects of gefitinib and CDDP cotreatment. Inthe combined treatment, gefitinib increased NTUB1/P cell sensi-tivity to CDDP (Fig. 2C, compare lanes 4 and 8) and promotedNTUB1 cell death (Fig. 2C, compare lanes 3 and 7). These resultsimply that EGFR plays a critical role in the CDDP resistance ofNTUB1/P cells.
Abrogating STAT3 activation inhibits CEBPD expression andenhances CDDP efficiency in CDDP-resistant UCUB
CDDP induced EGFR signaling in NTUB1/P cells (Fig. 2), andactivated p38 MAPK and STAT3 have been suggested toplay crucial roles in CEBPD transcriptional activation (8). There-fore, we analyzed STAT3 and p38 activation in the EGF-treatedNTUB1/P cells, revealing that EGF induced the activation ofSTAT3 (pY705) but not p38 MAPK in NTUB1/P cells (Fig. 2D).In addition to CEBPD suppression through attenuated STAT3activation, CEBPD was suppressed in gefitinib-treated NTUB1/Pcells (Supplementary Fig. S2A). To further determine whetherSTAT3 mediated the EGF-induced CEBPD expression in UCUBcells, we examined S3I-201 (29, 30). We first determined theeffects of S3I-201 on NTUB1/P cells treated with either CDDP or
EGF. Similar to our findings in gefitinib treatment in CDDP-treated NTUB1/P and J82 cells, S3I-201 inhibited CDDP- or EGF-induced STAT3 activation and CEBPD expression (Fig. 2E andSupplementary Fig. S2B).Moreover, an in vivoDNA-binding assayrevealed that the binding of activated STAT3 (pY705) in responseto CDDP treatment was attenuated in NTUB1/P cells treated withboth CDDP and S3I-201 (Fig. 2F).
CEBPD mediates CDDP-induced ABCB1 expressionAs previously mentioned, ABC transporters contribute to
drug resistance by increasing drug efflux, thereby reducingtoxicity. Therefore, we assessed which ABC transporters couldrespond to CEBPD induction in NTUB1/P cells. Among theABC transporters, ABCB1 and ABCC2 demonstrated specifictranscript upregulation in response to CEBPD treatment (Sup-plementary Fig. S3A). High ABCB1 levels were observed inpatients with UCUB (31, 32) and have been suggested to beinvolved in paclitaxel resistance. We observed that ABCB1transcripts were present at higher levels in NTUB1/P cells andwere further activated in NTUB1/P cells treated with CDDP;however, this activation did not occur in NTUB1 cells (Fig. 3A).Furthermore, exogenously expressed CEBPD induced ABCB1transcription in NTUB1/P cells (Fig. 3B), and CEBPD knock-down attenuated the CDDP-induced ABCB1 reporter activity(Fig. 3C). RT-PCR was used for determining whether exon 1 ofABCB1 could be transcribed in NTUB1/P cells (SupplementaryFig. S3B). Various 50-flanking regions and ABCB1 locus frag-ments were cloned into a reporter vector for CEBPD-responsiveregion identification (Fig. 3D, left, exon and intron mappingaccording to ABCB1 cDNA clone NM_000927.4). Six putativeCEBPD-binding motifs within the aforementioned fragmentswere predicted using the TFSEARCH program (http://www.cbrc.jp/research/db/TFSEARCH.html), and a reporter assayrevealed that the �21/þ386 bp region was significantly respon-sive to exogenous CEBPD expression (Fig. 3D). To determinewhether CEBPD can directly bind the CEBPD-responsive motifswithin the �21/þ386 bp region, we performed a ChIP assay.The results showed CEBPD binding, which was responsive toCDDP treatment in NTUB1/P cells (Fig. 3E). These resultssuggested that CDDP-induced CEBPD directly binds to ABCB1exon 1 and upregulates its transcription.
Gefitinib and S3I-201 reduce ABCB1 and ABCC2 transcriptlevels and enhance CDDP and paclitaxel sensitivity inCDDP-resistant NTUB1/P and J82 cells
We next assessed ABCC2 and ABCB1 mRNA levels followingthe treatment with either CDDP and gefitinib or CDDP and S3I-201 in NTUB1/P and J82 cells. The ABCC2 and ABCB1 transcriptlevels were responsive to CDDP treatment; however, they weresuppressed in NTUB1/P (Fig. 4A) and J82 cells (SupplementaryFig. S4A and S4B), following the treatment with either gefitinib orS3I-201. In addition, an efflux assay assessing the ABCB1 trans-porter activity was used for examining CDDP-induced paclitaxelresistance. The results revealed that gefitinib and S3I-201 inhibitfluorescent MDR indicator efflux in NTUB1/P and J82 cells(Fig. 4B and Supplementary Fig. S4C). These results suggestedthat gefitinib- and S3I-201–attenuated ABCB1 enhanced CDDPcytotoxicity in CDDP-resistant UCUB cells.
Our results suggested that the EGF/pSTAT3 signaling axismediates CDDP-induced CEBPD transcription and thatincreased CEBPD contributes to ABCB1 transcription and
Targeting CEBPD Prevents Drug Resistance in UCUB
www.aacrjournals.org Clin Cancer Res; 23(2) January 15, 2017 507
on April 8, 2021. © 2017 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from
Published OnlineFirst July 19, 2016; DOI: 10.1158/1078-0432.CCR-15-1169

expression in CDDP-resistant UCUB cells. We further demon-strated that gefitinib and S3I-201 increased NTUB1/P and J82cell sensitivity to CDDP and paclitaxel (Fig. 4C and D andSupplementary Fig. S4D, respectively). These results suggestedthat gefitinib and S3I-201 recover CDDP and paclitaxel sensi-tivity in CDDP-resistant cells. In addition, CDDP and paclitaxelare the substrate of ABCC2 and ABCB1, respectively (21). Tofurther assess the specific involvement of ABCB1 and ABCC2 incross-resistance, a cell viability assay was conducted in ABCB1or ABCC2 knockdown NTUB1/P cells. The results showed that
the loss of ABCC2 in NTUB1/P cells is sensitive to CDDP butminer to paclitaxel treatment (Fig. 4E). Moreover, the loss ofABCB1 in NTUB1/P cells is specifically sensitive to paclitaxelbut not to CDDP treatment (Fig. 4F). In addition, to verify thatCEBPD expression is correlated with pEGFR, pSTAT3, andABCB1 expression in patients with UCUB, we performedimmunohistochemical assays. The results revealed that CEBPDexpression was significantly associated with EGFR and STAT3activation (P ¼ 0.010 and <0.001, respectively). Moreover,ABCB1 expression in UCUB was significantly associated with
NTUCDDP - -+ +
NTP
pEGFR30
NTUNTP
20
10
CDDPGefitinib
EGF 0 0.5 1 2 4 h
pSTAT3
STAT3
pp38
p38
CEBPD
CEBPD
+1-279
-152-386-734-1320
-2,000pSTAT3
Gefitinib
S31-201CDDP
00
10
20
NTUNTP
Cel
l dea
th (%
)
1 5 10 (mmol/L)
STAT3
CEBPD
CDDPS3I-201
Input
-- -
-- - -
--
++
+++
+++
+apSTAT3 aIgG
A
B
AB
IP
a-Tubulin
pSTAT3
EGF
S3I-201 0 6 12 h
STAT3
CEBPD
a-Tubulin
a-Tubulin
+ +++-
---
+ ++-
--
Cel
l dea
th (%
)
0
EGFR
pSTAT3
STAT3
a-Tubulin
A C
DB
E F
Figure 2.
Abrogation of EGFR and STAT3 activationinhibits EGF and CDDP-induced CEBPDexpression in CDDP-resistant NTUB1/P cells.A, EGFR and STAT3 expression and activitylevels increased in NTUB1/P (NTP) cells.Western blot analysis was performed usinglysates from NTUB1 (NTU) and NTP cellstreated with CDDP for 24 hours and theindicated specific pEGFR (pY1085), pSTAT3(pY705), EGFR, STAT3, and a-tubulinantibodies. B, Gefitinib attenuated NTU andNTP cell survival. NTU and NTP cells weretreated with gefitinib at the indicated dosesfor 24 hours, and cell death was examinedusing propidium iodine staining. C, Gefitinibenhances NTU and NTP cell sensitization toCDDP. NTU and NTP cells were treated witheither gefitinib or CDDP for 24 hours, and celldeath was assayed using PI staining. D,CEBPD expression in NTP cells is associatedwith STAT3 activity following EGFtreatment. NTP cells were treated with EGFfor the indicated periods, and lysates wereharvested at the indicated time points.Antibodies recognizing pSTAT3 (pY705),STAT3, pp38 (T182/Y180), p38, CEBPD, anda-tubulin were used for Western blotanalysis. E, A STAT3 inhibitor, S3I-201,inhibits EGF and CDDP-activated CEBPDexpression. Western blot analysis wasperformed using the indicated antibodiesand lysates of cells pretreated with S3I-201for 1 hour and then stimulated with EGF orCDDP for 24 hours. F, CDDP enhancesbinding of STAT3 to the CEBPD promoter inNTP cells. Sonicated chromatin wasemployed in ChIP assays by using STAT3 orcontrol IgG antibodies and then subjected toPCR analysis after eliminating DNA-bindingproteins, as described in the Materials andMethods. In this figure, ��� , �� , and � denote asignificant difference with P < 0.001, <0.01,and <0.05, respectively.
Wang et al.
Clin Cancer Res; 23(2) January 15, 2017 Clinical Cancer Research508
on April 8, 2021. © 2017 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from
Published OnlineFirst July 19, 2016; DOI: 10.1158/1078-0432.CCR-15-1169

high CEBPD and pSTAT3 levels (P < 0.001; Table 1 andSupplementary Table S2; Fig. 5).
As previously mentioned, S3I-201 can abolish CEBPD acti-vation to promote the CDDP sensitivity of CDDP-resistantUCUB cells. To determine whether CEBPD and ABCB1 werealso responsive to CDDP treatment and were suppressed by
gefitinib and S3I-201, we studied a CDDP-resistant nasopha-ryngeal HONE1 cancer cell line, HONE1/R (SupplementaryFig. S4E). CEBPD and ABCB1 expression was higher inHONE1/R cells than in parental HONE1 cells (SupplementaryFig. S4F). As observed for NTUB1/P cells, S3I-201 also inhib-ited ABCB1 transporter efflux activity (Supplementary
64
3
2
Fold
s of
rela
tive ABCB1
mR
NA
Fold
s of
rela
tive
luci
fera
se a
ctiv
ity
Fold
s of
rela
tive ABCB1
mR
NA
1
0CTL CD CDDP
0
0
Luc
Luc
Luc
Luc
+917
+917
+917
+640
+386
-21
-1,000
-950 -657 -15
-21
+250
+163exon 1
+917
CDDP
AB
- + - -+ +Input
IP
A
B
aCEBPD aIgG
-21 +163 +917exon 1
exon 1
-1,000
-1,000 -1
10 20 30
CTLCDAB-C
AB-I
AB-II
AB-III
1
2
3 shLacZshCD2
Folds of relative luciferase activity
- +
CTLA B C
D
E
CDDP
4
2
0NTU NTP
Figure 3.
CEBPD upregulates ABCB1 expression following CDDP treatment. A, ABCB1 transcripts are abundant in NTUB1/P (NTP) cells. qPCR assays were performed usingtotal RNA harvested from NTUB1 (NTU) or NTP cells treated with CDDP for 24 hours. B, CEBPD induces ABCB1 transcription. qPCR assays were performedusing the total RNA harvested from NTP cells transfected with expression vectors with and without CEBPD cDNA [CD and control (CTL), respectively]. C, Loss ofCEBPD attenuates CDDP-induced ABCB1 reporter activity. NTP cells were pretreated with lentiviruses containing shLacZ or shCEBPD (shCD2), transfected withABCB1 reporter (AB-I, diagram shown as follows), and treated with or without CDDP. Luciferase assays were performed after 24 hours by using lysatesharvested from the experimental cells. D, Identification of the CEBPD-responsive region within the ABCB1 50-flanking region and intron 1. Schematic of the putativeCEBPD motifs (oval) within the human ABCB1 50-flanking region and exon/intron 1 (�1,000 to þ917 bp) and various ABCB1 reporters. For reporter assays,NTP cells were transfectedwith the indicatedABCB1 reporters and pCDNA3/HAwith or without CEBPD cDNA (CD and CTL, respectively). E, CDDP enhances CEBPDbinding to the ABCB1 region containing putative CEBPD-binding motifs in NTP cells. Sonicated chromatin was subjected to ChIP-PCR analysis by usingCEBPD or control IgG antibodies, as described in the Materials and Methods. In this figure, ��� denotes a significant difference (P < 0.001).
Targeting CEBPD Prevents Drug Resistance in UCUB
www.aacrjournals.org Clin Cancer Res; 23(2) January 15, 2017 509
on April 8, 2021. © 2017 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from
Published OnlineFirst July 19, 2016; DOI: 10.1158/1078-0432.CCR-15-1169

A3 ABCC2
ABCB1
2
Rel
ativ
e m
RN
A le
vels
(fo
ld)
Rel
ativ
e fl
uo
resc
ence
un
its
(x10
3, 4
85/5
20 n
m)
Cel
l via
bili
ty (
vs. c
on
tro
l)
Cel
l via
bili
ty (
vs. c
on
tro
l)
1
6
4
2
00CDDP
Gefitinib
S3I-201
CDDPGefitinib
S3I-201+
+++ + + + +
+
+- + ++
+
+
-- - -
----
- -
-
-
- --
-
-- -
-
CDDPGefitinib
S3I-201
PTXGefitinib
LacZ
shABCB1LacZ
shABCC2
S3I-201
++
+ + +
++
- + +++
+
-
PTX + +- -
- - ----
-- -
--
---
CDDP + +- -- -
--PTX + +- -
CDDP + +- -- -
--
--
B
C D
E F
1.5
1
0.5
0
1.5
1.0
0.5
0
Cel
l via
bili
ty (
vs. c
on
tro
l)
Cel
l via
bili
ty (
vs. c
on
tro
l)
1.5
1
0.5
0
1.5
1
0.5
0
Figure 4.
EGFR/STAT3 inhibition attenuatestransporter efflux and promotes CDDP-or paclitaxel (PTX)-induced death inNTUB1/P cells. A, Gefitinib and S3I-201inhibit CDDP-induced ABCC2 andABCB1 transcription. NTUB1/P (NTP)cellswere treatedwith CDDP alone or incombination with either S3I-201 orgefitinib for 24 hours. qPCR assayswereused for measuring ABCC2 and ABCB1transcript levels. B, Gefitinib and S3I-201–mediated the inhibition of ABCB1transporter pump activity in NTP cells.NTP cellswere treatedwith CDDP aloneor in combinationwith either gefitinib orS3I-201 for 24 hours. An MDR dye-loading solution was then added toeach well and incubated for 4 hours.Fluorescence intensity was thendetected using an ELISA reader. C andD, Gefitinib and S3I-201 enhance NTPcells sensitivity to CDDP and paclitaxel.NTP cells were treated with CDDP orpaclitaxel alone or combination with orwithout either gefitinib or S3I-201 for24 hours. Cell viability was measuredusing the CCK8 assay. In this figure,��� denotes a significant difference(P < 0.001). E and F, Attenuation ofABCC2 and ABCB1 in NTP cellssensitizes to CDDP and paclitaxel,respectively. NTP cells were pretreatedwith lentiviruses containing shLacZ(LacZ) or shABCC2 or shABCB1. After48 hours of incubation, experimentalcells were treated with CDDP orpaclitaxel for 24 hours and thenexamined for cell viability by CCK8assay. ��� , significant difference(P < 0.001); �� , significant difference(P < 0.05).
Wang et al.
Clin Cancer Res; 23(2) January 15, 2017 Clinical Cancer Research510
on April 8, 2021. © 2017 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from
Published OnlineFirst July 19, 2016; DOI: 10.1158/1078-0432.CCR-15-1169

Fig. S4G) and sensitized HONE1/R cells to CDDP (Supple-mentary Fig. S4H).
Gefitinib and S3I-201 significantly enhance CDDP therapeuticefficacy in CDDP-resistant UCUB
Our in vitro results suggested that gefitinib and S3I-201 treat-ment significantly sensitized CDDP-resistant UCUB cells toCDDP. To further evaluate these results in vivo, the effects ofCDDP, gefitinib, S3I-2-1, combinations of CDDP/gefitinib andCDDP/S3I-201 treatments were examined in a NTUB1/P-xeno-grafted NOD/SCID mouse model. In accordance with previousresults, gefitinib and S3I-201 have a lesser effect on NTUB1/Ptumor burden (58% and 52% inhibition, respectively, at day 9).Importantly, CDDP/gefitinib and CDDP/S3I-201 treatments sig-nificantly reduced the tumor burden (82% and 81% inhibition,respectively, at day 9) and weight (Fig. 5). Furthermore, pSTAT3,CEBPD, and ABCB1 levels in NTUB1/P tumors were verified. Asshown in Supplementary Fig. S6A and S6B, pSTAT3, CEBPD, andABCB1 levels and ABCC2 mRNA levels were reduced in theCDDP/gefitinib- and CDDP/S3I-201–treated NTUB1/P xeno-grafted NOD/SCID mice; this result was consistent with thereduction in tumor size.
DiscussionCDDP and paclitaxel are widely used in UCUB therapy. CDDP
ultimately triggers cell death and is commonly used in the clinicaltherapy of many cancers, including UCUB. Resistance can be pre-existing (intrinsic) or drug induced (acquired). As previouslymentioned, UCUB therapy remains associated with persistenthigh rates of local and distant failure resulting from the acquisi-tion of chemoresistance (33). CEBPD expression was higher inpatients withUCUB than in patients with normal urothelial tissue(34). Our findings indicated that CEBPD upregulated ABCB1 and
ABCC2 expression, revealing an increased drug efflux activity inCEBPD-abundant UCUB cells. These results suggested not onlythat CEBPD plays a functional role in intrinsic and acquiredresistance but also explained why patients with UCUB tend todevelop drug resistance and become desensitized to CDDP andpaclitaxel (35, 36).
CEBPD is a CDDP-responsive gene. In this study, we dem-onstrated that the EGFR/STAT3 signaling axis participates inCDDP-induced CEBPD transcription in UCUB cells, contribut-ing to CDDP and CDDP-induced cross-resistance to paclitaxelby activating ABCC2 and ABCB1 expression (Fig. 3). This studyalso provided a novel insight into the mechanism underlyingacquired resistance and linked EGFR signaling with ABCB1-mediated paclitaxel cross-resistance in CDDP-resistant UCUB.Furthermore, treatment with either gefitinib or S3I-201 reducedABCB1 expression in CDDP-resistant HONE1 cells and inCDDP-resistant UCUB cells. Thus, CEBPD plays a functionalrole in promoting drug resistance, and CEBPD inhibition maybe an effective approach for reducing the occurrence of drugresistance.
Forced CEBPD overexpression induces growth arrest and deathin cancer cells (9). Increased CEBPD levels have been observed inresponse to treatment with HMDB (10), dexamethasone (37),retinoic acid (37), and 5-azacytidine (38). Furthermore, CEBPDhave been suggested to contribute to apoptosis. It has beenrecently suggested to play dual roles in pro- and antitumorprocesses under certain conditions (39–41). Furthermore, upre-gulated caspase-3 and caspase-8 activity has been reported inresponse to CEBPD induction in prostate cancer (41), andCEBPDwas involved in antiapoptosis in astroglioma U373MG cells byupregulating the antiapoptotic gene ZNF179 (40). These observa-tions imply that CEBPD may serve dual roles in controlling cellsurvival and regulating cell fate in a cell-specific manner. Herein,we revealed that the EGFR/STAT3 pathway contributes to CEBPD
Table 1. Correlations between expression status of pEGFR, pSTAT3, CEBPD, and ABCB1 in muscle-invasive UCUB
pSTAT3 Exp. CEBPD Exp. ABCB1 Exp.Parameter Expression status Case no. Low High P Low High P Low High P
pEGFR Low Exp. 39 23 16 0.092 25 14 0.010a 25 14 0.010a
High Exp. 40 16 24 14 26 14 26
pSTAT3 Low Exp. 39 — — — 29 10 <0.001a 28 11 <0.001a
High Exp. 40 — — — 10 30 11 29
CEBPD Low Exp. 39 — — — — — — 32 7 <0.001a
High Exp. 40 — — — — — — 7 33aStatistically significant (bold values).
1,500 CTLCDDPGefitinibS3I-201CDDP+gefitinibCDDP+S3I-201
1,000
500
00 3 6 9
0.2
0
0.4
Tum
or w
eigh
t (g)
Tum
or s
ize
(mm
3 ) 0.6
0.8
(Days) CDDPGefitinibS3I-201
- ++
+
++
+
+--
-- -
- --- -
Figure 5.
Antitumor effects of CDDP treatment alone or incombination with either gefitinib or S3I-201 on NTUB1/P-xenografted NOD/SCID mice. Effects of the intraperitonealadministration of CDDP, gefitinib, S3I-201, CDDP/gefitinib,or CDDP/S3I-201 on NTUB1/P (NTP)-xenograftedNOD/SCID mice. A total of 24 NOD/SCID mice weresubcutaneously inoculated with NTP cells (1 � 107) anddivided into 6 groups. After solid tumor formation, theanimals were intraperitoneally injected with a controlvehicle or CDDP (5 mg/kg/2 days), gefitinib (10 mg/kg/2days), S3I-201 (10 mg/kg/2 days), CDDP and gefitinib, orCDDP and S3I-201.
Targeting CEBPD Prevents Drug Resistance in UCUB
www.aacrjournals.org Clin Cancer Res; 23(2) January 15, 2017 511
on April 8, 2021. © 2017 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from
Published OnlineFirst July 19, 2016; DOI: 10.1158/1078-0432.CCR-15-1169

activation and confers a drug-resistant activity that includes cross-resistance. In addition, a previous study showed that the over-expression of EGFR is sensitive to CDDP treatment through aligand-independent pathway in U87 glioma and MB468 breastcancer cells (42). In our system, the basal levels of pEGFR andpSTAT3 are higher and coincident with basal CEBPD abundancein NTUB1/P cells (Fig. 1A and Supplementary Fig. S7). Therefore,treatment of CDDP could further enhance the activities of pEGFRand pSTAT3 and coordinately CEBPD abundance in NTUB1/Pcells (Fig. 2A), indicating a unique but unknownprotein/pathwayis responsive to CDDP in the CDDP-resistant cells to strengthenthe activation of pEGFR and pSTAT3. More importantly, it alsoprovided a motivation for initiation of clinical studies withinhibition of the EGFR/STAT3 signaling pathwaywhen combinedwith CDDP to prevent the development of therapeutic resistanceand thus significantly impact overall survival in patients withUCUB.
P-gp was the first factor known to modulate MDR and is themost widely studied ATP-dependent drug efflux pump. P-gp isencodedbyABCB1 and is considered tomediateMDRby reducingthe intracellular accumulation of cytotoxic drugs and com-pounds. Many types of cancers express high ABCB1 levels andare unresponsive to chemotherapy (43, 44); ABCB1 expressionhas been considered a predictor of poor response to chemother-apy and overall survival (45, 46). Our results also indicated thatUCUB cells with high expression of CEBPD are strongly resistantto CDDP and paclitaxel because of ABCC2 and ABCB1 activation(Fig. 4), particularly in CDDP-resistant UCUB cells. Thus, wespeculated that CEBPD activation should, at least in part, con-tribute to the proapoptotic activity in response to anticancerdrugs; however, its activation conferred acquired resistance viaABCB1 and ABCC2, promoting the resistance of anticancer drug-induced cytotoxicity. In addition to forming a homodimer,CEBPD can heterodimerize with other C/EBP family members,including CEBPA and CEBPB (47). Moreover, ABCB1 is upregu-lated by CEBPD and CEBPB in anticancer drug-resistant breastcancer cells (48). In accordance with this study, our results alsosuggested the importance of examining the response and involve-ment of CEBPB in CDDP-treated and -resistant UCUB cells in thefuture. The association between ABC transporters with bladdercancers was previously established. Recent studies have reportedthe overexpression of ABC transporters in chemotherapeuticpatients with bladder cancer (32, 49). CEBPD is also responsiveto stimuli involving the activation of either p38 MAPK or STAT3.Therefore, it is reasonable to speculate that anticancer treatments,particularly those promoting CEBPD overexpression and p38MAPK and STAT3 activation, tend to lead to the development ofdrug-resistant cancers.
Chemoresistance in several cancer types has been linked toSTAT3activation. Recently, constitutively active STAT3was shownto lead to cellular transformation, suggesting an oncogenic role.STAT3 has also been suggested to confer an enhanced cellularsurvival following genotoxic treatments (11). Moreover, STAT3pathway inhibition has been shown to result in growth arrest,apoptosis, and chemosensitivity in several human malignancymodels (50).We previously demonstrated that CEBPD inhibitionincreased UCUB sensitivity to CDDP (19). In this study, wedemonstrated that CEBPD attenuation in UCUB and CDDP-resistant UCUB cells sensitized them to CDDP and CDDP-induced paclitaxel resistance (Fig. 1). We further revealed thenovel finding that ABCB1 responds to CEBPD upon CDDPtreatment by activating EGFR and STAT3 in UCUB and CDDP-resistantUCUB cells. Therefore, inactivating either EGFRor STAT3can sensitizeUCUBandCDDP-resistantUCUB cells toCDDPandeven to paclitaxel. In addition to gefitinib, we demonstrated thatS3I-201 inhibited CDDP-induced CEBPD and augmented CDDPand paclitaxel sensitivity.
Disclosure of Potential Conflicts of InterestNo potential conflicts of interest were disclosed.
Authors' ContributionsConception and design: W.-J. Wang, C.-F. Li, W.-C. Chang, J.-M. WangDevelopment of methodology: W.-J. Wang, Y.-Y. ChuAcquisition of data (provided animals, acquired and managed patients,provided facilities, etc.):W.-J. Wang, C.-F. Li, Y.-H. Wang, T.-C. Hour, C.-J. YenAnalysis and interpretation of data (e.g., statistical analysis, biostatistics,computational analysis): W.-J. Wang, C.-F. Li, W.-C. ChangWriting, review, and/or revision of the manuscript: W.-J. Wang, C.-F. Li,C.-J. Yen, J.-M. WangAdministrative, technical, or material support (i.e., reporting or organizingdata, constructing databases): W.-J. WangStudy supervision: W.-J. Wang, T.-C. Hour, J.-M. WangOther (helped to organize the communication of authors): W.-J. WangOther (collaborated with J.-M. Wang in this study and co-advisedW.-J. Wangwith J.-M. Wang): W.-C. Chang
Grant SupportThis work was supported from the grant MOST103-2321-B-006-031,
MOST103-2320-B-006-034-MY3 from the Ministry of Science and TechnologyandMOHW104- TDU-B-212–124-003,MOHW105-TDU-B-212-134-007 fromthe Ministry of Health and Welfare, Health and welfare surcharge of tobaccoproducts.
The costs of publication of this articlewere defrayed in part by the payment ofpage charges. This article must therefore be hereby marked advertisement inaccordance with 18 U.S.C. Section 1734 solely to indicate this fact.
Received May 25, 2015; revised June 28, 2016; accepted July 2, 2016;published OnlineFirst July 19, 2016.
References1. Wang D, Lippard SJ. Cellular processing of platinum anticancer drugs. Nat
Rev Drug Discov 2005;4:307–20.2. Bellmunt J, Guillem V, Paz-Ares L, Gonzalez-Larriba JL, Carles J, Albanell J,
et al. Gemcitabine/paclitaxel-based three-drug regimens in advancedurothelial cancer. Eur J Cancer 2000;36 Suppl 2:17–25.
3. Colquhoun AJ, Mellon JK. Epidermal growth factor receptor and bladdercancer. Postgrad Med J 2002;78:584–9.
4. Neal DE, Sharples L, Smith K, Fennelly J, Hall RR, Harris AL. The epidermalgrowth factor receptor and the prognosis of bladder cancer. Cancer1990;65:1619–25.
5. Rebouissou S, Bernard-Pierrot I, de Reynies A, Lepage ML, Krucker C,Chapeaublanc E, et al. EGFR as a potential therapeutic target for a subsetof muscle-invasive bladder cancers presenting a basal-like phenotype.Sci Transl Med 2014;6:244ra91.
6. Ciardiello F, Caputo R, Bianco R, Damiano V, Pomatico G, De Placido S,et al. Antitumor effect andpotentiationof cytotoxic drugs activity in humancancer cells by ZD-1839 (Iressa), an epidermal growth factor receptor-selective tyrosine kinase inhibitor. Clin Cancer Res 2000;6:2053–63.
7. Shrader M, Pino MS, Lashinger L, Bar-Eli M, Adam L, Dinney CP, et al.Gefitinib reverses TRAIL resistance in human bladder cancer cell lines via
Wang et al.
Clin Cancer Res; 23(2) January 15, 2017 Clinical Cancer Research512
on April 8, 2021. © 2017 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from
Published OnlineFirst July 19, 2016; DOI: 10.1158/1078-0432.CCR-15-1169

inhibition of AKT-mediated X-linked inhibitor of apoptosis protein expres-sion. Cancer Res 2007;67:1430–5.
8. Wang JM, Tseng JT, Chang WC. Induction of human NF-IL6beta byepidermal growth factor is mediated through the p38 signaling pathwayand cAMP response element-binding protein activation in A431 cells.Mol Biol Cell 2005;16:3365–76.
9. Ikezoe T, Gery S, Yin D, O'Kelly J, Binderup L, Lemp N, et al. CCAAT/enhancer-binding protein delta: a molecular target of 1,25-dihydroxyvita-min D3 in androgen-responsive prostate cancer LNCaP cells. Cancer Res2005;65:4762–8.
10. Pan YC, Li CF, Ko CY, PanMH, Chen PJ, Tseng JT, et al. CEBPD reverses RB/E2F1-mediated gene repression and participates in HMDB-inducedapoptosis of cancer cells. Clin Cancer Res 2010;16:5770–80.
11. Sanford DC, DeWille JW. C/EBPdelta is a downstream mediator of IL-6induced growth inhibition of prostate cancer cells. Prostate 2005;63:143–54.
12. Ko CY, Chang LH, Lee YC, Sterneck E, Cheng CP, Chen SH, et al. CCAAT/enhancer binding protein delta (CEBPD) elevating PTX3 expression inhi-bits macrophage-mediated phagocytosis of dying neuron cells. NeurobiolAging 2012;33:422.
13. Wang JM, Ko CY, Chen LC, Wang WL, Chang WC. Functional role of NF-IL6beta and its sumoylation and acetylation modifications in promoteractivation of cyclooxygenase 2 gene. Nucleic Acids Res 2006;34:217–31.
14. Ko CY, Hsu HC, Shen MR, Chang WC, Wang JM. Epigenetic silencing ofCCAAT/enhancer-binding protein delta activity by YY1/polycomb group/DNA methyltransferase complex. J Biol Chem 2008;283:30919–32.
15. Tang D, Sivko GS, DeWille JW. Promoter methylation reduces C/EBPdelta(CEBPD) gene expression in the SUM-52PE human breast cancer cell lineand in primary breast tumors. Breast Cancer Res Treat 2006;95:161–70.
16. Gery S, Tanosaki S, Hofmann WK, Koppel A, Koeffler HP. C/EBPdeltaexpression in a BCR-ABL-positive cell line induces growth arrest andmyeloid differentiation. Oncogene 2005;24:1589–97.
17. Wu SR, Li CF, Hung LY, Huang AM, Tseng JT, Tsou JH, et al. CCAAT/enhancer-binding protein delta mediates tumor necrosis factor alpha-induced Aurora kinase C transcription and promotes genomic instability.J Biol Chem 2011;286:28662–70.
18. Balamurugan K,Wang JM, Tsai HH, Sharan S, AnverM, Leighty R, et al. Thetumour suppressor C/EBPdelta inhibits FBXW7 expression and promotesmammary tumour metastasis. EMBO J 2010;29:4106–17.
19. Hour TC, Lai YL, Kuan CI, Chou CK,Wang JM, TuHY, et al. Transcriptionalup-regulation of SOD1 by CEBPD: a potential target for cisplatin resistanthuman urothelial carcinoma cells. Biochem Pharmacol 2010;80:325–34.
20. Taniguchi K, Wada M, Kohno K, Nakamura T, Kawabe T, Kawakami M,et al. A human canalicular multispecific organic anion transporter(cMOAT) gene is overexpressed in cisplatin-resistant human cancer celllines with decreased drug accumulation. Cancer Res 1996;56:4124–9.
21. Fletcher JI,HaberM,HendersonMJ,NorrisMD.ABC transporters in cancer:more than just drug efflux pumps. Nat Rev Cancer 2010;10:147–56.
22. Li CF, Wu WJ, Wu WR, Liao YJ, Chen LR, Huang CN, et al. The cAMPresponsive element binding protein 1 transactivates epithelial membraneprotein 2, a potential tumor suppressor in the urinary bladder urothelialcarcinoma. Oncotarget 2015;6:9220–39.
23. Fan EW, Li CC, Wu WJ, Huang CN, Li WM, Ke HL, et al. FGF7 overexpression is an independent prognosticator in patients with urothelialcarcinoma of the upper urinary tract and bladder. J Urol 2015;194:223–9.
24. Yu HJ, Tsai TC, Hsieh TS, Chiu TY. Characterization of a newly establishedhuman bladder carcinoma cell line, NTUB1. J Formos Med Assoc1992;91:608–13.
25. Yeh MY, Yu DS, Chen SC, Lin MS, Chang SY, Ma CP, et al. Establishmentand characterization of a human urinary bladder carcinoma cell line(TSGH-8301). J Surg Oncol 1988;37:177–84.
26. Chi JY, Hsiao YW, Li CF, Lo YC, Lin ZY, Hong JY, et al. Targetingchemotherapy-induced PTX3 in tumor stroma to prevent the progressionof drug-resistant cancers. Oncotarget 2015;6:23987–4001.
27. Li CF, Tsai HH, Ko CY, Pan YC, Yen CJ, Lai HY, et al. HMDB and 5-AzadCcombination reverses tumor suppressor CCAAT/enhancer-binding proteindelta to strengthen the death of liver cancer cells. Mol Cancer Ther2015;14:2623–33.
28. Kim WT, Kim J, Yan C, Jeong P, Choi SY, Lee OJ, et al. S100A9 and EGFRgene signatures predict disease progression in muscle invasive bladdercancer patients after chemotherapy. Ann Oncol 2014;25:974–9.
29. Lin L, Deangelis S, Foust E, Fuchs J, Li C, Li PK, et al. A novel smallmoleculeinhibits STAT3 phosphorylation and DNA binding activity and exhibitspotent growth suppressive activity in human cancer cells. Mol Cancer2010;9:217.
30. Siddiquee K, Zhang S, Guida WC, Blaskovich MA, Greedy B, Lawrence HR,et al. Selective chemical probe inhibitor of Stat3, identified throughstructure-based virtual screening, induces antitumor activity. Proc NatlAcad Sci U S A 2007;104:7391–6.
31. Clifford SC, Thomas DJ, Neal DE, Lunec J. Increased mdr1 gene transcriptlevels in high-grade carcinoma of the bladder determined by quantitativePCR-based assay. Br J Cancer 1994;69:680–6.
32. Tada Y, Wada M, Kuroiwa K, Kinugawa N, Harada T, Nagayama J, et al.MDR1 gene overexpression and altered degree of methylation at thepromoter region in bladder cancer during chemotherapeutic treatment.Clin Cancer Res 2000;6:4618–27.
33. Piulats JM, Jimenez L, Garcia del Muro X, Villanueva A, Vinals F, Germa-Lluch JR. Molecular mechanisms behind the resistance of cisplatin in germcell tumours. Clin Transl Oncol 2009;11:780–6.
34. Wang YH, Wu WJ, Wang WJ, Huang HY, Li WM, Yeh BW, et al. CEBPDamplification and overexpression in urothelial carcinoma: a driver oftumor metastasis indicating adverse prognosis. Oncotarget 2015;6:31069–84.
35. Yagoda A. Chemotherapy of urothelial tract tumors. Cancer 1987;60:574–85.
36. Gosland M, Lum B, Schimmelpfennig J, Baker J, Doukas M. Insights intomechanisms of cisplatin resistance and potential for its clinical reversal.Pharmacotherapy 1996;16:16–39.
37. Yeh WC, Cao Z, Classon M, McKnight SL. Cascade regulation of terminaladipocyte differentiation by three members of the C/EBP family of leucinezipper proteins. Genes Dev 1995;9:168–81.
38. Weiland T, Weiller M, Kunstle G, Wendel A. Sensitization by 5-azacytidinetoward death receptor-induced hepatic apoptosis. J Pharmacol Exp Ther2009;328:107–15.
39. Balamurugan K, Sterneck E. The many faces of C/EBPdelta and theirrelevance for inflammation and cancer. Int J Biol Sci 2013;9:917–33.
40. Wang SM, Lee YC, Ko CY, Lai MD, Lin DY, Pao PC, et al. Increase of zincfinger protein 179 in response to CCAAT/enhancer binding protein deltaconferring an antiapoptotic effect in astrocytes of Alzheimer's disease. MolNeurobiol 2015;51:370–82.
41. Chuang CH, Wang WJ, Li CF, Ko CY, Chou YH, Chuu CP, et al. Thecombination of the prodrugs perforin-CEBPD and perforin-granzyme Befficiently enhances the activation of caspase signaling and kills prostatecancer. Cell Death Dis 2014;5:e1220.
42. BenharM, Engelberg D, Levitzki A. Cisplatin-induced activation of the EGFreceptor. Oncogene 2002;21:8723–31.
43. Weinstein RS, Jakate SM, Dominguez JM, Lebovitz MD, Koukoulis GK,Kuszak JR, et al. Relationship of the expression of the multidrugresistance gene product (P-glycoprotein) in human colon carcinomato local tumor aggressiveness and lymph node metastasis. Cancer Res1991;51:2720–6.
44. Fojo AT, Shen DW, Mickley LA, Pastan I, Gottesman MM. Intrinsic drugresistance inhuman kidney cancer is associatedwith expressionof ahumanmultidrug-resistance gene. J Clin Oncol 1987;5:1922–7.
45. MelguizoC, Prados J, LuqueR,Ortiz R, RamaAR,CabaO, et al.Modulationof multidrug resistance gene expression in peripheral blood mononuclearcells of lung cancer patients and evaluation of their clinical significance.Cancer Chemother Pharmacol 2013;71:537–41.
46. Campa D, Muller P, Edler L, Knoefel L, Barale R, Heussel CP, et al. Acomprehensive study of polymorphisms in ABCB1, ABCC2 and ABCG2and lung cancer chemotherapy response and prognosis. Int J Cancer2012;131:2920–8.
47. Takiguchi M. The C/EBP family of transcription factors in the liver andother organs. Int J Exp Pathol 1998;79:369–91.
48. Conze D, Weiss L, Regen PS, Bhushan A, Weaver D, Johnson P, et al.Autocrine production of interleukin 6 causesmultidrug resistance in breastcancer cells. Cancer Res 2001;61:8851–8.
49. Nakagawa M, Emoto A, Nasu N, Hanada T, Kuwano M, Cole SP, et al.Clinical significance of multi-drug resistance associated protein and P-glycoprotein in patients with bladder cancer. J Urol 1997;157:1260–4.
50. Yu H, Pardoll D, Jove R. STATs in cancer inflammation and immunity: aleading role for STAT3. Nat Rev Cancer 2009;9:798–809.
www.aacrjournals.org Clin Cancer Res; 23(2) January 15, 2017 513
Targeting CEBPD Prevents Drug Resistance in UCUB
on April 8, 2021. © 2017 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from
Published OnlineFirst July 19, 2016; DOI: 10.1158/1078-0432.CCR-15-1169

2017;23:503-513. Published OnlineFirst July 19, 2016.Clin Cancer Res Wei-Jan Wang, Chien-Feng Li, Yu-Yi Chu, et al. Urinary BladderCross-resistance with Paclitaxel in the Urothelial Carcinoma of the Inhibition of the EGFR/STAT3/CEBPD Axis Reverses Cisplatin
Updated version
10.1158/1078-0432.CCR-15-1169doi:
Access the most recent version of this article at:
Material
Supplementary
http://clincancerres.aacrjournals.org/content/suppl/2016/07/19/1078-0432.CCR-15-1169.DC1
Access the most recent supplemental material at:
Cited articles
http://clincancerres.aacrjournals.org/content/23/2/503.full#ref-list-1
This article cites 50 articles, 19 of which you can access for free at:
Citing articles
http://clincancerres.aacrjournals.org/content/23/2/503.full#related-urls
This article has been cited by 1 HighWire-hosted articles. Access the articles at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications Department at
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://clincancerres.aacrjournals.org/content/23/2/503To request permission to re-use all or part of this article, use this link
on April 8, 2021. © 2017 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from
Published OnlineFirst July 19, 2016; DOI: 10.1158/1078-0432.CCR-15-1169