Embed Size (px)
Citation preview

Rapport de Master 2 Science de la Mer et du Littoral,
Mention Sciences biologiques Marine
Ingestion, efficacité d’a
taux d’absorption de quatre
microalgues pour le conditionnement
des géniteurs d’
Stage placé sous la responsabilité du Dr. René Robert et Ricardo González Araya
Laboratoire de Physiologie des Invertébrés Station Expérimentale d’ArgentonPresqu’île du vivier, 29280 Landunvez, France.
Rapport de Master 2 Science de la Mer et du Littoral,
Mention Sciences biologiques Marines
Année universitaire 2007-2008
Ingestion, efficacité d’absorption
absorption de quatre
microalgues pour le conditionnement
des géniteurs d’Ostrea edulis
Présenté par
Laure DAVID
Stage placé sous la responsabilité du Dr. René Robert et Ricardo González Araya
Laboratoire de Physiologie des Invertébrés Marins (UMR PE2M), IFREMERStation Expérimentale d’Argenton
280 Landunvez, France.
Rapport de Master 2 Science de la Mer et du Littoral,
bsorption et
absorption de quatre
microalgues pour le conditionnement
Ostrea edulis (L.)
Stage placé sous la responsabilité du Dr. René Robert et Ricardo González Araya
M), IFREMER

Remerciements :
Je remercie le Dr. René Robert, responsable de ce stage, pour son accueil et ses
conseils lors de l’écriture de ce rapport.
Merci également à Ricardo Gonz
collaboration et son soutien tout au long de mon stage.
Je voudrais aussi remercier l’ensemble de la Station expérimentale d’Argenton
(chercheurs, techniciens, doctorants et stagiaires),
Luc Lebrun qui m’ont approvisionnée en algues pour ces expériences.
~ 2 ~
Je remercie le Dr. René Robert, responsable de ce stage, pour son accueil et ses
conseils lors de l’écriture de ce rapport.
également à Ricardo González Araya, coordinateur de l’étude, pour sa
collaboration et son soutien tout au long de mon stage.
Je voudrais aussi remercier l’ensemble de la Station expérimentale d’Argenton
(chercheurs, techniciens, doctorants et stagiaires), et plus particulièrement Isabelle Queau et
Luc Lebrun qui m’ont approvisionnée en algues pour ces expériences.
Je remercie le Dr. René Robert, responsable de ce stage, pour son accueil et ses
lez Araya, coordinateur de l’étude, pour sa
Je voudrais aussi remercier l’ensemble de la Station expérimentale d’Argenton
et plus particulièrement Isabelle Queau et

Sommaire
INTRODUCTION ................................
MATÉRIELS ET MÉTHODES ................................
Matériels biologiques ................................
Protocoles expérimentaux
Mesures écophysiologiques
Calculs et statistiques ................................
RÉSULTATS ................................
Ajustement de l’apport nutritionnel journalier
Ecophysiologie des géniteurs d’Ostrea edulis
Taux de filtration ................................
Taux de consommation
Taux d’ingestion ................................
Efficacité d’absorption
Taux d’absorption ................................
Biométrie ................................
DISCUSSION ................................
Taux de filtration, taux de consommation, taux d’ingestion
Efficacité d’absorption, absorption
Biométrie ................................
Autres espèces ................................
Emissions larvaires ................................
CONCLUSION ................................
BIBLIOGRAPHIE ................................
ANNEXE 1 ................................
ANNEXE 2 ................................
ANNEXE 3 ................................
ANNEXE 4 ................................
ANNEXE 5 ................................
ANNEXE 6 ................................
ANNEXE 7 ................................
~ 3 ~
................................................................................................................................
................................................................................................
................................................................................................
expérimentaux ................................................................................................
Mesures écophysiologiques ................................................................................................
................................................................................................
................................................................................................................................
Ajustement de l’apport nutritionnel journalier ................................................................
Ecophysiologie des géniteurs d’Ostrea edulis................................................................
................................................................................................................................
Taux de consommation ................................................................................................
................................................................................................................................
Efficacité d’absorption ................................................................................................
................................................................................................................................
................................................................................................................................
................................................................................................................................
Taux de filtration, taux de consommation, taux d’ingestion ................................
Efficacité d’absorption, absorption ................................................................................................
................................................................................................................................
................................................................................................................................
................................................................................................
................................................................................................................................
................................................................................................................................
................................................................................................................................
................................................................................................................................
................................................................................................................................
................................................................................................................................
................................................................................................................................
................................................................................................................................
................................................................................................................................
......................................... 4
......................................................... 7
.............................................................. 7 ....................................................... 8
..................................................... 9 ............................................................ 10
.............................................. 12
..................................................... 12 ........................................................ 13
.......................................... 13 ................................................................ 13
........................................... 14 ................................................................. 14
........................................ 15 ...................................................... 15
............................................. 17
.................................................................... 17 .......................................... 17
.................................................. 18 ......................................... 18
................................................................... 19
........................................... 20
....................................... 21
................................................... 24
................................................... 25
................................................... 26
................................................... 27
................................................... 28
................................................... 29
................................................... 30

Introduction
L’huître plate Ostrea edulis
les eaux proches du Maroc, le long de la Méditerranée et dans la Mer Noire. C’est une huître
native européenne qui est utilisé
2008). Au cours des quarante
chute d'environ 30 000 tonnes
refringens et Bonamia ostrea
écloserie, ont été redirigés vers l’éradication et la lutte contre ces deux maladies. C’est dan
ce contexte que les chercheurs
(Naciri-Graven et al., 1998), mais
solutions reste le pré grossissement
afin d’obtenir des individus prêt à la
dues à ce parasite se produise
françaises (Robert et al., 1991)
Actuellement, la production
faible, avec une production moyenne de 6000 à
cette espèce commerciale emblématique
jusque dans les années 70, est considéré
luxueux, depuis l’augmentation de son
Cette dernière en 2005 est de 1300 tonnes
mondial (25% de la production mondiale
L’irrégularité du recrutement dans les zones d’exploitation, la difficulté de collecter
en milieu naturel certaines espèces de
pathologiques, ainsi que la dégradation de la qualité des eaux côtières, ont amené les
biologistes à développer les techniques de reproduction artificielle par la créatio
d’écloseries (Robert et al., 1996)
plate en écloserie s’avère plus
incube ses larves avant de les
~ 4 ~
Ostrea edulis (Linnaeus, 1758) se rencontre de la Norvège jusque dans
les eaux proches du Maroc, le long de la Méditerranée et dans la Mer Noire. C’est une huître
native européenne qui est utilisée depuis plusieurs siècles comme nourriture humaine
quarante dernières années, la production d’Ostrea edulis
000 tonnes, due principalement à l'impact de deux parasites
Bonamia ostreae). De ce fait, tous les efforts pour améliorer le rendement en
closerie, ont été redirigés vers l’éradication et la lutte contre ces deux maladies. C’est dan
que les chercheurs ont tenté de trouver une souche résistante à la Bonamiose
, mais pour l’instant, celle-ci n’a pas pleinement abouti. Une des
pré grossissement intensif des juvéniles au cours de la
prêt à la commercialisation avant leur 2 ans ½,
oduisent essentiellement à partir du troisième été
., 1991).
a production mondiale de l'huître plate européenne
une production moyenne de 6000 à 7000 tonnes entre 2002 et 200
espèce commerciale emblématique, car forte production française de l’huître plate
jusque dans les années 70, est considérée actuellement comme un produit alimentaire
x, depuis l’augmentation de son prix liée à la diminution de la production
n 2005 est de 1300 tonnes en France, ce qui la place au deuxième rang
e la production mondiale en 2005) derrière l’Espagne.
L’irrégularité du recrutement dans les zones d’exploitation, la difficulté de collecter
certaines espèces de juvéniles, l’augmentation des problèmes
pathologiques, ainsi que la dégradation de la qualité des eaux côtières, ont amené les
ologistes à développer les techniques de reproduction artificielle par la créatio
., 1996). Cependant, la production commerciale de
plus difficile car c’est une espèce larvipare, c'est
es larves avant de les libérer dans le milieu extérieur, contrairement à
) se rencontre de la Norvège jusque dans
les eaux proches du Maroc, le long de la Méditerranée et dans la Mer Noire. C’est une huître
depuis plusieurs siècles comme nourriture humaine (FAO,
edulis a montré une
parasites (Marteilia
pour améliorer le rendement en
closerie, ont été redirigés vers l’éradication et la lutte contre ces deux maladies. C’est dans
de trouver une souche résistante à la Bonamiose
nement abouti. Une des
des juvéniles au cours de la première année,
avant leur 2 ans ½, car les mortalités
partir du troisième été, dans les eaux
de l'huître plate européenne reste encore
7000 tonnes entre 2002 et 2005. En France
, car forte production française de l’huître plate
comme un produit alimentaire
oduction (Annexe 1).
ce qui la place au deuxième rang
L’irrégularité du recrutement dans les zones d’exploitation, la difficulté de collecter
l’augmentation des problèmes
pathologiques, ainsi que la dégradation de la qualité des eaux côtières, ont amené les
ologistes à développer les techniques de reproduction artificielle par la création
de naissain d’huître
, c'est-à-dire qu’elle
contrairement à

Crassostrea gigas, espèce ovipare
extérieur (Marteil, 1976).
Quel que soit l’effort mis en place pour rétab
impératif de passer par un bon conditionnement des géniteurs, afin d’o
de naissain relativement stable dans le temps
Dans une écloserie commerciale
des géniteurs d’une part et su
incubatrice, on peut s’interroger
développement larvaire ultérieur
adaptée était distribuée aux larves,
éléments nécessaires à la fabrication de larve initiale optimale
Figure 1 : Stratégie de conditionnement en écloserie commercial
Ostrea edulis
~ 5 ~
espèce ovipare, qui libère directement ses gamètes dans le milieu
que soit l’effort mis en place pour rétablir la production de cette espèce, il est
impératif de passer par un bon conditionnement des géniteurs, afin d’obtenir une quantité
relativement stable dans le temps (Frolov et al., 1992).
commerciale, les efforts sont concentrés sur le conditionnement
des géniteurs d’une part et sur celui des larves d’autre part. Or, pour Ostrea edulis
s’interroger sur l’influence de la nutrition des géniteurs sur le
ultérieur. En effet, le bénéfice serait réduit si une nourriture
adaptée était distribuée aux larves, sans que les géniteurs n’aient préalablement r
s à la fabrication de larve initiale optimale (Figure 1).
: Stratégie de conditionnement en écloserie commerciale et modèle hypothétique de stratégie à adopter
es gamètes dans le milieu
lir la production de cette espèce, il est
btenir une quantité
sur le conditionnement
Ostrea edulis, espèce
influence de la nutrition des géniteurs sur le
le bénéfice serait réduit si une nourriture
aient préalablement reçu les
pothétique de stratégie à adopter pour

Il est donc essentiel de
les paramètres biotiques et abiotiques au cours du conditionnement. Dans ce contexte, le
rôle de la température et de la salinité
al., 1995 ; Rodstrom et al., 2000)
apporter quotidiennement sont comprises entre
au poids secs des géniteurs (Millican
par Beiras et al. (1994). Dans tous les cas,
Stephenson (1973 in Utting et al
L’aspect qualitatif reste par ailleurs important
au bon développement des lar
et al., 1994 ; Jonsson et al., 1999
Il en est de même pour la compositi
qui conditionne fortement le
al., 1986 ; Ferreiro et al., 1990
(Wilson et al., 1996 ; Chaparro
été argumenté, et rien ne démontre dans ces travaux que ces algues
assimilées par les géniteurs, et que le transfert vers les œufs et les larves
opéré.
Un précédent travail (Gonzales, 2007)
géniteurs sur la fécondité, mais
comportement alimentaire.
C’est dans ce contexte qu
de l’efficacité d’absorption et
écloseries commerciales pour les mollusques en général
critère le meilleur régime nutritionnel
européenne.
~ 6 ~
de bien connaître la physiologie de cette espèce
les paramètres biotiques et abiotiques au cours du conditionnement. Dans ce contexte, le
rôle de la température et de la salinité a été précisé antérieurement (Mann, 1979
., 2000). En ce qui concerne la nutrition, les quantités d’aliment à
apporter quotidiennement sont comprises entre 3 et 7% du poids sec d‘
(Millican et al., 1994), bien supérieures aux 0
ans tous les cas, elles ne doivent pas dépasser
et al., 1997).
reste par ailleurs important car certains nutriments
au bon développement des larves d’O. edulis (Holland et al., 1973 ; Helm
., 1999 ; Labarta et al., 1999) et des juvéniles (Enright
omposition biochimique des microalgues apportées
conditionne fortement le développement larvaire ultérieur de cette espèce
., 1990 ; Frolov et al., 1992 ; Millican et al., 1994)
; Chaparro et al., 2006). Cependant, le choix de ces microalgues n’a pas
, et rien ne démontre dans ces travaux que ces algues sont
s par les géniteurs, et que le transfert vers les œufs et les larves
(Gonzales, 2007) a démontré le fort impact de la nutrition des
géniteurs sur la fécondité, mais ces performances n’ont pu être reliées à des différences de
C’est dans ce contexte qu’une étude approfondie de l’écophysiologie
et de le taux d’absorption de différents types d’algues utilisés
pour les mollusques en général, a été initiée, afin d’établir
eur régime nutritionnel pour le conditionnement
espèce afin d’optimiser
les paramètres biotiques et abiotiques au cours du conditionnement. Dans ce contexte, le
(Mann, 1979 ; Beiras et
. En ce qui concerne la nutrition, les quantités d’aliment à
d‘algues par rapport
aux 0,2 à 3% rapportés
épasser 9% d’après
nutriments sont essentiels
; Helm et al., 1991 ; Heras
(Enright et al., 1986).
on biochimique des microalgues apportées aux géniteurs
de cette espèce (Enright et
., 1994) et Ostrea chilensis
endant, le choix de ces microalgues n’a pas
sont bien ingérées et
s par les géniteurs, et que le transfert vers les œufs et les larves se soit réellement
fort impact de la nutrition des
ces performances n’ont pu être reliées à des différences de
écophysiologie de l’ingestion,
différents types d’algues utilisés en
, afin d’établir sur ce
de l’huître plate

Matériels et Méthodes
Matériels biologiques
Des mesures journalières
d’absorption et de le taux d’absorption
conditionnées sur quatre régimes monospécifiques,
Chaetoceros gracilis, Tetraselmis suecica
� Ostrea edulis
L’huître plate européenne
Anisomyaria, famille Ostreidae
branchies pour pomper les particules alimentaires présentes dans le milieu
(Marteil, 1976) ; c’est le processus d’ingestion
un certain tri des particules s’opère
en boulettes par du mucus,
mouvements secs de fermeture des valves
conduits jusqu’à l’orifice buccal
l’on appelle le taux d’absorption
essentiels tels que les acides
d’assimilation. Les restes non assimilés
(Marteil, 1976). La Figure 2 montre le
digestif des Ostreidae.
~ 7 ~
journalières de la consommation, de l’ingestion, d
le taux d’absorption chez des huîtres plates d
quatre régimes monospécifiques, Isochrysis affinis galbana
Tetraselmis suecica et Skeletonema marinoï ont été opéré
européenne (Mollusque, Lamellibranche, ordre Filibranchia
Ostreidae) est un bivalve filtreur qui utilise des cils présents sur
branchies pour pomper les particules alimentaires présentes dans le milieu
c’est le processus d’ingestion. Après être parvenu jusqu’au
ain tri des particules s’opère. Une partie de ces particules, généralement agglutiné
forme les pseudofèces qui sont rejetés à l’extérieur par des
mouvements secs de fermeture des valves (Marteil, 1976). Les autres
buccal pour être soumis au processus de digestion
d’absorption. Les particules sont ensuite transformées
essentiels tels que les acides aminés, les acides gras et les sucres
non assimilés sont rejetés par l’intestin et forment les fèces
montre le circuit des particules alimentaires à travers le système
Figure
des particules alimentairesCrassostrea gigas
Bernard (1974)
ingestion, de l’efficacité
d’environ 18 mois
Isochrysis affinis galbana clone t-Iso,
été opérées.
Filibranchia, sous-ordre
es cils présents sur ses
branchies pour pomper les particules alimentaires présentes dans le milieu extérieur
parvenu jusqu’aux palpes labiaux,
généralement agglutinée
rejetés à l’extérieur par des
Les autres éléments sont
au processus de digestion ; c’est ce que
transformées en nutriments
, les acides gras et les sucres ; on parle alors
sont rejetés par l’intestin et forment les fèces
es à travers le système
Figure 2: Schéma d'ingestion des particules alimentaires chez Crassostrea gigas, d'après Bernard (1974).

� Algues
Les quatre microalgues test
sont : Isochrysis affinis galbana
cellulaire : 40-50 µm3, poids sec
Bacillariophycée (volume cellulaire
Tetraselmis suecica, Prasinophycée
poids sec : 226 pg) ; Skeletonem
et al., 2005), Bacillariophycée
sec : 51 pg) d’après des mesures effectuées à l’écloserie avant
l’expérience.
Protocoles expérimentaux
Les huîtres de 18 mois
été préalablement stockées en salle de
avec un mélange 50/50 en volume
″standard″ Crassostrea gigas
les huîtres dans les bacs, elles
pendant sept à dix jours à la nourriture de l’expérience et
avec une augmentation de 1°C par jour
I. Détermination du seuil de déclenchement des pseudo
Pour ne pas sous-estimer les mesures
durant notre expérience doit
et les pseudofèces chez Ostrea edulis
facilement différenciés et récoltés en conséquence
Au cours de cette expérience
des observations. S’il y a une
estimée à une concentration en entrée de
concentration en entrée diminué
quatre semaines, afin d’obtenir la concentration algale pour laquelle la production de
pseudofèces a pu être considéré
détaillées dans l’Annexe 2).
~ 8 ~
testées pour la nourriture des huîtres
Isochrysis affinis galbana, clone t-Iso, Haptophycée (volume
, poids sec : 19 pg) ; Chaetoceros gracilis,
(volume cellulaire : 80 µm3, poids sec : 69 pg);
phycée (volume cellulaire : 300 µm3,
Skeletonema marinoï (anc. S. costatum, Sarno
Bacillariophycée (volume cellulaire : 85 µm3, poids
d’après des mesures effectuées à l’écloserie avant
huîtres de 18 mois originaires de la baie de Cancale (Ile et Vilaine, Bretagne)
s en salle de quarantaine à une température de 10
en volume de t-Iso et de Chaetoceros gracilis suivant le pr
: ration quotidienne ajustée à 1600 µm3.µl-1
tres dans les bacs, elles ont été débarrassées de leurs épibiontes
à la nourriture de l’expérience et à la température
1°C par jour.
du seuil de déclenchement des pseudofèces
estimer les mesures d’ingestion, la production de pseudofèces
être proche de zéro, car il est très difficile de séparer les fèces
Ostrea edulis, à l’inverse de Crassostrea gigas, où ils
facilement différenciés et récoltés en conséquence.
expérience, les concentrations ont donc été ajusté
a une production de pseudofèces pour une quantité standard,
à une concentration en entrée de 100%, l’expérience a été poursuivie
diminuée de moitié, et cette production a été évaluée
obtenir la concentration algale pour laquelle la production de
être considérée comme négligeable (ces différentes concentrations sont
Figure culture d'algues en cylindre
station I
laine, Bretagne), ont
à une température de 10°C et nourries
suivant le protocole
1. Avant de répartir
débarrassées de leurs épibiontes et acclimatées
température voulue (19°C)
, la production de pseudofèces
est très difficile de séparer les fèces
où ils peuvent être
ajustées en fonction
pour une quantité standard,
a été poursuivie avec une
a été évaluée pendant
obtenir la concentration algale pour laquelle la production de
(ces différentes concentrations sont
Figure 3: Photographie de la salle de culture d'algues en cylindre de 300L à la
station Ifremer d’Argenton (29)

Les huîtres ont été placé
traitement) en flux ouvert à un débit de
nourries avec deux régimes monospécifiques
L’apport journalier « standard
(Haure, 1999), qui a été établi
par jour, ce qui correspond à
(l’équivalence t-Iso, étant une équivalence
diminuée de moitié (soit 2,7.10
car les premières observations
indiquée dans la littérature. Le choix de deux microalgues différentes
qu’une même ration en volume
microalgues n’ayant pas le même volume cellulaire.
La ration alimentaire journalière
pseudofèces, a alors été
concentration « standard ».
II. Détermination du meilleur régime algal pour
écophysiologique
Trente huîtres ont été
traitement) en flux ouvert à un débit de
nourries avec quatre régimes monospécifiques différent
suecica ; S. marinoï) avec une ration alimentaire de
comme définie précédemment
différents bacs, c'est-à-dire qu’il n’y
Annexe 7) tant pour le poids frais
la longueur (longueur de la coquille moyenne d’un géniteur
Mesures écophysiologiques
Pour chaque expérience
la consommation algale des géniteurs et
inorganique particulaire contenu
~ 9 ~
placées dans des bacs de 50 litres (triplicats pour chaque
à un débit de 12 litres par heure, avec 15
s avec deux régimes monospécifiques : t-Iso et C. gracilis (Annexe 2)
standard » a été défini en fonction des données de la littérature
établi autour de 2 à 4.109 cellules de Skeletonema
par jour, ce qui correspond à un équivalent de 4 à 5.109 cellules de t-Iso par
Iso, étant une équivalence en volume (µm3)). Cette concentration
(soit 2,7.109 cellules de t-Iso par huître par jour) dès les premiers jours
les premières observations ont montré une consommation moins importante que celle
Le choix de deux microalgues différentes a permis
volume donne la même production de pseudofèces pour deux
microalgues n’ayant pas le même volume cellulaire.
ration alimentaire journalière choisie, pour laquelle il n’y a pas eu
utilisée dans l’expérience suivante comme la nouvelle
Détermination du meilleur régime algal pour Ostrea edulis
été placées dans des bacs de 50 litres (triplicats pour
à un débit de 12 litres par heure (Annexes 3).
régimes monospécifiques différents (I. aff. galbana
) avec une ration alimentaire de 2.109 cellule de t-
précédemment. Les huîtres ont été réparties de façon homogène
dire qu’il n’y a pas de différence inter-bac (p<0,05
pour le poids frais (poids frais moyen d’un géniteur : 25,13
de la coquille moyenne d’un géniteur : 5,39 ± 0,05 cm)
expérience, deux séries de mesures ont été effectuées,
des géniteurs et une mesure de la fraction de matière organique et
ulaire contenue dans les fèces. Une troisième série de m
(triplicats pour chaque
huîtres par bac et
(Annexe 2).
en fonction des données de la littérature
Skeletonema marinoï par huître
Iso par huître par jour
Cette concentration a été
dès les premiers jours
n moins importante que celle
permis de confirmer
la même production de pseudofèces pour deux
a pas eu production de
comme la nouvelle
Ostrea edulis du point de vue
(triplicats pour chaque
heure (Annexes 3). Elles ont été
galbana; C. gracilis ; T.
-Iso.géniteur-1.jour-1
s de façon homogène dans les
p<0,05 ; Tableau 5,
25,13 ± 8,74 g) que pour
cm).
effectuées, une mesure de
mesure de la fraction de matière organique et
dans les fèces. Une troisième série de mesures des poids

secs et des longueurs de coquille
fin de la deuxième expérience
Les mesures de la concentration algale
bacs de géniteurs ont été
Multisizer) deux fois par jour
consommation des géniteurs.
Pour la première expérience, les
servis à la détermination de
particulaire (MIP). Les biodépôts
brûlés à 450°C pendant 4 heures
balance de précision ± 0,01mg
appliqué mais sans distinction entre fèces et
étant égale à [(MS-MC)/MS], avec MS = Masse Sèche et MC = Masse C
Pour les mesures du poids sec
la fin) et de la longueur de coquille
préalablement conservés au
lyophilisation (Model CHRIST ALPHA 1
l’obtention d’un poids sec constant des échantillons.
Toutes ces mesures ont
d’ingestion, l’efficacité d’absorption
formules qui suivent. Les individus au cours de cette expérience
sains, au vue de la faible mortalité (onze huîtres
bac.
Calculs et statistiques
Le Taux de filtration en flux ouvert est dé
�� � �������� �� �, en litre.
Le Taux de consommation
cellule algale.g poids sec chair
(Beiras et al., 1994).
~ 10 ~
de coquille des huîtres ont été réalisée sur 30 individus au début et à la
expérience.
concentration algale de l’eau à l’entrée (C1) et à la sortie
opérées quotidiennement au compteur de
deux fois par jour (matin et soir), pour estimer le taux
ère expérience, les fèces et pseudofèces, produit
à la détermination de leur fraction de matière organique (MOP)
. Les biodépôts ont été séchés par étuvage à 60°C pendant 24 heures
à 450°C pendant 4 heures et pesés deux fois (avant et après combustion)
1mg. Pour la deuxième expérience, le même
appliqué mais sans distinction entre fèces et pseudofèces. La fraction de matière organique
, avec MS = Masse Sèche et MC = Masse Cendres.
poids sec (30 individus au début et 10 individus par traitement à
coquille (tous les individus de l’expérience), le
préalablement conservés au congélateur pendant 48 heures et
lyophilisation (Model CHRIST ALPHA 1-2) pendant 72 heures minimum à
l’obtention d’un poids sec constant des échantillons.
ont permis de calculer le taux de filtration, de
efficacité d’absorption, et le taux d’absorption d’Ostrea edulis
Les individus au cours de cette expérience ont été
de la faible mortalité (onze huîtres sur un total de 360 huîtres),
en flux ouvert est défini par la formule suivante
.g poids sec chair huître-1
.heure-1
(Beiras et al
Taux de consommation lui, est égal à : � � ��1 � �2 �� �g poids sec chair huître-1.heure-1 et en µm3.g poids sec de chair
sur 30 individus au début et à la
et à la sortie (C2) des
de particules (type
le taux journalier de
produits durant 24h, ont
(MOP) et inorganique
par étuvage à 60°C pendant 24 heures, puis
(avant et après combustion) sur une
le même protocole a été
La fraction de matière organique
endres.
et 10 individus par traitement à
es individus ont été
congélateur pendant 48 heures et déshydratés par
2) pendant 72 heures minimum à -60°C, jusqu’à
filtration, de consommation,
Ostrea edulis, grâce aux
considérés comme
sur un total de 360 huîtres), soit 10% par
fini par la formule suivante :
et al., 1994).
�� �, en nombre de
g poids sec de chair huître-1.heure-1

Si il y a production de
gramme de cellule algale sec.g
PF avec C en gramme de cellule
au poids sec des pseudofèces en g
1.heure-1). A l’inverse, si on considère que la production de pseudofèces est proche de zéro,
alors l’Ingestion est égale au taux de consommation en g
huître-1.heure-1.
L’Efficacité d’absorption
E correspondant à la fraction
respectivement (Conover, 1966)
Le Taux d’absorption est défini
huître-1
.heure-1
(Beiras et al., 1994).
L’efficacité d’absorption
les jours de prélèvements des fèces,
ces valeurs il a fallu préalablement vérifier qu’il n’y a
(p<0,05 ; Tableau 2, Annexe 4) entre les consommations et les ingestions obtenues lors de
ces cinq jours et celles obtenues lors des cinq
Analyse statistique : P
facteur (ANOVA), couplée à un test de comparaison des moyennes (Scheffé) a été réalisé
afin de comparer les proportions de pseudofèces dans les biodépôts à différentes
concentrations de t-Iso et C. gracilis
variances (test de Levene) ont
Pour la deuxième expérience,
pour tester l’effet du type d’algue sur la consommation et l’
une transformation préalable des données
Les données du taux de filtration, d’
analyse non paramétrique de type
règle d’homogénéité des variances
Enfin, pour la biométrie
comparer la croissance entre le début et la fin de l’expérience
Tous les tests statistiques
~ 11 ~
Si il y a production de pseudofèces (expérience préliminaire) alors l’
sec.g poids sec chair huître-1.heure-1 est calculée
ramme de cellule algale sec.g poids sec chair huître-1.heure
des pseudofèces en gramme poids sec de pseudofèces.g poids sec chair huître
A l’inverse, si on considère que la production de pseudofèces est proche de zéro,
est égale au taux de consommation en g poids sec algue
absorption, est définie comme suit : �� �% � � ��������
fraction de matière organique, en poids sec, des algue
1966).
est définie par � � � ��, en g poids sec algue
., 1994).
efficacité d’absorption et le taux d’absorption ont été estimées
les jours de prélèvements des fèces, soit cinq observations pour chaque
blement vérifier qu’il n’y a pas de différences significatives
, Annexe 4) entre les consommations et les ingestions obtenues lors de
rs et celles obtenues lors des cinq semaines de l’expérience.
Pour l’expérience préliminaire, une analyse de
facteur (ANOVA), couplée à un test de comparaison des moyennes (Scheffé) a été réalisé
proportions de pseudofèces dans les biodépôts à différentes
C. gracilis. La normalité des variables et l
ont été préalablement vérifiées avant l’analyse.
Pour la deuxième expérience, une ANOVA et un test de Scheffé
l’effet du type d’algue sur la consommation et l’efficacité d’absorption
transformation préalable des données ������ √ "#�$$% pour ea en %.
Les données du taux de filtration, d’ingestion et d’absorption ont été
de type Kruskal-Wallis, car ces variables ne respectaient pas
règle d’homogénéité des variances.
biométrie, une analyse de variance à deux facteurs a
le début et la fin de l’expérience pour chaque
statistiques ont été réalisés sur le logiciel STATISTICA, version 6.0.
pseudofèces (expérience préliminaire) alors l’Ingestion en
e comme suit : I = C-
.heure-1 (PF correspond
.g poids sec chair huître-
A l’inverse, si on considère que la production de pseudofèces est proche de zéro,
poids sec algue.g poids sec chair
��& 100 avec F et
algues et des fèces
algue.g poids sec chair
s uniquement pour
algue. Pour utiliser
de différences significatives
, Annexe 4) entre les consommations et les ingestions obtenues lors de
une analyse de la variance à un
facteur (ANOVA), couplée à un test de comparaison des moyennes (Scheffé) a été réalisée
proportions de pseudofèces dans les biodépôts à différentes
et l’homogénéité des
.
VA et un test de Scheffé ont été appliqués
cité d’absorption, après
été traitées par une
e respectaient pas la
a été effectuée pour
pour chaque régime.
réalisés sur le logiciel STATISTICA, version 6.0.

Résultats
Ajustement de l’apport nutritionnel journalier
La « proportion de pseudofèces dans les
gracilis) en fonction de l’apport
± erreur-type).
Pour C. gracilis, seule la valeur de
Tableau 3, Annexe 5) de pseudofèces (30%), par rapport à l’apport standard. A 75%, aucune
différence significative avec la
production de pseudofèces est proche de 15% du total des biodépôts
Lorsque les géniteurs
significativement plus forte (70% des biodépôts) lorsque la ration est de 125%, que les
de production pour une concentration standard (
De ce fait, les rations alimentaires
la ration alimentaire standard
huître par jour), pour s’assurer d’
possible, c'est-à-dire proche de 10%. Bien que
pseudofèces entre 75% et 100% ne soit pas significative
observations supplémentaires
et Tetraselmis suecica, nous ont
adaptée à tous les régimes.
Figure 4: Production de pseudofèces (moyenne géniteur et par jour : A. Production de pseudofèces pour le régime valeurs assignées des mêmes lettres ne sont pas statistiquement différentes (p<0,05ordonnées est différente pour les graphiques A et B.
0
10
20
30
40
75 100
Prp
op
rtio
ns
de
pse
ud
ofè
ces
dan
s le
s b
iod
ép
ôts
(%
)
Apport nutritionnel (%)
A : Chaetoceros gracilis
a
a
~ 12 ~
Ajustement de l’apport nutritionnel journalier
roportion de pseudofèces dans les biodépôts » par espèce d’
’apport nutritionnel journalier est représentée en
seule la valeur de 125% induit une production significative (p<0,05
5) de pseudofèces (30%), par rapport à l’apport standard. A 75%, aucune
différence significative avec la concentration standard n’est révélée et dans ce ca
production de pseudofèces est proche de 15% du total des biodépôts (Figure 4)
Lorsque les géniteurs sont alimentés avec t-iso, la production de pseudofèces est
significativement plus forte (70% des biodépôts) lorsque la ration est de 125%, que les
ur une concentration standard (p<0,05 ; Tableau 3, Annexe 5
fait, les rations alimentaires pour toutes les algues ont été stabilisées
la ration alimentaire standard (rappel 100% correspondait à 2,7.109 cellules de t
pour s’assurer d’une production de pseudofèces la plus négligeable
dire proche de 10%. Bien que cette différence de production
entre 75% et 100% ne soit pas significative en présence de
s opérées à ces deux concentrations pour Skeletonema marinoï
ont conduits à retenir la concentration de 75%
Production de pseudofèces (moyenne + erreur-type) dans les biodépôts mesurés pour différents apport: A. Production de pseudofèces pour le régime C. gracilis ; B. Production de pseudofèces pour le régime t
valeurs assignées des mêmes lettres ne sont pas statistiquement différentes (p<0,05 ; Tableau 3, Annexe 5). Npour les graphiques A et B.
125
Apport nutritionnel (%)
Chaetoceros gracilis
b
0
20
40
60
80
100
100
Pro
po
rtio
ns
de
pse
ud
ofè
ces
dan
s le
s b
iod
ép
ôts
(%
)
Apport nutritionnel (%)
B : Isochrysis affinis galbana
a
par espèce d’algue (t-Iso et C.
Figure 4 (moyenne
une production significative (p<0,05 ;
5) de pseudofèces (30%), par rapport à l’apport standard. A 75%, aucune
et dans ce cas, la
(Figure 4).
a production de pseudofèces est
significativement plus forte (70% des biodépôts) lorsque la ration est de 125%, que les 20%
, Annexe 5).
stabilisées à 75% de
cellules de t-Iso par
pseudofèces la plus négligeable
de production de
en présence de C. gracilis, des
Skeletonema marinoï
75% comme la mieux
type) dans les biodépôts mesurés pour différents apports nutritionnels par ; B. Production de pseudofèces pour le régime t-Iso. Les
, Annexe 5). Notez que l’axe des
125
Apport nutritionnel (%)
Isochrysis affinis galbana
b

Ecophysiologie des géniteurs d’Ostrea edulis
Taux de filtration
Les valeurs du taux de filtration
significative (p<0,05 ; Tableau 4
marinoï et C. gracilis avec des
1.h-1 respectivement) à ceux du
0,01 l.g-1.h-1 et 0,46 ± 0,01 l.g-
Figure 5: Taux de filtration (moyenne
valeurs assignées des mêmes lettres ne sont pas statistiquement différentes
Taux de consommation
La consommation journalière du groupe d
que celle de T. suecica et de t
1) pour S. marinoï et C. gracilis
t-Iso, d’autre part (Figure 6).
observée entre les deux diatomées et
diatomée.
0
0.2
0.4
0.6
0.8
1
C. gracilis
Tau
x d
e f
iltra
tio
n (
l.g-1
.h-1
)
0
2E+09
4E+09
6E+09
8E+09
C. gracilisTau
x d
e c
on
som
mat
oin
(µ
m3
.g-1
.h-1
)
Figure 6: Taux de consommation (moyenneLes valeurs assignées des mêmes lettres ne sont pas statistiquement différentes (p<0,05
~ 13 ~
Ecophysiologie des géniteurs d’Ostrea edulis
Les valeurs du taux de filtration rapportées en Figure 5 montrent
; Tableau 4, Annexe 6) entre deux groupes, celui
avec des taux deux fois supérieurs (0,78 ± 0,00 l.g-1.h
du groupe de flagellés composé de T. suecica
-1.h-1, respectivement).
oyenne ± erreur-type) O. edulis en fonction des régimes nutritionnels mêmes lettres ne sont pas statistiquement différentes (p<0,05 ; Tableau 4, Annexe 6
onsommation
a consommation journalière du groupe de diatomées est deux fois plus importante
t-Iso, avec des valeurs moyennes de 6,59 et 6,07 (
gracilis d’une part, et 3,41 et 3,54 (109 µm3.g-1.h-1
Une différence significative (p<0,05 ; Tableau
entre les deux diatomées et pour t-Iso et T. suecica par rapport à chaque
C. gracilis S. marinoï T. suecica t-Iso
Régime nutritionnel
aa
b b
C. gracilis S. marinoï T. suecica t-Iso
Régime nutritionnel
a b
cc
oyenne ± erreur-type) d’O. edulis (µm3.g
-1.h
-1) en fonction des régimes nutritionnels
Les valeurs assignées des mêmes lettres ne sont pas statistiquement différentes (p<0,05 ; Tableau 4, Annexe 6).
montrent une différence
elui des diatomées S.
.h-1 et 0,78 ± 0,00 l.g-
. suecica et de t-Iso (0,46 ±
nutritionnels (l.g-1
.h-1
). Les , Annexe 6).
deux fois plus importante
et 6,07 (109 µm3.g-1.h-
1) pour T. suecica et
Tableau 4, Annexe 6) est
par rapport à chaque
des régimes nutritionnels., Annexe 6).

Taux d’ingestion
C. gracilis est le régime nutriti
mg, suivi de S. marinoï (3,97
l’algue la plus faiblement ingérée (1,48
des taux d’ingestion significativement différents
Efficacité d’absorption
La meilleure efficacité d’absorption
(± 12,09), suivie de C. gracilis
différences non significatives (p<0,05
pas une moindre efficacité d’absorption
différente des trois régimes précédent
Figure 7: Taux d’ingestion (moyenne nutritionnels. Les valeurs assignées des mêmes lettres ne sont pas statistiquement différentes (p<0,05
Figure 8: Efficacité d’absorption (moyenneLes valeurs assignées des mêmes lettres ne sont pas statistiquement différentes (p<0,05
0
20
40
60
C. gracilis
Effi
caci
té d
'ab
sorp
tio
n (
%)
a
0
2
4
6
8
C. gracilis
Tau
x d
'inge
stio
n (
mg.
g-1.h
-1)
a
~ 14 ~
est le régime nutritionnel le mieux ingéré avec une valeur de
(3,97 ± 0,68 mg), de T. suecica (2,57 ± 0,08 mg) et
l’algue la plus faiblement ingérée (1,48 ± 0,05 mg : Figure 7) ; toutes ces algues
des taux d’ingestion significativement différents (p<0,05 ; Tableau 4, Annexe 6
Efficacité d’absorption
efficacité d’absorption (%) est relevée pour S. marinoï avec une valeur de
C. gracilis avec 38,76 (± 6,54) et de T. suecica 35,94
différences non significatives (p<0,05 ; Tableau 4, Annexe 6). A l’inverse, t
efficacité d’absorption avec une valeur de 1,46 (± 3,72
précédents (Figure 8).
± erreur-type) d’O. edulis exprimé en mg.g-1
.h-1
en fonction Les valeurs assignées des mêmes lettres ne sont pas statistiquement différentes (p<0,05
oyenne + erreur-type) d’O. edulis exprimé en % en fonction des régimes nutritionnelsLes valeurs assignées des mêmes lettres ne sont pas statistiquement différentes (p<0,05 ; Tableau 4, Annexe 6).
C. gracilis S. marinoï T. suecica t-Iso
Régime nutritionel
a
a
b
C. gracilis S. marinoï T. suecica t-Iso
Régime nutritionnel
b
c
d
onnel le mieux ingéré avec une valeur de 5,29 ± 0,70
mg) et de t-Iso qui est
toutes ces algues conduisent à
, Annexe 6).
avec une valeur de 48,50
94 (± 13,97), avec des
’inverse, t-Iso se caractérise
3,72), statistiquement
en fonction des régimes ; Tableau 4, Annexe 6).
des régimes nutritionnels., Annexe 6).
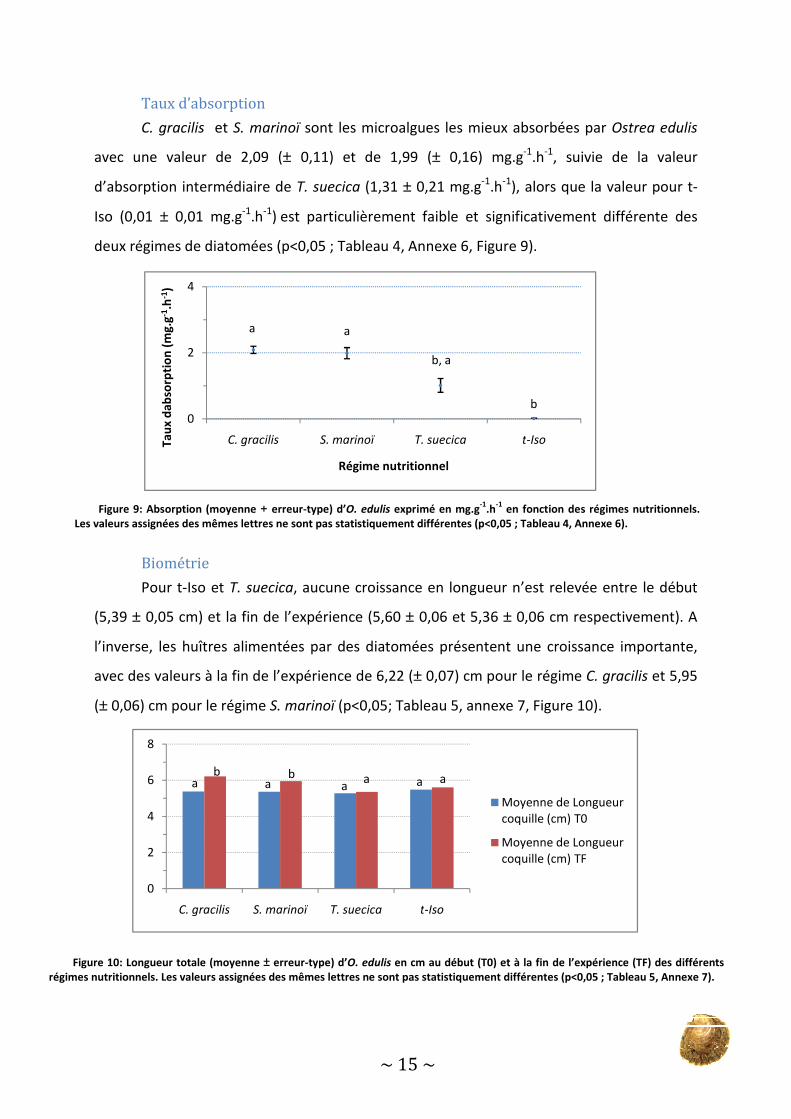
Taux d’absorption
C. gracilis et S. marinoï
avec une valeur de 2,09 (±
d’absorption intermédiaire de
Iso (0,01 ± 0,01 mg.g-1.h-1) est particulièrement
deux régimes de diatomées (p<0,0
Biométrie
Pour t-Iso et T. suecica
(5,39 ± 0,05 cm) et la fin de l’expérience (5,60
l’inverse, les huîtres alimentées par des diatomées présentent
avec des valeurs à la fin de l’expérience de 6,22 (
(± 0,06) cm pour le régime S. marinoï
Figure 9: Absorption (moyenne + erreurLes valeurs assignées des mêmes lettres ne sont pas statistiquement différentes (p<0,05
0
2
4
6
8
C. gracilis S. marinoï
a ab
Figure 10: Longueur totale (moyenne ± erreurrégimes nutritionnels. Les valeurs assignées des mêmes lettres ne sont pas statistiquement différentes (p<0
0
2
4
C. gracilisTau
x d
abso
rpti
on
(m
g.g-1
.h-1
)
a
~ 15 ~
S. marinoï sont les microalgues les mieux absorbée
± 0,11) et de 1,99 (± 0,16) mg.g-1.h-1, suivie
d’absorption intermédiaire de T. suecica (1,31 ± 0,21 mg.g-1.h-1), alors que la valeur pour
est particulièrement faible et significativement différente
(p<0,05 ; Tableau 4, Annexe 6, Figure 9).
T. suecica, aucune croissance en longueur n’est relevée
cm) et la fin de l’expérience (5,60 ± 0,06 et 5,36 ± 0,06 cm respectivement)
entées par des diatomées présentent une croissance importante,
avec des valeurs à la fin de l’expérience de 6,22 (± 0,07) cm pour le régime
S. marinoï (p<0,05; Tableau 5, annexe 7, Figure
erreur-type) d’O. edulis exprimé en mg.g-1
.h-1
en fonction des régimes nutritionnelsLes valeurs assignées des mêmes lettres ne sont pas statistiquement différentes (p<0,05 ; Tableau 4, Annexe 6).
S. marinoï T. suecica t-Iso
Moyenne de Longueur
coquille (cm) T0
Moyenne de Longueur
coquille (cm) TF
aa a ab
erreur-type) d’O. edulis en cm au début (T0) et à la fin de l’expérience (TF) des différents régimes nutritionnels. Les valeurs assignées des mêmes lettres ne sont pas statistiquement différentes (p<0
S. marinoï T. suecica t-Iso
Régime nutritionnel
a
b
b, a
mieux absorbées par Ostrea edulis
suivie de la valeur
, alors que la valeur pour t-
et significativement différente des
n’est relevée entre le début
cm respectivement). A
une croissance importante,
) cm pour le régime C. gracilis et 5,95
Figure 10).
des régimes nutritionnels., Annexe 6).
Moyenne de Longueur
coquille (cm) T0
Moyenne de Longueur
coquille (cm) TF
au début (T0) et à la fin de l’expérience (TF) des différents régimes nutritionnels. Les valeurs assignées des mêmes lettres ne sont pas statistiquement différentes (p<0,05 ; Tableau 5, Annexe 7).

A l’issue de l’expérience
peu marquées par rapport au poids d’origine (0,48
lorsqu’elles sont alimentées avec t
(0,62 ± 0,04 g). Les huîtres en présence de
en fin d’expérience (0,33 ±
statistiquement différentes des valeurs finales des autres régimes
Annexe 7, Figure 11).
Figure 11 : Poids sec de chair (moyenne l'expérience pour chaque régime nutritionneldifférentes (p<0,05 ; Tableau 5, Annexe 7).
L’ensemble de ces résultats
écophysiologiques étudiés pour chaque régime nutritionnel
obtenues avec les algues C. gracilis
régime T. suecica pour une majorité de v
de t-Iso ressort très nettement
Régimes
nutritionnels
Taux de
filtration
Taux de
consommation
C. gracilis + +
S. marinoï + +
T. suecica - -
t-Iso - -
Tableau 1: Classification de type ordinaleforte valeur , « - »montre la plus faible valeurvariable.
0
0.2
0.4
0.6
0.8
Témoins
Po
ids
sec
de
ch
air
(g) a
~ 16 ~
l’issue de l’expérience, les mesures de poids sec de chair présentent une évolution
par rapport au poids d’origine (0,48 ± 0,03 g) et statistiquement
alimentées avec t-Iso (0,58 ± 0,05 g), C. gracilis (0,56 ±
en présence de T. suecica présentent une décroissance
± 0,03 g) non significative avec les valeurs d’origine mais
statistiquement différentes des valeurs finales des autres régimes (p<0,05
oyenne ± erreur-type) des géniteurs d'O. edulis (g) au début (Témoins) et à la fin de régime nutritionnel. Les valeurs assignées des mêmes lettres ne sont pas statistiquement
nnexe 7).
de ces résultats (Tableau 1) rapporte les différences entre
pour chaque régime nutritionnel. Les meilleures réponses sont
C. gracilis et S. marinoï. De faibles valeurs sont observées pour le
pour une majorité de variables écophysiologiques, tandis que la
ressort très nettement.
Taux de
consommation
Taux
d’ingestion
Efficacité
d’absorption Absorption
++ + +
+ + +
- + + -
- - - -
lassification de type ordinale des différentes variables écophysiologiques étudiées »montre la plus faible valeur dans la série considérée, et « 0 » ne montre pas d’effet du régime sur la
Témoins C. gracilis S. marinoï T. suecica t-Iso
Régime nutritionnel
c, a
b, ab, a
b, a
, les mesures de poids sec de chair présentent une évolution
statistiquement identique
0,06 g), S. marinoï
une décroissance pondérale
non significative avec les valeurs d’origine mais
(p<0,05 ; Tableau 5,
au début (Témoins) et à la fin de mêmes lettres ne sont pas statistiquement
différences entre les paramètres
Les meilleures réponses sont
. De faibles valeurs sont observées pour le
, tandis que la médiocrité
Croissance
linéaire
Croissance
pondérale
+ 0
+ 0
0 -
0 0
bles écophysiologiques étudiées « + »montre la plus » ne montre pas d’effet du régime sur la

Discussion
L’influence de la qualité de la ration alimen
sur le conditionnement des géniteurs d’
semble nécessaire afin d’expliciter son
Taux de filtration, taux de consommation
Deux groupes se distinguent
de cette expérience. Les géniteurs nourris avec
deux fois plus fortes que celles enregistrées chez les
suecica. Le volume des cellules algales
différence entre ces deux groupes. En effet, les diatom
cellulaire équivalent (80-85 µm
celles de T. suecica, plus grandes
l’huître capte rapidement le volume
une plus grande quantité d’eau. Par contre, la faible taille de t
espèces étudiées) peut interférer
réponse purement mécanique
pas d‘indice de sélection qualitative des particules au niveau des branchies
sélection quantitative des particules
Le regroupement entre algue
mesures d’ingestion, où les quatre régimes
meilleurs taux d’ingestion concernent
discussion antérieure.
Efficacité d’absorption, absorption
La tendance précédente
taux d’absorption, et donc les diatomées se révèlent comme l
La forte valeur de T. suecica pour l’assimilation est
En effet cette Prasinophycée est composée
diatomées), les huîtres assimilent
via cette algue, mais peu d’acides gras essentiels, car cette dernière
~ 17 ~
L’influence de la qualité de la ration alimentaire (différents types
conditionnement des géniteurs d’Ostrea edulis d’un point de vue écophysiologique
afin d’expliciter son rôle potentiel sur la fécondité.
, taux de consommation, taux d’ingestion
se distinguent pour le taux de filtration et de consommatio
es géniteurs nourris avec C. gracilis et S. marinoï montr
que celles enregistrées chez les géniteurs nourris avec t
cellules algales paraît la meilleure explication pour
différence entre ces deux groupes. En effet, les diatomées ont toutes les
µm3) alors que les cellules de t-Iso sont plus petites (45 µm
grandes (300 µm3). Pour cette dernière il est
le volume d’algue nécessaire et ne nécessite donc pas
d’eau. Par contre, la faible taille de t-Iso (par rapp
peut interférer sur sa capacité de rétention par les branchies.
réponse purement mécanique a été déjà montrée par Barillé (1994), c'est
indice de sélection qualitative des particules au niveau des branchies
des particules s’exerce.
re algues observé précédemment, n’est pas
où les quatre régimes ont une influence différencié
taux d’ingestion concernent les deux diatomées, ce qui reste dans la logique de la
bsorption
précédente se confirme à nouveau pour l’efficacité d’absorption
es diatomées se révèlent comme les algues les
pour l’assimilation est surement liée à la nature de cette
phycée est composée à 88% de matière organique (
assimilent donc surement de grande quantité de matière organique
cette algue, mais peu d’acides gras essentiels, car cette dernière n’en contient
s d’apport d’algue)
d’un point de vue écophysiologique
pour le taux de filtration et de consommation au cours
montrent des valeurs
géniteurs nourris avec t-Iso et T.
xplication pour comprendre la
toutes les deux un volume
plus petites (45 µm3) et
donc possible que
et ne nécessite donc pas de filtrer
par rapport aux autres
es branchies. Cette
, c'est-à-dire qu’il n’y a
indice de sélection qualitative des particules au niveau des branchies et qu’une seule
pas confirmé pour les
ont une influence différenciée. Toutefois, les
deux diatomées, ce qui reste dans la logique de la
efficacité d’absorption et le
les plus bénéfiques.
à la nature de cette algue.
à 88% de matière organique (vs 77% pour les
rement de grande quantité de matière organique
en contient quasiment

pas (Enright et al., 1986). Cette relativement forte
d’être mieux absorbée que t-Iso,
Le taux d’absorption quasiment nulle de t
avec sa faible efficacité d’absorption
Les valeurs d’efficacité d’absorption
valeurs rapportées pour Crassostrea gigas
cette espèce, pendant la même période. Une
a été déterminée chez Crassost
(Boglino, 2008, com. pers) ce qui est nettement plus fort que la valeur
pour Ostrea edulis. Les données d’
également très différentes puisque des valeurs de 5% (Boglino, 2008, com. pers) sont
rapportées chez cette espèce (
paraissent tout à fait surprenant
Crassostrea gigas (70-80%, Bourles
Ces différences d’efficacité d’absorption
fortement que ces deux espèces pourtant proches possèdent
enzymatique différents, et qu’elles n’utili
deux espèces d’huîtres apprécient les diatomées, l’une rejette
et l’autre t-Iso (Ostrea edulis).
Biométrie
Les biométries révèlent
pour les diatomées. Les géniteurs
absorbé pour assurer une croissance pondérale
qu’absorbée, la médiocre qualité nutritive d
l’absence d’accroissement pondéral
se traduit donc pas par un bénéfice pour l’animal mais pourra
son activité basale (comme la respiration).
Autres espèces
Le lien entre le conditionnement
d’autres espèces, comme chez
~ 18 ~
ette relativement forte efficacité d’absorption permet à
Iso, pourtant ingérée dans les mêmes proportions.
quasiment nulle de t-Iso, pour Ostrea edulis, est à mettre en relation
efficacité d’absorption.
efficacité d’absorption pour O. edulis sont cependant à comparer avec les
Crassostrea gigas, à travers des travaux du même type conduits sur
, pendant la même période. Une efficacité d’absorption de 67% pour
Crassostrea gigas dans des conditions expérimentales similaires
ce qui est nettement plus fort que la valeur
données d’efficacité d’absorption de T. suecica
différentes puisque des valeurs de 5% (Boglino, 2008, com. pers) sont
s chez cette espèce (vs 38% chez O. edulis). De même, les résultats pour t
tout à fait surprenants par rapport aux fortes valeurs enregistrées chez
80%, Bourles, 2008 com. pers).
efficacité d’absorption observées entre O. edulis et C. gigas
spèces pourtant proches possèdent des systèmes de digestion
fférents, et qu’elles n’utilisent donc pas les algues de la même manière.
deux espèces d’huîtres apprécient les diatomées, l’une rejette Tetraselmis suecica
).
Les biométries révèlent également une meilleure croissance linéaire
géniteurs nourris avec le régime t-Iso utilisent le
croissance pondérale, dont peut être la reproduction
qualité nutritive de T. suecica pour la croissance
pondérale des géniteurs qui en sont nourris. Cette
pas par un bénéfice pour l’animal mais pourrait être utilisé pour maintenir
me la respiration).
Le lien entre le conditionnement des géniteurs et la nourriture a déjà été étudié chez
hez C. gigas, où Chàvez (2001) montre une relation
permet à T. suecica
dans les mêmes proportions.
est à mettre en relation
endant à comparer avec les
des travaux du même type conduits sur
de 67% pour S. marinoï
dans des conditions expérimentales similaires
ce qui est nettement plus fort que la valeur (50%) déterminée
chez C. gigas sont
différentes puisque des valeurs de 5% (Boglino, 2008, com. pers) sont
De même, les résultats pour t-Iso
aux fortes valeurs enregistrées chez
C. gigas suggèrent
des systèmes de digestion
ent donc pas les algues de la même manière. Si les
Tetraselmis suecica (C. gigas)
croissance linéaire et pondérale
ent le peu de nourriture
peut être la reproduction. Bien
pour la croissance est montrée par
Cette microalgue ne
être utilisé pour maintenir
déjà été étudié chez
e relation entre le

conditionnement et la gamétogénèse (production d’ovocytes) mais pas entre le
conditionnement et le rendement larvaire.
plasticité en réponse aux variations de nourritu
d’être facilement cultivée en écloserie
de nourrir les géniteurs, car l’accumulation des réserves pendant l’hiver servent plus que le
conditionnement trophique pour l’élaboration de la gamétogénèse
qualité (Cannuel et al., 2005).
Pour Ostrea chilensis, autre espèce incubatrice
meilleur régime par rapport à
même auteur montre aussi une
et sur le développement des larves. Il faut
larves incubées qui sont capable
manteau de leur mère, ce qui est
Chez Pecten maximus, la gamétogenèse dépend du régime
pas le développement larvaire ultérieur
Emissions larvaires
En fin d’expérience, des larves
premières émissions (1 million
géniteurs nourris avec t-Iso montre
émises, ce qui est vraiment surprenant au vue de la mauvaise
durant toute l’expérience. Il est probable que ces émissions aient été obtenues à partir des
réserves initiales mais l’intensité de la ponte est plus dif
cette même hypothèse d’intervention des réserves peut être avancée,
particulièrement faible (300 000).
(1997) qui montre que les géniteurs d’
performantes que celles émises par les huîtres ayant reçu
de plus, que les larves émises
ne sont pas plus performante
régime basé sur T. suecica ne sembl
nos résultats puisqu’une faible absorption
qualité des larves n’ayant pas
~ 19 ~
conditionnement et la gamétogénèse (production d’ovocytes) mais pas entre le
conditionnement et le rendement larvaire. Comme cette espèce possède une certaine
plasticité en réponse aux variations de nourriture (quantitatif et qualitatif)
en écloserie (Uriarte et al., 2004). De plus, il n’
de nourrir les géniteurs, car l’accumulation des réserves pendant l’hiver servent plus que le
pour l’élaboration de la gamétogénèse et des larves de bonne
autre espèce incubatrice, (Wilson et al., 1996) t-
par rapport à C. gracilis ce qui est complètement opposé à nos résultats. Ce
une influence de la nourriture des géniteurs sur la gamétogénèse
ppement des larves. Il faut toutefois porter attention à la nourriture des
capables de capter les particules en suspension
ce qui est confirmé par Chaparro (2006).
gamétogenèse dépend du régime nutritionnel
pas le développement larvaire ultérieur (Utting et al., 1998).
fin d’expérience, des larves ont été émises pour tous les régimes
1 million de larves) étant apparues chez S. marinoï
Iso montrent cependant la meilleure fécondité avec 1
, ce qui est vraiment surprenant au vue de la mauvaise absorption
Il est probable que ces émissions aient été obtenues à partir des
réserves initiales mais l’intensité de la ponte est plus difficile à expliquer.
cette même hypothèse d’intervention des réserves peut être avancée,
particulièrement faible (300 000). Cette hypothèse a déjà été formulée par
que les géniteurs d’O. edulis non nourris émettent des larves aussi
ises par les huîtres ayant reçu un supplément d’algue.
émises par des géniteurs d’O. edulis nourris avec Tetraselmis s
pas plus performantes que les larves émises par des géniteurs non nourris
ne semble pas favoriser le conditionnement, ce qui concorde avec
puisqu’une faible absorption de cette algue est démontrée ici
qualité des larves n’ayant pas été étudiée cette question reste encore entière.
conditionnement et la gamétogénèse (production d’ovocytes) mais pas entre le
possède une certaine
(quantitatif et qualitatif) cela lui permet
De plus, il n’est pas nécessaire
de nourrir les géniteurs, car l’accumulation des réserves pendant l’hiver servent plus que le
des larves de bonne
-Iso s’avère être le
opposé à nos résultats. Ce
influence de la nourriture des géniteurs sur la gamétogénèse
attention à la nourriture des
les particules en suspension à l’intérieur du
nutritionnel des géniteurs mais
émises pour tous les régimes nutritionnels, les
S. marinoï et C. gracilis. Les
avec 1 400 000 larves
absorption de cette algue
Il est probable que ces émissions aient été obtenues à partir des
ficile à expliquer. Pour T. suecica
cette même hypothèse d’intervention des réserves peut être avancée, la fécondité étant
par Berntsson et al.
nt des larves aussi
un supplément d’algue. Il souligne,
Tetraselmis suecica
par des géniteurs non nourris. Donc un
, ce qui concorde avec
est démontrée ici. Cependant, la
cette question reste encore entière.

Conclusion
Les meilleurs régimes
marinoï. Au vue de nos résultats
dans le régime des géniteurs d’
d’expliquer pourquoi cette algue est
fécondité enregistrée est importante.
Ces résultats soulignent aussi qu’
être conditionnées de la même façon en écloseries.
Pour une meilleure interprétation de ces premiers résultats, des analyses
biochimiques, tant des géniteurs que des larves
potentiel transfert des éléments biochimiques contenus dans les algues jusqu’aux géniteurs,
et des géniteurs aux larves.
régimes les mieux adaptés pour
~ 20 ~
algaux pour les géniteurs d’O. edulis sont donc
résultats, nous pouvons dire que T. suecica n’est
dans le régime des géniteurs d’O. edulis. Pour t-Iso, un doute subsiste car
quoi cette algue est médiocre sur le plan écophysiologique alors que la
fécondité enregistrée est importante.
Ces résultats soulignent aussi qu’Ostrea edulis et Crassostrea gigas
être conditionnées de la même façon en écloseries.
une meilleure interprétation de ces premiers résultats, des analyses
biochimiques, tant des géniteurs que des larves émises, sont en cours pour évaluer un
potentiel transfert des éléments biochimiques contenus dans les algues jusqu’aux géniteurs,
Ceci servira de plus à établir plus spécifiquement le ou les
régimes les mieux adaptés pour le conditionnement des géniteurs de l’huître plate
donc C. gracilis et S.
pas recommandée
subsiste car il est très difficile
sur le plan écophysiologique alors que la
Crassostrea gigas ne doivent pas
une meilleure interprétation de ces premiers résultats, des analyses
émises, sont en cours pour évaluer un
potentiel transfert des éléments biochimiques contenus dans les algues jusqu’aux géniteurs,
à établir plus spécifiquement le ou les
onditionnement des géniteurs de l’huître plate.

Bibliographie
Barillé, L., 1994. Observations of feeding structures involved in preingestive mechanisms in the
Japanese oyster Crassostrea gigas
Beiras, R., Camacho, A.P., Albentosa, M., 1994.
performance of Ostrea edulis
system. Marine Biology, 119(2), 227
Beiras, R., Camacho, A.P., Albentosa, M., 1995.
budget of young oyster
Experimental Marine Biology and Ecology
Bernard, F.R., 1974. Particle sorting and labial palp function in
(Thunberg, 1795). Biological Bulletin
Berntsson, K.M., Jonsson, P.R., Wangberg, S.A., Carlsson, A.S.,
fatty acid composition, survival and growth rates in larvae of the
edulis. Aquaculture, 154(2), 139
Cannuel, R., Beninger, P.G., 2005.
oocyte quality predictors and validators in
35-43.
Chaparro, O.R., Navarrete, L.R.,
oyster Ostrea chilensis and the utilisation of biochemical energy reserves during development:
An extreme case of the brooding habit.
Chàvez Villalba, J.E., 2001. Condi
Conover, R.J., 1966. Assimilation of organic matter zooplankton.
338-345.
Enright, C.T., Newkirk, G.F., Craigie, J.S., Castell, J.D.,
juvenile Ostrea edulis L. Journal of Experimental Marine Biology and Ecology
FAO, 2006. Ecloserie de bivalves.
Ferreiro, M.J., Perez-Camacho, A., Labarta, U., Beiras, R., Planas, M., Fernandez
Changes in the biochemical
Marine Biology, 106(3), 395
Frolov, A.V., Pankov, S.L., 1992.
biochemical point-of-view.
Molecular Biology, 103(1), 161
Gonzales, R., 2007. Incidence de la nutrition pour la reproduction, l'
métamorphose de l'huître plate
~ 21 ~
, L., 1994. Observations of feeding structures involved in preingestive mechanisms in the
Crassostrea gigas. Haliotis, 23, 125-137.
Albentosa, M., 1994. Comparison of the scope for growth
edulis seed reared at different food concentrations in an open
, 119(2), 227-233.
Albentosa, M., 1995. Short-term and long-term alterations in the
budget of young oyster Ostrea edulis L. in response to temperature
Experimental Marine Biology and Ecology, 186(2), 221-236.
Bernard, F.R., 1974. Particle sorting and labial palp function in the Pacific oyster
Biological Bulletin, 146(1), 1-10.
, Jonsson, P.R., Wangberg, S.A., Carlsson, A.S., 1997. Effects of broodstock diets on
fatty acid composition, survival and growth rates in larvae of the European
, 154(2), 139-153.
Beninger, P.G., 2005. Is oyster broodstock feeding always necessary? A study using
oocyte quality predictors and validators in Crassostrea gigas. Aquatic Living Resources
, Thompson, R.J., 2006. The physiology of the larva of the Chilean
and the utilisation of biochemical energy reserves during development:
An extreme case of the brooding habit. Journal of Sea Research, 55(4), 292
vez Villalba, J.E., 2001. Conditionnement experimental de l'huître Crassostrea gigas
Conover, R.J., 1966. Assimilation of organic matter zooplankton. Limnology and Oceanography
Newkirk, G.F., Craigie, J.S., Castell, J.D., 1986. Evaluation of phyto
Journal of Experimental Marine Biology and Ecology
Ecloserie de bivalves. Un manuel pratique.
amacho, A., Labarta, U., Beiras, R., Planas, M., Fernandez
Changes in the biochemical-composition of Ostrea edulis larvae fed on diff
, 106(3), 395-401.
Pankov, S.L., 1992. The reproduction strategy of oyster ostrea
view. Comparative Biochemistry and Physiology B
, 103(1), 161-182.
Incidence de la nutrition pour la reproduction, l'éleva
tre plate Ostrea edulis. Mémoire Master 2.
, L., 1994. Observations of feeding structures involved in preingestive mechanisms in the
Comparison of the scope for growth with the growth-
erent food concentrations in an open-flow
term alterations in the energy
in response to temperature-change. Journal of
c oyster Crassostrea gigas
ects of broodstock diets on
European flat oyster, Ostrea
Is oyster broodstock feeding always necessary? A study using
Aquatic Living Resources, 18(1),
The physiology of the larva of the Chilean
and the utilisation of biochemical energy reserves during development:
, 55(4), 292-300.
Crassostrea gigas. Thèse.
Limnology and Oceanography, 11(3),
plankton as diets for
Journal of Experimental Marine Biology and Ecology, 96(1), 1-13.
amacho, A., Labarta, U., Beiras, R., Planas, M., Fernandez-Reiriz, M.J., 1990.
fferent food regimes.
The reproduction strategy of oyster ostrea-edulis l from the
Comparative Biochemistry and Physiology B-Biochemistry &
levage larvaire et la

Haure, J., 1999. Testage de souches d'huitres plates
élevage contrôlé, écophysiologie et mod
Helm, M.M., Holland, D.L., Utting, S.D., East, J.
larvae of the european flat oyster,
of the United Kingdom, 71(3), 691
Heras, H., Keanhowie, J., Ackman, R.G., 1994.
supplements for adult ostrea
Holland, D.L., Spencer, B.E., 1973.
during larval development, metamorphosis and early spat growth.
Biological Association of the United Kingdom
Jonsson, P.R., Berntsson, K.M., Andre, C., Wandgberg, S.A.,
European oyster (Ostrea edulis
composition. Marine Biology
Labarta, U., Fernandez-Reiriz, M.,
development, metamorphosis an
Biochemistry and Physiology a
Mann, R., 1979. Some biochemical and physiological aspects
Crassostrea gigas and ostre
Marine Biological Association of the United Kingdom
Marteil, L., 1976. [Shellfish culture in France. Part 2. Oyster and mussel biology].
Institut Peches Maritimes, Nantes, 40(2), 149
Millican, P.F., Helm, M.M., 1994.
Ostrea edulis. Aquaculture
Naciri-Graven, Y., Martin, A.G., Baud, J.P., Renault, T., Gerard, A.,
Ostrea edulis (L) for survival when infected with the parasite
Experimental Marine Biology and Ecology
Robert, R., Borel, M., Pichot, Y., Trut, G.,
edulis in the Bay of Arcachon (France).
Robert, R., Trintignac, P., 1996. Microalgae and larval feeding in mollusc hatchery.
Rodstrom, E.M., Jonsson, P.R., 2000.
function of temperature and salinity with implications for culture policies on the Swedish west
coast. Journal of Shellfish Research
Sarno, D., Kooistra, Whcf., Medlin, L.K.,
Skeletonema (Bacillariophyceae). II. An assessment of the taxonomy of
~ 22 ~
Haure, J., 1999. Testage de souches d'huitres plates Ostrea edulis tolérantes
cophysiologie et modélisation de la croissance. Thèse.
Helm, M.M., Holland, D.L., Utting, S.D., East, J., 1991. Fatty-acid composition of early nonfeeding
at oyster, Ostrea edulis. Journal of the Marine Biological A
, 71(3), 691-705.
Ackman, R.G., 1994. The potential use of lipid microspheres as nutritional
supplements for adult ostrea-edulis. Aquaculture, 123(3-4), 309-322.
Spencer, B.E., 1973. Biochemical changes in fed and starved oysters,
during larval development, metamorphosis and early spat growth. Journal of the Marine
Biological Association of the United Kingdom, 53(2), 287-298.
, Berntsson, K.M., Andre, C., Wandgberg, S.A., 1999. Larval growth and settlement of the
Ostrea edulis) as a function of food duality measured as fatty acid
Marine Biology, 134(3), 559-570.
Reiriz, M., Perez-Camacho, A., 1999. Dynamics of fatty acids in the larval
development, metamorphosis and post-metamorphosis of Ostrea edulis
Biochemistry and Physiology a-Molecular & Integrative Physiology, 123(3), 249
Mann, R., 1979. Some biochemical and physiological aspects of growth and gametogenesis in
and ostrea-edulis grown at sustained elevated-temperatures.
Marine Biological Association of the United Kingdom, 59(1), 95-110.
sh culture in France. Part 2. Oyster and mussel biology].
, Nantes, 40(2), 149-346.
Helm, M.M., 1994. Effects of nutrition on larvae production in the european
Aquaculture, 123(1-2), 83-94.
, Martin, A.G., Baud, J.P., Renault, T., Gerard, A., 1998. Selecting the
Ostrea edulis (L) for survival when infected with the parasite Bonamia ostreae
Experimental Marine Biology and Ecology, 224(1), 91-107.
, Borel, M., Pichot, Y., Trut, G., 1991. Growth and mortality of the Euro
edulis in the Bay of Arcachon (France). Aquatic living resources. Nantes, 4(4), 265
Trintignac, P., 1996. Microalgae and larval feeding in mollusc hatchery.
Jonsson, P.R., 2000. Survival and feeding activity of oyster spat (
function of temperature and salinity with implications for culture policies on the Swedish west
sh Research, 19(2), 799-808.
, Kooistra, Whcf., Medlin, L.K., Percopo, I., Zingone, A., 2005. Diversity in the genus
Skeletonema (Bacillariophyceae). II. An assessment of the taxonomy of
rantes à Bonamia ostrae:
acid composition of early nonfeeding
Journal of the Marine Biological Association
The potential use of lipid microspheres as nutritional
Biochemical changes in fed and starved oysters, Ostrea edulis L.
Journal of the Marine
Larval growth and settlement of the
) as a function of food duality measured as fatty acid
Camacho, A., 1999. Dynamics of fatty acids in the larval
Ostrea edulis (L.). Comparative
, 123(3), 249-254.
of growth and gametogenesis in
temperatures. Journal of the
sh culture in France. Part 2. Oyster and mussel biology]. Revue Travaux
ects of nutrition on larvae production in the european flat oyster,
Selecting the flat oyster
Bonamia ostreae. Journal of
Growth and mortality of the European oyster Ostrea
, 4(4), 265-274.
Trintignac, P., 1996. Microalgae and larval feeding in mollusc hatchery. Haliotis, 26, 1-13.
al and feeding activity of oyster spat (Ostrea edulis L.) as a
function of temperature and salinity with implications for culture policies on the Swedish west
Diversity in the genus
Skeletonema (Bacillariophyceae). II. An assessment of the taxonomy of S. costatum-like

species with the description of four new species.
Uriarte, I., Farias, A., Hernandez, J., Schafer, C., Sorgeloos., 2004.
Chilean scallop (Argopecten
enriched diets. Aquaculture
Utting, S.D., Millican, P.F., 1997. Techniques for the hatchery conditioning of bivalve broodstocks and
the subsequent effect on egg quality and larval v
Utting, S.D., Millican, P.F., 1998.
review. Aquaculture, 165(3
Wilson, J.A., Chaparro, O.R., Thompson, R.J., 1996.
viability of larvae and spat in the Chilean oyster
~ 23 ~
species with the description of four new species. Journal of Phycology, 41(1), 151
ernandez, J., Schafer, C., Sorgeloos., 2004. Reproductive conditioning of
Chilean scallop (Argopecten purpuratus) and the Pacific oyster (Crassostrea gigas
Aquaculture, 230, 349-357.
Millican, P.F., 1997. Techniques for the hatchery conditioning of bivalve broodstocks and
ect on egg quality and larval viability. Aquaculture, 155(1
Millican, P.F., 1998. The role of diet in hatchery conditioning of Pecten maximus
, 165(3-4), 167-178.
Thompson, R.J., 1996. The importance of broodstock nutrition on the
viability of larvae and spat in the Chilean oyster Ostrea chilensis. Aquaculture
, 41(1), 151-176.
Reproductive conditioning of
Crassostrea gigas): effects of
Millican, P.F., 1997. Techniques for the hatchery conditioning of bivalve broodstocks and
, 155(1-4), 45-54.
Pecten maximus L.: a
The importance of broodstock nutrition on the
Aquaculture, 139(1-2), 63-75.

ANNEXE 1
0
5000
10000
15000
20000
25000
30000
35000
19
50
19
53
19
56
19
59
Production en tonnes
Figure 12: Production mondiale et française de l'huitre plate européenne (au kilogramme en France (source FAO).
~ 24 ~
19
59
19
62
19
65
19
68
19
71
19
74
19
77
19
80
19
83
19
86
19
89
19
92
19
95
19
98
20
01
prix $US/kg
années
Production Mondiale (tonnes)Production Française (tonnes)prix moyen en France $US/kilo
: Production mondiale et française de l'huitre plate européenne (Ostrea edulis
au kilogramme en France (source FAO).
0
1
2
3
4
5
6
7
8
20
04
prix $US/kg
Ostrea edulis), et prix moyen

ANNEXE 2
Algues Concentration (%)
C. gracilis
t-Iso
Figure 13: Protocole expérimentale pour l’ajustement de
1ère semaine
Acclimatation à la
température (T° exp.=
19°C) et à la nourriture
[100%]
2ème semaine expérience
[100%]
3ème semaine ré-
acclimatation à la
nourriture
[100%]
4ème semaine expérience [100%]
Isochrysis affinis galbana
Dispositif
salle expérience
Bac
(15huîtres)
X3
Bac de 50L,
alimenté en flux
ouvert
~ 25 ~
Concentration (%) Apport journalier (cellule algale.géniteur
-1.jour
-1)
75% 0,825.109
100% 1,1.109
125% 1,375.109
100% 2,7.109
125% 3,375.109
Protocole expérimentale pour l’ajustement de l’apport nutritionnel journalier
[100%] [100%] [100%]
[100%] [100%] [75%]
[100%] [100%] [100%]
[100%]
[125%]
[100%]
Isochrysis affinis galbana,
clone t-Iso Chaetoceros gracilis
Bac
(15huîtres)
X3
Bac
(15huîtres)
X3
Bac
(15huîtres)
X3
l’apport nutritionnel journalier
[100%]
[100%]
[100%]
[125%]
Chaetoceros gracilis
Bac
(15huîtres)
X3

ANNEXE 3
Entrée Eau de mer
Sortie Eau de mer
Filtre
5µm
Filtre
1µm
Isochrysis
affinis
galbana,
clone t
Skeletonema
marinoï
30
huîtres
30
huîtres
30
huîtres
30
huîtres
30
huîtres
30
huîtres
~ 26 ~
Figure 14: Schéma de la salle d'expérience
Filtre
UV
Filtre
µm
Isochrysis
affinis
galbana,
clone t-Iso
Tetraselmis
suecica
Chaetoceros
gracilis
Skeletonema
marinoï
Mesure de la
concentration
algale :
C1
C2
Sortie Eau de mer
30
huîtres
30
huîtres
30
huîtres
30
huîtres
30
huîtres
30
huîtres

ANNEXE 4
Tableau 2: Test t entre le taux de consommation
de prélèvement de pseudofèces, pour chaque régime nutritionnel.
t-Iso Moyenne
Groupe 1
Moyenne
Groupe 2
Consommation 3,151.109 2,585.10
9
Ingestion 0,002 0,002
C. gracilis Moyenne
Groupe 1
Moyenne
Groupe 2
Consommation 6,073.109 6,101.10
9
Ingestion 0,006 0,006
S. marinoï Moyenne
Groupe 1
Moyenne
Groupe 2
Consommation 6,583.109 6,812.10
9
Ingestion 0,002 0,002
T. suecica Moyenne
Groupe 1
Moyenne
Groupe 2
Consommation 3,401.109 3,480.10
9
Ingestion 0,002 0,002
~ 27 ~
: Test t entre le taux de consommation et le taux d'ingestion des 5 semaines de l'expérience et des 5 joursde prélèvement de pseudofèces, pour chaque régime nutritionnel.
Test t pour des échantillons indépendants
Valeur
t dl p
N actifs
Groupe
1
N actifs
Groupe
2
Ec-type
Groupe 1
Ec-type
Groupe 2
1,547 89 0125 79 12 1,238.109 0,634.10
1,078 89 0,284 79 12 0,001 0,001
Test t pour des échantillons indépendants
Valeur
t dl p
N actifs
Groupe
1
N actifs
Groupe
2
Ec-type
Groupe 1
Ec-type
Groupe 2
-0,124 117 0,901 104 15 0,820.109 0,855.10
0,453 117 0,651 104 15 0,001 0,001
Test t pour des échantillons indépendants
Valeur
t dl p
N actifs
Groupe
1
N actifs
Groupe
2
Ec-type
Groupe 1
Ec-type
Groupe 2
-0,658 88 0,512 81 9 1,037.109 0,204.10
-1,653 88 0,102 81 9 0,000 0,000
Test t pour des échantillons indépendants
Valeur
t dl p
N actifs
Groupe
1
N actifs
Groupe
2
Ec-type
Groupe 1
Ec-type
Groupe 2
-0,253 76 0,801 67 11 0,974.109 0,890.10
-1,054 76 0,295 67 11 0,001 0,001
et le taux d'ingestion des 5 semaines de l'expérience et des 5 jours
type
Groupe 2
Ratio F
variances
P
variance
634.109 3,814 0,018
001 1,994 0,206
type
Groupe 2
Ratio F
variances
P
variance
855.109 1,087 0,757
001 1,389 0,344
type
Groupe 2
Ratio F
variances
P
variance
204.109 25,811 0,000
000 4,136 0,037
type
Groupe 2
Ratio F
variances
P
variance
890.109 1,197 0,807
001 2,911 0,069

ANNEXE 5
Tableau 3: Tests statistiques de Levene, ANOVA, Scheffé pour l'expérience
Test de Levene d’Homogénéité des Variances
Variable: Proportion de Pseudofèces dans les biodépôts
Algues SC Effet Dl
Effet MC Effet
Pour C.
gracilis 5,555 2
Pour t-Iso 52,593 1
Variable: Proportion de Pseudofèces dans les biodépôts
Algues SC Effet Dl
Effet MC Effet
Pour C.
gracilis 5897,230 2 293
Pour t-Iso 5270,741 1 5270
Différences Variable: Proportion de Pseudofèces dans les
Ci C. gracilis
75 0
100
Ci t-Iso
100 0
~ 28 ~
statistiques de Levene, ANOVA, Scheffé pour l'expérience d’ajustement de l’apport journalier
Test de Levene d’Homogénéité des Variances Effet significatifs marquées à p < 0,05
Variable: Proportion de Pseudofèces dans les biodépôts
MC Effet SC Erreur Dl
Erreur MC
Erreur F
2,776 96,347 11 8,759 0,317 0,
52,593 1123,822 11 102,166 0,515 0,
Analyse de la Variance Effet significatifs marquées à p < 0,05
Variable: Proportion de Pseudofèces dans les biodépôts
MC Effet SC
Erreur Dl
Erreur MC
Erreur F
293,615 340,281 11 30,935 9,491 0,
5270,741 2677,083 11 243,371 21,657 0
Test Scheffé Différences significatives marquées à p < 0,05 Variable: Proportion de Pseudofèces dans les
biodépôts
100 125
0,293 0,004
0,048
125
0,001
d’ajustement de l’apport journalier
p
0,735
0,488
p
0,004
0,001

ANNEXE 6
Tableau 4: Tests statistiques de Levene, ANOVA, de filtration, taux de consommation, taux d'ingestion,
Test de Levene d’Homogénéité des Variances / Effet
Variables SC Effet Dl Effet
Taux de filtration
(l.g-1
.h-1
) 0,328 3
Taux de
consommation
(µm3.g
-1.h
-1)
1,488.1018
3
Taux d’ingestion (g.g-
1.h
-1)
0,000 3
Efficacité
d’absorption ������ √ "#
�$$% 0,030 3
Absorption (g.g-1
.h-1
) 0,000 3
Analyse de la Variance / Effet
Variables SC Effet Dl Effet
Taux de
consommation
(µm3.g
-1.h
-1)
5,630.1020
3
Efficacité
d’absorption������ √ "#
�$$% 3,141 3
Test Scheffé ; Différences significatives marquées à p < 0,
ALGUES C. gracilis
t-Iso 0,000
C. gracilis
S. marinoï
Test Scheffé ; Différences significatives marquées à p < 0,
C. gracilis
t-Iso 0,000
C. gracilis
S. marinoï
Test Kruskal-Wallis : H(3, N=290)=209,2252 p=0,
ALGUES C. gracilis
t-Iso 0,000
C. gracilis
S. marinoï
Test Kruskal-Wallis : H(3, N=290)=24
ALGUES C. gracilis
t-Iso 0,000
C. gracilis
S. marinoï
Test Kruskal-Wallis : H(3, N=40)=
ALGUES C. gracilis
t-Iso 0,000
C. gracilis
S. marinoï
~ 29 ~
: Tests statistiques de Levene, ANOVA, Scheffé, Kruskal-Wallis pour l'expérience sur l'écophysiologie (taux , taux d'ingestion, efficacité d’absorption, absorption) des géniteurs d'Ostrea edulis.
Test de Levene d’Homogénéité des Variances / Effet significatifs marquées à p < 0,
Dl Effet MC Effet SC Erreur Dl
Erreur MC Erreur
3 0,109 0,418 286 0,001
3 4,960.1017
8,256.1019
286 2,887.10
3 0,000 0,000 286 0,013
3 0,010 0,145 36 0,004
3 0,000 0,000 36 0,000
Analyse de la Variance / Effet significatifs marquées à p < 0,05
Dl Effet MC Effet SC Erreur Dl
Erreur MC Erreur
3 1,877.1020
2,295.1020
286 8,023.10
3 1,047 0,481 36 0,013
gnificatives marquées à p < 0,05 ; Variable : Taux de consommation (µm
C. gracilis S. marinoï
000 0,000
0,003
gnificatives marquées à p < 0,05 ; Variable : Efficacité d’absorption
C. gracilis S. marinoï
000 0,000
0,312
: H(3, N=290)=209,2252 p=0,00 ; Valeurs p des Comparaisons MultiplesVariable : Taux de filtration (l.g
-1.h
-1)
C. gracilis S. marinoï
000 0,000
1,000
: H(3, N=290)=247,4502 p=0,00 ; Valeurs p des Comparaisons MultiplesVariable : Taux d’ingestion (g.g
-1.h
-1)
C. gracilis S. marinoï
000 0,000
0,000
: H(3, N=40)=28,65118 p=0,00 ; Valeurs p des Comparaisons MultiplesVariable : Absorption (g.g
-1.h
-1)
C. gracilis S. marinoï
000 0,000
1,000
Wallis pour l'expérience sur l'écophysiologie (taux , absorption) des géniteurs d'Ostrea edulis.
significatifs marquées à p < 0,05
MC Erreur F p
001 74,95 0,000
.1017
1,718 0,163
13 5,638 0,001
004 2,498 0,075
000 9,079 0,000
MC Erreur F p
.1017
233,878 0,000
013 78,334 0,000
Taux de consommation (µm3.g
-1.h
-1)
T. suecica
0,888
0,000
0,000
Efficacité d’absorption ������ √ "#�$$%
T. suecica
0,000
0,923
0,133
Valeurs p des Comparaisons Multiples
T. suecica
1,000
0,000
0,000
Valeurs p des Comparaisons Multiples
T. suecica
0,003
0,000
0,000
Valeurs p des Comparaisons Multiples
T. suecica
0,175
0,108
0,238

ANNEXE 7
Tableau 5: Test de Levene, ANOVA, Scheffé pour les biométries
Analyse de la Variance en fonction des Algues et de la Date
Variables SC Effet
Longueur coquille (cm) 59,340
Poids Frais (g) 1151,189
Variables SC Effet
Poids sec chair (g) 0,640
Test Scheffé ; Différences
ALGUES/DATE C. gracilis
t0
t-Iso t0 0,973
C. gracilis t0
S. marinoï t0
T. suecica t0
t-Iso tF
C. gracilis tF
S. marinoï tF
Test Scheffé ; Différences s
ALGUES/DATE C. gracilis
t0
t-Iso t0 1,000
C. gracilis t0
S. marinoï t0
T. suecica t0
t-Iso tF
C. gracilis tF
S. marinoï tF
Test Scheffé ; Différences significatives marquées à p<0,05Variable
Algues t-Iso tF
Témoin t0 0,652
t-Iso tF
C. gracilis tF
S. marinoï tF
Test de Levene d’Homogénéité des Variances
Variables SC Effet
Longueur coquille (cm) 0,770
Poids Frais (g) 323,275
Poids secs chair (g) 0,0281
~ 30 ~
: Test de Levene, ANOVA, Scheffé pour les biométries
Analyse de la Variance en fonction des Algues et de la Date Effet significatifs marquées à p < 0,05
Dl Effet
MC Effet
SC Erreur Dl Erreur MC Erreur
7 8,477 185,950 638 0,
7 164,456 46137,19 638 72
Analyse de la Variance en fonction de t0 et des AlguesEffets significatifs marquées à p<0,05
Dl Effet
MC Effet
SC Erreur Dl Erreur MC Erreur
4 0,160 2,103 70 0,
Différences significatives marquées à p < 0,05 ; Variable : Longueur (cm)
S. marinoï
t0
T. suecica t0
t-Iso tF C. gracilis
tF
0,964 0,534 0,955 0,000
1,000 0,988 0,415 0,000
0,992 0,382 0,000
0,054 0,000
0,000
Différences significatives marquées à p < 0,05 ; Variable : Poids frais total (g)
S. marinoï
t0
T. suecica t0
t-Iso tF C. gracilis
tF
1,000 1,000 0,986 0,531
1,000 1,000 0,993 0,609
0,988 0,984 0,517
0,977 0,470
0,984
Différences significatives marquées à p<0,05 Variable : Poids sec chair (g)
C. gracilis tF S. marinoï tF T. suecica tF
0,790 0,339 0,143
0,999 0,994 0,024
0,960 0,034
0,006
Test de Levene d’Homogénéité des Variances / Effet significatifs marquées à p < 0,05
Dl Effet MC Effet SC Erreur Dl Erreur MC Erreur
7 0,110 68,343 638 0
7 46,182 18419,85 638 28
4 0,007 0,917 70 0
MC Erreur F p
,291 29,085 0,00
72,315 2,274 0,027
Analyse de la Variance en fonction de t0 et des Algues Effets significatifs marquées à p<0,05
MC Erreur F p
,030 5,325 0,001
: Longueur (cm)
S. marinoï
tF
T. suecica
tF
0,000 0,964
0,000 1,000
0,000 1,000
0,000 0,996
0,042 0,412
0,279 0,000
0,000
: Poids frais total (g)
S. marinoï
tF
T. suecica
tF
0,982 0,987
0,991 0,975
0,979 0,989
0,970 0,993
1,000 0,676
0,987 0,106
0,993
marquées à p < 0,05
MC Erreur F p
0,107 1,026 0,411
28,871 1,600 0,132
0,013 0,537 0,709

Abstract: The incidence of food quality and quantity
bivalves species remains poorly known nowadays. Suspension feeding bivalves collect their food
from seawater by retaining suspended organic particles. Because particles retention depends on
microalgae species, ingestion rate, assimilation efficiency and absorption
been studied in Ostrea edulis broodstock to determine the ideal diet for its conditioning. Oysters
were conditioned at 19 °C and fed four single diets:
gracilis, Skeletonema marinoï an
90% for all experimental diets which gave confidence to the results reported here. The highest
ingestion rates were recorded with
T. suecica and t-Iso were two fold lower (2.5 and 1.5 mg g
also exhibited highest assimilation efficiency (38 and 48%) and absorption (2.0 and 1.9 mg g
respectively). In contrast, similar values were recorded with
while t-Iso led to very low values (1.3% for EA and 0.01
growth was recorded during this
relative dry weight decrease leading to a significant difference between diets at the end of
experiment. These results spotlight that diatoms are essential for
are inefficient on a physiological point of view.
Keywords: Ostrea edulis, Feeding, Ingestion rate, Assimilation efficiency, Absorption,
Conditioning.
Résumé: L'incidence de la qualité et quantité de la nourriture des géniteurs sur les larves de
différentes espèces de bivalves reste mal connus de nos jours. Les bivalves
nourriture à partir de l’eau de mer, en retenant les pa
rétention des particules dépend des espèces de microalgue, le taux
d’absorption et le taux d’absorption
d’Ostrea edulis pour déterminer le régime
été conditionnées à 19°C et nourri
galbana (t-Iso), Chaetoceros gracilis
l’expérience, la survie était supéri
confiance dans les résultats reportés ici.
gracilis et S. marinoï (5,3 et 4,0) alors que ceux relevés
faible (2,5 et 1,5 mg.g-1.h-1 respectivement).
efficacité d’absorption (38 et 48%) et d’absorption (2,0 et 1,9 mg.g
similaires ont été enregistrées avec
faibles sont notés pour t-Iso (1,3% pour l’EA
croissance significative n’a été enregistrée
suecica révèle une relative décroissance en poids
régimes à la fin de l’expérience. Ces résultats montre
pour Ostrea edulis par rapport à
écophysiologique.
Mots clés: Ostrea edulis, Nutrition, Taux d’ingestion,
Conditionnement.
~ 31 ~
: The incidence of food quality and quantity from larvae to adults of some
species remains poorly known nowadays. Suspension feeding bivalves collect their food
from seawater by retaining suspended organic particles. Because particles retention depends on
microalgae species, ingestion rate, assimilation efficiency and absorption of four microalgae have
broodstock to determine the ideal diet for its conditioning. Oysters
were conditioned at 19 °C and fed four single diets: Isochrysis affinis galbana
and Tetraselmis suecica. At the end of experiment, survival was high
90% for all experimental diets which gave confidence to the results reported here. The highest
ingestion rates were recorded with C. gracilis and S. marinoï (5.3 and 4.0) while those record
Iso were two fold lower (2.5 and 1.5 mg g-1 h-1 respectively). C. gracilis
also exhibited highest assimilation efficiency (38 and 48%) and absorption (2.0 and 1.9 mg g
respectively). In contrast, similar values were recorded with T. suecica (36% for EA and 1.3 mg g
w values (1.3% for EA and 0.01 mg g-1 h-1). Whatever the diets
this 5 week period experiment. However, T. suecica
relative dry weight decrease leading to a significant difference between diets at the end of
experiment. These results spotlight that diatoms are essential for O. edulis while
ficient on a physiological point of view.
, Feeding, Ingestion rate, Assimilation efficiency, Absorption,
L'incidence de la qualité et quantité de la nourriture des géniteurs sur les larves de
de bivalves reste mal connus de nos jours. Les bivalves filtreurs
nourriture à partir de l’eau de mer, en retenant les particules organique en suspension. Parce que la
rétention des particules dépend des espèces de microalgue, le taux d’ingestion, l’
le taux d’absorption de quatre microalgues ont été étudiées pour les géniteurs
pour déterminer le régime le plus adapté pour leur conditionnement. Les huîtres
été conditionnées à 19°C et nourries avec quatre régimes monospécifiques
Chaetoceros gracilis, Skeletonema marinoï et Tetraselmis suecica
supérieure à 90% pour tous les régimes, ce qui donne une certaine
ance dans les résultats reportés ici. Les plus forts taux d’ingestion ont été enregistrés pour
4,0) alors que ceux relevés pour T. suecica et t-Iso sont deux fois plus
respectivement). C. gracilis et S. marinoï montre
(38 et 48%) et d’absorption (2,0 et 1,9 mg.g-1.h-1). Au contraire, des valeurs
s avec T. suecica (36% pour l’EA et 1,3 mg.g-1.h-1) alors des valeu
Iso (1,3% pour l’EA et 0,01 mg.g-1.h-1). Quelle que soit le régime, aucune
ignificative n’a été enregistrée durant les cinq semaines d’expérience. Cependant
relative décroissance en poids sec montrant une différence par rapport aux autres
régimes à la fin de l’expérience. Ces résultats montrent donc que les diatomées sont essentielles
par rapport à T. suecica et t-Iso qui sont inefficaces
, Nutrition, Taux d’ingestion, Efficacité d’absorption
from larvae to adults of some marine
species remains poorly known nowadays. Suspension feeding bivalves collect their food
from seawater by retaining suspended organic particles. Because particles retention depends on
of four microalgae have
broodstock to determine the ideal diet for its conditioning. Oysters
Isochrysis affinis galbana (t-Iso), Chaetoceros
. At the end of experiment, survival was high
90% for all experimental diets which gave confidence to the results reported here. The highest
(5.3 and 4.0) while those recorded with
C. gracilis and S. marinoï
also exhibited highest assimilation efficiency (38 and 48%) and absorption (2.0 and 1.9 mg g-1 h-1
(36% for EA and 1.3 mg g-1 h-1)
ever the diets no significant
T. suecica exhibited some
relative dry weight decrease leading to a significant difference between diets at the end of the
while T. suecica and t-Iso
, Feeding, Ingestion rate, Assimilation efficiency, Absorption,
L'incidence de la qualité et quantité de la nourriture des géniteurs sur les larves de
filtreurs collectent leur
ticules organique en suspension. Parce que la
d’ingestion, l’efficacité
été étudiées pour les géniteurs
pour leur conditionnement. Les huîtres ont
s avec quatre régimes monospécifiques : Isochrysis affinis
Tetraselmis suecica. A la fin de
régimes, ce qui donne une certaine
ont été enregistrés pour C.
Iso sont deux fois plus
montrent aussi une forte
). Au contraire, des valeurs
) alors des valeurs très
que soit le régime, aucune
durant les cinq semaines d’expérience. Cependant T.
rant une différence par rapport aux autres
donc que les diatomées sont essentielles
d’un point de vue
Efficacité d’absorption, Absorption,

~ 32 ~





























