Embed Size (px)
Citation preview

Int J Colorect Dis (1989)4:75-90
Review
~j ........ ~ j o o l ( oliii ecta Disease
�9 Springer-Verlag 1989
Inflammatory intermediaries in inflammatory bowel disease
K. Lauritsen ~, L.S. Laursen ~, K. Bukhave 2 and J. Rask-Madsen 3
1 Department of Medical Gastroenterology, Odense University Hospital, Odense z Department of Biochemistry and Nutrition, The Technical University of Denmark 3 Section of Gastroenterology, Department of Medicine G, Bispebjerg Hospital, University of Copenhagen, Copenhagen, Denmark
The chronic inflammatory bowel diseases (IBD) include ulcerative colitis (UC) and Crohn's disease (CD). Their causes and precise relations remain obscure which is why these conditions are com- monly designated as "nonspecific" or "idiopathic". The current concept of their pathogenesis involves an interaction between host responses, immunolo- gic genetic influences, and environmental agents [1-3]. In the absence of decisive knowledge of the causes of IBD no consistently curative medical treatment exists and the foundation of drug treat- ment remains empirical. Within the last decade, however, increased research interest has been fo- cused on soluble intermediaries of the inflamma- tory response with the major object of uncovering specific targets for pharmacologic manipulation.
The inflammatory response
Inflammation is a term used to describe a series of responses of vascularized tissues to injury. The clin- ical signs of this phenomenon, acknowledged since ancient times (Celsus), can be related to increased local blood flow (calor and rubor), increased vascu- lar permeability and cellular infiltration (tumor), and release of a variety of substances at the site of inflammation that induces pain (dolor). Later, loss
Abbreviations." 5-ASA, 5-aminosalicylic acid; C, complement- derived peptide; CD, Crohn's disease; FMLP, N-formyl- methionyl-leucyl-phenylalanine; HETE, hydroxyeicosatetra- enoic acid; HPETE, hydroxypcroxyeicosatetraenoic acid; IBD, inflammatory bowel diseases; IL, interleukin; LT, leucotriene; NSAID, nonsteroidal anti-inflammatory drug; PAF-acether, platelet-activating factor; PG, prostaglandin; ROS, reactive oxy- gen species; TNF, tumour necrosis factor, TX, thromboxane; UC, ulcerative colitis
of function was added by Virchow as an additional cardinal sign of inflammation (functio laesa).
The mechanisms of inflammation depend great- ly on the site and the stimulus and are influenced by genetic, nutritional, as well as hormonal factors. Furthermore, the processes themselves represent an extensive network of interacting mechanisms, me- diators, and cells.
In principle inflammation is a beneficial reac- tion of tissues to injury, which normally leads to removal of the inciting agent and repair of the in- jured site. In contrast, the uncontrolled inflamma- tory reactions occuring in IBD may induce perma- nent tissue destruction.
The term "inflammatory intermediary" should, in essence, be reserved for chemical substances, which signal information between different cells in- volved in the inflammatory process. More than a hundred compounds appear to mediate some aspect or other of the inflammatory process, and this number will increase considerably if the current rate of discovery is maintained [4]. Inflammation thus becomes ever more complex, but over the last few years evidence for an ordered control of the inflammatory response has begun to emerge in the sense that inflammation is one of the integral parts of the body defence mechanisms.
The criteria used to determine whether an endo- genous substance serves a primary role in mediat- ing inflammation has been considered by Vane [5]: (i) the substance should be detectable, at the site of inflammation, in amounts adequate to account for the effect under consideration; (ii) the substance, when administered in concentrations of the order of those found in the lesion, should produce the observed effect; (iii) specific blocking agents or an- tagonists of the effects of the substance should pre- vent or attenuate the effect; (iv) prevention of re- lease of the substance should abolish or prevent the

76
MEMBRANE PHOSPHOLIPID
Phosphollpases (A2, C)
ARACHIDONIC ACID
C y c l o o x y g e d ~ 15-HPETE /Pathway k "Lipo~enase
/ ~ S " - ~ 12-HPETE CYCLIC ENDOPEROXIDEL ~ 5-HPETE
PGI S / I ~ "~ TXA 2 5-HETE " I " PoGD' 1 PGF2" [
OH OH I ~ , PGE 2 ~ OH
6-keto-PGF1. TXB 2 LTB 4
', 15-HETE
, 12-HETE
H O LTA.
jr LTC4'-e.. LTD4-e.. LTE 4
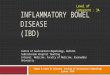
Fig. 1. Biosynthetic pathways ofeicosanoid metabolism leading to formation of prostaglandins (PGs), prostacyclin (PGI2), thromboxanes (TXs), hydroxyperoxyeicosatetraenoic acids (HPETEs), hydroxyeicosatetraenoic acids (HETEs), and leuco- trienes (LTs)
effect; and (v) agents or procedures preventing the breakdown or removal of the substances should prolong or potentiate the effect.
Eicosanoids
Chemistry and cellular origin
These compounds (eicosa derived from the Greek term denoting 20) constitute a group of biologically active lipid mediators derived from 20-carbon "es- sential" fatty acids. In mammals, the most im- portant is arachidonic acid (5,8,11,14-eicosa- tetraenoic acid), which is released from cellular phospholipids by the activity of various phospholi- pases [6, 7] and subsequently metabolized either through the cyclo-oxygenase pathway [8] to prosta- glandins (PGs), prostacyclin (PGI2), and throm- boxanes (TXs) or through the lipoxygenase path- ways [9, 10] to hydroxyperoxyeicosatetraenoic acids (HPETEs), hydroxyeicosatetraenoic acids (HETEs), and leucotrienes (LTs) (Fig. 1). PGs are designated A through F depending on the exact structure of the cyclopentane ring. The subscript number (for example, 1, 2, or 3) denotes the num- ber of double bonds in the side chain. The Greek
letter that follows (either alpha or beta) describes the stereochemistry at the carbon 9 position. Also for LTs the subscript figure (3, 4, or 5) indicates the number of double bonds, three of which occur at alternate carbon atoms (a conjugated "triene").
Eicosanoid formation in mammalian tissues is ubiquitious, but the profile of eicosanoids varies dependent on species, tissue, and cell population within the tissue. Most studies on the regulation of eicosanoid synthesis have concluded that the avail- ability of arachidonic acid is the limiting factor for the total output of its metabolites [6]. Arachidonic acid may be released by the Ca2+-dependent en- zyme, phospholipase A2, from phosphatidyl- choline, phosphatidylethanolamine, and phospha- tidylinositol [6,7]. However, release of arachidonic acid can also be derived from inositol phospho- lipids through alternative reactions catalyzed by phospholipase C and diacylglycerol [11]. Very little is known about the regulation of further steps ei- ther in the cyclo-oxygenase or the lipoxygenase pathways. Several mechanisms and routes of inacti- vation and degradation are available, either locally or in the lungs, but neither oxidation nor substi- tution or uptake by excretory organs are known to be under specific control. On stimulation substan- tial amounts of eicosanoids may be released, but no indications are presently available for storage and only little is known about the mechanism of excre- tion from the producing cells.
Different cell types have different potentials for arachidonic acid metabolism. Whether receptor- mediated regulation of oxidative metabolism of

77
arachidonic acid relates to the distribution and ac- tion of membrane phospholipases and lipases or to the presentation of phospholipid substrates or both is not known. Whereas the oxidative metabolism of arachidonic acid to cyclooxygenase products via five or more terminal synthetases [12] with differen- tial cellular prominence is characteristic of most cell types, 5-1ipoxygenation appears to be less widely distributed, mainly to neutrophils, eosino- phils, monocytes, macrophages, and mast cells. These cells originate in the bone marrow and prob- ably derive from the same stem-cell. Further, among these cell types relevant to host inflamma- tory response, the generation of LTs, quantitatively and qualitatively, exibits remarkable cellular speci- ficity [13]. Thus in the stimulated neutrophil, the major route of arachidonic acid metabolism along the 5-1ipoxygenase pathway accounts for produc- tion of 5-HETE and LTB 4 [14]. With eosinophils, the preferentially generated compound is LTC 4 [15]. Monocytes and macrophages are able to synthesize both LTB~ and the sulfidopeptide-lipid LTs [12].
Methodological problems
Materials for studying the role of eicosanoids in IBD have comprised not only patients with the disorders and healthy controls, but also a variety of animal models [16, 17]. The pattern of arachidonic acid metabolism in these models (and the effect of medication) varies considerably with the model and the experimental conditions [18-20] and the results are interpreted in the light of the notion that lack of a reliable model for IBD has usually been consid- ered a hall-mark of these disorders. The experimen- tal designs applied include various in vitro meth- ods, such as determination of contents of eicosa- noids or activities of enzymes in tissue homo- genates and generation of eicosanoids in organ cul- tures or in leucocytes derived from blood or intes- tinal tissues. These approaches may define the ca- pacity of the tissue to produce different eicosa- noids, and thus the profile of arachidonic acid me- tabolites. Ex vivo organ perfusions have also been applied. The in vivo methods include equilibrium and nonequilibrium dialysis against rectal mucosa or faeces. These approaches appear presently to provide a more reliable index of the balance be- tween intestinal mucosal eicosanoid formation and degradation in vivo [21-23], since they limit arti- ficial eicosanoid formation.
Currently available analytical methods are bioassays, thin-layer chromatography with auto- radiography, radioimmunoassays, high pressure
liquid chromatography, and gas chromatography- mass spectrometry [8-10, 21].
Formation of eicosanoids in IBD
Since Gould [24] first reported raised concentra- tions of PG-like material in stools from patients with an acute attack of UC, in vitro experiments with fresh biopsies of colonic mucosa obtained from patients with IBD have demonstrated PGE2- , LTB4- , and 5-HETE concentrations three, six and ten times higher than in normal tissues [25, 26], in addition to increased activities of PG synthetase [27]. The amounts of PGE z, PGF2~, 6-keto-PGFl~ (that is, the stable breakdown product of PGI2) , TXB 2 (that is, the stable breakdown product of T X A 2 ) , 12-HETE, 15-HETE, LTB4, and LTC4 generated by cultures of rectal mucosa have also been reported abnormally increased [25, 26, 28-301.
Similarly, the accumulation o fPGE 2 and TXB 2 in the medium of cultured peripheral blood mono- nuclear cells [31] and intestinal mononuclear cells [32] from patients with CD, but not in the medium of cells from patients with UC [31, 32], has been reported significantly increased compared with that measured in the medium of cells from controls. By contrast, neutrophils activated with the calcium ionophore A23187 [33] appear to have increased capacity for release of LTBa in both groups of pa- tients, but not if activated during phagocytosis of artificial immune complexes [34]. Also the forma- tion of LTB 4 and LTC 4 in organ cultures obtained from rectal biopsies of patients with CD is in- creased compared to control cultures [30, 35] and the ileal mucosa from a small group of patients with CD has shown a trend toward increased phos- p h o l i p a s e A 2 activity, even in unaffected areas [36].
Measurements of tissue concentrations of eico- sanoids are suspect, however, because these com- pounds are generally not stored. The effect of ma- nipulating (for example, by biopsy) could be to activate membrane bound phospho!ipases with subsequent artificial eicosanoid formation [20-22, 37, 38], which would also reflect increased arachi- donic acid contents in the phospholipids as ob- served in patients with UC [39]. Similar limitations apply to tissue culture techniques [23]. Also the conclusions based on profiles of products obtained in response to ionophore activation should be tem- pered due to the limited data available for cells in response to transmembrane physiologic stimuli [13]. Finally, the findings that intestinal phagocytes appear to behave differently from similar cells in

78
peripheral blood originating in patients with IBD have emphasized the limitation of data obtained from studies of phagocytes in blood [40].
Measurements of blood or plasma levels of eicosanoids as well as urinary excretions are inap- propriate for estimation of local production. Also results of plasma determinations of eicosanoids are often artifactual because synthesis may occur dur- ing bloodletting. Consequently, plasma concentra- tions may vary by orders of magnitude [21, 41].
In vivo formation of eicosanoids has been esti- mated, therefore, by measuring the rate of PGE 2 accumulating in dialysis bags placed in the empty rectum [42-46] or the equilibrium concentrations of PGE 2 and LTB 4 [47-53] by use of the same technique or the original method [54] of in vivo dialysis of faeces [48, 51, 52, 55]. The named meth- ods have demonstrated that the luminal concentra- tions of eicosanoids are always raised in active dis- ease, but decrease toward normal levels in inactive disease. Also profiles of eicosanoids have been de- termined by equilibrium in vivo dialysis of the rec- tum in consecutive untreated patients with IBD and Clostridium difficile associated colitis [50]. PGE2, PGF2~ and TXB2, but not 6-keto-PGFl~, were abnormally high in all types of inflammation, whereas extreme rises in PGE 2 and TXB/ levels were observed in patients with UC. By contrast, the luminal concentrations of LTB4 were significantly increased only in the group of patients with UC, although abnormally high values were observed in single cases of Crohn's colitis and Clostridium diffi- cile colitis with rectal ulceration [50].
Eicosanoids and the activity o f IBD
Positive correlations between the luminal concen- tration of PGE 2 or LTB 4 and disease activity as judged by clinical, endoscopic, or histologic grad- ings have been demonstrated by means of the meth- od of equilibrium in vivo dialysis of the rectum in patients with distal UC [47]. A decrease in raised concentrations of the named eicosanoids toward the levels observed in healthy controls occurred only in patients who responded to treatment. Fur- thermore, pretreatment PGE 2 and LTB 4 levels seemed to be significantly higher in patients not responding to therapy than in those who improved [47], Approximately 15 % of patients with UC con- fined to the rectum and sigmoid colon are resistant to most forms of drug treatment [56]. If pretreat- ment levels of PGE2 and LTB4 reflect local disease activity, they may prove more useful predictors of
the outcome of medical treatment in these patients than clinical indices of disease activity [48, 51].
Finally, it has been suggested that a rise in PGE2 concentrations in rectal dialysis fluid from patients with UC in complete remission precedes the development of clinical symptoms in those who later suffer a relapse [52]. In vivo dialysis of the rec- tum for determination of luminal PGE 2 concentra- tions may, therefore, improve decision-making in symptom-free patients, who wish to stop sulphasa- lazine medication, but relatively frequent measure- ments are desirable [52].
Potential role in inflammation
The role of eicosanoids in various aspects of intes- tinal function, such as transmucosal ion transport, mucosal blood flow, microvasculature, and motil- ity, in addition to mucus composition and secretion has been reviewed elsewhere [57-60]. The present paper will consider only the potential role of eicosa- noids in inflammation.
Prostaglandins enhance vasodilation and oedema formation, acting synergistically with other mediators of inflammation (for example, histamine and bradykinin), whereas LTs promote leucocyte migration and stimulate aggregation and degranu- lation of neutrophils and the release of lysosomal enzymes and superoxide production [14, 61-66]. Hence, LTB 4 is the most potent chemotactic (i.e., stimulating directional movements of neutrophils) and chemokinetic (i.e., stimulating random move- ment of neutrophils) eicosanoid and its activity ex- ceeds that of other eicosanoids 100-fold [67]. The cell motion and secretory activities of LTB 4 could be due to changes in calcium and sodium flux across the cell membranes of neutrophils [68]. LTB 4 induces neutrophil chemotaxis in vitro (69) at con- centrations of 3 ng/ml with a maximum response at 30 ng/ml - comparable to concentrations observed in rectal dialysates from patients with UC [50]. Also 5-HETE is a weak chemotactic agent, whereas LTC4, LTD 4 and LTE4 (collectively accounting for the biological activity of "slow-reacting substance of anaphylaxis") have vasoconstrictor activity and contribute to oedema by increasing capillary per- meability.
There is increasing evidence that platelets, in addition to their function in haemostasis, play a role in inflammation by exhibiting a platelet- leucocyte cell-to-cell interaction [70]. Indeed, the platelet eicosanoid, 12-HPETE, formed during pla- telet aggregation leads to activation of human neu- trophil 5-1ipoxygenase up to levels obtained upon

79
stimulation of the cells with the calcium ionophore A 23187 [12]. Considering the findings of markedly enhanced in vivo LTB 4 formation in UC [47] to- gether with the known biological effects of the com- pound (especially its chemotactic and chemokinetic properties) is suggestive of a triggering role of this substance in the inflammatory process. Hypothet- ically, tissue macrophages and neutrophils present in the intestinal mucosa would be activated by phagocytic and chemotactic stimuli, such as the complement-derived peptide C5a [71], and by me- diators released by aggregating platelets, in addition to various kinins [72]. LTB 4 would then be released and act as a chemoattractant to stimulate further the migration of neutrophils, thus accelerating the development of the inflammatory reaction. The essential function of LTB 4 would, therefore, be to amplify or stabilize the chemotactic gradients [68, 73].
Platelet-activating factor (PAF-acether)
PAF-acether is a phospholipid mediator [74] formed by eosinophils, macrophages, neutrophils, platelets, and vessel endothelium. Like eicosanoids, but unlike other potential mediators of inflamma- tion, PAF-acether is not stored as such in the cells, but is present as a precursor phospholipid (1-0- alkyl- 2 (R) acyl- glycero- 3- phosphatidylcholine) in cell membranes. It is released by de-acylation to lyso-PAF, following activation of phospholipase A 2 and subsequent acetylation by an acetyl trans- ferase. PAF-acether is catabolized by at least two different acetyl hydrolases, a cytosolic and a plas- ma enzyme [75], the latter being a very effective means of removing circulating PAF-acether.
Formation of PAF-acether accompanies the stimulation of most inflammatory cells as well as platelets and vascular endothelial cells. Further- more, PAF-acether activates eosinophils, neutro- phils and platelets [76]. The pharmacological ac- tions include a variety of in vivo effects related to inflammation, particularly to immediate hyper- sensitivity [75, 77]. Increased formation of PAF- acether determined by bioassay and characterized by its migration on thin layer chromatography has been reported in incubated colonic mucosa ob- tained from patients with UC [78], in addition to tissue levels in various models of colitis [79]. Similar methodological constraints as described for eicosa- noids must be acknowledged in interpreting these findings.
Different PAF-acether antagonists have been synthesized [75] but their role in experimental and
human colitis has not been worked out. In any case, the burst in PAF-acether antagonists will help to unravel the situation and is likely to provide new tools and more important, possible new remedies.
Biogenie amines (histamine and 5-hydroxytryptamine)
Histamine (2-(4-imidazolyl)ethylamine) and 5-hy- droxytryptamine are naturally occurring amines stored within the body and capable of eliciting ma- jor changes in a variety of bodily functions [80-82]. Currently, it appears that 5-hydroxytryptamine is a mediator of acute inflammation in rodents only [83]. Histamine is formed by decarboxylation of histidine and is stored primarily in mast cells, al- though histidine decarboxylase activity has been found in most tissues.
Release of histamine from mast cells immedi- ately after injury has been reported in response to a number of stimuli ranging from anaphylaxis to both physical and chemical insults. Local adminis- tration of histamine elicits classical vascular re- sponses of acute inflammation, such as increase in blood flow and vascular permeability, oedema for- mation, and pain and itching. The histamine H 1 receptors are considered to be responsible for the pro-inflammatory activities. By contrast, sustained release of histamine in chronic inflammation, such as in IBD, has not been proven, but histamine may serve as a co-mediator to PGs in potentiating an increase in vascular permeability. Studies with his- tamine antagonists are consistent with the role of histamine in the early phase of acute inflammation and not in more sustained inflammation.
Kinins
Kinins are a group of small peptides formed in blood and biological fluids by the actions of kalli- kreins (kininogenases) on alpha 2 globulins, the kininogens [84 86]. In general, the concentration of kinins in circulating blood and tissue is low since the kallikreins are coupled to inhibitors and kept as inactive precursors, the prekallikreins. When an appropriate physiological or pathological stimulus activates the prekallikreins, the nonapeptide bra- dykinin is formed in the blood by the action of plasma kallikreins on high molecular weight kinin- ogens (plasma pathway). These proteins are also required for the intrinsic blood coagulation cascade [86]. Similarly, the decapeptide kallidin (lysyl bra- dykinin) is released in tissues by the actions of tis-

80
Surface HMW-Kininogen HF ~ aHF
IT I [ HMW_Kininogen
Prekallikrein ~ Kallikrein
HMW-Kininogen
Factor XI "Pro"-Kallikrein
Intracellular Enzymes Plasmin
~ P a s m a Kallikrein
Tissue Kallikrein Factor XIA ~, LMW-Kininogen
]1 Coagulation ~ Lysyl Bradykinin I, [ Bradykinin ] (Kallidin)
Carboxypeptidase N IAminopeptidas e (CPN)
Des_ArgO_Bradykini n ~ CPN I Bradykinin] Angiotensin Converting Enzyme
l[Otherf P asma Proteases
Inactive Peptides, Amino Acids
Fig. 2. Mechanisms of bradykinin for- mation by plasma and tissue pathways. HF, Hageman factor (coagulation factor XII); aHF, activated Hageman factor; HMW, high molecular weight; LMW, low molecular weight
sue kallikreins on low molecular weight kininogens (tissue pathway) (Fig. 2). Inactivation of the kinins occurs locally and in the lungs and the kidneys.
Only in recent years, with improved assay tech- nologies and an increased awareness of the mecha- nisms regulating kinin levels, meaningful direct evi- dence to support a role for the kinin system in human inflammatory disorders has begun to accu- mulate. The kinins exert a variety of biological ac- tions by activating at least two different receptor types. B 1 and B 2 receptors for kinins have been identified with agonists and antagonists in classical pharmacological assays [84]. The existence of the two receptor types has been confirmed with bind- ing assays, using labelled bradykinin and des Arg 9 bradykinin, its octapeptide metabolite following C-terminal arginine removal [87].
Antagonists for kinins active on B1 receptors have been identified and further developed to fairly potent, specific and competitive blockers [88]. B 2 receptor antagonists have also been found [89] and they are specific, but rather weak and nonselective, since they also block B 1 receptors. B 2 receptors mediate a large number of rapidly occurring bio- logical effects, particularly the symptoms and signs of inflammation [87], while B 1 receptors appear to be involved in some retarded, long lasting effects of kinins, such as collagen synthesis and cell multipli- cation [90].
Kinins are among the naturally occurring agents involved in inflammatory reactions, for ex- ample, vasodilation, increase of vascular permea- bility, venoconstriction, and mobilization of blood and tissue cells. The kinins generated locally con- tribute to the acute and possibly the chronic phase of the inflammatory reaction by producing vasodi-
lation, local oedema, and pain. Kinins may also modulate migration of blood and tissue leucocytes that take part in the inflammatory process.
Several of the biological effects of the kinins are mediated by eicosanoids [84]. Hence kinins are among the most potent activators of PG formation [87] and the major actions, for instance the endo- thelium mediated vasodilation, the smooth muscle contraction or relaxation, and the production of pain are associated with release of PGs. In models of IBD increased mucosal generation of kinins is associated with the induction of a special subtype of bradykinin receptors in inflamed colonic tissue [91], which might contribute to the increased PG formation by bradykinin-induced release of arachi- donic acid [92, 93]. Kinin-induced PGE 2 seems to be derived principally from cells in the lamina pro- pria and not from the epithelial cells [94]. In the healthy colonic mucosa this eicosanoid release ap- pears to mediate the bradykinin-stimulated secre- tion [92]. In an ex vivo model of colitis, increased vascular resistance has been observed following bradykinin infusion and was associated with an exaggerated generation of TXs. The increase in per- fusion pressure was blunted, but not abolished, by concomitant infusion of selective TX inhibitors [93]. Although the exaggerated eicosanoid release may function to modulate colonic blood flow, and contribute to manifestations of inflammation [17], the relevance of these observations for IBD is spec- ulative.
Complement-derived peptides
The complement system of blood plasma and ex- travascular tissue fluid plays an important role in

81
many immune defence reactions and absence of a functional complement system reduces many in- flammatory reactions [95]. The system consists of about 20 different proteins [96-98] which, once the system is activated, interact in an orderly, sequen- tial and regulated series of reactions [99]. Comple- ment activation promotes acute inflammation, re- cruitment of leucocytes, and killing of pathogens by phagocytosis, lysis, or release of toxic products [71, 96, 100]. The central event in complement acti- vation is cleavage of C3 with the liberation of C3a and its metabolite C3a des Arg. This is accom- plished by activation of either of two pathways (Fig. 3), the classical and alternative pathways, to produce C3 convertase. The classical pathway is activated by IgG or IgM antibodies, once they have formed a complex with the antigen. The alternative pathway [101] is typically activated by polysaccha- rides. The large fragment of C3 cleavage, C3b, binds covalently to the activator and promotes im- mune adherence and phagocytosis. Addition of C3b to either of the C3 convertases results in the formation of a C5 convertase which cleaves C5 into C5a and C5b. This is the last enzymatic step in the complement cascade, because the larger fragment, C5b, and later components, C6-C9, form the lytic complex (membrane attack complex) by a series of hydrophobic interactions [98]. C5a plays no further direct role in the complement cascade, but contri- butes to the inflammatory response. It is rapidly converted by a carboxypeptidase, found in plasma and extracellular tissue fluid, to C5a des Arg by loss of the carboxyl terminal arginine.
C3a and C5a are anaphylatoxins, because they release histamine and stimulate smooth muscle contraction [96]. C5a has probably the most impor- tant biological activities [71]. More important, however, the C5-derived peptides have potent effects on leucocytes in vitro and promote neutro- phil-endothelial interactions in vivo, which lead to neutrophil accumulation and associated oedema formation [100]. In addition to initiating neutro- phil-endothelial cell interactions, C5a and C5 des Arg may also contribute to inflammation by stimu- lating the release of other mediators. Apart from the release of histamine, C5a appears to release interleukin-1 [102], PGs [103], LTs [104], and PAF- acether [105]. Thus, inflammatory responses begun by complement activation may be prolonged and potentiated by the actions of other intermediaries released by C5a.
Although there are few studies showing an ef- fect of complement-derived peptides on the gastro- intestinal tract, the biological properties of C5a and C3a suggest that these compounds play a role in
Classical Pathway IgG / IgM-Antigen
Complexes -o ~
Ciqrs ' .
C4b2~o ' .
C3
C5
Alternative Pathway Biological Effect Surface Contact
(Polysaccharides) Z
oO. " D
."~ C3bBh
- ~ J= C3a. .
~ C3b . . . . . . . . . . . . . . . ~ . . . . . . ~ [ Phagocytosis ]
C4b2*b3b C313b3b . . . .,~
v [ 1 �9 C5a . . . . . . . . . . . ~ I A c u t e I n f l a m m a t i o n [
C5b
C7
C8
C9
C5b6789 . . . . . . . . . . . . . . . . . ~ I - ' - - I
Fig, 3. Activation of complement. The central event, cleavage of the third component to produce C3a and C3b, is mediated by either of two distinct pathways, known as the classical and alternative pathways. Proteolytic activity is denoted by *
IBD. Complement activation is normally an appro- priate and beneficial reaction to inflammation. In theory, it may be desirable to be able to inhibit complement activation in inappropriate inflamma- tory states like IBD, though a beneficial effect is by no means certain. Alternatively, the effects of a C5a receptor antagonist may be beneficial, but such compounds are not available at present. The pharmacological manipulation of the complement system has been reviewed elsewhere [106].
Lymphokines
Lymphokines or 'cytokines' are polypeptides that act on leucocytes and other tissue targets. These compounds activate, modulate, and control vari- ous aspects of body defence and repair. The group includes the interleukins (ILs) and tumour necrosis factor (TNF). Interleukin-1 (IL-1), produced by macrophages in response to a large number of stim- uli [107], activates lymphocytes and also induces several components of the acute-phase response to infection and injury. Its most dramatic biological property is its ability to induce eicosanoid forma- tion [108]. Interleukin-2 (IL-2), produced by T lym- phocytes, is a growth factor for helper, suppressor,

82
and cytotoxic T lymphocytes. In contrast to IL-1, IL-2 is a single substance, a true growth factor with seemingly the single purpose of expanding T-cell populations [109], whereas IL-1 is a family of poly- peptides that mediates several components of the acute inflammation, in addition to affecting a num- ber of nonleucocytic targets, such as the liver, pan- creas, bone, muscle, synovial fibroblasts, and brain [108, 110].
Two forms of IL-I have been cloned, IL-l-beta from human monocytes [111] and IL-l-alpha from a murine macrophage line [112]. The two forms had originally been identified as pI 7 (beta) and pI 5 (alpha) on isoelectric focusing [107]. The two forms of IL-1 are initially synthesized as 31 kiloDalton precursor peptides and share only small stretches of amino acid homology. Depending on the stimulus, intracellular levels of precursor IL-1 can be high, whereas some cell activators result in large amounts of processed IL-1. Precursor IL-1 is cleaved into a 17.5 kiloDalton peptide, which is the predominant extracellular form. Another macrophage product - TNF - has been cloned and shown to be identical to a substance termed cachectin, which mediates cachexia associated with various diseases [113]. Many of the biological effects of IL-1 and TNF overlap, and the compounds often act synergist- ically [107].
IL-1 induces eicosanoids and lymphocyte acti- vation as well as many different biological activities including fever, acute-phase protein synthesis, fi- broblast proliferation, and other defence and repair functions [107, 108, 110]. Most if not all of these effects are mediated by an IL-1 stimulated increase in phospholipase A 2 activity and subsequent re- lease and metabolism of arachidonic acid from cel- lular phospholipids in the target cells. Hence the eicosanoids provide the second signal for IL-1 in- duced cell activation [108].
It is conceivable that IL-1 plays a role in the inflammation in IBD [114]. Whereas the peripheral blood IL-2 pathway appears normal in patients with IBD [115] increased serum concentrations of IL-1 have been reported in some patients with UC and CD [116] and intestinal mucosal levels [117, 118] and production of IL-I in colonic tissue organ culture [118] are increased in patients with IBD compared with controls.
Chemotactic oligopeptides
As already described a number of substances are chemotactic for leucocytes, for example LTs and
C5a. Another class of chemoattractants is peptides elaborated by bacteria [119, 120]. The best studied of these oligopeptides is N-formyl-methionyl- leucyl-phenylalanine (FMLP), a neutrophil chemo- tactic factor produced by E. coli [121]. All chemo- tactic peptides appear to produce their effects by binding to cell-surface receptors, and FMLP recep- tors on human neutrophils have been characterized [119]. Hence both high- and low-affinity forms of the oligopeptide receptor have been identified by direct binding studies.
The chemotactic peptides induce directed mi- gration of inflammatory cells toward their targets, and cause degranulation with lysosomal release, superoxide production, phagocytosis, and eicosa- noid formation [119, 122]. Leucotriene B 4 poten- -tiates the response elicited. In perfused rabbit colon FMLP stimulates release of eicosanoids [123] and intrarectal administration of high concentrations of FMLP has been used as a model of UC [124]. Thus, it may be speculated that bacterial oligopep- tides, originating in the colonic lumen, breach mucosal defences and initiate colonic inflamma- tion.
Neuropeptidcs
The involvement of the nervous system in the gen- eration of inflammatory responses is well estab- lished [125-127]. Polymodal nociceptor activation generates axon-reflexes in the terminal arborisa- tions of primary afferent neurones. These reflexes, in turn, cause the C fibres to release neuropeptides, for example, the undecapeptide, substance P [128, 129]. Neurokinin A (substance K), neurokinin B (neuromedin K), vasoactive intestinal polypeptide (VIP) and calcitonin gene-related peptide (CGRP) are other possible candidates [125].
The neuropeptides are believed to exert their effects on vascular smooth muscles and endothe- lium to increase flow and permeability, in addition to altering the motility of the muscularis mucosae. It has been suggested that the effects of substance P at least in part are mediated by IL-1 [130].
So far, in contrast to other tissues, little is known about the role and mechanisms of neuro- peptides in intestinal inflammation. Increased mucosal/submucosal concentrations of substance P have been observed in patients with UC, but not CD [131], whereas others have reported high con- centrations of substance P receptor binding sites in colonic tissue obtained from both groups of pa- tients [132].

83
Free radicals - reactive oxygen species (ROS)
Free radicals are chemical species (atoms, ions or molecules) with one or more unpaired electrons in their outer orbital, rendering the compound chemi- cally reactive. The odd electron is often represented in the formula as a dot. The univalent reduction of oxygen by certain oxidases produces the superoxide anion radical (O2). Other reactive oxygen species (ROS) are hydrogen peroxide (H202) and the hy- droxyl radical (OH). HzO 2 is formed by the diva- lent reduction of 02 (or the univalent reduction of O2 ) to yield the non-radical species 022 which is protonated at physiological pH to HzO 2. In the presence of iron II or copper I it is reduced and the hydroxide ion ( O H ) and the hydroxyl radical (OH) are formed. Several enzymes, including xanthine oxidase, aldehyde oxidase, and NADPH oxidase, are known to produce superoxide directly or indirectly as a reaction product [133]. Phagocytic leucocytes, when stimulated, produce large am- ounts of ROS via the NADPH oxidase system [134, 135] and intestinal microvascular and mucosal epi- thelium also contains various oxidases, for exam- ple, xanthine oxidase, a cytosolic enzyme normally present in its dehydrogenase form, but converted to an oxidase form during ischaemia [136].
The cellular toxicity associated with superoxide and hydrogen peroxide is generally ascribed to their role as a precursor of hydroxyl radicals and other secondary ROS [133]. The hydroxyl radicals oxidize sulphhydryl compounds that may result in inactivation of essential enzymes or membrane transport proteins. Furthermore, the radicals will react with DNA to cause strand scission [137] or other abnormalities, and attack of membrane asso- ciated polyunsaturated fatty acids results in the peroxidation of lipids [138]. Lipid peroxidation of- ten precedes lysis of target cells and is also associat- ed with increased rates of protein fragmentation and changes in composition [139] theoretically as- sociated with changes in antigenicity. Finally, free radical production may have a potentiating effect on PG synthesis, initiating lipid peroxidation and providing the hydrogenperoxide required for the early stages of the PG cascade.
Since radical reactions are self-perpetuating un- less two radicals interact, the net effect of generat- ing ROS in vivo will be widespread and un- predictable. The production of ROS is essential to normal metabolism, but they are theoretically de- structive unless tightly controlled. The endogenous defence is provided by catalytic free radical removal, for example, by various enzymes, such as superoxide dismutase, catalase, and gluthathione
peroxidase, by physiological non-enzymatic "free radical scavengers", such as alpha-tocopherol and ascorbate, and by removal of free transition iron and copper ions [133 135] through binding to transferrin or coeruloplasmin.
Although all cells possess numerous antioxi- dant enzymes and scavengers to protect themselves from ROS, the rate of production of ROS may exceed the capacity of the antioxidant defences, thereby resulting in substantial tissue damage. In animal experiments ROS mediate much of the mi- crovascular injury associated with reperfusion of ischaemic intestine [140, 141]. It has also been sug- gested that transient ischaemia in patients with UC initiates a cascade of self-perpetuating cycles or ROS formation, inflammation and, ultimately, mucosal injury [133].
Implication for therapy in IBD
It is hoped that a better understanding of the inter- mediaries and mechanisms of inflammation will lead toward a better therapeutic control of un- desirable elements of inflammation. It should be borne in mind, however, that the more potent the therapeutic intervention the more likely there is to be undesirable side effects. The current drug thera- py of IBD established through controlled trials mainly relies on the use of corticosteroids and sul- phasalazine [142].
Corticosteroids
Despite the widespread use of the corticosteroids it had for long been difficult to conceptualize their anti-inflammatory effects in any single model [143]. In recent years, however, a considerable amount of evidence points to a modulation of eicosanoid for- mation. Corticosteroids have also been shown to inhibit the release of non-arachidonic acid media- tors from many inflammatory cell types (for exam- ple, histamine release from basophils). Because ac- tivation of these cell types may require phospholi- pase activity, it is not yet clear whether the corti- costeroids inhibit these cellular processes via a sim- ilar mechanism [144]. Unlike aspirin and other nonsteroidal anti-inflammatory drugs (NSAIDs), which block the cyclo-oxygenase pathway [145], the corticosteroids do not inhibit arachidonic acid me- tabolism in cell-free systems [145, 146]. In contrast, these drugs are considered to act by preventing formation of free arachidonic acid for eicosanoid formation [147 149]. This effect is believed to be

84
mediated by promotion of a natural phospholipase A2 inhibitor, commonly denoted as lipocortin [150]. Recently, the principal biological species in the lipocortin family in both rat and human, a 40 kiloDalton protein, has been isolated, purified to homogeneity, sequenced, and cloned [151, 152]. The described effect of corticosteroids is fairly re- producible in various in vitro systems [153] and in most experimental models of inflammation in animals, although in some experimental conditions exceptions to this concept have been found [154, 155]. Such failure of corticosteroids to prevent eico- sanoid formation suggests that multiple functional pools of phospholipase are operative in relation to the arachidonic acid cascade [154]. Also basal PG production in vivo has been reported to be un- affected by pharmacological doses of corticoste- roids [156, 157]. However, studies on the profile of eicosanoids in the inflamed rectum [49] in patients with severely active UC have demonstrated that prednisolone (1.5mg/kg/day orally) promptly (i. e., within 72 h) decreases luminal concentrations of both PGE 2 and LTB4, whereas indomethacin (150 mg/day orally) reduces PGE 2 concentrations in spite of unchanged, or even increased, LTB 4 levels. These prompt reductions are likely to be caused by a direct drug action, rather than being secondary to decreased tissue damage, and accord with the lipocortin theory [150].
Sulphasalazine and 5-aminosalicylic acid (5-ASA)
Interpretation of results obtained in vitro on the mode of action of sulphasalazine and 5-ASA has been hampered by the lack of knowledge of the therapeutically relevant concentrations of the drugs. Such results include inhibition of PG synthe- sis [24, 25, 28], increased intestinal "cytoprotec- tion" [158] due to inhibition of PG degradation [159-162], and stimulation of PG synthesis [163], inhibition of TX synthesis [164, 165], reduction of LT formation [30, 165-169], modulation of poly- morphonuclear leucocyte function [170-173], im- paired receptor-binding of chemotactic peptides [174], and impairment of folate absorption and me- tabolism [175]. A common feature of such in vitro experiments is the use of concentrations derived from in vivo measurements of so-called "therapeu- tic serum concentrations" (20-50 pg/ml) of sul- phapyridine [176]. This may explain at least some of the conflicting data in the papers referred to. Although the exact concentrations of 5-ASA in the mucosa are unknown, the therapeutically relevant
concentrations seem to be those achieved intralu- minally. This notion is supported by the observa- tions that 5-ASA acts topically after rectal instilla- tion [177-180] and that the concentration of 5-ASA in serum is negligible [181,182] in the pres- ence of high luminal concentrations after oral ad- ministration [182].
That sulphasalazine and 5-ASA exert their ther- apeutic benefit through modulation of eicosanoid formation is a suggestion underlying a substantial amount of experimental work. Hence there has been considerable discussion whether the therapeu- tic effect relies on inhibition of the cyclooxygenase activity, suppression of the activity of PG-degrad- ing enzymes, or reduction of the lipoxygenase ac- tivity.
The hypothesis, that sulphasalazine acts through promotion of endogenous cytoprotective PGs [183] is based on in vitro observations of in- creased PGE 2 and PGI 2 production by normal co- lonic mucosa in the presence of 5-ASA, in addition to decreased enzymatic degradation of PGs in the presence of sulphasalazine (but not 5-ASA) [159-163], Nevertheless, the effect of sulphasa- lazine on PG formation may vary depending on substrate arachidonic acid availability [184], in ad- dition to drug concentration [163]. Evidence against the proposed mechanisms of drug action is provided by experiments in humans, which demon- strate that therapeutic doses of the drug (2 g/day) result in concentrations of 5-ASA, which are 100 to 1,000 times higher [182] than those stimulating PG synthesis in vitro [163]. Furthermore, both sulpha- salazine (2 g/day) and olsalazine (2 g/day) normal- ize significantly raised intraluminal PGE 2 levels, even in patients with inactive UC. These findings suggest that the named drugs modify colonic PG production in the absence of clinical and endo- scopic disease activity [55].
A number of in vitro experiments [25, 28, 30, 185, 186], in addition to observations in vivo [55], have suggested that 5-ASA is a weak cyclooxygen- ase inhibitor. The latter trait is most likely un- related to its therapeutic efficacy, because more potent cyclo-oxygenase inhibitors, such as NSAIDs, are ineffective in this respect [43, 45, 185, 187]. Both sulphasalazine and 5-ASA block the soybean lipoxygenase [167, 168] (although this ef- fect is disputed as regards sulphasalazine [188]) and inhibit the formation of 5-HETE and LTB 4 by hu- man leucocytes [166, 169], in addition to inhibiting the colonic lipoxygenase [26, 30]. The inhibitory effect (for example, the drug concentration neces- sary for half-maximal reduction in the formation of lipoxygenase products), the target level in the path-

85
way, and the differential effect of sulphasalazine and 5-ASA appear, however, to depend on the sys- tem used and the experimental conditions.
Finally, there is some evidence to indicate that interventions directed toward oxidant scavenging and xanthine oxydase inhibition may provide a new approach to the management of IBD. 5-ASA is an extremely efficient oxygen radical scavenger, suggesting that the beneficial effect of salicylates in IBD may be due, in part at least, to scavenging of enterocyte- or phagocyte-derived oxidants [189 192].
New drug development
As mentioned previously, the rationale for seeking pharmacotherapeutic agents to limit LT biosyn- thesis and/or end-organ effects is based upon the increased generation of these compounds in IBD, their potent proinflammatory actions, and the lack of clinical efficacy by blocking one of the two major routes of arachidonic acid metabolism (i. e., the cyclooxygenase pathway), in addition to the con- trastingly remarkable efficacy of corticosteroids that inhibit the formation of oxygenation products via both routes. It is possible that such compounds possess corticosteroid-like therapeutic efficacy but are free from steroid-related toxicity. Even if this is over optimistic, the inhibition of LT synthesis must be seen as a promising concept and if one or more of these compounds ultimately prove to benefit pa- tients with IBD, it would concomitantly substan- tiate the belief that LTs are mediators of primary importance for the inflammatory process.
Prospects
Progress in therapy of IBD is most likely to come from a better understanding of the role of the solu- ble intermediaries of inflammation. In particular, it appears likely that lipoxygenase products of arachi- donic acid are potent intermediaries that may prove to be important in the pathogenesis of IBD. To confirm or refute this belief requires the demon- stration of clinical efficacy by selective lipoxyge- nase inhibitors or LT receptor antagonists through controlled clinical trials.
Improved knowledge of the role of inflamma- tory intermediaries has allowed reinterpretation of the rationale behind current therapy. Cortico- steroids and sulphasalazine, the drugs with estab- lished efficacy in IBD, affect arachidonic acid re-
lease and metabolism, which may prove to be the scientific basis of IBD therapeutics.
References
1. Kirsner JB, Shorter RG (1982) Recent developments in nonspecific inflammatory bowel disease. N Engl J Med 306:775-785, 837 848
2. Elson CO, Kagnoff MF, Fiocchi C, Befus AD, Targan S (1986) Intestinal immunity and inflammation: recent pro- gress. Gastroenterology 91 : 746- 768
3. Strober W, James SP (1986) The immunologic basis of inflammatory bowel disease. J Clin Immunol 6:415-432
4. Majno G (1985) Inflammatory mediators: where are they going? In: Higgs GA, Williams TH (eds) Inflammatory mediators. MacMillan, Basingstoke, pp 1-6
5. Vane JR (1972) Prostaglandins in the inflammatory re- sponse. In: Lepow IH, Ward PA (eds) Inflammation: mechanisms and control. Academic Press, New York, pp 261-279
6. Irvine RF (1982) How is the level of free arachidonic acid controlled in mammalian cells? Biochem J 204:3-16
7. Blackwell GJ, Flower RJ (1983) Inhibition of phospholi- pase. Br Med Bull 39:260 264
8. Samuelsson B, Goldyne M, Granstr6m E, Hamberg M, Hammerstr6m S, Malmsten C (1978) Prostaglandins and thromboxanes. Annu Rev Biochem 47:997-1029
9. Hammerstr6m S (1983) Leukotrienes. Annu Rev Biochem 52:355-377
10. Borgeat P, Nadeau M, Salari H, Poubelle P, de Laclos BF (1985) Leukotrienes: biosynthesis, metabolism, and analy- sis. Adv Lipid Res 21:47 77
t 1. Majerus PW, Wilson DB, Conolly TM, Bross TE, Neufeld EJ (1985) Phosphoinositide turnover provides a link in stimulus-response coupling. Trends Biochem Sci 10: 168-171
12. Needleman P, Turk J, Jakschik BA, Morrison AR, Lefko- with JB (1986) Arachidonic acid metabolism. Annu Rev Biochem 55:69 102
13. Lewis RA, Austen KF (1984) The biologically active leu- kotrienes. Biosynthesis, metabolism, receptors, functions, and pharmacology. J Clin Invest 73:889-897
14. Samuelsson B (1983) Leukotrienes: mediators of immedi- ate hypersensitivity reactions and inflammation. Science 220:568 - 575
15. Weller PF, Lee CW, Foster DW, Corey EJ, Austen KF, Lewis RA (1983) Generation and metabolism of 5-1ipoxygenase pathway leukotrienes by human eosino- phils: predominant production of leukotriene C 4. Proc Natl Acad Sci USA 80:7626-7630
16. Boughton-Smith NK, Whittle BJR (1988) The role ofeico- sanoids in animal models of inflammatory bowel disease. In: Goebell H, Peskar BM, Malchow H (eds) Inflammato- ry bowel diseases. Basic research and clinical implications. MTP Press Lancaster, pp 175-199
17. Brown JA, Zipser RD (1987) Prostaglandin regulation of colonic blood flow in rabbit colitis. Gastroenterology 92: 54- 59
18. Sharon P, Stenson WF (1985) Metabolism of arachidonic acid in acetic acid colitis in rats: similarity to human in- flammatory bowel disease. Gastroenterology 88:55-63
19. Boughton-Smith NK, Whittle BJR (1985) Increased me- tabolism of arachidonic acid in an immune model of colitis in guinea pigs. Br J Pharmacol 86:439-446
20. Zipser RD, Nast CC, Lee M, Kao HW, Duke R (1987) In vivo production of leukotriene B 4 and leukotriene C 4 in

86
rabbit colitis. Relationship to inflammation. Gastroenter- ology 92:33 39
21. Rask-Madsen J, Bukhave K (1984) The difficulties of establishing the pathophysiological role of prostaglandins in secretion. In: Skadhauge E, Heintze K (eds) Intestinal absorption and secretion. MTP Press, Lancaster, pp 453 468
22. Zipser RD, Laffi G (1985) Prostaglandins, thromboxanes and leukotrienes in clinical medicine. West J Med 143:485-497
23. Rigas B (1986) Eicosanoids and the gastrointestinal tract: promising but no verdict yet. Am J Gastroenterol 81:218-221
24. Gould SR (1975) Prostagtandins, ulcerative colitis, and sulphasalazine. Lancet ii:988
25. Sharon P, Ligumsky M, Rachmilewitz D, Zor U (1978) Role of prostaglandins in ulcerative colitis. Enhanced pro- duction during active disease and inhibition by sulfasala- zine. Gastroenterology 75:638-640
26. Sharon P, Stenson WF (1984) Enhanced synthesis of leu- kotriene B 4 by colonic mucosa in inflammatory bowel dis- ease. Gastroenterology 86:453 460
27. Harris DW, Smith PR, Swan CHJ (1978) Determination of prostaglandin synthetase activity in rectal biopsy material and its significance in colonic disease. Gut 19:875-877
28. Ligumsky M, Karmeli F, Sharon P, Zor U, Cohen F, Rachmilewitz D (1981) Enhanced thromboxane A 2 and prostacyclin production by cultured rectal mucosa in ulce- rative colitis and its inhibition by steroids and sulfasalazi- he. Gastroenterology 81:444 449
29. Boughton-Smith NK, Hawkey CJ, Whittle BJR (1983) Biosynthesis of lipoxygenase and cyclo-oxygenase pro- ducts from ~4C-arachidonic acid by human colonic mucosa. Gut 24:1176-1182
30. Peskar BM, Dreyling KW, Peskar BA, May B, Goebell H (1986) Enhanced formation of sulfidopeptide-leukotrienes in ulcerative colitis and Crohn's disease: inhibition by sulfasalazine and 5-aminosalicylic acid. Agents Actions 18:381 383
31. Rachmilewitz D, Ligumsky M, Haimowitz A, Treves AJ (1982) Prostanoid synthesis by cultured peripheral blood mononuclear cells in inflammatory diseases of the bowel. Gastroenterology 82:673-679
32. Zifroni A, Treves AJ, Sachar DB, Rachmilewitz D (1983) Prostanoid synthesis by cultured intestinal epithelial and mononuclear cells in inflammatory bowel disease. Gut 24:659-664
33. Nielsen OH, Ahnfelt-Ronne I, Elmgreen J (1987) Abnor- mal metabolism of arachidonic acid in chronic inflamma- tory bowel disease: enhanced release of leucotriene B 4 from activated neutrophils. Gut 28:181-185
34. Nielsen OH, Elmgreen J, Thomsen BS, Ahnfelt-Ronne I (1986) Release of leukotriene B 4 and 5-hydroxyeicosa- tetraenoic acid during phagocytosis of artificial immune complexes by peripheral neutrophils in chronic inflamma- tory bowel disease. Clin Exp Immunol 65:465-471
35. Hawkey C J, Karmeli F, Rachmilewitz D (1983) Imbalance of prostacyclin and thromboxane synthesis in Crohn's dis- ease. Gut 24:881-885
36. Bolin T, Heuman R, Sj6dahl R, Tagesson C (1984) De- creased lysophospholipase and increased phospholipase A 2 activity in ileal mucosa from patients with Crohn's disease. Digestion 29:55 59
37. Bennett A, Stamford IF, Stockley HL (1977) Estimation and characterization of prostaglandins in the human gastrointestinal tract. Br J Pharmacol 61:579-586
38. Gr6en K, Aly A, Johanson C (1981) Measurements of prostaglandin biosynthesis in the gastrointestinal tract: biochemical and technical problems. Prostaglandins 21 [Suppl]: 1 -7
39. Nishida T, Miwa H, Shigematsu A, Yamamoto M, Iida M, Fujishima M (1987) Increased arachidonic acid composi- tion of phospholipids in colonic mucosa from patients with active ulcerative colitis. Gut 28:1002-1007
40. Hermanowicz A, Gibson PR, Jewell DP (1985) The role of phagocytes in inflammatory bowel disease. Clin Sci 69:241-249
41. Metz SA, McRae JR, Robertson RP (1981) Prostaglandins as mediators of paraneoplastic syndromes: review and up- date. Metabolism 30:299 316
42. Rampton DS, Sladen GE (1981) The effect of sulphasala- zine withdrawal on rectal mucosal function and prosta- g l a n d i n E 2 release in inactive ulcerative colitis. Scand J Gastroenterol 16:157-159
43. Rampton DS, Sladen GE (1981) Prostaglandin synthesis inhibitors in ulcerative colitis: flurbiprofen compared with conventional treatment. Prostaglandins 21:417 425
44. Rampton DS, Sladen GE (1981) Relapse of ulcerative proctocolitis during treatment with non-steroidal anti- inflammatory drugs. Postgrad Med J 57:297 299
45. Rampton DS, Sladen GE (1984) Relationship between rec- tal mucosal prostaglandin production and water and elec- trolyte transport in ulcerative colitis. Digestion 30:13-22
46. Rampton DS, Sladen GE, Youlten LJF (1980) Rectal mu- cosal prostaglandin E 2 release and its relation to disease activity, electrical potential difference, and treatment in ulcerative colitis. Gut 21 : 591 - 596
47. Lauritsen K, Laursen LS, Bukhave K, Rask-Madsen J (1986) Effects of topical 5-aminosalicylic acid and predni- solone on prostaglandin E= and leukotriene B 4 levels deter- mined by equilibrium in vivo dialysis of rectum in relapsing ulcerative colitis. Gastroenterology 91 : 837- 844
48. Lauritsen K, Laursen LS, Bukhave K, Rask-Madsen J (1987) Intraluminal colonic levels of arachidonic acid me- tabolites in ulcerative colitis. Adv Prostaglandin Throm- boxane Leukotriene Res 17:347-352
49. Lauritsen K, Laursen LS, Bukhave K, Rask-Madsen J (1987) In vivo effects of orally administered prednisolone on prostaglandin and leucotriene production in ulcerative colitis. Gut 28:1095 1099
50. Lauritsen K, Laursen LS, Bukhave K, Rask-Madsen J (1988) In vivo profiles of eicosanoids in ulcerative colitis, Crohn's colitis, and Clostridium difficile colitis. Gastroen- terology 95:11 17
51. Lauritsen K, Laursen LS, Bukhave K, Rask-Madsen J (1988) Longterm olsalazine treatment: pharmacokinetics, tolerance and effects on local eicosanoid formation in ulce- rative colitis and Crohn's colitis. Gut 29:974-982
52. Lauritsen K, Laursen LS, Bukhave K, Rask-Madsen J (1988) Use of colonic eicosanoid levels as predictors of relapse in ulcerative colitis: double blind placebo control- led study on sulphasalazine maintenance treatment. Gut 29:1316-1321
53. Lauritsen K, Laursen LS, Bukhave K, Rask-Madsen J (1987) Does vitamin E supplementation modulate in vivo arachidonate metabolism in human inflammation? Phar- macol Toxicol 61:246-249
54. Wrong O, Metcalfe-Gibson A, Morrison RBI, Ng ST, Howard AV (1965) In vivo dialysis of faeces as a method of stool analysis. I. Technique and results in normal sub- jects. Clin Sci 28:357-375
55. Lauritsen K, Hansen J, Bytzer P, Bukhave K, Rask- Madsen J (1984) Effects of sulphasalazine and disodium

azodisalicylate on colonic PGE 2 concentrations determin- ed by equilibrium in vivo dialysis of faeces in patients with ulcerative colitis and healthy controls. Gut 25:1271-1278
56. Farmer RG (1985) Ulcerative colitis. Medical manage- ment. In: Berk JE, Haubrich WS, Kalser MH, Roth JLA, Schaffner F (eds) Bockus gastroenterology. Saunders, Phi- ladelphia, pp 2199 2207
57. Rampton DS, Hawkey CJ (1984) Prostaglandins and ulce- rative colitis. Gut 25:1399-1413
58. Hawkey CJ, Rampton DS (1985) Prostaglandins and the gastrointestinal mucosa: are they important in its function, disease, or treatment? Gastroenterology 89:1162-1188
59. Rask-Madsen J (1986) Eicosanoids and their role in the pathogenesis of diarrhoeal diseases. Clin Gastroenterol 15:545 566
60. Lauritsen K, Laursen LS, Bukhave K, Rask-Madsen J (1987) Role of eicosanoids in inflammatory bowel disease. In: J/irnerot G (ed) Inflammatory bowel disease. Raven Press, New York, pp 83-94
61. Goetzl EJ (1981) Oxygenation products of arachidonic acid as mediators of hypersensitivity and inflammation. Med Clin North Am 65:809-828
62. Davies P, Bailey P J, Goldenberg M, Ford-Hutchinson AW (1984) The role of arachidonic acid oxygenation products in pain and inflammation. Annu Rev Immunol 2:335 357
63. Higgs GA, Moncada S, Vane JR (1984) Eicosanoids in inflammation. Ann Clin Res 16:287-299
64. Piper PJ (1984) Formation and actions of leukotrienes. Physiol Rev 64:744-761
65. Sirois P (1985) Pharmacology of the leukotrienes. Adv Lipid Res 21:79 101
66. Parker CW (1987) Lipid mediators produced through the lipoxygenase pathway. Annu Rev Immunol 5:65 84
67. Marcus AJ (1985) Eicosanoids as bioregulators in clinical medicine. Am J Med 78:805-810
68. Naccache PH, Sha'afi RI (1983) Arachidonic acid, leuko- triene B4, and neutrophil activation. Ann NY Acad Sci 414:125-139
69. Goetzl EJ, Pickett WC (1980) The human PMN leukocyte chemotactic activity of complex hydroxy-eicosatetraenoic acids (HETEs). J Immunol 125:1789-1791
70. Borgeat P, Sirois P, Braquet P, Rola-Pleszczynski M (1986) Leukotrienes and inflammation. In: Cohen MM (ed) Biological protection with prostaglandins, vol I. CRC Press, Cleveland, pp 13-26
71. Jose PJ (1987) Complement-derived peptide mediators of inflammation. Br Med Bull 43:336 349
72. Donowitz M (1985) Arachidonic acid metabolites and their role in inflammatory bowel disease. An update re- quiring addition of a pathway. Gastroenterology 88: 580 587
73. Lobos EA, Sharon P, Stenson WF (1987) Chemotactic activity in inflammatory bowel disease. Role of leukotriene B 4. Dig Dis Sci 32:1380-1388
74. Snyder F (1985) Chemical and biochemical aspects of pla- telet activating factor: a novel class of acetylated ether- linked choline-phospholipids. Med Res Rev 5:107 140
75. Vargaftig BB, Braquet PG (1987) PAF-acether today - relevance for acute experimental anaphylaxis. Br Med Bull 43:312-335
76. Pinckard RN, McManus LM, Hanahan DJ (1982) Chem- istry and biology of acetyl glyceryl ether phosphoryl- choline (Platelet-activating factor). In: Weissmann G (ed) Advances in inflammation research. Raven Press, New York, 4:147 180
77. Bach MK (1982) Mediators of anaphylaxis and inflamma- tion. Annu Rev Microbiol 36:317-413
87
78. Wengrower D, Eliakim R, Karmeli F, Razin E, Rachmile- witz D (1987) Pathogenesis of ulcerative colitis (UC): en- hanced colonic formation of inositol phosphates (IP) and platelet activating factor (PAF) (abstract). Gastroenter- ology 92:1691
79. Boughton-Smith NK, Whittle BJR (1988) Formation of the proinflammatory mediator, PAF-acether in different models of colitis (abstract). Gastroenterology 94:A45
80. Austen KF (1979) Biological implications of the structural and functional characteristics of the chemical mediators of immediate-type hypersensitivity. Harvey Lect 73:93-161
81. Plaut M, Lichtenstein LM (1982) Histamine and immune responses. In: Ganellin CR, Parsons ME (eds) Pharmacol- ogy of histamine receptors. Wrigt PSG, Bristol, pp 392-435
82. Bradley PB, Fozard JR, Humphrey PPA, Staughan DW (eds) (1984) Proceedings of a symposium: 5-HT, peripheral and central receptors and function. Neuropharmacology 23:1465-1569
83. Owen DAA (1987) Inflammation - histamine and 5-hydroxytryptamine. Br Med Bull 43:256-269
84. Regoli D, Barab~ J (1980) Pharmacology of bradykinin and related kinins. Pharmacol Rev 32:1-46
85. Regoli D (1987) Kinins. Br Med Bull 43:270 284 86. Proud D, Kaplan AP (1988) Kinin formation: mechanisms
and role in inflammatory disorders. Annu Rev Immunol 6:49-83
87. Marceau F, Lussier A, Regoli D, Giroud JP (1983) Phar- macology of kinins: their relevance to tissue injury and inflammation. Gen Pharmacol 14:209-229
88. Regoli D, Drapeau G, Rovero P, Dion S, D'Ortbans-Juste P (1986) The actions of kinin antagonists on B 1 and B a receptor systems. Eur J Pharmacol 123:61-65
89. Vavrek R J, Stewart JM (1985) Competitive antagonists of bradykinin. Peptides 6:161-164
90. Goldstein RH, Wall M (1984) Activation of protein for- mation and cell division by bradykinin and des-Arg 9- bradykinin. J Biol Chem 259:9263-9268
91. Kachur JF, Allbee W, Gaginella TS (1986) Effect ofbrady- kinin and des-Argg-bradykinin on ion transport across normal and inflamed rat colonic mucosa (abstract). Gastroenterology 90:1481
92. Musch MW, Kachur JF, Miller RJ, Field M (1983) Bradykinin-stimulated electrolyte secretion in rabbit and guinea pig intestine. Involvement of arachidonic acid metabolites. J Clin Invest 71:1073-1083
93. Zipser RD, Patterson JB, Kao HW, Hauser C J, Locke R (1985) Hypersensitive prostaglandin and thromboxane re- sponse to hormones in rabbit colitis. Am J Physiol 249:G457-463
94. Hoult JRS, Phillips JA (1986) Kinin-induced prostaglan- din release in rat colon does not display serosal/mucosal 'sidedness' after epithelial removal. Br J Pharmacol 88:3-5
95. Schur PH (1986) Inherited complement component abnor- malities. Annu Rev Med 37:333 346
96. Hugli TE, Miiller-Eberhard HJ (1978) Anaphylatoxins: C3a and C5a. Adv Immunol 26:1-53
97. Joiner KA, Brown EJ, Frank MM (1984) Complement and bacteria: chemistry and biology in host defense. Annu Rev Immunol 2:461-491
98. Mfiller-Eberhard HJ (1986) The membrane attack com- plex of complement. Annu Rev immunol 4: 503- 528
99. Perlmutter DH, Colten HR (1986) Molecular immunbio- logy of complement biosynthesis: a model of single-cell control of effector-inhibitor balance. Annu Rev Immunol 4:231 251

88
100. Webster RO, Hong SR, Johnston RB, Henson PM (1980) Biological effects of the human complement fragments C5a and C5a des Arg on neutrophil function. Immuno- pharmacology 2:201 219
101. Fearon DT, Austen KF (1980) Current concepts in immu- nology: the alternative pathyway of complement - a sy- stem for host resistance to microbial infection. N Engl J Med 303:259-263
102. Goodman MG, Chenoweth DE, Weigle WO (1982) Induc- tion of interleukin 1 secretion and enhancement of humo- ral immunity by binding of human C5a to macrophage surface C5a receptors. J Exp Med 156:912 917
103. Rampart M, Bult H, Herman AG (1983) Activated com- plement and anaphylatoxins increase the in vitro produc- tion of prostacyclin by rabbit aorta endothelium. Naunyn Schmiedebergs Arch Pharmacol 322:158-165
104. Stimler NP, Bach MK, Bloor CM, Hugli TE (1982) Re- lease of leukotrienes from guinea pig lung stimulated by C5a des Arg anaphylatoxin. J Immunol 128:2247-2252
105. Camussi G, Aglietta M, Coda R, Bussolino F, Piacibello W, Tetta C (1981) Release of platelet-activating factor (PAF) and histamine. II. The cellular origin of human PAF: monocytes, polymorphonuclear neutrophils and ba- sophils. Immunology 42:191-199
106. Asghar SS (1984) Pharmacological manipulation of com- plement system. Pharmacol Rev 36:223-244
107. Billingham MEJ (1987) Cytokines as inflammatory media- tors. Br Med Bull 43:350-370
108. Dinarello CA (1984) Interleukin-1. Rev Inf Dis 6:51-95 109. Smith KA (1984) Interleukin-2. Annu Rev Immunol
2:319-333 110. Durum SK, Schmidt JA, Oppenheim JJ (1985) Inter-
leukin-l: an immunological perspective. Annu Rev Im- munol 3:263-267
111. Auron PE, Webb AC, Rosenwasser LJ, Mucci SF, Rich A, Wolff SM, Dinarello CA (1984) Nucleotide sequence of human monocyte interleukin-1 precursor cDNA. Proc Natl Acad Sci USA 81:7907-7911
112. Lomedico PT, Gubler U, Hellman CP, Dukovich M, Giri JG, Pan YC, Collier K, Semionow R, Chun AO, Mizel SB (1984) Cloning and expression of murine interleukin-1 in Escherichia coli. Nature 312:458-462
113. Beutler B, Cerami A (1986) Cachectin and tumor necrosis factor: two sides of the same biological coin. Nature 320:584 588
114. Cominelli F, Nast CC, Gentilini P, Zipser RD (1988) Re- combinant interleukin-1 induces colonic prostaglandin production: lymphokines may have an important role in colitis (abstract). Gastroenterology 94:A75
115. Ming RH, Atluru D, Spellman CW, Imir T, Goodwin JS, Strickland RG (1987) Peripheral blood mononuclear-cell interleukin-2 production, receptor generation and lympho- kine-activated cytotoxicity in inflammatory bowel disease. J Clin Immunol 7:59-63
116. Wandall JH, Bendtzen K, Valerius NH, Nielsen AM, Hax- holdt H (1986) Serum concentrations of interleukin-1 in ulcerative colitis (UC) and in Crohn's disease (CD) (ab- stract). Scand J Gastroenterol [Suppl] 21:41
117. Sartor RB, Chapman EJ, Schwab JH (1988) Increased interleukin-lbeta concentrations in resected inflammatory bowel disease (IBD) tissue (abstract). Gastroenterology 94:A399
118. Ligumsky M, Simon PL, Karmeli F, Rachmilewitz D (1988) Interleukin-1 -possible mediator of the inflamma- tory response in ulcerative colitis (UC) (abstract). Gastro- enterology 94:A263
119. Snyderman R, Pike MC (1984) Chemoattractant receptors on phagocytic cells. Annu Rev Immunol 2:257-281
120. Nast CC, LeDuc LE (1988) Chemotactic peptides. Mecha- nisms, functions, and possible role in inflammatory bowel disease. Dig Dis Sci [Suppl] 33: 50S-57S
121. Marasco WA, Phan SH, Krutzsch H, Showell HJ, Feltner DE, Nairn R, Becker EL, Ward PA (1984) Purification and identification of formyl-methionyl-leucyl-phenylalan- ine as the major peptide neutrophil chemotactic factor produced by Escherichia coli. J Biol Chem 259:5430 5439
122. Bokoch GM, Reed PW (1980) Stimulation of arachidonic acid metabolism in the polymorphonuclear leukocyte by an N-formylated peptide. Comparison with ionophore A23187. J Biol Chem 255:10223-10226
123. Zipser RD, Patterson JB, LeDuc LE (1987) Chemotactic peptide stimulation of leukotrienes from healthy and in- flamed rabbit colons. J Pharmacol Exp Ther 241:218 222
124. Chester JF, Ross JS, Malt RA, Weitzman SA (1985) Acute colitis produced by chemotactic peptides in rats and mice. Am J Pathol 121:282-290
125. Foreman JC (1987) Peptides and neurogenic inflamma- tion. Br Med Bull 43:386-400
126. Shanahan F, Anton P (1988) Neuroendocrine modulation of the immune system. Possible implications for inflamma- tory bowel disease. Dig Dis Sci [Suppl] 33:41S 49S
127. Mayer EM, Raybould H, Koelbel C (1988) Neuropepti- des, inflammation, and motility. Dig Dis Sci [Suppl] 33:71S-77S
128. Pernow B (1983) Substance P. Pharmacol Rev 35: 85-141 129. Barth6 L, Holzer P (1985) Search for a physiological role
of substance P in gastrointestinal motility. Neuroscience 16:1 32
130. Lotz M, Vaughan JH, Carson DA (1988) Effect of neuro- peptides on production of inflammatory cytokines by hu- man monocytes. Science 241:1218 - 1221
131. Koch TR, Carney JA, Go VLW (1987) Distribution and quantitation of gut neuropeptides in normal intestine and inflammatory bowel diseases. Dig Dis Sci 32:369 376
132. Mantyh CR, Gates TS, Zimmerman RP, Welton ML, Pas- saro EP, Vigna SR, Maggio JE, Kruger L, Mantyh PW (1988) Receptor binding sites for substance P, but not substance K or neuromedin K, are expressed in high con- centrations by arterioles, venules, and lymph nodules in surgical specimens obtained from patients with ulcerative colitis and Crohn's disease. Proc Natl Acad Sci USA 85:3235 -3239
133. Grisham MB, Granger DN (1988) Neutrophil-mediated mucosal injury. Role of reactive oxygen metabolites. Dig Dis Sci [Suppl] 33:6S-15S
134. Fantone JC, Ward PA (1982) Role of oxygen-derived free radicals and metabolites in leukocyte-dependent inflam- matory reactions. Am J Pathol 107:397-418
135. Blake DR, Allen RE, Lunec J (1987) Free radicals in bio- logical systems - a review orientated to inflammatory processes. Br Med Bull 43:371 385
136. McCord JM (1985) Oxygen-derived free radicals in post- ischemic tissue injury. N Engl J Med 312:159-163
137. Birnboim HC, Kanabus-Kaminska M (1985) The produc- tion of DNA strand breaks in human leukocytes by super- oxide may involve a metabolic process. Proc Natl Acad Sci 82: 6820- 6824
138. Esterbauer H (1986) Lipid peroxidation products: forma- tion, chemical properties and biological activities. In: Poli G, Cheeseman KH, Dianzani MU, Slater TF (eds) Free radicals in liver injury. IRL Press, Oxford, pp 29-45
139. Freeman BA, Crapo JD (1982) Biology of disease: free radicals and tissue injury. Lab Invest 47:412-426

89
140. Granger DN, Rutili G, McCord JM (1981) Superoxide radicals in feline intestinal ischemia. Gastroenterology 81:22 29
141. Parks DA, Granger DN (1983) Ischemia-induced vascular changes: role of xanthine oxidase and hydroxyl radicals. Am J Physiol 245:G285-G289
142. Riis P (1980) A critical survey of controlled studies in the treatment of ulcerative colitis and Crohn's disease. Clin Gastroenterol 9:351-369
143. Parillo JE, Fauci AS (1979) Mechanisms of glucocorticoid action on immune processes. Annu Rev Pharmacol Toxi- col 19:179-201
144. Scheimer RP (1985) The mechanisms of antiinflammatory steroid action in allergic diseases. Annu Rev Pharmacol Toxicol 25:381 412
145. Vane JR (1971) Inhibition of prostaglandin synthesis as a mechanism of action for aspirin-like drugs. Nature New Biol 231:232-235
146. Lewis GP, Piper PJ (1975) Inhibition of release of prostag- landins as an explanation of some of the actions of antiin- flammatory corticosteroids. Nature 254:308-311
147. Blackwell GJ, Carnuccio R, DiRosa M, Flower RJ, Pa- rente L, Persico P (1980) Macrocortin: a polypeptide caus- ing the anti-phospholipase effect of glucocorticoids. Na- ture 287:147 149
148. Hirata F (1981) The regulation of lipomodulin, a phospho- lipase inhibitory protein in rabbit neutrophils by phospho- rylation. J Biol Chem 256:7730-7733
149. Cloix JF, Colard O, Rothhut B, Russo-Marie F (1983) Characterization and partial purification of 'renocortins': two polypeptides formed in renal cells causing the anti- phospholipase-like action of glucocorticoids. Br J Pharma- col 79:313-321
150. DiRosa M, Flower RJ, Hirata F, Parente L, Russo-Marie F (1984) Nomenclature announcement. Anti-phospho- lipase proteins. Prostaglandins 28:441 442
151. Wallner BP, Mattaliano RJ, Hession C, Cate RL, Tizard R, Sinclair LK, Foeller C, Chow EP, Browing JL, Rama- chandran KL, Pepinsky RB (1986) Cloning and expression of human lipocortin, a phospholipase A 2 inhibitor with potential anti-inflammatory activity. Nature 320:77-81
152. Pepinsky RB, Sinclair LK, Browning JL, Mattaliano RJ, Smart JE, Chow EP, Falbel T, Ribolini A, Garwin JL, Wallner BP (1986) Purification and partial sequence analy- sis of a 37-kDa protein that inhibits phospholipase A z activity from rat peritoneal exudates. J Biol Chem 261:4239-4246
153. Hawkey CJ, Truelove SC (1981) Effect ofprednisolone on prostaglandin synthesis by rectal mucosa in ulcerative coli- tis: investigations by laminar flow bioassay and radioim- munoassay. Gut 22:190-193
154. Hoult JRS, Berry CN, Timms E (1985) Failure of anti- inflammatory steroids to inhibit prostaglandin release from the hydronephrotic rabbit kidney. Agents Actions 17: 304- 307
155. Hales CA, Brandstetter RD, Neely CF, Peterson MB, Kong D, Wattkins WD (1986) Methylprednisolone on cir- culating eicosanoid and vasomotor tone after endotoxin. J Appl Physiol 61 : 185-191
156. Nfiray-Fejes-T6th A, Fejes-T6th G, Fischer C, Fr61ich JC (1984) Effect of dexamethasone on in vivo prostanoid pro- duction in the rabbit. J Clin Invest 74:120-123
157. Rosenkrantz B, Nfiray-Fejes-T6th A, Fejes-T6th G, Fischer C, Sawada M, Fr61ich JC (1985) Dexamethasone effect on prostanoid formation in healthy man. Clin Sci 68:681 685
158. Robert A (1979) Cytoprotection by prostaglandins. Gas- troenterology 77:761-767
159. Hoult JRS, Moore PK (1978) Sulphasalazine is a potent inhibitor of prostaglandin 15-hydroxydehydrogenase: pos- sible basis for therapeutic action in ulcerative colitis. Br J Pharmacol 64:6-8
160. Hoult JRS, Moore PK (1980) Effects of sulphasalazine and its metabolites on prostaglandin synthesis, inactiva- tion and actions on smooth muscle. Br J Pharmacol 68:719 730
161. Hillier K, Mason PJ, Pacheco S, Smith CL (1982) Ulcera- tive colitis: effect of sulphasalazine, its metabolites and indomethacin on the ability of human colonic mucosa to metabolize prostaglandins in vitro. Br J Pharmacol 76:157-161
162. Berry CN, H o u r JRS, Peers SH, Agback H (1983) Inhibi- tion of prostaglandin 15-hydroxydehydrogenase by sul- phasalazine and a novel series of potent analogs. Biochem Pharmacol 32:2863-2871
163. Hoult JRS, Page H (1981) 5-Aminosalicylic acid, a co- factor for colonic prostacyclin synthesis? Lancet ii:255
164. Stenson WF, Lobos E (1983) Inhibition ofplatelet throm- boxane synthetase by sulfasalazine. Biochem Pharmacol 32:2205 2209
165. Hawkey CJ, Boughton-Smith NH, Whittle BJR (1985) Modulation of human colonic arachidonic acid metabo- lism by sulfasalazine. Dig Dis Sci 30:1161 - 1 1 6 5
166. Stenson WF, Lobos E (1982) Sulfasalazine inhibits the synthesis of chemotactic lipids by neutrophils. J Clin Invest 69:496-497
167. Sircar JC, Schweuder CF, Carthers ME (1983) Inhibition of soybean lipoxygenase by sulfasalazine and 5-amino- salicylic acid: a possible mode of action in ulcerative colitis. Biochem Pharmacol 32:170-172
168. Allgayer H, Eisenburg J, Paumgartner G (1984) Soy- bean lipoxygenase inhibition: studies with sulphasalazine metabolites N-acetylaminosalicylic acid, 5-aminosalicylic acid and sulphapyridine. Eur J Clin Pharmacol 26: 449 451
169. Nielsen OH, Bukhave K, Elmgreen J, Ahnfelt-Ronne I (1987) Inhibition of 5-1ipoxygenase pathway of arachido- nic acid metabolism in human neutrophils by sulfasalazine and 5-aminosalicylic acid. Dig Dis Sci 32:577-582
170. Rubinstein A, Das KM, Melamed J, Murphy RA (1978) Comparative analysis of systemic immunological parame- ters in ulcerative colitis and idiopathic proctitis: effects of sulphasalazine in vivo and in vitro. Clin Exp Immunol 33:217-224
171. Molin L, Stendahl O (1979) The effect of sulfasalazine and its active components on human polymorphonuclear leu- kocyte function in relation to ulcerative colitis. Acta Med Scand 206:451 457
172. Rhodes JM, Bartholomew TC, Jewell DP (1981) Inhibi- tion of leucocyte motility by drugs used in ulcerative coli- tis. Gut 22:642-647
173. MacDermott RP, Kane MG, Steele LL, Stenson WF (1986) Inhibition of cytotoxicity by sulfasalazine. I. Sulfa- salazine inhibits spontaneous cell-mediated cytotoxicity by peripheral blood and intestinal mononuclear cells from control and inflammatory bowel disease patients. Immu- nopharmacology 11:101 109
174. Stenson WF, Mehta J, Spilberg I (1984) Sulfasalazine inhi- bition of binding of N-formyl-methionyl-leucyl-phenyl- alanine (FMLP) to its receptor on human neutrophils. Biochem Pharmacol 33:407-412

90
175. Baum CL, Selhub J, Rosenberg IH (1981) Antifolate ac- tions of sulfasalazine on intact lymphocytes. J Lab Clin Med 97:779 784
176. Das KM, Eastwood MA, McManus JPA, Sircus W (1973) The metabolism of salicyl-azosulphapyridine in ulcerative colitis. I. The relationship between metabolites and the response to treatment in inpatients. II. The relationship between metabolites and the progress of the disease studied in outpatients. Gut 14:631 641
177. Azad Khan AK, Piris J, Truelove SC (1977) An experi- ment to determine the active therapeutic moiety of sulpha- salazine. Lancet ii:892 895
178. van Hees PAM, Bakker JH, van Tongeren JHM (1980) Effect of sulphapyridine, 5-aminosalicylic acid, and pla- cebo in patients with idiopathic proctitis: a study to deter- mine the active therapeutic moiety of sulphasalazine. Gut 21:632-635
179. Campieri M, Lanfranchi GA, Bazzocchi G, Brignola C, Sarti F, Franzin G, Battocchia A, Labo G, DalMonte PR (1981) Treatment of ulcerative colitis with high-dose 5-aminosalicylic acid enemas. Lancet ii:270-271
180. Danish 5-ASA Group (1987) Topical 5-aminosalicylic acid versus prednisolone in ulcerative proctosigmoiditis. A ran- domized, double-blind multicenter trial. Dig Dis Sci 32:598 602
181. Azad Khan AK, Truelove SC (1980) Circulating levels of sulphasalazine and its metabolites and their relation to the clinical efficacy of the drug in ulcerative colitis. Gut 21:706 710
182. Lauritsen K, Hansen J, Ryde M, Rask-Madsen J (1984) Colonic azodisalicylate metabolism determined by in vivo dialysis in healthy volunteers and patients with ulcerative colitis. Gastroenterology 86:1496-1500
183. Hour JRS, Moore PK (1983) Prostaglandins and the mo- de of action of sulphasalazine in ulcerative colitis: two opposing viewpoints. In: Rachmilewitz D (ed) Inflamma- tory bowel diseases. Martinus Nijhoff, The Hague, pp 174-189
184. Schlenker T, Peskar BM (1981) Dual effect of sulphasala- zine on colonic prostaglandin synthetase. Lancet ii: 815
185. Gould SR, Brash AR, Conolly ME, Lennard-Jones JE (1981) Studies of prostaglandins and sulphasalazine in ul- cerative colitis. Prostaglandins Med 6:165 182
186. Hawkey CJ, Truelove SC (1983) Inhibition of prostaglan- din synthetase in human rectal mucosa. Gut 24:213-217
187. Campieri M, Lanfranchi GA, Bazzocchi G, Brignola C, Corazza G, Cortini C, Michelini M, Labo G (1978) Salicy- late other than 5-aminosalicylic acid ineffective in ulcerati- ve colitis. Lancet ii:993
188. Marcinkiewicz E, Duniec Z, Robak J (1985) Salazosulfa- pyridine and non-steroidal anti-inflammatory drugs do not inhibit soy-bean lipoxygenase. Biochem Pharmacol 34:148 149
189. Craven PA, Pfanstiel J, Saito R, DeRubertis FR (1987) Actions of sulfasalazine and 5-aminosalicylic acid as reac- tive oxygen scavengers in the suppression of bile acid- induced increases in colonic epithelial cell loss and prolife- rative activity. Gastroenterology 92:1998 2008
190. Dull BJ, Salata K, Langehove A, Goldman P (1987) 5-Aminosalicylate: oxidation by activated leukocytes and protection of cultured cells from oxidative damage. Bio- chem Pharmacol 36:2467-2472
191. Miyachi Y, Yoshioka A, Imamura S, Niwa Y (1987) Effect of sulphasalazine and its metabolites on the generation of reactive oxygen species. Gut 28:190-195
192. Ahnfelt-Ronne I, Nielsen OH (1987) The anti-inflam- matory moiety of sulfasalazine, 5-aminosalicylic acid, is a radical scavenger. Agents Actions 21:191 194
Dr. J. Rask-Madsen Section of Gastroenterology Department of Medicine G Bispebjerg Hospital DK-2400 Copenhagen NV Denmark