Embed Size (px)
Citation preview

INDUCED HEREDITARY CHANGE IN DROSOPHILA BY INJECTED EXTRACTS OF NEOPLASTIC FLIES, MICE, OR HUMANS
Morris Henry Harnly Craduate School of Arts and Science, Nev York University, Nev York, N.Y.
Introduction Purified extracts were prepared from (1) tissues of tu-e Drosophila larvae (a
spontaneous hereditary melanotic tumor strain) ; (2) tissues of spontaneous leukemic AKR and virus induced leukemic C3H(f)* mice; and (3) serum of Hodgkin's disease patients (extraction procedure: Burton and Friedman, 1956; Burton et al., 1959; Kaplan et al., in press). Each of these extracts was injected into 96-hour larvae of the tumor-free (less than 1 'tumorous larva/250,000 larvae) wild 51-52 strain of D. melanogaster. The emerged adult hosts having melanotic tumors in each experiment were mated. Although no tumors were observed in any of the three F1 progenies, these FI adults were randomly mated within each of the test lines. In the Fz of each test, approximately 1 percent of the adults showed melanotic tumors. Those showing tumors within each line were isolated and randomly mated ( i e . nonvirgin) within the line. In the Fa of each line the incidence of adults with melanotic tumors increased to ap- proximately 25 per cent. This higher incidence permitted the selection and mating of tumorous males with tumorous virgin females. Thereafter virgin sib matings of tumorous flies were made for 50+ generations, and the tumor inci- dence rose to and is a t present 90+ per cent. Each of these 3 induced melanotic tumor lines has been kept isolated and maintained as an individual line: ft, an induced tumor line by Drosophda tu-e extract; mt, an induced tumor line by mouse extract; and ht, an induced tumor line by human extract. Since each of these 3 lines produces regularly ninety-odd per cent melanotic tumorous off- spring in each generation, apparently an hereditary change has been induced by the single specific injection in the PI generation; then the 90-odd per cent value would be the penetrance (the capacity of the hereditary condition to express itself) of this hereditary condition irrespective of whether the matings have been made between tumorous or nontumorous individuals within the hereditary line. This raises the question of the method of inheritance in each of these injection-induced hereditary tumor lines. The experiments reported here are an analysis of the method of inheritance in the human extract-induced tumor strain.
Materials and Methods All animals were reared in half-pint milk bottles on the standard corn meal,
molasses, and brewers' yeast medium at 25' C. The genetic strains of D. melanogaster used to determine the method of in-
heritance were the following: (1) the human extract-induced tumor line (sym- bol ht); (2) the mouse extract-induced tumor line (symbol mt); (3) the fly ex- tract-induced tumor line (symbol ft) ; (4) the nontumorous wild type 51-52 strain, a genotype developed years ago from the original Oregon-wild by special
* C3H(f) are foster-nursed strain and presumably free of the milk agent.
817
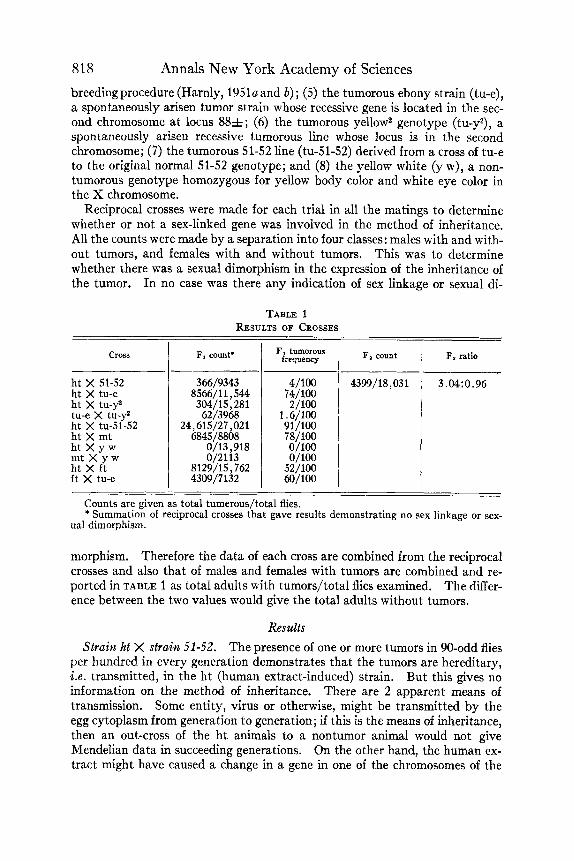
818
h t X 51-52 h t X tu-e ht X tu-y' tu-e X tu-y* ht X tu-51-52 ht X mt h t X y w mt X y w ht X ft ft X tu-e
Annals New York Academy of Sciences
- 366/9343 4/100 4399/18,031 3.04:0.96
8566/11,544 74/100 304/15,281 2/100 62/3968 1.6/100
24,615/27,021 91/100 6845/8808 78/100
0/13,918 0/100 0/2113 0/100
8129/15,762 52/100 4309/1132 60/100
breeding procedure (Hardy, 1951aand b) ; (5) the tumorous ebony strain (tu-e), a spontaneously arisen tumor strain whose recessive gene is located in the sec- ond chromosome at locus 8 8 f ; (6) the tumorous yellow2 genotype (tu-yz), a spontaneously arisen recessive tumorous line whose locus is in the second chromosome; (7) the tumorous 51-52 line (tu-51-52) derived from a cross of tu-e to the original normal 51-52 genotype; and (8) the yellow white (y w), a non- tumorous genotype homozygous for yellow body color and white eye color in the X chromosome.
Reciprocal crosses were made for each trial in all the matings to determine whether or not a sex-linked gene was involved in the method of inheritance. All the counts were made by a separation into four classes: males with and with- out tumors, and females with and without tumors. This was to determine whether there was a sexual dimorphism in the expression of the inheritance of the tumor. In no case was there any indication of sex linkage or sexual di-
TABLE 1 RESULTS OF CROSSES
cross F, count" 1 ~j~~~~~ I F, count 1 F, ratio I
morphism. Therefore the data of each cross are combined from the reciprocal crosses and also that of males and females with tumors are combined and re- ported in TABLE 1 as total adults with tumors/total flies examined. The differ- ence between the two values would give the total adults without tumors.
Results Strain ht X strain 51-52. The presence of one or more tumors in 90-odd flies
per hundred in every generation demonstrates that the tumors are hereditary, i.e. transmitted, in the ht (human extract-induced) strain. But this gives no information on the method of inheritance. There are 2 apparent means of transmission. Some entity, virus or otherwise, might be transmitted by the egg cytoplasm from generation to generation; if this is the means of inheritance, then an out-cross of the ht animals to a nontumor animal would not give Mendelian data in succeeding generations. On the other hand, the human ex- tract might have caused a change in a gene in one of the chromosomes of the

Harnly : Induced Hereditary Change 819 injected animals; in this case, an out-cross of the ht flies to a nontumorous line would give a Mendelian ratio in the F2 generation.
In the F1 generation 9343 adults were examined and 366 of these had one or more tumors. This demonstrates that the ht condition is due to a recessive gene that can break through (penetrate) in the heterozygous condition with 51-52 in a frequency of 4/100 animals. The F1 males and females were mated with each other and a total of 18,031 Fz adults were examined. Melanotic tumors were observed in 4399 of the 18,031 flies, 13,632 being nontumorous. The ratio is 3.04 nontumorous to 0.96 tumorous. This is a typical Mendelian monohybrid ratio. Since the reciprocal crosses show it is not sex-linked, this demonstrates that the ht condition is due to a recessive gene carried in one of the 3 pairs of autosomes.
Strain ht X strain tu-e. The ht flies were crossed to tu-e flies. In the F1 generation 11,544 adults were examined and of these 8566 had one or more tumors. The frequency of 74 tumorous/100 imagos indicates that the ht genes and the tu-e gene are alleles, which means that the human extract had caused a genetic change of the normal allele at the tu-e locus in the second chromosome when injected into the normal 51-52 larvae. This raises the question of whether there is more than 1 tumor-producing locus in the second chromosome; if there is, then the extract was highly selective in its action. The tumors of the tu-y2 (spontaneous mutant) are due to a recessive gene in the second chromosome (Benjaminson, 1961) and make possible an experimentally determined answer to this question.
Strain ht X strain tu-y2. The ht flies were crossed with the twy2 flies, and in the F1 generation 15,281 imagos were examined. Of these, 304 had 1 or more tumors, This is a frequency of 2 tumorous/100 adults and demonstrates that the ht gene is not an allele of the tu-y2 gene. The 2/100 tumorous animals would result from the slightly incomplete dominance of the normal ht gene in- troduced by the tu-yz parent and the slightly incomplete dominance of the nor- mal tu-yz gene introduced by the ht parent. This indicates the influence of 2 different loci in the second chromosome. A cross between the 2 spontaneously arisen tumor strains in the second chromosome would determine whether 1 locus or 2 different loci in the second chromosome were involved.
Strain tu-e was crossed to tu-y2 and 3968 F1 adults were examined. Of these only 62 were tumor-bearing, a frequency of 1.6 tumorous/100 adults. This demonstrates that there are at least 2 different normal loci in the second chromosome, a spontaneous mutation of either one producing melanotic tumors (there are other known tumor-producing loci in the chromosomes of D. melanogaster in addition to these 2 loci). In the data reported above i t has been demonstrated that the ht gene is an allele of the tu-e gene but not an allele of the tu-y2 gene, and that the spontaneous mutants tu-e and tu-y2 in the second chromosome are not alleles of each other. The results answer the original question and demonstrate that the human extract was highly selective in the locus it affected. Evidence of the same selectivity of the other 2 extracts for the tu-e locus will be presented below.
In connection with other experiments the tu-e
To decide this point, h t was crossed to the nontumorous 51-52 (TABLE 1).
Strain tu-e X strain tu-yz.
Strain ht X strain tu-51-52.

820 Annals New York Academy of Sciences flies had been crossed to normal 51-52, and through breeding and selection the tumor gene of the tu-e had been incorporated into the 51-52, producing the tu-51-52 tumorous line. It was decided to test the ht gene heterozygous with the original tu-e gene but in a double dose of other 51-52 genes. The ht animals carrying all of the 51-52 genes except a t the tumor locus were crossed to this tu-51-52 strain. In the F1 generation 27,021 adults were examined and of these 24,615 had one or more tumors. This is a frequency of 91 tumorous/100 adults and the highest frequency in the 5 crosses reported here in which there was an heterozygous condition for 2 different tumor-producing alleles at the tu-e locus. The original 51-52 strain has proved to be unique in its lability in our work on hereditary tumors in D. melanogaster. Extracts of malignant tissue from hu- man, mouse, chicken, fish, fly, clover leaf tumor, and crown gall bacteria all produce tumorous hosts when injected into normal 51-52 larvae.
The ht flies were crossed to the mt individuals and 8808 F1 adults were examined. Of these 6845 had one or more tumors. This is a frequency of 78 tumorous/100 adults and would indicate that either the ht and mt genes are alleles or the mt gene is a dominant. The following two crosses were made to determine the correct alternative.
Strain ht x strain y w. They w is a nontumorous strain carrying yellow body color and white eye color in the X chromosome. The ht flies were crossed to the y w individuals and 13,918 FI adults were examined for tumors. There were no tumorous adults among all these F1 imagos, emphasizing two points: (1) ht is a recessive gene; and (2) the 51-52 strain is unique, since in the cross of ht by 51-52 the F1 showed a 4/100 frequency in a count of 9343 adults.
Strain mt x strain y w. The cross mt x y w was made and 2113 F1 adults were examined. Not one of them had a tumor. This demonstrates that mt is a recessive, and since the ht X mt cross gave a frequency of 78 tumorous/100 adults, the genes ht and mt must be alleles.
Strain ht X strain f t . The human extract-induced mutant tumor flies were crossed with the fly extract-induced mutant tumor flies and 15,762 F1 adults were examined. Of these, 8129 had one or more tumors, a frequency of 52 tumorous/100 adults. Since ft is known to be a recessive from earlier work and ht has already been proved to be a recessive, this data demonstrates that ht and ft are allelic genes.
Strain ft X strain tu-e. The cross ft X tu-e was made and 7132 FI adults were examined. Of these, 4309 were tumorous, giving a frequency of 60 tu- morous/100 adults. This frequency in the F1 generation demonstrates that the f t gene is behaving as an allele of the tu-e gene. From the data presented here and above, 6 conclusions follow: (1) the induced mutant genes, ht, mt, and ft are all recessives; (2) these 3 genes are alleles of each other; (3) 2 of the 3 have been demonstrated to be alleles of the spontaneously arisen mutant gene tu-e; (4) all 3 extracts (human, mouse, and fly) were selective for, and active in, in- ducing mutations only at the tu-e locus of all the possible tumor loci in the nor- mal 51-52 larvae; (5) all 3 extracts, 2 from different vertebrate sources and 1 from an invertebrate, produced the same general effect a t the same locus, namely, a change from a dominant normal allele to a recessive tumor-producing allele; and (6) the differences in the F1 heterozygous tumor frequencies of
Strain ht x strain mt.

Harnly : Induced Hereditary Change 82 1 52/100, 60/100, 74/100, 78/100, and 91/100 (TABLE 1) demonstrates that the ht, mt, ft, and tu-e genes are not the same gene, but are alleles of each other.
Discussion Previous work has established that only extracts from tumorous larvae will
induce melanotic tumors in the host. Extracts prepared from the wild 51-52 nontumorous strain injected into larvae of the nontumorous yellow white strain resulted in nontumorous adult hosts. Similarly the extract from the non- tumorous yellow white strain injected into the wild 51-52 nontumorous strain larvae resulted in tumor-free adult hosts (Harnly et al., 1954). Also Wadding- ton’s insect Ringer’s solution was injected, as a control material, into the wild 51-52 strain of nontumorous larvae. Following injection of this solution, none of the 357 surviving hosts developed melanotic tumors. A histological survey of Drosaphila that had been “injured” or injected with this salt solution demonstrated that the spindle cells (basic component of the tumors) were not present in the test animals. This indicated that the injection of tumor factor was necessary for the appearance of the spindle cells and tumor formation in the host animals (Burton et al., 1956). On the other hand, melanotic inclusions may appear in larvae from improper injection techniques, i.s., faulty needles, injury to internal larval organs, and melanization of chitinous exoskeleton pushed into the body cavity. Histological examination of these artifacts dem- onstrates that they are nontumorous in structure. Such artifacts, as well as the external melanized injection scar, are larval characters not usually found in the emerged adult.
In the work reported above to determine the mode of inheritance of the hu- man extract-induced tumor strain (ht) of D. melanogaster, 10 different genetic crosses were made. Six of these were with known second chromosome loci. Approximately 132,500 animals were examined for the presence of melanotic tumors. The results constitute a thorough and adequate, in terms of numbers, analysis of the mode of inheritance of this tumor. The cross of ht with normal 51-52 produced in the Fz a ratio of 3.04 normal to 0.96 tumorous in a count of over 18,000 flies. This is a typical Mendelian monohybrid ratio and critically proves that the ht tumor is due to a single recessive gene. The ht cross to tu-e producing 74 tumorous/100 adults in an FI count of over 11,500 animals proves that the ht gene is an allele of the tu-e gene and is carried at the locus (88rt) in the second chromosome. The tu-e gene was a spontaneous mutant in the second chromosome at locus 8 8 f . The cross of ht with tu-51-52 produced 91 tumorous/100 F1 adults in a count of about 27,000 imagos and is further critical proof that h t and tu-e are allelic genes, both being recessives. These 3 crosses prove that the human extract-induced hereditary tumor ht is due to a single recessive gene, ht, carried at locus 88=t in the second chromosome. The ht X tu-y2 cross giving a 2 tumorous/100 F1 adults count in nearly 15,300 flies proves that ht is not an allele of tu-y2, another recessive, spontaneously mutated, tumor-producing gene in the second chromosome. Therefore, the extract in- jected was highly selective in the locus at which i t induced a mutation. The crosses ht x y w and mt X y w produced no tumorous F1 animals in a total count of some 16,000 animals, proving that nit as well as ht is a recessive pheno-

822 Annals New York Academy of Sciences type. The mt tumor is an hereditary tumor induced by the mouse extract. The h t X mt cross gave a frequency of 78 tumorous/100 in an F1 count of 8800 adults. This demonstrates that the mt tumor is caused by a recessive allele of the ht gene and is also carried a t locus 8 8 f in the second chromosome. Finally the crosses ht X ft with an F1 tumor frequency of 52/100 in count of over 15,700 and ft X tu-e with an F1 tumor frequency of 60/100 in count of over 7100 demonstrate that the fly extract-induced hereditary tumor ft is also due to a recessive allele a t locus 8 8 f in the second chromosome. These crosses prove that the human, mouse, and fly extracts have each induced a recessive mutation a t the same locus in the second chromosome causing the same melano- tic tumor phenotype.
Since all three extracts induced a recessive mutation for the same melanotic tumors a t the same locus in chromosome (2 of all the possible tumor-produc- ing loci in D. melanogaster), these extracts were highly selective and specific in their action. This would indicate that these three extracts contain the same tumor-inducing and tumor-mutation-inducing entity.
Any interpretation of the above facts should be based also on all the other known facts regarding these extracts:
(1) Human tissue extracts of Hodgkin’s disease, lymphosarcoma, leukemic tissue, lung carcinoma, ovarian carcinoma, clear-cell kidney tumor, myxoma, and mammary carcinoma injected into normal 51-52 D. melanogaster larvae cause the typical Drosophila melanotic tumors in the hosts.
(2) Similarly, human serum extracts of all of the above and also melanoma induce melanotic tumors in the 51-52 hosts.
(3) Mouse leukemic tissue extract injected into 51-52 larvae gave the typical Drosophila tumors but when injected into the malignancy free C3H(f) mice, produced typical parotid tumors. These induced mouse parotid tumors were extracted, and the extract injected into 51-52 larvae and into the C3H(f) mice and again produced melanotic tumors in the Drosophila hosts and parotid tumors in the mice hosts.
(4) Mouse serum extracts of spontaneous and induced leukemia, spontaneous and induced mammary carcinoma, and induced fibrosarcoma injected into the 51-52 larvae produced the typical melanotic tumors in the hosts.
(5) The mouse hepatitis extract was injected into normal 51-52 larvae and allowed an incubation period of 18 to 24 hours, re-extracted, diluted, and in- jected in new 51-52 larvae for 4 passages in normal 51-52 larvae and then the final extract injected into the malignancy-free strain of mice, with the conse- quent induction of hepatitis in these host mice. In 4 larval passages the dilu- tion factor becomes so great (dilution factor greater than that the original entity injected would have disappeared; hence either this entity could reproduce itself in the D. melanogaster larvae or induce them to produce it.
(6) Chickentissue extracts of Rous sarcoma, erythroblastosis, lymphomatosis, and kidney tumor injected into 51-52 larvae caused the typical Droso$hiZa tumors.
(7) The fish hybrid swordtail melanoma material* injected into 51-52 larvae
* Obtained from the Genetics Laboratory of the New York Aquarium.

Harnly : Induced Hereditary Change 82 3 induced the typical Drosophila tumors (Sandra Kassel, personal communica- tion).
(8) The tu-e larval extract injected into the normal 51-52 larvae produces the typical Drosophila melanotic tumors in the hosts, but injected into the malignancy-free C3H(f) strain of mice produces in the hosts the typical mouse neoplasms, i.e. parotid and mammary carcinomas and fibrosarcomas (Kassel el al., 1961).
(9) The crown gall bacteria extract produces the typical melanotic tumors in the 51-52 hosts.
(10) The clover leaf tumor extract injected into 51-52 larvae induces the usual melanotic tumors.
Here we have one common extraction procedure but a number of different sources for the extract: 2 vertebrate mammalian sources, human and mouse; 1 vertebrate Aves source, chicken; 1 vertebrate Pisces; 1 invertebrate, Insecta; and two sources from the plant kingdom, plant crown gall bacteria and clover leaf. Extracts from each of these 7 phylogenetic sources injected into D. melanogastw normal 5 1-52 larvae produces the same effect, namely, the typical melanotic tumor of Drosophila; but injected into mice, each produces typical mouse malignancies. Irrespective of whether or not the extracts from different sources contain the same entity, the type of response is, in terms of structure produced, etc., species specific. Furthermore a vertebrate mouse entity (from hepatitis) may be passed four times through the invertebrate Drosophila larvae and then be returned to normal mice and produce the original typical mouse response (hepatitis). In a current histological and pathological study of in- duced and spontaneously arisen melanomas in D. melanogaster, A. Rottino of St. Vincent’s Hospital, New York, N.Y., has found that the tumors of the ht , mt, and the spontaneous tu-e are histologically similar. It is evident that this common extraction procedure isolates an entity (or entities) causing a definite response in the injected host, the nature of the response depending not on the source of the extract but upon the host. Finally, three extracts (human, mouse, fly) have caused an hereditary change (mutation) a t the same locus in normal 51-52 larvae, producing in all three cases the same phenotypic change, melanotic tumors.
The work of the past few years with this extraction procedure demonstrates that from a given neoplastic source a standard entity can be extracted that will cause a standard response in a given host. Upon injection into the host it can do two things:
(1) The host, mouse or fly, may develop a neoplasm typical for the specific host.
(2) In the case of normal 51-52 D. melanogaster the human, mouse, and fly extracts are able to induce an hereditary change (mutation) a t the tu-e locus in the second chromosome. The entity injected is highly selective, acting only a t the tu-e locus of all the possible tumor loci. This change is the same for ex- tracts from all three sources, a mutation from a normal nontumorous dominant allele to a recessive allele producing melanotic tumors.
These data from extensive sampling of various types of malignancy from 7 different phylogenetic sources including both the animal and plant kingdoms

824 Annals New York Academy of Sciences lead to certain conclusions: (1) the extracts are repeatable, i.e., the extract from a given phylogenetic source and type of malignancy is constant; (2) the response to this entity is independent of its source and is host species specific, i .e. , de- termined strictly by the hosts genotype; and (3) three of these extracts, from human, mouse, and fly, have been found to have an affinity for the tu-e locus, and the tu-e locus only, in the wild 51-52 strain of I). melunogaster, causing a mutation at that locus in the second chromosome. These facts lead to the con- clusion that the entity extracted is a constant, i.e., the same entity irrespective of its source in the animal or plant kingdom. I t is an entity causing cell multi- plication and abnormal growth. This conclusion-that it is the same entity irrespective of its source-should not be surprising. It is not a unique phenom- enon. For example, ATP is common to the animal and plant kingdom and, irrespective of its animal or plant source, has the same action in a given or- ganism, the typical host’s response or action.
Since the data demonstrate that this entity is capable of inducing a mutation to a recessive tumor-producing gene a t a single specific locus in the D. meluno- guster, the question arises: How might such a mutation be brought about? One may assume that the mutagenic injected material is a chemical entity and that the gene in the chromosome is also a chemical entity. If by chance a molecule of the mutagenic substance gets into a nucleus of a gonadal cell, it might take a position near the tu-e locus of the second chromosome. Then one of two things may happen. At the time the normal tu-e allele is duplicating itself some kind of chemical reaction between the gene and the molecule could occur, causing a failure in the exact duplication and the consequent production of the mutant allele. This would be a chemically induced mutation. Since this oc- curs only at the tu-e locus, the entity and the reaction with the duplicating gene must be highly specific. Irrespective of its source (human, mouse, or fly) we have here a chemical entity specific for only one locus of all the loci in D. melunogaster causing a mutation to a tumor-producing allele. The induced mutant genes ht and mt and the spontaneously mutant gene tu-e all produce histologically similar melanotic tumors. Since the locus affected, the type of mutation produced, and the tumor are the same, then the entity injected would appear to be the same or similar irrespective of its source.
A second way in which such a mutation might be brought about would also be chemical. Assume again that a molecule of the extract entity gets into the nucleus of a gonadal cell and comes near the tu-e locus. The structure of this molecule is such that it has an affinity for the tu-e locus and is incorporated into the second chromosome, replacing the nontumor allele at the tu-e locus. This might occur as the result of the extract molecule having a template relation to the nontumor gene, covering it and being duplicated thereafter instead.
The data presented here and the conclusion drawn from it raises 2 general questions fundamental to the cancer problem. Counting serum and tissue as different sources from the same neoplasm, 17 human extracts, 7 mouse extracts, four chicken extracts, 1 fish extract, 1 insect extract, 1 crown gall extract, and 1 clover leaf tumor extract-a total of 32 extracts from 7 different phylogenetic sources in the animal and plant kingdoms-have been produced by the same extraction procedure. These 32 extracts have been injected into the normal

Harnly : Induced Hereditary Change 82 5 51-52 strain of D . melanogaster. In every case the same typical melanotic tumor was induced in the hosts. Some of these extracts were injected into mice and in the mouse hosts induced typical mouse neoplasms. Since with one common extraction procedure all the diverse forms with pathological conditions tested in the animal and plant kingdoms produced a positive response, there must be a common tumor-inducing denominator in these extracts. This raises the question: Is there some common fundamental entity underlying and in- volved in the development of neoplasms in all species, both animal and plant? We have here demonstrated positive tests on different neoplasms from 7 widely diverse species ranging from man to clover. This would suggest that in the course of time it may be found that there is some common denominator, some common factor in neoplastic development, perhaps the common entity in the various extracts used here.
Three of these extracts induced the same mutation from a dominant normal gene to a recessive tumor gene condition at the tu-e locus in the normal 51-52 strain of D. melanogaster. This has been tried with only 3 of the 32 extracts that produced tumors in the hosts injected and tried in only 1 host genotype, but in all three cases tested (human, mouse, and fly) this mutation was induced. A fourth extract (fish) is now being tested in the normal 51-52 strain. Pre- liminary results indicate that a genetic change has occurred (Sandra Kassel, personal communication). The mutagenic action of this common extract en- tity in the case of this specific locus of the normal 51-52 strain of Drosophila raises the question: Is this mutagenic action or transformation a more or less universal phenomena, both somatically (induction of melanotic tumors) and germinally, and what bearing does i t have on the problem of neoplasia? In a number of forms lower than man the genetic basis of certain malignancies has been conclusively demonstrated. Once a given hereditary neoplasm has orig- inated by spontaneous mutation it will be perpetuated in the direct line of descent. But in terms of this extract entity’s mutagenic action by injection, as demonstrated here, a new situation might arise. If there is any vector by which this entity could be transferred to other genetic lines, then new genetic lines of the same neoplasm might arise by induced mutation or transformation.
Szlmmar y (1) By the same extraction procedure 32 different neoplastic extracts from
seven widely divergent phylogenetic species, including both animals and plants, have been injected into the wild type 51-52 strain of D. melanogaster. All 32 extracts induced melanotic tumors in the hosts. Several of these extracts have been injected into the tumor-free strain C3H (f) mice and produced typical mouse neoplasms in the hosts. The tumor response always reflects host tissue pro- liferation irrespective of the extract source.
(2) The melanotic tumorous Drosophila hosts of three of these extracts (hu- man, mouse, fly) have been bred separately. Each of the three lines, ht, mt, and f t has been found by extensive genetic crosses to carry a mutant gene pro- ducing a melanotic tumor phenotype.
(3) This change is from a normal dominant gene to a recessive gene producing melanotic tumors.

826 Annals New York Academy of Sciences (4) Each of these three extracts has induced a mutation at the same locus
in the right arm of the second chromosome of the wild type 51-52 strain of D. melanogaster. Since only the tu-e locus of all possible tumor producing loci is affected, the extract entity is highly selective and specific.
(5) These are “chemically” induced mutants. All 32 extracts cause the same type of tumor response in a given host, and the 3 that have been tested as mutagenic agents were specific for the same second chromosome locus. It then follows that there must be a common entity in these diverse extracts.
References BENJAMINSON, M. A. 1961. The genetic analysis of an hereditary tumor in the tu-y2 vg
BURTON, L. & F. FRIEDMAN. 1956. The purification of an inherited tumor-inducing factor
BIJRTON. L.. F. FRIEDMAN. R. KASSEL. M. L. KAPLAN & A. ROTTINO. 1959. The Durifica-
strain of Drosophila melanogaster. Masters thesis. N.Y. Univ. New York, N.Y.
in Drosophila melanogaster. Cancer Research. 16: 880-884. ~.
tion and action of tumor factor ‘extracted from mouse and human neoplastir tissue. Trans. N.Y. Acad. Sci. Ser. I1 21(8): 700-707.
1956. The activity of a tumor factor in Dro- BURTON, L.. M. H. HARNLY & M. J. KOPAC. sophila’development. Cancer Research. 16: 402-507.
HARNLY, M. of Drosophila melanogaster. J. Exptl. Zool. 118: 21-36.
HARNLY, M. H. of Drosoplzda melanogaster. J. Exptl. Zool. 118: 37-70.
HARNLY, M. H., L. BURTON, F. FRIEDMAN & M. J. KOPAC. Drosophila melanogaster by injection of e tu larval fluids.
KAPLAN, M. L., R. KASSEL, L. BURTON, F. FRIEDMAN & A. ROTTINO. cinogenic activity of refined tumor factor isolated from human neoplasia.
KASSEL, R. et al. malian neoplasia. In press.
1951a. Size of parts present vs. number of parts present in the development
An optimum-temperature hy othesis for the heat reaction curves
1954. Tumor induction in
1961-1962. Car- In press.
Utilization of induced Drosophila melanoma in the study of mam-
1951b.
Science. la0: 225-227.
1961.
Discussion of the Paper J. COHEN (University of Birmingham, Birmingham, England) : Have you
tried comparable extracts from normal animals for their mutogenic properties? M. H. HARNLY (New York Uniwersity, New York, N.Y.) : This has been tried,
and, as far as I know, was negative. COHEN: In your F-2’s, in the first series, you mentioned that you obtained 1
per cent tumors. Did you mate these animals for your subsequent line, and did you also mate the animals that did not have tumors in that generation?
HARNLY: No, but if they had been mated i t is possible that some might have produced a low percentage of tumors in the F-3 generation and others not, depending on whether there were mutating genes as opposed to nonmutating genes.
SPEAKER FROM THE FLOOR: Since you did not use various normal tissues, what was your control system?
HARNLY: All that I have done is the genetic aspect. R. KASSEL (St. Vincent’s Hospital, New York, N .Y . ) : In the course of a
series of earlier experiments, we injected a variety of materials into drosophilae- protein, enzymes, nucleic acids, carboxypeptidases-a whole variety of differ- ent things, and we never got any tumor induction with them.





























