Embed Size (px)
Citation preview

Advances in Enzyme Regulation 50 (2010) 112–118
Contents lists available at ScienceDirect
Advances in Enzyme Regulationjournal homepage: www.elsevier .com/locate/
advenzreg
Indirect mechanisms of carcinogenesis via downregulationof PTEN function
Nick R. Leslie, Laura Spinelli, Priyanka Tibarewal, Georgios Zilidis,Nimmi Weerasinghe, Joseph C. Lim, Helene Maccario, C. Peter Downes*
Division of Molecular Physiology, College of Life Sciences, University of Dundee, Dundee, DD1 5EH, UK
Introduction
PTEN (EC 3.1.3.67) has been well established as one of the most important tumour suppressors inhuman cancer (Salmena et al., 2008). The PTEN gene appears to be the second most frequently mutatedtumour suppressor gene in human cancers after TP53, with loss of at least one allele occurring in asmany as 30% of sporadic tumours (Keniry and Parsons, 2008). Patients inheriting one mutant PTENallele display a rather diverse set of symptoms, usually including multiple benign intestinal polyps, orhamartomas, and suffer from an increased risk of cancers, especially of the breast, endometrium andthyroid (Eng, 2003). Similarly, mice with one wild-type and one null Pten allele succumb to a broadrange of tumour types, and tissue specific deletion of both Pten alleles in mice leads to the formation ofaggressive tumours in most tissues analysed (Suzuki et al., 2008). Despite this ability to drive tumourinitiation in many mouse tissues, in most human tumour types, loss of PTEN appears to be a relativelylate event in tumour progression (Keniry and Parsons, 2008). There are however, notable exceptions tothis, such as endometrial carcinoma, in which PTEN loss is a very early event occurring in the majorityof these tumours (Levine et al., 1998; Maxwell et al., 1998; Mutter et al., 2000).
An important aspect to PTEN tumour biology is that, at least in the tissues studied, PTEN appears toact as a haploinsufficient tumour suppressor, such that loss of one allele greatly increases tumouroccurrence (Salmena et al., 2008; Trotman et al., 2003). A logical speculation based upon these data isthat other mechanisms that lead to the partial suppression of PTEN expression or activity may also playimportant roles in tumour formation. Thus, a potential result of PTEN’s dominant role in controllingtumour formation is that changes to other proteins in tumours that affect PTEN expression or activityindirectly might also drive tumorigenesis. Recent data regarding several novel PTEN interactingproteins strongly support this hypothesis and will be dealt with in more detail here.
The PTEN protein is a lipid phosphatase that controls phosphoinositide 3-kinase (PI3K EC 2.7.1.153)dependent signalling through dephosphorylation of the lipid second messenger PIP3 (Leslie and
* Corresponding author. Tel.: þ44 1382 385156.E-mail address: [email protected] (C.P. Downes).
0065-2571/$ – see front matter � 2009 Elsevier Ltd. All rights reserved.doi:10.1016/j.advenzreg.2009.10.015

N.R. Leslie et al. / Advances in Enzyme Regulation 50 (2010) 112–118 113
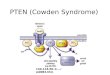
Downes, 2002). In this way, PTEN has been shown to control cellular processes including growth,survival and migration that are believed to be the major contributors to its tumour suppressor action(Keniry and Parsons, 2008; Leslie and Downes, 2002; Salmena et al., 2008). Over the last few yearsa variety of proteins have been identified that appear to interact with PTEN and regulate its functionby different mechanisms, such as controlling its phosphorylation (Okahara et al., 2004), ubiquitina-tion (Van Themsche et al., 2009; Wang et al., 2007), localisation (Feng et al., 2008; Takahashi et al.,2006; van Diepen et al., 2009; von Stein et al., 2005) and oxidation (Cao et al., 2009). Here we discussthe significance and roles of many of these PTEN interacting proteins (Fig. 1). Such an analysissupports the idea that many of these proteins represent clinically significant oncogenes and tumoursuppressors that are modified during carcinogenesis, yet mediate their tumour promoting effectsindirectly through controlling PTEN function. In this regard, the evidence is probably strongest forPICT-1/GLTSCR2, P-Rex-2, RAK/FRK, Prdx1 and NEDD4.1.
Peroxiredoxin 1
PTEN activity and downstream signalling can be regulated by oxidation of its catalytic cysteineresidue by Reactive Oxygen Species (ROS) that are generated in many circumstances, including whencells are stimulated by several growth factors (Kwon et al., 2004; Leslie et al., 2003). Despite the fact thatmost tumours display greatly elevated levels of ROS, it has been difficult to determine both moleculardetails and the importance of PTEN oxidation in tumour formation and in its physiological functions.
It has been known for a few years that mice lacking the hydrogen peroxide scavenging enzymePeroxiredoxin 1 (EC 1.11.1.15) develop a broad range of tumours as they age (Neumann et al., 2003). Instudies to determine the mechanisms by which Prdx1 null mice develop tumours, Cao et al. (2009)showed that MEFs lacking Prdx1 displayed elevated levels of Akt activation, but only if they alsoexpressed PTEN. Detailed analysis of the connection between PTEN and Prdx1 indicates that theproteins physically interact in a redox dependent manner, and that this interaction protects PTEN from
P P
C N
PDZ binding site PTEN 403aa
C2 Phosphatase
Prdx1
Phosphorylation sites
MyoV Pict1
PAR3 NHERF MAGI
P-Rex2
VEEPSNPEASSSTSV T PDV S DNEPDHYRY S D TT D S DP ENEPFDEDQHTQI TK V *
GSK3 CK2 CK 2
P P P P P P P P P P P P MSP58
Fig. 1. PTEN binding partners with known binding positions on the phosphatase. The 403 amino acid PTEN phosphatase is shown,including a detailed expansion of the 52 amino acid C-terminal tail that includes an extreme C-terminal PDZ binding consensus and2 clusters of phosphorylation sites, which are phosphorylated by CK2 and GSK3 as shown. Where known, the implicated bindingsites on PTEN for its binding partners are shown. The PDZ domain containing proteins, PAR-3, NHERF and MAGI bind to theC-terminal PDZ binding consensus and MSP58 binds to Thr366 when this residue is phosphorylated. Myosin V, Peroxiredoxin 1 andP-Rex-2 bind to the C-terminal tail of PTEN in a less well defined manner. For diagrammatic purposes, each binding partner isrepresented as a small oval. The sizes of the binding partners in humans according to NCBI Entrez Protein and Unigene (http://www.ncbi.nlm.nih.gov/protein) are: P-Rex-2, 1605 amino acids; Pict1/GLTSCR2, 1317 amino acids; Myosin Va, 1854 amino acids; Prdx1, 199amino acids; MAGI-1, 1461 amino acids; NHERF1, 356 amino acids; PAR-3, 1244 amino acids and MSP58, 462 amino acids.

N.R. Leslie et al. / Advances in Enzyme Regulation 50 (2010) 112–118114
oxidation. The data also show that Prdx1 null mice develop more H-Ras induced mammary tumoursthan mice expressing Prdx1 and that these tumours display both elevated Akt phosphorylation andPTEN oxidation (Cao et al., 2009). These data not only provide the strongest current evidence for theimportance of redox regulation in controlling PTEN’s tumour suppressor status, but also implicatehydrogen peroxide itself as an important mediator of PTEN oxidation in vivo.
Myosin V
It is clear that in order to function correctly, especially in large cells such as neurons, not just largecomponents like vesicles and mRNA, but also many soluble proteins must be actively trafficked aroundthe cell. A recent report from van Diepen et al. (in press) has provided the first evidence supporting theidea that PTEN protein does not simply move around the cell by diffusion, but rather can be activelytransported to sites of action. The authors identified Myosin Va (EC 3.6.4.1) in a screen for PTEN inter-acting proteins in brain extract and verified the interaction both by co-immunoprecipitation of theendogenous proteins and fluorescence lifetime measurements of the expressed tagged proteins. Theinteraction appeared to be shared by Myosins Vb and Vc, was mediated by, or at least required, a chargedcluster of amino acids in the Myosin V globular cargo binding domain and was most efficient withphosphorylated PTEN that is believed to take up a closed conformation. Significantly, the authors thenshowed that blocking the interaction, by over-expressing this cargo binding domain alone or working incells with reduced endogenous expression of both Myosins Va and Vb, increased neuronal cell size andribosomal protein S6 phosphorylation in a PI3K dependent manner (van Diepen et al., in press).
Currently there is very little data to link Myosin 5 function with cancer. Amongst over 2000mutations identified in colon and breast tumours, Sjoblom et al. (2006) identified 2 mutations inMyosin 5c in colon tumours although there is no evidence for the significance of these mutations intumour development and Kuang et al. (2008) identified promoter methylation of Myosin5b selectivelyin Acute Lymphoblastic Leukemia (ALL). If PTEN function in many cell types requires trafficking to sitesof action, one would predict that interfering with this could be significant for tumour development,although redundancy between the 3 human Myosin 5 isoforms or specific roles in large differentiatedcells may be significant in this regard.
PTEN ubiquitin E3 ligases – NEDD4.1 and XIAP
PTEN expression in cultured cell systems can be controlled strongly by changes in protein stabilitymediated by the ubiquitin-proteasome system (Leslie et al., 2008). This has led to great interest in theenzymes that control the ubiquitination of PTEN and the somewhat controversial proposals that PTENstability can be regulated through the direct actions of the E3 ubiquitin ligases NEDD4.1 (Wang et al.,2007) and XIAP (Van Themsche et al., 2009). NEDD4.1 is a HECT domain containing ubiquitin E3 ligase(EC 6.3.2.19). It’s connection to PTEN was identified by the biochemical purification of a UbcH5cdependent PTEN E3 ligase activity. NEDD4.1 acts very efficiently in these conditions as a PTEN E3 ligasein vitro and influenced PTEN expression levels in cultured cells when over-expressed or knocked down(Wang et al., 2007). Moreover, the authors revealed a significant inverse correlation between NEDD4.1and PTEN mRNA expression levels in a group of bladder cancers and that NEDD4.1 knockdown sup-pressed xenograft tumour formation in a PTEN expressing prostate cancer cell line (DU145), but not ina line lacking PTEN (PC3). These data and subsequent work analysing NEDD4.1 over-expression ingastric and colorectal tumours (Kim et al., 2008a) and confirming an interaction between NEDD41.1and PTEN (Ahn et al., 2008; Yim et al., 2009) come together to support a model whereby NEDD4.1 over-expression in tumours leads to the ubiquitination and degradation of PTEN, driving carcinogenesis.However, the generality of this model has been called into question, firstly by a study that saw no effectof NEDD4.1 deletion on PTEN expression or localisation in studies using MEFs or cardiac tissue fromNEDD4.1 knockout mice (Fouladkou et al., 2008).
A further study that has called into question a general role for NEDD4.1 in the control of PTENexpression through ubiquitination and proteasomal degradation has recently proposed a different,RING domain containing Ub E3 ligase, XIAP, in this role (Van Themsche et al., 2009). This report saw noeffect of NEDD4.1 knockdown on PTEN expression in MCF7 cells and failed to detect an interaction

N.R. Leslie et al. / Advances in Enzyme Regulation 50 (2010) 112–118 115
between the proteins. However, in these experiments, knockdown of XIAP led to a strong increase inPTEN expression in MCF7 cells and an associated decrease in downstream Akt phosphorylation that didnot occur if PTEN itself was knocked down (Van Themsche et al., 2009). The authors then went on toshow in vitro poly-ubiquitination of PTEN mediated by XIAP. These results suggest the interestingpossibility that the well documented over-expression of XIAP in many tumours, that has made ita target for cancer therapy, might be in part driven by effects on PTEN abundance in addition to itsindependent well documented role in the control of apoptosis (Galban and Duckett, 2009).
An important point in the consideration of PTEN ubiquitination is its likely complexity. There isalready evidence for PTEN mono- and poly-ubiquitination, with different functional consequences onat least two regions of the phosphatase (Trotman et al., 2007). It is currently unknown whether all PTENpoly-ubiquitination involves Lys48 linked chains that are usually believed to target proteins for pro-teasomal degradation, or whether other chain linkages are found. It seems likely therefore thatdifferent E3 and possibly E2 enzymes will be involved in controlling PTEN function in different ways.
PDZ-dependent binding partners: PAR-3, NHERF, MAST and MAGI proteins
PDZ domains are modular protein domains that most commonly mediate interactions with smallC-terminal peptide binding sequences in other proteins, but can also bind to internal sequences andeven lipids (Garner et al., 2000; Zimmermann, 2006). The first PTEN interacting proteins proposedwere several containing PDZ domains, including DLG, MAST205, and the MAGI proteins, 1, 2 and 3,which initially were identified in yeast 2-hybrid screens (Adey et al., 2000; Wu et al., 2000a,b). Theseinteractions all depend upon the extreme C-terminus of the PTEN protein, ending in the amino acids(-QITKV), which form a consensus binding sequence for a subfamily of PDZ domains. Although forsome of these initial proteins, this significance is uncertain as an interaction with PTEN has not yet beendemonstrated using endogenous proteins, the evidence for an interaction between PTEN and bothMAGI-1 and MAGI-2 is quite strong, along with the implication that this interaction can control cellmigration and invasion (Hu et al., 2007; Kotelevets et al., 2005; Wu et al., 2000a).
More recently interactions have been shown for PTEN with both the polarity determinant PAR-3and the scaffold NHERF, confirmed using endogenous proteins. Knockdown of NHERF was shown tolead to enhanced stimulation of both Akt phosphorylation and membrane ruffling in response to PDGF(Takahashi et al., 2006), fitting with the ability of NHERF also to bind the PDGF-R and of PTEN to opposethese processes. Studies in polarised MDCK canine kidney epithelial cells have shown that knockdownof either PAR-3 or PTEN lead to repolarisation defects in a calcium switch assay in which cadherindependent cell junctions are disrupted by removal of extracellular calcium (Feng et al., 2008). Thisdefect could be rescued only by PAR-3 and PTEN proteins competent for binding to each other.Significantly, this study not only used a PTEN deletion mutant lacking the extreme C-terminal minimalPDZ binding mutant, which prevents the interaction of PTEN with any PDZ domain containing bindingpartner, but also used a PTEN protein carrying a mutation in a neighbouring acidic pocket that isrequired for the efficient PTEN-PAR-3 interaction, but does not appear to affect other PDZ dependentPTEN interactions (Feng et al., 2008).
These studies agree with earlier work showing that the PDZ dependent targeting of PTEN is notrequired for its ability to control overall cellular PIP3 levels or Akt, but appears more important for theregulation of functions related to cytoskeletal remodelling such as membrane ruffling, cell spreadingand migration, which presumably reflect localised pools of PIP3 (Leslie et al., 2001, 2007). However,further work is required to dissect the contributions of different PDZ domain containing bindingpartners to the mechanisms by which PTEN controls these functions and to tumour suppression. Anintriguing addition to the potential complexity of this regulation was the recent discovery that PTENcan be acetylated on lysine residues (Okumura et al., 2006) and that acetylation of the penultimateC-terminal amino acid, lysine 402, may be able to enhance the interaction of PTEN with PDZ domaincontaining binding partners (Ikenoue et al., 2008).
MSP58/MCRS1
MSP58, also known as MCRS1 and the human homolog of the avian TOJ3 oncogene, was identifiedin a yeast 2-hybrid screen for PTEN interacting proteins and the two proteins were shown able to

N.R. Leslie et al. / Advances in Enzyme Regulation 50 (2010) 112–118116
interact through the phospho-specific binding FHA domain of MSP58 and C-terminal phosphorylationsites on PTEN, almost certainly Thr366. It was also shown that MSP58 could transform PTEN null MEFs,but that this transformation could be blocked by either wild-type or catalytically inactive PTEN(Okumura et al., 2005). Unlike some of the other PTEN interacting proteins, MSP58 appears to be able todrive transformation only when it is not bound by PTEN, and it will be interesting to learn more in thefuture about the mechanisms underlying these data.
PICT-1/GLTSCR2
The PICT-1/GLTSCR2 protein is encoded by a candidate tumour suppressor gene and has beenshown to bind to PTEN, appearing to promote its phosphorylation and stability (Okahara et al., 2004).More recent data show increases in cultured cell proliferation and resistance to apoptosis in cells withknocked down PICT1 expression, but not if PTEN was also knocked down. This indicates that PICT-1/GLTSCR2 suppresses cell proliferation and causes sensitivity to apoptosis specifically through PTEN(Okahara et al., 2006). Finally in this study, PICT1 expression was found to be reduced in 41% of a cohortof neuroblastoma tumour samples, associating with reduced PTEN protein levels with retained mRNA.The demonstration of reduced expression of PICT-1/GLTSCR2 in glioblastoma samples and the iden-tification of mutations and deletions within the gene in these tumours (Kim et al., 2008b) providesstrong evidence that it may be a bona fide tumour suppressor at least in some tissues and that thisactivity may be mediated in large part through its effects on PTEN expression (Okahara et al., 2004,2006; Yim et al., 2007).
P-Rex-2
An exciting new study has revealed an interaction between PTEN and the rac activator P-Rex-2(Fine, 2009). Small GTPases, like those of the ras superfamily, have an active conformation when theyare bound to GTP and change to an inactive form upon hydrolysis of the bound GTP to GDP. Therefore,they are activated by guanine nucleotide exchange factors (GEFs) that promote the dissociation of GDPand the binding of GTP which is present in the cell at higher concentrations. P-Rex-2 is a GEF for thesmall GTPase rac1 which appears to activate rac when both PI 3-kinase and G-protein coupled receptorsignalling pathways are activated (Welch et al., 2002).
P-Rex-2 was initially identified in a screen for PTEN interacting proteins in cell extracts froma glioblastoma cell line that lacks PTEN, using a GST-PTEN column to pull down binding partners.After the interaction had been verified using endogenous proteins, the authors showed that purifiedP-Rex-2 protein was able to inhibit PTEN lipid phosphatase activity in vitro. This agreed with datafrom cell lines, in which over-expression or knockdown of P-Rex-2 respectively inhibited orincreased both Akt phosphorylation and proliferation, but these effects were only observed in cellsexpressing wild-type PTEN (MCF7 and MCF10A), not in cells lacking the phosphatase (BT549). Finallythe authors presented evidence that P-Rex-2 could be a clinically significant oncogene. P-Rex-2mRNA is over-expressed in many tumours relative to normal tissue, the P-Rex-2 gene is located ina region frequently amplified in breast prostate and colorectal cancers and finally with 9 differentmutations having been described in a range of tumour types, P-Rex-2 appears to be one of the mostfrequently mutated GEFs in human cancers (Fine, 2009). Since the inhibitory activity of P-Rex-2appeared to require the GEF active regions, yet a GEF inactive point mutant of P-Rex-2 had the sameeffect on PTEN and Akt phosphorylation, these data support a model by which P-Rex-2, bindsdirectly to PTEN through its GEF domains (DH-pH) to inhibit its phosphatase activity and activatedownstream PI3K dependent signalling. It seems possible that this inhibitory activity might also becontrolled conformationally by PI3K and GPCR signalling as P-Rex-2 GEF activity is, but this must bethe subject of future study.
RAK
The RAK protein tysoine kinase, also known as FRK, was recently identified in a screen for PTENinteracting proteins (Yim et al., 2009). Its expression tightly correlated with PTEN in breast cancer

N.R. Leslie et al. / Advances in Enzyme Regulation 50 (2010) 112–118 117
tissue samples and knockdown or induced expression of RAK led to a loss or gain of PTEN expressionrespectively. Similarly, the stability of the PTEN protein seemed reduced and its ubiquitinationenhanced when RAK expression was reduced by RNA interference. Finally, over-expression of RAK inbreast cancer MCF7 cells suppressed proliferation and xenograft tumour formation in mice, whereasknockdown of RAK expression in untransformed MCF10A mammary epithelial cells led to enhancedproliferation and tumour formation (Yim et al., 2009). This work proposes RAK/FRK as a novel tumoursuppressor, which fits with the location of the FRK gene at chromosome 6q21, a region of commondeletion in many tumour types, including breast, and the previous demonstration of growth inhibitionin breast cancer caused by RAK/FRK over-expression (Meyer et al., 2003).
Summary
The principle of significant oncogenes and tumour suppressors acting through a dominant down-stream tumour suppressor has been well established with p53. Here, the MDM2 oncoprotein isa ubiquitin ligase that targets p53 for proteasomal degradation and also appears to directly inhibit itstranscriptional activity. The ARF tumour suppressor in turn is able to bind to MDM2 and block itseffects on p53. Also, a variety of tumour viruses encode oncoproteins that disrupt this system (O’Sheaand Fried, 2005). In this way, expression of the MDM2 oncogene or any of several viral oncogenes, ormutations in the ARF tumour suppressor, have been shown to promote carcinogenesis indirectlythrough suppression of p53 expression and function (Vogelstein et al., 2000).
Evidence is emerging that, unlike p53, PTEN’s tumour suppressor functions act in a dose dependentmanner. Because complete loss of PTEN appears not to be required to drive tumour development,binding partners that are able to control PTEN activity are likely to be of great interest as indicated bythe recent reports reviewed in this article. Future research into these mechanisms should reveal thetrue significance of PTEN as a dominant mediator of changes in other novel oncogenes and tumoursuppressors.
Acknowledgements
NRL is a Research Councils UK Academic Fellow. LS and PT are supported by studentships from theBBSRC and GZ and NW are supported by studentships from the Wellcome Trust. Research in the InositolLipid Signalling laboratory is funded by the UK Medical Research Council, the Association for Inter-national Cancer Research and a consortium of pharmaceutical companies comprising Astra Zeneca,Boehringer Ingelheim, GlaxoSmithKline, Merck-Serono and Pfizer.
References
Adey NB, Huang L, Ormonde PA, Baumgard ML, Pero R, Byreddy DV, et al. Threonine phosphorylation of the MMAC1/PTEN PDZbinding domain both inhibits and stimulates PDZ binding. Cancer Res 2000;60:35–7.
Ahn Y, Hwang CY, Lee SR, Kwon KS, Lee C. The tumour suppressor PTEN mediates a negative regulation of the E3 ubiquitin-protein ligase Nedd4. Biochem J 2008;412:331–8.
Cao J, Schulte J, Knight A, Leslie NR, Zagozdzon A, Bronson R, et al. Prdx1 inhibits tumorigenesis via regulating PTEN/AKTactivity. EMBO J 2009;28:1505–17.
Eng C. PTEN: one gene, many syndromes. Hum Mutat 2003;22:183–98.Feng W, Wu H, Chan LN, Zhang M. PAR-3-mediated junctional localization of the lipid phosphatase PTEN is required for cell
polarity establishment. J Biol Chem 2008;283:23440–9.Fine Bea. Activation of the PI3K pathway in cancer through inhibition of PTEN by exchange factor P-REX2a. Science 2009.Fouladkou F, Landry T, Kawabe H, Neeb A, Lu C, Brose N, et al. The ubiquitin ligase Nedd4-1 is dispensable for the regulation of
PTEN stability and localization. Proc Natl Acad Sci U S A 2008;105:8585–90.Galban S, Duckett CS. XIAP as a ubiquitin ligase in cellular signaling. Cell Death Differ 2009.Garner CC, Nash J, Huganir RL. PDZ domains in synapse assembly and signalling. Trends Cell Biol 2000;10:274–80.Hu Y, Li Z, Guo L, Wang L, Zhang L, Cai X, et al. MAGI-2 Inhibits cell migration and proliferation via PTEN in human hep-
atocarcinoma cells. Arch Biochem Biophys 2007;467:1–9.Ikenoue T, Inoki K, Zhao B, Guan KL. PTEN acetylation modulates its interaction with PDZ domain. Cancer Res 2008;68:6908–12.Keniry M, Parsons R. The role of PTEN signaling perturbations in cancer and in targeted therapy. Oncogene 2008;27:5477–85.Kim SS, Yoo NJ, Jeong EG, Kim MS, Lee SH. Expression of NEDD4-1, a PTEN regulator, in gastric and colorectal carcinomas. APMIS
2008a;116:779–84.Kim YJ, Cho YE, Kim YW, Kim JY, Lee S, Park JH. Suppression of putative tumour suppressor gene GLTSCR2 expression in human
glioblastomas. J Pathol 2008b;216:218–24.

N.R. Leslie et al. / Advances in Enzyme Regulation 50 (2010) 112–118118
Kotelevets L, van Hengel J, Bruyneel E, Mareel M, van Roy F, Chastre E. Implication of the MAGI-1b/PTEN signalosome instabilization of adherens junctions and suppression of invasiveness. FASEB J 2005;19:115–7.
Kuang SQ, Tong WG, Yang H, Lin W, Lee MK, Fang ZH, et al. Genome-wide identification of aberrantly methylated promoterassociated CpG islands in acute lymphocytic leukemia. Leukemia 2008;22:1529–38.
Kwon J, Lee SR, Yang KS, Ahn Y, Kim YJ, Stadtman ER, et al. Reversible oxidation and inactivation of the tumor suppressor PTENin cells stimulated with peptide growth factors. Proc Natl Acad Sci U S A 2004;101:16419–24.
Leslie NR, Batty IH, Maccario H, Davidson L, Downes CP. Understanding PTEN regulation: PIP2, polarity and protein stability.Oncogene 2008;27:5464–76.
Leslie NR, Bennett D, Gray A, Pass I, Hoang-Xuan K, Downes CP. Targeting mutants of PTEN reveal distinct subsets of tumoursuppressor functions. Biochem J 2001;357:427–35.
Leslie NR, Bennett D, Lindsay YE, Stewart H, Gray A, Downes CP. Redox regulation of PI 3-kinase signalling via inactivation ofPTEN. EMBO J 2003;22:5501–10.
Leslie NR, Downes CP. PTEN: the down side of PI 3-kinase signalling. Cell Signal 2002;14:285–95.Leslie NR, Yang X, Downes CP, Weijer CJ. PtdIns(3,4,5)P3-dependent and -independent roles for PTEN in the control of cell
migration. Curr Biol 2007;17:115–25.Levine RL, Cargile CB, Blazes MS, van Rees B, Kurman RJ, Ellenson LH. PTEN mutations and microsatellite instability in complex
atypical hyperplasia, a precursor lesion to uterine endometrioid carcinoma. Cancer Res 1998;58:3254–8.Maxwell GL, Risinger JI, Gumbs C, Shaw H, Bentley RC, Barrett JC, et al. Mutation of the PTEN tumor suppressor gene in
endometrial hyperplasias. Cancer Res 1998;58:2500–3.Meyer T, Xu L, Chang J, Liu ET, Craven RJ, Cance WG. Breast cancer cell line proliferation blocked by the Src-related Rak tyrosine
kinase. Int J Cancer 2003;104:139–46.Mutter GL, Lin MC, Fitzgerald JT, Kum JB, Baak JP, Lees JA, et al. Altered PTEN expression as a diagnostic marker for the earliest
endometrial precancers. J Natl Cancer Inst 2000;92:924–30.Neumann CA, Krause DS, Carman CV, Das S, Dubey DP, Abraham JL, et al. Essential role for the peroxiredoxin Prdx1 in
erythrocyte antioxidant defence and tumour suppression. Nature 2003;424:561–5.O’Shea CC, Fried M. Modulation of the ARF-p53 pathway by the small DNA tumor viruses. Cell Cycle 2005;4:449–52.Okahara F, Ikawa H, Kanaho Y, Maehama T. Regulation of PTEN phosphorylation and stability by a tumor suppressor candidate
protein. J Biol Chem 2004;279:45300–3.Okahara F, Itoh K, Nakagawara A, Murakami M, Kanaho Y, Maehama T. Critical role of PICT-1, a tumor suppressor candidate, in
phosphatidylinositol 3,4,5-trisphosphate signals and tumorigenic transformation. Mol Biol Cell 2006;17:4888–95.Okumura K, Mendoza M, Bachoo RM, DePinho RA, Cavenee WK, Furnari FB. PCAF modulates PTEN activity. J Biol Chem 2006;
281:26562–8.Okumura K, Zhao M, Depinho RA, Furnari FB, Cavenee WK. Cellular transformation by the MSP58 oncogene is inhibited by its
physical interaction with the PTEN tumor suppressor. Proc Natl Acad Sci U S A 2005;102:2703–6.Salmena L, Carracedo A, Pandolfi PP. Tenets of PTEN tumor suppression. Cell 2008;133:403–14.Sjoblom T, Jones S, Wood LD, Parsons DW, Lin J, Barber TD, et al. The consensus coding sequences of human breast and
colorectal cancers. Science 2006;314:268–74.Suzuki A, Nakano T, Mak TW, Sasaki T. Portrait of PTEN: messages from mutant mice. Cancer Sci 2008;99:209–13.Takahashi Y, Morales FC, Kreimann EL, Georgescu MM. PTEN tumor suppressor associates with NHERF proteins to attenuate
PDGF receptor signaling. EMBO J 2006;25:910–20.Trotman LC, Niki M, Dotan ZA, Koutcher JA, Di Cristofano A, Xiao A, et al. Pten dose dictates cancer progression in the prostate.
PLoS Biol 2003;1:E59.Trotman LC, Wang X, Alimonti A, Chen Z, Teruya-Feldstein J, Yang H, et al. Ubiquitination regulates PTEN Nuclear Import and
tumor suppression. Cell 2007;128:141–56.van Diepen M, Parsons M, Downes CP, Leslie NR, Hindges R, Eickholt BJ. Myosin V controls PTEN function and neironal cell size.
Nat Cell Biol, in press.Van Themsche C, Leblanc V, Parent S, Asselin E. X-linked Inhibitor of apoptosis protein (XIAP) regulates PTEN ubiquitination,
Content, and Compartmentalization. J Biol Chem 2009;284:20462–6.Vogelstein B, Lane D, Levine AJ. Surfing the p53 network. Nature 2000;408:307–10.von Stein W, Ramrath A, Grimm A, Muller-Borg M, Wodarz A. Direct association of Bazooka/PAR-3 with the lipid phosphatase
PTEN reveals a link between the PAR/aPKC complex and phosphoinositide signaling. Development 2005;132:1675–86.Wang X, Trotman LC, Koppie T, Alimonti A, Chen Z, Gao Z, et al. NEDD4-1 is a Proto-Oncogenic ubiquitin ligase for PTEN. Cell
2007;128:129–39.Welch HC, Coadwell WJ, Ellson CD, Ferguson GJ, Andrews SR, Erdjument-Bromage H, et al. P-Rex1, a PtdIns(3,4,5)P3- and
Gbetagamma-regulated guanine-nucleotide exchange factor for Rac. Cell 2002;108:809–21.Wu X, Hepner K, Castelino-Prabhu S, Do D, Kaye MB, Yuan XJ, et al. Evidence for regulation of the PTEN tumor suppressor by
a membrane- localized multi-PDZ domain containing scaffold protein MAGI-2. Proc Natl Acad Sci U S A 2000a;97:4233–8.Wu Y, Dowbenko D, Spencer S, Laura R, Lee J, Gu Q, et al. Interaction of the tumor suppressor PTEN/MMAC with a PDZ domain of
MAGI3, a novel membrane-associated guanylate kinase. J Biol Chem 2000b;275:21477–85.Yim EK, Peng G, Dai H, Hu R, Li K, Lu Y, et al. Rak functions as a tumor suppressor by regulating PTEN protein stability and
function. Cancer Cell 2009;15:304–14.Yim JH, Kim YJ, Ko JH, Cho YE, Kim SM, Kim JY, et al. The putative tumor suppressor gene GLTSCR2 induces PTEN-modulated cell
death. Cell Death Differ 2007;14:1872–9.Zimmermann P. The prevalence and significance of PDZ domain-phosphoinositide interactions. Biochim Biophys Acta 2006;
1761:947–56.