Embed Size (px)
Citation preview

I
Psychopharmacology© by Springer-Verlag 1978
0033-3158/78/0060/0001/$ 02.20
of development (Lerner and Nose, 1977; Asper et aI.,1973) of dopaminergic supersensitivity.
Although the time-course of development of dopaminergic supersensitivity has received some attention,there is little or no information on the rate of development of dopamine/neuroleptic receptors in the firstdays of neuroleptic treatment. For example,Christensen et al. (1976) reported an increase in sensitivity to apomorphine-induced stereotypies within aday or two after single injection of chlorpromazine orhaloperidol; similar results were reported on climbingbehavior by Costentin et al. (1977) and Martres etal.(1977). However, detailed information on the timecourse of development of the receptor alterations afterrepeated neuroleptic administration has not yet beenreported. The shortest treatment schedule hithertoreported was by Burt et al. (1977), who treated rats withhaloperidol for 7 days and then withdrew them for fivedays. By that time it was found that the 3H-haloperidolreceptors had already achieved their maximum increase(Table 1).
This rapid development of dopaminergic supersensitivity in animals (albeit at massive doses) is fasterthan the rate of development of tardive dyskineticsymptoms in patients. This is one of the main reasonswhy Tarsy and Baldessarini (1977) feel thatneuroleptic-induced dopaminergic supersensitivity (inanimals) may not be an appropriate model for tardivedyskinesia.
According to Crane (1973), the development oftardive dyskinesia within the first 6 months of treatment is unusual and most of the patients with. tardivedyskinesia developed their symptoms after neuroleptictreatment for one year or more. Tarsy and Baldessarini(1977) suggest, therefore, that the do.paminergic supersensitivity seen after repeated neuroleptic treatment ofanimals is a better model for acute dyskinesia. Thisdyskinesia appears within 2- 5 days after the initiationof the neuroleptic treatment (Fig. 1).
.. - ... .~- --.....,.., .. - ~~ ---~. ..,...~
tment of Pharmacology. Faculty of Medicine, University of Toronto. Toronto. Ontario, Canada, M5S 1A8
. I Muller and Philip Seeman
:words: Tardive dyskinesia - Dopamine receptors~~ereotypy:le!
e-Course of Development of Dopaminergicersensitivity
re appears to be a correlation between the time. :se of development of tolerance to a neuroleptic''I. the rate of development" of dopaminergiclrsensitivity. For example, according to Ezrin
.aters and Seeman (1977), tolerance of catalepsy toQperidol develops rapidly over the first five days and
.. ~.develops more slowly. While the development of}olerance may to some extent be accounted for byning from test to test, it correlates well with the rate
ract. It is known that a single dose of a neurolepticelicit dopaminergic supersensitivity in animals. OnI ther hand, the clinical syndrome of tardive dyski-.~ takes many months or years to develop. To:Ive this apparent discrepancy, it is possible that'linical or latent tardive dyskinesia is fully comi ated in most patients taking neuroleptics. In:rs, where the tardive dyskinesia is full-blown and
Iy apparent, the dopaminergic supersensitivity" be decompensated. Such compensatory and de'pensatory phases have been proposed earlier bypykiewicz (1974), in the case of Parkinson's,ase.popaminergic supersensitivity persists for a periodportional to the length of the neuroleptic treatment.., not yet clear whether the relation between the'th of treatment and the persistence of super-
. itivity holds for very long treatments, but in""bple the relationship might account for the per
ence of tardive dyskinesia after years of neuroleptic'·1treatment.J
inalInvestigations
:.Jtaminergic Supersensitivity After Neuroleptics:~!'e-Course and Specificity

"1!~!"
Psychopharmacology 60 (197
242220
LENGTH OF TREATMENTWITH NEUROLEPTIC
patients may have a latent or subclinical dyskiwhich is fully compensated by certain adaptati "the brain (see Fig. 2). ,
This suggestion of a latent compensated Ntardive dyskinesia is analogous to the earlypensated phase of Parkinson's Disease, as prop6Hornkiewicz (1974). In this early stage of Park!Disease, it is thought that the dopaminergic celicounterbalanced by several compensatory cha~
18
LONG
1614
_0 _" CATALEPSYTOLERANCE
~DOPAMINERGIC SUPERSENSITIVITYUNMASKED BY DA AGONISTS
12
~ Patients unlikelyto develop Tardive dyskinesia
About 2 yearsSHORT
lCOMPENSATEDIn the presence
_ :~:~u~::~t::_
IDECOMPENSATED(-20% 01 patients)
IS?I
ACUTE
~~SKINESIAS~
%RAT7)0-- ---080 GNAWING" DOPAMINEo II TURNOVER1 TOLERANCE
::/1//2°V/~
o 1 2 3 4 5 6 7 8 9 10
enCOJenQ;Cl.:J
(f)
.':!OlQ;c
Fig.2. Compensation of dopaminergic supersensitivity in tardive dyskinesia - a model. Decompensated dopaminergic supersensitivity J,spontaneous appearance of dyskinetic symptoms. Dopaminergic supersensitivity compensated in the presence of neuroleptics will be clidormant until neuroleptics are discontinued or dose is lowered. Fully compensated dopaminergic supersensitivity could be precipitadopamine agonists or anticholinergic drugs
In order to demonstrate behavioural dopaminergicsupersensitivity in rats which have received long-termneuroleptics, it is necessary to challenge them witheither dopamine-mimetic drugs or anticholinergicdrugs (Tarsy and Baldessarini, 1974; Gianutsosand ,Lal, 1976). This is because such rats do not spontaneously show stereotypy. Similarly, many patients onlong-term neuroleptic},may not spontaneously exhibitany obvious dyskinetlc signs in the early stages. Such
E'"Q.
oo
Days neuroleptic treatment
Fig. I. Time-course of dopaminergic supersensitivity. Maximal observed cbange from tbe control was taken as 100 ::~. Rat gnawing: Christen',et al. (1976). Acute dyskinesias: Marsden et al. (1975). Turnover tolerance: Lerner and Nose (1977). Catalepsy tolerance: Ezrin-WatersSeeman (1977)
2
DEVELOPMENT OF DOPAMINERGIC SUPERSENSITIVITY:COMPARISON WITH ACUTE DYSKINESIA
E!c 100oo
EgOJOlC
'".co'"Ex
'"2
' .. ;-'':,J
3
50
®
100
OJ 80OJcC1l£:0 60CiiE)(
40C1l~
"" 20
NS
STRIATALNEUROLEPTICBINDING (K)
f-------7-o.-'o.:::~- STRIATAL
\
\----__ NEUROLEPTIC'\ ---.q BINDING (B)
\ \\ \
\ \\ \
\ \\ \o \ \
\ \\ \
\ \. . \ \
TURNING \ " \(SA) 0 ,,\
STEREOTYPY~\ \(SM) \ \\ \
\ \\ \
\ \
\ \o. 0
o 5 10 15Days after neuroleptic
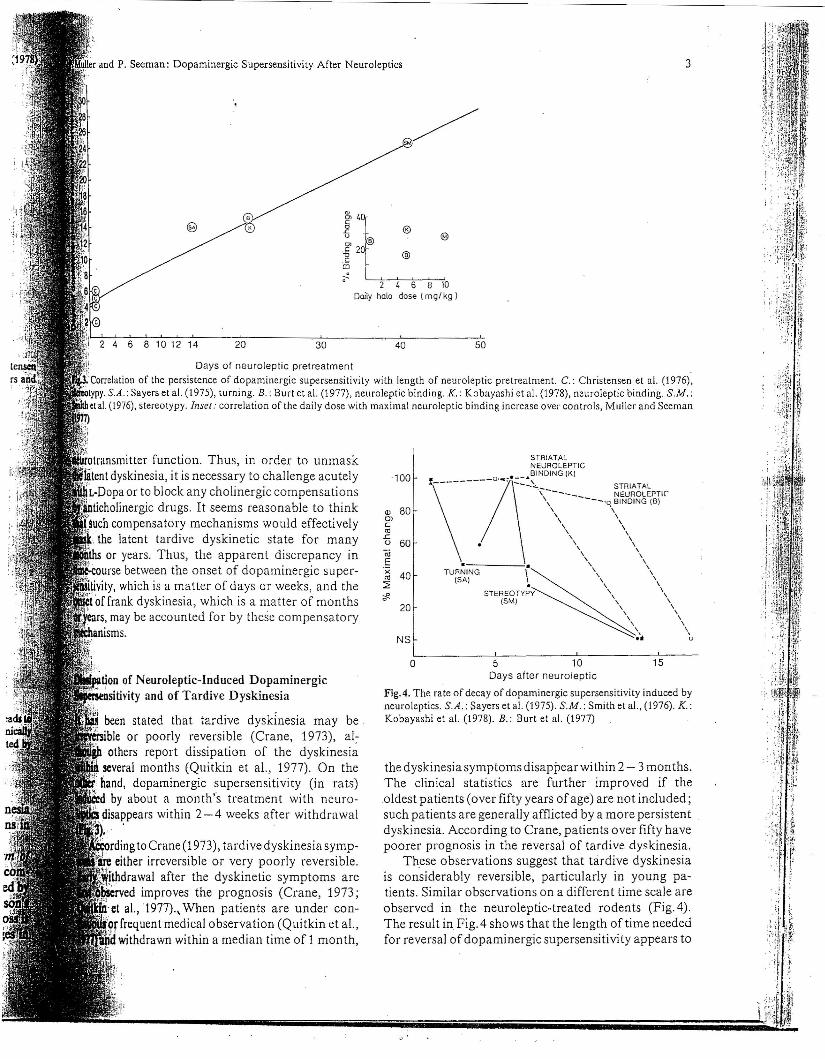
Fig.4. The rate of decay of dopaminergic supersensitivity induced byneuroleptics. S.A.: Sayers et al. (1975). S.M.: Smith el aI., (1976). K.:Kobayashi et al. (1978). B.: Burt et al. (1977)
the dyskinesia symptoms disappear within 2 - 3 months.The clinical statistics are further improved if theoldest patients (over fifty years of age) are not included;such patients are generally afflicted by a more persistentdyskinesia. According to Crane, patients over fifty havepoorer prognosis in the reversal of tardive. dyskinesia.
These observations suggest that tardive dyskinesiais considerably reversible, particularly in young patients. Similar observations on a different time scale areobserved in the neuroleptic-treated rodents (Fig. 4).The result in Fig. 4 shows that the length oftime neededfor reversal ofdopaminergic supersensitivity appears to
40
t'}01 B
~ 20 @
~ I , , .. ,• 2'6810
Daily halo dose Imgt kg)
30204 6 8 10 12 14
and P. Seeman: Dopaminergic Supersensitivity After Neuroleptics
tioo of Neuroleptic-Induced DopaminergicI sitivity and of Tardive Dyskinesia
been stated that tardive dyskinesia may be.ible or poorly reversible (Crane, 1973), al~
. others report dissipation of the dyskinesiaseveral months (Quitkin et a!., 1977). On thehand, dopaminergic supersensitivity (in rats)
by about a month"s treatment with neurodisappears within 2-4 weeks after withdrawal
.).''... rding to Crane (1973), tardive dyskinesia symp-I. either irreversible or very poorly reversible.'ilhdrawal after the dyskinetic symptoms are'rved improves the prognosis (Crane, 1973;ct aI., 1977).\ When patients are under con
or frequent medical observation (Quitkin et a!.,../:"
" withdrawn within a median time of 1 month,
Days of neuroleptic pretreatment
Correlation of the persistence of dopaminergic supersensitivity with length of neuroleptic pretreatment. C.: Christensen et at. (1976),ypy. S.A.: Sayers et al. (1975), turning. B.: Burt et at. (1977), neuroleptic binding. K.: Kobayashi et al. (1978), neuroleptic binding. S.M.:etal. (1976), stereotypy. Inset: correlation of the daily dose with maximal neuroleptic binding increase over controls, Muller and Seeman
otransmitter function. Thus, in order to unmask.. lent dyskinesia, it is necessary to challenge acutelyL-Dopa or to block any cholinergic compensationsticholinergic drugs. It seems reasonable to think
:such compensatory mechanisms would effectively. the latent tardive dyskinetic state for many
or years. Thus, the apparent discrepancy inurse between the onset of dopaminergic super-
'vity, which is a matter of days or weeks, and thet of frank dyskinesia, which is a matter of months
rs, may be accounted for by these compensatorynisms.

6
•Cfji@
6 : 6i~l~! 8 ! 4
~14
i~~: 3 1
i'1I!
7 8
o,.$ .....~ .~
~c::: C)-o,
&- '::' ,§o,~ <? -9
~;;:- ~
~ ~ /? <:Q ~'1> '>:" 4' 0; ~ <:-
".:<: i ".:z:: J ".:<: ".:<: ".:<: ".:<: i1 I II I I.. I I I
~I J I
I II II II I
[jJI 1i . 1
- I
t1JI,
- 4 I1~ Gi:ii' I
I E&l I8 7 I 7 3 4 I
I I
80[6040
20
ON
~ 20[
t °::: N
Effects of Long-Term Nonneuroleptic Drugson Dopamine Transmission
From the drugs summarized in Table 2, opiates aretdrugs closest to neuroleptics in terms of their effectthe dopaminergic system after repeated.admirtistratioTolerance to the cataleptic effects as well as to dpamine turnover occurs with both opiatesneuroleptics (Gessa and Tagliamonte, 1975)'1
Reports ofcross tolerance between neurolepticsopiates have been published (Puri and Lal, 1974; Ez'Waters arid Seeman, 1977), even though the two studdo not agree whether the cross tolerance is one-waytwo-way. Acutely, both opiates and neuroJeptics in'acatalepsy, even though there are differences in'pearance of the animal, as well as different pathinvolved (Costall and Naylor, 1973); furtherm'
Fig.5. Percentage changes in· neurotransmitter receptors (rat bregions) following long-term haloperidol treatment (10 mg/kgJdi"for at least 3 weeks). Height of the bars: mean percentage chan.Shaded portioll: SEM. The number of completely independentlY-1l$sayed membrane preparations (from 2 to 15 rats per preparatilIsed in each experiment (N) is indicated below each bar (* P < O.OS** P < 0.02, using Wilcoxon's rank test). Long-term haloperidtreatment selectively increased the dopamine/neuroleptic bindil1without significant effects on acetylcholine receptors eH-QNalpha-adrenergic receptors eH-WB-4101), or opiate receptors enaloxone)
Psychopharmacology 60 (I
significant increase in 3H-dopamine binding to tlimbic areas, even though the results show a large tretowards such increase. Tolerance to catalepsy aftrepeated neuroleptic administration was demonstra(by some, but not others (Table 1).
Effects of Long-Term Neuroleptic Treatmenton Dopamine Receptors (Table 1)
Soon after the development of the 3H-haloperidol assaymethod for neurolepticfdopamine receptors (Seeman etal., 1975; Burt et al., ·1975), Muller and Seemanreported that these receptors increased after long-termneuroleptic treatment (Muller and Seeman,1976; Burtet al., 1977). This has been recently more fully examined(Kobayashi et al., 1978; Muller and Seeman, 1977;Burt et al., 1977; Friedhoff et al., 1977). The maximumincrease appears to be around 30 % for the 3H_neuroleptic receptor (striatum) and about 65 %for the3H-apomorphine sites. This increase is characteristicfor all neuroleptics studied, except for clozapine in thestudy by Kobayashi et al. (1978). Clozapine hasgenerally yielded conf1icting results. Chronic treatmentwith this drug resulted in stimulated locomotion (Smithand Davis, 1976; Gianutsos and Moore, 1977) andstereotypy in the hands of Smith and Davis (1976) butnot of Gnegy et al. (1977). Chronic clozapine pretreatment had no effect on dopamine turnover (vonStralendorff et al., 1976; Gianutsos and Moore, 1977)in different brain areas, but large decreases ofdopamineturnover after long-term clozapine was observed in thestriatum and the olfactory tubercle (Gianutsos andMoore, 1977). There was also no effect on the adenylatecyclase (Gnegy et al., 1977). .
The increase in locomotor behaviour after repeatedneuroleptics suggests that dopaminergic supersensitivity occurs in the mesolimbic areas. Locomotion hasbeen shown to be' associated primarily with the mesolimbic rather than striatal dopaminergic system(Jackson et al., 1975a, b; Costall and Naylor, 1975;Pijnenburg et al., 1976; Creese and Iversen, 1974).Jackson et al. (1975 a) have shown that long-termpenfluridol potentiates the locomotor response to dopamine administered to the nucleus accumbens but notto the striatum. These results are supported by ourfindings (Fig. 5) of dopaminergic supersensitivity in themesolimbic areas. Klawans et al. (1977) did not show a
Dose-Dependency in Long-Term Neuroleptic Treatment
The experiments with chronic neuroleptic showed littledose-dependency (Fig.3, inset). It is probable that theeffect could be dose-dependent at lower neurolepticconcentrations. Interestingly, according to Crane(1973), no consistent dose-dependency can be demonstrated for the incidence of tardive dyskinesia.
correlate with the duration of neuroleptic administration, regardless of the type and dose of neurolepticemployed, and regardless of the type of supersensitiveproperty monitored.
",~:" ':
~~ J.:
i~~~L:"
~~'j'i
"~ .

Table 3. The effect of chronic administration of neuroleptics on non-dopaminergic neurotransmission
there are biochemical differences (Kuschinsky andHo~nykiewicz, 1972; Leysen et al., 1977) a~ well aspharmacological :differences (Ezri'n-Waters et al.,1976). Both groups of drugs block amphetamineindhced ;stereotypy (Sasame et al., 1972) as well asincrease dopamine turnover (Ahtee and! Kaariainen,
~l .
.• 1Muller and Seeman (1977)'Muller and Seeman (1977)
Kobayashi et al. (1978)
Kobayashi et al. (1978)
Muller and Seeman (1977)
Kobayashi et al. (1978)
Muller and Seeman (1977). ;':!
Heal et al. (1976)
Muller and Seeman (1977) ,
Kobayashi et al. (1978)
Kobayashi et al. (1978)
Muller and Seeman (1971)
Jl,[;
Dustan and Jackson (1976)Dustan and Jackson (197~
Muller and Seeman (1977) :I
Heal et al. (1976)
Heal et al. (1976)
Dolphin et al. (1977)Dustan and Jackson (197Muller and Seeman (19
Dolphin etal. (1977)Dolphin et al. (1977)
References
st 10 nM GABA± 0.5 nM GABAhippo 10 nM GABA± 0.5 pM GABA
st 1 nM QNB± 1 ~IM atrop.st I nM QNB± 100 nM scop.hippo 1 nM QNB± 1 pM atrop.hippo 1 nM QNB± 1 pM atrop.hippo 1 nM QNB± 0.1 pM scop.mli 1 nM QNB± 0.1 pM scop.cx 1 nM QNB± 0.1 pM scop.
locomotionwith L-tryptophanlocomotionwith L-trypophanlocomotionwith L-trypophanst 3 nM 5-HT ± 100 oMhippo 3 nM 5-HT± 100 nM 5-HTex 3 oM 5-HT± 100 nM 5-HT
Assay conditions
adenylate cyclase limbicadenylate cyclase limbic;100 ~IM NAc1onidine-locomotionc1onidine-Iocomotioncx 1 nM WB-4101± 1 ~IM phenox.st 1 nM WB-4101± 1 ~IM phenox.
benztropine-Iocomotionreserpine-blockedlocomotion'atropine-Iocomotionpilocarp-blockedlocomotiondexetimide-locomotion
1973). Chronic neuroleptic treatment does notany significant change in the striatal or corloxone binding (Fig. 5). The morphine pHii'profile also differs from the neuroleptic profimorphine does not produce supersensitivity tQphine (Table 2). .
1-14 d
2d
2 d
3-5d
2d
1-7d
4d3-5d
2d
3-5d
4d4d
2d
3 d
2d2 d
2 d
3 d4d2d
3 d3 d
14 d
14 d
18 h
W
18 h
3 wks
3 wks
3 wks
3 wks
3 wks
3 wks
3 wks
3 wks
3 wks
4 d
3 wks
4d
3 wks3 wks
3 wks3 wks
3 wks
7 d3 wks3 wks
5 d
7 d7 d
24 d
2 wks24 d
R,
Haloperidol 10 rug/kg p.o.
Haloperidol 10 rug/kg p.o.
Haloperidol 5 mg/kg i. p.
Haloperidol 10 rug/kg p.o.
Haloperidol 10 mg{kg p.o.
Haloperidol 5 mg/kg i.p.
Haloperidol 5 mg/kg i.p.
Haloperidol 5 mg/kg i.p.
Haloperidol 3 mg/kg in waterHaloperidol2.5-10 mg/kg i.p.Haloperidol2.5-10 mg{kg i.p.Haloperidol 5 mg{kg i.p.
Haloperidol 10 mg/kg p.o.Haloperidol 10 mg/kg p.o.
Haloperidol 10 mg/kg p.o.
Haloperidol 10 mg/kg i.p.
Chlorpromazine 30 mg/kg i.p.
Spiroperidol 1 mg/kg i.p.
-31 %
NS
266%58%
73%
NS
NS
NS
NS
NS
NS
GABA:
NS
%Ll Drug/dose/day
Acetylcholine:
NS Haloperidol 3 mg/kg i.p.-17 % Haloperidol 3 mg/kg in water
NS
NS
Serotonin:
100%
NS Haloperidol 10 mg/kg p.o.
NS Chlorpromazine 5 mg/kg i.p.167 % Haloperidol 3 mg/kg i.p113 % Haloperidol 10 mg/kg p.o.
,
Noradrenaline:
NS Chlorpromazine 5 mg/kg i.p.60 % Chlorpromazine 5 mg/kg i.p.
120%NS
8
NS
.;~ ,.

9
AcknolVledgements. Supported by the Ontario Mental HealthFoundation and the Medical Research Council of Canada.
References
Ahtee, L., Kaariainen, 1.: The effect of narcotic analgesics on thehomovanillic acid content of rat nucleus caudatus. Eur. J.Pharmacol. 22, 216-218 (1973)
Ahtee, L.: Catalepsy and stereotypies in the rats treated withmethadone: relation to striatal dopamine. Eur. J. Pharmacol. 27,221-230 (1974)
Anden, N.-E., Butcher, S. G., Corrodi, H., Fuxe, K., Ungerstedt, U.:Receptor activity and turnover of dopamine and noradrenalineafter neuroleptics. Eur. J. Pharmacal. 11,303-314 (1970)
Asper, H., Baggiolini, M., Burki, H. R., Lauener, H., Ruch, W.,Stille, G.: Tolerance phenomena with neuroleptics. Catalepsyapomorphine stereotypies and striatal dopamine metabolism inthe rat after single and repeated administration of loxapine andhaloperidol. Eur. J. Pharmacol. 22, 287 - 294 (1973)
Bowers, M. B., Jr., Rozitis, A.: Regional differences in homovanillicacid concentrations after acute and chronic administration ofantipsychotic drugs. J. Pharm. Pharmacol. 26,743-745 (1974)
Burkard, W. P., Bartholini, G.: Changes in activation of adenylatecyclase and of dopamine turnover in rat striatum duringprolonged haloperidol treatment. Experimentia 30, 685 (1974)
Burt, D. R., Creese, 1., Snyder, S. H.: Antischizophrenic drugs:"chronic treatnlent elevates dopamine receptor binding in brain.Science 196, 326 - 328 (1977)
adenylate cyclase activity was unchanged by the treatment. Stimulation of locomotion with clonidine wasmore pronounced in the study of Dustan and Jackson(1976), but not in that of Dolphin et al. (1977). We havereported an increase in alpha-adrenergic receptors inthe rat cortex but not in the striatum (Muller andSeeman, 1977; Fig. 5). Such a possible noradrenergicsupersensitivity might be due to blockade of noradrenergic receptors by neuroleptics (U'Prichard et aI., 1977;Anden et aI., 1970; Keller et al., 1973).
Chronic neuroleptic treatment does not potentiatethe locomotor response to L-tryptophan. Of the neuroleptics studied by Heal et al. (1976), chlorpromazinewas the only one which caused apparent behaviouralsupersensitivity to L-tryptophan. In our binding studies, 3H-serotonin binding was increased in thestriatum but not in the cortex or hippocampus afterrepeated haloperidol treatment. Since we saw the sameeffect in the striatum of rats treated chronically withethanol, we thin,k that the neuroleptic-induced increasein the striatal binding might not be a specific effect(Muller and Seeman, 1977).
Table 3 shows no consistent changes in the cholinergic or GABA sites after repeated haloperidol administration. The apparent cholinergic hyposensitivity of thecholinergic system reported by Gianutsos and Lal(1976) could be accounted for by an increase in tonicdopaminergic action, thus swinging the cholinergicdopaminergic balance towards dopamine even if thesensitivity of the cholinergic system remainedunchanged.
":ofLong-Term Neuroleptics on Nondopaminergic'ssion
al studies indicate that there may be a possiblerenergic supersensitivity following repeated
jleptic administration (Table 3). Dolphin et a\.'n report supersensitivity of the limbic adenylate
to 10 11M noradrenaline while the baseline
er and P. Seeman: Dopaminergic Supersensitivity After NeurolepticsJ,i-i'
i~mg-term neuroleptic treatment consistently ele-;,~3H~neuroleptic receptors (Table 1); dopamine"ve adenylate cyclase, however, is inconsistently
d (Table 1; Burkard and Bartholini, 1974).fthe other drugs reviewed, the only drug showing'similarity to neuroleptics is ethanol. It causeslation of locomotion (Table 2), but there is no
nce of ethanol-induced dopaminergic supersensi", Phenobarbital, diazepam, promethazine, andutaclamol also do not produce dopaminergicsensitivity.e tolerance in dopamine turnover (striatum) after
'Ieptics has been noted by several labs with onetion (Puri and Lal, 1974); there is no agreementether such tolerance occurs in the limbic areas.ver, the degree of tolerance in turnover is of
"p~rable magnitude to the increase in 3H_61eptic receptors.:'hronic administration of dopamine agonists (ammine, L-Dopa or bromocryptine) induce apparent
inergic 'behavioural-facilitation', as monitored'1 :"
,;Stereotypy (Klawans and Margolin, 1975; Klawansf'l,' 1977; Fuxe et aI., 1973). Chronic treatment with
'agonists produces tolerance to their acute, ine-turnover-reducing effect. This agrees with
,proposal that chronic agonist pretreatment pros dopaminergic subsensitivity of dopaminergic'receptors and thus produces apparent behavioural, 'nergic facilitation (Muller and Seeman, inJation).'e have found that the binding of 3H_
morphine in the striatum is reduced in the chroniapomorphine- or amphetamine-treated rat, while
?H-haloperidol sites (displaceable by a low con"~tion of pimozide) is not altered by the same
atment.. Apomorphine is thought to have somelection for presynaptic sites (Carlsson, 1975) and
f.~zide prefers postsynaptic receptors (Gianutsos et76; Walters and Roth, 1976). We thus interpret.oP in 3H-apomorphine sites as a reduction in'ceptors after repeated administration of amphet; and apomorphine. The findings of Burt et al.
and Friedhoffet a\. (1977) did not detect changeschronic agonist pretreatment, possibly becausemethods did not distinguish between pre- andnaptic binding.

10
Burt, D. R., Enna, S. J., Creese, 1., Snyder, S. H.: Dopamine receptorbinding in the corpus striatum of mammalian brain. Proc. Natl.Acad. Sci. U.S.A. 72, 4655-4659 (1975)
Carlsson, A.: Receptor-mediated control of dopamine metabolism.In: Pre- and postsynaptic receptors. E. Usdin and W. E. Bunney,eds., pp. 49-63. New York: Marcel Dekker 1975
Christensen, A. V., Fjalland, B., M9Iller Nielsen, 1.: On the supersensitivity of dopamine receptors induced by neuroleptics.Psychopharmacology 48, 1- 6 (1976)
Clouet, D. H., Iwatsubo, K.: Dopamine-sensitive adenylate cyclaseof the caudate nucleus of rats treated with morphine. Life Sci. 17,35-40 (1975)
Costall, B., Naylor, R. J.: Neuroleptic and non-neuroleptic catalepsy.Arzneim. forsch. 23, 674-683 (1973)
Costall, B., Naylor, R. J.: The behavioural effects of dopamineapplied intracerebrally to areas of mesolimbic system. Eur. J.Pharmacol. 32, 87 - 92 (1975)
Costentin, J., Marcais, H" Protais, P., Baudry, M., Delabaume, S.,Matres, M. P., Schwartz, J. C.: Rapid devclopment of hypersensitivity of striatal dopamine receptors induced by alfamethylparatyrosine and its prevention by protein synthesis inhibitors.Life Sci. 21, 307-314 (1977)
Cox, B., Ary, M., Lomax, P.: Changes in sensitivity to apomorphineduring morphine dependence and withdrawal in rats. J.Pharmacol. Exp. Ther, 196,637-641 (1975)
Crane, G. E.: Persistent dyskinesia. Br. J. Psychiatry 122, 395 -405(1973)
Creese, 1., Iversen, S. D.: The role of forebrain dopamine systems inamphetamine induced stereotyped behaviour in the rat.Psychopharmacologia (Berl.) 39, 345 - 357 (1974)
Dolphin, A., Sawaya, M. C. B., Jenner, P., Marsden, C. D.:Behavioural and biochemical effects of chronic reduction ofcerebral noradrenaline receptor stimulation. NaunynSchmiedeberg's Arch. Pharmacol. 299, 163 -173 (1977)
Dustan, R., Jackson, D. M.: The demonstration of a change inadrenergic receptor sensitivity in the central nervous system ofmice after withdrawal from long-term treatment with haloperidol. Psychopharmacology 38, 105 -114 (1976)
Dustan, R., Jackson, D, M.: The demonstration of a change inresponsiveness of mice to physostigmine and atropine afterwithdrawal from long-term haloperidol pretreatment, J. NeuralTransm. 40, 181-189 (1977)
Engel, J., Liljequist, S.: The effect of long-term ethanol treatment onthe sensitivity of the dopamine receptors in the nucleus accumbens. Psychopharmacology 49,253-257 (1976)
Ezrin-Waters, c., Muller, P., Seeman, P.: Catalepsy induced bymorphine or haloperidol effects of apomorphine and anticholinergic drugs. Can. J. Physiol. Pharmacol. 54, 516-519(1976)
Ezrin- Waters, c., Seeman, P.: Tolerance to haloperidol catalepsy.Eur. J. Pharmacol. 41, 321-327 (1977)'
Ezrin-Waters, c., Seeman, P.: Haloperidol-induced tolerance tomorphine catalepsy. Life Sci. 21, 419-422 (1977)
Friedhoff, A. J., Bonnet, K., Rosengarten, H.: Reversal of twomanifestations of dopamine receptor supersensitivity by administration of L-DOPA. Res. Commun. Chern. Pathol. Pharmacol.16,411-423 (1977)
Fukui, K., Takagi, H.: Effect of morphine on cerebral contents ofmetabolites of dopamine in the normal and tolerant mice: itspossible relation to analgesic action. Br. J. Pharmacol. 44, 4551 (1972)
Fuxe, K.: Tools in the treatment of Parkinson's disease: studies onnew types of dopamine receptor stimulating agents. In: Progressin the treatment of Parkinsonism. Advances in neurology, Vol. 3,D. B. Caine, ed., pp. 273-279. New York: Raven 1973
Gessa, G. L" Tagliamonte, A.: Effect of methadone and dextromoramide on dopamine metabolism: comparison with halo-
peridol and amphetamine. Neuropharmacology 14, 913-':9(1975) il
Gianutsos, G., Drawbaugh, R. B.; Hynes, M. D., Lal"Behavioural evidence for dopaminergic supersensitivity ,chronic haloperidol. Life Sci. 14, 887 - 898 (1974) , t
Gianutsos, G., Lal, H.: Alteration in the action of cholinergic'anticholinergic drugs after chronic haloperidol: indirectdence for cholinergic hyposensitivity. Life Sci. 18, 515-5(1976) ,.
Gianutsos, G., Thornburg, J. E., Moore, K. E.: Differential actioof dopaminc agonists and antagonists on y-butyrolactoinduced increase in mouse brain dopamine. Psychpharmacology 50, 225 - 229 (1976)'1
Gianutsos, G., Moore, K. E.: Dopaminergic supersensitivityi'striatum and olfactory tubercle following chronic administratioof haloperidol or clozapine. Life Sci. 20, 1585 - J592 (1977)'
Gnegy, M., Uzunov, P., Costa, E.: Participation of an endogenoCa + + -binding protein activator in the development of dtinduced supersensitivity of strialal dopamine receptors.Pharmacol. Exp. Ther. 202, 558 - 564 (1977)
Griffiths, P. J., Littleton, J. M., Ortiz, A.: Changes in monoamiconcentrations in mousc brain associated with ethanol dpendence and withdrawal. Br. J. Pharmacol. 50, 489-494(197
Gyorgy, L., Pfeifer, K, A., Hajlman. B,: Modification of cert .central nervollS effects of haloperidol during long-term treament in the mouse and rat. Psychopharmacologia (Berl.) 1223 - 233 (1969)
Heal, D. J., Green, A. R., Boullin, D. J., Graham-Smith, D. G.Single and repeated administration of neuroleptic drugs to rateffect on striatal dopamine-sensitive adenylate cyclase anlocomotor activity produced by tranylcypromine and':.tryptophan or L-dopa. Psychopharmacology 49, 287-3(1976) •.
Hicks, J., Strong, R., Smith, R. C., Samorajski, T.: Behavioursupersensitivity to bromocriptine in mice. Society iiiNeuroscience abstract, III, p. 44J, No. 1406 (1977) i"
Hoffman, P. L., Tabakoff, B.: Alterations in dopamine recepfsensitivity by chronic ethanol treatment. Nature 268, 551-55(1977) i
Hornykiewicz, 0.: The mechanisms of action of L-DOPA'Parkinson's disease. Life Sci. 15,1249-1259 (1974)
Iwatsubo, K., Clouet, D. H.: Dopamine-sensitive adenylate cye!of the caudate nucleus of rats treated with morphinehaloperidol. Biochem. Pharmacol. 24, 1499-1503 (1975) ",
Jackson, D. M., Anden, N.-E., Engel, J., Liljequist, S.: The effect'long-term penfluridol treatment on the sensitivity of the ,dpamine receptors in the nucleus accumbens and the corp
. striatum. Psychopharmacologia (Bert.) 45, 151 -155 (1975) .;Jackson, D. M., Anden, N.-E., Dahlstrom, A.: A functional effect
dopamine in the nucleus accumbens and in some olhdopamine-rich parts of the rat brain. Psychopharmacol9'(Berl.) 45,139-149 (1975) :
Keller, H. H., Bartholini, G., Pletscher, A.: Increase of3-methoxhydroxyphenylethylene glycol in rat brain by neuroleptic d"Eur. J. Pharmacol. 23,183-186 (J973)f
Klawans, H. L., Crossett, P., Dana, N.: Effect of chronic amp'amine exposure on stereotyped behaviour: implications rpathogenesis of L-Dopa-induced dyskinesias. In: Dopamine'mechanisms. Advances in neurology, Vol. 9, D. Caine,Chase and A. Barbeau, eds., pp. 105 -112. New York: R1975 r
Klawans, H. L., Margolin, D. 1.: Amphetamine-induced dopamigic hypersensitivity in guinea-pigs. Arch. Gen. Psychiatry!725 -732 (1975) :
Klawans, H. L., Goetz, c., Nausieda, P. A., Weiner, W.:',Levodopa-induced dopamine receptor hypersensitivity. A'Neurology 2,125-129 (1977) I',
1£

Received February 24, 1978
11
Sasame, H. A., Perez-Cruet, J.: Evidence that methadone blocksdopamine receptors in the brain. J. Neurochem. 19, 1953 -1957(1972)
Sayers, A. C., Biirki, H. R., Ruch, W. Asper, H.: Neurolepticinduced hypersensitivity of striatal dopamine receptors in the ratas a model of tardive dyskinesias. Effects of c1ozapine, haloperidol, loxapine and chlorpromazine. Psychopharmacologia(Berl.) 41, 97 -104 (1975)
Scatton, B., Glowinski, J., Julou, L.: Dopamine metabolism in themesolimbic and mesocortical dopaminergic systems after singleor repeated administrations of the neuroleptics. Brain Res. 109,184-189 (1976)
Seeber, U., Kuschinsky, K.: Dopamine-sensitive adenylate cyclase inhomogenates of rat striata during ethanol and barbituratewithdrawal. Arch. Toxicol. (Berl.) 35, 247-253 (1976)
Seeman, P., Chau-Wong, M., Tedesco, J., Wong, K.: Brain receptorsfor antipsychotic drugs and dopamine: direct binding assays.Proc. Natl. Acad. Sci. U.S.A. 72, 4376 - 4380 (1975)
Smith, R. c., Davis, J. M.: Behavioural evidence for supersensitivityafter chronic administration of haloperidol, c10zapine andthioridazine. Life Sci. 19, 725 - 732 (1976)
Tang, L. c., Cotzias, G. C.: L-3,4-Dihydroxyphenylalanine-inducedhypersensitivity simulating features of denervation. Proc. Natl.Acad. Sci: U.SA 74, 2126-2129 (1977)
Tarsy, D., Baldessarini, R. J.: Behavioural supersensitivity toapomorphine following chronic treatment with drugs whichinterfere with synaptic function of catecholamines.Neuropharmacology 13, 927 - 940 (1974)
Tarsy, D., Baldessarini, R. J.: The pathophysiologic basis of tardivedyskinesia. BioI. Psychiatry 12, 431-450 (1977)
U'Prichard, D. c., Greenberg, D. A., Snyder, S. H.: Bindingcharacteristics of radiolabeled agonist and antagonist at centralnervous system alpha noradrenergic receptors. Mol. Pharmacol.13,454-473 (1977)
Von Stralendorff, V. B., Ackenheil, M., Zimmermann, J.: Akute undchronische Wirkung von Carpipramin, Clozapin; Haloperidolund Sulpirid auf den Stoffwechsel biogener Amine imRattengehirn. Arzneim. Forsch. 26, 1096 -1098 (1976)
Von Voigtlander, P. F., Losey, E. G., Triezenberg, H. J.: Increasedsensitivity .to dopaminergic agents after chronic neuroleptictreatment. J, Pharmacol. Exp. Ther. 193,88-94 (1975)
Walters, J. R., Roth, R. H.: Dopamine neurons: an in vivo system formeasuring drug interactions with presynaptic receptors .Naunyn-Schmiedeberg's Arch. Pharmacol. 296, 5 -14 (1976)
Worms, P., Scatton, B.: Tolerance to stereotyped behaviour and todecrease in striatal homovanillic acid levels after repeatedtreatment with apomorphine dipivaloyl ester. Eur. J. Pharmacol.45,395-396 (1977)
Yarborough, G.: Supersensitivity of caudate neurones after repeatedadministration of haloperidol. Eur. J. Pharmacol. 31, 367 - 369(1975)
Zarcone, V., Barchas, J., Hoddes, E., Montplaisir, J., Sack, R.,Wilson, R.: Experimental ethanol ingestion: sleep variables andmetabolites of dopamine and serotonin in the cerebrospinalfluid. In: Alcohol intoxication and withdrawal. Vol. 2, M. M.Gross, ed., pp. 431-451. New York: Plenum 1975
yashi, R. M., Fields, J. Z., Hruska, R. E., Beaumont, K.,amamura, H. 1.: Brain neurotransmitter receptors and chronictipsychotic drug treatment: a model for tardive dyskinesia. In:
nimal models in psychiatry. E. Usdin, ed., pp. 405-409. Newark: Pergamon 1978'nsky, K., Hornykiewicz, 0.: Morphine catalepsy in rat:ation to striatal dopamine metabolism. Eur. J. Pharmacol. 19,9-122 (1972) ,nsky, K.: Dopamine receptor sensitivity after repeated mor
administrations to rats. Life Sci. 17, 43 - 48 (1975)r, P., Nose, P.: Haloperidol: effect of long-term treatment ontstriatal dopamine synthesis and turnover. Science 197, 181
83 (1977)n, 1., Tollenaere, J. P., Koch, M. H. J" Laduron, P.:iITerentiation of opiate and neuroleptic receptor binding in ratrain. Eur. J. Pharmacal. 43, 253 - 267 (1977)en, C. D., Tarsy, D., Baldessarini, R. J.: Spontaneous andg-induced movement disorders in psychotic patients. In:chiatric aspects of neurologic disease. D. F. Benson and D.mer, eds., pp. 219 - 265. New York: Grune & Stratton 1975,M. P., Costentin, J., Baudry, M., Marcais, H., Protais, P.,
hwartz, J. C.: Long-term changes in the sensitivity of pre- andstsynaptic dopamine receptors in mouse striatum evidenced
. behavioural and biochemical studies. Brain Res. 136, 3197 (1977)Z., Singhal, R. L., Hrdina, P. D., Lirig, G: M.: Changes in'n cyclic AMP metabolism and acetylcholine and dopamine
ring narcotic dependence and withdrawal. Life Sci. 16,188994 (1975): Nielsen, 1., Fjalland, B., Pedersen, V., Nymark, M.:"rmacology of neuroleptics upon repeated administration.chopharmacologia (Berl.) 34, 95 -1 04 (1974)
,P" Seeman, P.: Increased specific neuroleptic binding afterronic haloperidol in rats. Soc. Neurosci. Abstr. 2, 874 (1976),P., Seeman, P.: Brain neurotransmitter receptors after long-
haloperidol: dopamine, acetylcholine, serotonin, (X
radrenergic and naloxone receptors. Life Sci. 21, 1751-1758977)
urg, A. J. J., Honing, W. M. M., Van Der Heyden, J. A. M.,Rossum, J. M.: Effects of chemical stimulation of the
"olimbic dopamine system upon locomotor activity. Eur. J.rmaca!. 35, 45 - 58 (1976)
.K., Lal, H.: Tolerance to the behavioural and neurochemicaleets of haloperidol and morphine in rats chronically treated
morphine or haloperidol. Naunyn-Schmiedeberg's Arch.rmacol. 282, 155 -170 (1974)
·K., Volicer, L., Cochin, J.: Changes in the striatal adenylatease activity following acute and chronic morphine treatment
, during withdrawal. J. Neurochem. 27, 1551-1554 (1976)F., Rifkin, A., Gochfeld, L., Klein, D. F.: Tardive
. iDesia: are first signs reversible? Am. J. Psychiatry 134,84(1977)W. H., Gerald, M. C.: The effect of chronic administration
withdrawal of ( +)-amphetamine on seizure threshold and.ogenous catecholamine concentrations and their rates ofynlhesis in mice. Psychopharmacology 51,175-179 (1977),J., Friedman, E., Gershon, S.: Striatal adenylate cyclase
\'ity following reserpine and chronic chlorpromazine admin-lion in rats. Life Sci. 1'7, 563 -568 (1975)
er and P. Seeman: Dopaminergic Supersensitivity After NeurolepticsF





























