Embed Size (px)
Citation preview

Vol. 41, No. 3JOURNAL OF VIROLOGY, Mar. 1982, p. 1055-10620022-538X/82/031055-08$02.00/0
In Vitro Reassembly of Vesicular Stomatitis Virus SkeletonsWILLIAM W. NEWCOMB, GREGORY J. TOBIN, JOHN J. McGOWAN, AND JAY C. BROWN*
Department of Microbiology, University of Virginia School of Medicine, Charlottesville, Virginia 22908
Received 14 August 1981/Accepted 19 October 1981
Vesicular stomatitis virus (VSV) has been disrupted with nonionic detergentplus 0.5 M NaCl under conditions which result in solubilization of the viralglycoprotein (G), matrix protein (M), and lipids, leaving the nucleocapsid in ahighly extended state. Dialysis of these suspensions to remove NaCl was found toresult in reassociation of nucleocapsids with M protein. Reassociated structureswere highly condensed and similar in appearance to "native" VSV skeletonsproduced by extraction of virions with detergent at low ionic strength. Forinstance, electron microscopic analysis revealed that, like "native" skeletons,"reassembled" skeletons were cylindrical in shape, with diameters in the range of51.0 to 55.0 nm and cross-striations spaced approximately 6.0 nm apart along thelength of the structure. Like native skeletons, reassembled skeletons were foundby sodium dodecyl sulfate-polyacrylamide gel electrophoresis to contain the viralN and M proteins, but they lacked the glycoprotein entirely. Both native andreassembled skeletons were found to be capable of in vitro RNA-dependent RNAsynthesis (transcription). In vitro skeleton assembly required the presence of Mprotein and nucleocapsids. No skeleton-like structures were formed by dialysis ofnucleocapsids in the absence of M protein or of M protein in the absence ofnucleocapsids. These results provide strong support for the view that the VSV Mprotein plays a functional role in condensing the viral nucleocapsid in vitro andraise the possibility that it may play a similar role in vivo.
Recent experiments in our laboratory haveemphasized the fact that NaCl can profoundlyaffect the disruption of vesicular stomatitis virus(VSV) by nonionic detergents (14). Extraction ofnative virions with 60 mM octylglucoside or 1%Triton X-100 at low ionic strength results insolubilization of the viral glycoprotein (G) andlipids, leaving insoluble structures called "skele-tons" (1, 12). Skeletons contain the viral RNAcomplexed with the N and M proteins in highlycondensed and very regular structures that havethe same overall cylindrical shape and striatedappearance as the native virus. Similar struc-tures are produced by extraction with digitonin(18) or with Tween 80 plus diethylether (2). Incontrast, detergent extraction in the presence of0.5 M NaCl results in solubilization of the Gprotein, the M protein, and lipids, liberating theviral nucleocapsid in an irregular and highlyextended state. One molar KCl was found tohave a similar effect on the extraction of Sendaivirus with 2% Triton X-100 (15).The dramatic effect of NaCl on the detergent-
induced disruption of VSV has motivated us toask whether its removal from detergent-highsalt-solubilized virus might promote some de-gree of reassociation of viral components. Intactvesicular stomatitis virions were, therefore, sol-ubilized in nonionic detergent containing 0.5 M
NaCl and then dialyzed against a low-ionic-strength buffer to remove NaCl. The results ofthis basic experiment, described below, showthat dialysis is accompanied by the reassemblyof VSV nucleocapsids and M protein to formcondensed structures quite similar to "native"VSV skeletons.
MATERIALS AND METHODSCell and virus growth. All experiments were per-
formed with the Mudd-Summers strain of VSV (Indi-ana), which was grown on monolayer cultures ofBHK-21 cells. Cells were propagated at 37°C in 150-cm2 plastic tissue culture flasks containing 40 ml ofDulbecco modified minimal essential medium supple-mented with 10% calf serum, 10% tryptose phosphatebroth, 100 U of penicillin per ml, and 0.1 mg ofstreptomycin per ml. Virus stocks were grown at a lowmultiplicity of infection as previously described (5)and were shown by rate-velocity ultracentrifugation tobe free from defective interfering particles. Viral pro-teins were radioactively labeled by including 5 p.Ci ofL-[3H]leucine (60 Ci/mmol; Amersham Corp.) per mlin the virus growth medium. VSV was purified fromthe infected cell medium by a two-step procedureinvolving rate-zonal followed by equilibrium ultracen-trifugation, essentially as described by Hunt and Wag-ner (9). The purified virus preparations were judged tobe free from cellular contamination by sodium dodecylsulfate (SDS)-polyacrylamide gel analysis which re-vealed only the five VSV proteins (N, NS, M, G, and
1055
on June 25, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

1056 NEWCOMB ET AL.
L); no contaminating cellular proteins could be detect-ed.
Disruption of VSV. Freshly purified VSV to beextracted with detergent was gently suspended in 0.01M Tris-hydrochloride buffer, pH 7.4, and centrifugedfor 10 min at 1,000 x g to remove virus aggregates.The supernatant virus suspension was then extractedwith either octylglucoside (1-O-n-octyl-P-D-glucopy-ranoside; Calbiochem, La Jolla, Calif.) or Triton X-100in the presence of 0.5 M NaCl. Octylglucoside extrac-tion was carried out by adjusting the virus suspensionto 60 mM octylglucoside, 0.5 M NaCI, 5 mM dithio-threitol, 10% glycerol, and 10 mM Tris-hydrochloridebuffer, pH 7.8 (octylglucoside-0.5 M NaCl dissocia-tion medium) at 0°C, and a concentration of 0.1 to 0.2mg of viral protein per ml. Similar conditions wereemployed for Triton X-100 extraction except that 1%Triton X-100 was substituted for 60 mM octylgluco-side. In both cases virus suspensions were thoroughlybut gently mixed with detergent plus NaCl and allowedto stand on ice (0°C) for 30 min before further opera-tions were performed. The initially turbid virus sus-pensions were clarified immediately upon addition ofdetergent plus NaCI, and the suspensions remainedclear during the 30-min period allowed for disruption.Detergent extraction of VSV in the absence of NaCIwas carried out as described previously (14).Reassembly conditions. VSV components were al-
lowed to reassociate during dialysis against 0.01 MTris-hydrochloride (pH 7.4)-S5 mM dithiothreitol (TB).Four-milliliter samples of VSV disrupted in detergentplus 0.5 M NaCI as described above were dialyzedagainst TB for 16 h at 4°C and for an additional 12 h at25°C. If dialyzed solutions were not analyzed immedi-ately, they were stored on ice for a maximum of 4 h.
Ultracentrifugation. Insoluble material was pelletedfrom suspensions of dissociated VSV or from reassem-bly mixtures by ultracentrifugation in 5-ml BeckmanSW50.1 nitrocellulose tubes. Four-milliliter samples ofdissociated or reassembled viral components werelayered on top of a 0.5-ml "cushion" of glycerol andwere centrifuged at 38,000 rpm (130,000 x g) for 2 h at4°C. The fluid above and including the interface of theglycerol pad was aspirated while the remainder of thepad, containing the pelleted material, was mixed with0.5 ml of TB and dialyzed overnight at 4°C to removeglycerol. Dialyzed samples were used directly forelectron microscopic and transcription analyses orwere lyophilized prior to SDS-polyacrylamide gel elec-trophoresis.Both rate-velocity and sedimentation equilibrium
sucrose gradient methods were employed to purifysubviral structures from dissociated and reassociatedVSV components. Rate-velocity analyses began with0.5 ml of sample which was layered onto the top of 10to 70% (wt/vol) linear sucrose gradients prepared in 5-ml Beckman SW50.1 nitrocellulose tubes. Stock su-crose solutions were prepared by dissolving ultrapuresucrose (Schwarz/Mann) in TB. Gradients were centri-fuged at 38,000 rpm for 90 min at 4°C in an SW50.1rotor. After centrifugation, the gradients were frac-tionated from the bottom by use of a peristaltic pumpand an LKB-7000 fraction collector. The radioactivitypresent in each fraction was determined by dissolvinga 20-piu sample in 0.5 ml of NCS tissue solubilizer(Amersham Corp.) containing 10 ml of toluene-basedscintillation cocktail (Research Products International,
Grove Village, Ill.) and counting in a Packard model3320 liquid scintillation spectrometer. Gradient frac-tions to be analyzed further were dialyzed against TBovernight at 4°C to remove sucrose and then werestained directly for electron microscopy or lyophilizedfor SDS-polyacrylamide gel analysis. Equilibrium ul-tracentrifugation was carried out by the same methoddescribed for rate-velocity centrifugation except thatcentrifugation was for 16 h rather than 90 min. Thedensity of each gradient fraction was determined byweighing a 100-,ul sample on a Mettler H18 analyticalbalance.
In vitro RNA-dependent RNA synthesis and poly-acrylamide gel analysis of RNA. Native and reassem-bled skeletons were tested for the presence of endoge-nous RNA-dependent RNA polymerase activity by theassay ordinarily employed for intact VSV (4). Assaymixtures contained 0.14 M NaCl, 0.2% Triton X-100,7.5 mM MgCl2, 0.01 M Tris-hydrochloride, pH 7.9, 1mM dithiothreitol, 0.3 mM ATP, GTP, and CTP, and0.03 mM [a-32P]UTP (0.45 Ci/mmol) plus 25 to 35 ,g ofviral protein in a total volume of 30 ,lI. Incubations forup to 3 h were carried out at 31°C, after which 5-RIsamples of the reaction mixture were precipitated in 1ml of 5% trichloroacetic acid. Precipitated RNA wascollected by filtration on 24-mm filters (MilliporeCorp.), dried, and counted at an efficiency of 65% intoluene-based liquid scintillation cocktail. Resultswere expressed as nanomoles of [32P]UMP incorporat-ed per minute per milligram of protein.
Other methods. Electron microscopic analysis wascarried out with subviral structures negatively stainedwith 2% phosphotungstic acid (pH 7.0) as describedpreviously (14). Particle measurements were made onpositive enlargements of electron microscope nega-tives. SDS-polyacrylamide gel electrophoresis and gelstaining with Coomassie blue were performed on 2-mm-thick slab gels as described by Nagpal and Brown(13). Seven-millimeter lanes were loaded with samplescontaining 50 to 100 ,ug of viral protein. All gelscontained a lane, labeled "std," for electrophoresis ofsolubilized VSV, and positions of the five VSV pro-teins (L, G, N, NS, and M) are indicated. All proteinconcentrations were determined by the Lowry method(11).
RESULTSDisruption of VSV in nonionic detergent plus
NaCl. Vesicular stomatitis virions were disrupt-ed in nonionic detergent plus 0.5 M NaCl underconditions shown previously to solubilize theviral envelope components and release the nu-cleocapsid in an uncondensed form (14). Theextent of detergent-NaCl-induced virus disas-sembly was monitored by electron microscopy,by sedimentation velocity ultracentrifugation,and by SDS-polyacrylamide gel electrophoresis.Figure 1A shows a negatively stained prepara-tion of insoluble structures pelleted from VSVdisrupted with 60 mM octylglucoside-0.5 MNaCI by centrifugation for 2 h at 130,000 x g asdescribed in Materials and Methods. Pelletedmaterial was found to contain only extendedviral nucleocapsids (6); no intact VSV or con-densed VSV skeletons could be detected. It
J. VIROL.
on June 25, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

REASSEMBLY OF VSV SKELETONS 1057
C
FIG. 1. Electron micrographs of detergent-extracted VSV and reassembled VSV skeletons. (A) Nucleocap-sids isolated by centrifugation of solutions produced by extracting VSV with 60 mM octylglucoside plus 0.5 MNaCl. (B) Skeletons isolated by centrifugation of suspensions produced by extracting VSV with 60 mMoctylglucoside. (C) Reassembled skeletons (arrows) identified by direct observation of the suspension resultingfrom dialysis of VSV solubilized with 60 mM octylglucoside-0.5 M NaCl. (D) Reassembled skeletons isolated bycentrifugation of the suspension resulting from dialysis of VSV solubilized with 60 mM octylglucoside-0.5 MNaCl. (A) x105,000; (B, C, and D) x125,000. Bar = 0.2 ,.m.
must be, therefore, that all or nearly all intactvirus particles were disassembled.
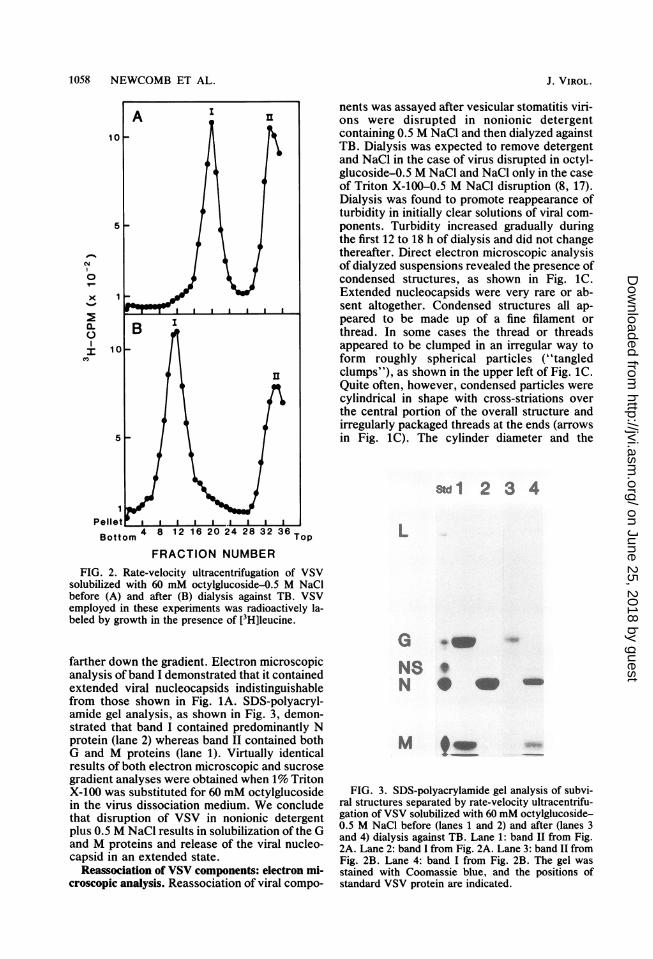
Figure 2A shows the results obtained whenVSV, grown in the presence of [3H]leucine, wasdisrupted in 60 mM octylglucoside plus 0.5 M
NaCl and analyzed by rate-velocity sedimenta-tion on 10 to 70% sucrose gradients. Two bandsof 3H-labeled viral proteins were observed, one(band II) at the top of the gradient where solubleproteins should occur and the other (band I)
VOL. 41, 1982
on June 25, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from
1058 NEWCOMB ET AL.
04N0
X 10
5
a. BI 10_
5 -
1PelletBottom Top
FRACTION NUMBERFIG. 2. Rate-velocity ultracentrifugation of VSV
solubilized with 60 mM octylglucoside-0.5 M NaCIbefore (A) and after (B) dialysis against TB. VSVemployed in these experiments was radioactively la-beled by growth in the presence of [3H]leucine.
farther down the gradient. Electron microscopicanalysis of band I demonstrated that it containedextended viral nucleocapsids indistinguishablefrom those shown in Fig. 1A. SDS-polyacryl-amide gel analysis, as shown in Fig. 3, demon-strated that band I contained predominantly Nprotein (lane 2) whereas band II contained bothG and M proteins (lane 1). Virtually identicalresults of both electron microscopic and sucrosegradient analyses were obtained when 1% TritonX-100 was substituted for 60 mM octylglucosidein the virus dissociation medium. We concludethat disruption of VSV in nonionic detergentplus 0.5 M NaCl results in solubilization of the Gand M proteins and release of the viral nucleo-capsid in an extended state.
Reassociation of VSV components: electron mi-croscopic analysis. Reassociation of viral compo-
nents was assayed after vesicular stomatitis viri-ons were disrupted in nonionic detergentcontaining 0.5 M NaCl and then dialyzed againstTB. Dialysis was expected to remove detergentand NaCl in the case of virus disrupted in octyl-glucoside-0.5 M NaCl and NaCl only in the caseof Triton X-100-0.5 M NaCl disruption (8, 17).Dialysis was found to promote reappearance ofturbidity in initially clear solutions of viral com-ponents. Turbidity increased gradually duringthe first 12 to 18 h of dialysis and did not changethereafter. Direct electron microscopic analysisof dialyzed suspensions revealed the presence ofcondensed structures, as shown in Fig. 1C.Extended nucleocapsids were very rare or ab-sent altogether. Condensed structures all ap-peared to be made up of a fine filament orthread. In some cases the thread or threadsappeared to be clumped in an irregular way toform roughly spherical particles ("tangledclumps"), as shown in the upper left of Fig. 1C.Quite often, however, condensed particles werecylindrical in shape with cross-striations overthe central portion of the overall structure andirregularly packaged threads at the ends (arrowsin Fig. 1C). The cylinder diameter and the
Std1 2 3 4
L
G urnNS'N *
M9 -
FIG. 3. SDS-polyacrylamide gel analysis of subvi-ral structures separated by rate-velocity ultracentrifu-gation of VSV solubilized with 60 mM octylglucoside-0.5 M NaCl before (lanes 1 and 2) and after (lanes 3and 4) dialysis against TB. Lane 1: band II from Fig.2A. Lane 2: band I from Fig. 2A. Lane 3: band II fromFig. 2B. Lane 4: band I from Fig. 2B. The gel wasstained with Coomassie blue, and the positions ofstandard VSV protein are indicated.
J. VIROL.
on June 25, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

REASSEMBLY OF VSV SKELETONS 1059
spacing between cross-striations were found tobe similar to those observed for "native" VSVskeletons, as shown in Table 1. The length of thecross-striated region was variable but nevergreater than the length of native skeletons. Themorphological similarity of native and reassem-bled skeletons was emphasized by electron mi-croscopic analysis of insoluble material pelletedfrom reassembly mixtures by ultracentrifugationat 130,000 x g for 2 h. Reassembled skeletonsharvested in this way were indistinguishablefrom native skeletons, as shown in Fig. 1D andB.
Control experiments showed that VSV nu-cleocapsids and soluble components must bepresent together for striated skeletons to formduring dialysis. These experiments involved dis-assembly of native virions in 60 mM octylgluco-side plus 0.5 M NaCl as described above andthen centrifugation to remove the nucleocapsidsfrom solution. The supernatant containing solu-bilized viral G protein, M protein, and lipids(soluble components) was then dialyzed againstTB under the same conditions employed forreassembly of viral skeletons. Similarly, theisolated nucleocapsids were resuspended at aconcentration of 0.1 mg of protein per ml inoctylglucoside-0.5 M NaCl dissociation mediumand dialyzed against TB. Electron microscopicanalysis of the two dialyzed fractions revealedthat neither contained skeletons or skeleton-likestructures. Isolated nucleocapsids gave imagessimilar to Fig. 1A both before and after dialysis.Dialysis of soluble components resulted in for-mation of a precipitate which pelleted aftercentrifugation at 600 x g for 5 min. SDS-poly-acrylamide gel analysis showed that the precipi-tate contained M protein only (data not shown);the glycoprotein remained in the supernatant.
Protein composition of reassembled skeletons.The protein composition of reassembled skele-tons was determined by SDS-polyacrylamide gelanalysis of structures purified from reassemblymixtures by sucrose density gradient ultracen-trifugation. Preparation of reassembled skele-tons began with VSV grown in the presence of[3H]leucine. Purified virions were disrupted innonionic detergent plus 0.5 M NaCl, and theresulting solutions were dialyzed to reassembleskeletons as described above. Reassembledskeletons were purified from the dialyzed sus-pensions by rate-velocity ultracentrifugation inone set of experiments and by sedimentationequilibrium in another. Figure 2B shows theresults of rate-velocity purification performedwith skeletons reassembled after disruption ofVSV with 60 mM octylglucoside plus 0.5 MNaCl. 3H-labeled viral components were foundto sediment in two bands, one (band II) near thetop of the gradient and the other (band I) cen-
TABLE 1. Dimensions of native and reassembledVSV skeletonsa
Spacing ofSkeletons Length (nm) Diam (nm) striations
(nm)
Native 204.4 ± 6.00 51.7 ± 3.00 5.7 ± 0.75Reas-
sembled Variable' 53.7 ± 2.70 6.2 ± 0.32a Results are reported as the mean value + 1 stan-
dard deviation for at least 20 measurements of repre-sentative particles. Data for native skeletons are fromNewcomb and Brown (14).
b The length of the cylindrical, striated portion ofreassembled skeletons was variable but never greaterthan 200.0 nm.
tered at fraction 12. Electron microscopic analy-sis revealed the presence of reassembled skele-tons in band I, but not in band II (micrographsnot shown). SDS-polyacrylamide gel analysis ofband I from Fig. 2B (reassembled skeletons)demonstrated that it contained the viral M and Nproteins, as shown in Fig. 3, lane 4. No Gprotein was detected in reassembled skeletonpreparations; G protein was found in band II atthe top of the gradient (see Fig. 3, lane 3).Results essentially identical to those shown inFig. 2 and 3 were obtained when 1% Triton X-100 was substituted for 60 mM octylglucoside inthe virus dissociation medium.When dialyzed reassociation mixtures were
centrifuged to equilibrium, two bands of 3H-labeled viral components were observed, one(band I) with a buoyant density of approximately1.26 g/cm3 and the other (band II) near the top ofthe gradient. Electron microscopic analysisdemonstrated the presence of reassembled skel-etons in band I; none were found in band II. SDS-polyacrylamide gel analysis of band I materialshowed that M and N were the predominantprotein components present (see Fig. 4, lanes 1and 3). As in the case of reassembled skeletonspurified by rate-velocity centrifugation, little orno G protein could be detected in reassembledskeletons purified by equilibrium sedimentation.Instead, G protein was found in band II at thetop of equilibrium gradients, as shown in Fig. 4,lanes 2 and 4. The identity of the disruptingdetergent did not substantially affect either thebuoyant density or the protein composition(compare lanes 1 and 3 in Fig. 4) of reassembledskeletons. Both parameters were virtually thesame in skeletons reassembled from virions dis-rupted with octylglucoside-NaCl and with Tri-ton X-100-NaCl.RNA-dependent RNA polymerase activity. In
vitro assays showed that both native and reas-sembled skeletons contained endogenous RNA-dependent RNA polymerase activity. Their spe-cific polymerase activities were found to be
VOL. 41, 1982
on June 25, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

1060 NEWCOMB ET AL.
Std 1 2
L
GNSN
_
4
M d-
FIG. 4. SDS-polyacrylamisembled skeletons and solublby sedimentation equilibritLane 1: skeletons (band I) isolubilized with 60 mM octyLane 2: soluble componentsreassembly of skeletons frommM octylglucoside-0.5 M N(band I) reassembled from VTriton X-100-0.5 M NaCl. Inents (band II) remaining afttons from VSV solubilized wiM NaCl. Gels were stained wthe positions of standard VSN
comparable to the activitnucleocapsids, as shown irtially lower than the specvirions assayed under t]Polyacrylamide gel analyspattern of similarity in thesized by reassembled sktons, and native VSV. IRNA (3) and many hipRNA species were syishown).
DISCUSSThe results reported I
formation of VSV skelet(reassembly took place duiof solutions produced by dular stomatitis virions in ntaining 0.5 M NaCl. Thesembled under these conbe virtually indistinguisiVSV skeletons isolated brus with nonionic deterger(14). For example, both n,skeletons were found to bders, with diameters of 5tions spaced approximattnative and reassembled siviral N protein, M proteil
3 4 Std protein entirely.
tons, reassembled skeletons were found to beL capable of in vitro RNA-dependent RNA syn-
thesis. There can be little question, therefore,that reassembled skeletons are faithful replicasof their native counterparts.
It is most likely that removal of salt, notremoval of detergent, was responsible for pro-
G moting skeleton formation in vitro. Skeletonsoz1 i NS formed abundantly during dialysis of virus solu-
N bilized with octylglucoside-NaCl when both de-tergent and NaCl were removed. They alsoformed normally from virus solubilized with
so_ Triton X-100-NaCl when only NaCl was re-
_ -111 M moved. The presence of detergent, therefore,did not appear to affect skeleton formation invitro, but the presence of NaCl clearly did. Salt
de gel analysis of reas- was found to have a similar effect on the abilitye components separated of Semliki Forest virus glycoproteins to asso-
reassembledntfrfoumga iSnV ciate with Semliki Forest virus nucleocapsids in
eglucoside-0.5 M NaCl. vitro. Glycoprotein-nucleocapsid interaction
(band II) remaining after was observed in 24 mM octylglucoside-10 mMVSV solubilized with 60 sodium phosphate buffer, pH 6.8, but not in thelaCl. Lane 3: skeletons same solution containing 0.1 M NaCl at pH 8.0'SV solubilized with 1% (7).Lane 4: soluble compo- Although mature skeletons were the predomi-ter reassembly of skele- nant structures formed during dialysis of solubi-ith 1% Triton X-100-0.5 lized VSV, other related structures were alsoith Coomassie blue, and observed at lower frequency. These included theV proteins are indicated. "tangled clumps" described above and tangled
clumps containing central cross-striations. Tan-ty observed for VSV gled clumps appeared to consist of a fine threadiTable 2, but substan- wound in an irregular way to form a roughly:ific activity of intact spherical particle. The thread diameter washe same conditions. smaller than the diameter of VSV nucleocap-is revealed an overall sids, and no nucleocapsids were found to pro-RNA species synthe- trude from tangled clumps. Cross-striated tan-eletons, native skele-n all cases "leader"gher-molecular-weightnthesized (data not
SIONhere demonstrate theons in vitro. Skeletonring overnight dialysislissolving native vesic-onionic detergent con-skeletons which reas-ditions were found tohable from "native"y extracting intact vi-at at low ionic strength,ative and reassembled)e cross-striated cylin-il to 55 nm and stria-ely 6 nm apart. Bothkeletons contained then, and RNA, but both
TABLE 2. RNA-dependent RNA synthesis bynative skeletons, reassembled skeletons, and
nucleocapsidsaSpecific % of native
Preparation polymeraseactivityb VSV
Native VSV 1.78 100Native VSV skeletons 0.45 25Nucleocapsids 0.44 25Reassembled skeletons 0.38 21
(Triton X-100)Reassembled skeletons 0.89 50
(octylglucoside)a Native skeletons and nucleocapsids were prepared
by disruption of VSV in 60 mM octylglucoside in theabsence of salt and in the presence of 0.5 M NaCl,respectively, as described previously (14). Reassem-bled skeletons were prepared by dialysis of VSVsolubilized with Triton X-100-NaCl or octylglucoside-NaCl as described in Materials and Methods.
b Expressed as nanomoles of [32P]UMP incorporat-ed per minute per milligram of protein.
J. VIROL.
I
on June 25, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

REASSEMBLY OF VSV SKELETONS 1061
gled clumps were similar in appearance totangled clumps except that they contained acentral cylindrical region with regularly spacedstriations and tangled clumplike material at bothends. The central cylindrical region was of vari-ous lengths in different particles, but its diameterand striation spacing were the same as thosefound in mature skeletons. Particles of this typecan be seen in Fig. 1C (arrows). We presumethat the minor components (tangled clumps andcross-striated tangled clumps) found in reassem-bly mixtures are intermediates in the skeletonself-assembly process. Their structural relation-ship to mature skeletons suggests that skeletonreassembly in vitro took place in the followingway. Nucleocapsids and M protein aggregatedto form tangled clumps in which condensednucleocapsids were surrounded by a fine threadcomposed of M protein. Cross-striations werethen formed by subsequent internal rearrange-ments in the tangled clump structure. This pro-posed pathway for skeleton reassembly is shownschematically in Fig. 5. Further experimentalstudies will be required to substantiate its role inskeleton reassembly.The results described in this paper indicate
that M protein must have a very strong affinityfor VSV nucleocapsids. This can be appreciatedfrom the fact that very few, if any, free nucleo-capsids remained after dialysis of virions solubi-lized with detergent-0.5 M NaCl; virtually allwere complexed with M protein in condensedstructures (see Fig. 2B). Similarly, nearly all Mprotein became associated with nucleocapsids.Very little remained in the soluble fraction, asshown in Fig. 3, lane 3, or Fig. 4, lanes 2 and 4.This contrasts to the behavior of the glycopro-tein which was present during dialysis of solubi-lized VSV, but was not incorporated into reas-sembled skeletons. The interaction ofnucleocapsids with M protein must, therefore,be very strong and at least somewhat specific.The abundant formation of VSV skeletons in
vitro raises the possibility that similar structuresmay be formed in vivo. Calculations based onthe data of Simonsen et al. (16) show that themolar concentrations of nucleocapsids and Mprotein in VSV-infected HeLa cells are compa-rable to or greater than their concentrations inthe reassembly experiments described here. De-pending on the values assumed for the cellvolume and the time after infection, the concen-tration of negative-strand nucleocapsids in in-fected cells falls in the range of 2 x 10-9 to 4 x10-9 M and that ofM protein is approximately 2x 10-6 to 5 x 10- M. The concentrationsemployed for in vitro skeleton reassembly in ourexperiments were 1 x 10-9 to 2 x 10- M fornucleocapsids and 1.2 x 10-6 to 2.4 x 10-6 Mfor M protein. The concentrations of nucleocap-
a. b. C. d.
FIG. 5. Proposed pathway for in vitro reassemblyofVSV skeletons. (a) Tangled clumps, (b and c) cross-striated tangled clumps, and (d) reassembled skele-tons.
sids and M protein are, therefore, high enough tosupport skeleton formation in infected cells.Experimental results of Knipe et al. (10) also
suggest that M protein may associate with VSVnucleocapsids in vivo. These authors examinedCHO cells infected with a strain of VSV,tsM601, which carries a temperature-sensitivelesion in the N protein. An unusually low levelof VSV nucleocapsids accumulated in cells in-fected with tsM601 at the nonpermissive tem-perature (39°C); N protein was rapidly degraded,and little full-length (42S) viral RNA was synthe-sized. In contrast, G protein was made in normalamounts and it migrated normally to the plasmamembrane. M protein was also synthesized innormal amounts, but it accumulated in solubleform in the cytoplasm rather than becomingattached to membranes as it does in cells infect-ed with wild-type VSV. This is the result thatwould be expected if M protein could associatewith membranes only after it had bound tonucleocapsids.
ACKNOWLEDGMENTSWe thank Nancy Salomonsky and Margie Phipps for techni-
cal assistance.This work was supported by National Institutes of Health
contract N01-NS-8-2391 and by Public Health Service grantHD-06390 from the National Institutes of Health.
LITERATURE CITED
1. Cartwright, B., C. J. Smale, and F. Brown. 1970. Dissec-tion of vesicular stomatitis virus into the infective ribonu-cleoprotein and immunizing components. J. Gen. Virol.7:19-32.
2. Cartwright, B., P. Talbot, and F. Brown. 1970. Theproteins of biologically active sub-units of vesicular sto-matitis virus. J. Gen. Virol. 7:267-272.
3. Colonno, R. J., and A. K. Banerjee. 1976. A unique RNAspecies involved in initiation of vesicular stomatitis virusRNA transcription in vitro. Cell 8:197-204.
4. Emerson, S. U., and R. R. Wagner. 1972. Dissociation andreconstitution of the transcriptase and template activitiesof vesicular stomatitis B and T virions. J. Virol. 10:297-309.
5. Fong, B. S., R. C. Hunt, and J. C. Brown. 1976. Asymmet-ric distribution of phosphatidylethanolamine in the mem-brane of vesicular stomatitis virus. J. Virol. 20:658-663.
6. Heggeness, M. H., A. Scheid, and P. W. Choppin. 1980.Conformation of the helical nucleocapsids of paramyxo-viruses and vesicular stomatitis virus: reversible coilingand uncoiling induced by changes in salt concentration.Proc. Natl. Acad. Sci. U.S.A. 77:2631-2635.
7. Helenius, A., and J. Kartenbeck. 1980. The effects ofoctylglucoside on the Semliki Forest Virus membrane.Eur. J. Biochem. 106:613-618.
VOL. 41, 1982
AI&-- c.l 1- -
qw,T
on June 25, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

1062 NEWCOMB ET AL.
8. Helenius, A., and K. Simons. 1975. Solubilization ofmembranes by detergents. Biochim. Biophys. Acta415:29-79.
9. Hunt, D. M., and R. R. Wagner. 1975. Inhibition byaurintricarboxylic acid and polyethylene sulfonate ofRNA transcription of vesicular stomatitis virus. J. Virol.16:1146-1153.
10. Knipe, D. M., D. Baltimore, and H. F. Lodish. 1977.Maturation of viral proteins in cells infected with tempera-ture-sensitive mutants of vesicular stomatitis virus. J.Virol. 21:1149-1158.
11. Lowry, 0. H., N. J. Rosebrough, A. L. Farr, and R. J.Randall. 1951. Protein measurement with the Folin phenolreagent. J. Biol. Chem. 193:265-275.
12. Miller, D. K., B. I. Fever, R. Vanderoef, and J. Lenard.1980. Reconstituted G protein-lipid vesicles from vesicu-lar stomatitis virus and their inhibition of VSV infection.J. Cell Biol. 84:421-429.
13. Nagpal, M. L., and J. C. Brown. 1980. Protein andglycoprotein components of phagosome membranes de-
rived from mouse L cells. Int. J. Biochem. 11:127-138.14. Newcomb, W. W., and J. C. Brown. 1981. Role of the
vesicular stomatitis virus matrix protein in maintainingthe viral nucleocapsid in the condensed form found innative virions. J. Virol. 39:295-299.
15. Shimizu, K., and N. Ishida. 1975. The smallest protein ofSendai virus: its candidate function of binding nucleocap-sid to envelope. Virology 67:427-437.
16. Simonsen, C. C., S. Batt-Humphries, and D. F. Summers.1979. RNA synthesis of vesicular stomatitis virus-infectedcells: in vivo regulation of replication. J. Virol. 31:124-132.
17. Stubbs, G. W., H. G. Smith, Jr., and B. J. Litman. 1976.Alkyl glucosides as effective solubilizing agents for bovinerhodopsin: a comparison with several commonly useddetergents. Biochim. Biophys. Acta 426:46-56.
18. Wagner, R. R., and C. A. Schnaitman. 1970. Proteins ofvesicular stomatitis virus, p. 655-671. In R. D. Barry andB. W. J. Mahy (ed.), The biology of large RNA viruses.Academic Press, Inc., New York.
J. VIROL.
on June 25, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from





























