Embed Size (px)
Citation preview

Research Collection
Doctoral Thesis
A molecular study of enterotropic murine coronavirus
Author(s): Zhang, Linong
Publication Date: 1996
Permanent Link: https://doi.org/10.3929/ethz-a-001702361
Rights / License: In Copyright - Non-Commercial Use Permitted
This page was generated automatically upon download from the ETH Zurich Research Collection. For moreinformation please consult the Terms of use.
ETH Library

CIss-ETH e>c.
Dissertation No. 11872
A molecular study of enterotropic murine
coronavirus
A dissertation submitted to the SWISS FEDERAL INSTITUTE OF TECHNOLOGY
ZURICH
For the degree of Doctor of Natural Science
Presented by Linong Zhang
Master Degree of Medicine from Fujian Medical College, P. R. China
Bom on 25th of October, 1961
Citizen of China
Accepted on the recommendation of
Prof. Dr. F. Wurgler, examiner
Dr. F. R. Homberger, co - examiner
Prof. Dr. P. E. Thomann, co - examiner
ZOrich 1996
ETHICS ETH-BW
00100002920942

1. Introduction
The Coronaviridae were first recognized as a distinct virus family by Tyrrell and
coworkers in 1968 (Tyrrell et a/., 1968). The family comprises two genera,
coronavirus and torovirus, which share similarities in the organization and
expression of their genomes and the structure of the virial gene products (Table 1).
The name "coronavirus" is derived from the solar corona-like ("corona" is Latin for
"crown") appearance of virus particles in negatively stained electron micrographs.
Table 1. Coronaviridae
Natural host virus Acronym
Coronavirus
Chicken
Cattle
Dog
Man
Man
Cat
Mouse
Pig
Pig
Pig
Turkey
Torovirus
Horse
Cattle
Avian infectious bronchitis virus IBV
Bovine coronavirus BCV
Canine coronavirus CCV
Human coronavirus 229E HCV 229E
Human coronavirus OC43 HCV OC43
Feline infectious peritonitis virus FIPV
Murine hepatitis virus MHV
Porcine epidemic diarrhea virus PEDV
Porcine hemagglutinating encephalomyelitis virus HEV
Porcine transmissible gastroenteritis virus TGEV
Turkey coronavirus TCV
Berne virus BEV
Breda virus BRV

The spectrum of diseases in humans and animals caused by coronaviruses
ranges from subclinical infection to rhinitis, enteritis, hepatitis, peritonitis,
encephalomyelitis, and death. One of the best-studied members of the coronavirus
family is mouse hepatitis virus (MHV). MHV is the most prevalent and probably the
most important viral pathogen in contemporary research mouse colonies (Homberger
and Thomann, 1994; Kraft and Meyer, 1990; Lindsey, 1986). Some MHV strains
produce primary respiratory infections, and many MHV strains are enterotropic
(Homberger and Thomann, 1994).
In this chapter a concise overview of the molecular biology of coronaviruses is
presented with the emphasis on MHV.
The virion
Coronaviruses are enveloped viruses, containing a single stranded, positive-
sensed and nonsegmented genome of 27-30 kb in length. A schematic
representation of a coronavirion is shown in Figure 1. The lipid envelope surrounds
a helicoidal nucleocapsid which consists of nucleocapsid phosphoprotein (N) and
the genomic RNA Several glycoproteins are present in the envelope, the membrane
protein (M) which serves as a bridge between the nucleocapsid and the viral
envelope; the spike protein (S) which forms the characteristic peplomers on the
virion surface; the small membrane protein (E); and in some coronaviruses (e.g.
MHV-JHM, BCV, HCV-OC43), the hemaglutinin esterase (HE).
Genome organization
In all coronaviruses the genes encoding the RNA-dependent RNA polymerase
(POL) and the structural proteins are present in the same order. 5' POL - S - sM - M
- N 3' (de Groot et a/., 1989). These genes, each having one or more open reading
frames (ORFs), are separated by conserved stretches of 8 to 10 nucleotides, termed
-4-

FIG. 1. Schematic representation of a coronavirion. Indicated are the RNA genome (RNA), the
nucleocapsid protein (N), the spike protein (S), the membrane protein (M), the small membrane
protein (E) and the hemagglutin-esterase protein (HE) (only present in some coronavirus).
intergenic sequences. So far, three coronavirus genomes have been completely
sequenced, i.e. MHV, HCV-229E and IBV (Boursnell et al., 1987; Bredenbeek et al
1990; Herold etai, 1993; Lee etal., 1991). Genome organizations of these three are
shown in Fig. 2. Some coronaviruses like TGEV, CCV, FIPV and FECV contain one
or two additional ORFs downstream of the N gene (de Groot et al., 1988; Vennema
etal., 1992).
Replication
The first step in the viral replication cycle is the binding of the virus to its cellular
receptor via its S glycoprotein peplomers or HE glycoproteins. This step is believed
-5-

Pol la
Pol la
Pol la
2 HE
MHV
5«L£
S M N
m
A 1113aM5a IBV3b\ \l5b
E
HCV
E N229E
FIG. 2. Genomic organizations of MHV, IBV and HCV 229E. The genomic open reading frames
(ORFs) encoding structural proteins (HE, S, M, N and E, i.e. 5b, 5 and 3c for MHV, HCV and IBV
respectively) are lightly dotted; those encoding nonstructural proteins (ORF 1a and ORF 1b) are filled.
to be one of the major determinants of viral tropism. Following binding of the virus to
the cell, fusion of the viral and cellular membranes takes place and the viral
nucleocapsid enters the host cytosol. The entire replication cycle takes place in the
cytoplasm of the host cell and nuclear functions are unnecessary. Coronaviruses are
positive-stranded RNA viruses and do not have a virion associated polymerase.
After the virion has entered the cell and the genome has been uncoated, gene 1 of
the viral genome is translated to produce the viral RNA-dependent RNA
polymerase(s) (Lai, 1990). Thus, the viral genomic RNA serves as a mRNA for the
RNA polymerase and then as a template for the synthesis of negative-stranded RNA.
From the new negative strands, full-length genomic positive strands as well as, when
transcription starts, the subgenomic mRNAs are synthesized. From these
subgenomic messenger RNAs, secondary translation of the nonstructural and
structural proteins occurs. The N protein is synthesized in the cytoplasm and is
-6-

phosphorylated on serine residues (Stohlman and Lai, 1979). As the only protein
constituent of the nucleocapsid, the N protein binds to the viral genomic RNA. The
M, S, and HE glycoproteins are synthesized on the rough endoplasmic reticulum and
transported to the membrane between the rough endoplasmic reticulum (RER) and
Golgi (reviewed in Spaan et al., 1988). Nucleocapsids are enveloped in the
RER/Golgi via interactions of the N, M and E proteins. Virus particles are released
from cells via exocytosis or cytolysis.
Transcription
The mechanism of coronavirus mRNA synthesis is a matter of considerable
debate. The first RNA believed to be transcribed by the RNA dependent RNA
polymerase is the full-length negative-stranded template (antigenome). The initial
model of coronavirus transcription was based on the observation that only a full-
length copy of the genome was present (Lai et al., 1982). Consequently, this was
believed to be the only template for all positive-stranded RNA species of
coronavirus. The primary transcript in this model was the leader, of 58-90 nt,
encoded at the 3' end of the antigenome (Brown et al., 1984; Lai et al., 1983; Spaan
et al., 1983). The leader/polymerase complex would disassociate from the template
and then realign at a complementary sequence (5'-AAUCUAAAC 3' or a closely
related sequence) in the intergenic regions on the antigenome (Baker and Lai, 1990;
Baric et al., 1983; Budzilowicz et al., 1985; Makino etal., 1986; Spaan etal., 1983).
This complex then primes transcription of the mRNAs. Since the intergenic regions
preceede every transcription unit, in MHV there are 6, this model of leader-primed
transcription thus leads to the nested set of coronavirus RNAs (Cheley et al., 1981;
Lai 1990; Lai etal., 1981; Leibowitz etal., 1981; Spaan etal., 1988). Recently it was
shown that other negative stranded RNAs are made during coronavirus transcription:
full-length copies of each mRNA, containing antileader and poly(U) (Hofman et al.,
1990; Sawicki and Sawicki 1990; Sethna et al., 1989, 1991). It has been
-7-

demonstrated that these subgenomic negative strands are transcriptionally active
(Sawicki and Sawicki, 1990; Schaad and Baric, 1994). Two new models were put
forward to explain these observations. In one, the leader-primed transcription is
considered to occur as thought previously. However, each mRNA that is synthesized
is then a template for minus-strand synthesis, leading to replicons for mRNA
amplification (Sethna etal., 1989). In the second model, subgenomic minus-strands
are believed to be transcribed first from the genomic RNA. The intergenic regions
now serve as leaky transcription termination signals (Sawicki and Sawicki, 1990).
This leads to a nested set of minus stranded RNAs, which is then the template for
the nested set of plus stranded RNAs. Addition of the leader occurs either by leader-
priming, as in the classic model, or by translocation of the minus strand/polymerase
complex to the leader sequence at the extreme end of the genome and continuation
of transcription.
Viral proteins and their genes
The structural proteins S, M and N, and if present, HE are the most abundantly
produced proteins. The S protein is encoded by mRNA 3 in MHV and is a 158 to 200
K glycoprotein (Siddell ef al., 1982). The S protein induces cell fusion in vitro and in
vivo, binds to the cell surface receptor and elicitates neutralizing antibody as well as
cell-mediated immunity (Boyle et al., 1987; Collins et al., 1982; Frana ef al., 1985;
Holmes et al., 1981; Sturman and Holmes 1981; Sturman ef al., 1985). For some
coronaviruses, the mature S protein is cleaved into two 90 KDa subunites S1 and S2
(e.g. MHV and BCV), whereas cleavage does not occur for others (e.g. FIPV, TGEV)
(reviewed in Spaan et al., 1988). The role of S protein cleavage in MHV
pathogenesis is controversial.
The M protein is encoded by mRNA 6 and is a 20 to 30 KDa integral membrane
protein (Siddell ef al., 1982). It is synthesized on the RER/Golgi membranes
(Armstrong et al., 1984; Rottier et al., 1986). The M protein is never transported to
-8-

the plasma membrane of the cell and accumulates in the RER/Golgi where it
interacts with the S protein. Monoclonal antibodies specific for the M protein
neutralize virus infectivity only in the presence of complement (Collins ef al., 1982).
The N protein is encoded by mRNA7 and is a 43 to 60 KDa phosphorylated
protein (Stohlman and Lai, 1979). The N protein probably encapsidates the RNA
genome by interacting with the leader sequence and many other sites of the viral
RNA to form a helical nucleocapside (Stohlman et al., 1988). The nucleocapsid then
interacts with the M protein and buds into the RER/Golgi where it is enveloped. The
interaction between the M protein and the nucleocapsid may determine the site of
virus budding (Machamer and Rose, 1987; Rottier and Rose, 1987; Tooze and
Tooze, 1985; Tooze ef al., 1984). The N protein has been shown to bind to the M
protein and also to the virion RNA in vitro (Baric et al., 1988; Masters et al., 1992;
Robbins etal., 1986; Stohlman etal., 1988; Sturman etal., 1980).
The HE protein is present only in some coronaviruses. It is encoded by mRNA 2-
1 in MHV and is present in the virus envelope as a 120 to 140 KDa disulfide-linked
dimer (King etal., 1985; Shieh etal., 1989; Yokomori etal., 1989). The HE protein is
responsible for hemagglutination by binding to 9-0-acetylated neuraminic acid
residues on red blood cells (Vlasak et al., 1988). The HE protein also has an
esterase activity which cleaves the acetyl group from 9-0-acetylated neuraminic acid
(Pfleiderer ef al., 1991; Schultzeef a/., 1991; Vlasak ef a/., 1988). The HE proteins of
coronaviruses have regions of high homology with the influenza C virus surface
glycoprotein, HEF (Luytjes ef al., 1988). The role of the HE protein in MHV
pathogenesis is unknown.
Recently, a 10 KDa protein encoded by the gene 5b has been determined to be
another envelope protein of MHV (Yu ef al., 1994). This protein has been named
small membrane (sM) or envelope protein (E). It has been shown clearly that E
protein plays an important role in virus budding (Bos etal., 1996).
-9-

RNA recombination
The ability to exchange genetic information may allow RNA viruses to adapt to a
changing environment and to overcome potential deleterious effects caused by the
high error frequency of the RNA polymerase. Viruses with segmented genomes can
undergo RNA reassortment. However, the ability of RNA viruses with nonsegmented
genomes to exchange genetic elements is more limited. Only a few RNA viruses,
including picornaviruses (Cooper 1968,1977; Cooper etal., 1975; King et al., 1982,
1985, 1987; Lake ef al., 1975), coronaviruses (Lai ef al., 1985; ), cowpea chlorotic
mottle virus 3a (Allison ef al., 1990), alphaviruses (Hahn ef al., 1988; Weiss and
Schlesinger, 1991) and bromoviruses (Bujarski and Kaesberg, 1986) have been
reported to undergo RNA-RNA recombination at various efficiencies. During a mixed
infection of two MHV strains, RNA recombination occurs at a high frequency both in
vivo and in vitro (Keck ef al., 1988; Lai ef al., 1985; Makino et al., 1986). RNA
recombination is believed to occur through a copy-choice mechanism in which the
polymerase and partially replicated genome dissociate from the orginal template,
associate with a different viral RNA molecule, and then complete the genome
replication (Fig 3). Based on the analysis of a set of ts-mutants, the frequency of
recombination was found to be very high for coronaviruses (up to 25% over the
whole genome) (Baric ef al., 1990). It is also shown that crossovers can occur more
than once within the genome (Baric et al., 1990). Recombination is almost random
and previously identified hot spots resulted from selection of recombinant viruses
(Banner ef a/., 1991).
Coronaviruses also undergo non-homologous recombination. The original
example of this is the HE pseudogene of MHV-A59 (Luytjes ef al., 1988). This gene
bears significant sequence similarities at the amino acid level with the HEF
sequence of influenza C virus (Luytjes ef al., 1988), and is believed to originate from
a non-homologous recombination event with influenza C HEF RNA. Defective
-10-
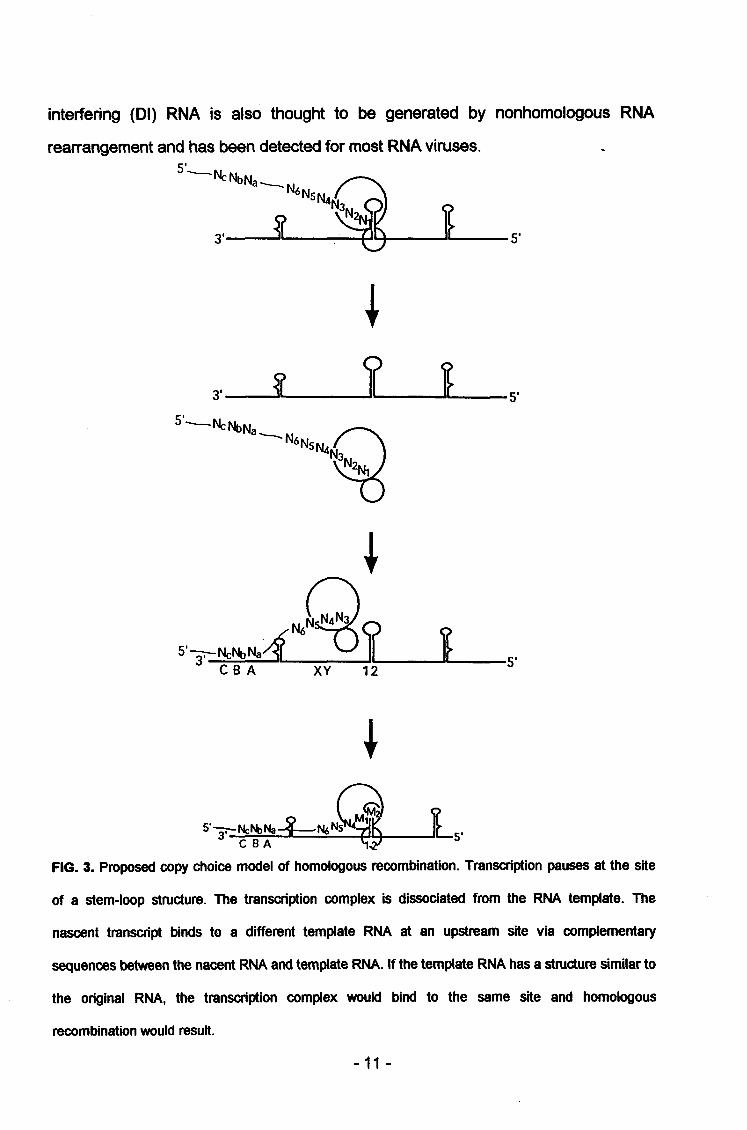
interfering (Dl) RNA is also thought to be generated by nonhomologous RNA
ibeen
NcNbNa.
rearrangement and has been detected for most RNA viruses
5'.
i
•N6n
3" Jl » 1 5'
5'—**N._M,_.
5'^-NcNbNa „ „ „
CBA XY 12
FIG. 3. Proposed copy choice model of homologous recombination. Transcription pauses at the site
of a stem-loop structure. The transcription complex is dissociated from the RNA template. The
nascent transcript binds to a different template RNA at an upstream site via complementary
sequences between the nacent RNA and template RNA. If the template RNA has a structure similar to
the original RNA, the transcription complex would bind to the same site and homologous
recombination would result.
-11-

Defective interfering RNAs
The genomes of defective interfering (Dl) particles have lost most of their coding
sequences but have retained the essential cis-acting replication and encapsidation
signals (Huang and Baltimore, 1970). Dl particles can only be replicated and
packaged in the presence of the standard viruses. As a result of the competition for
viral proteins and host factors, Dl viruses interfere with the replication of the
standard viruses (reviewed in Holland, 1991).
Coronavirus Dl RNAs have first been analyzed for MHV strain JHM (Makino ef
al., 1984, 1985). Several Dl RNAs with different sizes were found in virus infected
cells, but only the near-genome length DIssA was packaged into Dl particles (Makino
ef al., 1984). The other smaller Dl RNA, DIssE, apparently lacked a specific RNA
packaging signal since it was packaged inefficiently (Makino and Lai, 1989; Makino
ef al., 1988). So it was considered to be less suitable for molecular genetic studies.
Van der Most ef al. used a natural MHV-A59 Dl RNA (Dl-a) as a template to
construct a Dl cDNA clone, pMIDI. Molecular cloning and sequence analysis showed
that MIDI genome consists of three discontinuous fragments from the viral genome
(Fig. 4) (Van der Most et al., 1991). Dl RNAs transcribed from this clone and from its
derivatives, are replicated upon transfection into MHV-infected cells. The synthetic
Dl RNAs have proven to be excellent tools to investigate viral replication,
transcription, genome packaging and recombination.
Pathogenesis of MHV
MHV strains can be classified into two basic biotypes: Respiratory (polytropic)
and enterotropic (Barthold and Smith, 1984). After oronasal inoculation of infant mice
all of the prototype strains including MHV-1, -3, -A59, -JHM, and others, infect upper
respiratory (nasal) mucosa (Barthold, 1986; Compton ef al., 1993). If the infecting
MHV strain is sufficiently virulent or the mouse is susceptible due to age
-12-

I I MtZTJl AAAAn MHVA 59 genomic RNA2 3 45 6/7 32 KB
J—-P^AAAAn pMIDI RNA
3890 4689 5494 5.5 KB
FIG. 4. A comparison of the structures of pMIDI RNA and the MHV-A59 genomic RNA. Black bars
indicate sequences derived from the 5' end of the genome; the dotted bars represent sequences
derived from ORF1b, and the cross-hatched bars indicate sequences derived from the 3' end.
(preweanling), genotype, or immune status, the virus will disseminate readily to
multiple other organs (Barthold, 1986; Compton ef al., 1993). MHV-JHM has been
used as a model for the respiratory group of viruses because it is polytropic and
moderately virulent. After oronasal inoculation of genetically susceptible BALB/c
mice, virus initiates replication in the nasal epithelium. Virus apparently disseminates
via local lymphatics and blood, with replication in pulmonary vascular endothelium
and secondary viremia to multiple organs (Barthold and Smith, 1992). Virus can be
detected in a variety of tissues including nose, brain, vascular endothelium (lung and
elsewhere), bone marrow, lymphoid tissue (e.g., thymus, lymph node, spleen,
mucosa-associated lymphoid tissues), liver, uterus, placenta, peritoneum, and other
sites. Virus is detectable in intestine but is restricted to gut-associated lymphoid
tissue rather than mucosal epithelium. By contrast, genetically resistant SJL mice
develop significantly less severe infections and clear virus earlier than do BALB/c
mice, with significant virus dissemination and disease occurring only in mice infected
at 1 week of age. Respiratory strains of MHV can infect brain hematogenously during
disseminated infection in immunocompromised or infant mice or directly via olfactory
pathways from the nose to brain, even in the apparent absence of virus
-13-

dissemination to other organs (Barthold, 1988; Barthold ef al., 1986). Several
virulent and avirulent respiratory MHV strains can enter the brain via olfactory
pathways. Encephalitis is common among oronasally inoculated mice but
demyelination and chronic infection are largely experimental phenomena (Barthold,
1988; Barthold ef a/., 1986).
Enterotropic strains of MHV such as MHV-RI, -Y and -DVIM differ significantly
from respiratory MHV strains in their biologic behavior but cannot yet be
differentiated antigenically or genetically. Enterotropic MHV strains replicate
preferentially in intestinal mucosal epithelium and disseminate minimally to other
organs such as liver or brain (Barthold, 1987; Barthold ef al., 1993). This is in
contrast to respiratory strains of MHV which rarely infect intestinal mucosal
epithelium. Mice of all ages are susceptible to active enterotropic MHV infection, but
disease occurs only in very young mice. Oral inoculation of BALB/C and SJL mice at
1, 3, or 6 weeks of age with enterotropic MHV-Y results in nearly equal levels of
virus replication in intestinal mucosa regardless of age or genotype. Disease, in the
form of enteritis, is minimal in mice inoculated at 3 or 6 weeks of age and less severe
in SJL mice. Lesions may be restricted to the presence of a few enterosyncytia in the
surface mucosa of the ascending colon. The ascending colon produces the highest
virus titers and develops the most severe lesions in MHV-Y-infected mice. This also
seems to be true for other enterotropic MHV strains. Mice recover from infection with
no apparent carrier state. Thus, the resistance of SJL mice to respiratory MHV-JHM-
induced disease correlates with low virus titers in target tissues (Barthold and Smith,
1987). In contrast, SJL mice support high titer replication of enterotropic MHV with
only mild disease. Disease and virus titers peak at around 5 days in MHV-JHM-
infected mice, whereas MHV-Y-infected mice develop peak virus titers by 2 days
after inoculation. Enterotropic MHV strains tend not to disseminate from intestine to
other organs, but involvement of mesenteric lymph nodes occurs frequently.
Hepatitis may occur but not to the same extent as with respiratory strains of MHV.
Neonates, less than 48 hours old, infected with enterotropic MHV develop severe
-14-

necrotizing enterocolitis with high mortality within 2 days after inoculation. Epizootics
of enterotropic MHV in naive breeding populations can result in 100% mortality
among neonatal mice (Barthold etal., 1982).
Although the molecular mechanism of the MHV tissue tropism is still poorly
understood, several studies have shown an important role of the S protein in
determining the viral virulence and pathogenesis. The S protein appears to be one of
the most variable components of MHV. In addition to molecular diversity in S from
the various MHV strains, variability in the MHV S protein arises during passage
through rodents, generating antigenic and pathogenic variants (Dalziel ef al., 1986;
Fleming ef a/., 1986; Gallagher ef al., 1990; Haspel etal., 1978; Knobleref a/., 1982;
La Monica ef a/., 1991; Massa ef al., 1988; Matsubara ef a/., 1991; Morris ef al.,
1989; Parker et al., 1989; Robb etal., 1979; Taguchi etal., 1985, Wang etal., 1992).
A study on the MHV strains MHV-1, -3, -S, -JHM, and A59 demonstrated the
presence of five antigenic sites, A through E, among which B is involved with
neurotropism and C with variance (Talbot and Buchmeier, 1985). Such analyses also
uncovered polymorphism in the S protein of MHV-JHM variants isolated after
escaping neutralization by MAb (Buchmeier etal., 1988; Dalziel etal., 1986). Some
isolates obtained in this manner were attenuated and others more virulent when
compared with the parental JHM. Attenuation was manifested through a requirement
for larger inocula to initiate central nervous system disease and a change in
preferential tropism from neurons to oligodendrocytes. Thus, a paralytic,
demyelinating disease rather than an acute encephalitis was elicited (Buchmeier ef
al., 1988).
Aim of the research and scope of the thesis
The goal of this research was to gain insight into the molecular aspects of
enterotropic MHV strains, focusing on the role of S protein in determining tissue
tropism.
-15-

In chapter 2 the S genes of enterotropic strains MHV-RI and -Y were cloned and
sequenced. A high degree of homology was found between the MHV-RI and MHV-
JHM S proteins, whereas the homology that was found between the MHV-Y S protein
and the S proteins of other MHV strains was much lower. These results indicate that
if the spike protein is indeed determining the biotype of a strain only a few amino
acid changes may be responsible for tissue tropism. The fact that the S protein of
MHV-Y is inefficiently cleaved may be due to the presence of several amino acid
changes upstream from the predicted cleavage site.
In chapter 3 the presence and position of open reading frames (ORFs) 4, 5a and
5b of three enterotropic strains were determined. They were found to be very similiar
to those of MHV-JHM, except that due to a frameshift ORF 5a of MHV-Y terminated
after merely 63 nucleotides. The proteins, if present, were found to be highly
conserved among all strains. No biotype-specific pattern could be detected.
In chapter 4 the MHV-RI S gene cDNA was inserted in-frame into pMIDI, a full
length cDNA clone of a MHV-A59 Dl, yielding pDPRIS. RNA in vivo transcribed from
pDPRIS could be replicated and passaged efficiently in MHV-A59 infected L cells. In
the absence of positive selection pressure, a highly specific and sensitive RT-PCR
was used for the detection of recombinant RNAs. Recombinant genomic RNA was
detected in intracellular RNA from pDPRIS transfected and MHV-A59 infected cells
in total lysates as well as in agarose gel purified genomic RNA from these fractions.
Six total RNA PCR products and five virion RNA PCR products were cloned and
sequenced. Crossovers between A59 and Rl RNA were found ranging from 112
nucleotides from the 5' border to 89 nts from the 3' border of sequence homologous
between A59 and Rl S genes. Homologous RNA recombination occurred between
the genomic RNA template and the synthetic Dl RNA template at different locations,
generating a series of MHV recombinant RNAs with chimeric S genes.
In chapter 5 a number of applications of PCR in the study of enterotropic MHV
are described. A universal diagnostic PCR was developed which detected all of 11
different MHV strains. This fast and reliable method was also able to differentiate
-16-

MHV from other non-murine coronaviruses. On the basis of this assay a quantitative
PCR was designed using a mutant template containing a point mutation which
competed for the PCR primers. The amplication and cloning of the structural protein
genes of enterotropic MHV strains in plasmid vectors for subsequent sequencing is
described.
In chapter 6 I describe a strategy which takes advantage of RNA recombination
occurring at high frequency between MHV helper virus and synthetic interfering (Dl)
RNA. A synthesic Dl containing MHV-RI S gene cDNA clone, pDPRIS, was
constructed. RNA in vivo transcribed from the plasmid was passaged in MHV-A59
infected cells. Using a highly specific and sensitive RT-PCR, recombinant genomic
RNA was detected in both intracellular and virion RNA. In addition, the same
intracellular RNA was isolated and separated on a low melting point agarose gel.
The band containing the MHV genomic RNA was excised and the subsequently
eluted RNA yielded a positive signal in the recombinant-specific RT-PCR. Since the
purified genomic RNA did not contain DPRIS RNA, it confirmed that homologous
recombination between MHV-A59 RNA and synthetic Dl RNA had indeed taken
place.
-17-






![2016 [Advances in Virus Research] Coronaviruses Volume 96 __ Interaction of SARS and MERS Coronaviruses with the Antivir](https://img.dokumen.tips/doc/110x75/613ca6cf9cc893456e1e874c/2016-advances-in-virus-research-coronaviruses-volume-96-interaction-of-sars.jpg)










![2016 [Advances in Virus Research] Coronaviruses Volume 96 __ Feline Coronaviruses](https://img.dokumen.tips/doc/110x75/613ca6ce9cc893456e1e874a/2016-advances-in-virus-research-coronaviruses-volume-96-feline-coronaviruses.jpg)











