Embed Size (px)
Citation preview

THE ANATOMICAL RECORD 243:109-119 (1995)
lmmunocytochemical Localization of the Protein Reactive to Human p- 1 ,4-G alactosy I t ransf e rase Anti bodies During Chick Embryonic
Skin Differentiation YOSHIHIRO AKIMOTO, AKIKO OBINATA, HIROYOSHI ENDO,
KIYOSHI FURUKAWA, DAISUKE AOKI, SHIRO NOZAWA, AND HIROSHI HIRANO Department of Anatomy, Kyorin University School of Medicine, Mitaka, Tokyo (Y.A. ,
H.H.); Department of Physiological Chemistry, Faculty of Pharmaceutical Sciences, Teikyo University, Sagamiko, Kanagawa (A.O., H.E.); Department of Obstetrics and Gynecology, Keio University School of Medicine, Shinanomachi, Shinjuku-ku, Tokyo (DA. , S.N.); and Department of Biosignal Research, Tokyo Metropolitan Znstitute of Gerontology, Sakae-cho,
Itabashi-ku, Tokyo (K.F.); Japan
ABSTRACT Background P-1, 4-Galactosyltransferase (GalTase) trans- fers galactose from UDP-galactose to terminal N-acetylglucosamine in gly- coconjugates and is located both in the Golgi apparatus and in the plasma membrane. The cell surface GalTase is thought to be involved in cell-to-cell recognition and cell-to-extracellular matrix interaction.
Methods: By the use of specific monoclonal antibodies against human GalTase, changes in cell surface localization of the protein reactive to the antibodies in chick embryonic skin during its differentiation in vivo and in vitro were detected immunohistochemically at both light- and electron mi- croscopic levels. The distribution of glycoconjugates having terminal N-acetylglucosamine residues was detected by staining with succinylated wheat germ agglutinin (s-WGA).
Results: Under the light microscope, intense immunostaining was ob- served in the keratinized epidermis, particularly in the intermediate layer. Marked changes in the localization of the staining were observed in vitamin A-induced mucus-secreting skin, in which keratinization was suppressed. The localization of the immunostaining was in parallel with that of glyco- conjugates having terminal N-acetylglucosamine residues.
Immunoelectron microscopically the immunostaining was located on the cell surface and in the intercellular space of the desmosomes in the inter- mediate cells of the keratinized epidermis. However, the staining was not present on the cell surface but was detected on the limiting membrane of the mucous granules, in the mucous metaplastic epidermis. In contrast, the staining was always found in the Golgi apparatus in all of the cells. Conclusions: These results suggest that the protein reactive to human
GalTase antibody may be involved in chick epidermal differentiation. 0 1995 Wiley-Liss, Inc.
Key words: Antibody against human 0-1, 4-Galactosyltransferase, Immu- nocytochemistry, Chick embryonic skin, Differentiation, Keratinization, Mucous metaplasia
In our investigations of the differentiation processes of chick embryonic tarsometatarsal skin in vivo and in vitro, we observed marked changes in the desmosome (Takata et al., 1981), basement membrane (Hirano et al., 1985; Akimoto et al., 1988), plasma membrane (Takata and Hirano, 1983), and intercellular matrix (Akimoto et al., 1992). By using plant lectins as a probe, changes in the glycoconjugates on the cell sur- face of epidermis have also been shown to occur during differentiation of chick embryonic skin (Takata and Hirano, 1983). Moreover, we found that changes in
localization of endogenous (3-galactoside-binding lec- tins occur during the skin differentiation in association with those in distribution of glycoconjugates on the cell surface (Hirano et al., 1988; Oda et al., 1989; Akimoto et al., 1992,1993). During the period of the skin differ-
Received March 24, 1994; accepted March 2, 1995. Address reprint requests to Professor Hiroshi Hirano, Department
of Anatomy, Kyorin University School of Medicine, Mitaka, Tokyo 181, Japan.
0 1995 WILEY-LISS, INC

110 Y. AKIMOTO ET AL.
entiation, in which keratinization starts from day 13 in ovo (stage 39; Hamburger and Hamilton, 1951) and is completed by day 17 in ovo (stage 43), the changes in glycoconjugates on the cell surface are closely related to the skin differentiation processes.
There have been reported several mechanisms by which cells interact with either neighboring cells or extracellular matrix or with both (Takeichi, 1988; Sharon and Lis, 1989; Lasky et al., 1992). One of these is mediated by the cell surface carbohydrates. Glyco- syltransferases (Roseman, 1970) and lectins (Sharon and Lis, 1989) including selectins (Lasky et al., 19921, are known to be receptors for this type of the cell rec- ognition.
p-1, 4-Galactosyltransferase (GalTase, EC 2. 4. 1, 22), which is an enzyme that transfers galactose from UDP-galactose to terminal N-acetylglucosamine in glycoconjugates (Trayer and Hill, 1971; Aoki et al., 19901, is located both in Golgi apparatus, more pre- cisely in its trans region, and in the plasma membrane (Pestalozzi et al., 1982; Roth and Berger, 1982; Roth et al., 1985; Hartel-Schnenke et al., 1991; Suganuma et al., 1991; Watzele et al., 1991; Aoki et al., 1992; Cooke and Shur, 1994; Kawano et al., 1994). Several lines of evidence suggest that the cell surface GalTase is in- volved in cell to cell recognition in the early embryo and cell to extracellular matrix interaction such as cell migration and fertilization (Romagnano et al., 1990; Miller et al., 1991; Shur, 1991; Eckstein and Shur, 1992).
Little is known about the cell surface expression and distribution of GalTase during skin differentiation. In the present study, we detected the protein reactive to human GalTase antibodies on the cell surface during epidermal keratinization but not mucous metaplasia induced in chick embryonic skin in vivo andlor in vitro. The similar distribution of glycoconjugates having ter- minal N-acetylglucosamine residues was also observed using s-WGA (succinylated wheat germ agglutinin) as a probe during the differentiation.
MATERIALS AND METHODS Skin
Tarsometatarsal skin of 13- and 17-day-old chick em- bryos was used. In some cases, skin explants from the tarsometatarsal region of 13-day-old chick embryos were cultured for 9 days in a chemically defined me- dium, BGJb (Biggers et al., 1961), containing 5% deli- pidized fetal calf serum (dFCS) and 20 nM hydrocorti- sone hemisuccinate (Japan Upjohn, Ltd., Tokyo) by the Millipore filter-roller-tube method (Sugimoto et al., 1974). Under these conditions, keratinization of the epidermis occurs in vitro similarly as in ovo (Takata et al., 1981). Or the skin explants were first cultured for 1 day in BGJb medium containing 5% dFCS, 20 nM hydrocortisone, and 20 pM vitamin A (Sigma, St. Louis, MO), and then in BGJb supplemented with 2 mM Bt,cAMP (Sigma, St. Louis, MO) for 8 days by the same method. As reported previously, keratinization was suppressed and mucous metaplasia was induced by the addition of vitamin A in vitro (Hirano et al., 1985; Obinata et al., 1991). Moreover, vitamin A-induced epi- dermal mucous metaplasia was accelerated by the ad- dition of Bt,cAMP (Obinata et al., 1991). The medium was renewed every other day.
Monoclonal Antibody We used mouse monoclonal antibodies (MAb 8513
and MAb 8628) against human GalTase (Uejima et al., 1992; Uemura et al., 1992). Uejima et al. (1992) re- ported previously on the epitopes of these monoclonal antibodies by using the recombinant GalTase ex- pressed in Escherichia coli. The epitopes of the mono- clonal antibodies resides on the protein moiety of GalTase but not on the carbohydrate moiety (Uejima et al., 1992).
lrnmunoblotting Procedures Seventeen-day-old chick embryonic tarsometatarsal
skin was dissected, homogenized in PBS containing 4 mM 2-mercaptoethanol, mixed with Laemmli sampling solution, and boiled for 3 min as described previously (Akimoto et al., 1992). The homogenate was subjected to SDS-polyacrylamide gel electrophoresis (PAGE, 12.5% gel) and transferred to nitrocellulose sheets (Towbin et al., 1979). The sheets after blocking with bovine serum albumin were incubated with a mouse monoclonal antibody against human GalTase (Uemura et al., 1992) or normal mouse IgG or IgM, followed by horseradish peroxidase (HRP)-conjugated goat anti- mouse IgG or IgM. The protein band was visualized by immersion in 3,3’-diaminobenzidine.4HCl (DAB, 0.2 mg/ml)-H,O, (0.005%) for 5 min at room tempera- ture.
lmrnunoprecipitation and GalTase Assay Since the monoclonal antibody used in the present
study was raised against human GalTase (Uemura et al., 19921, whether or not the antibody binds to the chick embryo transferase was studied. Livers (3 g) from 17-day-old chick embryos were homogenized in 3 ml of phosphate-buffered saline (PBS), pH 7.2, containing 1% Triton X-100. The homogenate was centrifuged at 1,500g for 15 min. An aliquot (100 p1) of the super- natant was incubated with mouse monoclonal anti- body 8628 (100 pl containing 3.5 mg protein) at 4°C for 60 min. The mixture was further incubated with pro- tein A-Sepharose suspension (v/v, 1:l) (200 p1) at 4°C for 60 min. After centrifugation at 600g for 1 min, the immunoprecipitates were washed with PBS. The final pellet was suspended in 50 p1 of 20 mM Mes buffer, pH 6.5. GalTase activity was determined as described previously (Furukawa et al., 1990). In brief, the transferase assays were conducted in 50 p1 of 20 mM Mes buffer, pH 6.5, containing 3 mM 5’-AMP, 5 mM 2, 3-dimercapto-1-propanol, 0.05% Nonidet P-40, 10 mM MnCl,, 100 pM UDP-L3H1Gal (323 pcilmmol), 250 pg asialo-agalacto-transferrin as an acceptor, and the immunoprecipitated material as an enzyme source. Mixtures were incubated at 37°C in a shaking water bath for 60 min, and the reaction was terminated by adding 70 p1 of bovine serum albumin (1 mg) and 30 p1 of 50% trichloroacetic acid (TCA). After standing at 0°C for 60 min, reaction mixtures were transferred to Whatman micro glass filters, and the filters were washed thoroughly with 5% TCA. Radioactivities on the filters were determined by a liquid scintillation counter as described previously (Furukawa and Roth, 1985).

LOCALIZATION OF THE PROTEIN REACTIVE TO GALTASE ANTIBODY 111
lmmunostaining for Light Microscopic Observation Tarsometatarsal skin specimens of 13- and 17-day-
old chick embryos were fixed in 4% formaldehyde in 0.1 M phosphate buffer (pH 7.3) for 1 h at 4°C. The speci- mens were subsequently immersed in 2.3 M sucrose-10 mM PBS, and then frozen in liquid nitrogen. Frozen sections of 4 pm thickness were cut with a coldtome CM-41 (Sakura, Ltd, Tokyo), washed with PBS, and treated for 10 min with 1% bovine serum albumin (BSA) in PBS. Sections were incubated with a mouse monoclonal antibody against GalTase or with normal mouse IgG or IgM for 1 h at room temperature, washed with PBS, incubated with the HRP-conjugated goat anti-mouse IgG or M for 1 h, washed again with PBS, and then immersed in DAB (0.2 mg/ml)-H,Oz (0.005%) for 5 min at room temperature. Finally sections were rinsed in distilled water, dehydrated, cleared, and mounted. Nonspecific staining was checked by the in- cubation with the HRP-conjugated goat anti-mouse IgG or IgM alone. To detect endogenous peroxidase ac- tivity, other sections were incubated with DAB-H,O, solution alone. To further confirm the specificity of the staining, we also carried out pre-absorption and com- petition experiments. The antibody (10 pg/ml) was pre- incubated with excess human milk GalTase (50 pg/ml, Sigma, St. Louis, MO) for 24 h a t 4°C and then applied to the sections.
Lectin Staining for Light Microscopic Observation Specimens were fixed overnight with Bouin’s solu-
tion and embedded in paraffin. Four-micrometer sec- tions were cut, deparaffinized with xylene, and dehy- drated through a graded series of ethanols. Lectin binding sites were visualized with biotinylated lectin (s-WGA: succinylated wheat germ agglutinin, Vector Lab, Inc., Burlingame, CA) and avidin-biotin-peroxi- dase complex (ABC) reagent (Vector Lab Inc., Burlin- game, CA). In this procedure the sections were first treated for 10 min with 1% BSA in PBS and then in- cubated with biotinylated lectin at a concentration of 25 pg/ml in 0.1% BSA-PBS for 30 min at room temper- ature. After a wash in PBS, the sections were incu- bated with ABC reagent for 30 rnin at room tempera- ture, washed again with PBS, and then immersed in DAB-H202 for 5 rnin at room temperature. Finally the sections were rinsed in water, dehydrated, cleared, and mounted. To confirm the specificity of lectin staining, some sections were preincubated with an appropriate hapten sugar (N-acetylglucosamine) at a concentration of 0.2 M for 30 min and then incubated with biotinyl- ated lectin in the presence of 0.2 M hapten sugar. To detect endogenous peroxidase activity, other sections were incubated with DAB-H2O2 solution alone. Non- specific binding of ABC reagent was also checked by incubation, first with ABC reagent, and then with DAB-H,02 solution.
lmmunostaining for Electron Microscopy Specimens were fixed in 4% formaldehyde in 0.1 M
phosphate buffer (pH 7.3) for 1 h at 4°C. After washing, the specimens were frozen and cut into 40 pm slices with a cryomicrotome. Slices were incubated overnight at 4°C with the antibody, washed with PBS, and incu- bated for 1 h at room temperature with HRP-conju-
gated goat anti-mouse IgG or IgM. For a cytochemical control, normal mouse IgG or IgM was used. The spec- imens were washed with PBS again and fixed in 2.5% glutaraldehyde in 0.1 M phosphate buffer, pH 7.4, for 10 rnin to obtain the better preservation of the ultra- structure and to immobilize the HRP-conjugates. After another washing with PBS, the specimens were im- mersed in DAB-H,02 for 5 min at room temperature, and then washed with distilled water. Finally the spec- imens were osmicated for 30 min, dehydrated through a graded series of ethanols, and embedded in Epon 812. Ultrathin sections were cut, stained with lead citrate, and observed in a JEM-1200EX electron microscope. Nonspecific staining was checked by the incubation with the secondary antibody-HRP conjugate alone. Some skin explants were fixed in 2.5% glutaraldehyde and processed for conventional electron microscopic ob- servation.
nzwc rs Western Blot Analysis Using the Antibody
In order to characterize the protein reactive to hu- man GalTase antibody, 17-day-old chick embryonic skin homogenate solubilized with Triton-X 100 was subjected to SDS-PAGE and then immunoblotted (Fig. 1). The immunoblot result showed that the anti- GalTase antibody detected a 68-kDa protein (Fig. la), which is in good agreement with the molecular weight of chick embryonic liver GalTase reported previously (Furukawa and Roth, 1985; Hathaway and Shur, 1992). No protein band was detected with preimmune mouse IgG or IgM instead of the specific antibody (Fig. lb).
lmmunoprecipitation of the Protein With Monoclonal Antibody
Since the monoclonal antibody used in the present study was raised against human GalTase (Uemura et al., 19921, whether or not the protein reactive to the antibody contains the GalTase activity was investi- gated. Solubilized liver homogenate prepared from chick embryos was incubated with the antibody, and GalTase activity was assayed using the immuno- precipitated material. Only a small amount of the radioactivity was incorporated into the asialo-agalacto- transferrin acceptor (data not shown). Tritium galacto- sylated transferrin glycopeptides were isolated by ConA-Sepharose column chromatography after ex- haustive digestion of reaction mixtures with pronase. Tritium-labeled galactose was released by digestion of the glycopeptides with diplococcal p-galactosidase, which cleaves the Galpl-4GlcNAc but not the Galpl- 3GlcNAc linkage (Glasgow et al., 1977), indicating that galactose was transferred to N-acetylglucosamine residues via pl-4 linkage (data not shown). Since the antibody is specific to human GalTase and since the immunoprecipitated enzyme activity of chick embryo is low with this antibody, whether or not the protein re- active to the antibody is chicken GalTase has to be further determined.
Light Microscopic Observation With the Antibody In skin specimens obtained from 13-day-old chick
embryos, immunostaining was observed at the apical surface of the epidermis and, a t a very low level, in the

112 Y. AKIMOTO ET AL.
116K- 97K- 66K-
45K-
29K-
a b
Fig. 1. Immunoblotting of skin lysates with the use of anti-GalTase antibody (MAb 8513). Seventeen-day-old chick embryonic skin ex- plants were lysed, electrophoresed on 12.5% SDS polyacrylamide gel under reducing conditions, and transferred to nitrocellulose. Proteins were immunostained with the antibody. A single 68-kDa band is de- tected (a). No staining was observed with normal mouse IgG or IgM (b).
cell surface of the epidermis (Fig. 2a). As the embryo developed, the staining became more intense; and the location of the protein was more clearly detected on the cell surface. In the epidermis keratinized both in vivo (17-day-old chick embryo) and in vitro, intense stain- ing was observed along the cell surface of cells in the superficial and intermediate layers (Fig. 2b,c), while staining was weak in keratinized and the basal cell layers. The epidermal-dermal junctional zone also showed moderate staining. A positive staining was ob- served in the superficial cells of the epidermis in which keratinization had been suppressed and mucous meta- plasia had been induced by vitamin A (Fig. 2d).
When the antibodies were omitted or replaced with the preimmune mouse IgG or IgM, no positive staining was observed. All the positive staining was disap- peared when 17-day-old chick embryonic skin was in- cubated with the antibody in the presence of human milk GalTase (Fig. 2e).
Light Microscopic Observation With s- WGA In order t o elucidate the localization of glycoconju-
gates which contain N-acetylglucosamine residues at the non-reducing termini, lectin staining was con- ducted using s-WGA which specifically recognizes a terminal N-acetylglucosamine residue. Staining was observed at the apical surface of the epidermis and, at a very low level, along the cell surface of the epidermis of 13-day-old chick embryo (Fig. 3a). In the skin kera- tinized both in vivo and in vitro, the superficial and intermediate cell layers were positively stained (Fig. 3b,c). However, both keratinized and basal cells were
not stained (Fig. 3b,c). The epidermal-dermal junc- tional zone was scarcely stained. In the mucous meta- plastic epidermis, the cytoplasm of the superficial cells was intensely stained, while intermediate and basal cells were not stained (Fig. 3d). The lectin binding was completely inhibited by the addition of the haptenic sugar, N-acetylglucosamine (Fig. 3e). Neither endoge- nous peroxidase activity nor other non-specific binding were observed.
Electron Microscopic Observation With the Antibody The precise localization of the protein reactive to the
antibody was studied by electron microscope. In the 13-day-old chick embryo, the epidermis consisted of 3-5 cell layers and was not keratinized yet (Fig. 4a). The cell surface of intermediate cells were stained weakly (Fig. 4b). In contrast, in the 17-day-old chick embryonic epidermis, in which several layers of the superficial cells were keratinized (Fig. 5a), staining in- tensity was increased significantly. In the superficial and intermediate cell layers, intense staining was ob- served along the cell surface and in the intercellular space of the desmosomes (Fig. 5b,c). The cell surface of the basal cell layer was stained at its basal side as well as the basement membrane (Fig. 5d).
The staining pattern in the epidermis in which kera- tinization had been induced in vitro by hydrocortisone was the same as that of the 17-day-old embryonic epi- dermis in vivo (Fig. 6a,b). In the epidermis in which mucous metaplasia had been induced by vitamin A, many mucous granules were observed in the cytoplasm of cells in the superficial layer (Fig. 6c). Positive stain- ing was detected on the limiting membrane of the mu- cous granules, in the Golgi apparatus, and on a part of the nuclear membrane (Fig. 6d). The limiting mem- brane of mucous granules was more intensely stained by MAb 8513 than MAb 8628. In the intermediate layer, staining was scarcely observed or the cell surface or in the intercellular space of the desmosomes.
Positive staining was not observed when incubated with preimmune mouse IgG or IgM instead of the spe- cific antibody (Fig. 7a,b). Neither endogenous peroxi- dase activity nor non-specific binding of the secondary antibody was observed.
DISCUSSION GalTase is mainly localized in Golgi apparatus
where it is involved in the synthesis of complex carbo- hydrates by concerted actions of many other glycosyl- transferases. GalTase is also found on cell surfaces of a variety of cells, and implicated to be involved in cell to cell and cell to matrix interactions (Pierce et al., 1980). In fact, the cell surface GalTase is shown to promote cell adhesion as well as cell migration (Shur, 1989; Hathaway and Shur, 1992). GalTase in the plasma membrane of mouse sperm binds to polylactosamino- glycans of the zona pellucida glycoproteins (Lopez et al., 1989; Macek et al., 1991; Miller et al., 1991). In the early stage of mouse embryonic development, cell sur- face GalTase is involved in compaction and the adhe- sion between blastomeres (Bayna et al., 1988; Hatha- way et al., 1989). Binding between cell surface GalTase and polylactosaminoglycans is considered to be in- volved in the implantation of embryos to the uterus

LOCALIZATION OF THE PROTEIN REACTIVE TO GALTASE ANTIBODY 113
Fig. 2. Immunohistochemical localization by light microscopy of the protein reactive to the antibody in chick embryonic skin from 13- (a) and 17-day-old (b) embryos, and in the skin of 13-day-old embryos cultured in the presence of hydrocortisone for 9 days (c) or hydrocorti- sone and vitamin A for 1 day and then Bt,cAMP for 8 days (d). Stained with mouse anti-GalTase antibody and with HRP-conjugated goat anti-mouse IgG or IgM. Arrowheads indicate the basal surface of the epidermis. P, periderm. (a) In the undifferentiated skin from a 13-day-old embryo, positive staining is observed a t a low level. In 17-day-old (b) and hydrocortisone-treated (c) chick embryonic epider-
mis, in which keratinization had taken place, intense staining is ob- served mainly in the intermediate layers. Positive staining is also seen in the epidermal-dermal junction. d: Positive staining is found in the superficial cells of the mucous metaplastic epidermis induced by vitamin A treatment in vitro. In the superficial cells of the epidermis, patchy staining (arrows) is observed in the cytoplasm. e: Seventeen- day-old chick embryonic skin was preincubated with antigen, then incubated with anti-GalTase antibody in the presence of antigen. Posi- tive staining is noticeably diminished. x 900.

114 Y. AKIMOTO ET AL.
Fig. 3. Staining of 13- and 17-day-old chick embryonic skin (a, b) and cultured skin explants (c, d) with s-WGA. a: Thirteen-day-old embryonic skin. The epidermal cells are scarcely stained. b: Seven- teen-day-old embryonic skin. The superficial and intermediate cells are intensely stained. c: Skin that was keratinized in vitro by treat- ment with hydrocortisone for 9 days. The cell membrane of interme- diate cells is intensely positive for the s-WGA reaction. d Mucous
metaplastic skin that was induced in vitro by treatment with hydro- cortisone and vitamin A for 1 day and then with Bt,cAMP for 8 days. The cytoplasm of superficial cells is positively stained (arrows). e: A cytochemical control. Seventeen-day-old embryonic skin was incu- bated with s-WGA in the presence of 0.2 M N-acetylglucosamine. Positive reaction is completely inhibited. Arrowheads indicate the basal surface of the epidermis. P, periderm. x 800.
(Dutt et al., 1987) and in cell adhesion of embryonal carcinoma (Shur, 1983).
In the present study, the localization of the protein
reactive to human GalTase antibody was found in the Golgi apparatus and on the cell surface of chick embry- onic skin. Changes in the cell surface localization of the

LOCALIZATION OF THE PROTEIN REACTIVE TO GALTASE ANTIBODY 115
Fig. 4. Electron micrographs of epidermis from the tarsometatarsal region of 13-day-old chick embryos. a: Osmium-fixed and Epon-em- bedded section of the epidermis. The epidermis consists of 3-5 cell layers and it not keratinized yet. S, superficial cells; I, intermediate cells; B, basal cells; P, peridermal cells, and D, dermis. b: Immuno- cytochemical localization by electron microscopy of the protein in the
intermediate cells. Formaldehyde-fixed and frozen sections were re- acted with mouse anti-GalTase antibody, then, with HRP-conjugated goat anti-mouse IgG or IgM, and processed for transmission electron microscopy. Plasma membrane of intermediate cells is stained weakly (arrows). a, X 1,600; b, X 24,000.
protein were observed immunocytochemically in the chick embryonic tarsometatarsal skin in association with the differentiation and metaplasia both in vivo and in vitro. The localization of the protein in the kera- tinized epidermis was the same as that of the endoge- nous P-galactoside-binding lectins previously reported (Akimoto et al., 1992, 1993). The protein was also de- tected in the limiting membrane of mucous granules in chick embryonic mucous metaplastic epidermis.
In general, changes in carbohydrate structures of glycoconjugates have important roles in embryonic de- velopment and differentiation (Kawai et al., 1979; Barnes, 1988). These changes have been also observed in developing chick embryonic epidermis (Takata and Hirano, 1983). In the undifferentiated epidermis of the 13-day-old embryo, glycoconjugates terminating with N-acetylglucosamine, galactose, and N-acetylgalacto- samine residues are not detected by cytochemical stud- ies. As the epidermis develops toward keratinization, however, these glycoconjugates accumulate on the cell surface of the intermediate cells. The expression of these glycoconjugates chronologically correspond to the appearance of the protein reactive to human GalTase and P-galactoside-binding lectin on the cell surface. The protein is also present in desmosomes which are well developed in the keratinized epidermis (Takata et al., 1981). In the desmosomes which are not well devel- oped in the mucous metaplastic epidermis (Hirano et
al., 1985; Obinata et al., 1991), glycoconjugates con- taining terminal N-acetylglucosamine residues do not appear on the cell surface, and the protein is hardly detected on the cell surface or in the intercellular space of desmosomes. These results indicate that in chick em- bryonic skin the antibody reactive protein and glyco- conjugates terminating N-acetylglucosamine residues might play important roles in the skin differentiation probably by mediating specific cellular interactions.
The protein was also located in the epidermal-dermal junctional region. However, s-WGA staining was scarcely observed in the epidermal-dermal junctional region. This might be due to the masking of terminal N-acetylglucosamine residues with other sugar resi- dues at this region.
Immunostaining of the protein in the superficial cells of the epidermis was increased in association with the mucous metaplastic changes in vitro. Immunoelec- tron microscopic observation revealed that the protein was located in the limiting membrane of the mucous granules in addition to the Golgi apparatus in the mu- cous metaplastic epidermis.
Positive staining was also observed on the nuclear membranes of the metaplastic epidermis cultured in the presence of vitamin A. This was only detected in the cultured epidermis. The localization of GalTase in the nuclear membranes was already reported in cul- tured F9 embryonal cells (Suganuma et al., 1991). Al-

116 Y. AKIMOTO ET AL.
Fig. 5. Electron micrographs of epidermis from the tarsometatarsal region of 17-day-old chick embryos. a: Osmium-fixed and Epon-em- bedded sections of the epidermis. Several layers of the superficial cells (S) are keratinized. Peridermal cells (P) are above the keratinized layer (K). I, intermediate cells; B, basal cells; and D, dermis. b-d. Immunocytochemical localization by electron microscopy of the pro- tein in the epidermis. Formaldehyde-fixed and frozen sections were reacted with mouse anti-GalTase antibody, then, reacted with HRP-
conjugated goat anti-mouse IgG or IgM, and processed for transmis- sion electron microscopy. In the superficial (b) and intermediate (c) layers, the plasma membrane (arrows in b) and intercellular spaces of the desmosomes (arrows in c) are stained. d: At the boundary between the epidermis and the dermis, the basal plasma membrane of the basal cells, the basal lamina (arrows), and immediate environs (ar- rowheads) are stained. B, basal cell; CF, collagen fibers; N, nucleus. a , x 1,700; b, ~25 ,000 ; c, ~20 ,000 ; d, ~31 ,000 .

LOCALIZATION O F THE PROTEIN REACTIVE TO GALTASE ANTIBODY 117
Fig. 6. Electron micrographs of epidermis from the tarsometatarsal region of chick embryonic epidermis cultured in the presence of hy- drocortisone for 9 days (a and b), or hydrocortisone and vitamin A for 1 day and then Bt,cAMP for 8 days (c and d). Immunocytochemical localization by electron microscopy of the protein (b and d). Formal- dehyde-fixed and frozen sections were reacted with mouse anti- GalTase antibody and then reacted with HRP-conjugated goat anti- mouse IgG or IgM, and processed for transmission electron microscopy. a: Osmium-fixed and Epon-embedded section of the kera- tinized epidermis. Epidermis consists of superficial (S), intermediate (I), and basal (B) layers. Keratinized cells (K) are seen in the super- ficial layer of the epidermis. D, dermis. b: In the intermediate layer of
the epidermis, in which keratinization was induced by hydrocorti- sone, the plasma membrane (arrows) is intensely stained. c: Osmium- fixed and Epon-embedded section of the epidermis. Epidermis consists of superficial (S), intermediate (I), and basal (B) layers. No kerati- nized layer is seen in the epidermis. D, dermis. d: Positive staining is detected in the limiting membrane of the mucous granules (MI, in the Golgi complex (G), and in part of the nuclear membranes (arrow- heads) of the superficial cells of the mucous metaplastic epidermis. Desmosomes (DE) and the cell surface are scarcely stained. a, x 1,800; b, x 9,300; c, x 1,700; d, x 14,000. Inset Staining is positive in the Golgi apparatus (G) as indicated by arrows. x 27,500.
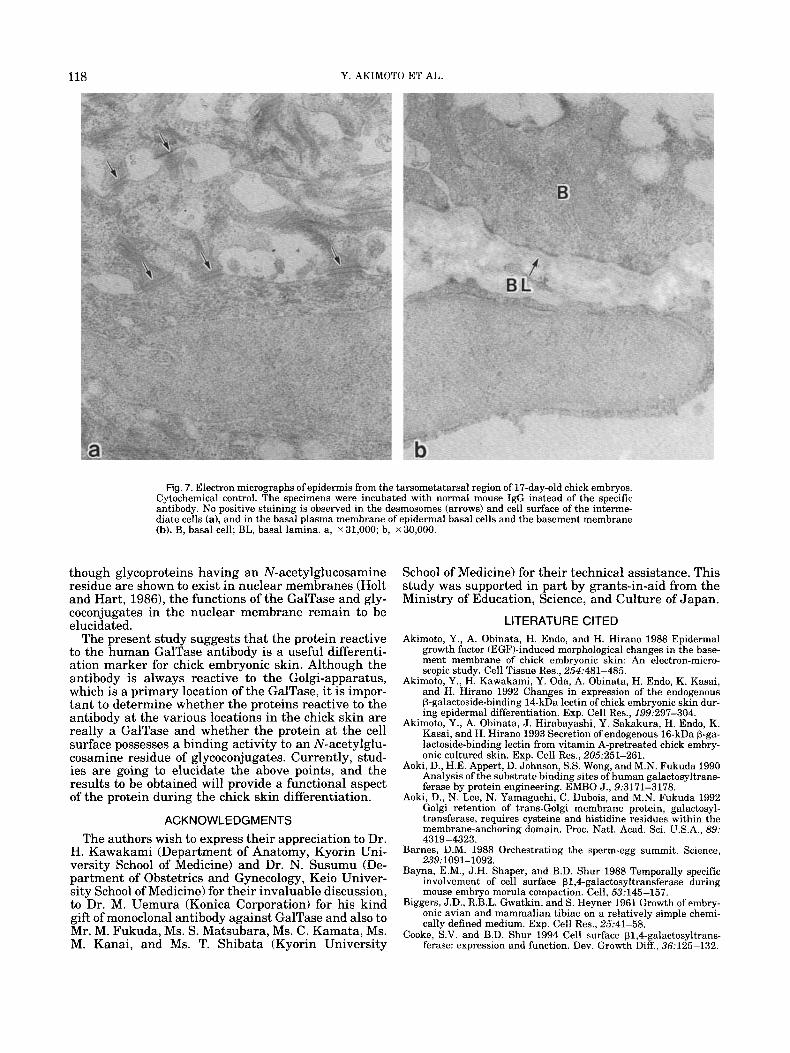
118 Y. AKIMOTO ET AL.
Fig. 7. Electron micrographs of epidermis from the tarsometatarsal region of 17-day-old chick embryos. Cytochemical control. The specimens were incubated with normal mouse IgG instead of the specific antibody. No positive staining is observed in the desmosomes (arrows) and cell surface of the interme- diate cells (a), and in the basal plasma membrane of epidermal basal cells and the basement membrane (b). B, basal cell; BL, basal lamina. a, x 31,000; b, x 30,000.
though glycoproteins having an N-acetylglucosamine residue are shown to exist in nuclear membranes (Holt and Hart, 1986), the functions of the GalTase and gly- coconjugates in the nuclear membrane remain to be elucidated.
The present study suggests that the protein reactive to the human GalTase antibody is a useful differenti- ation marker for chick embryonic skin. Although the antibody is always reactive to the Golgi-apparatus, which is a primary location of the GalTase, i t is impor- tant to determine whether the proteins reactive to the antibody a t the various locations in the chick skin are really a GalTase and whether the protein at the cell surface possesses a binding activity to an N-acetylglu- cosamine residue of glycoconjugates. Currently, stud- ies are going to elucidate the above points, and the results to be obtained will provide a functional aspect of the protein during the chick skin differentiation.
ACKNOWLEDGMENTS
The authors wish to express their appreciation to Dr. H. Kawakami (Department of Anatomy, Kyorin Uni- versity School of Medicine) and Dr. N. Susumu (De- partment of Obstetrics and Gynecology, Keio Univer- sity School of Medicine) for their invaluable discussion, to Dr. M. Uemura (Konica Corporation) for his kind gift of monoclonal antibody against GalTase and also to Mr. M. Fukuda, Ms. S. Matsubara, Ms. C . Kamata, Ms. M. Kanai, and Ms. T. Shibata (Kyorin University
School of Medicine) for their technical assistance. This study was supported in part by grants-in-aid from the Ministry of Education, Science, and Culture of Japan.
LITERATURE CITED Akimoto, Y., A. Obinata, H. Endo, and H. Hirano 1988 Epidermal
growth factor (EGFI-induced morphological changes in the base- ment membrane of chick embryonic skin: An electron-micro- scopic study. Cell Tissue Res., 254:481-485.
Akimoto, Y., H. Kawakami, Y. Oda, A. Obinata, H. Endo, K. Kasai, and H. Hirano 1992 Changes in expression of the endogenous p-galactoside-binding 14-kDa lectin of chick embryonic skin dur- ing epidermal differentiation. Exp. Cell Res., 199~297-304.
Akimoto, Y., A. Obinata, J . Hirabayashi, Y. Sakakura, H. Endo, K. Kasai, and H. Hirano 1993 Secretion of endogenous 16-kDa p-ga- lactoside-binding lectin from vitamin A-pretreated chick embry- onic cultured skin. Exp. Cell Res., 205:251-261.
Aoki, D., H.E. Appert, D. Johnson, S.S. Wong, and M.N. Fukuda 1990 Analysis of the substrate binding sites of human galactosyltrans- ferase by protein engineering. EMBO J., 9:3171-3178.
Aoki, D., N. Lee, N. Yamaguchi, C. Dubois, and M.N. Fukuda 1992 Golgi retention of trans-Golgi membrane protein, galactosyl- transferase, requires cysteine and histidine residues within the membrane-anchoring domain. Proc. Natl. Acad. Sci. U.S.A., 89: 4319-4323.
Barnes, D.M. 1988 Orchestrating the sperm-egg summit. Science, 239:1091-1092.
Bayna, E.M., J.H. Shaper, and B.D. Shur 1988 Temporally specific involvement of cell surface pl,4-galactosyltransferase during mouse embryo morula compaction. Cell, 53t145-157.
Biggers, J.D., R.B.L. Gwatkin, and S. Heyner 1961 Growth of embry- onic avian and mammalian tibiae on a relatively simple chemi- cally defined medium. Exp. Cell Res., 25:41-58.
Cooke, S.V. and B.D. Shur 1994 Cell surface pl,4-galactosyltrans- ferase: expression and function. Dev. Growth Diff., 36:125-132.

LOCALIZATION OF THE PROTEIN
Dutt, A., J.-P. Tang, and D.D. Carson 1987 Lactosaminoglycans are involved in uterine epithelial cell adhesion in vitro. Dev. Biol., 119:27-37.
Eckstein, D.J. and B.D. Shur 1992 Cell surface p-1,4-galactosyltrans- ferase is associated with the detergent-insoluble cytoskeleton on migrating mesenchymal cells. Exp. Cell Res., 201:83-90.
Furukawa, K., K. Matsuta, F. Takeuchi, E. Kosuge, T. Miyamoto, and A. Kobata 1990 Kinetic study of a galactosyltransferase in the B cells of patients with rheumatoid arthritis. Internat. Immunol., 2:105-112.
Furukawa, K. and S. Roth 1985 Co-purification of galactosyltrans- ferases from chick-embryo liver. Biochem. J., 227r573-582.
Glasgow, L.R., J.C. Paulson, and R.L. Hill 1977 Systematic purifica- tion of five glycosidases from Streptococcus (Diplococcuus) pneu- moniue. J . Biol. Chem., 252:8615-8623.
Hamburger, V. and H.L. Hamilton 1951 A series of normal stages in the development of the chick embryo. J . Morphol., 88:49-92.
Hartel-Schnenk, S., N. Minnifield, W. Reutter, C. Hanski, C. Bauer, and D.J. Morre 1991 Distribution of glycosyltransferases among Golgi apparatus subfractions from liver and hepatomas of the rat. Biochim. Biophys. Acta, 1115:108-122.
Hathaway, H.J., L.C. Romagnano, and B.S. Babiarz 1989 Analysis of cell surface galactosyltransferase activity during mouse trophec- todermal differentiation. Dev. Biol., 134:351-361.
Hathaway, H.J. and B.D. Shur 1992 Cell surface pl,4-galactosyl- transferase functions during neural crest cell migration and neu- rulation in vivo. J . Cell Biol., 117:369-382.
Hirano, H., Y. Akimoto, H. Kawakami, Y. Oda, and K. Kasai 1988 Localization of endogenous p-galactoside-binding lectin in em- bryonic chick epidermis. In: Glycoconjugates in Medicine. Ohvama M. Muramatsu T. eds. Professional Posteraduate Ser- vicis, Tokyo, pp. 8-13. ’
Hirano, H., N. Oishi, A. Obinata, and H. Endo 1985 Metaplastic changes in chick embryonic skin induced by vitamin A. In: Base- ment Membranes. Shibata s, ed. Elsevier, Amsterdam, pp. 395- 406.
-
Holt, G.D. and G.W. Hart 1986 The subcellular distribution of termi- nal N-acetylglucosamine moieties. J . Biol. Chem., 261:8049- 8057.
Kawai, N., F. Nishiyama, and H. Hirano 1979 Changes of lectin- binding sites on the embryonic muscle cell surface in the devel- oping ascidian, Hulocynthiu uuruntium. Exp. Cell Res., 122:293- 304.
Kawano, J., S. Ide, T. Oinuma, and T. Suganuma 1994 A protein- specific monoclonal antibody to rat liver pl-4 galactosyltrans- ferase and its application to immunocytochemistry. J . Histochem. Cytochem., 42~363 -369.
Lasky, L.A. 1992 Selectins: Interpreters of cell-specific carbohydrate information during inflammation. Science, 258:964-969.
Lopez, L.C., C.M. Maillet, K. Oleszkowicz, and B.D. Shur 1989 Cell surface and golgi pools of ~-1,4-galactosyltransferase are differ- entially regulated during embryonal carcinoma cell differentia- tion. Mol. Cell Biol., 9:2370-2377.
Macek, M.B., L.C. Lopez, and B.D. Shur 1991 Aggregation of p-1,4- galactosyltransferase on mouse sperm induces the acrosome re- action. Dev. Biol., 147:440-444.
Miller, D.J., M.B. Macek, and B.D. Shur 1991 Complementarity be- tween sperm surface p-1,4- galactosyltransferase and egg-coat ZP3 mediates sperm-egg binding. Nature, 357:589-593.
Obinata, A,, Y. Akimoto, H. Hirano, and H. Endo 1991 Stimulation by Bt,cAMP of epidermal mucous metaplasia in retinol-pretreated chick embryonic cultured skin, and its inhibition by herbimycin A, an inhibitor for protein-tyrosine kinase. Exp. Cell Res., 193: 36-44.
Oda, Y., Y. Ohyama, A. Obinata, H. Endo, and K. Kasai 1989 Endog-
REACTIVE TO GALTASE ANTIBODY 119
enous p-galactoside-binding lectin expression is suppressed in retinol-induced mucous metaplasia of chick embryonic epidermis. Exp. Cell Res., 182:33-43.
Pestalozzi, D.M., M. Hess, and E.G. Berger 1982 Immunohistochem- ical evidence for cell surface and Golgi localization of galactosyl- transferase in human stomach, jejunum, liver, and pancreas. J . Histochem. Cytochem., 30:1146-1152.
Pierce, M., E.A. Turley, and S. Roth 1980 Cell surface glycosyltrans- ferase activities. Int. Rev. Cytol., 65:l-47.
Romagnano, L. and B. Babian 1990 The role of murine cell surface galactosyltransferase in trophoblast: Laminin interactions in uztro. Dev. Biol., 241.254-261.
Roseman, S. 1970 The synthesis of complex carbohydrate by multi- glycosyltransferase systems and their potential function in inter- cellular adhesion. Chem. Phys. Lipids, 5:270-297.
Roth, J . and E.G. Berger 1982 Immunocytochemical localization of galactosyltransferase in Hela cells: codistribution with thiamine pyrophosphatase in trans-Golgi cisternae. J . Cell Biol., 93:223- 229.
Roth, J., M.J. Lentze, and E.G. Berger 1985 Immunocytochemical demonstration of ecto-galactosyltransferase in absorptive intes- tinal cells. J . Cell Biol., 100:118-125.
Sharon, N. and H. Lis 1989 Lectins as cell recognition molecules. Science, 246:227-234.
Shur, B.D. 1989 Expression and function of cell surface galactosyl- transferase. Biochem. Biophys. Acta 988:389-409.
Shur, B.D. 1991 Mini Review: Cell surface p1,4 galactosyltransferase: Twenty years later. Glycobiology, 1:563-575.
Shur, B.D. 1983 Embryonal carcinoma cell adhesion: The role of sur- face galactosyltransferase and its 90K lactosaminoglycan sub- strate. Dev. Biol., 99:360-372.
Suganuma, T., H. Muramatsu, T. Muramatsu, K. Ihida, J. Kawano, and F. Murata 1991 Subcellular localization of N-acetylglu- cosaminide p1+4 galactosyltransferase revealed by immunoelec- tron microscopy. J . Histochem. Cytochem., 39:299-309.
Sugimoto, M., K. Tajima, A. Kojima, and H. Endo 1974 Differential acceleration by hydrocortisone of the accumulation of epidermal structural proteins in the chick embryonic skin growing in a chemically defined medium. Dev. Biol., 39:295-307.
Takata, K., A. Obinata, H. Endo, and H. Hirano 1981 Induction of the a-type keratinization by hydrocortisone in embryonic chick skins grown in a chemically defined medium: An electron microscopic study. Dev. Biol., 85:370-379.
Takata, K. and H. Hirano 1983 Changes in soybean agglutinin (SBA) and peanut agglutinin (PNA) binding pattern in the epidermis of the developing chick embryo. Dev. Growth. Diff., 25r299-305.
Takeichi, M. 1988 The cadherins: Cell-cell adhesion molecules con- trolling animal morphogenesis. Development, 102:639-655.
Towbin, H., T. Staehelin, and J . Gordon 1979 Electrophoretic transfer of proteins from polyacrylamide gels to nitrocellulose sheets: Pro- cedure and some applications. Proc. Natl. Acad. Sci. U.S.A., 76: 4350-4354.
Trayer, I.P. and R.L. Hill 1971 The purification and properties of the A protein of lactose synthetase. J . Biol. Chem., 246:6666-6675.
Uejima, T., M. Uemura, S. Nozawa, and H. Narimatsu 1992 Comple- mentary DNA cloning for galactosyltransferase associated with tumor and determination of antigenic epitopes recognized by spe- cific monoclonal antibodies. Cancer Res., 52:6158-6163.
Uemura, M., T. Sakaguchi, T. Uejima, S. Nozawa, and H. Narimatsu 1992 Mouse monoclonal antibodies which recognize a human (pl- 4)galactosyltransferase associated with tumor in body fluids. Cancer Res, 52:6153-6157.
Watzele, G., R. Bachofner, and E.G. Berger 1991 Immunocytochemi- cal localization of the Golgi apparatus using protein-specific an- tibodies to galactosyltransferase. Eur. J . Cell Biol., 56:451-458.





























