Embed Size (px)
Citation preview

1
1 Identification of a 5’-Deoxyadenosine Deaminase in Methanocaldococcus 2 jannaschii and its Possible Role in Recycling the Radical SAM Enzyme 3
Reaction Product 5’-Deoxyadenosine 4 5 6 7 Danielle Miller, Kaitlin O’Brien, Huimin Xu and Robert H. White# 8 9 10 11 12 13 Department of Biochemistry, Virginia Polytechnic Institute and State University, 14 Blacksburg, Virginia 24061 15 16 17 #To whom correspondence should be addressed. Telephone: (540) 231-6605, Fax: 18 (540) 231-9070, e-mail: [email protected] 19 20 Running title: a new pathway for recycling SAM radical enzyme reaction products 21 22
JB Accepts, published online ahead of print on 27 December 2013J. Bacteriol. doi:10.1128/JB.01308-13Copyright © 2013, American Society for Microbiology. All Rights Reserved.
on July 29, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

2
ABSTRACT 23 Here we characterize the MJ1541 gene product from Methanocaldococcus 24 jannaschii, an enzyme that was annotated as a 5’-methylthioadenosine/S-25 adenosylhomocysteine deaminase (EC 3.5.4.31/3.5.4.28). The MJ1541 gene product 26 catalyzes the conversion of 5’-deoxyadenosine to 5’-deoxyinosine as its major product, 27 but will also deaminate 5’-methylthioadenosine, S-adenosylhomocysteine, and adenosine 28 to a small extent. On the basis of these findings we are naming this new enzyme 5’-29 deoxyadenosine deaminase (DadD). The KM for 5’-deoxyadenosine was found to be 14.0 30 ± 1.2 μM with a kcat/KM of 9.1 x 109 M-1s-1. Radical SAM enzymes account for nearly 31 2% of the M. jannasshii genome, where the major SAM derived products is 5’-32 deoxyadenosine. Since 5’-dA has been demonstrated to be an inhibitor of radical SAM 33 enzymes; a pathway for removing this product must be present. Here we propose that 34 DadD is involved in the recyclization of 5’-deoxyadenosine where upon the 5’-35 deoxyribose moiety of 5’-deoxyinosine is further metabolized to deoxyhexoses used for 36 the biosynthesis of aromatic amino acids in methanogens. 37 38
on July 29, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

3
ABBREVIATIONS 39 5’-deoxyadenosine (5’-dA), 5’-methylthioadenosine (MTA), S-adenosylhomocysteine 40 (SAH), 5’-deoxyadenosine deaminase (DadD), 5’-deoxyinosine (5’-dI), 5’-41 methylthioadenosine/S-adenosylhomocysteine deaminase (MtaD), 5’-42 methylthioadenosine deaminase (MtaD), S-adenosylhomocysteine deaminase (SAHD), S-43 adenosylmethionine (SAM), 5’-methylthioinosine (MTI), S-inosylhomocysteine (SIH), 44 methylthioadenosine/S-adenosylhomocysteine nucleosidase (MtaN), adenosine (Ad), 45 inosine (In), ethylenediaminetetraacetic acid (EDTA), sodium dodecyl sulfate- 46 polyacrylamide gel electrophesis (SDS-PAGE), phosphoenolpyruvate (PEP), 5-47 deoxyribose 1-phosphate (5-dRP), erythrose 4-phosphate (E-4-P), 3-dehydroquinate 48 (DHQ), 6-deoxy-5-ketofructose-1-phosphate (DKFP). 49 50 51 on July 29, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

4
INTRODUCTION 52 On the basis of sequence comparison, the MJ1541 gene product has been 53
annotated as a 5’-methylthioadenosine/S-adenosylhomocysteine deaminase (EC 3.5.4.31/ 54 3.5.4.28) (MtaD) (1) and would thus be expected to catalyze the deamination of the 55 adenine-containing nucleotide products of some SAM radical enzymes. Homologues of 56 this enzyme from Thermotoga maritima, Pseudomonas aeruginosa, and Streptomyces 57 flocculus (1-3) have each been enzymatically characterized using 5’-methylthioadenosine 58 (MTA), S-adenosyl-L-homocysteine (SAH), and adenosine (Ad) as substrates, but never 59 5’-deoxyadenosine (5’-dA). Here we show that the MJ1541 gene encodes a metal-60 dependent 5’-deoxyadenosine deaminase (DadD) that specifically catalyzes the 61 deamination of 5’-dA (Figure 1). 62 In a recent search of the Methanocaldococcus jannaschii genome for possible 63 radical SAM enzymes we found that 35 of the 1811 protein encoding genes (almost 2% 64 of the genome) contain the canonical CX3CX2C motif indicating a possible radical SAM 65 enzyme. Most of these gene products have no annotated function other than being 66 identified as radical SAM enzymes. Radical SAM enzymes are found in many aspects of 67 metabolism such as the biosynthesis of cofactors, antibiotics, and in the repair of DNA 68 (4). Since a major portion of radical SAM enzymes proceed via a mechanism that leads 69 to the formation of 5’-dA (4), then in an organism with 35 radical SAM enzymes it seems 70 likely that there would be a salvage pathway for the metabolism of the 5’-dA. The 71 presence of a salvage pathway is important because it has been shown that both biotin 72 synthase (BioB) and lipoyl synthase (LipA) are strongly inhibited by 5’-dA (4). 73
on July 29, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

5
The importance of DadD in the salvage of this radical SAM enzyme reaction 74 product is that in methanogens the MTA/SAH nucleosidase [EC 3.2.2.9], which normally 75 metabolizes these compounds, is absent. The MTA/SAH nucleosidase works to remove 76 adenine from the modified ribose sugar. Without the presence of this enzyme, 77 methanogens instead perform an initial deamination step to salvage the byproducts of the 78 radical SAM reactions (Figure 1). 79 MATERIALS AND METHODS 80 Chemicals. 5’-Deoxyadenosine, 2’-deoxyadenosine, 5’-methylthioadenosine, S-81 adenosylhomocysteine, and adenosine were obtained from Sigma/Aldrich. 82 Cloning, Overexpression, and Purification of M. jannaschii MJ1541 Gene 83 Product in E. coli. The MJ1541 gene (Swiss-Prot accession number Q58936) was 84 amplified by PCR from genomic DNA using oligonucleotide primers MJ1541-Fwd: 5'-85 GGTCATATGATATTGATAAAAAATG-3' and MJ1541-Rev: 5'-86 GCTGGATCCTTAGCTTCTCAAAATC-3'. PCR amplification was performed as 87 described previously (5) using a 45 oC annealing temperature. Purified PCR product was 88 digested with Nde1 and BamH1 restriction enzymes and ligated into compatible sites in 89 vector pT7-7. Sequence of the resulting plasmid, pMJ1541, was verified by DNA 90 sequencing. pMJ1541 was transformed into E. coli strain BL21-CodonPlus(DE3)-RIL 91 cells (Stratagene). The transformed cells were grown in LB medium (200 mL) 92 supplemented with 100 µg/mL ampicillin at 37 oC with shaking until they reached an 93 OD600 of 1.0. Recombinant protein production was induced by addition of lactose to a 94 final concentration of 28 mM (5). After an additional 2 hours of culture, the cells (200 95 mL) were harvested by centrifugation (4,000 x g, 5 min) and frozen at -20 oC. Induction 96
on July 29, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

6
of the desired protein was confirmed by SDS-polyacrylamide gel electrophoresis (SDS-97 PAGE) analysis of total cellular proteins. 98 The frozen E. coli cell pellet containing the desired protein (~0.4 g wet wt) was 99 suspended in 3 mL of extraction buffer (50 mM N-[tris(hydroxymethyl)methyl]-2-100 aminoethanesulfonic acid (TES), pH 7.0, 10 mM MgCl2, 20 mM DTT) and lysed by 101 sonication. The resulting expressed protein was found to remain soluble after heating the 102 resulting cell extracts for 10 min at 70 oC followed by centrifugation (16,000 x g) for 10 103 min. This heating step allowed for the purification of the recombinant enzymes from the 104 majority of E. coli proteins, which denature and precipitate under these conditions. The 105 next step of purification was performed by anion-exchange chromatography of the 70 oC 106 soluble fractions on a MonoQ HR column (1 x 8 cm; Amersham Bioscience) using a 107 linear gradient from 0 to 1 M NaCl in 25 mM Tris buffer (pH 7.5), over 55 min at a flow 108 rate of 1 mL/min. Fractions of 1 mL were collected and the desired protein was 109 identified through SDS-PAGE analysis of the fractions. Protein concentrations was 110 determined by Bradford analysis (6). 111
Recombinant Expression of the MJ1541 gene product in the presence of 112 Ni(II), Zn(II), Co(II), and Fe(III). The metal containing protein was recombinantly 113 produced by overexpression in E. coli grown in Luria-Bertani medium (200 mL), as 114 described above, supplemented with either 1 mM NiCl2, ZnCl2, CoCl2, or Fe(NH4)2SO4. 115 Expression under these conditions allowed for production of much larger amounts of 116 soluble enzyme as judged by the intensity of the protein band upon SDS-PAGE analysis 117 of the cell extracts. 118 Site Directed Mutagenesis. The MJ1541 C294S, Y136R, E150R, and 119
on July 29, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

7
Y136R/E150R were amplified by PCR from genomic DNA using oligonucleotide 120 primers: MJ1541-C294S-Fwd : 5-CTTAGGAACTGATGGAAGTG-121 GAAGTAACAACAAC-3', MJ1541-C294S -Rev : 5-GTTGTTGTTACTTCCACTTC-122 CATCAGTTCCTAAG-3', MJ1541-Y136R-Fwd : 5'-GAGGGCAGTTTTAGC-123 CCGCGGAATGATTGATTTATTTG-3', MJ1541-Y136R-Rev : 5'-124 CAAATAAATCAATCATTCCGCGGGCTAAAACTGCCCTC-3', MJ1541-E150R-Fwd 125 : 5'- GAGGAGAGAAGGGAGAGACGGCTTAAAAATGCTGAGAAG - 3', MJ1541-126 E150R-Rev : 5'-CTTCTCAGCATTTTTAAGCCGTCTCTCCCTTCTCTC-CTC-3, 127 MJ1541-Y136R/E150R-Fwd : using the MJ1541 -Y136R as the template and using 128 MJ1541-E150R-Fwd. as the primer, MJ1541 -Y136R/E150R -Rev. using MJ1541 -129 Y136R as the template using MJ1541 -E150R -Rev. as the primer. Overexpression of the 130 mutant proteins was done as described above. 131
MALDI-MS Identification of SDS-PAGE Protein Bands. The protein bands 132 corresponding to the predicted molecular mass of the MJ1541 gene product were excised 133 from the polyacrylamide gel, the cut gel bands were de-stained with 50 mM ammonium 134 bicarbonate and water (LC-MS grade) twice for 3 hours. Then the gel pieces were 135 dehydrated in acetonitrile (LC-MS grade) for another 5 hours. Finally, the acetonitrile 136 was removed by centrifugation. Dehydrated gel pieces were rehydrated on ice using 5 μl 137 proteomics-grade porcine trypsin10 ng/μl in 25 mM ammonium bicarbonate. Once the 138 gel pieces had rehydrated, the digests were incubated approximately 4 hours at 37 oC. 139 Liquid was collected in the bottom of the tube by centrifugation at 10 x g for 5 minutes at 140 room temperature. Digests were then incubated in a sonicating water bath for 15 minutes 141 and the liquid was again collected at the bottom of the tube by centrifugation. 142
on July 29, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

8
Approximately 1 μl of each digest was spotted onto a stainless steel MALDI plate. Once 143 dry the digests were overlaid with 1 μl matrix solution. The matrix solution consisted of 144 4 mg/ml α-cyano-4-hydroxycinnamic acid prepared in a 50:50 mixture of LC-MS grade 145 solvents acetonitrile and water (Spectrum Chemical) supplemented with 0.1% (v/v) 146 trifluoroacetic acid and 10 mM ammonium chloride. 147
Once the matrix had dried the digests were analyzed by MALDI using an Applied 148 Biosystems (AB) Sciex 4800 MALDI Tof/Tof mass spectrometer. First an MS spectrum 149 was obtained for each digest for the m/z range of 800 to 4000 using the MS reflector 150 positive operating mode. The MS spectra were typically the average of 1000 laser shots. 151 A data dependent acquisition of the MS/MS spectra for the 10 most intense peaks was 152 then performed utilizing the MS/MS 1kV positive operating mode. Each MS/MS 153 spectrum was typically the average of 3500 laser shots. The MALDI software (4000 154 Series Explorer, AB Sciex) was then used to generate peak lists for searching on the 155 public Mascot server (Matrix Science). 156
Measurement of Native Molecular Weight of DadD. The native molecular 157 weight of DadD was determined by size exclusion chromatography as described 158 previously using a Superose 12 HR column (7). 159
Metal Analysis of Metal Expressed DadD. Metal analysis of DadD was 160 performed at the Virginia Tech Soil Testing Laboratory using inductively coupled plasma 161 emission spectrophotometry. Instrumentation included a Spectro CirOS VISION made 162 by Spectro Analytical Instruments equipped with a Crossflow nebulizer with a Modified 163 Scott spray chamber, nebulizer rate was 0.75 L/min. A 50 mg/L yttrium internal standard 164
on July 29, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

9
was introduced by peristaltic pump. Samples were analyzed for cobalt, copper, iron, 165 manganese, nickel, and zinc. 166
Analysis of DadD Activity and DadD Mutants. The enzymatic activity of 167 DadD was determined by incubating 9 ng of the expressed DadD, Y136R, E150R, 168 Y136R/E150R mutants, and each metal expressed DadD with 0.4 mM 5’-deoxyadenosine 169 (5’-dA) in 50 mM BIS TRIS propane (pH 9.0) in a total volume of 100 µL for 10 min at 170 60 oC. The enzymatic reaction was stopped by the addition 5 µL 2 M perchloric acid 171 followed by centrifugation (16,000 x g) for 5 min. The separated soluble material was 172 then analyzed by HPLC. To determine if the activity of DadD is linear with time, a time 173 course experiment was done. In this experiment the standard assay was used to measure 174 DadD activity with incubation times of 5, 10, 15, 20, and 30 minutes. 175
The chromatographic analysis of 5’-dA and 5’-dI was performed on a Shimadzu 176 HPLC System with a C18 reverse phase column (Varian Pursuit XRs 250 × 4.6 mm, 5 177 μm particle size, operated at RT) equipped with photodiode array detection. The elution 178 profile consisted of 5 min at 95% sodium acetate buffer (25 mM, pH 6.0, 0.02% NaN3) 179 and 5% methanol followed by a linear gradient to 50% sodium acetate buffer/50% 180 methanol over 25 min at 0.5 mL/min. Under these chromatographic conditions the 181 following nucleotides eluted in the order: S-inosylhomocysteine (SIH) at 10.8 minutes, 182 inosine at 12.3 minutes, S-adenosylhomocysteine (SAH) at 14.5 minutes, 5’-dI at 16 183 minutes, adenosine (Ad) at 17.2 minutes, 5’-dA at 20 minutes, methylthioinosine (MTI) 184 at 22 minutes, and 5’-methylthioadenosine (MTA) at 26 minutes. Quantitation was based 185 on absorbance at 260 nm for the adenosine and 248 nm for the inosine containing 186
on July 29, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

10
compounds using ε260 = 14900 M-1cm-1 for adenosine and ε250 = 12300 M-1cm-1 for 187 inosine (8). 188
The chromatographic separation of 5’-dA and 5’-dI was also performed on the 189 same Shimadzu HPLC System with a Phenomex Kinetex (C18 reverse phase column 100 190 x 4.6 mm, 2.6 µm particle size) with an elution profile that consisted of 1 min at 98% 191 sodium acetate buffer and 2% methanol followed by a linear gradient to 70% sodium 192 acetate buffer/30% methanol over 19 minutes, then another linear gradient to 20% 193 sodium acetate buffer/80% methanol over 5 minutes, followed by an isocratic flow at 194 20% sodium acetate buffer/80% methanol for 5 minutes at 0.5 mL/min. Under these 195 conditions 5’-dI eluted at 12 minutes and 5’-dA eluted at 16 minutes. The quantitation 196 was based on peak areas. 197
Temperature Stability of DadD and Mutants. To determine the temperature 198 stability of the enzymes, each was pre-incubated for 10 min at 25, 50, 60, and 70 °C and 199 the remaining activity was assayed using the standard assay at 60 °C. 200
Determination of pH Optimum for the wild-type DadD. The pH optium was 201 determined over the pH range 6.5 to 12.5 using three different buffer systems: 50 mM 202 BIS TRIS propane, pH 6.5 to 9.0, 50 mM diethanolamine, pH 9.0 to 10.0, and 50 mM 203 sodium phosphate, pH 11.0 to 12.5, using the standard assay with 0.2 mM 5’-dA. 204 Effect of added divalent metals on DadD activity. To determine if the activity 205 of DadD, was affected by the addition of metals, the enzyme was assayed in the presence 206 of 0.05 mM ZnCl2, MnCl2, CoCl2, NiCl2, CuCl2, or Fe(NH4)2(SO4)2· 6H2O added to the 207 standard assay mixture as aqueous solutions. 208 Substrate Specificity of DadD Activity. To establish the substrate specificity of 209
on July 29, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

11
the native DadD the enzyme and its mutants were assayed with different concentrations 210 of 5’-dA, MTA, SAH, and Ad under the standard assay conditions. 211 Effect of DTT on DadD Activity. To determine if DTT had an effect on the 212 activity of the native DadD and C294S mutant the standard assay was performed with the 213 addition of 0.02 M DTT to the assay. This concentration of DTT has been used 214 previously with other metal-dependent DTT dependent enzymes (7). 215
Effect of Chelating Agents on DadD Activity. To determine if the presence of a 216 chelating agent would have an effect on the native and nickel expressed DadD activity, 217 25 mM ethylenediaminotetracetic acid (EDTA) was added to the standard assay. The 218 native and nickel-expressed DadD were also pre-incubated for 52 hours at RT in the 219 presence of 25 mM EDTA prior to the activity assay to establish if the metal had been 220 removed from the protein by monitoring the activity of the enzyme before and after the 221 incubation. The nickel-expressed DadD was also dialyzed in Buffer A (50 mM 2-(N-222 morpholino)ethanesulfonic acid sodium salt, 1 mM EDTA, 50 mM sodium sulfate, and 223 25 mM dipicolinic acid) for three days at RT with stirring. The buffer was switched out 224 for fresh Buffer A after 24 and 48 hours. Then Buffer B (same as dialysis buffer A 225 except without dipicolinic acid) was exchanged for Buffer A after 36 hours. Fresh Buffer 226 B was exchanged for the old buffer 3 times at 12 hours intervals to ensure the complete 227 removal of dipicolinic acid from sample. The activity of the dialyzed enzyme was then 228 checked following the standard assay described above. This procedure was modified 229 from that reported by Chistopherson et al.(9). 230 Phylogenetic Analysis of DadD and its homologues. The evolutionary history 231 was inferred using the Neighbor-Joining method (10). The optimal tree with the sum of 232
on July 29, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

12
branch length = 21.00517872 is shown (next to the branches). The evolutionary distances 233 were computed using the Poisson correction method (11) and are in the units of the 234 number of amino acid substitutions per site. The analysis involved 22 amino acid 235 sequences. All positions containing gaps and missing data were eliminated. There were a 236 total of 408 positions in the final dataset. Evolutionary analyses were conducted in 237 MEGA5 (12). 238 RESULTS: 239
Cloning, Overexpression, and Purification of MJ1541 gene product in E. coli. 240 The MJ1541 derived protein was highly expressed in E. coli as measured from the SDS-241 PAGE analysis of the total proteins in the E. coli cells after expression. MALDI analysis 242 of the tryptic peptides recovered from this spot confirmed the presence of the desired 243 protein. After sonication of the E. coli cells (3 mL) suspended in extraction buffer and 244 centrifugation (14,000 × g) 5 min, SDS-PAGE analysis of the soluble material and the 245 pellet demonstrated that most of the expressed MJ1541 protein was found in the soluble 246 extract. Heating portions of the resulting crude soluble extract for 10 min at different 247 temperatures indicated that DadD precipitated at temperatures above 70 °C based on 248 SDS-PAGE analysis. Thus, the first step in the purification of the native enzyme was 249 heating of the sonicated cell extract to 70 °C for 10 min prior to purification of the extract 250 on MonoQ. DadD purified by this method showed one peak of activity eluting at 0.42 M 251 NaCl. SDS-PAGE analysis showed the peak was greater than 80 % pure. 252
Recombinant Expression of DadD in the presence of Ni(II), Zn(II), Fe(III), or 253 Co(II). In order to try to increase the amount of soluble protein expressed and alter the 254 proteins metal content, the expression of the protein was conducted in LB medium to 255
on July 29, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

13
which either 1 mM Ni(II), Zn(II), Fe(II), or Co(II) had been added. On the basis of the 256 measured enzymatic activity in the purified enzymes, the total DadD activity was found 257 to be the same when expressed without additional metals, or expressed in the presence of 258 Ni(II). The activity found in the enzymes expressed with added Zn(II) and added Fe(II) 259 were 2-fold higher than expressed with Ni(II) or without additional metal. The reason for 260 the two fold higher activity remains unclear. 261
Each metal expression produced more soluble protein than with no added metal. 262 This was confirmed by SDS-PAGE analysis of the protein extracts of the cells that 263 showed much more soluble protein in the extract expressed in the presence of added 264 metal. The protein from all the separate expressions was then purified on MonoQ as 265 described above for subsequent characterization. No cell growth was observed in the 266 Co(II) expression. 267
Cloning, Overexpression, and Purification of MJ1541 Mutant Gene Products 268 in E. coli. The MJ1541 mutants were highly expressed in E. coli as measured from the 269 SDS-PAGE analysis of the total proteins in the E. coli cells after expression. After 270 sonication of the cells suspended in extraction buffer and centrifugation (14,000 × g) 5 271 min, SDS-PAGE analysis of the pellet and soluble portion of the extract showed that 272 most of the Y136R, E150R, and Y136R/E150R expressed proteins were in the soluble 273 extract. Heating portions of the resulting crude soluble extracts for 10 min at different 274 temperatures followed by removal of the insoluble proteins by centrifugation, indicated 275 that a small band in the soluble protein fraction, that could be the DadD mutants, 276 precipitated at temperatures above 70 °C. Thus, the first step in the purification was 277 heating of the sonicated cell extract to 70 °C for 20 min prior to separation of the extract 278
on July 29, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

14
on MonoQ. This separation produced one peak of DadD activity eluting at 0.44 M NaCl 279 for the Y136R mutant, 0.46 M NaCl for the E150R mutant, and 0.46 M NaCl for the 280 Y136R/E150R mutant. 281
Y136 and E150 were selected for site directed mutagenesis because of the X-ray 282 structure of the T. maritima enzyme (1) identified two arginines in the position of the 283 Y136 and E150 of the M. jannaschii sequence. These residues were identified in the X-284 ray structure to be involved in binding the substrate, particularly in the stabilization of the 285 carboxyl moiety of SAH. We wanted to use site directed mutagenesis to determine if by 286 changing the Tyr and Glu to Arg we could improve DadD’s ability to deaminate SAH by 287 increasing DadD’s ability to hydrolyze this substrate. 288 Measurement of Native Molecular Weight of DadD. The molecular mass of 289 DadD was measured to be ~230 kDa using size exclusion chromatography. This is 290 consistent with the enzyme being a tetramer with a monomeric subunit mass of 57.5 kDa 291 (data shown in Supplemental Figure 1). 292 Temperature Stability of Wild-type DadD and Mutants. The temperature 293 stability of the DadD was found to be different for the purified wild-type and the DadD 294 mutants. The wild-type and the C294S mutant were found to be stable to heating for 10 295 min at 60 oC. The Y136R and E150R DadD mutants were found to be stable for only 10 296 min at 50 oC, showing a significant loss of activity at 60 oC, Table 1. The Y136R/E150R 297 DadD double mutant showed the most significant change in temperature stability, only 298 being stable at 25 oC. The wild-type DadD, metal expressed, and C127S mutant were 299 assayed at 60 oC, the Y126R and E150R mutants were assayed at 50 oC, and the double 300 mutant was assayed at 25 oC for the standard assay. 301
on July 29, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

15
pH optimum of wild-type DadD. The pH optimum of the DadD was found to be 302 pH 9.0 in the BIS TRIS propane buffer (Supplemental Figure 2). Above pH 11.0, the 303 enzyme no longer showed measurable activity due to denaturation of the enzyme. 304 Denaturation of the enzyme was established by changing the pH of the MonoQ purified 305 fraction from 7.5 to 11 by the addition of 2.2 µL 50 mM sodium phosphate pH 12.5 to 13 306 μL of a solution of the purified DadD (0.6 μg). An assay done on the resulting sample at 307 pH 9 showed no enzymatic activity, which lead us to conclude that after exposure to pH 308 11.0 DadD denatures. 309
Dependence DadD Activity on Added Metals. The activity of DadD was tested 310 with several different divalent cations. The addition of metal to the assay had no effect 311 on DadD activity, indicating that the catalytic metal(s) is already coordinated in the active 312 site. 313 Effect of Chelating Agents on DadD Activity. The addition of EDTA to the 314 standard assay stimulated the activity of DadD by 20%. This result is a similar to that 315 seen when urease was incubated with EDTA (13) and indicates that the catalytic metal is 316 not removed by EDTA. The stimulation could be a result of the removal of bound metal 317 that is present on the enzyme that was causing inhibition (14). Dialysis of the wild-type 318 DadD for 3 days against a dipicolinic acid containing buffer also failed to show any 319 lowering enzyme activity, again indicating that chelation is not able to remove the bound 320 metal in the active site. 321 DadD Activity with Alternate Substrates. DadD is annotated as a MTA/SAH 322 deaminase so we tested its activity to deaminate 5’-dA, MTA, SAH, and Ad as substrates 323 using the purified enzyme and each mutant. The results of the kinetic analysis for each of 324
on July 29, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

16
these are summarized in Table 2. The results clearly showed that the preferred substrate 325 is 5’-dA, where the catalytic efficiency of the enzyme for 5’-dA is 103 fold higher than 326 for MTA, and 104 fold higher than with SAH or Ad (Table 2). 327 Effect of DTT on the DadD Activity. The effect of DTT on the activity of 328 DadD was tested by the addition of 0.02 M DTT in the standard assay. This experiment 329 was performed as a result of there being four conserved cysteines in the amino acid 330 sequences of most of the enzymes from the methanogens and it was possible that their 331 oxidation to disulfides during the isolation of the enzyme could have altered the activity. 332 The presence of 0.02 M DTT had no effect on the enzymatic activity. 333 Phylogenetic Analysis of DadD and its homologues. Phylogenetic analysis of 334 the DadD homologues showed they are a widely distributed among archaea and bacteria 335 (Figure 2) and are all annotated as MtaD enzymes. Figure 2 shows that there are three 336 major branches. The M. jannaschii DadD is located in the lower branch (Figure 2, 337 bottom bracket) of the tree along with the Thermotoga maritima homolog, with a 338 sequence identity of 38% to M. jannaschii. Only two other methanogens, M. infernus 339 and M. fervens belong in the same branch. In the top branch (Figure 2, top bracket) 340 contains the P. aeruginosa MTAD enzyme that was found to be specific for MTA (15) 341 and not SAH or Ad, the middle branch (Figure 2, middle bracket) contains the S. griseus 342 SAHD which was found to be specific for SAH and not MTA or Ad (2). This could 343 represent three subfamilies that have varying degrees of substrate specificity, but it is 344 likely that all will catalyze the deamination of 5’-dA, MTA, SAH, and Ad to some degree 345 of efficiency. The specificity of the enzymes may have evolved from the availability of 346 5’-dA, MTA, or SAH that is present in each subfamily of MTA/SAH deaminases. All of 347
on July 29, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

17
the homologs in the phylogenetic tree contain a sequence identity to M. jannaschii DadD 348 ranging from 20 to 92%. 349 DISCUSSION 350
5’-Deoxyadenosine deaminase (DadD) was originally annotated as a 5’-351 methylthioadenosine/S-adenylylhomocysteine deaminase (MtaD, E.C. 3.5.4.31/ 3.5.4.28) 352 (16). At the present time only the enzymes from Thermotoga maritima (17), 353 Pseudomonas aeroginosa (3), and Streptomyces flocculus (18) have been studied. None 354 of these enzymes were ever tested with 5’-deoxyadenosine (5’-dA) as the substrate. The 355 enzyme from T. maritima has had its X-ray crystal structure determined and was shown 356 to have a nickel ion bound to the enzyme at the catalytic site (PDB 1p1m), however, 357 subsequent work done with excess ZnCl2 present during the crystallization showed the 358 protein to have Zn(II) in the active site during analysis of its structure and activity (17). 359 In most cases the presence of 5’-methylthioadenosine/S-adenosylhomocysteine 360 nucleosidase (MtaN) is responsible for maintaining low levels of MTA so that the radical 361 SAM enzymes can remain active (19). MtaN is an enzyme that is known to be involved 362 in the recycling of MTA produced from SAM during the biosynthesis of the polyamines, 363 spermidine, and spermine (20). MtaN works by catalyzing the first step of the 364 methionine salvage pathway (21), where MTA is converted to 5’-methylthioribose-1-P. It 365 could also just as well function where the MTA is deaminated into 5’-methylthioinosine 366 before it undergoes phosphorolysis to 5’-methylthioribose-1-P. Here we propose that the 367 same series of reactions are occurring but the product is 5’-deoxyribose-1-P, from 5’-dI, 368 which is converted into a precursor for aromatic amino acid biosynthesis instead of 369 methionine. 370
on July 29, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

18
Determination of DadD substrate specificity and amino acid residues 371 involved in substrate binding. Substrate specificity was determined by comparing the 372 catalytic efficiency of the wild-type enzyme with each of the four substrates indicated in 373 Table 2. The substrate binding was determined by incubating the wild-type and mutant 374 DadD with varying concentrations of the different substrates and measuring the catalytic 375 efficiency of DadD for each substrate. Our results showed that 5’-dA was the preferred 376 substrate for the enzyme, where the catalytic efficiency (kcat /KM) of the enzyme for 5’-dA 377 was 103 fold higher than for MTA, and 104 fold higher than with SAH or Ad (Table 2). 378 These data indicate that the in vivo substrate is likely 5’-dA, the major product of many 379 radical SAM enzymes (4). 380 The X-ray structure from the T. maritma MtaD gave us insight as to what residues 381 may be important in binding and stabilizing the different substrates in the active site. The 382 X-ray crystal structure showed that two arginines (Figure 3, green) were involved in 383 stabilizing the carboxylate moiety of SAH (1). Sequence alignments of the T. maritima 384 MtaD and other homologous enzymes, including DadD, showed that these two arginines 385 were not highly conserved, in DadD the residues in the position of the two arginines are 386 completely different, this led us to postulate that these arginines maybe what imparts the 387 T. maritima MtaD to have an increased ability to deaminate SAH in comparison with 388 DadD. The KM for SAH from T. maritima MtaD is 210 μM which is lower than that of 389 the M. jannaschii (1100 ± 1300 μM), indicating that the presence of the two arginines in 390 T. maritima may play an important role in binding of SAH (Figure 3). The KM of the 391 wild-type DadD for SAH is much higher than for the other substrates (Table 2) indicating 392 that the loss of the two arginines in M. jannaschii DadD (Figure 3, green) diminished its 393
on July 29, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

19
ability to deaminate SAH, because of DadD’s inability to stabilize the carboxylate moiety 394 of the SAH in the active site. The single mutants in which, Y136 and E150 were replaced 395 by arginine, and the double mutant Y136R/E150R, did not show an increase in the ability 396 to bind or deaminate SAH (Table 2). Interestingly, the kcat/KM values for each of the 397 single mutants of DadD showed that 5’-dA and MTA were the preferred substrates 398 (Table 2). Surprisingly the double mutant Y136R/E150R failed to have any activity 399 towards Ad (Table 2). 400
Taken together the data indicates that the two arginines stabilizing the carboxylate 401 moiety of SAH are not the only residues involved, as previously believed for T. maritima 402 (1). This is consistent with the data reported for the MtaD from P. aeruginosa which 403 showed no activity with SAH as a substrate and it too lacks two arginines in the same 404 position as the arginines from T. maritima (Figure 3) (3). Determining the kinetic 405 parameters of the Y136R, E150R, and Y136R/E150R mutants the data indicates that the 406 stabilization of the substrates, especially SAH, is much more complicated than previously 407 suggested for the T. maritima MtaD based on X-ray analysis of the active site with SAH 408 bound. 409 Characterization of the metal dependence of the M. jannaschii DadD. The 410 amidohydrolase superfamily is known to have a metal bound in the active site that is 411 responsible for generating the nucleophilic water required for the deamination reaction 412 (22). In MtaD a mononuclear metal center is likely coordinated by three histidines and 413 an aspartate (Figure 3, cyan) as is seen in all DadD homologs. The original crystal 414 structure from T. maritima showed that the metal was nickel, however, upon further 415 characterization of the T. maritima enzyme it was co-crystallized with Zn(II) (1). The 416
on July 29, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

20
MTA deaminase (MTADA) from P. aeroginosa was also shown to use zinc for its 417 catalytic metal (3). The catalytic metal in SAH deaminase from S. flocculus was not 418 identified because the authors were unable to remove any metal through the use of 419 several different chelating agents. From this work they concluded that the SAH 420 deaminase did not use a specific divalent cation (2). We were also unable to remove the 421 metal from DadD or establish the nature of the catalytic metal (see Supplemental Data). 422 Proposed route for Re-incorporation of 5’-dA into Aromatic Amino Acids. 423 The conversion of the SAM radical product, 5’-dA, into the deoxyhexose used for 424 aromatic amino acid biosynthesis would first proceed through the deamination of 5’-dA 425 to 5’-deoxyinosine (5’-dI). Next, 5’-dI would undergo phosphorolysis to make 426 hypoxanthine and 5’-deoxyribose-1-phosphate (5-dR-P) (Figure 4). This step has been 427 confirmed to be catalyzed by the MJ0060 gene product (White, R.H. unpublished). The 428 MJ0060 gene product is annotated as a 5’-methylthioadenosine phosphorylase (MTAP) 429 which shares 47% sequence identity to the characterized MTAP from Pyrococcus 430 furiosus. The P. furiosus MTAP was found to catalyze the phosphorolysis of MTA and 431 Ad, but it could also catalyze the phosphorolysis of inosine with a kcat/KM of 9.74 x 103 432 M-1s-1 (23). Recently, a 5’-methylthioinosine phosphorylase (MTIP) from 433 Pseudomonoas aeruginosa has been identified and characterized (15). Interestingly, 434 MTIP was found to be competitively inhibited by 5’-methylthioadensine, the annotated 435 substrate. The kcat /KM was found to be 1.8 ± 0.3 x 106 M-1s-1 for MTI with inosine being 436 the only other substrate, but with 300 fold lower catalytic efficiency (15). However, the 437 sequence identity between MTIP and the MJ0060 gene product is only 39%, lower than 438 that of the MJ0060 gene product with MTAP (47%). We are currently studying the 439
on July 29, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

21
substrate specificity of the MJ0060 gene product. 440 The canonical aromatic amino acid starts with the reaction of PEP with erythrose 441
4-phosphate (E-4-P), a product the pentose phosphate pathway, to produce 3-deoxy-D-442 arabino-heptulosonate -7-phosphate catalyzed by 3-deoxy-D-arabino-heptulosonate 7-443 phosphate synthase (24). Not only are the genes encoding for these enzymes absent in 444 archaeal genomes, so is the canonical pentose phosphate pathway (25). Without the 445 pentose phosphate pathway, E-4-P is unable to be produced necessitating methanogens to 446 have another method for the production of ribose-5-phosphate and/or E-4-P. The 447 archaeal biosynthetic route for the production of 3-dehydroquinate (DHQ), the second 448 intermediate in the shikimate pathway, is also different than the typical biosynthesis of 449 DHQ in bacteria and eukaryotes. The archaeal pathway begins with the condensation of 450 6-deoxy-5-keto-fructose-1-phosphate (DKFP) and L-aspartate semialdehyde (26). A 451 transaldolase reaction transfers the intact C3 hydroxyacetone fragment (carbons 4, 5, and 452 6) to the L-aspartate semialdehyde, forming 2-amino-3,7-dideoxy-D-threo-hept-6-453 ulosonic acid. This amino sugar then undergoes an NAD-dependent oxidative 454 deamination to produce 3,7-dideoxy-D-threo-hept-2,6-diulosonic acid, which cyclizes to 455 DHQ (27). DKFP’s biosynthesis was proposed to arise via transaldolase reaction between 456 methylglyoxal and fructose-1,6-bis-P (28) with the methylglyoxal arising from 457 lactaldehyde (29). We propose that the origin of the methylglyoxal and/or DKFP arises 458 from the 5’-deoxyribose-1-P the product of the MJ0060 gene. 459
The hypoxanthine product could be incorporated into IMP by reaction with 460 phosphoribosyl pyrophosphate catalyzed by a hypoxanthine phosporibosyltransferase. 461 The 5-dRP product could be incorporated into the deoxyhexoses required for aromatic 462
on July 29, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

22
amino acid biosynthesis without coming from the traditional pentose pathway shunt. 463 This could involve an isomerase (MJ0454 gene product) that is annotated to use the 5’-464 methylthio derivative of 5-dRP to produce 5-deoxyribulose-1-phosphate typically 465 involved in the methionine salvage pathway (30, 31). This compound could be further 466 metabolized by several yet to be identified enzymes to produce 6-deoxy-5-ketofructose, 467 the required precursor for aromatic amino acid (Figure 4). The identification of the 468 enzymes and pathway for the biosynthesis of deoxyhexoses is currently being studied. 469 Conclusions. 470
In conclusion we have shown that the annotated 5’-methylthioadenosine/ S-471 adenosyl-L-homocysteine deaminase MJ1541 gene product from M. jannaschii 472 preferentially uses 5’-deoxyadenosine as its substrate compared to 5’-473 methylthioadenosine, S-adenosylhomocysteine, and adenosine. From this analysis we 474 rename this enzyme 5’-deoxyadenosine deaminase (DadD). Here we also suggest that 475 other members of this family may be using 5’-deoxyadenosine as a preferred substrate as 476 well, based on phylogenetic analysis with other homologs. Unfortunately, we were 477 unable to fully identify the in vivo catalytic metal, but based on our data we propose it is 478 either Ni(II) or Zn(II). 479 Acknowledgments 480
This work was supported by the National Science Foundation Grant 481 MCB1120346 supported this work. 482 483
on July 29, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

23
REFERENCES 484 1. Hermann JC, Marti-Arbona R, Fedorov AA, Fedorov E, Almo SC, Shoichet 485 BK, Raushel FM. 2007. Structure-based activity prediction for an enzyme of 486 unknown function. Nature 448:775-779. 487 2. Zulty JJ, Speedie MK. 1989. Purification and characterization of S-488 adenosylhomocystine deaminase from streponigrin-producing Streptomyces 489 flocculus. Journal of Bacteriology 171:6840-6844. 490 3. Guan R, Ho M-C, Frohlich RFG, Tyler PC, Almo SC, Schramm VL. 2012. 491 Methylthioadenosine deaminase in an alternative quorum sensing pathway 492 in Pseudomonas aeurginosa. Biochemistry 51:9094-9103. 493 4. Booker SJ, Grove TL. 2010. Mechanistic and functional versatility of radical 494 SAM enzymes. F1000 Biology Reports 2 (52). 495 5. Graham. D. E. X, H., and White, R. H. . 2002. Identification of coenzyme M 496 biosynthetic phosphosulfolactate synthase: a new family of sulfonate 497 biosynthesizing enzymes. J. Biol. Chem. 277:13421-13429. 498 6. Bradford MM. 1976. A rapid and sensitive method for the quantitation of 499 microgram quantities of protein utilizing the principle of protein dye-500 binding. Anal. Biochem. 72:248-257. 501 7. Miller DV, Xu H, White RH. 2012. A new subfamily of agmatinases in 502 methanogenic Archaea is Fe(II) dependent. Biochemistry 51:3067-3078. 503 8. Volkin E, Cohn WE. 1954. Estimation of nucleic acids. Methods Biochem. 504 Anal. I:287-305. 505
on July 29, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

24
9. Huang DT, Thomas MA, Christopherson RI. 1999. Divalent metal 506 derivatives of the hamster dihydroorotase domain. Biochemistry 38:9964-507 9970. 508 10. Saitou N, Nei M. 1987. The neighbor-joining method: A new method for 509 reconstructing phylogenetic trees. Mol. Biol. Evol. 4:406-425. 510 11. Zuckerkandl E, Pauling L. 1965. Evolutionary divergence and convergence 511 in proteins. Evolving Genes and Proteins 97:97-166. 512 12. Tamura K, Peterson D, Peterson N, Stecher G, Nei M, Kumar S. 2011. 513 MEGA5: Molecular Evolutionary Genetics Analysis using Maximum 514 Likelihood, Evolutionary Distance, and Masximum Parsimony Methods. Mol. 515 Biol. Evol. 28:2731-2739. 516 13. Park IS, Hausinger RP. 1996. Metal ion interaction with urease and UreD-517 urease apoproteins. Biochemistry 35:5345-5352. 518 14. Mahadevan S, Sauer FD, Erfle JD. 1977. Purification and Properties of 519 Urease from Bovine Rumen. Biochemical Journal 63:495-501. 520 15. Guan R, Ho M-C, Almo SC, Schramm VL. 2011. Methylthioinosine 521 Phosphorylase from Pseudomonas aeroginosa. Structure and Annotaion of a 522 Novel Enzyme in Quorum Sensing. Biochemistry 50:1247-1254. 523 16. Bult CJ, White O, Olsen GJ, Zhou L, Fleischmann RD, Sutton GG, Blake JA, 524 FitzGerald LM, Clayton RA, Gocayne JD, Kerlavage AR, Dougherty BA, 525 Tomb J-F, Adams MD, Reich CI, Overbeek R, Kirkness EF, Weinstock KG, 526 Merrick JM, Glodek A, Scott JL, Geoghagen NSM, Weidman JF, Fuhrmann 527 JL, Nguyen D, Utterback TR, Kelley JM, Peterson JD, Sadow PW, Hanna 528
on July 29, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

25
MC, Cotton MD, Roberts KM, Hurst MA, Kaine BP, Borodovsky M, Klenk 529 H-P, Fraser CM, Smith HO, Woese CR, Venter JC. 1996. Complete Genome 530 Sequence of the Methanogenic Archaeon, Methanococcus jannaschii. Science 531 273:1058-1073. 532 17. Herman JC, Marti-Arbona, R., Fedorov, A. A., Fedorov, E., Almo, S. C., 533 Shoichet, B. K., and Raushel, F. M. 2007. Structure- based activity 534 prediction for an enzyme of unknown function. Nature 448:775-779. 535 18. Speedie MK, Zulty, J.J., and Brothers, P. 1988. S-Adenosylhomocysteine 536 Metabolism in Streptomyces flocculus. J. Bacteriol. 170:4376-4378. 537 19. Challand MR, Ziegert T, Douglas P, Douglas P, Wood RJ, Kriek M, Shaw 538 NM, Roach PL. 2009. Product inhibition in the radical S-adenosylmethionine 539 family. FEBS Lett. 583:1358-1362. 540 20. Parveen N, Cornell KA. 2011. Methylthioadenosine/S-541 adenosylhomocysteine nucleosidase, a critical enzyme for bacterial 542 metabolism. Molecular Microbiology 79:7-20. 543 21. Pirkov I, Norbeck J, Gustafsson L, Albers E. 2008. A complete inventory of 544 all enzymes in the eukaryotic methionine salvage pathway. FEBS J. 545 275:4111-4120. 546 22. Seibert CM, Raushel FM. 2005. Structural and catalytic diversity within the 547 amidohydrolase superfamily. Biochemistry 44:6383-6391. 548 23. Cacciapuoti G, Bertoldo C, Brio A, Zappia V, Porcelli M. 2003. Purificatin 549 and characterization of 5'-methylthioadenosine phosphorylase from the 550 hyperthermophilic archaeon Pyrococcus furiosus. Extremphiles 7:159-168. 551
on July 29, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

26
24. Schoner R, Herrmann KM. 1976. 3-Deoxy-D-arabino-heptulosonate 7-552 phosphate sunthase. J Biol Chem 251:5440-5447. 553 25. Grochowski L, Xu H, White RH. 2005. Ribose-5-phosphate biosynthesis in 554 Methanocaldococcus jannaschii occurs in the absence of a pentose phosphate 555 pathway. Journal of Bacteriology 187:7382-7389. 556 26. White RH. 2004. L-Aspartate semialdehyde and a 6-deoxy-5-ketohexose 1-557 phosphate are the precursors to the aromatic amino acids in 558 Methanocaldococcus jannaschii. Biochemistry 43:7618-7627. 559 27. White RH. 2004. L-Aspartate Semialdehyde and 6-Deoxy-5-ketofructose 1-560 Phosphate Are the Precursors to the Aromatic Amino Acids in 561 Methanocaldococcus jannaschii. Biochemistry 43:7618-7627. 562 28. White RH, Xu H. 2006. Methylglyoxal is an intermediate in the biosynthesis 563 of 6-deoxy-5-ketofructose-1-phosphate: a precursor for aromatic amino acid 564 biosynthesis in Methanocaldococcus jannaschii. Biochemistry 45:12366-565 12379. 566 29. White RH. 2008. Biochemical origins of lactaldehyde and hydroxyacetone in 567 Methanocaldococcus jannaschii. Biochemistry 47:5037-5046. 568 30. Saito Y, Ashida H, Kojima C, Tamura H, Matsumura H, Kai Y, Yokota A. 569 2007. Enzymatic Characterization of 5-Methylthioribose 1-Phosphate 570 Isomerase from Bacillus subtilis. Biosc. Biotechnol. Biochem. 71:2021-2028. 571 31. Tamura H, Saito y, Ashida H, Inoue T, Kai Y, Yokota A, Matsumura H. 572 2008. Cyrstal structure of 5-methylthioriobse 1-phosphate isomerase 573
on July 29, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

27
product complex from Bacillus subtilis: Implications for catalytic mechanism. 574 Protein Science 17:126-135. 575 576 577
on July 29, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

28
Table 1. Temperature stability of purified wild-type and DadD mutants 578 DadD Temperature
Percent of Retained
Activity*
WT 60 90%
70 34%
Y136R 50 80%
60 40%
E150R 50 96%
60 25%
Y136R&E150R double
mutant
25 100%
50 35%
*after heating for 10 minutes at the indicated temperature 579 580 581 582 583 584 585 586 587 588 589 590
on July 29, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

29
591 Table 2. Kinetic parmeters of DadD and mutants with 5’-dA, MTA, SAH, and Ad 592
DadD Substrate KM (mM) kcat (sec-1) kcat /KM
(M-1sec-1)
WT 5’-dA 0.014 ± 0.0012 1.2 x 105 9.1 x 109
MTA 0.11 ± 0.028 1.6 x 102 1.1 x 106
SAH 1.1 ± 1.3 1.3 x 103 4.4 x 106
Ad 0.15 ± 0.0047 1.1 x 102 7.5 x 105
Y136R 5’-dA 0.032 ± 0.033 3.3 x 102 1.7 x 107
MTA 0.022 ± 0.0019 2.2 x 102 1.1 x 107
SAH 0.16 ± 0.13 24.7 2.3 x 105
Ad 0.072 ± 0.0018 1.4 x 102 2.9 x 106
E150R 5’-dA 0.021 ± 0.0061 2.1 x 102 9.8 x 107
MTA 0.048 ± 0.0029 2.0 x 102 4.2 x 106
SAH 3.9 ± 5.2 58.2 2.1 x 104
Ad 0.15 ± 0.047 1.1 x 102 7.5 x 105
Y136R/E150R 5’-dA 0.05 ± 0.046 1.12 3.1 x 104
MTA 0.073 ± 0.0054 1.39 1.9 x 104
SAH 0.4 ± 0.1 2.41 5.3 x 103
Ad Nm* Nm* Nm*
*Nm: Not Measurable 593 594 595
on July 29, 2018 by guesthttp://jb.asm
.org/D
ownloaded from
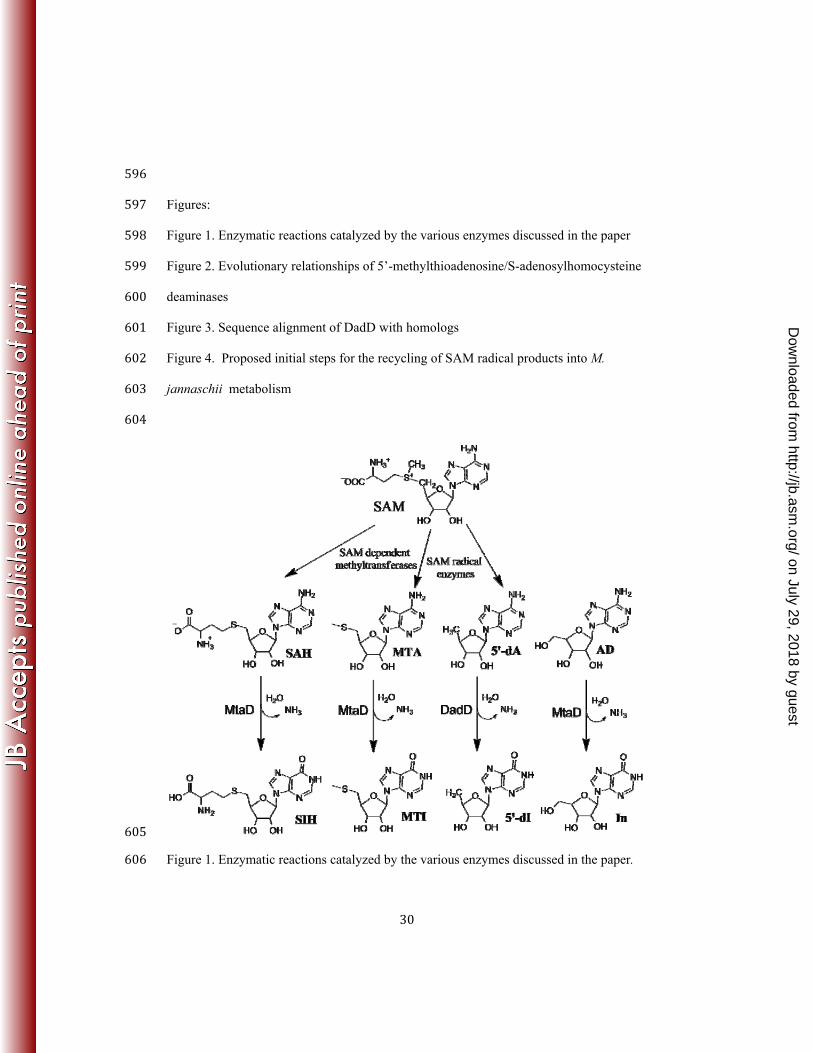
30
596 Figures: 597 Figure 1. Enzymatic reactions catalyzed by the various enzymes discussed in the paper 598 Figure 2. Evolutionary relationships of 5’-methylthioadenosine/S-adenosylhomocysteine 599 deaminases 600 Figure 3. Sequence alignment of DadD with homologs 601 Figure 4. Proposed initial steps for the recycling of SAM radical products into M. 602 jannaschii metabolism 603 604
605 Figure 1. Enzymatic reactions catalyzed by the various enzymes discussed in the paper. 606
on July 29, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

31
607 608 Figure 2. Evolutionary relationships of 5’-methylthioadenosine/S-adenosylhomocysteine 609 deaminases to M. jannaschii DadD. MtaD represents the homologs that do not have 610 confirmed gene annotation, those confirmed are MTAD, SAHD, MtaD, and DadD. The 611 organisms in the top bracket represent those that have the closest homology to MTAD 612 and likely use MTA as their preferred substrate. The organisms in the middle bracket 613 share homology with SAHD and likely use SAH as their preferred substrate. Those in 614 the bottom bracket share homology with DadD of M. jannaschii (highlighted in the box) 615 and we propose use 5’-dA as their preferred substrate. 616
Thermococcus onnurineus mtaD
Pyrococcus kodakaraensis mtaD
Aquifex aeolicus mtaD
Methanococcus aeolicus mtaD
Pelosinus fermentans mtaD
Methanothermobacter thermautotrophicus mtaD
Pseudomonas fluorescens mtaD
Pseudomonas aeruginosa MTAD
Streptomyces griseus SAHD
Clostridium ultunense mtaD
Bacillus halodurans mtaD
Methanobrevibacter ruminantium mtaD
Methanopyrus kandleri mtaD
Moorella thermoacetica mtaD
Clostridium botulinum mtaD
Methanococcus maripaludis mtaD
Archaeoglobus veneficus mtaD
Pyrococcus furiosus mtaD
Methanocaldococcus infernus mtaD
Thermotoga maritima MtaD
Methanocaldococcus jannaschii DadD
Methanocaldococcus fervens mtaD
0.6601
0.6876
0.0759
1.3546
0.5024
0.6962
0.0286
0.0889
1.0095
0.9867
0.9387
0.9243
0.7301
0.0505
1.0121
1.2318
0.7164
1.2397
1.2303
0.1424
1.3554
0.10061.2036
0.8062
0.4293
0.2166
0.3111
0.4597
0.2116
0.2339
0.2946
0.2834
0.1755
0.1462
0.0988
0.1029
0.0995
0.0679
0.0419
0.0293
0.0266
0.0036
on July 29, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

32
The accession number for the proteins used in the construction of the tree in order 617 from the top to the bottom are as follows: B6YUF8, Q5JER0, O66851, A6UUG9, 618 I9MDY4, O27549, Q4K8M5, A6V2Q5, G0PU20,M1Z1T6, Q9KC82, D3DYL9, 619 Q8TYD4, Q2RJW1, B1BCH5, A4FW32, F2KTB3, Q8U0P7, D5VT65, Q9X034, 620 Q58936, C7P5A2. 621 622
on July 29, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

33
623 Figure 3. Sequence alignment of DadD from M. jannaschii with homologs from other 624 methanogens as well as with previously characterized enzymes. The residues are 625 highlighted in the following way: cyan background with black lettering are those residues 626 coordinating the metal, a blue background with white letters are residues interacting with 627 the ribose moiety of the substrate, the pink background with black letters are those 628 interacting with the purine base, the purple background with white letters are those 629 involved in forming a hydrophobic pocket to bind the 5’-methylthio moiety of MTA, the 630
30 78 M. jannaschii DadD KEEIEN--AEIIDGKNKIAIPGLINTHTHIPMTLFRGVADDLPLMEWLNN M. maripaludis ISEVSKDETEIIDGKNCVLIPGLVNTHTHVPMSLFRGVADDIPLMEWLSG M. thermautotrophicus NTLSPGDADTVIDGTGKLLIPGLVNTHTHLSMTLFRGIADDLPLDRWLND T. maritima mtaD R-VLQGEVKVDLDLSGKLVMPALFNTHTHAPMTLLRGVAEDLSFEEWLFS P. aeruginosa MTADA EQAMRHGATEIRELPGMLLAPGLVNAHGHSAMSLFRGLADDLPLMTWLQD S. griseus SAHD ELTRTHPGLHAVRLPGHLLMPGFVDAHHHLTQSFGSALAFGEPSEIFRR- 79 123 M. jannaschii DadD YIWPMEAKLN-EEIVYWGTLLGCIEMIRSGTTTFNDM----YFFLEGIAK M. maripaludis HIWPMESKLN-EKIVYAGTLLGTVEMIKSGTTAFNDM----YFFLDSIIK M. thermautotrophicus HIWPAEARLN-GDYCYAGALLGCIEMIRSGTTSFNDM----YFYMDHVAR T. maritima mtaD KVLPIEDRLT-EKMAYYGTILAQMEMARHGIAGFVDM----YFHEEWIAK P. aeruginosa MTADA HIWPAEGQWVSEDFIRDGTELAIAEQVKGGITCFSDM----YFYPQAICG S. griseus SAHD VWVPLEGALD-EEAAYAAAKLAALEALRGGFTTVTDAGTRADVDVEVVAS 124 136 150 157 M. jannaschii DadD AVDESGMRAVLAYGMIDLFDEERRERELKNA----------------EKY M. maripaludis AVDETGIRSTIAYGMIDLFDEEKREKELKTA----------------RKS M. thermautotrophicus AVEEAGLRCVISHGMIDLGDTEKMTAELRES----------------RRI T. maritima mtaD AVRDFGMRALLTRGLVDSNGDDG--GRLEEN----------------LKL P. aeruginosa MTADA VVHDSGVRAQVAIPVLDFPIPGARDSAEAIR----------------QGM S. griseus SAHD AARDAGIRCVLGLVCNDVTGDPASTAGPSTAGDSTAGESASTAAVLERAE 155 207 M. jannaschii DadD INYINSLNNSRIMPALGPHAPYTCSKELLMEVNNLAKKYNVPIHIHLNET M. maripaludis LETIKNLNNSRITGALGPHAPYTCSKELLESTNTLAREYNVPIHIHMNET M. thermautotrophicus IKECHGMADDRIRVALGPHSPYTCSEELLKETAALADKNDLMIHIHVSET T. maritima mtaD YNEWNGFEG-RIFVGFGPHSPYLCSEEYLKRVFDTAKSLNAPVTIHLYET P. aeruginosa MTADA ALFDDLKHHPRIRIAFGPHAPYTVSDDKLEQILVLTEELDASIQMHVHET S. griseus SAHD KHLARYAGDPLIHASLAVSIPEAATARTLHRTARLAAEAGAIVQIHVNEH 258 307 M. jannaschii DadD INVSHNPISNLKLASGVAPIPKLLAEGINVTLGTDGCGSNNNLNLFEEIK M. maripaludis IFAAHNPVSNLKLASGVSPVLKLLENNVPVTLGTDGCGSNNNMNLFEEIK M. thermautotrophicus VKVSHNPSSNMKLASGVSPVARLLQRGVNVSLGTDGAASNNNLDMFQEMK T. maritima mtaD FFVSHNPASNLKLGNGIAPVQRMIEHGMKVTLGTDGAASNNSLNLFFEMR P. aeruginosa MTADA SSVIHCPESNLKLASGFCPVEKLWQAGVNVAIGTDGAASNNDLDLLGETR S. griseus SAHD TAVSYNPVASAWKGNAVAPATALAERGIRFGLGTDGTRGDG-FRLTDAAE
on July 29, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

34
grey background with black letters are interacting with the 5’-methylthio moiety of MTA, 631 and the arginines residues interacting with the SAH carboxylate are highlighted in a 632 green background with black letters and are underlined. M. jannaschii numbering is 633 used. 634
635 Figure 4. Proposed route for the incorporation 5’-dA into deoxyhexoses for the 636 biosynthesis of aromatic amino acids in Methanocaldococcus jannaschii 637 638
on July 29, 2018 by guesthttp://jb.asm
.org/D
ownloaded from



![Cinetica-Biochimica2012 Branchini [modalit compatibilit ]m.docente.unife.it/francesco.bernardi/materiale-didattico... · Costante catalitica (kcat) o Numero di turnover Per il modello](https://img.dokumen.tips/doc/110x75/5c65c04e09d3f29b6e8d3435/cinetica-biochimica2012-branchini-modalit-compatibilit-m-costante-catalitica.jpg)











![Interacțiunea enzimă-substratCinetica reactiilor enzimatice - teoria starii stationare k2 = k cat vmax = [E] 0 kcat v = vmax [S] Km + [S] Ecuatia Michaelis-Menten. Introduction to](https://img.dokumen.tips/doc/110x75/6105ebdb36b89e4800454342/interaciunea-enzimf-substrat-cinetica-reactiilor-enzimatice-teoria-starii-stationare.jpg)





![Enzymes II - Oregon State Universityoregonstate.edu/instruct/bb450/450material/Keynotes/10Enzymes.pdfMichaelis)Menten,Kinetics • Vmax and Kcat Vmax [Enzyme Used] = [Product] [Enzyme](https://img.dokumen.tips/doc/110x75/5e75841310de0e0b173bc785/enzymes-ii-oregon-state-u-michaelismentenkinetics-a-vmax-and-kcat-vmax-enzyme.jpg)







