Embed Size (px)
Citation preview

CLINICAL AND VACCINE IMMUNOLOGY, Mar. 2008, p. 402–411 Vol. 15, No. 31556-6811/08/$08.00�0 doi:10.1128/CVI.00366-07Copyright © 2008, American Society for Microbiology. All Rights Reserved.
Identification of 19 Polymorphic Major Outer Membrane ProteinGenes and Their Immunogenic Peptides in Ehrlichia ewingii
for Use in a Serodiagnostic Assay�†Chunbin Zhang, Qingming Xiong, Takane Kikuchi, and Yasuko Rikihisa*
Department of Veterinary Biosciences, College of Veterinary Medicine, The Ohio State University,1925 Coffey Rd., Columbus, Ohio 43210-1093
Received 3 September 2007/Returned for modification 30 October 2007/Accepted 5 December 2007
Ehrlichia ewingii, a tick-transmitted rickettsia previously known only as a canine pathogen, was recentlyrecognized as a human pathogen. E. ewingii has yet to be cultivated, and there is no serologic test available todiagnose E. ewingii infection. Previously, a fragment (505 bp) of a single E. ewingii gene homologous to 1 of 22genes encoding Ehrlichia chaffeensis immunodominant major outer membrane proteins 1 (OMP-1s)/P28s wasidentified. The purposes of the present study were to (i) determine the E. ewingii omp-1 gene family, (ii)determine each OMP-1-specific peptide, and (iii) analyze all OMP-1 synthesized peptides for antigenicity.Using nested touchdown PCR and a primer walking strategy, we found 19 omp-1 paralogs in E. ewingii. Thesegenes are arranged in tandem downstream of tr1 and upstream of secA in a 24-kb genomic region. Predictedmolecular masses of the 19 mature E. ewingii OMP-1s range from 25.1 to 31.3 kDa, with isoelectric points of5.03 to 9.80. Based on comparative sequence analyses among OMP-1s from E. ewingii and three other Ehrlichiaspp., each E. ewingii OMP-1 oligopeptide that was predicted to be antigenic, bacterial surface exposed, uniquein comparison to the other E. ewingii OMP-1s, and distinct from those of other Ehrlichia spp. was synthesizedfor use in an enzyme-linked immunosorbent assay. Plasmas from experimentally E. ewingii-infected dogsreacted significantly with most of the OMP-1-specific peptides, indicating that multiple OMP-1s were ex-pressed and immunogenic in infected dogs. The results support the utility of the tailored OMP-1 peptides asE. ewingii serologic test antigens.
Ehrlichia ewingii, a granulocytotropic Ehrlichia species (1),was recognized in 1999 as a human pathogen belonging to thefamily Anaplasmataceae in the United States, following Ehrli-chia chaffeensis and Anaplasma phagocytophilum (6). Between1996 and 2001, approximately 10 confirmed cases of humangranulocytic ehrlichiosis caused by E. ewingii infection wereidentified in Missouri and Oklahoma (17). The numbers of E.ewingii cases being significantly lower than the numbers of A.phagocytophilum and E. chaffeensis cases (9) may be due par-tially to difficulty in diagnosing E. ewingii infection. Clinicalsigns of patients infected with E. ewingii, such as fever, head-ache, myalgia, leukopenia, and thrombocytopenia, are similarto those for human monocytic ehrlichiosis caused by E.chaffeensis and human granulocytic anaplasmosis caused by A.phagocytophilum (4, 6, 31). Hence, clinical features alone can-not distinguish these causative agents.
A further complication in the diagnosis of ehrlichiosis infec-tions is that E. ewingii and E. chaffeensis also share the samevector tick species and animal reservoirs. Experimentally, theLone Star tick (Amblyomma americanum) is a competent vec-tor for E. ewingii and E. chaffeensis infection (2, 10). Thewhite-tailed deer (Odocoileus virginianus) is suspected to beand considered an important reservoir for E. ewingii and E.
chaffeensis, respectively, and the dog is another possible reser-voir of both Ehrlichia spp. (8, 10, 21, 48). Consequently, E.ewingii and E. chaffeensis have similar seasonal and geographicdistributions. In contrast, A. phagocytophilum uses Ixodes spe-cies of ticks as vectors and wild rodents as primary reservoirs,thus having a different seasonal and geographic distributionfrom E. ewingii. However, in addition to human travel, there isenough crossover in both vector and reservoir distributions tocause confusion in attempting to properly distinguish E. ewingiifrom the other agents of ehrlichioses, warranting developmentof better laboratory testing.
To distinguish infections, direct tests such as culture isola-tion, PCR, and microscopic observation of morulae (micro-colonies of ehrlichiae) are useful if blood specimens are avail-able at acute stages of infection. However, unlike E. chaffeensisand A. phagocytophilum, which can be culture isolated in my-elocytic leukemia and tick cell lines, E. ewingii is currently notcultivable. PCR tests based on the E. ewingii-specific partialsequence of a 16S rRNA gene and a partial p28-19 sequencehave been reported (8, 16, 17, 21, 26, 48), yet the sensitivitiesand specificities of E. ewingii PCR tests of clinical specimensare unknown, as there are no other definitive tests with whichto compare. The microscopic observation of morulae inRomanovsky dye-stained peripheral blood granulocytes pro-vides definitive proof of ehrlichial infection. Unfortunately,this test cannot be used as a single diagnostic test for E. ewingiiinfection because it cannot distinguish E. ewingii morulae fromother granulocytic agents, such as A. phagocytophilum. Fur-thermore, negative results from Romanovsky dye staining can-not rule out E. ewingii infection, owing to high false-negative
* Corresponding author. Mailing address: Department of Veteri-nary Biosciences, College of Veterinary Medicine, The Ohio StateUniversity, 1925 Coffey Rd., Columbus, OH 43210-1093. Phone: (614)292-5661. Fax: (614) 292-6473. E-mail: [email protected].
† Supplemental material for this article may be found at http://cvi.asm.org/.
� Published ahead of print on 19 December 2007.
402
on October 14, 2018 by guest
http://cvi.asm.org/
Dow
nloaded from

rates caused by sample conditions and the low sensitivity of theassay. These setbacks in prior diagnostic testing necessitate anadditional test to properly identify E. ewingii infection.
Given the shortcomings in direct testing, indirect testing maybe the answer for identification of ehrlichioses. Since ehrlichialinfections induce significant antibody titers in nonimmuno-compromised patients and since nonexposed people seldomhave antibodies reactive to Ehrlichia spp., serologic tests areconsidered the most reliable tests for confirmation of ehrli-chioses, especially when ruling out the possibility of infection.Although an indirect fluorescent-antibody (IFA) test using cul-tivated bacteria is widely used for E. chaffeensis and A. phago-cytophilum serodiagnosis (9), this assay is not applicable for thediagnosis of E. ewingii infection due to the lack of culturedbacteria. As an alternative to whole bacteria, several immuno-dominant proteins, including major outer membrane proteinsof Ehrlichia and Anaplasma spp., have been cloned and ex-pressed as immunoreactive fusion proteins (5, 18, 22–24, 29,30, 46, 50, 53). We previously reported that dot immunoblotsor enzyme-linked immunosorbent assays (ELISAs) of dog andhuman sera with the recombinant 30-kDa major outer mem-brane protein (rP30) of Ehrlichia canis and the recombinant44-kDa major outer membrane protein (rP44) of A. phagocy-tophilum, respectively, provide diagnostic sensitivities andspecificities comparable to those of their whole bacteria IFAtest counterparts (29, 40, 53). In the present study, we there-fore proposed to create a similar test for E. ewingii.
In order to develop a serologic test using major antigens ofE. ewingii, genes encoding these proteins must first be identi-fied. Several members of the genus Ehrlichia have immuno-dominant surface proteins, e.g., outer membrane protein 1(OMP-1)/P28s in E. chaffeensis, P30s in E. canis, and Map 1 inEhrlichia ruminantium, which have been used as serodiagnosticantigens (12, 29, 32, 41, 43, 51). These surface proteins areencoded by a polymorphic multigene family and are immuno-logically highly cross-reactive to each other (25, 28–30, 33, 45,49). A partial sequence (505 bp) for a single gene of the majorOMP-1 gene family called p28-19 (ortholog of E. chaffeensisp28 [30]) is known for E. ewingii (17). In the present study, wesystematically identified the entire E. ewingii OMP-1 genomiclocus, which contains multiple omp-1 paralogs.
While there is no small laboratory animal model for E.ewingii infection, dogs can be used as an infection model. Infact, E. ewingii was originally discovered as a granulocytic vari-ant of Ehrlichia canis that typically infects canine monocytes(11). Canine infection with E. ewingii has been detected morefrequently and in broader geographic regions than human in-fection (8, 16, 21, 26, 27). In the present study, therefore,plasmas from experimentally E. ewingii-, E. chaffeensis-, and E.canis-infected dogs were used to test the potential utility of theE. ewingii OMP-1 peptides as serodiagnostic antigens byELISA.
MATERIALS AND METHODS
E. ewingii omp-1 cluster amplification, sequencing, and assembly. An EDTA-treated whole-blood specimen (�200 �l) collected in April 2005 from an 8-week-old male German Shepherd mixed-breed dog in Ohio was used for DNA extrac-tion. DNA was extracted using a QIAamp blood kit (Qiagen, Valencia, CA) andused as the template for the entire amplification and sequencing process. E.ewingii infection of the dog was confirmed by PCR and sequencing of the 16S
rRNA of E. ewingii as well as by observation of bacterial inclusions (morulae) ingranulocytes in the blood and joint fluid smear. PCR analysis showed that thedog was negative for infection by A. phagocytophilum, E. chaffeensis, and E. canis(Q. Xiong, W. Bao, and Y. Rikihisa, unpublished data).
The omp-1 fragments were amplified first by touchdown PCR (37) with theprimer pairs F1 and R7, F8 and R14, and F15 and R21 (see Table S1 in thesupplemental material). The PCR mix (50 �l) included 0.5 �l template DNAcorresponding to 4 �l of the original blood sample, 10 pmol of each primer, 0.2mM deoxynucleoside triphosphate mixture, 2.5 U high-fidelity Taq polymerase(Invitrogen, Carlsbad, CA), and 1.5 mM MgCl2. Amplification was performedwith the following program: 94°C for 3 min; a gradient over 10 cycles of 94°C for0.5 min, 64°C for 0.5 min, and 72°C for 2 min, with the annealing temperaturedecreased by 1°C/cycle; 35 cycles of 94°C for 0.5 min, 55°C for 0.5 min, and 68°Cfor 9 min; and, finally, 68°C for 9 min. Nested PCRs were performed using thefirst PCR products as templates, with 21 pairs of degenerate primers, creatingamplicons of approximately 1,500 bp that each overlapped approximately 200 bpaccording to the E. chaffeensis and E. canis omp-1 clusters (Fig. 1; see Table S1in the supplemental material). The conditions of the nested PCR were similar tothose of the first PCR, except that Taq polymerase was used and the elongationstep was done at 72°C for 2 min. The nested PCR products were run in a 1%agarose gel with TAE buffer (40 mM Tris-acetate, 1 mM EDTA, pH 8.0). Theamplified DNA fragments were recovered from the gel with a QIAEX II gelextraction kit (Qiagen) and directly sequenced with the nested PCR primers (Fig.1). For fragment 3, two touchdown PCRs with high-fidelity Taq polymerase wereperformed, using infected dog blood DNA as the template and one of thefollowing primer pairs: forward primer P28-19F and primer R21 or a forwardprimer designed based on the 3� end of fragment 2 (Specific 4F) and reverseprimer P28-19R (see Table S1 in the supplemental material). Nested amplifica-tion of these two PCR products and direct sequencing were used for subsequentdesign of new specific primers. Direct sequences obtained ranged from 250 to800 bp. The poly(G/C) or poly(A/T) region (Fig. 1) was cloned using a TAcloning kit (Invitrogen), and the plasmid insert was sequenced. All sequencingdata were assembled using the SeqMan program of DNASTAR software(DNASTAR Inc., Madison, WI).
E. ewingii omp-1 cluster analysis. Artemis (38) was used to identify the openreading frames (ORFs) in the newly obtained E. ewingii omp-1 cluster. TheORFs encoding more than 100 amino acids were searched against the NCBIGenBank database to find homologs. The Artemis comparison tool (7) was usedto analyze the synteny of the E. ewingii omp-1 cluster to those of E. chaffeensis,E. canis, and E. ruminantium. To search for repeat regions in the E. ewingii omp-1cluster and between the E. ewingii omp-1 cluster and that of E. chaffeensis, E.canis, or E. ruminantium, dot plot analysis was performed with the Java Dot PlotAlignments program (JDotter; http://athena.bioc.uvic.ca/index.php).
Phylogenetic analysis. The deduced amino acid sequences of E. chaffeensisOMP-1s/P28s, E. canis P30s, E. ruminantium MAP1s, and E. ewingii OMP-1swere aligned using the MegAlign program of DNASTAR software. Phylogeneticanalysis was then performed with PHYLIP software (version 3.66) (13). Thephylogram was constructed using the neighbor-joining method with Kimura’sformula, and 1,000 bootstrap replications were conducted to evaluate the reli-ability of the tree (13).
Peptide synthesis and peptide-pin ELISA analysis. Peptide libraries weresynthesized using noncleavable multipin synthesis technology and 9-fluorenyl-methoxy carbonyl chemistry (Mimotopes Pty. Ltd., Victoria, Australia) (15).After stripping off any previously bound proteins from the peptide pins with 0.1M sodium phosphate buffer containing 1% sodium dodecyl sulfate (pH 7.2) and0.1% �-mercaptoethanol at 60°C for 30 min, followed by two water washes for 10min each, nonspecific binding sites were blocked with 200 �l of 3% skim milk(Becton Dickinson and Co., Sparks, MD) in phosphate-buffered saline (PBS)–Tween 20. Blocking was carried out in 96-well plates for 1 h at room tempera-ture. Sets of peptide-bound pins were washed once with PBS containing 0.1%(vol/vol) Tween 20 for 10 min and then incubated in the blocking solution (1:100dilution) with plasmas from E. ewingii-, E. chaffeensis-, or E. canis-infected dogsor with preinfection dog plasma at 4°C overnight. Samples from dogs 2119, 2185,and 2405 were collected on days 206, 109, and 123, respectively, after infectionwith E. ewingii. Samples from dog CTUALJ (E. chaffeensis IFA titer, 1:2,560),dog 1425 (E. chaffeensis IFA titer, 1:320), and dog 3918815 (E. chaffeensis IFAtiter, 1:2,560) were collected on days 41, 121, and approximately 210, respec-tively, after infection with E. chaffeensis. Samples from dogs 246, 433, and 688were collected 21 days after infection with E. canis, when the IFA titers were�1:2,560. Dog 2405 serum from the indicated time points (0, 21, 28, and 123 daysafter infection with E. ewingii) was selected for time course assays. Preinfectionplasmas from dogs 2119, 2185, 1425, 246, 433, and CTUALJ were used asnegative controls. After being washed four times as described above, the peptide
VOL. 15, 2008 IMMUNOREACTIVE E. EWINGII OMP-1s 403
on October 14, 2018 by guest
http://cvi.asm.org/
Dow
nloaded from
pins were placed in wells filled with horseradish peroxidase-labeled goat anti-dogimmunoglobulin G (heavy plus light chains) (Kirkegaard & Perry Laboratories,Gaithersburg, MD) diluted at 1:1,000 in PBS–Tween 20 and incubated for 1 h atroom temperature. Samples were washed four times, and then the peptide pinswere incubated for 20 min at room temperature with the horseradish peroxidasesubstrate 2,2�-azido-di-(3-ethyl)-benzthiazoline-6-sulfonic acid (Sigma, St. Louis,MO) in 70 mM citrate buffer (pH 4.2) applied to a new plate. Absorbance valuesat 405 and 492 nm were measured in an ELISA plate reader (Molecular Devices,Sunnyvale, CA). Each assay was repeated at least three times. The cutoff value(optical density at 405 nm [OD405] � OD492) for positive reaction was set as themean OD405 � OD492 � 3 standard deviations of the negative control plasma.
Nucleotide sequence accession number. The final sequence assembled fromthe entire E. ewingii omp-1 locus was deposited in GenBank under accession no.EF116932.
RESULTS
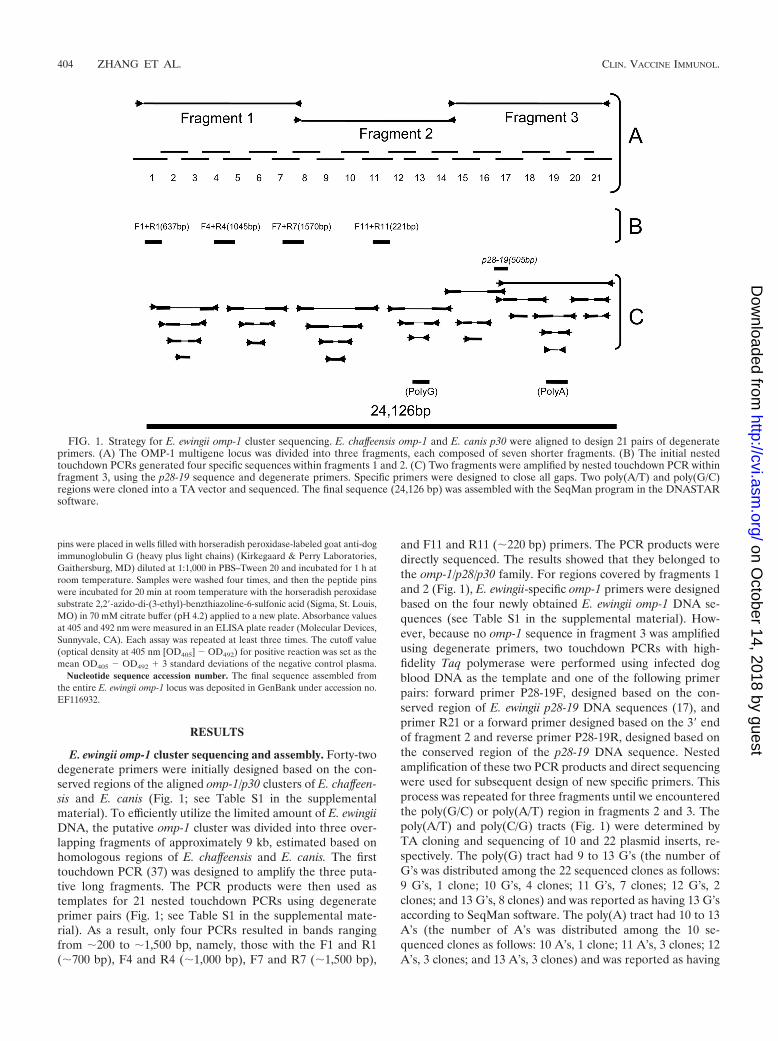
E. ewingii omp-1 cluster sequencing and assembly. Forty-twodegenerate primers were initially designed based on the con-served regions of the aligned omp-1/p30 clusters of E. chaffeen-sis and E. canis (Fig. 1; see Table S1 in the supplementalmaterial). To efficiently utilize the limited amount of E. ewingiiDNA, the putative omp-1 cluster was divided into three over-lapping fragments of approximately 9 kb, estimated based onhomologous regions of E. chaffeensis and E. canis. The firsttouchdown PCR (37) was designed to amplify the three puta-tive long fragments. The PCR products were then used astemplates for 21 nested touchdown PCRs using degenerateprimer pairs (Fig. 1; see Table S1 in the supplemental mate-rial). As a result, only four PCRs resulted in bands rangingfrom �200 to �1,500 bp, namely, those with the F1 and R1(�700 bp), F4 and R4 (�1,000 bp), F7 and R7 (�1,500 bp),
and F11 and R11 (�220 bp) primers. The PCR products weredirectly sequenced. The results showed that they belonged tothe omp-1/p28/p30 family. For regions covered by fragments 1and 2 (Fig. 1), E. ewingii-specific omp-1 primers were designedbased on the four newly obtained E. ewingii omp-1 DNA se-quences (see Table S1 in the supplemental material). How-ever, because no omp-1 sequence in fragment 3 was amplifiedusing degenerate primers, two touchdown PCRs with high-fidelity Taq polymerase were performed using infected dogblood DNA as the template and one of the following primerpairs: forward primer P28-19F, designed based on the con-served region of E. ewingii p28-19 DNA sequences (17), andprimer R21 or a forward primer designed based on the 3� endof fragment 2 and reverse primer P28-19R, designed based onthe conserved region of the p28-19 DNA sequence. Nestedamplification of these two PCR products and direct sequencingwere used for subsequent design of new specific primers. Thisprocess was repeated for three fragments until we encounteredthe poly(G/C) or poly(A/T) region in fragments 2 and 3. Thepoly(A/T) and poly(C/G) tracts (Fig. 1) were determined byTA cloning and sequencing of 10 and 22 plasmid inserts, re-spectively. The poly(G) tract had 9 to 13 G’s (the number ofG’s was distributed among the 22 sequenced clones as follows:9 G’s, 1 clone; 10 G’s, 4 clones; 11 G’s, 7 clones; 12 G’s, 2clones; and 13 G’s, 8 clones) and was reported as having 13 G’saccording to SeqMan software. The poly(A) tract had 10 to 13A’s (the number of A’s was distributed among the 10 se-quenced clones as follows: 10 A’s, 1 clone; 11 A’s, 3 clones; 12A’s, 3 clones; and 13 A’s, 3 clones) and was reported as having
FIG. 1. Strategy for E. ewingii omp-1 cluster sequencing. E. chaffeensis omp-1 and E. canis p30 were aligned to design 21 pairs of degenerateprimers. (A) The OMP-1 multigene locus was divided into three fragments, each composed of seven shorter fragments. (B) The initial nestedtouchdown PCRs generated four specific sequences within fragments 1 and 2. (C) Two fragments were amplified by nested touchdown PCR withinfragment 3, using the p28-19 sequence and degenerate primers. Specific primers were designed to close all gaps. Two poly(A/T) and poly(G/C)regions were cloned into a TA vector and sequenced. The final sequence (24,126 bp) was assembled with the SeqMan program in the DNASTARsoftware.
404 ZHANG ET AL. CLIN. VACCINE IMMUNOL.
on October 14, 2018 by guest
http://cvi.asm.org/
Dow
nloaded from
12 A’s according to SeqMan software. The predominant in-frame sequences in each region were deposited in GenBank. Thefinal sequence assembled from the entire E. ewingii omp-1 locuscontained 24,126 bp. The G�C content of the E. ewingii omp-1cluster was 28.74%, which is similar to those of the E. canis, E.chaffeensis, and E. ruminantium clusters (29.36%, 30.95%, and27.19%, respectively). Sequence identities of the entire E. ewingiiomp-1 cluster relative to E. canis, E. chaffeensis, and E. ruminan-tium were 28.4%, 22.2%, and 14.8%, respectively.
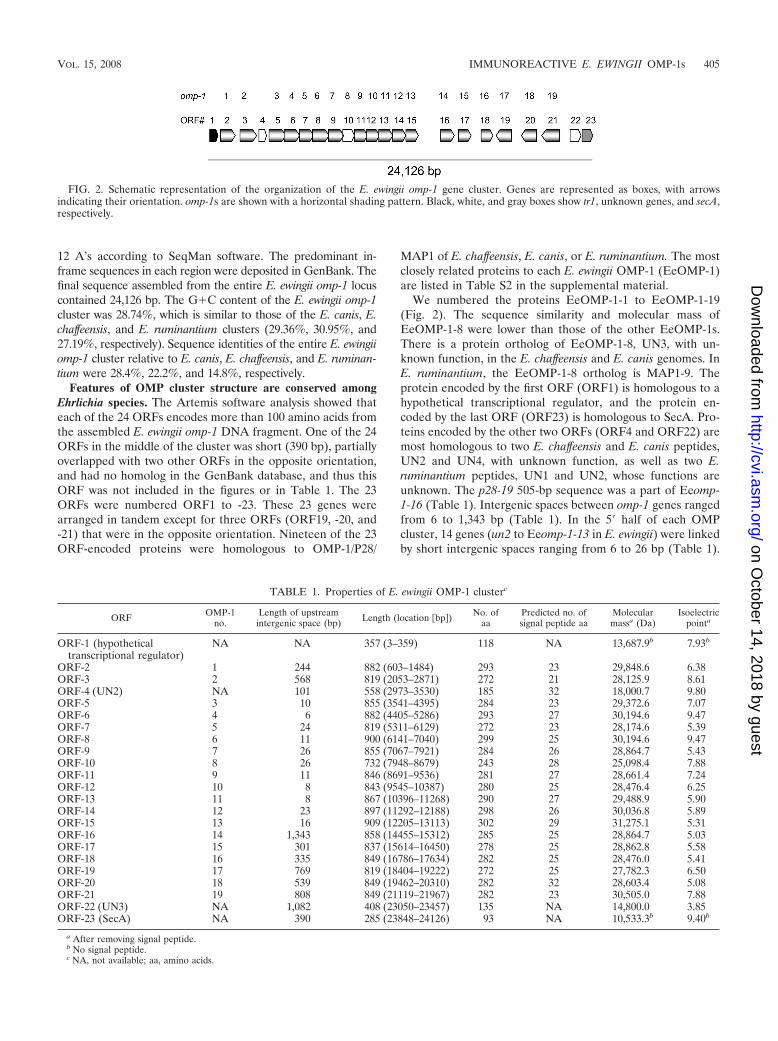
Features of OMP cluster structure are conserved amongEhrlichia species. The Artemis software analysis showed thateach of the 24 ORFs encodes more than 100 amino acids fromthe assembled E. ewingii omp-1 DNA fragment. One of the 24ORFs in the middle of the cluster was short (390 bp), partiallyoverlapped with two other ORFs in the opposite orientation,and had no homolog in the GenBank database, and thus thisORF was not included in the figures or in Table 1. The 23ORFs were numbered ORF1 to -23. These 23 genes werearranged in tandem except for three ORFs (ORF19, -20, and-21) that were in the opposite orientation. Nineteen of the 23ORF-encoded proteins were homologous to OMP-1/P28/
MAP1 of E. chaffeensis, E. canis, or E. ruminantium. The mostclosely related proteins to each E. ewingii OMP-1 (EeOMP-1)are listed in Table S2 in the supplemental material.
We numbered the proteins EeOMP-1-1 to EeOMP-1-19(Fig. 2). The sequence similarity and molecular mass ofEeOMP-1-8 were lower than those of the other EeOMP-1s.There is a protein ortholog of EeOMP-1-8, UN3, with un-known function, in the E. chaffeensis and E. canis genomes. InE. ruminantium, the EeOMP-1-8 ortholog is MAP1-9. Theprotein encoded by the first ORF (ORF1) is homologous to ahypothetical transcriptional regulator, and the protein en-coded by the last ORF (ORF23) is homologous to SecA. Pro-teins encoded by the other two ORFs (ORF4 and ORF22) aremost homologous to two E. chaffeensis and E. canis peptides,UN2 and UN4, with unknown function, as well as two E.ruminantium peptides, UN1 and UN2, whose functions areunknown. The p28-19 505-bp sequence was a part of Eeomp-1-16 (Table 1). Intergenic spaces between omp-1 genes rangedfrom 6 to 1,343 bp (Table 1). In the 5� half of each OMPcluster, 14 genes (un2 to Eeomp-1-13 in E. ewingii) were linkedby short intergenic spaces ranging from 6 to 26 bp (Table 1).
FIG. 2. Schematic representation of the organization of the E. ewingii omp-1 gene cluster. Genes are represented as boxes, with arrowsindicating their orientation. omp-1s are shown with a horizontal shading pattern. Black, white, and gray boxes show tr1, unknown genes, and secA,respectively.
TABLE 1. Properties of E. ewingii OMP-1 clusterc
ORF OMP-1no.
Length of upstreamintergenic space (bp) Length (location �bp) No. of
aaPredicted no. ofsignal peptide aa
Molecularmassa (Da)
Isoelectricpointa
ORF-1 (hypotheticaltranscriptional regulator)
NA NA 357 (3–359) 118 NA 13,687.9b 7.93b
ORF-2 1 244 882 (603–1484) 293 23 29,848.6 6.38ORF-3 2 568 819 (2053–2871) 272 21 28,125.9 8.61ORF-4 (UN2) NA 101 558 (2973–3530) 185 32 18,000.7 9.80ORF-5 3 10 855 (3541–4395) 284 23 29,372.6 7.07ORF-6 4 6 882 (4405–5286) 293 27 30,194.6 9.47ORF-7 5 24 819 (5311–6129) 272 23 28,174.6 5.39ORF-8 6 11 900 (6141–7040) 299 25 30,194.6 9.47ORF-9 7 26 855 (7067–7921) 284 26 28,864.7 5.43ORF-10 8 26 732 (7948–8679) 243 28 25,098.4 7.88ORF-11 9 11 846 (8691–9536) 281 27 28,661.4 7.24ORF-12 10 8 843 (9545–10387) 280 25 28,476.4 6.25ORF-13 11 8 867 (10396–11268) 290 27 29,488.9 5.90ORF-14 12 23 897 (11292–12188) 298 26 30,036.8 5.89ORF-15 13 16 909 (12205–13113) 302 29 31,275.1 5.31ORF-16 14 1,343 858 (14455–15312) 285 25 28,864.7 5.03ORF-17 15 301 837 (15614–16450) 278 25 28,862.8 5.58ORF-18 16 335 849 (16786–17634) 282 25 28,476.0 5.41ORF-19 17 769 819 (18404–19222) 272 25 27,782.3 6.50ORF-20 18 539 849 (19462–20310) 282 32 28,603.4 5.08ORF-21 19 808 849 (21119–21967) 282 23 30,505.0 7.88ORF-22 (UN3) NA 1,082 408 (23050–23457) 135 NA 14,800.0 3.85ORF-23 (SecA) NA 390 285 (23848–24126) 93 NA 10,533.3b 9.40b
a After removing signal peptide.b No signal peptide.c NA, not available; aa, amino acids.
VOL. 15, 2008 IMMUNOREACTIVE E. EWINGII OMP-1s 405
on October 14, 2018 by guest
http://cvi.asm.org/
Dow
nloaded from

Eight genes in the 3� half (Eeomp-1-14 to Eeomp-1-19) wereconnected by longer intergenic spaces, ranging from 301 to 808bp. Thus, features of the OMP cluster structure were con-served among E. ewingii, E. canis, E. chaffeensis, and E. rumi-nantium, with the exception of the opposite orientation ofthree genes at the 3� end in E. ewingii instead of one gene (E.canis and E. ruminantium) or two genes (E. chaffeensis).
After removal of the signal peptide sequence, predicted mo-lecular masses of mature E. ewingii OMP-1s ranged from 25.1to 31.3 kDa. The predicted signal peptides ranged from 21 to32 amino acids. The predicted isoelectric points of the matureOMP-1s were 5.03 to 9.80. Properties of the ORFs of the E.ewingii omp-1 cluster, including predicted signal peptidelengths, molecular masses of mature proteins, and isoelectricpoints, are shown in Table 1.
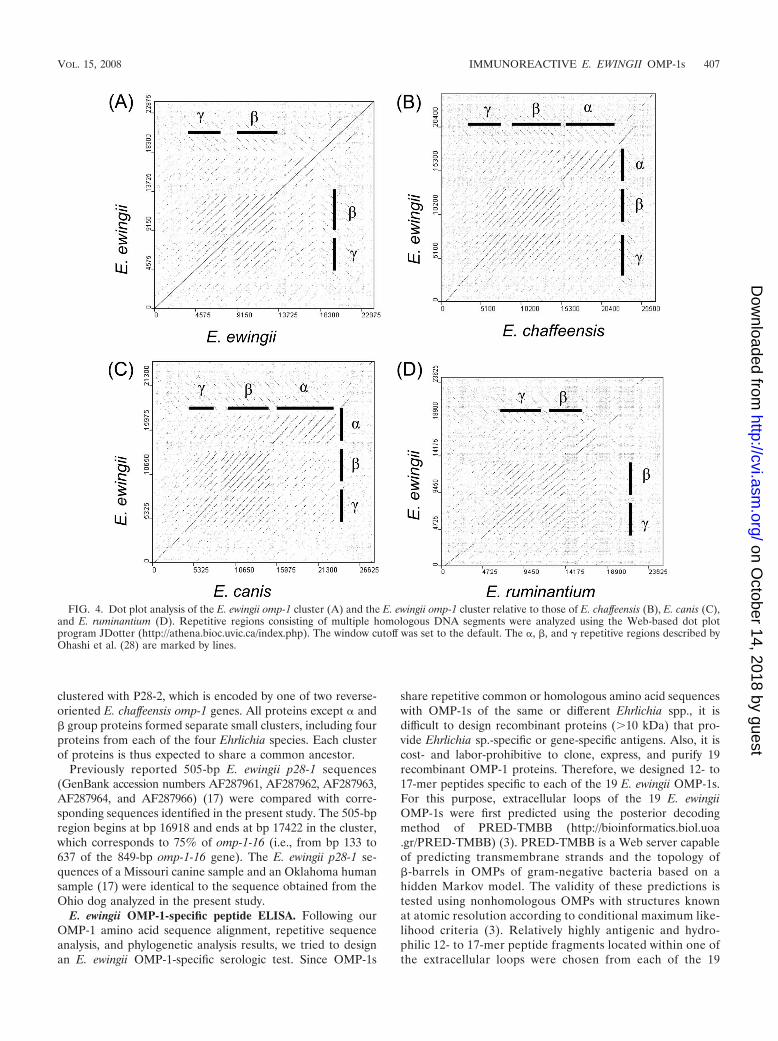
omp-1/p28/map1 gene clusters display synteny at the 5� end.The synteny among entire OMP-1 gene clusters of E. ewingii andthree related Ehrlichia species was analyzed by the Artemis com-parison tool, and the results are shown in Fig. 3. The genes at the5� ends of the omp-1 clusters were more highly conserved thanwere genes in the central region or the 3� end (Fig. 3). Previously,we defined three repeat sequence regions, , �, and �, in omp-1clusters of E. chaffeensis and E. canis (28). The dot plot analysis ofthe E. ewingii omp-1 cluster and the dot plot between E. ewingii
and E. ruminantium revealed only � and � repeat regions, as the regions of these bacteria are too short (Fig. 4). The � repeatregion in E. ruminantium was shorter than those of E.chaffeensis and E. canis. The dot plot analyses between E.ewingii and E. chaffeensis and between E. ewingii and E. canisshowed three clear repeat regions, indicating that the region is expanded in E. canis and E. chaffeensis.
Phylogenetic analysis of all 79 OMPs of E. ewingii, E.chaffeensis, E. canis, and E. ruminantium is shown in Fig. 5. Thepreviously defined and �1 regions in the E. chaffeensis omp-1cluster (28) encoded five (P28, OMP-1F, -1D, -1C, and -1E)and four (OMP-1H, -1A, -1S, and -1Z) proteins, respectively,and the and �1 regions in the E. canis p30 cluster encoded six(P30, P30-1, P30-2, P30-3, P30-4, and P30a) and four (P30-6,P30-5, P30-7, and P30-8) proteins, respectively. However, in E.ewingii, the and �1 regions each encoded two proteins(EeOMP-1-15/EeOMP-1-16 and EeOMP-1-12/EeOMP-1-13,respectively). In E. ruminantium, the region encoded onlyone protein (MAP1) and the �1 region encoded two proteins(MAP1-2 and MAP1-3) (Fig. 5).
EeOMP-1-8 and MAP1-5 were far removed from the re-maining OMP-1s, raising the possibility that they do not belongto the OMP cluster (Fig. 5). EeOMP-1-18 and EeOMP-1-19,which are encoded by genes in the reverse orientation, were
FIG. 3. Synteny analysis of the E. ewingii (Ee) omp-1 cluster relative to those of E. chaffeensis (Ech), E. canis (Eca), and E. ruminantium (Eru),using the Artemis comparison tool.
406 ZHANG ET AL. CLIN. VACCINE IMMUNOL.
on October 14, 2018 by guest
http://cvi.asm.org/
Dow
nloaded from
clustered with P28-2, which is encoded by one of two reverse-oriented E. chaffeensis omp-1 genes. All proteins except and� group proteins formed separate small clusters, including fourproteins from each of the four Ehrlichia species. Each clusterof proteins is thus expected to share a common ancestor.
Previously reported 505-bp E. ewingii p28-1 sequences(GenBank accession numbers AF287961, AF287962, AF287963,AF287964, and AF287966) (17) were compared with corre-sponding sequences identified in the present study. The 505-bpregion begins at bp 16918 and ends at bp 17422 in the cluster,which corresponds to 75% of omp-1-16 (i.e., from bp 133 to637 of the 849-bp omp-1-16 gene). The E. ewingii p28-1 se-quences of a Missouri canine sample and an Oklahoma humansample (17) were identical to the sequence obtained from theOhio dog analyzed in the present study.
E. ewingii OMP-1-specific peptide ELISA. Following ourOMP-1 amino acid sequence alignment, repetitive sequenceanalysis, and phylogenetic analysis results, we tried to designan E. ewingii OMP-1-specific serologic test. Since OMP-1s
share repetitive common or homologous amino acid sequenceswith OMP-1s of the same or different Ehrlichia spp., it isdifficult to design recombinant proteins (�10 kDa) that pro-vide Ehrlichia sp.-specific or gene-specific antigens. Also, it iscost- and labor-prohibitive to clone, express, and purify 19recombinant OMP-1 proteins. Therefore, we designed 12- to17-mer peptides specific to each of the 19 E. ewingii OMP-1s.For this purpose, extracellular loops of the 19 E. ewingiiOMP-1s were first predicted using the posterior decodingmethod of PRED-TMBB (http://bioinformatics.biol.uoa.gr/PRED-TMBB) (3). PRED-TMBB is a Web server capableof predicting transmembrane strands and the topology of�-barrels in OMPs of gram-negative bacteria based on ahidden Markov model. The validity of these predictions istested using nonhomologous OMPs with structures knownat atomic resolution according to conditional maximum like-lihood criteria (3). Relatively highly antigenic and hydro-philic 12- to 17-mer peptide fragments located within one ofthe extracellular loops were chosen from each of the 19
FIG. 4. Dot plot analysis of the E. ewingii omp-1 cluster (A) and the E. ewingii omp-1 cluster relative to those of E. chaffeensis (B), E. canis (C),and E. ruminantium (D). Repetitive regions consisting of multiple homologous DNA segments were analyzed using the Web-based dot plotprogram JDotter (http://athena.bioc.uvic.ca/index.php). The window cutoff was set to the default. The , �, and � repetitive regions described byOhashi et al. (28) are marked by lines.
VOL. 15, 2008 IMMUNOREACTIVE E. EWINGII OMP-1s 407
on October 14, 2018 by guest
http://cvi.asm.org/
Dow
nloaded from

EeOMP-1 amino acid sequences, based on DNASTAR Pro-tean analysis. Using the program BLAST, these peptidesequences were compared with the entire E. ewingii omp-1locus and the E. chaffeensis, E. canis, and E. ruminantiumgenome sequences to synthesize one peptide specific to eachof the 19 EeOMP-1s (Table 2).
Plasmas from three dogs experimentally infected with E. ewin-gii and preinfection plasmas from four dogs were then tested inELISAs containing the 19 EeOMP-1-specific peptides. Thirteenpeptides (EeOMP-1-1, EeOMP-1-2, EeOMP-1-3, EeOMP-1-4,EeOMP-1-5, EeOMP-1-8, EeOMP-1-9, EeOMP-1-10, EeOMP-1-13, EeOMP-1-14, EeOMP-1-15, EeOMP-1-16, and EeOMP-1-19) were consistently recognized with plasmas from three dogs
experimentally infected with E. ewingii compared with preinfec-tion dog plasmas (Fig. 6).
By peptide-pin ELISA, eight peptides (EeOMP-1-1, EeOMP-1-3, EeOMP-1-4, EeOMP-1-5, EeOMP-1-13, EeOMP-1-14,EeOMP-1-16, and EeOMP-1-19) with high sensitivities (indi-cated by the ratio of E. ewingii plasma reactivity to control plasmareactivity) did not cross-react with plasmas from E. chaffeensis-infected dogs (indicated by the ratio of E. chaffeensis plasmareactivity to control plasma reactivity [�1.00]) (Fig. 6). The sameELISA was performed using three E. canis-infected dog plasmas.Based on the ratios of E. ewingii plasma reactivity to controlplasma reactivity and E. canis plasma reactivity to control plasmareactivity, EeOMP-1-9, EeOMP-1-11, EeOMP-1-12, EeOMP-1-
FIG. 5. Phylogram of OMPs of E. ewingii, E. chaffeensis, E. canis, and E. ruminantium. A total of 39 OMPs were segregated into 10 clusters withfour or three Ehrlichia species each, but 40 remaining proteins were not. The tree was constructed using the neighbor-joining (NEIGHBORprogram from PHYLIP) method, based on an alignment generated with CLUSTAL V; 1,000 bootstrap replications were performed. The nodessupported by bootstrap values of �60% are labeled. The OMPs encoded by the three repetitive regions in Fig. 4 are indicated by , �1, and �2.Branch lengths are proportional to percent divergence. The calibration bar is on the lower left.
408 ZHANG ET AL. CLIN. VACCINE IMMUNOL.
on October 14, 2018 by guest
http://cvi.asm.org/
Dow
nloaded from

13, EeOMP-1-14, and EeOMP-1-15 showed high sensitivities andgood specificities (data not shown). The temporal reactivities ofE. ewingii-infected dog sera to these peptides showed thatEeOMP-1-13, EeOMP-1-14, EeOMP-1-15, and EeOMP-1-16gave higher sensitivities than the remaining peptides and gavesignals above that of control dog plasma when E. ewingii PCRbecame positive (at day 21 postexposure) (Fig. 7). While morespecimens need to be tested and peptide antigens can be modi-fied, several of these EeOMP-1 peptides may serve as good can-didate antigens for E. ewingii serodiagnosis to distinguish E. ew-ingii infection from both E. chaffeensis and E. canis infections.
DISCUSSION
In the present study, for the first time, the entire 24-kb E.ewingii omp-1 locus, containing 19 omp-1 genes, was se-quenced. Since the only available source of E. ewingii DNA wasa small amount of an infected dog blood specimen, we em-ployed touchdown PCR. This method has been used previouslyto amplify a small amount of fragmented Aegyptianella pullo-rum DNA from archival paraffin sections on glass slides (36).Incorrect base calls resulting from amplification or sequencingerrors were minimized in the present study because large poolsof PCR products were directly sequenced. In addition, multi-ple overlapping regions throughout the sequences ensured thereliability of sequencing results. So far, only a few E. ewingiigenes, including the 16S rRNA gene (1, 16, 48), groESL (39),p28-19 (17), dsbA (GenBank accession no. DQ902688), gltA(GenBank accession no. DQ365879), and the disulfide oxi-doreductase gene (19), have been reported. Applying a similarapproach to that used here, it would be possible to obtain
TABLE 2. E. ewingii OMP-1 peptide sequences used in ELISA
OMP-1 no. Amino acid sequencePeptide length
(aaa �aa position inthe protein sequence)
OMP-1-1 SCTEQEMKPAQQNGSSK 17 (151–167)OMP-1-2 EQHFALASELDTNGNQ 16 (130–145)OMP-1-3 VSAPSGYDDNIYAYFSI 17 (123–139)OMP-1-4 FAIPRDTFFNNSIPY 15 (144–158)OMP-1-5 NSSSLISSNNHYTQLY 16 (129–144)OMP-1-6 KDNNQVQPKAHDSSSTD 17 (148–164)OMP-1-7 KDPKDCSVKDAFRHL 15 (130–144)OMP-1-8 TEDKYLTSEQEVNDY 15 (120–134)OMP-1-9 ICKENDKPTPKEKKY 15 (145–159)OMP-1-10 YRYFAIAREMNSSSNNQ 17 (137–153)OMP-1-11 KNSGHSSIDAHR 12 (136–147)OMP-1-12 IESDQNKFQPKNANSNS 17 (154–170)OMP-1-13 SESSKEPQPKNPNSAGN 17 (159–175)OMP-1-14 KYYGLFREGTPQEEEH 16 (146–161)OMP-1-15 SRKDNANIGTTPQDKK 16 (141–156)OMP-1-16 KIEDNQVQNKFTISNY 16 (76–91)OMP-1-17 QFYREGSNNYKF 12 (123–134)OMP-1-18 VQDTKSHIVDDNYR 14 (121–134)OMP-1-19 SKQDNLNSDYVTLIN 15 (171–185)
a aa, amino acids.
FIG. 6. ELISA analysis of E. ewingii- and E. chaffeensis-infected dogs with the 19 EeOMP-1 oligopeptides. Preinfection control and postin-fection plasmas from dogs were allowed to react with the 19 synthesized EeOMP-1-specific peptides. The y axis shows the OD405 – OD492 values.A reaction was considered positive when the plasmas from infected dogs yielded an OD405 – OD492 value greater than the mean OD405 – OD492value for preinfection control plasma plus 3 standard deviations (dashed line with closed triangles). Representative data from three to five assaysare shown. Reactivity ratios of E. ewingii/control plasma values (EW/Cont.) and E. chaffeensis/control plasma values (ECH/Cont.) were calculatedbased on the averages for three E. ewingii-positive and three E. chaffeensis-positive samples, respectively, and four negative control samples.EeOMP-1 peptides that showed good sensitivity and specificity for detecting E. ewingii infection are underlined.
FIG. 7. Temporal development of antibodies to EeOMP-1 pep-tides in an E. ewingii-infected dog. Plasmas collected on 0, 21, 28, and123 days postinfection were assayed using selected EeOMP-1 peptides13, 14, 15, and 16 by ELISA. Nested PCR using E. ewingii-specificprimers was negative on day 0 and positive on days 21, 28, and 123postinfection (47). The y axis shows OD405 – OD492 values.
VOL. 15, 2008 IMMUNOREACTIVE E. EWINGII OMP-1s 409
on October 14, 2018 by guest
http://cvi.asm.org/
Dow
nloaded from

DNA sequences of other genomic regions to further our un-derstanding of this uncultivable emerging zoonotic pathogen.
Because E. ewingii infects granulocytes, the distinction be-tween E. ewingii and strains of A. phagocytophilum was unclearprior to the molecular era. However, in concordance with the16S rRNA and groESL sequence-based classification of thisbacterium (1, 39), our finding of the complete OMP-1 clusterstructure flanked with tr1 and secA clearly demonstrated thatE. ewingii belongs to the genus Ehrlichia. Synteny analysissuggests that the OMP clusters existed in a common ancestorof the present-day four Ehrlichia species. Furthermore, thelocus appears to have been partially scrambled as speciesevolved. The E. ewingii OMP-1 cluster has greater synteny withmonocytotropic E. chaffeensis and E. canis than with endothe-liotropic E. ruminantium. Whether OMP-1s and host cell typespecificity coevolved remains to be studied.
The present study revealed 19 E. ewingii OMP-1 amino acidsequences and 19 E. ewingii immunogenic amino acid se-quences. Studies of E. chaffeensis have shown an important rolefor OMP-1s/P28s in the stimulation of the host immune re-sponse and protection of the host from infection. Immuniza-tion with recombinant P28 (one of the major OMP-1/P28 fam-ily members) protected mice from E. chaffeensis challenge (30).A monoclonal antibody against OMP-1g (P28) mediated pro-tection of SCID mice from fatal E. chaffeensis infection (20).While antibodies against a single OMP-1 protein confer partialprotection, the existence of multiple homologous surface pro-teins has been suspected to allow evasion of the host immuneresponse. A recent proteomic study showed that 18 of 21 E.chaffeensis OMP-1/P28 family proteins are indeed bacterialsurface exposed, supporting the idea of immunoevasion (14).The number of E. ewingii omp-1 genes found in the OMP-1cluster (19 copies) was similar to those for E. canis (22 copies,but there is an additional locus with three p30s) (28), E.chaffeensis (22 copies), and E. ruminantium (16 copies). Inaddition, there is extensive diversification among omp-1 genesof E. ewingii, similar to the case for other Ehrlichia spp., sup-porting the hypothesis that multiple omp-1/p28 paralogspresent in Ehrlichia spp. are involved in immunoavoidance.Thus, these studies suggest that incorporation of immunogenicpeptides of multiple OMP-1s in a vaccine preparation wouldprovide better protection against Ehrlichia infection than theuse of a single OMP-1 in a vaccine.
Multiple OMP-1/P28 and P30 mRNAs are expressed by E.chaffeensis and E. canis during experimental infections of dogswith these bacteria (42, 44). All 22 E. chaffeensis P28 recom-binant antigens were recognized by sera from two dogs exper-imentally infected with E. chaffeensis (52). Similarly, thepresent results suggest that all 19 EeOMP-1 peptides wererecognized by the plasmas from three E. ewingii-infected dogs.Thus, the lack of immunological cross-reactivity of E. canis andE. chaffeensis OMP-1/P28/P30 with plasmas from human pa-tients or dogs infected with E. ewingii in previous studies (6, 34,35, 43) was likely due to divergence of the amino acid se-quences of the E. ewingii OMP-1s from those of the E. canisand E. chaffeensis OMP-1/P28/P30 proteins expressed in cellculture. It is also most likely for E. ewingii-infected humans anddogs that different combinations of multiple OMP-1s are ex-pressed at different stages of infection and under differentimmune and health statuses of animals. Considering these
observations, for serodiagnosis of E. ewingii infection in bothhumans and animals, the use of a combination of EeOMP-1s asthe antigen is expected to provide more sensitive and specificserodiagnosis than the use of a single EeOMP-1 antigen. Fur-thermore, all of the EeOMP-1 amino acid sequences obtainedin the present study would help in optimizing peptide antigensto provide desired specificity and sensitivity to detect poten-tially diverse E. ewingii strains in the field. The entireEeOMP-1 DNA sequence data also obtained in the presentstudy should help in improving diagnostic PCRs for human anddog E. ewingii infections to make this direct test more reliablefor all infective species. Lastly, the sequencing and compara-tive analysis of all of the E. ewingii OMP-1s in the present studyshould greatly advance E. ewingii epidemiologic and immuno-logic studies.
ACKNOWLEDGMENTS
This work was supported in part by National Institutes of Healthgrants R01AI47407 and R01AI30010.
We appreciate K. Gibson for assistance with preparation of themanuscript.
REFERENCES
1. Anderson, B. E., C. E. Greene, D. C. Jones, and J. E. Dawson. 1992. Ehrlichiaewingii sp. nov., the etiologic agent of canine granulocytic ehrlichiosis. Int. J.Syst. Bacteriol. 42:299–302.
2. Anziani, O. S., S. A. Ewing, and R. W. Barker. 1990. Experimental trans-mission of a granulocytic form of the tribe Ehrlichieae by Dermacentorvariabilis and Amblyomma americanum to dogs. Am. J. Vet. Res. 51:929–931.
3. Bagos, P. G., T. D. Liakopoulos, I. C. Spyropoulos, and S. J. Hamodrakas.2004. PRED-TMBB: a web server for predicting the topology of beta-barrelouter membrane proteins. Nucleic Acids Res. 32:W400–W404.
4. Bakken, J. S., and J. S. Dumler. 2006. Clinical diagnosis and treatment ofhuman granulocytotropic anaplasmosis. Ann. N. Y. Acad. Sci. 1078:236–247.
5. Bell-Sakyi, L., E. B. Koney, O. Dogbey, K. J. Sumption, A. R. Walker, A.Bath, and F. Jongejan. 2003. Detection by two enzyme-linked immunosor-bent assays of antibodies to Ehrlichia ruminantium in field sera collectedfrom sheep and cattle in Ghana. Clin. Diagn. Lab. Immunol. 10:917–925.
6. Buller, R. S., M. Arens, S. P. Hmiel, C. D. Paddock, J. W. Sumner, Y.Rikhisa, A. Unver, M. Gaudreault-Keener, F. A. Manian, A. M. Liddell, N.Schmulewitz, and G. A. Storch. 1999. Ehrlichia ewingii, a newly recognizedagent of human ehrlichiosis. N. Engl. J. Med. 341:148–155.
7. Carver, T. J., K. M. Rutherford, M. Berriman, M. Rajandream, B. G. Barrelland Julian Parkhill. 2005. ACT: the Artemis comparison tool. Bioinformat-ics 21:3422–3423.
8. Dawson, J. E., K. L. Biggie, C. K. Warner, K. Cookson, S. Jenkins, J. F.Levine, and J. G. Olson. 1996. Polymerase chain reaction evidence ofEhrlichia chaffeensis, an etiologic agent of human ehrlichiosis, in dogs fromsoutheast Virginia. Am. J. Vet. Res. 57:1175–1179.
9. Demma, L. J., R. C. Holman, J. H. McQuiston, J. W. Krebs, and D. L.Swerdlow. 2005. Epidemiology of human ehrlichiosis and anaplasmosis inthe United States, 2001–2002. Am. J. Trop. Med. Hyg. 73:400–409.
10. Ewing, S. A., J. E. Dawson, A. A. Kocan, R. W. Barker, C. K. Warner, R. J.Panciera, J. C. Fox, K. M. Kocan, and E. F. Blouin. 1995. Experimentaltransmission of Ehrlichia chaffeensis (Rickettsiales: Ehrlichieae) amongwhite-tailed deer by Amblyomma americanum (Acari: Ixodidae). J. Med.Entomol. 32:368–374.
11. Ewing, S. A., W. R. Roberson, R. G. Buckner, and C. S. Hayat. 1971. A newstrain of Ehrlichia canis. J. Am. Vet. Med. Assoc. 159:1771–1774.
12. Faburay, B., S. Munstermann, D. Geysen, L. Bell-Sakyi, A. Ceesay, C.Bodaan, and F. Jongejan. 2005. Point seroprevalence survey of Ehrlichiaruminantium infection in small ruminants in The Gambia. Clin. Diagn. Lab.Immunol. 12:508–512.
13. Felsenstein, J. 1989. PHYLIP—phylogeny inference package (version 3.2).Cladistics 5:164–166.
14. Ge, Y., and Y. Rikihisa. 2007. Surface-exposed proteins of Ehrlichia chaffeen-sis. Infect. Immun. 75:3833–3841.
15. Geysen, H. M. 1990. Molecular technology: peptide epitope mapping and thepin technology. Southeast Asian J. Trop. Med. Public Health 21:523–533.
16. Goldman, E. E., E. B. Breitschwerdt, C. B. Grindem, B. C. Hegarty, J. J.Walls, and J. S. Dumler. 1998. Granulocytic ehrlichiosis in dogs from NorthCarolina and Virginia. J. Vet. Intern. Med. 12:61–70.
17. Gusa, A. A., R. S. Buller, G. A. Storch, M. M. Huycke, L. J. Machado, L. N.Slater, S. L. Stockham, and R. F. Massung. 2001. Identification of a p28 gene
410 ZHANG ET AL. CLIN. VACCINE IMMUNOL.
on October 14, 2018 by guest
http://cvi.asm.org/
Dow
nloaded from

in Ehrlichia ewingii: evaluation of gene for use as a target for a species-specific PCR diagnostic assay. J. Clin. Microbiol. 39:3871–3876.
18. Ijdo, J. W., W. Sun, Y. Zhang, L. A. Magnarelli, and E. Fikrig. 1998. Cloningof the gene encoding the 44-kilodalton antigen of the agent of humangranulocytic ehrlichiosis and characterization of the humoral response. In-fect. Immun. 66:3264–3269.
19. Labruna, M. B., J. W. McBride, L. M. Camargo, D. M. Aguiar, M. J. Yabsley,W. R. Davidson, E. Y. Stromdahl, P. C. Williamson, R. W. Stich, S. W. Long,E. P. Camargo, and D. H. Walker. 2007. A preliminary investigation ofEhrlichia species in ticks, humans, dogs, and capybaras from Brazil. Vet.Parasitol. 143:189–195.
20. Li, J. S., F. Chu, A. Reilly, and G. M. Winslow. 2002. Antibodies highlyeffective in SCID mice during infection by the intracellular bacteriumEhrlichia chaffeensis are of picomolar affinity and exhibit preferential epitopeand isotype utilization. J. Immunol. 169:1419–1425.
21. Liddell, A. M., S. L. Stockham, M. A. Scott, J. W. Sumner, C. D. Paddock,M. Gaudreault-Keener, M. Q. Arens, and G. A. Storch. 2003. Predominanceof Ehrlichia ewingii in Missouri dogs. J. Clin. Microbiol. 41:4617–4622.
22. Lodes, M. J., R. Mohamath, L. D. Reynolds, P. McNeill, C. P. Kolbert, E. S.Bruinsma, D. R. Benson, E. Hofmeister, S. G. Reed, R. L. Houghton, andD. H. Persing. 2001. Serodiagnosis of human granulocytic ehrlichiosis byusing novel combinations of immunoreactive recombinant proteins. J. Clin.Microbiol. 39:2466–2476.
23. Magnarelli, L. A., J. W. Ijdo, A. E. Van Andel, C. Wu, and E. Fikrig. 2001.Evaluation of a polyvalent enzyme-linked immunosorbent assay incorporat-ing a recombinant p44 antigen for diagnosis of granulocytic ehrlichiosis indogs and horses. Am. J. Vet. Res. 62:29–32.
24. McBride, J. W., R. E. Corstvet, E. B. Breitschwerdt, and D. H. Walker. 2001.Immunodiagnosis of Ehrlichia canis infection with recombinant proteins.J. Clin. Microbiol. 39:315–322.
25. McBride, J. W., X. J. Yu, and D. H. Walker. 2000. A conserved, transcrip-tionally active p28 multigene locus of Ehrlichia canis. Gene 254:245–252.
26. Murphy, G. L., S. A. Ewing, L. C. Whitworth, J. C. Fox, and A. A. Kocan.1998. A molecular and serologic survey of Ehrlichia canis, E. chaffeensis, andE. ewingii in dogs and ticks from Oklahoma. Vet. Parasitol. 79:325–339.
27. Ndip, L. M., R. N. Ndip, S. N. Esemu, V. L. Dickmu, E. B. Fokam, D. H.Walker, and J. W. McBride. 2005. Ehrlichial infection in Camerooniancanines by Ehrlichia canis and Ehrlichia ewingii. Vet. Microbiol. 111:59–66.
28. Ohashi, N., Y. Rikihisa, and A. Unver. 2001. Analysis of transcriptionallyactive gene clusters of major outer membrane protein multigene family inEhrlichia canis and E. chaffeensis. Infect. Immun. 69:2083–2091.
29. Ohashi, N., A. Unver, N. Zhi, and Y. Rikihisa. 1998. Cloning and character-ization of multigenes encoding the immunodominant 30-kilodalton majorouter membrane proteins of Ehrlichia canis and application of the recombi-nant protein for serodiagnosis. J. Clin. Microbiol. 36:2671–2680.
30. Ohashi, N., N. Zhi, Y. Zhang, and Y. Rikihisa. 1998. Immunodominantmajor outer membrane proteins of Ehrlichia chaffeensis are encoded by apolymorphic multigene family. Infect. Immun. 66:132–139.
31. Paddock, C. D., and J. E. Childs. 2003. Ehrlichia chaffeensis: a prototypicalemerging pathogen. Clin. Microbiol. Rev. 16:37–64.
32. Peter, T. F., C. J. O’Callaghan, G. F. Medley, B. D. Perry, S. M. Semu, andS. M. Maha. 2001. Population-based evaluation of the Ehrlichia ruminantiumMAP 1B indirect ELISA. Exp. Appl. Acarol. 25:881–897.
33. Reddy, G. R., C. R. Sulsona, A. F. Barbet, S. M. Mahan, M. J. Burridge, andA. R. Alleman. 1998. Molecular characterization of a 28 kDa surface antigengene family of the tribe Ehrlichiae. Biochem. Biophys. Res. Commun. 247:636–643.
34. Rikihisa, Y., S. A. Ewing, and J. C. Fox. 1994. Western immunoblot analysisof Ehrlichia chaffeensis, E. canis, or E. ewingii infections in dogs and humans.J. Clin. Microbiol. 32:2107–2112.
35. Rikihisa, Y., S. A. Ewing, J. C. Fox, A. G. Siregar, F. H. Pasaribu, and M. B.Malole. 1992. Analyses of Ehrlichia canis and a canine granulocytic Ehrlichiainfection. J. Clin. Microbiol. 30:143–148.
36. Rikihisa, Y., C. Zhang, and B. M. Christensen. 2003. Molecular character-
ization of Aegyptianella pullorum (Rickettsiales, Anaplasmataceae). J. Clin.Microbiol. 41:5294–5297.
37. Roux, K. H., and K. H. Hecker. 1997. One-step optimization using touch-down and stepdown PCR. Methods Mol. Biol. 67:39–45.
38. Rutherford, J., J. Parkhill, J. Crook, T. Horsnell, P. Rice, M. Rajandream,and B. Barrell. 2000. Artemis: sequence visualization and annotation. Bioin-formatics 16:944–945.
39. Sumner, J. W., G. A. Storch, R. S. Buller, A. M. Liddell, S. L. Stockham, Y.Rikihisa, S. Messenger, and C. D. Paddock. 2000. PCR amplification andphylogenetic analysis of groESL operon sequences from Ehrlichia ewingii andEhrlichia muris. J. Clin. Microbiol. 38:2746–2749.
40. Tajima, T., N. Zhi, Q. Lin, Y. Rikihisa, H. W. Horowitz, J. Ralfalli, G. P.Wormser, and K. E. Hechemy. 2000. Comparison of two recombinant majorouter membrane proteins of the human granulocytic ehrlichiosis agent foruse in an enzyme-linked immunosorbent assay. Clin. Diagn. Lab. Immunol.7:652–657.
41. Unver, A., S. Felek, C. D. Paddock, N. Zhi, H. W. Horowitz, G. P. Wormser,L. C. Cullman, and Y. Rikihisa. 2001. Western blot analysis of sera reactiveto human monocytic ehrlichiosis and human granulocytic ehrlichiosis agents.J. Clin. Microbiol. 39:3982–3986.
42. Unver, A., N. Ohashi, T. Tajima, R. W. Stich, D. Grover, and Y. Rikihisa.2001. Transcriptional analysis of p30 major outer membrane multigene fam-ily of Ehrlichia canis in dogs, ticks, and cell culture at different temperatures.Infect. Immun. 69:6172–6178.
43. Unver, A., Y. Rikihisa, N. Ohashi, L. C. Cullman, R. Buller, and G. A.Storch. 1999. Western and dot blotting analyses of Ehrlichia chaffeensisindirect fluorescent-antibody assay-positive and -negative human sera byusing native and recombinant E. chaffeensis and E. canis antigens. J. Clin.Microbiol. 37:3888–3895.
44. Unver, A., Y. Rikihisa, R. W. Stich, N. Ohashi, and S. Felek. 2002. The omp-1major outer membrane multigene family of Ehrlichia chaffeensis is differen-tially expressed in canine and tick hosts. Infect. Immun. 70:4701–4704.
45. van Heerden, H., N. E. Collins, K. A. Brayton, C. Rademeyer, and B. A.Allsopp. 2004. Characterization of a major outer membrane protein multi-gene family in Ehrlichia ruminantium. Gene 330:159–168.
46. van Vliet, A. H., F. Jongejan, M. van Kleef, and B. A. van der Zeijst. 1994.Molecular cloning, sequence analysis, and expression of the gene encodingthe immunodominant 32-kilodalton protein of Cowdria ruminantium. Infect.Immun. 62:1451–1456.
47. Xiong, Q., W. Bao, Y. Ge, and Y. Rikihisa. Ehrlichia ewingii infection delaysspontaneous neutrophil apoptosis through stabilization of mitochondria.J. Infect. Dis., in press.
48. Yabsley, M. J., A. S. Varela, C. M. Tate, V. G. Dugan, D. E. Stallknecht, S. E.Little, and W. R. Davidson. 2002. Ehrlichia ewingii infection in white-taileddeer (Odocoileus virginianus). Emerg. Infect. Dis. 8:668–671.
49. Yu, X., J. W. McBride, X. Zhang, and D. H. Walker. 2000. Characterizationof the complete transcriptionally active Ehrlichia chaffeensis 28 kDa outermembrane protein multigene family. Gene 248:59–68.
50. Yu, X. J., P. Crocquet-Valdes, L. C. Cullman, and D. H. Walker. 1996. Therecombinant 120-kilodalton protein of Ehrlichia chaffeensis, a potential di-agnostic tool. J. Clin. Microbiol. 34:2853–2855.
51. Yu, X. J., P. A. Crocquet-Valdes, L. C. Cullman, V. L. Popov, and D. H.Walker. 1999. Comparison of Ehrlichia chaffeensis recombinant proteins forserologic diagnosis of human monocytotropic ehrlichiosis. J. Clin. Microbiol.37:2568–2575.
52. Zhang, J. Z., H. Guo, G. M. Winslow, and X. J. Yu. 2004. Expression ofmembers of the 28-kilodalton major outer membrane protein family ofEhrlichia chaffeensis during persistent infection. Infect. Immun. 72:4336–4343.
53. Zhi, N., N. Ohashi, Y. Rikihisa, H. W. Horowitz, G. P. Wormser, and K.Hechemy. 1998. Cloning and expression of the 44-kilodalton major outermembrane protein gene of the human granulocytic ehrlichiosis agent andapplication of the recombinant protein to serodiagnosis. J. Clin. Microbiol.36:1666–1673.
VOL. 15, 2008 IMMUNOREACTIVE E. EWINGII OMP-1s 411
on October 14, 2018 by guest
http://cvi.asm.org/
Dow
nloaded from




























