Embed Size (px)
Citation preview

MORPHOLOGICAL AND A N A T O M I C A L INVESTIGATIONS ON ARTOCARPUS FORST.
I. Vegetative Organs 1, 2
BY M. R. SHARMA
(School of Plant Morphology, Meerut)
Received June 2, 1962
(Communicated by Dr. V. Puri, F.A.SC.)
INTRODUCTION
Artocarpus Forst. (Artos--Bread; Carpos--fruiO is a unique genus which, on account of its peculiar and edible fruits in certain species, has attracted attention from very remote past. It is essentially a tropical genus with about 50 species (Comer, 1952 and Jarrett, 1959, a). They are distri- buted from India to China. The Indian species are: A. heterophyllus Lamk., A. lakoocha Roxb., A. altilis (Park) Fosb., A. choplasha Roxb. and A. hirsutus Lain. It is said to be the only Angiosperm whose inflorescence pieces are reported from cretaceous reeks of Greenland (Seward, 1941). It belongs to the subfamily Artocarpoide~e, tribe Artocarp~e which is included in Urticace~e (Hooker, 1885; Ridley, 1924 and Comer, 1952) or in Moracea: (Engler and Prantl, 1881; Hutchinson, 1926; Rendle, 1938; Bailey, 1950 and Lawrence, 1951). Jarrett (1959, a, b) has made a revision of Artocarpus and allied genera touching upon some points of morphological interest also.
Wood anatomy of the Morace~e and allied families has been studied in detail by Tippo (1938) and Record and Hess (1940, 1943). Tippo is of the opinion that anatomical specialization in general has proceeded from Moroide~e to Artocarpoidem, Cephaloide~e and Cannaboide~e (Cannabinace~e) and that evolution of floral structures is correlated with the evolutionary development of anatomical structures.
The wood of many species of Artocarpus is of great economic importance. A. hirsutus yields one of the best types of wood called ' Aini ", A. chaplasha yields ' Chaplash' and other species yield wood used for various purposes.
1 Based on a part of the thesis accepted for the Ph.D. degiee of the Agra Universily. Research contribution No. 42 ftorn the School of Plant Mc~p~clc~. /¢,~1~1Cc]lete,
Meerut. 243

244 M . R . SI-IARMA
The genus thus has considerable taxonomic, paleobotanical, anatomical, morphological and commercial interest about it, and can bear some more anatomical and morphological work. In the present communication, atten- tion is focused on the morphological and anatomical features of vegetative organs.
MATERIAL AND METHODS
The material of vegetative parts of A. heterophyllus Lamk. [syn. A. integrifolius auct., non Linn. f., A. integra (Thunb.) Merr.] and A. lakooeha Roxb., was collected and fixed in F.A.A. from trees cultivated in local gardens and also from Calcutta and Saharanpur respectively. A. ehaplasha Roxb. was collected from Calcutta and Mr. Virendra Prakash Gupta fixed some for me from the gardens of F.R.I., Dehra Dun. Some fixed material of A. altilis (Park) Fosb. (syn. A. ineisa Linn.) was very kindly obtained for me by my colleague Dr. Y. S. Murty from Mr. Veerabhadra Rao of Visakha- patnam. The material of five Malayan species [A. heterophyllus Lamk., A. altilis (Park) Fosb., A. rigidus Blume. (syn. A. eehinatus Roxb.), A. integer (syn. A. champedon Spring.) and A. odoratissimus Blanco.] was very kindly procured through the courtesy of Dr. J. Sinclair of Singapore.
The material was dehydrated, cleared and embedded in the usual manner and serial microtome sections of 10-14 microns were cut. Crystal-violet- erythrosin and safranin-fast-green combinations gave satisfactory results.
OBSERVATIONS
Almost all species of Artocarpus are trees of moderate height of about 50 feet but A. chaplasha may reach 100-150 feet. The main trunk (clean bole) which varies from 6-20 feet in length is thick and stout. Its surface may be rough or smooth, with dark brown or dull grey bark splitting irregu- larly. The main trunk and old branches of A. heterophyllus have irregularly distributed warty excrescences of variable shape and size. Stem of young branches is smooth in A. heterophyllus and A. altilis or rough with wiry long hairs in A. integer or densely hairy in A. rigidus, A. chaplasha and A. odoratissimus, etc.
The leaves are stipulate, petiolate, simple and arranged (spirally) on young branches with different angles of divergence. In A. heterophyllus, A. integer, A. altilis and A. chaplasha, etc., the alternate phyllotaxy is of 2/5 type with angular divergence of 144 °. In A. lakoocha the leaves are alternately arranged on slender and very much elongated branches which have clear dorsiventral symmetry. They are distichous but the angular divergence
Morphological and Anatomical Investigations on A r t o c a r p u s Forst.
(~d; ARTOCARPUS HETEROPHYLLUS
~i .~i ~''~,~j ( ? ~ , . ,.~, ....,-'-,~ ,--,....
245
/
I~k ~ ~,~ I:NF
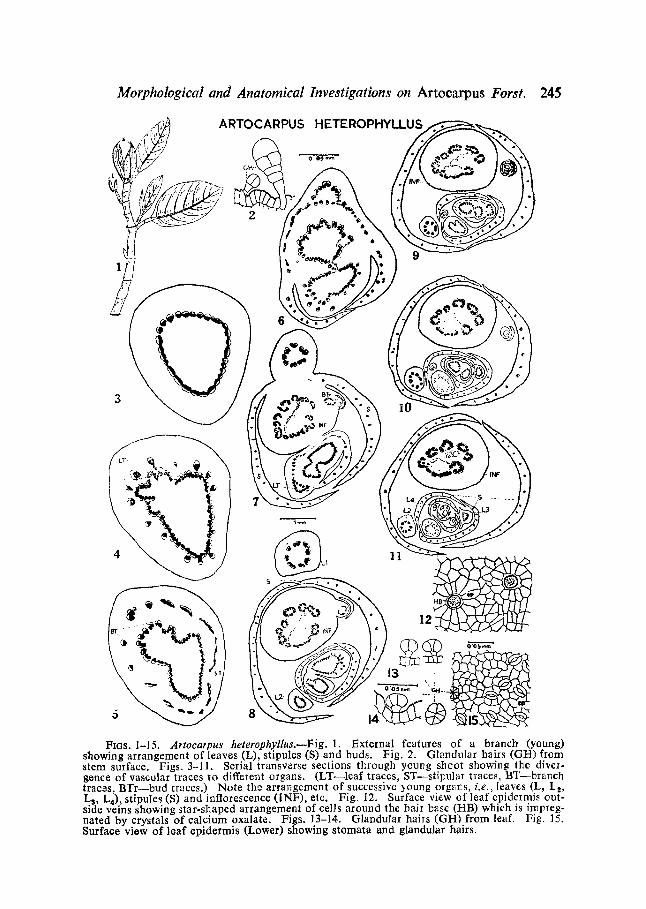
Ftas. 1-15. Artocatpus heterophyllus.wFig. 1. External features of a branch (young) showing arrangement of leaves (L), stipules (S) and buds. Fig. 2. Glandular bails (G/q) from stem surface. Figs. 3-11. Serial transverse sections through young shoot showing the diver- gonce of vascular traces to different organs. (LT--leaf traces, ST--stil~ular traces, BT brailch traczs, BTr bud traces.) Note the arrangement of successive young orgars, i.e., leaves (L, I_2, Ls, L4), stipules (S) and inflorescence (INF), etc. Fig. 12. Surface view of leaf epidelmis out- side veins showing star-shaped arrangement of cells around the hair base (/qB) which is impreg- nated by crystals of calcium oxalate. Figs. 13-14. Glandular hails (GH) from leaf. Fig. 15. Surface view of loaf epidermis (Lox~er) showing stomata and glandular hairs.
c ~ . "G ~
Lt

246 M . R . SHARMA
between first and second, and third and fourth leaves is 120 ° and that between second and third, and fourth and fifth is 240 ° and so on. Therefore, the leaves on the dorsal side of the branch are separated from one another by half the distance of what separates them on the ventral side of the branch (Fig. 51). In A. heterophyllus, the leaves are evergreen, thick and coriaceous. In young plants they may be lobed but in adult ones the margin is entire. In A. altilis the leaves are evergreen in Malaya but deciduous in monsoon countries and southerly pacific isles with mild winter (Corner, 1952). In A. lakoocha and A. chaplasha the leaves are deciduous, coriaceous or sub-eoriaceous and larger in size than those of A. heterophyllus. In A. altilis they may be 30-90 cm., deeply dissected or incised into several pointed lobes. Surprisingly enough the leaves appear to be dimorphic in A. chaplasha. The juvenile leaves are up to 90 cm. long, lobed or pinnatifid while in adult condition they are almost elliptical in shape, coriaceous in texture and are not more than 25 cm. in length (Bor, 1953).
Each leaf has two caducous and amplexicaul stipules which may be large and spathaceous (A. heterophyllus, A. integer and A. altilis) (Figs. 1, 16 and 25) or small (somewhat semi-amplexieaul) enclosing the bud only for a short duration and becoming free lateral soon after (A. lakoocha) (Figs. 51, 55 and 56). They are mostly hairy and pubescent or glabrous (A. heterophyllus). These stipules protect the young vegetative or flowering buds like a bud scale and soon fall off leaving a prominent characteristic, annular cieatrice (scar) on the node (Figs. 1, 16, 25, 32, etc.). The smaller stipules (A. lakoocha) have small line of attachment and correspondingly their cicatrices are small and when mature do not form a ring.
Anatomical features: Internode.--The internode of vegetative shoot is cylindrical and oval or circular i,n cross-section. In flowering shoot it becomes flattened and triangular at a node. The epidermal cells are mostly squarish or radially elongated (A. heterophyllus). The cuticle is thin in A. heterophyllus, A. integer and A. altilis but becomes very thick in A. lakoocha, A. chaplasha, A. odoratissimus and A. rigidus. Some of the epidermal cells are full of latex, more so in A. heterophyllus but many are produced into unicellular and multi- cellular glandular hairs (Figs. 2, 17, 28 and 52). In A. heterophyllus both these types of hairs are small, microscopic and deciduous but in other species only the multicellular glandular hairs are microscopic and deciduous and found on young stem only. They develop from a single epidermal cell by transverse division that cuts off a basal cell. The upper cell divides again transversely and vertically twice and forms a glandular club-shaped head of eight or more cells (Fig. 2). These cells may be full of latex and have thick wall. Some- times a stalk subtends a peltate glandular head of many cells found

Morphological and Anatomical Investigations on Artocarpus Forst. 247
mostly in leaves (Figs. 13 and 14). The unicellular hairs of other species, i.e., A. chaplasha, A. rigidus, A. lakoocha and A. odoratissimus, etc., are macro- scopic, dense, rough and may be straight or hooked with pointed tips (Fig. 48). In A. integer they are also tough and pointed but are wiry and much elongated. Because of the dense growth of hairs, the epidermis in these species is irregular or very oftert becomes obscure (Figs: 48 and 50). The cell-wall of these hairs is thick and lignified and has very fine excrescences of cutin and silica of different magnitudes. Thus the hairs appear to have some systematic value.
In A. rigidus, A. chaplasha and others with dense hairs, there is an earlier origin of cork cambium. The hypodermis is collenchymatous and chlorenchymatous and has a few layers. Some of their cells have latex in them. Cortex and pith regions are made of parenchymatous cells with conspicuous intercellular spaces. The laticifers are abundant in cortex and pith in A. heterophyllus but their frequency diminishes gradually in A. rigidus, A. altilis, A. integer, A. chaplasha, A. lakoocha and A. odoratissimus. Sph~erocrystals are common in the cortex and pith regions of all the species. In A. heterophyllus, there are present elongated fibres scattered in the cortex. There are also present sclereids having pitted thickenings.
The vascular cylinder has 20-30 or more vascular bundles arranged in a ring (Fig. 3). The bundles as in dicotyledons are conjoint, collateral and unequal in size. No endodermis is visible but sclerenchymatous pericycle in the form of bundle cap may occur outside the phloem in A. heterophyllus, A. altilis, A. chaplasha, etc. Phloem parenchyma is rich and has a number of sph~erocrystals. In flowering shoots, the sieve tubes of this region are more prominent than those found in normal vegetative shoots and they are also richer in food contents. A. heterophyllus, the species worked out in detail, has, just before the spring season, regular cambium with 3-5 layers of elongated cells that are rectangular in transverse section.
The Wood.--In A. heterophvllus the wood is somewhat diffused porous, pores being of conspicuous size and are uniformly distributed or may form wavy bands. The primary xylem is made of spiral and annular tracheids, and vessels are 15-30 microns in diameter but secondary xylem has larger vessels and narrower tracheids. The mature vessels are round, 40-90 microns across and have pitted thickenings. In A. lakoocha, however, the xylem vessels vary in size from 20-140 microns in diameter. There are prominent tyloses in the old vessels of A. heterophyllus. The fibres are innumerable and interspersed in the wood. They are 14-21 microns across and 0-35- 1.0 mm. in length. The distribution of wood parenchyma is paratracheal
248 M . R . SHARMA
and aliform or aliform to confluent. Medullary rays are prominent, 2-5 layers in width and 10-30 layers in depth, the cells measuring 14-28 microns across. They broaden considerably towards the cortex. The laticifers may also traverse through the rays.
16
17 o 0 5 ~ - IE
ARTOCARPU5 INTEGER
~~ - '~ I ~' 1 ~
, < s ~ '- ' ~ARTOCARPUS ALTILIS
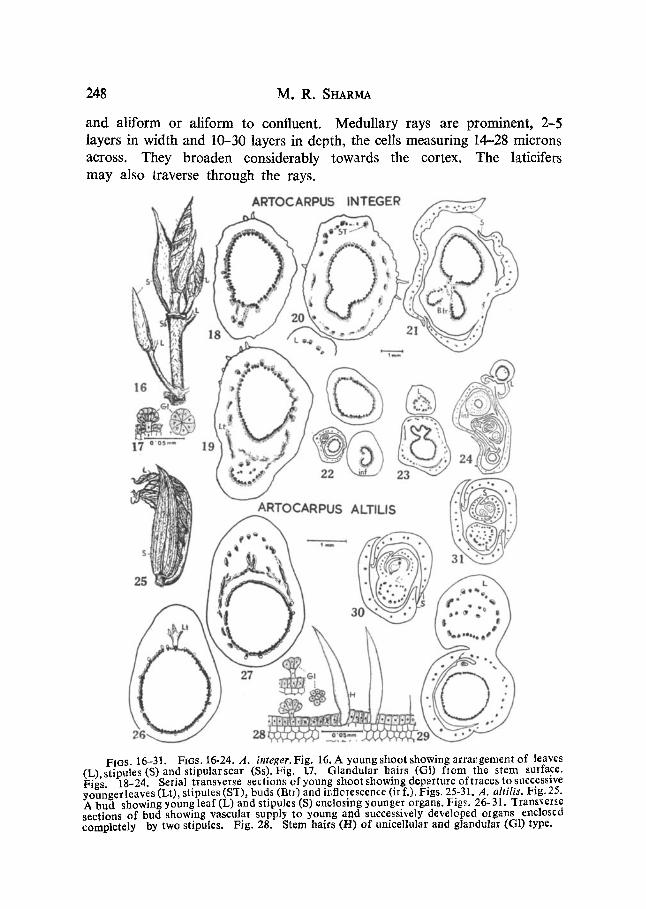
FIGS. 16-31. FIGS. 16-24. A. integer.Fig. 16. A young shoot showing arrangement of leaves (L))stipules (S)and stipularscar (Ss).Fig. 1.7. Glandular hairs (Gl) from lhe stem surface. Figs 18-24. Serial transverse sections of young shoot showing depattme of traces to successive youngerleaves (Lt), stipules (S~[), buds (Bt)) and ir, flo~¢scence (ir f.), Figs. 25-31. ,4, altilia, l~ig. 25. A bud showing young loaf (L) and stipu]es (S) enclosing younger organs. Figs. 26-31. ~Iransve1~e sections of bud showing vascular supply to young and successively developed organs enclosed completely by two stipules. Fig. 28. Stem hairs (H) of unicellular and glandular (G1) type.

Morphological and Anatomical Investigations on Artocarpus Forst. 249
It is interesting to note that the internodes of flowering shoots even in their primary structure are thicker than those of ordinary vegetative shoots in which some secondary growth also has taken place. This difference is apparently due to increase in the size of the individual cortical cells which store large quantities of food material and remain more or less turgid. The phloem region of flowering shoot is also better developed than that of the vegetative shoot.
The Node.--The node o f vegetative shoot is circular in outline while that of flowering shoot is somewhat flattened and triangular in A. heterophyllus and A. integer and this distinction is reflected to a greater or less extent in the form of their vascular cylinder (Figs. 3, 18 and 19). In the sub-nodal region the stelar bundles on the side of next leaf become prominent and begin to send off vascular traces successively (Figs. 4, 18, 26, 33, 38, 45 and 53, etc.). The median trace is the first to pass out and is soon followed by more traces on either side (about 4-6 in A. heterophyllus, 6 in A. integer, 8 i n A. rigMus and A. lakoocha, 10-12 in A. altilis and A. chaplasha and even more up to 20 in A. odoratissimus) in successive order (Figs. 4, 5, 19, 27, 34, 37, 39, 42, 46 and 54, etc.). These are the leaf traces which may branch further, arrange in a depressed ring, or semicircular arch (A. lakoocha) in the petiolar region and traverse further to supply the leaf (Figs. 6-8, 20, 23, 29-31, 35, 36, 40, 41, 47, 49, 55-57, etc.).
Subsequent to the leaf traces there depart many traces (about 12 in A. heterophyllus), all around the stele. These supply the two stipules which are fused to both sides of leaf-base and are in the form of a mantle and cover the nodal and internodal regions completely (Figs. 8-11, 21, 24, 30, 31, 35-37, 43, 44, 56 and 57). Their stipular traces divide into three each and produce many branches traversing parallelly. After receiving the vascular supply, the overlapping margins of the two stipules begin to separate off from leaf- base. They surround the axis and occupy apparently an axillary position with respect to the leaf, except in A. lakoocha, where the stipules surround only the young bud for a short distance and higher up become free and take a lateral position in contrast to the condition in other species (Figs. 55-57). A notable feature about the development of stipules and buds in this species is that they stand almost at right angles to the parent stele and thus cut longi- tudinally or obliquely in transverse section (Fig. 56).
Subsequent to the divergence of leaf and stipular traces, branch traces begin to set off from the bundles of the main stele. They occupy an axillary position and form a lunar-shaped pouch of vascular tissue even before the actual separation of the stipules (Figs. 5, 20, 34, 43, 46 and 54). This small
B5

250 M . R . SHARMA
stele constitutes the vascular cylinder of an ordinary vegetative bud, dormant or active or of a reproductive nature that may develop into a male or female inflorescence (Figs. 7-9, 21-24 and 35-37). In flowering shoots, before entering its destination, the branch-stele generally may split and supply an accessory bud that develops conspicuously on old shoots. In A. integer, A. rigidus and others, the axillant semicircular pouch of a bud in flowering shoots becomes invaginated in the middle region and forms a ~oove which deepens gadually till the pouch gets divided into two compressed and elliptical steles one of which (right side in Text-figs. 21 and 22)supplies the developing young inflorescence while the other one (left side in Text-figs. 35-37) enters the accessory bud. The course of further vascular supply from the main stele to the leaf, stipules and young buds in all species is in the same sequence as described above (Figs. 7-11).
The Leaf.--The leaf of A. heterophyllus has been studied in some detail. A transection of petiole shows a circular outline with a small groove or some- what flat surface towards adaxial side. The epidermis is thick-waUed and has thin cuticle. Some of its cells are produced into very small unicellular or multicellular and glandular hairs. The latter are peltate and have 8-10 cells. In A. rigidus, A. chaplasha and A. odoratissimus the epidermis becomes very irregular because of dense growth of thick-walled hairs. Next to the epidermis there are a few layers of collenchymatous cells, which gradually increase in size inward. Some of these cells contain sphaerocrystals. The parenchymatous cortex and pith has many scattered laticifers, the latter being also common in the medullary rays.
Leaf traces traversing the petiole divide and increase from 15-20 in number. It is interesting to note that some of the petiolar bundles as they enter the leaf-base turn inward from the open end or adaxial side (.4. altilis, A. rigidus, A. chaplasha and A. lakoocha, etc.). They gradually begin to arrange in such a way as to form a complete or incomplete ring inside the parent ring of the petiolar bundles whose orientation remains normal, i.e., endarch (Figs. 29, 36, 49 and 55). In A. odoratissimus, there are only a few medullary bundles and in A. heterophyllus they are only rarely present. There is a distinct sclerenchymatous sheath around the phloem of outer as well as inner bundles. In the mid-vein, as also in some prominent veinlets, this course of double ring of bundles may persist for some distance.
As lamina is approached, the fiat side of the petiole becomes more depressed and may form a deeper groove. The lateral sides also become flattened and gradually the laminar tissue starts developing. The two lateral flattened wings receive their vascular supply successively from the vascular
Morphological and Anatomical Investigations on Artoearpus Forst. 251
bundles situated at the two lateral ends of the mid-vein. Gradually the vascular bundles of the petiole (now mid-vein) become compact and further up the vascular traces diverge out one after another oft either side forming lateral veins for the lamina whose margins are slightly convoluted in young
~s L ARTOCARPUS RIGIDUS / / ~ t ~
14 ''----,~ 35 36
~ A R T O C A R P U S ODORATISSIMUS
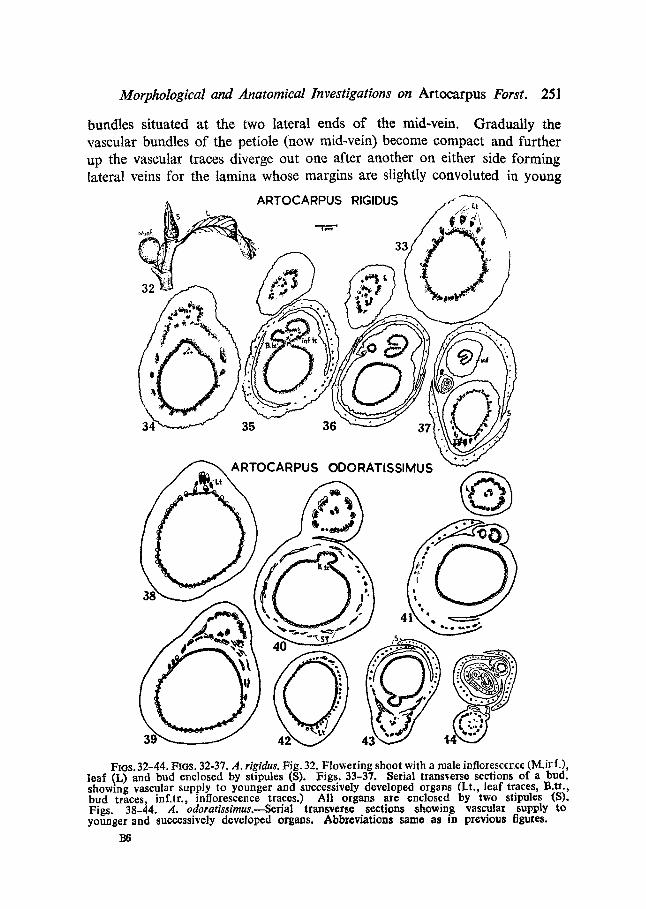
FIOS. 32-44. FIGS. 32-37. A. rigidus. Fig. 32. Flowering shoot with a male infloresccr~ce (M.irf.), leaf (L) and bud enclosed by stipules (S). Figs. 33-37. Serial transverse sections of a bud. showing vascular supply to younger and successively developed organs (Lt., leaf traces, B.ti., bud traces, inf.tr., inforescence traces.) All organs are enclosed by two stipules (S). Figs. 38--44. A. odoratissimus.--Serial transver~© sections showing vascular supply to younger and successively developed organs. Abbr©viation8 same as in previous figures.
]36

252 M . R . SHARMa
condition. In old leaves the groove of vascular ring may disappear and become circular in outline (,4. heterophyllus).
The lower and upper epidermis of the lamina are similar in surface view and vertical sections. In A. heterophyllus the epidermal cells have slightly wavy anticlinal walls and these undulations are present only on the outer side. The epidermal cells occurring on upper and lower sides of the veins and veinlets are very regular, compact and elongeated as observed in surface view (Figs. 12 and 15). Some of these cells are produced into uni- and multi- cellular hairs. The epidermal cells surrounding such hairs are arranged in a beautiful stellate or star-like pattern (Fig. 12). These hairs are broad at the base and pointing at the apex. They are deciduous and visible only in very young leaves. In old leaves they are represented only by the broad base of the hair in level with other epidermal cells. These occasionally show cystolith-like structures which are probably sealed off from outside by thick depositions of cutin or similar material (Fig. 12). The multicellular hairs are of glandular type and are developed by transverse division of the initiating young epidermal cell. The lower cell forms the foot and the upper cell further divides transversely. Of these latter, the lower cell elongates and becomes the stalk while the upper cell after 2, 3 or more vertical divisions produces 6-10 or more celled head (Figs. 13-15). Their cells appear to be peltate in vertical sections and have some chloroplasts. Such hairs remain projecting from the epidermal surface in young leaves but in well-developed ones they usually occur in characteristic pits with their tips in flush with the general leaf surface. There is dense growth of unicellular and persistent hairs in A. integer, A. altilis, A. rigidus, A. odoratissimus, A. chaplasha and A. lakoocha, etc.
The lower epidermis has innumerable stomata, i.e., about 700 per sq. cm. in A. heterophyllus, 500-600 per sq. cm. in A. altilis. They measure (including guard cells) about 25:420 microns in A. heterophyllus and A. altilis, 25 ×16 microns in A. lakoocha. There are two or three layers of palisade cells with abundant chloroplasts. These are followed by a few layers of loose, spongy parenchyma with large number of air-spaces. These tissues are occasionally interrupted by distinct collenchymatous and sclerenchymatous tissue on both sides inner to the vein or veinlets. The vascular bundles of midwein or their veinlets are normal and sph~erocrystals are very common in the phloem parenchyma. A large number of laticifers are present around the veins and also in the cortex and pith region of the midrib.
The Stipule.--It has thick and well-developed cuticle on the outer (dorsal) epidermis that consists of elongated thick-walled rectangular or columnar
Morphological and Anatomical Investigations on A r t o c a r p u s Forst. 253
E
ARTOCARPUS CHAPI.ASHA
' =~' ° F
46
J
~ °
e ~ o e j
L9
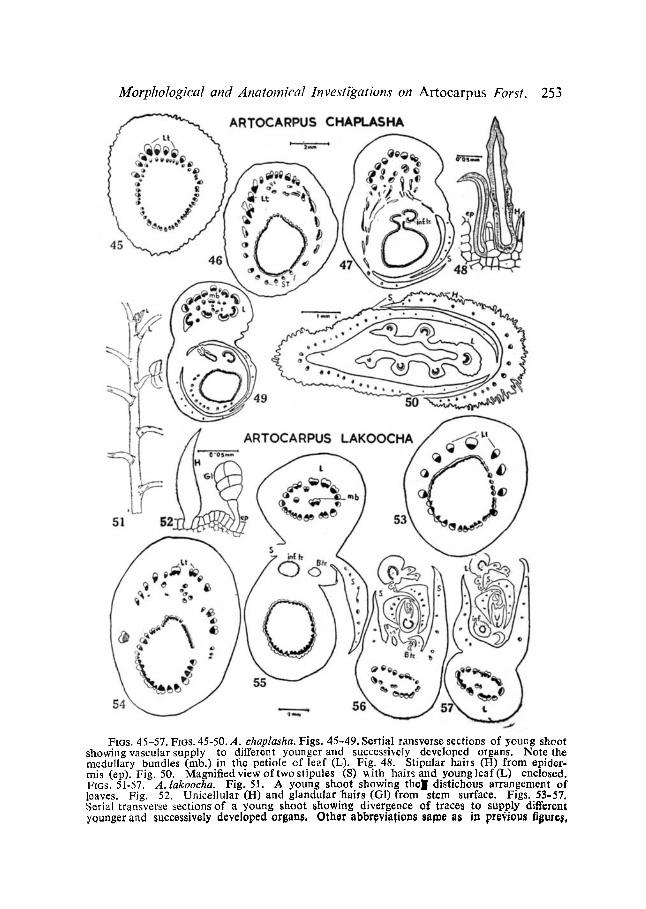
FIGS. 45-57. FIGS. 45-50. A. ehaplasha. Figs. 45-49. Sertial ransverse sections of young shoot showing vascular supply to different younger and successively developed organs. Note the medullary bundles (nab.) in the petiole of leaf (L). Fig. 48. Stipular hails (H) from epider- mis (ep). Fig. 50. Magnified view of two slipules (S) ~ith hairs and youngleaf(L) enclosed. FIGS. 51-57. A. lakoocha. Fig. 51. A young shoot showing tholl distichous arrangement of loaves. Fig. 52. Unicellular (H) and glandular hairs (G1) from stem surface. Figs. 53-57. Serial transverse sectionsof a young shoot showing divergence of traces to supply different younger and successively' developed organs, Other abbr~)vi~fion~ same as in pt0~iou~ figute~,

254 M . R . SHARMA
cells arranged compactly. Epidermis of adaxial (ventral) side is smooth and is devoid of hairs. It is indistinct in A. heterophyllus, but in others it is distinct and regular. The outer (dorsal) epidermis develops the usual two types of hairs, i.e., unicellular (long) and glandular (short), which are sparse and ephemeral in A. heterophyllus but in others they are well developed. The unicellular hairs usually remain persistent due to which the outer epidermis is immensely deformed and becomes irregular, indistinct or even obscure (.4. chaplasha, A. odoratissimus and A. rigidus, etc.) (Figs. 37, 48 and 50). The ground tissue of stipule is made of parenchymatous cells which are larger in the centre and gradually become smaller towards outside. There develop air-spaces as the stipule matures. In A. chaplasha, A. rigidus, A. odoratissimus, large irregular cavities may develop from small air-spaces. There are sph~erocrystals (druses) and laticifers, and 6-8 vascular bundles travel at the base of each stipule which divide and increase in number (12-20) and run parallel converging at the tip.
DISCUSSION AND CONCLUSION
As a rule all species of Artocarpus as revealed by morphological and anatomical studies have more or less the same structural plan. There are, however, some minor differences. For instance, while most of the species have 2/5 type of phyllotaxy, A. lakoocha is distinct in having distichous arrangement with peculiar angular divergence. This character is significant because it forms the basis for the division of Artocarpus into Jaca and Pseudojaca (see Treed, 1847, quoted from Jarrett, 1959 a). Recently, Jarrett (1959 a) has maintained the separation of two subgenera on the basis of this character and some others. He replaces the name Jaca by subgenus Artocarpus while subgenus Pseudojaca is maintained. This change of name appears to be unnecessary. King (1889) while emphasizing floral and fruit characters also divided the genus into two groups without assigning names.
Another character that Jarrett (1959 a, b) has used for distinguishing two subgenera is the number of cells in a glandular hair. This too does not appear to hold good since A. lakoocha (put in subgenus Pseudojaca) like others of the subgenus Artocarpus has hairs with many-celled head on young stem, leaves and stipules. As they are microscopic and ephemeral, they become obscure in the dense growth of unicellular hairs. Jarrett's subgenus Pseudojaca with compact mesophyll and absence of resin cells also does not have a sharp line of demarcation since A. lakoocha (Pseudojaca) may have loose mesophyll in well-developed leaves and has laticifers though sparsely distributed.

Morphological and Anatomical Investigations on Artocarpus Forst. 255
A study of the nodal anatomy reveals considerable uniformity in various species of Artoearpus. The node is uniformly multilacunar with minor variations in the number of traces and gaps. In this respect the genus is similar to Ficus which also has multilacunar nodes, but differs from other members of Urticales that are essentially trilacunar (Sinnott, 1914).
The Stipules.--The two conspicuous stipules present in Artocarpus are characteristic and are well suited to protect the young foliar buds. They resemble closely those of various species of Ficus. Goebel (1905) describes them as axillary stipules and derives them from lateral ones by fusion. He considers their derivation from those of Fieus carica that are free and inserted up along the upper side of the leaf-base. He imagines that if this zone of insertion is raised upon a common base, we may expect an axillary stipule with free upper ends and open on outer side as is the case in Artoearpus. The study of stipules in different species of Artocarpus reveals uniformity in their development and structure. A. lakoocha appears to be slightly different in this respect. Its stipules just after their development become free and lateral and enclose the young bud only for a very short duration. Jarrett (1959 a, b) has exploited this character of stipule (i.e., amplexicaul, leaving annular sear in Artocarpus and non-amplexicaul or lateral, leaving a lateral or intrapetiolar scar in Pseudo]aea) to divide the genus into two subgenera. This character too is not very sharp and demarcated as in very young buds of A. lakoocha. The stipules are similar to those with amplexicaul and therefore it is a matter of degree and duration to cover the young buds and is not a distinct character. King (1889) also mentions that stipules are variable in size and degree but the principle of attachment in all species is similar.
The nature of stipules in Angiosperms had been the subject of much discussion although now it is claimed to have been settled in favour of leaf- base divergence. Sinnott and Bailey (1914) concluded that there is an inti- mate connection between the type of nodal anatomy and the occurrence of stipules, etc., and the latter should be regarded as integral part of the leaf. Mitra and Majumdar (1952) believed that the development of a free base in the form of a sheath or a pair of stipules depends on the course and behaviour of trace bundles through axial component. They discuss the conditions for the derivation of different types of stil~ules and conclude that if in case of a multilacunar node, the laterals branch during their horizontal course towards the median, the stipules formed are either ochrea (Polygonum) or bud scales (species of Fieus and Artocarpus). Majumdar (1955, 1956) concluded that all stipules, stiples and ligules without exception are outgrowths of the leaf-base. He emphasized the importance of leaf-base which in primitive

256 M . R . SHARMA
angiospermous l e~ was wholly incorporated in the axis as the axial component and the lamina with broad base directly inserted on the axis. As the petiole developed, a pair of stipules came into being and the node became trilacunar. Meanwhile, the axial component extended laterally around the apical dome and finally completely enveloped the latter and the node thus became multi- lacunar. Then it gave rise to the sheathing stipules of the type of bud scales as found in species of Ficus and Artocarpus integrifolia (cf. Mitra, 1950). Cross (1937), however, considered the stipular scar in Morus alba as cauliae. He interpreted that the region from which the stipules arise is not the base of the leaf but "might be considered leaf stem transition region ". Parkin (1948) regards the free stipules as cauline and the adnate ones as petiolar. He concludes that a pair of free stipules is an original accompaniment of angiospermous leaf and from it all other kinds of stipular outgrowths can be theoretically derived.
Whatever may be the trends of specialization by which the sheathing stipules of Artocarpus type are derived, it appears to the present author that the discussion on the nature of stipules whether cauline or part of leaf-base is somewhat frivolous. Even in Artocarpus it is very difficult to distinguish between the leaf and the stipular traces. So, the best thing is to consider the stipules as part and parcel of the leaf.
SUMMARY
1. The morphological and anatomical features of seven species of Artocarpus have been studied and described at some length.
2. All species are trees of moderate or lofty habits. They are of uni- form patterns; however, each species has certain peculiarities.
3. While most of the species have 2/5 type of phyllotaxy, A. lakoocha is distinct in having distichous arrangement with peculiar angular divergence.
4. There are unicellular as well as multicellular (glandular) hairs of different magnitudes in stem, leaves and stipules. They have some taxonomic significance.
5. The nodal anatomy reveals considerable uniformity in all species of Artocarpus studied here. The node is uniformly multilacunar, like that of Ficus but differs from those members of Urticales that are essentially trilacunar.
6. The two stipules are characteristic and well suited to protect the young buds. They have been described and their morphological nature has been discussed at some length,

Morphological and Anatomical Investigations on Artocarpus Forst. 257
ACKNOWLEDGEMENTS
I take this opportunity to express my deep sense of gratitude to Prof0 V. Puri, o.sc., F.A.SC., F.N.I., for suggesting the problem, sympathetic guidance and help in various ways. I also thank Dr. Y. S. Murty for sugges- tions and Mr. Radhey Shiam for drawing a few diagrams of external features.
REFERENCES
Bailey, L. H.
Bet, N. L. Corner, E. J. H.
Cross, G. L.
Engler, A. and Prantl, K. Goebel, K.
Hooker, J. D.
Hutchinson, J.
Jarrett, F. M.
King, G.
Lawrence, 0 . H. M. Majumdar, G. P.
Mitra, G. C.
- - - - - - and Majumdar, G . P . . .
Parkin, J. ..
Record, S. J. and Hess, R. W . . .
Rendle, A.B. • •
.. The Standard Cyclopedia of Horticulture, New Ycik, 1950.1.
.. Manual of Indian Forest Botany, Oxford, 1953. .. Wayside Trees of Malaya, Sing~pole. 1952, 1, 193-99.
.. "The origin and development of Ike foliage leaves and stipules of Motus alba," Bull. Tcttey Bet. Club, 1937, 64, 145-63.
.. Die Nutiirlichen Pflanzenfamilien.,Leipzig, 1888, 18, 80-93. .. Organography of Plants, English Edition by I. B. Balfour,
Oxford. 1905, 2.
.. The Flora of British lndia, London, 1885, 5.
.. The Families of Flowering Plants, Dicotyledons, London. 1926, 1.
.. "Studies in Artocarpus and allied genera--I," Your. Arnold Arb., 1959a, 40, 1-29.
.. "Studies in Artoeatpu~ and allied genera--Ill . 1rid., 1959b, 40, 113-55 and 298-368.
.. "The species of Attoeatpus indigenous to Brili~h India," Ann. Bet. Gard. Calcutta, 1889, 2, 1-16.
.. Taxonomy of Vascular Plants, New York, 1951.
.. "The complete foliage leaf," Prec. Ind. Acad. ScL, 1955, 42 B, 65-72.
.. "Stipules, ligules and leaf-sheath," Ibid., 1956, 43B, 9-22.
.. "Developmental studies. A comparative account of the origin, development and morphology of t~e stipules of Attocarpus integrifolia L., Ficus religiosa L. and 17. elaatica, Roxb.," Prec. Nat. lust. Sci. lndia, 1950, 16, 15%68.
"The leaf-base and the internode--theit true morphology," The Palceobotanist, 1952, 1, 351-67.
"The stipules considered phylogenetically," The North- Western Naturalist, March-December 1948, pp. 63-82.
"American woods of the family Morace~e," Trop. Woods, 1940, 61, 11-54.
Timbers of New World, New Haven, 1943.
The Classification of Flowering Plants," Dicotyledons, Cambridge. 1938, 2.

258 M . R . SrtagMh
Ridley, I-I. N.
Se,~ard, A. C7. Sinnott, E. W.
and Bailey, I. W.
Tippo, O.
*Tr~cul, A.
* Not soon in original.
. . The Flora o f the Malay Peninsula, London, 1924, 3.
. . Plant Life through the Ages, Cambridge, 1941.
. . "Investigations on tl:e phylogeny of the Angiospelms. I. The anatomy of the node as an aid in classification of Angiosperms," Amer. Jour. Bet., 1914, 1, 303-22.
. . "Investigations on the phylogeny of Angiosperms. 1II. Nodal anatomy and the morphology of stipules," Ibid., 1914, 1~ 441-53.
• . "Comparative anatomy of the Morace~0 and their presumed allies," Bet. Gaz., 1938, 100, 1-99.
. . "M6moire sur la famille des Artocarpees." Ann. Sei. Nat. Bat., 1847, 8(3), 38-157.
696--62. Printed at The Bangalore Press, Bangalore City, by T. K. Balakrishnan, Superintendent. and Published by B. S. Venkatachar, Editor, " Proceedings of the Indian Academy of Sciences ",
Bangalore





























