Embed Size (px)
Citation preview

Kidney International, Vol. 59 (2001), pp. 2182–2191
ION CHANNELS – MEMBRANE TRANSPORT – INTEGRATIVE PHYSIOLOGY
Hypertonic activation of the renal betaine/GABA transporteris microtubule dependent
JARED C. BASHAM, ALEXANDRA CHABRERIE, and STEPHEN A. KEMPSON
Department of Cellular and Integrative Physiology, Indiana University School of Medicine, Indianapolis, Indiana, USA
Hypertonic activation of the renal betaine/GABA transporter structural polarity because typically their minus ends re-is microtubule dependent. main bound in a perinuclear centrosome near the center
Background. Epithelial cells in the renal inner medulla accu- of the cell, while their plus ends are free and extend towardmulate osmolytes such as betaine to maintain normal cell volumethe cell periphery. The total surface area of the microtu-during prolonged extracellular hypertonic stress. Betaine accu-bules within a cell may be equivalent to the surface areamulation is the result of activation of transcription of the BGT1
transporter gene followed by increased betaine transport. of the plasma membrane, and it provides an enormousMethods. We studied the possible role of microtubules in area for protein–protein interactions [1]. Microtubule-
this adaptive mechanism using renal cells in culture. associated proteins serve both to stabilize microtubulesResults. In cultured renal cell lines [Madin-Darby canineagainst disassembly and to mediate their interaction withkidney (MDCK) and mouse inner medullary collecting ductother cell components. Some proteins are microtubule(mIMCD-3)], up-regulation of BGT1 activity was maximal after
24 to 30 hours in growth medium made hypertonic (510 mOsm/kg) motors that use the energy of adenosine 59-triphosphateby the addition of sucrose or NaCl. Up-regulation was reversed (ATP) hydrolysis to move along microtubules. Specificwithin 24 to 36 hours after returning cells to isotonic medium. examples of motor molecules are the kinesins and cyto-Both cycloheximide (20 mmol/L) and nocodazole (20 mmol/L)
plasmic dyneins, which move toward the plus and minusblocked the hypertonic up-regulation of BGT1. Nocodazoleends, respectively, and provide a system for directionalwas partially effective even when added 16 to 20 hours after
the switch to hypertonic medium. Recovery from nocodazole flow of organelles, vesicles, and other cargo proteinsaction was rapid, and there was full activation of BGT1 trans- within the cytoplasm [1–3].port within three to six hours after nocodazole removal, sug- In a wide variety of cells, an intact microtubule net-gesting rapid trafficking to the cell surface once microtubules
work is essential for insertion of transport proteins andrepolymerized. Hypertonic activation of BGT1 transport wasreceptors into the plasma membrane in response to spe-detected in an isolated membrane fraction and was blocked
by cycloheximide but not by nocodazole. Confocal microscopy cific stimuli. The cAMP-mediated translocation of theconfirmed the increased abundance of BGT1 proteins in the bile acid transporter in liver cells [4], the carbachol stimu-plasma membrane of hypertonic cells and showed that BGT1 lation of GLUT1 activity in glioma cells [5], the hypertonicremained intracellular during nocodazole treatment.
up-regulation of system A amino acid transport in smoothConclusions. Hypertonic activation of BGT1 in renal cellsmuscle cells [6], and the insulin-stimulated translocationrequires de novo protein synthesis and microtubule-dependent
trafficking of additional transporters to the cell surface. The of the oligopeptide transporter in intestinal cells [7] areapparent resistance of membrane BGT1 to nocodazole block- specific examples. Similarly, in renal cells, the disruptionade is likely due to the presence in the membrane fraction of of microtubules impaired the acute up-regulation of typean increased intracellular pool of active BGT1 transporters. IIa Na/Pi cotransporter abundance [8] and the apical
targeting of the A1 adenosine receptor [9] and severaltransport proteins [10].
In nonmotile tissue cells, microtubules fill the cyto- Cells of the mammalian nephron that are located inplasm and usually occupy the space between the nucleus the renal medulla face prolonged osmotic stress causedand the plasma membrane. Microtubules have an inherent by the hypertonic environment required for the concen-
trating function of the kidney. They survive and maintaina normal cell volume by accumulating osmolytes suchKey words: cyloheximide, colchicine, nocodazole, BGT1 transport,
MDCK cells, mIMCD-3 cells, plasma membrane, confocal imaging. as sorbitol, myo-inositol, and betaine [11, 12]. Accumu-lation of betaine occurs by increased uptake of extracel-Received for publication June 16, 2000lular betaine, a process mediated by the Na1- and Cl2-and in revised form December 7, 2000
Accepted for publication December 26, 2000 dependent betaine/GABA transporter (BGT1) locatedprimarily in the basolateral plasma membrane [11–13]. 2001 by the International Society of Nephrology
2182

Basham et al: Microtubules in hypertonic response of BGT1 2183
The BGT1 clones from MDCK cells and human kidney MDCK and mIMCD-3 cells. The uptake values werecorrected for surface binding and trapping of solute byencode a protein of 614 amino acids that transports both
betaine and GABA and belongs to the gene family of subtraction of blank values.Madin-Darby canine kidney membrane fractions wereneurotransmitter transporters [14, 15]. The presence of
sorting signals and basolateral retention motifs have re- prepared by using nitrogen cavitation to disrupt cells fol-lowed by differential centrifugation to recover mem-cently been identified in the cytosolic C-terminal tail of
BGT1 [16, 17]. The 59 flanking region of the BGT1 gene branes, as described previously for MDCK and endo-thelial cells [22, 25]. Cells were collected by scrapingcontains two tonicity-responsive enhancers (TonE1 and
TonE2) that mediate increased transcription of BGT1 in in Tris-buffered saline (pH 7.4) that was either isotonicor hypertonic, as appropriate. After two saline washes,response to prolonged hypertonic stress in both Madin-
Darby canine kidney (MDCK) cells and in mouse kidney the cells were quickly suspended in 300 mmol/L manni-tol, 5 mmol/L egtazic acid (EGTA), 0.2 mmol/L pheny-in vivo [18–20]. This leads to a 15- to 20-fold increase in
the abundance of BGT1-specific mRNA in MDCK cells methylsulfonyl fluoride, and 12 mmol/L Tris-HCl (pH 7.1),which was diluted 1:1 with water followed immediately[21]. Since this step is likely followed by increased synthe-
sis of BGT1 transporter proteins, we studied the possible by cavitation. The final membrane pellet was resuspendedin 300 mmol/L mannitol, 12 mmol/L Tris-HCl (pH 7.4)role of microtubules in the up-regulation of BGT1 trans-
port in renal cells. at a protein concentration of 5 mg/mL. BGT1 transportactivity in isolated membranes was determined by therapid filtration procedure [22, 25]. The medium for up-
METHODStake contained 100 mmol/L NaCl, 100 mmol/L mannitol,
Canine MDCK cells (CCL-34) and mouse inner med- 10 mmol/L Tris (pH 7.4 with HEPES), 0.05 mmol/Lullary collecting duct (mIMCD-3) cells (CRL-2123; Amer- [3H]GABA, and 0.05 mg membrane protein. When char-ican Type Culture Collection, Rockville, MD, USA) acterizing the ion dependency of GABA uptake, eitherwere used between passages 15 and 30. They were plated KCl or sodium isethionate (100 mmol/L) was includedin 35 mm plastic dishes (2.5 3 106 cells/dish) for cell instead of NaCl.uptake studies and in 75 cm2 flasks (4 3 106 cells/flask) Tubulin was extracted from cell monolayers usingfor membrane transport studies. The growth medium microtubule-stabilizing buffer, as described elsewherewas a 1:1 mixture of Dulbecco’s modified Eagle’s me- [26]. Cells in a T-75 flask were washed twice in phos-dium (DMEM) and Ham’s F-12K containing 10% bovine phate-buffered saline (PBS) and incubated for 15 min-calf serum, 10 mmol/L HEPES, 25 mmol/L NaHCO3 utes at 378C in 5 mL buffer containing 0.1% Triton(pH 7.4), and penicillin G (100 U/mL), as in previous X-100, 2.0 mol/L glycerol, 1 mmol/L MgSO4, 1 mmol/Lstudies [22, 23]. Cultures were maintained at 378C in ethylenediaminetetraacetic acid (EDTA), 0.1 mol/Lan atmosphere of 5% CO2 in air and were used when Pipes, pH 6.75. The buffer and cell residue were collectedconfluent. The osmolality of the growth medium was 285 by scraping, and the suspension was centrifuged atmOsm/kg as determined by freezing point depression in 48,000 3 g for 10 minutes to separate the Triton-solublean osmometer. Hypertonic growth medium was made by fraction from the Triton-insoluble residue.addition of either sucrose (MDCK) or NaCl (mIMCD-3) Protein samples for sodium dodecyl sulfate-polyacryl-to achieve a final osmolality of 510 mOsm/kg. Colchicine, amide gel electrophoresis (SDS-PAGE) were heated fornocodazole, and cycloheximide were added to the re- three minutes at 958C in (final concentrations) 2% SDS,quired concentration from 5 mmol/L stock solutions in 10% glycerol, 100 mmol/L dithiothreitol, 1 mmol/L EDTA,dimethyl sulfoxide stored at 2208C. De novo protein 100 mmol/L Tris, pH 6.8. Equal amounts of differentsynthesis in cell monolayers was assessed as [3H]leucine protein samples were loaded (20 mg/lane) and separatedincorporation that resisted extraction by 5% trichloro- on a 10% polyacrylamide gel at 10 mA for two hoursacetic acid, as described previously [24]. and electrotransferred to nitrocellulose paper at 80 mA
Since the BGT1 transporter accepts both betaine and overnight using the BioRad minigel system, as in previ-g-aminobutyric acid (GABA) as substrates [14], BGT1 ous studies [27]. After blocking with saline containingtransport activity in cell monolayers was determined as 10 mmol/L Tris (pH 7.6), 0.05% Tween-20, and 5% Car-Na1- and Cl2-dependent [3H]GABA uptake as described nation milk powder, Western blots were performed with apreviously [22]. The final concentration of GABA in the mouse monoclonal antibody against rat b-tubulin (Sigma,uptake solution was 0.01 mmol/L. Control cells main- St. Louis, MO, USA) at a 1:5000 dilution. Binding oftained in isotonic growth medium were processed in par- the primary antibody was visualized using goat anti-allel with the cells in hypertonic medium, using isotonic mouse IgG (1:5000) conjugated to horseradish peroxi-or hypertonic uptake and wash solutions as appropriate dase as secondary antibody and enhanced chemilumines-[22, 23]. All uptakes were determined at a ten-minute in- cence (ECL; Amersham, Arlington Heights, IL, USA)
detection reagents.cubation time, which lies within the linear range for both

Basham et al: Microtubules in hypertonic response of BGT12184
Immunofluorescence studies were performed on MDCKcell monolayers grown on glass coverslips. Hypertonicgrowth medium (containing either 20 mmol/L nocoda-zole or vehicle) was added 48 hours after plating whenthe cells were still subconfluent. In these studies, the os-molality of the hypertonic medium was reduced to450 mOsm/kg [22] to minimize cell detachment duringsubsequent fixation and immunostaining. After 24 hoursof hypertonicity, the cells were washed twice in ice-coldTris-buffered saline (TBS; 150 mmol/L NaCl, 50 mmol/LTris-HCl, pH 7.6) containing sucrose to maintain theosmolality at 450 mOsm/kg. The washed cells were fixedby immersion for 10 minutes at room temperature in100% methanol previously chilled to 2208C [16]. Cellsmaintained in isotonic media throughout were processedin the same way. A 30-minute preincubation in 2% gela-tin in TBS was followed by a two-hour incubation withaffinity-purified anti-dog BGT1 antibody [16, 17] diluted1:100 in 1% gelatin/TBS. The primary antibody wasdetected by a 60-minute incubation in affinity-purifiedFITC-conjugated goat anti-rabbit IgG (Jackson Immuno-Research, West Grove, PA, USA), at a 1:100 dilution in1% gelatin/TBS. All incubations were at 378C. In controlincubations, either the primary or secondary antibody wasomitted. Cells were mounted in mowiol solution (9%wt/wt mowiol 4-88, 24% wt/wt glycerol, 90 mmol/L Tris-HCl, pH 8.5). Fluorescence images were acquired with Fig. 1. Hypertonic up-regulation and recovery of BGT1 transport in
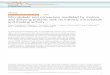
mIMCD-3 cell monolayers. Na1/Cl2-dependent [3H]GABA uptake wasa Zeiss LSM 510 confocal microscope using a water im-measured at various times, and data are mean values of two experiments.mersion 340 lens with 1.2 numerical aperture and 220 (A) Cells grown in isotonic medium for three days were switched toeither fresh isotonic (s; Iso, 285 mOsm) or hypertonic (d; Hyp,mm working distance. The excitation wavelength was 488510 mOsm) at time zero. (B) Following exposure to hypertonic mediumnm, and emission was collected at 505 to 520 nm. Thefor 30 hours, hypertonic cells were returned to isotonic conditions at
optical section thickness was 1 mm, and the instrument time zero. Iso cells were maintained in isotonic medium throughout.gain and offset were the same for all samples.
Na,K-ATPase, alkaline phosphatase, and protein weredetermined by colorimetric assays as previously [22].
of BGT1 in mIMCD-3 cells was similar to that previouslya-Mannosidase II activity was determined at 378C usingreported for MDCK cells [21, 22].5 mmol/L p-nitrophenyl a-D-mannopyranoside as sub-
Hypertonic up-regulation of BGT1 in mIMCD-3 cellsstrate in 50 mmol/L MES buffer (pH 6.0) containingwas sensitive to blockade by cycloheximide, as expected.
0.5 mmol/L EDTA to inhibit mannosidase I [28, 29]. AllCells were pretreated for two hours with 20 mmol/L cyclo-
experiments were analyzed in triplicate and repeated at heximide prior to switching to hypertonic medium. Theleast three times. Control cells and hypertonic cells were hypertonic medium also contained cycloheximide but atalways compared within the same experiment, and re- a lower concentration. The combination of 20 mmol/L pre-sults were analyzed with the Student t-test. Differences treatment and a 1 mmol/L sustaining dose for 24 hourswere considered significant at P , 0.05. produced complete blockade (Fig. 2). This action of
cycloheximide correlated directly with its effectiveness inblocking [3H]-leucine incorporation into acid-insoluble
RESULTS protein, a measure of de novo protein synthesis. A 35%Hypertonic up-regulation of BGT1 transport in reduction in leucine incorporation was accomplished
mIMCD-3 cells reached a peak after 30 hours and was by the 20 mmol/L pretreatment alone, 60% by the 20/reversed after returning the cells to isotonic medium. 0.2 mmol/L combination, and 80% by the 20/1.0 mmol/LRecovery was complete within 36 hours (Fig. 1). Al- combination.
Inclusion of colchicine in the hypertonic mediumthough slower to up-regulate and recover, this response

Basham et al: Microtubules in hypertonic response of BGT1 2185
Fig. 3. Dose-dependent inhibition of hypertonic up-regulation ofBGT1 transport by colchicine. mIMCD-3 cells were incubated for
Fig. 2. Blockade of hypertonic up-regulation of BGT1 transport in 24 hours in hypertonic medium containing colchicine at concentrationsmIMCD-3 cells by cycloheximide (CHX). Cell monolayers were incu- in the range of 0 to 30 mmol/L. Data are expressed as percentage ofbated for two hours in isotonic medium containing 20 mmol/L CHX and the Na1/Cl2-dependent GABA uptake determined in the absence ofthen transferred to isotonic (I) or hypertonic (H) medium containing 0, colchicine because of variations in absolute values. Each point is mean 60.2, or 1.0 mmol/L CHX for 24 hours. Data for Na1/Cl2-dependent SE from three experiments.GABA uptake are mean 6 SE from three experiments. *Significantlydifferent (P , 0.05) compared with uptake in isotonic controls.
produced a dose-dependent inhibition of BGT1 up-regulation. Maximum inhibition (50%) was achieved at10 mmol/L, and higher doses (30 mmol/L) had no addi-tional effect (Fig. 3). Nocodazole treatment had a similareffect but was much more effective. Doses in the range20 to 30 mmol/L completely prevented up-regulation(Fig. 4) and maintained BGT1 activity at the level ofisotonic controls (data not shown). The concentrationsof colchicine and nocodazole that blocked BGT1 up-regulation were within the range shown previously to beeffective in disrupting microtubules in cultured renalcells [26, 30]. For nocodazole, this was checked directlyusing MDCK cells. BGT1 activity was 40 6 10 pmolGABA/mg/10 min in isotonic control MDCK cells com-pared with 288 6 50 pmol/mg/10 min (P , 0.01) after24 hours of hypertonicity. Inclusion of 20 mmol/L noco-dazole in the hypertonic medium reduced BGT1 activityto 97 6 31 pmol/mg/10 min (mean 6 SE, N 5 3), alevel not significantly different from isotonic controls.Western blots of Triton extracts of MDCK cells revealedthat the Triton-insoluble tubulin fraction, representingintact microtubules [26], was eliminated almost com-pletely within two hours after the addition of 20 mmol/L
Fig. 4. Dose-dependent inhibition of hypertonic up-regulation ofnocodazole. This effect persisted as long as nocodazoleBGT1 transport by nocodazole. mIMCD-3 cells were incubated for
was present for up to 24 hours (Fig. 5). A corresponding 24 hours in hypertonic medium containing nocodazole in the range of0 to 50 mmol/L. Each data point is mean 6 SE from three experiments.increase in the Triton-soluble tubulin fractions was not

Basham et al: Microtubules in hypertonic response of BGT12186
Fig. 5. Western blot showing nocodazole action on b-tubulin distribu-tion in MDCK cells. The cell monolayers were incubated in isotonicmedium in the absence (C) or presence (NZ) of 20 mmol/L nocodazolefor either 2 or 24 hours prior to lysis in Triton buffer, as described inthe Methods section. Rat brain supernatant was run on the gel as apositive control.
Fig. 7. Hypertonic up-regulation of BGT1 transport recovered rapidlyafter nocodazole (NZ) blockade was removed. MDCK cell monolayerswere incubated in isotonic (Iso) or hypertonic (Hyp) growth mediumin the absence or presence of 20 mmol/L NZ. After 24 hours, all cellswere rapidly washed and transferred to NZ-free hypertonic medium(time zero). Up-regulation of Na1/Cl2-dependent GABA transport wasmonitored in each group for a further 24 hours. Data are mean 6 SEfrom three experiments. *Uptake was significantly different (P , 0.05)from 24-hour uptake in the same group.
tent with rapid nocodazole action to depolymerize micro-tubules and, presumably, disrupt trafficking of newly syn-thesized BGT1 proteins to the cell surface. Delayedaddition of nocodazole is effective because hypertonicup-regulation of BGT1 in MDCK cells is not maximal
Fig. 6. Delayed addition of nocodazole (NZ; 20 mmol/L) produceduntil 24 hours [13, 21, 22].partial blockade of hypertonic up-regulation. MDCK cells were
switched to hypertonic (Hyp) medium at time zero, and NZ was added Since microtubules repolymerize rapidly (within 10either at the same time or later. The delay in NZ addition ranged from minutes) after the removal of nocodazole [31], we next0 to 20 hours. Na1/Cl2-dependent GABA uptake is expressed as the
determined whether cells previously treated with noco-percentage of uptake in cells incubated for 24 hours in Hyp mediumin the absence of NZ. Data are mean 6 SE from three experiments. dazole during exposure to hypertonic stress would ex-
hibit a more rapid hypertonic up-regulation of BGT1once the drug was removed. The rationale was that syn-thesis of additional BGT1 transporters would alreadydetected, presumably because the size of this pool ishave occurred, but they would remain trapped in an intra-relatively large. These findings indicate that there wascellular pool in the presence of nocodazole. This provedrapid depolymerization of microtubules on addition ofto be the case (Fig. 7). Within four to six hours afternocodazole and, no repolymerization occurred duringtransfer to nocodazole-free hypertonic medium, BGT1long-term (24 hours) nocodazole treatment.transport activity had increased to a level not signifi-In the preceding studies, nocodazole was applied tocantly different from the level at 24 hours. In contrastthe cells at the time of switching to hypertonic medium.and as shown previously [13, 21, 22], cells not previouslyWhen nocodazole (20 mmol/L) addition was delayed forexposed to hypertonic stress did not exhibit the full adap-several hours after applying hypertonic stress, there wastive response until at least 18 hours after switching tostill significant impairment of BGT1 up-regulation. Forhypertonic medium. Previous treatment with nocodazoleexample, only 60 to 65% of the full up-regulation occurredunder isotonic conditions did not alter the subsequentwhen nocodazole was added as late as 16 to 20 hours
after the start of hypertonic stress (Fig. 6). This is consis- full adaptive response to hypertonicity (Fig. 7), confirm-

Basham et al: Microtubules in hypertonic response of BGT1 2187
Fig. 8. Ion dependence of [3H]GABA uptake in membrane fractionsfrom MDCK cells. Ise, isethionate. Data are mean 6 SE of triplicate Fig. 9. Cycloheximide (CHX) blocked hypertonic up-regulation ofdeterminations with a single membrane preparation. BGT1 transport in MDCK membranes. Prior to fractionation, cells
were incubated for 24 hours in either isotonic or hypertonic growthmedium, and CHX was applied as described in the text. Data aremean 6 SE from three experiments. NS, no significant difference (P .0.05) in GABA uptake determined in NaCl uptake medium compareding the reversible action of nocodazole on microtubular with uptake when NaCl was replaced by KCl.
organization.Because of closely similar responses of MDCK and
mIMCD-3 cells, the remaining studies were confined toMDCK cells. Plasma membrane markers, both basolat- 24 hours (20 mmol/L pretreatment plus 1 mmol/L sus-
taining dose) completely blocked hypertonic up-regula-eral and apical, are well-established in the MDCK cell[22], and the subsequent studies focused on the adaptive tion of BGT1 at the membrane level (Fig. 9), as in intact
cells (Fig. 2). There was no Na1-dependent GABA up-response of BGT1 at the membrane level. The purposewas to determine whether BGT1 activity in the absence take in membranes isolated from cells that had been
subjected to hypertonic stress for 24 hours in the pres-of possible intracellular regulatory factors correlatedwith the responses observed in intact cells. The mem- ence of cycloheximide. The overall uptake was similar
to that in cycloheximide-treated controls that were main-brane fraction from MDCK cells contained both apicaland basolateral plasma membranes, based on a 5.0- and tained in isotonic growth medium (Fig. 9).
The addition of nocodazole to MDCK cells at the same4.1-fold enrichment of alkaline phosphatase and Na,K-ATPase, respectively, in the final membrane fraction time as the growth medium was made hypertonic (no
delay) was completely ineffective in blocking the up-compared with the initial cell homogenate. In contrast,there was only a 1.1-fold increase in the activity of a-man- regulation of BGT1 activity in the membrane fraction
(Fig. 10), in contrast to nocodazole action on BGT1nosidase II, a Golgi marker [32, 33], in the membranefraction. It was important to recover both apical and transport determined in whole cells (Fig. 6).
The apparent discrepancy between whole cell data andbasolateral membranes since BGT1 is exclusively baso-lateral under isotonic conditions, but some apical activity membrane data was investigated further by studying the
time course of BGT1 down-regulation when cells wereis induced during hypertonic stress [13]. BGT1 activity inthis membrane fraction was detected as Na1/Cl2-depen- returned to isotonic medium. BGT1 activity in MDCK
cells was down-regulated to the level in isotonic controlsdent GABA uptake (Fig. 8). Both Na1 and Cl2 ionswere required for optimal GABA uptake and only the when the cells were allowed to recover for 24 hours in
isotonic medium (Fig. 11A). In contrast, BGT1 activityNa1- and Cl2-dependent component was up-regulatedby hypertonic stress, confirming the characteristic prop- in the membrane fraction was not down-regulated after
24-hour recovery in isotonic medium. Up to 48 hours oferties of the BGT1 transporter [18]. GABA uptake de-termined in the absence of either Na1 or Cl2 was not recovery was required for complete down-regulation of
BGT1 in the membrane fraction (Fig. 11B).different between isotonic and hypertonic groups (Fig. 8).Treatment of MDCK cells with cycloheximide for Immunofluorescence studies were undertaken in

Basham et al: Microtubules in hypertonic response of BGT12188
Fig. 10. Action of nocodazole (NZ) on up-regulation of BGT1 trans-port in MDCK membranes. Prior to fractionation, cells were incubatedfor 24 hours in either isotonic (Iso) or hypertonic (Hyp) growth mediumin the presence or absence of 20 mmol/L NZ. Data for Na1/Cl2-depen-dent GABA uptake are mean 6 SE from three experiments. *Uptakewas significantly different (P , 0.02) compared with uptake in the Isocontrol group.
MDCK cells to determine why nocodazole appeared tohave no effect on BGT1 up-regulation at the membranelevel. An affinity-purified antibody to dog BGT1 was kindlyprovided by Dr. Grazia Pietrini (University of Milan, Mi-
Fig. 11. Rate of down-regulation of BGT1 transport in MDCK cellslan, Italy). This antibody was used previously to demon- after 24 hours of hypertonic stress. The isotonic (Iso) controls werestrate basolateral membrane targeting of BGT1 in trans- maintained in isotonic growth medium throughout. (A) At time zero,
cells previously incubated in hypertonic (Hyp) medium for 24 hoursfected MDCK cells [16, 17]. In wild-type MDCK cellswere switched to isotonic medium, and Na1/Cl2-dependent GABAunder isotonic conditions, no fluorescence was observed uptake was determined at various times ranging from 0 to 24 hours.
to be associated with the plasma membrane (Fig. 12A) Data are mean 6 SE from three experiments. *Uptake was significantlydifferent (P , 0.05) from the uptake in the same group after 24 hoursas expected, based on the low level of BGT1 transportof recovery. (B) BGT1 transport in membrane fractions obtained afterin isotonic MDCK cells [13]. Although cells tended to 24 and 48 hours of isotonic recovery following 24 hours of hypertonicity.
round up after the addition of nocodazole to the isotonic Data are mean 6 SE from three experiments. *Uptake in NaCl uptakemedium was significantly different (P , 0.02) from uptake when NaClmedium, no plasma membrane fluorescence was detectedwas replaced by KCl.
(Fig. 12C). In marked contrast, there was pronouncedfluorescence associated with the plasma membrane fol-lowing hypertonic stress for 24 hours (Fig. 12B). Opticalsections in the x-z plane (data not shown) revealed that
was most noticeable in the perinuclear region (Fig. 12D),the fluorescence was confined to the basolateral plasmawhich likely includes the endoplasmic reticulum [17].membrane. No membrane fluorescence was detectedSome of the intracellular fluorescence may also be duewhen either the primary or secondary antibody was omit-to BGT1 present in the Golgi, since this structure isted, strongly suggesting that the membrane-associateddispersed into multiple fragments throughout the cell byfluorescence was due to specific interactions between10 to 20 mmol/L nocodazole [34, 35]. These data confirmantibodies and the BGT1 protein. Hypertonic cells treatedthat the abundance of BGT1 protein in the basolateralwith nocodazole showed no plasma membrane fluores-
cence. Instead, the fluorescence was intracellular and plasma membrane is up-regulated in response to hyper-

Basham et al: Microtubules in hypertonic response of BGT1 2189
Fig. 12. Effect of nocodazole (NZ) on BGT1protein distribution in MDCK cells exposedto either isotonic growth medium (A and C)or hypertonic medium (B and D) for 24 hours.Under isotonic conditions, there was no plasmamembrane fluorescence (A, arrow), and thiswas not changed by the addition of 20 mmol/LNZ (C, arrow). Following hypertonic stress,there was marked plasma membrane fluores-cence (B, arrow), but this was absent afterNZ treatment. Instead, the fluorescence wasintracellular, principally in the perinuclear re-gion (D, arrow). Original magnification 31500(A, B, and D) and 31000 (C).
tonic stress. However, in the absence of intact microtu- but key events may be the activation of the p38 kinasepathway and the binding of a transcription factor (TonEbules, the newly synthesized BGT1 proteins remain in-binding protein) to TonE [39]. This factor becomes moretracellular and are not inserted in the basolateral plasmaabundant and shifts into the nucleus in response to hy-membrane. The intracellular BGT1 proteins are likely topertonicity [40].be recovered in the membrane fraction from nocodazole-
Studies on the aldose reductase gene in PAP-HT25treated cells, since the preparative procedure does notcells showed that the increase in aldose reductase mRNAdiscriminate between plasma membranes and intracellu-during hypertonic stress was due primarily to increasedlar membranes. This may help explain why nocodazoletranscription rather than increased mRNA stability [11].appears to have no effect on up-regulation of BGT1 atIncreased transcription also played a key role in thethe membrane level (Fig. 10).increase in BGT1 mRNA level in osmotically stressedMDCK cells [21]. The events that follow transcription
DISCUSSION and accumulation of BGT1 mRNA are largely unknown.Both MDCK cells [11] and mIMCD-3 cells [36] accu- Blockade of the hypertonic up-regulation of BGT1 activ-
mulate betaine in response to prolonged hypertonic stress. ity by cycloheximide (Fig. 2) indicates that de novo pro-In MDCK cells, the increase in intracellular betaine is tein synthesis is required. A similar blockade of BGT1due to increased transport into the cell through the activ- up-regulation by actinomycin D and cycloheximide hasity of the BGT1 transporter [11–13]. The hypertonic up- been reported in chondrocytes [41]. These data stronglyregulation and recovery of BGT1 activity in mIMCD-3 suggest that the Vmax change in BGT1 transport in MDCKcells (Fig. 1) suggest that increased transport plays an cells during hypertonic stress [21] is due to an increaseimportant role in betaine accumulation in these cells in the number of BGT1 transport proteins. The blockadealso. Exposure of renal cells to hypertonic solutions con- by colchicine and nocodazole (Figs. 3 and 4) is consistenttaining NaCl rapidly activates multiple families of mito- with a role for intact microtubules in trafficking newlygen-activated protein kinases (MAPK), stimulates tran- synthesized proteins to the plasma membrane.scription of genes responsible for accumulating organic The transport studies with isolated MDCK mem-osmolytes, and enhances expression of molecular chap- branes indicate that BGT1 up-regulation involves an in-erones [37]. Neither the MAPK cascade [38] nor protein trinsic change at the membrane level and is not duetyrosine phosphorylation [39] appears to be involved simply to altered ion gradients in intact cells. Only the
Na1- and Cl2-coupled component of GABA uptake wasdirectly in transcriptional activation of the BGT1 gene,

Basham et al: Microtubules in hypertonic response of BGT12190
altered, indicating that there were no nonspecific changes APPENDIXin membrane permeability during hypertonic stress. The Abbreviations used in this article are: BGT1, betaine/GABA trans-
porter; cAMP, cyclic adenosine monophosphate; GABA, g-aminobu-most likely explanation for the up-regulation is an in-tyric acid; MAPK, mitogen-actived protein kinase; MDCK, Madin-crease in the number of BGT1 transport proteins sinceDarby canine kidney; mTAP, mictotubule-associated proteins; TonE,
the membrane adaptation was blocked by pretreating tonicity-responsive enhancers.cells with cycloheximide (Fig. 9). The membrane adapta-tion was not affected by nocodazole (Fig. 10). This is REFERENCESprobably an artifact of the membrane preparation, which
1. Gundersen GG, Cook TA: Microtubules and signal transduction.is enriched in plasma membranes, but also contains mem- Curr Opin Cell Biol 11:81–94, 1999
2. Mironov AA, Luini A, Buccione R: Constitutive transport be-branes from intracellular organelles such as Golgi. Iftween the trans-Golgi network and the plasma membrane ac-BGT1 transporters accumulate intracellularly during ex-cording to the maturation model: A hypothesis. FEBS Lett 440:99–
posure to nocodazole, they would be recovered in the 102, 19983. Brown D, Breton S: Sorting proteins to their target membranes.membrane fraction and, if functional, would contribute
Kidney Int 57:816–824, 2000to the overall transport activity of the fraction. Indirect 4. Dranoff JA, McClure M, Burgstahler AD, et al: Short-termevidence supporting intracellular accumulation of BGT1 regulation of bile acid uptake by microfilament-dependent translo-
cation of rat ntcp to the plasma membrane. Hepatology 30:223–229,transporters is provided by the rapid up-regulation of1999BGT1 activity in three to six hours following removal 5. Singh SP, Gao Y, Singh LD, et al: Role of microtubules in glucose
of nocodazole from hypertonic medium (Fig. 7), likely uptake by C6 glioma cells. Pharmacol Toxicol 83:83–89, 19986. Chen J-G, Hinesley R, Kempson SA: Dual action of colchicine onbecause of insertion of preformed BGT1 transporter
hypertonic activation of system A amino acid transport in vascularproteins in the plasma membrane. These conclusions smooth muscle cells. Life Sci 61:29–37, 1997were confirmed by the immunofluorescence and confocal 7. Thamotharan M, Bawani SZ, Zhou X, Adibi SA: Hormonal
regulation of oligopeptide transporter pept-1 in a human intestinalmicroscopy data, which showed a marked increase incell line. Am J Physiol 276:C821–C826, 1999plasma membrane BGT1 abundance in response to hy- 8. Lotscher M, Kaissling B, Biber J, et al: Role of microtubules in
pertonicity (Fig. 12). When nocodazole was present dur- the rapid regulation of renal phosphate transport in response toacute alterations in dietary phosphate content. J Clin Invest 99:ing hypertonic stress, however, newly synthesized BGT11302–1312, 1997proteins accumulated intracellularly and were not in- 9. Saunders C, Limbird LE: Disruption of microtubules reveals two
serted in the basolateral plasma membrane. independent apical targeting mechanisms for G-protein-coupledreceptors in polarized renal epithelial cells. J Biol Chem 272:19035–Like the nocodazole data, studies on recovery from hy-19045, 1997pertonic stress also showed a divergence between whole 10. Breton D, Brown D: Cold-induced microtubule disruption andrelocalization of membrane proteins in kidney epithelial cells. J Amcell data showing complete recovery within 24 hoursSoc Nephrol 9:155–166, 1998(Fig. 11A) and membrane data showing that recovery
11. Garcia-Perez A: Molecular regulation of organic osmolytes inrequired 48 hours (Fig. 11B). This suggests that recovery renal cells, in Molecular Nephrology, edited by Schlondorff D,
Bonventre JV, New York, Marcel Dekker Inc., 1995, pp 451–458occurs in part via endocytic internalization of BGT1 pro-12. Burg MB, Kwon ED, Kultz D: Regulation of gene expressionteins, which then remain functional for several hours
by hypertonicity. Annu Rev Physiol 59:437–455, 1997while in an intracellular membrane pool. This is in 13. Yamauchi A, Kwon HM, Uchida S, et al: Myo-inositol and betaine
transporters regulated by tonicity are basolateral in MDCK cells.marked contrast to the NaPi type IIa transporter, whichAm J Physiol 261:F197–F202, 1991is degraded rapidly (within 1 h) after internalization [42]. 14. Yamauchi A, Uchida S, Kwon HM, et al: Cloning of a Na1 and
In summary, hypertonic up-regulation of BGT1 activ- Cl–-dependent betaine transporter that is regulated by hypertonic-ity. J Biol Chem 267:649–652, 1992ity in renal cells involves an intrinsic change at the cell
15. Rasola A, Galietta LJ, Barone V, et al: Molecular cloning andsurface that is mediated by de novo protein synthesis functional characterization of a GABA/betaine cotransporter fromand microtubule-dependent trafficking of additional human kidney. FEBS Lett 373:229–233, 1995
16. Perego C, Bulbarelli A, Longhi R, et al: Sorting of two polytopictransporters to the plasma membrane.proteins, the g-aminobutyric acid and betaine transporters, in po-larized epithelial cells. J Biol Chem 272:6584–6592, 1997
17. Perego C, Vanoni C, Villa A, et al: PDZ–mediated interactionsACKNOWLEDGMENTSretain the epithelial GABA transporter on the basolateral surface
This work was supported by a grant from the National Kidney of polarized epithelial cells. EMBO J 18:2384–2393, 1999Foundation of Indiana. We are indebted to Dr. G. Pietrini (University 18. Takenaka M, Preston AS, Kwon HM, Handler JS: The tonicity-of Milan, Milan, Italy) for generously providing antibodies to dog sensitive element that mediates increased transcription of the beta-BGT1. We thank Drs. M. Montrose and S. Chu for assistance with ine transporter gene in response to hypertonic stress. J Biol Chemconfocal microscopy, Dr. F. Pavalko for tubulin antibodies, Dr. J. 269:29379–29381, 1994McAteer for cell culture facilities, and Dr. J.-G. Chen for helpful 19. Miyakawa H, Rim JS, Handler JS, Kwon HM: Identification ofdiscussions. This study was published in part in abstract form in J Am the second tonicity-responsive enhancer for the betaine transporterSoc Nephrol 11:42A, 2000. (BGT1) gene. Biochim Biophys Acta 1446:359–364, 1999
20. Kaneko T, Takenaka M, Okabe M, et al: Osmolarity in renalReprint requests to Stephen Kempson, Ph.D., Medical Sciences 374, medulla of transgenic mice regulates transcription via 59-flanking
Indiana University School of Medicine, 635 Barnhill Drive, Indianapo- region of canine BGT1 gene. Am J Physiol 272:F610–F616, 1997lis, Indiana 46202-5120, USA. 21. Uchida S, Yamauchi A, Preston AS, et al: Medium tonicity regu-
lates expression of the Na1 and Cl–-dependent betaine transporterE-mail: [email protected]

Basham et al: Microtubules in hypertonic response of BGT1 2191
in Madin-Darby canine kidney cells by increasing transcription of 33. Naim HY, Joberty G, Alfalah M, Jacob R: Temporal associationof the N- and O-linked glycosylation events and their implication inthe transporter gene. J Clin Invest 91:1604–1607, 1993the polarized sorting of intestinal brush border sucrase-isomaltase,22. Kempson SA: Differential activation of system A and betaine/aminopeptidase N, and dipeptidyl peptidase IV. J Biol ChemGABA transport in MDCK cell membranes by hypertonic stress.274:17961–17967, 1999Biochim Biophys Acta 1372:117–123, 1998
34. Shima DT, Cabrera-Poch N, Pepperkok R, Warren G: An or-23. Chen J-G, Coe M, McAteer JA, Kempson SA: Hypertonic activa-dered inheritance strategy for the Golgi apparatus: Visualizationtion and recovery of system A amino acid transport in renal MDCKof mitotic disassembly reveals a role for the mitotic spindle. J Cellcells. Am J Physiol 270:F419–F424, 1996Biol 141:955–966, 199824. Abraham MI, McAteer JA, Kempson SA: Insulin stimulates phos-
35. Storrie B, White J, Rottger S, et al: Recycling of golgi-residentphate transport in opossum kidney epithelial cells. Am J Physiolglycosyltransferases through the ER reveals a novel pathway and258:F1592–F1598, 1990provides an explanation for nocodazole-induced Golgi scattering.25. Kempson SA, Hoshaw MJ, Hinesley RS, McAteer JA: Hyperos-J Cell Biol 143:1505–1521, 1998motic stress up-regulates amino acid transport in vascular endothe-
36. Rauchman MI, Nigam SK, Delpire E, Gullans SR: An osmoti-lial cells. Kidney Int 52:1332–1339, 1997cally tolerant inner medullary collecting duct cell line from an26. Pasdar M, Li Z, Krzeminski KA: Desmosome assembly in MDCKSV40 transgenic mouse. Am J Physiol 265:F416–F424, 1993epithelial cells does not require the presence of functional microtu-
37. Santos BC, Chevaile A, Hebert M-J, et al: A combination ofbules. Cell Motil Cytoskeleton 23:201–212, 1992NaCl and urea enhances survival of IMCD cells to hyperosmolality.27. Kempson SA, Lotscher M, Kaissling B, et al: Parathyroid hor-Am J Physiol 274:F1167–F1173, 1998mone action on phosphate transporter mRNA and protein in rat
38. Kwon HM, Itoh T, Rim JS, Handler JS: The MAP kinase cascaderenal proximal tubules. Am J Physiol 268:F784–F791, 1995is not essential for transcriptional activation of osmolyte trans-28. Rabouille C, Kuntz DA, Lockyer A, et al: The Drosophila GMIIporter genes. Biochem Biophys Res Commun 213:975–979, 1995gene encodes a Golgi a-mannosidase II. J Cell Sci 112:3319–3330, 39. Atta MG, Dahl SC, Kwon HM, Handler JS: Tyrosine kinase1999 inhibitors and immunosuppressants perturb the myo-inositol but
29. Gonzalez DS, Karaveg K, Vandersall-Nairn AS, et al: Identifi- not the betaine cotransporter in isotonic and hypertonic MDCKcation, expression and characterization of a cDNA encoding cells. Kidney Int 55:956–962, 1999human endoplasmic reticulum mannosidase I. J Biol Chem 274: 40. Woo SK, Dahl SC, Handler JS, Kwon HM: Bidirectional regula-21375–21386, 1999 tion of tonicity-responsive enhancer binding protein in response
30. Morris RG, Tousson A, Benos DJ, Schafer JA: Microtubule to changes in tonicity. Am J Physiol 278:F1006–F1012, 2000disruption inhibits AVT-stimulated Cl2 secretion but not Na1 re- 41. de Angelis E, Petronini PG, Borghetti P, et al: Induction ofabsorption in A6 cells. Am J Physiol 274:F300–F314, 1998 betaine-g-aminobutyric acid transport activity in porcine chondro-
31. Manandhar G, Sutovsky P, Joshi HC, et al: Centrosome reduction cytes exposed to hypertonicity. J Physiol (Lond) 518:187–194, 1999during mouse spermiogenesis. Dev Biol 15:424–434, 1998 42. Lotscher M, Scarpetta Y, Levi M, et al: Rapid downregulation of
32. Herscovics A: Importance of glycosidases in mammalian biosyn- rat renal Na/Pi cotransporter in response to parathyroid hormoneinvolves microtubule rearrangement. J Clin Invest 104:483–494, 1999thesis. Biochim Biophys Acta 1473:96–107, 1999