Embed Size (px)
Citation preview

THE Jommnr, OB BIOLOQICAL CIIEMIBTRY Vol. 243, No. 1. Iawe of January 10, pp. 160-166.1968
Printed in U.S.A.
Human Alkaline Phosphatase
IMMUNOCHEMICAL IDENTIFICATION OF ORGAN-SPECIFIC ISOENZYMES
(Received for publication, July 31, 1967)
HOWARD H. SUMMAN,* PARKER A. SMALL, JR.,$ AND ERNEST COTLOVE
From the Clinical Pathology Department, Clinical Center, National Institutes of Health, and the Laboratory of Neurochemistry, National Institute of Mental Health, Bethesda, Maryland SO014
SUMMARY
The antigenic distinctiveness of individual organ-specific isoenzymes of human alkaline phosphatase was shown by immunochemical means. A two-step method involving two separate antigen-antibody reactions was utilized to identify and measure the isoenzymes. The first reaction consisted of reaction of the isoenzyme with its antiserum; the second reaction consisted of precipitating the enzyme-antibody complex with anti-y-globulin. The second reaction was nec- essary because the enzyme was neither inhibited nor pre- cipitated by specific antibody. By this method, individual isoenzymes could be identified from mixtures of isoenzymes.
The results clearly show that antibody to liver phosphatase and antibody to placental phosphatase were specific for their respective homologous antigens, and did not react with alkaline phosphatase from bone, neutrophils, kidney, or intestine. Thus, there are at least three antigenic types of alkaline phosphatases: one derived from liver, one from placenta, and one or more from other organs.
There is evidence that human alkaline phosphatase exists in multiple forms and that the forms found in different organs pos- sess distinct properties by which they may be recognized. Iden- tification has been based upon immunochemical (l-7) and elec- trophoretic (8-14) properties, thermal stability (10, 13, 15), relative activity with a variety of inhibitors (16-l@, and kinetic analyses (10, 19, 20). These studies, however, have not pro- vided conclusive criteria for identifying the isoenzymes. Inter- pretations based on these data, consequently, have not always been in agreement on the existence or definition of specific organ forms (9, 21). Some of the difficulties may have been due to insufficiently purified enzyme preparations and to the limitations of using physical properties or differential substrate activity to define molecular differences.
* To whom requests for reprints should be addressed. $ Present address, Department of Microbiology, University of
Florida Medical School, Gainesville, Florida.
In the present study, an immunochemical method capable of specific and quantitative measurement of individual molecular forms was used to examine alkaline phosphatases isolated from individual organs.’ Evidence derived from these immunochemi- cal studies is presented for the existence of antigenically distinct, organ-specific alkaline phosphatase isoenzymes.
MATERIALS AND METHODS
The chromatographic materials used were DEAE-cellulose (Bio-Rad), Sephadex G-200 (Pharmacia), and hydroxylapatite (Hypatite, Lot 6281, Clarkson Chemical Company). Acrylam- ide gel reagents were obtained from Canalco, and p-nitrophenyl phosphate was a Sigma product.
Enzyme Assay-Enzyme activity was assayed at 37” with p-nitrophenyl phosphate as the substrate, and the release of p-nitrophenol was measured at 410 rnp (23). Enzyme activity is expressed as the change in absorbance per unit of time at that wave length. Specific activities are expressed as micromoles of p-nitrophenol released per min per mg of protein with a measured molar absorbance of 1.84 X 104 for &nitrophenol in 0.02 N NaOH. Protein was measured as absorbance at 280 rnp, with the use of an extinction coefficient of 1.0 for a 1.0 mg per ml aqueous solu- tion read in a l-cm light path.
Polyacrylamide Gel Disc ElectrophoresisElectrophoresis was conducted at room temperature. A 5% upper gel and a 7.5% lower gel were used with 0.005 M Tris-HCl-0.04 M glycine buffer, pH 8.3, in both upper and lower electrode compartments; 2 ma per disc were applied (24). Enzyme protein was detected by incubating the gel cylinder in p-nitrophenyl phosphate for 10 to 15 set at room temperature and observing the development of yellow color, indicating release of p-nitrophenol in the region of enzyme activity. The boundaries of enzyme activity were marked with two razor cuts immediately after color development. The gel was then counterstained with Amid0 black to delineate the relationship between enzyme activity and protein bands.
Purijicatim of Enzymes-Alkaline phosphatases were extracted and purified from the following: liver, p!acenta, kidney, bone, intestine, and neutrophils. Although the initial preparation of each tissue was different, subsequent extraction with n-butyl
1 A preliminary report on this subject has been presented (22).
160
by guest on May 21, 2018
http://ww
w.jbc.org/
Dow
nloaded from
Issue of January 10, 1968 H. H. Xussman, P. A. Small, Jr., and E. Cotlove 161
alcohol and initial processing during purification were similar for all.
Liver and Kidneys-Livers and kidneys were obtained within 24 hours after death from cadavers of patients who had died of trauma or of myocardial infarction. Nonparenchymal tissues were removed, and excess blood was washed out.
Placenta-Placentas were initially processed by N. Kontaxis (Stanford Research Institute, Menlo Park, California), who kindly provided an acetone powder of the extracted enzyme. Prior to extraction, the placentas had been washed and then per- fused with cold 0.25 M sucrose and heparin to remove blood.
Bone-Human bone and cartilage were collected at autopsy from the costochondral junction, patella, tibia1 and femoral epiphyses at the knee, and the pelvis of adults and children who had died of trauma or heart disease. The bones were stored at -2O”, and 300 g of tissue were processed.
NeutrophilsNeutrophils were obtained from the b&y coat of multiple units of centrifuged venous blood from normal sub- jects. Two hundred milliliters of buffy coat were used for en- zyme preparation. The erythrocytes present were lysed by osmotic shock (25), and the neutrophils were washed with 0.01 M Tris-HCl buffer, pH 7.4.
Intestine-A small intestine was obtained at autopsy from a 20-year-old white man who had died of trauma 8 hours previ- ously. The luminal surface was washed with running tap water, and the mucosal cells were scraped off and frozen. Two days later the cells were thawed and washed three times with 20 vol- umes of cold 0.14 M NaCl solution. Microsomes were prepared by differential centrifugation after the cells had been homoge- nized in 0.01 M Tris-HCl, pH 7.4, with a motor-driven Teflon pestle with a ground glass homogenizer. The initial, 600 x g supernatant was centrifuged at 20,000 x g for 20 min to remove mitochondria. The supernatant from this step was centrifuged at 105,000 x g for 2 hours to obtain the microsomal pellet.
Enzyme Extraction and Pur$ication-Alkaline phosphatase was extracted from each tissue by a modification of Morton’s procedure (26). Each tissue and the intestinal microsomes were homogenized in 0.25 M sucrose and 20% (v/v) n-butyl alcohol at 4”. The resulting homogenate was incubated at 37” for 1 hour, kept overnight at 4”, and centrifuged at 1000 x g for 2 hours. The enzyme activity was recovered in the aqueous phase. The sediment was re-extracted with butyl alcohol and homoge- nized as before, and the aqueous phase from this was pooled with that from initial extraction.
Bone enzyme was extracted by a modification of the method of Volkin (27). Fragments were allowed to autolyze in Tris-HCl buffer at room temperature for 2 weeks prior to homogenization with butyl alcohol.
Fractional acetone precipitation of the tissue extracts was con- ducted at 4”. Enzyme was precipitated in the 30 to 50% acetone fraction. In the case of placental phosphatase, an acetone powder was prepared from a single 60% precipitation. The pre- cipitates were then suspended in 0.01 M Tris-HCl buffer, pH 7.4, and dialyzed free of butyl alcohol and acetone.
Chromatography and Gel Filtration-The liver, placental, and kidney enzymes were further purified following acetone precipita- tion by chromatography and gel filtration; neutrophil, bone, and intestinal phosphatase preparations were not. All chromato- graphic procedures were conducted at 4”.
DEAE-cellulose was equilibrated with 0.01 M Tris-HCl, pH 7.4, Liver and placental enzymes were purified on columns (5.5 x
I.750 -
1.500 -
1.250-
1.000 -
.750 -
,500 -
,250 -
A\
20 - I I I“
l6- ,’ ‘1 , \ /
I \ / - .06
400 1200 2000 2600 3600 4400
go-
75 -
60-
45-
00 160 240 320
‘i;:: 160 480 800 1120 1440 1760
EFFLUENT VOLUMEtml)
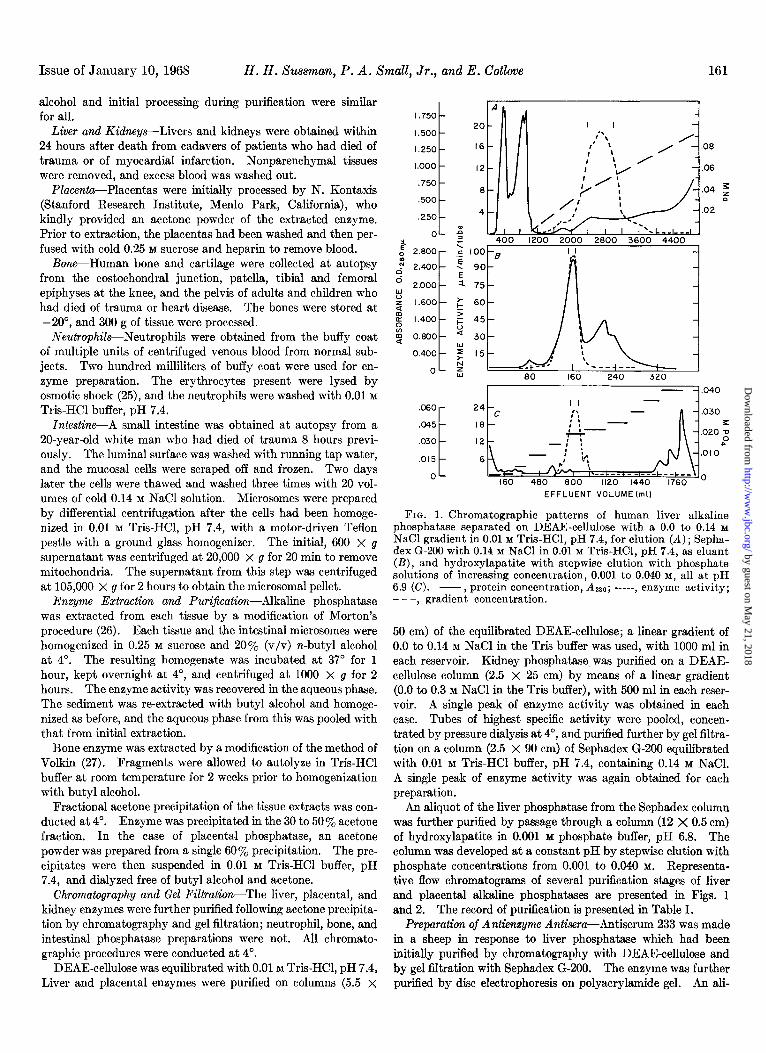
FIG. 1. Chromatogranhic patterns of human liver alkaline phosphatase separated &r DEAE-cellulose with a 0.0 to 0.14 M NaCl gradient in 0.01 M Tris-HCl. nH 7.4. for elution IA) : Senha- dex Gl200 with 0.14 M NaCl in O.diM Tris-HCI, pH 7.4, is e&ant (B), and hydroxylapatite with stepwise elution with phosphate solutions of increasing concentration, 0.001 to 0.040 M, all at pH 6.9 (C). -, protein concentration, 4280; -----, enzyme activity; - - -, gradient concentration.
50 cm) of the equilibrated DEAE-cellulose; a linear gradient of 0.0 to 0.14 M NaCl in the Tris buffer was used, with 1000 ml in each reservoir. Kidney phosphatase was purified on a DEAE- cellulose column (2.5 X 25 cm) by means of a linear gradient (0.0 to 0.3 M NaCl in the Tris buffer), with 500 ml in each reser- voir. A single peak of enzyme activity was obtained in each case. Tubes of highest specific activity were pooled, concen- trated by pressure dialysis at 4’, and purified further by gel filtra- tion on a column (2.5 X 90 cm) of Sephadex G-200 equilibrated with 0.01 M Tris-HCI buffer, pH 7.4, containing 0.14 M NaCl. A single peak of enzyme activity was again obtained for each preparation.
An aliquot of the liver phosphatase from the Sephadex column was further purified by passage through a column (12 X 0.5 cm) of hydroxylapatite in 0.001 M phosphate buffer, pH 6.8. The column was developed at a constant pH by stepwise elution with phosphate concentrations from 0.001 to 0.040 M. Representa- tive flow chromatograms of several purification stages of liver and placental alkaline phosphatases are presented in Figs. 1 and 2. The record of purification is presented in Table 1.
Preparation of Antienzyme Antisera-Antiserum 233 was made in a sheep in response to liver phosphatase which had been initially purified by chromatography with DEAE-cellulose and by gel filtration with Sephadex G-200. The enzyme was further purified by disc electrophoresis on polyacrylamide gel. An ali-
by guest on May 21, 2018
http://ww
w.jbc.org/
Dow
nloaded from

162 Alkaline Phosphatase Isoenzymes Vol. 243, No. 1
3.500 r 1~
96
72
24
1600 3600 5400 7200
100 250 310
EFFLUENT VOLUME (ml)
FIG. 2. Chromatographic patterns of human placental alkaline phosphatase separated on DEAE-cellulose with a 0.0 to 0.09 M NaCl gradient in 0.01 M Tris-HCl, pH 7.4, used for elution (A) and Sephadex G-200 with 0.14 M NaCl in 0.01 M Tris-HCl, pH 7.4, as eluent (B). -, protein concentration, &so; -----, enzyme ac- tivity; - - -, gradient concentration.
TABLE I Puri$cation of phosphatase from human liver and placenta
Sequential purification procedure Total en- yyme units
Specific activity
Human liver enzyme Acetone precipitate,
30-5oyc. . . . . . . . . . . . . . DEAE-cellulose chro-
matography . . . . . . . . Sephadex G-200 gel fil-
tration . . . . . . . . . . . . . Hydroxylapatite chro-
matography . . . . . . . Human placenta enzyme
Acetone precipitate, 60% . . . . . . . . . . . . . . .
DEAE-cellulose chro- matography . . . . . .
Sephadex G-200 gel fil- tration . . . . . . . . . . . . .
II ‘otal pro. Relative t ein, A280 purity
.-
%z moles/min moles/min jrn)
7,000 455 0.065 1.0
77 430 5.5 85
26 183 7.0 108
- -a 14.7 226
8,800 25,920 3.0 1.0
240 13,200 55 18
27 8,856 328 109
a An aliquot rather than the total sample was purified by this procedure.
quot containing 120 /Lg of enzyme protein (specific activity, 7 pmoles per min per mg) was placed on each analytical gel cylm- der, and after electrophoresis the site of enzyme activity in the gel cylinder was determined as previously described. The en- zyme-containing bands were immediately cut out from the gels and stored at -20” until used. The remainder of the cylinder was stained for protein to ascertain if the removed section in- cluded any non-enzyme protein (Fig. 3). Only sections free of non-enzyme protein were used for immunization. Twenty gel
sections were used for primary and booster immunizations. The sections were placed in a tuberculin syringe and fragmented by slowly squeezing them through a No. 25 needle. The fragments were suspended in 0.5 ml of complete Freund’s adjuvant prior to inoculation. Approximately 0.2 mg of protein was used for each immunization.
Antiserum 234 was made in sheep in response to liver phospha- tase which had been purified with DEAE-cellulose and Sephadex G-200 (specific activity, 7 pmoles per min per mg). Both pri- mary and booster immunizations contained 2.0 mg of protein in 0.5 ml of NaCl emuslified with 0.5 ml of complete Freund’s adjuvant.
Antiserum 333 to the purified placental phosphatase (specific activity, 328 pmoles per min per mg) was made in sheep. This material had a single, coinciding enzyme protein band on disc electrophoresis. The primary immunization contained 0.8 mg of protein in 0.5 ml of NaCl emulsified with 0.5 ml of complete Freund’s adjuvant. An initial booster containing 0.3 mg of protein emulsified with the same amount of Freund’s adjuvant did not initiate an adequate antienzyme response. A satisfac- tory response was obtained with a second booster containing 2.7 mg of protein.
The antigen emulsions were injected intramuscularly at multi- ple sites. Antibody activity was evaluated in 6 weeks by tech- niques to be described. The animals were bled between 10 and 16 days after the booster injections, vhen antibody was present in a satisfactory titer.
Preparation of Burro Anti-sheep y-Globulin--y-Globulin was prepared from sheep serum by sodium sulfate precipitation (28). Three milligrams of the sheep y-globulin in 0.5 ml of NaCl were
FIG. 3. Disc electrophoretic method of obtaining isoenzymes for immunization. Aliquots of liver phosphatase purified through the Sephadex G-200 gel filtration step were separated by poly- acrylamide gel disc electrophoresis. Top, cylinder stained to show protein bands; middle, cylinder incubated with p-nitrophenyl phosphate, and the resulting band of enzyme activity demarcated with razor cuts prior to protein staining; bottom, cylinder stained for prot,ein after removal of the enzyme band for immunization.
by guest on May 21, 2018
http://ww
w.jbc.org/
Dow
nloaded from
Issue of January 10, 1968 H. H. Sussman, P. A. Small, Jr., and E. Cotlove 163
TABLE II
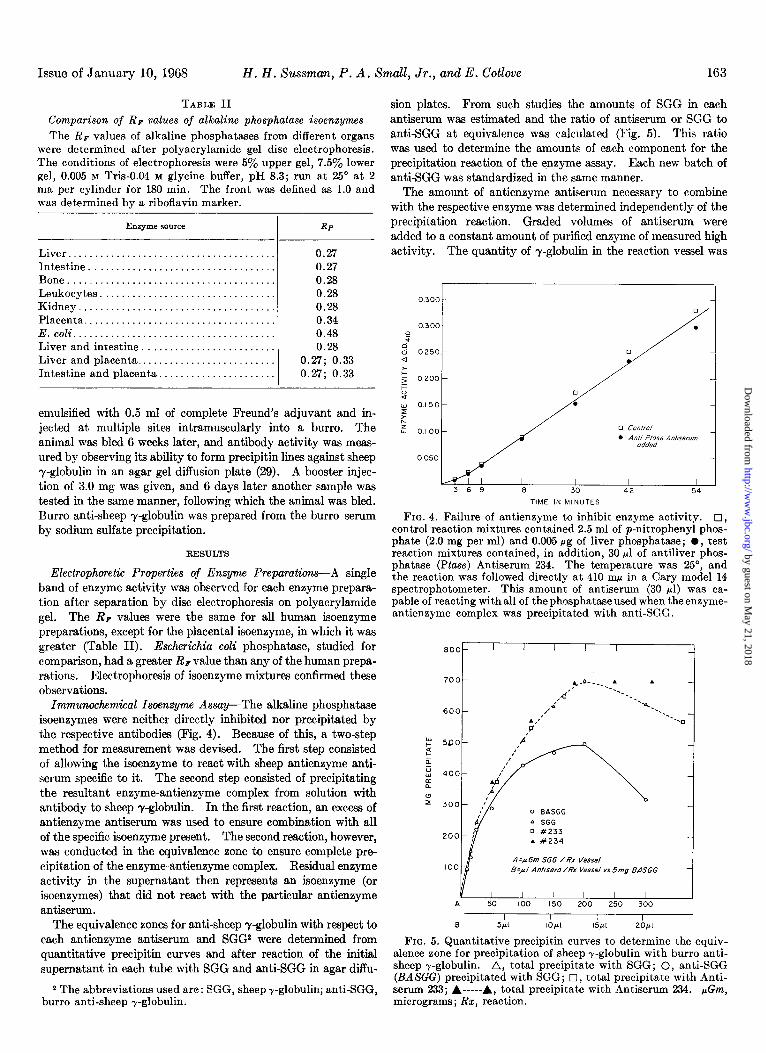
Comparison of Rp values of alkaline phosphatase isoenzymes
The RF values of alkaline phosphatases from different organs were determined after polyacrylamide gel disc electrophoresis. The conditions of electrophoresis were 5% upper gel, 7.5% lower gel, 0.005 M Tris-0.04 M glycine buffer, pH 8.3; run at 25’ at 2 ma per cylinder for 180 min. The front was defined as 1.0 and was determined by a riboflavin marker.
Enzymesource RF
Liver....................................... 0.27 Intestine................................... 0.27 Bone....................................... 0.28 Leukocytes. . . . . . . . . . . . . . . 0.28 Kidney..................................... 0.28 Placenta.................................... 0.34 E.coli...................................... 0.48 Liver and intestine. . . . . . . . . . . . . . . 0.28 Liver and placenta.......................... 0.27; 0.33 Intestine and placenta.. . . . . . . . . . . . 0.27; 0.33
emulsified with 0.5 ml of complete Freund’s adjuvant and in- jected at multiple sites intramuscularly into a burro. The animal was bled 6 weeks later, and antibody activity was meas- ured by observing its ability to form precipitin lines against sheep y-globulin in an agar gel diffusion plate (29). A booster injec- tion of 3.0 mg was given, and 6 days later another sample was tested in the same manner, following which the animal was bled.
Burro anti-sheep y-globulin was prepared from the burro serum by sodium sulfate precipitation.
RESULTS
Electrophoretic Properties of Enzyme Preparations-A single band of enzyme activity was observed for each enzyme prepara- tion after separation by disc electrophoresis on polyacrylamide gel. The RF values were the same for all human isoenzyme preparations, except for the placental isoenzyme, in which it was greater (Table II). Escherichiu coli phosphatase, studied for comparison, had a greater RF value than any of the human prepa- rations. Electrophoresis of isoenzyme mixtures confirmed these observations.
Immunochemical Isoenzymc Assay-The alkaline phosphatase isoenzymes were neither directly inhibited nor precipitated by the respective antibodies (Fig. 4). Because of this, a two-step method for measurement was devised. The first step consisted of allowing the isoenzyme to react with sheep antienzyme anti- serum specific to it. The second step consisted of precipitating the resultant enzyme-antienzyme complex from solution with antibody to sheep y-globulin. In the first reaction, an excess of antienzyme antiserum was used to ensure combination with all of the specific isoenzyme present. The second reaction, however, was conducted in the equivalence zone to ensure complete pre- cipitation of the enzyme-antienzyme complex. Residual enzyme activity in the supernatant then represents an isoenzyme (or isoenzymes) that did not react with the particular antienzyme antiserum.
The equivalence zones for anti-sheep y-globulin with respect to each antienzyme antiserum and SGG2 were determined from quantitative precipitin curves and after reaction of the initial supernatant in each tube with SGG and anti-SGG in agar diffu-
2 The abbreviations used are: SGG, sheep r-globulin; anti-SGG, burro anti-sheep r-globulin.
sion plates. From such studies the amounts of SGG in each antiserum was estimated and the ratio of antiserum or SGG to anti-SGG at equivalence was calculated (Fig. 5). This ratio was used to determine the amounts of each component for the precipitation reaction of the enzyme assay. Each new batch of anti-SGG was standardized in the same manner.
The amount of antienzyme antiserum necessary to combine with the respective enzyme was determined independently of the precipitation reaction. Graded volumes of antiserum were added to a constant amount of purified enzyme of measured high activity. The quantity of y-globulin in the reaction vessel was
03cm-
0.300 -
:
: 0250 - 0
E $ 0.200 -
:: s 0150-
L Ls 0.1 00 -
0050-
3 6 9 18 30 42 54 TIME IN MINUTES
FIG. 4. Failure of antienzyme to inhibit enzyme activity. IJ, control reaction mixtures contained 2.5 ml of p-nitrophenyl phos- phate (2.0 mg per ml) and 0.005 pg of liver phosphatase; l , test reaction mixtures contained, in addition, 30 ~1 of antiliver phos- phatase (Ptase) Antiserum 234. The temperature was 25”, and the reaction was followed directly at 410 nm in a Cary model 14 spectrophotometer. This amount of antiserum (30 ~1) was ca- pable of reacting with all of the phosphatase used when the enzyme- antienzyme complex was precipitated with anti-SGG.
0 #233 . #234
A =A Gm SGG /Rx Vessel B=,aI Antisera /Rx Vessel vs 5mg BASGG
Y I I I I I I A 50 100 150 200 250 300
I I I I s 5Pl IOpl 15pl 20,ul
FIG. 5. Quantitative precipitin curves to determine the equiv- alence zone for precipitation of sheep r-globulin with burro anti- sheep r-globulin. A, total precipitate with SGG; 0, anti-SGG (BASGG) precipitated with SGG; 0, total precipitate with Anti- serum 233; A-----& total precipitate with Antiserum 234. pGm, micrograms; Rx, reaction.
by guest on May 21, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Alkaline Phosphatase Isoenzymes Vol. 243, No. 1 164
,233
.I67
: D d
z .I 33
5 F Y .I 00
Y L 6 ,067
.033
pl ANTISERUM #233 /REACTION VESSEL
FIG. 6. Determination of the amount of antienzyme Antiserum 233 necessary to combine with all of a constant amount of enzyme of high activity. The equivalence zone was maintained by the addition of appropriate amounts of nonantibodJ- SGG to each reaction vessel to ensure complete precipitation of the enzyme- antienzyme complex. Residual activity in the supernatant repre- sents the excess of enzyme for each amount of antienzyme added.
II 0
z 100 r .
0 90 P
d d 80 .- r= E 70
= 60 3
5 50 CI i
IO 20 30 40 50 60 70 60 90 100 II0
ENZYME ACTIVITY ADDED PER REACTION VESSEL,milli 0.D.4,0/MlN
FIG. 7. The range of activity of liver phosphatase isoenzymes measurable in the final assay system following precipitation reac- tion with anti-SGG. l , enzyme activity without added anti- enzyme antiserum; 0, enzyme activity with added antienzyme antiserum. Resu!ts are shown for Antiserum 233; similar relations were observed for Antisera 234 and 333. The units expressed as milli O.D.d,o/MIN represent (change in absorbance at 410 rnp per min) X lOma.
kept constant by the addition of reciprocal, graded amounts of nonspecific sheep y-globulin. This procedure maintained the SGG:anti-SGG ratio within the equivalence zone for the precipi- tation reaction. The entire enzyme-antienzyme complex is precipitated within this equivalence zone, and any residual ac- tivity can be interpreted as isoenzyme present in excess of that
TABLE III
SpeciJicity of antiphosphatase antisera
The results are from multiple experiments in which a range of enzyme activities was used.
Antiserum
Antiliver No. 233. . . Antiliver No. 234. . . . Antiplacenta No. 333..
1.
Phosphatase from
Liver Bone ,‘eka ?$i Kidney ‘$~~- E. coli
--~ --~-
+= 0 0 0 0 0 0
+ 0 0 + +/o 0 0 oo+oooo
0 Key: + = positive reaction; more than 90% of the enzyme activity removed from solution by the precipitation reaction. +/O = partial reaction; approximately 15 to 20yo of the enzyme activity removed from solution by the precipitation reaction. 0 = negative reaction; less than 10% of the enzyme activity re- moved from solution by the precipitation reaction.
which reacted with the amount of antienzyme used. Fig. 6 displays the results of such an experiment, in which Antiserum 233 was evaluated. Antisera 234 and 333 were evaluated simi- larly. These studies also show the independence of the two
reactions, antienzyme complex formation and precipitation. The range of isoenzyme activity which would react with the
volume of antienzyme antiserum used in the finally selected assay system was also determined (Fig. 7). In this assay system, in a total volume of 400 ~1, 10 ~1 of sheep antiserum and 5 mg of anti- SGG were used with Antisera 234 and 333; with Antiserum 233 the amounts were 25 ~1 and 12.5 mg. The concentration of y-globulin in each antiserum was similar, as determined by quan- titative precipitin reactions, and so the ratio of antiserum to anti- SGG in the reaction was the same in all instances.
The assay procedure consisted first of allowing the enzyme to react with antienzyme antiserum for 30 min at room tempera-
ture. Next, anti-SGG was added in amounts to give maximum precipitation, and 0.9% NaCl solution was added to bring the mixture to a constant volume. The reaction mixture was then
incubated at 37” for 30 to 60 min and kept at 4” overnight. The next day the precipitate was separated by centrifugation at 800 x 9 for 20 mm at 4”, and an aliquot of the supernatant was assayed for alkaline phosphatase activity. The background phosphatase activities of the antienzyme antiserum and of the anti-SGG were measured in control tubes which had been taken through all steps in the assay, and the values obtained were sub- tracted from the appropriate specimens. In most instances the background activity was small (5 to 10%) compared to the sam- ple enzyme activity. With specimens of lowest enzyme activity, the background was as high as 40 %.
Specijicity of Antisera-The specificity of each antienzyme antiserum was determined by allowing it to react with the purified phosphatase preparations made from the various organs. The subsequent removal of activity from the supernatant depended entirely upon the specificity of the enzyme-antienzyme reaction, because the precipitation reaction was conducted in the equiva- lence zone and was complete. The precipitation reaction was independent of the enzyme-antienzyme reaction (Fig. 6). The data are presented in Table III. Antiserum 233 was specific for liver phosphatase, and Antiserum 333 was specific for placental phosphatase. Antiserum 234 was not specific; it reacted com-
by guest on May 21, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Issue of January 10, 1968 H. H. Sussman, P. A. Small, Jr., and E. Cotlove 165
TABLE IV
Discrimination experiments: ability of antisera to identify specific organ isoenzymes from mixtures
Data are mean values from duplicate analyses performed on separate days. Incubation times: 30 and 33 min; temperature, 37". Assay vessels contained 0.10 ml of supernatant from pre- cipitation reaction; 0.50 ml of p-nitrophenyl phosphate, 6.0 mM; and 0.05 ml of MgC&, 8.0 mM.
Isoenzyme in reaction vessel
Individual values Bone . . . . . . Liver........................... Neutrophils. . . . Kidney. . . . . Placenta........................
Sum of individually measured values........................
Measured value of a mixture of the listed isoenzymes . .
Control
After reaction with antiliver phosphatase
antisera
No. 234 No. 233
AArdmin X 10-a
4.22 10.45
5.85 8.90 0.35
29.77
34.40
4.22 4.22 0 0.80 0 5.85 7.50 8.00 0.35 0.35
12.47 19.27
Discrimination Experiments-The experiments described above showed that each specific antiserum reacted with its complemen- tary isoenzyme when individually tested. Experiments also were performed to determine whether each antiserum could discriminate its complementary isoenzyme from mixtures of isoenzymes, or whether cross-reaction or coprecipitation might occur. Results of these experiments showed that each specific
antiserum selectively identified its homologous isoenzyme in mix- tures of isoenzyme (Table IV). To test the possibility that iso- enzymes might be bound to a common carrier substance in serum,
similar studies were performed in which human serum and iso- enzyme were incubated at 37” for 30 min prior to immunoassay. If a common carrier existed, coprecipitation in the second reac- tion could occur and negate the specificity of the enzyme-anti- enzyme reaction in assay of serum or other biological fluids. These experiments showed that such was not the case, for no co- precipitation occurred (Table V). Residual activity in the supernatant was quantitatively similar to that in experiments without added serum. Measurement of enzyme activity in the precipitate confirmed that only the specific isoenzyme was pre- cipitated.
13.50 17.25 DISCUSSION
TABLE V
Discrimination experiments: eflect of preincubation of isoenzymes with human serum on immunoassay specijkity
Human serum, in which phosphatase activity was minimized by heating to 56” for 45 min, was incubated with the isoeneymes at 37” for 30 min prior to immunoassay. The same amount of each organ phosphatase was used in all experiments, whet,her alone or mixed. The enzyme activity in the total precipitate was measured after it had been washed three times with 0.5 ml of cold 0.14 M NaCl solution. Then 0.5 ml of p-nitrophenyl phosphate substrate was added to each tube, and the precipitate was stirred; it readily dissolved in the alkali-buffered substrate. The tubes were incubated at 37” for 30 min. The values are corrected to allow for the precipitate being equivalent to 0.2 ml of the original volume while 0.1 ml of the supernatant was used for analysis.
Liver
Bone
Liver + bone
Placenta Placenta + liver
1
--
-
Wienzyme antiserum
233 234
233 234
233 234
234
nzyme activit: n supernatant
YE nzyme activity in precipitate
AAm AAm
0.150 0.000
0.031 0.125 0.000 0.160
0.130 0.000 0.125 0.000 0.130 0.000
0.260 0.000 0.150 0.115 0.130 0.165
0.135 0.140
0.000 0.170
I
pletely with both liver and neutrophil phosphatases and partially (20%) with kidney phosphatase. None of the antisera reacted with the phosphatases isolated from human bone or intestine or from E. coli.3
3 E. coli enzyme was the gift of Dr. R. W. Brockman, National
In these studies antigenically distinct, organ-specific isoen- zymes of human alkaline phosphatase have been found. Indi- vidual isoenzymes were identified on the basis of antigenic differ- ences by a two-step immunochemical method. The first step, reaction of antienzyme antiserum with isoenzyme, was shown to be specific. The second step, precipitation of the resulting en- zyme-antienzyme antibody complex with anti-y-globulin, was necessary because the antienzyme antibody did not inhibit or precipitate the enzyme. The second step was independent of the first, and did not interfere with its specificity.
Organ-specitic forms of a number of mammalian enzymes have been found. Although human alkaline phosphatase has been assumed to be heterogeneous, evidence for specific organ-derived forms has been contradictory. With the exception of placental phosphatase, electrophoretic and chromatographic characteristics of human alkaline phosphatase isoenzymes are similar, so that these techniques have not been entirely satisfactory for distin- guishing individual forms (8-14, 19, 30, 31). Possible explana- tions for this difficulty are that the properties measured by these techniques are unaltered by molecular substitutions which no not affect size or charge, but can be altered by molecular binding to other components in biological fluids as well as by buffer effects. Although differential inhibition and differential substrate ac- tivity have been reported for some phosphatase isoenzymes (1618), the results are inconclusive, possibly because alkaline phosphatase is a relatively nonspecific enzyme capable of mono- phosphate hydrolase activity with a variety of substrates. Ki- netic measurements have not been satisfactory for distinguishing phosphatase organ isoenzymes because of similar K, values (10, 19, 20). The low K, values and the multiple steps in the over-all reaction reduce the discriminatory ability of this type of study. Furthermore, for comparison of K, data to be of value, uniform conditions of enzyme measurement must be used (32, 33).
Immunochemical methods can be specific and precise for dis-
Institute of Arthritis and Metabolic Diseases. The material was purified with osmotic shock by a method which has not been pub- lished.
by guest on May 21, 2018
http://ww
w.jbc.org/
Dow
nloaded from

166 Alkaline Phosphatase Isoenzymes Vol. 243, No. 1
tinguishing structural differences. If molecules are antigenically distinct, they have different structures; if they are antigenically indistinct, the structures may or may not be the same. In previ- ous studies, antigenic differences of organ-specific mammalian enzymes, including human alkaline phosphatases, have been shown by various immunochemical methods (l-7). In the studies of human alkaline phosphatase, however, cross-reactions prevented conclusive distinction of organ isoenzymes. It was not possible to determine whether these cross-reactions were due to antigenic similarity of isoenzymes, to contaminants, or to non- specific antisera. The present study was designed to circumvent this difficulty by making specific antisera and by testing these against purified isoenzyme preparations. This required purifica- tion of isoenzyme from the designated organs, both for immuniza- tion and for subsequent determination of specificity.
The efficacy of using purified enzyme to make specific antisera was established when no cross-reactivity was observed with anti- liver phosphatase Antiserum 233 and antiplacental phosphatase Antiserum, 333. In contrast, Antiserum 234, prepared from a less pure liver phosphatase preparation than was 233, completely cross-reacted with neutrophil alkaline phosphatase and partially cross-reacted with kidney phosphatase. These observations suggest that the cross-reactivity found in previous studies may have been due to nonspecificity of antisera or to mixtures of iso- enzymes in the assay standards, rather than to common anti- genicity. .. The present data, based on immunochemical identification of Isoenzymes, establish that alkaline phosphatases from human liver and placenta can be distinguished from each other and also from alkaline phosphatase isolated from human bone, intestine, neutrophils, or kidney. The antigenic specificity of liver and placental enzymes suggests that these are separate molecular species. Bone, intestinal, and kidney phosphatases were not isolated in sufficient quantities to make specific antisera. Bone and intestinal phosphatases did not react with any of the anti- sera, whether specific (for liver, No. 233; or placenta, No. 333) or nonspecific (No. 234). Kidney phosphatase reacted partially with Antiserum 234, but not with the other two. These findings suggest that phosphatases of bone, intestine, and kidney might constitute additional distinct molecular species. The partial reaction of kidney phosphatase with antiliver phosphatase Anti- serum 234 may have been due to heterogeneity of the kidney enzyme preparation as well as the antiserum. The cross-reac- tion of Antiserum 234 with neutrophil phosphatase probably was due to heterogeneity of the liver enzyme preparation used for immunization to make that antiserum. The heterogeneity could have resulted from phosphatase of neutrophils if present in the liver in sufficient quantity, or from common antigenicity between neutrophil phosphatase and phosphatase of fixed or free hepatic macrophages. Either case suggests the existence of another phosphatase isoenzyme. None of the antisera reacted with E. coli phosphatase, which was to be expected in view of the phylo- genetic difference.
From these data it is concluded that there are at least three antigenically distinct, organ-specific molecular forms of alkaline phosphatase, i.e. liver, placental, and one or more others. In- direct evidence suggests the existence of more than three anti- genitally distinct isoenzymes, since Antiserum 234 reacted fully with neutrophil phosphatase and partially with kidney phospha- tase, and phosphatase of bone and intestine reacted with none of the antisera.
Acknowledgments-We wish to thank Dr. Raymond Zinn for advice and assistance in immunizing the animals, and Dr. Arlan J. Gottlieb for reviewing the manuscript.
REFERENCES 1. 2.
3. 4.
5.
6. 7. 8.
9.
SCHLAMOWITZ, M., Ann. N. Y. Acad. Sci., 76, 373 (1958). SCHLAMOWITZ, M., AND BODANSKY, O., J. Biol. Chem., 234,
1433 (1959). BOYER. S. H.. Science. 134. 1002 (1961). PELIC~OVA, fX., KRA~L, j., CHI&LA; M., AND VECEREK, B.,
Folia Biochim. (Prague), 11, 208 (19651. NISSELBAUM, J. A., SCHLAMOWITZ, k.: AND BODANSKY, O.,
Ann. N. Y. Acad. Sci.. 94, 970 (19611 CINNADER, B., Ann. N. ?. Acad. Sci.,‘iOS, 495 (1963). BOYER, S. H., Ann. N. Y. Acad. Sci., 103, 938 (1963). CUNNINGHAM, V. R., AND RIMER, J. G., Biochem. J., 89, 50~
(1963). HODSON, A. W., LATNER, A. L., AND RAINE, L., Clin. Chim.
Acta, 7, 255 (1962). 10. Moss, D. W., AND KING, E. J., Biochem. J., 84, 192 (1962). 11. ROSENBERG, I. N., J. Clin. Invest., 38, 630 (1959). 12. CHIANDUSSI, L., GREENE, S. F., AND SHERLOCK, S., Clin. Sci.,
22, 425 (1962). 13. WARNOCK, M., Clin. Chim. Acta, 14, 156 (1966). 14. HAIJE, W. B., AND DE JONG, M., Clin. Chim. Acta, 8,620 (1963). 15. POSEN, S., NEALE, F. C., AND CLUBB, J. S., Ann. Intern. Med.,
62, 1234 (1965). 16. FISHMAN, W. H., GREEN, S., AND INGLIS, N. I., Nature, 198,
685 (1963). 17. KREISHER, J. H., CLOSE, V. A., AND FISHMAN, W. H., Clin.
Chim. Acta, 11, 122 (1965). 18. MORTON, R. K., Biochem. J., 61, 232 (1955). 19. LANDAU, W., AND SCHLAMOWITZ, M., Arch. Biochem. Biophys.,
96, 474 (1961). 20. Moss, D. W., Biochem. J., 81, 441 (1961). 21. GUTMAN, A. B., Amer. J. Med., 27, 875 (1959). 22. SUSSMAN, H. H., AND COTLOVE, E., Fed. Proc., 26, 279 (1967). 23. BESSEY, 0. A., LOWRY, 0. H., AND BROCK, M. J., J. Biol. Chem.,
164, 321 (1946). 24. DAVIS, B. J., Ann. N. Y. Acad. Sci., 121, 404 (1964). 25. FALLON, H. J., FREI, E., DAVISDON, J. D., AND TRIER, J. S.,
J. Lab. Clin. Med., 69, 779 (1962). 26. MORTON, R. K., Biochem. J., 67, 594 (1954). 27. VOLKIN, E., in S. P. COLOWICK AND N. 0. KAPLAN (Editors),
Methods in enzymology, Vol. II, Academic Press, New York, 1955, p. 539.
28. KECKWICK, R. A., Biochem. J., 34, 1246 (1940). 29. OUCHTERLONY, O., Acta Pathol. Microbial. Stand., 26, 507
(1949). 30. ESTBORN, B., Z. Klin. Chem., 2, 53 (1964). 31. BUTTERWORTH, P. J., Biochem. H., 96, 74~ (1965). 32. MOTZOK, I., AND BRANION, H. D., Biochem. J., 72, 177 (1959). 33. JENKINS, W. T., AND D’ARI, L., J. Biol. Chem., 241, 295 (1966).
by guest on May 21, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Howard H. Sussman, Parker A. Small, Jr. and Ernest CotloveORGAN-SPECIFIC ISOENZYMES
Human Alkaline Phosphatase: IMMUNOCHEMICAL IDENTIFICATION OF
1968, 243:160-166.J. Biol. Chem.
http://www.jbc.org/content/243/1/160Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/243/1/160.full.html#ref-list-1
This article cites 0 references, 0 of which can be accessed free at
by guest on May 21, 2018
http://ww
w.jbc.org/
Dow
nloaded from





























