Embed Size (px)
Citation preview

Hugan Qingzhi medication ameliorates hepatic steatosis by activatingAMPK and PPARα pathways in L02 cells and HepG2 cells
JinJin Yin a, YanQin Luo b, HouLiang Deng a, ShuMin Qin c, WaiJiao Tang a,Lu Zeng a, BenJie Zhou a,n
a Center for Drug Research and Development, Zhujiang Hospital, Southern Medical University, Guangdong, Guangzhou 510282, PR Chinab Department of Pharmacy, Nanfang Hospital, Southern Medical University, Guangzhou 510515, PR Chinac Institute of Digestive Disease, Longhua Hospital, Shanghai University of Traditional Chinese Medicine, Shanghai 200032, PR China
a r t i c l e i n f o
Article history:Received 15 November 2013Received in revised form15 March 2014Accepted 5 April 2014Available online 13 April 2014
Chemical compounds studied in this article:Quercetin (PubChem CID: 5280343)23-O-acetylalisol B (PubChem CID:3084460)Ursolic acid (PubChem CID: 64945)Nuciferine (PubChem CID: 10146)Isorhamnetin-3-O-neohesperidoside(PubChem CID: 24204448)
Keywords:Hugan Qingzhi tabletHepatic steatosisOxidative stressAdiponectinAMPKPPARα
a b s t r a c t
Ethnopharmacological relevance: Hugan Qingzhi tablet (HQT), a lipid- lowering traditional Chinesemedicine formula, has been used for the prevention and treatment of nonalcoholic fatty liver (NAFLD).Aim of the study: This study was realized to evaluate the effects of HQT-medicated serum on hepaticsteatosis using in vitro experiments with cells and explore the relevant mechanisms with method ofserum pharmacology.Materials and methods: A model of hepatic steatosis in the L02 and HepG2 cells was induced by free fattyacid (FFA). The components in the HQT-medicated serum were assayed by high-performance liquidchromatography. Intracellular lipid droplets were detected by Oil Red O staining, and their ultrastructurewas examined by transmission electron microscope. The biochemical parameters, including triglyceride(TG), lactate dehydrogenase (LDH), aspartate aminotransferase (AST) and alanine aminotransferase (ALT),total antioxidant capacity (T-AOC), malondialdehyde (MDA), superoxide dismutase (SOD) and glu-tathione (GSH), were measured with commercial kits. Furthermore, the expression of adiponectin, AMP-activated protein kinase (AMPK) phosphorylation, sterol regulatory element-binding protein 1 (SREBP-1),peroxisome proliferator activated receptor-α (PPARα), carnitine palmitoyltransferase 1 (CPT-1), andacetyl-CoA oxidase 1 (ACOX1) was analyzed by Western blot and/or quantitative reverse transcription–polymerase chain reaction (qRT–PCR).Results: Moderate- and high-dose HQT-medicated serum reduced (Po0.05 or Po0.01) the accumulationof lipid droplets and the cellular TG content in L02 and HepG2 cells. They caused significant reductions(Po0.01) in LDH, AST, ALT and MDA and significant increase (Po0.05 or Po0.01) in T-AOC in the culturemedium. They also caused increase (Po0.05 or Po0.01) in GSH level and SOD activity in FFA-inducedsteatotic L02 and HepG2 cells. Furthermore, moderate- and high-dose HQT-medicated serum enhanced(Po0.01) adiponectin expression in a concentration-dependent manner and increased (Po0.05 orPo0.01) the phosphorylation of AMPK and the expression of PPARα, CPT-1, and ACOX1, and reduced(Po0.05 or Po0.01) the expression of SREBP-1.Conclusion: The results suggested that HQT-medicated serum exerts a preventive effect against hepaticsteatosis, and the potential mechanism might be activation of AMPK and PPARα pathways.
& 2014 Elsevier Ireland Ltd. All rights reserved.
1. Introduction
With the increasing numbers of adults and childrenwho are obeseor overweight, nonalcoholic fatty liver disease (NAFLD) is a highlyprevalent chronic liver disease (Hesham, 2009; Trauner et al., 2010). Itencompasses a wide spectrum of liver disorders and damages charac-terized by hepatic steatosis without significant alcohol consumption,including simple steatosis, inflammatory non-alcoholic steatohepatitis(NASH), liver cirrhosis and hepatocellular carcinoma (Contos et al.,2004). Although the pathogenesis of NAFLD is not completely under-stood, hepatic lipid metabolic disorders are likely to play importantroles in its initiation and development (Torres et al., 2012). Therefore,
Contents lists available at ScienceDirect
journal homepage: www.elsevier.com/locate/jep
Journal of Ethnopharmacology
http://dx.doi.org/10.1016/j.jep.2014.04.0110378-8741/& 2014 Elsevier Ireland Ltd. All rights reserved.
Abbreviations: ACOX1, acetyl-CoA oxidase 1; ALT, alanine aminotransferase;AMPK, AMP-activated protein kinase; AST, aspartate aminotransferase; ChREBP,carbohydrate-responsive element-binding protein; CPT-1, carnitine palmitoyl-transferase 1; FFA, free fatty acid; GSH, glutathione; HPLC, high-performance liquidchromatography; HQT, Hugan Qingzhi tablet; LDH, lactate dehydrogenase; MDA,malondialdehyde; NAFLD, nonalcoholic fatty liver disease; NASH, non-alcoholicsteatohepatitis; PPARα, peroxisome proliferator activated receptor-α; qRT –PCR,quantitative reverse transcription–polymerase chain reaction; SOD, superoxidedismutase; SREBP-1, sterol regulatory element-binding protein 1; T-AOC, totalantioxidant capacity; TG, triglyceride
n Corresponding author. Tel.: þ86 02061634549; fax: þ86 02084300639.E-mail addresses: [email protected], [email protected] (B. Zhou).
Journal of Ethnopharmacology 154 (2014) 229–239

understanding the molecular mechanisms involved in lipid metabo-lism is an essential step in the prevention and treatment of obesity-related NAFLD.
Mounting evidence suggests that AMP-activated protein kinase(AMPK) and peroxisome proliferator activated receptor-α (PPARα) aretwo critical regulators of hepatic lipid metabolism and may betherapeutic targets in the treatment of NAFLD (Brown and Goldstein,1997; Zhou et al., 2001; Lee et al., 2010). AMPK, an intracellular sensorof energy homeostasis, mediates lipid metabolism through severallipid-metabolism-related transcription factors, such as sterol regula-tory element-binding protein 1 (SREBP-1) and carbohydrate-respon-sive element-binding protein (ChREBP) (Long and Zierath, 2006). Andactivation of AMPK leads to the inhibition of lipogenesis and stimula-tion of fatty acid oxidation, glucose production and protein synthesisin the liver (Viollet et al., 2009). PPARα, a ligand-activated nuclearhormone receptor, regulates the expression of lipid oxidation genes,such as acyl-CoA oxidase 1 (ACOX1) and carnitine palmitoyltransferase1 (CPT-1) (Tailleux et al., 2012). Inhibition of PPARα has been observedin patients with hepatic steatosis, and PPARα agonists have been usedto alleviate this condition (Seo et al., 2008). Therefore, AMPK andPPARα have emerged as particularly promising therapeutic targets forthe prevention of fatty liver disease.
Traditional Chinese medicines have long been used to treat NAFLD,with few adverse effects (Chen et al., 2011; Xie et al., 2012; Feng et al.,2013). Hugan Qingzhi tablet (HQT), a Chinese herbal formula, has along history of use to alleviate NAFLD in clinical practice. The formulamainly consists of five plant materials listed in Table 1. The majorcomponents of HQT, such as Rhizoma Alismatis and Fructus Crataegi,had been reported to improve hyperlipidemia (Dan et al., 2011; Niu etal., 2011), and our previous animal study showed that HQT reducedthe liver index and serum triglycerides, and increased the antioxidantcapacity in a NAFLD rat model (Tang et al., 2013). However, themolecular mechanism involved in its inhibition of hepatic lipidaccumulation requires further investigation.
Many researchers believe that serum pharmacology is morescientific and more suitable for traditional Chinese medicines thantraditional pharmacology (Tang et al., 2009; Bochu et al., 2005).Considering the key roles of AMPK and PPARα activation in theregulation of lipid metabolism and the potential capacity of HQT toprevent hepatic steatosis, we hypothesized that HQT ameliorateshepatic steatosis through the AMPK and PPARα signaling pathways.To test our hypothesis, we systematically evaluated the therapeuticeffects of HQT on steatotic L02 and HepG2 cells induced with freefatty acid (FFA), and explored the mechanisms involved using themethod of serum pharmacology.
2. Materials and methods
2.1. Plant material and preparation of HQT
The component herbs of HQT were purchased from theGuangzhou Chinese Crude Drug Co. (Guangzhou, China) andauthenticated by Professor Chuanming Liu, School of Chinese
Medicine, Southern Medical University, Guangzhou, China. Vou-cher specimens (1209011YP, 1209012YP, 1209013YP, 1209014YPand 1209015YP for Rhizoma Alismatis, Fructus Crataegi, PollenTyphae, Folium Nelumbini and Radix Notoginseng, respectively)have been deposited in Southern Medical University, Guangzhou510515, PR China.
HQT (batch no. 121023) was supplied by Zhujiang Hospital,Southern Medical University (Guangzhou, China) in October 2012.30% Rhizoma Alismatis, 30% Fructus Crataegi, 15% Pollen Typhaeand 20% Folium Nelumbinis were mixed and 70% ethanol (6:1, v/w)were added to the mixture. The mixture was heated and refluxedfor 2 h. The ethanol extract was collected, and the residue wasfiltered and extracted twice with the method introduced above. Theethanol extracts were pooled and evaporated to dryness underreduced pressure. The yield of the dried extract from the startingcrude materials was 14.45% (w/w). Then, 5% Radix Notoginseng wasgrinded, sifted, and added to the dried extract to produce HQT. Thequalitative and quantitative assays of ursolic acid (a major compo-nent of HQT) were used to check the basic quality of the HQT withthin-layer chromatography and high-performance liquid chromato-graphy (HPLC) analyses (Zhou et al., 2012).
2.2. Experimental animals
Thirty Sprague Dawley rats (all female), weighing 200720 g,were purchased from the Animal Center of Southern MedicalUniversity, Guangzhou, China. The rats were housed in plasticcages at a room temperature of 2271 1C under a 12 h light–darkcycle. Food and water were available ad libitum. The experimentwas approved by the Animal Ethics Committee of our universityand was performed in strict accordance with the recommenda-tions in the Guide for the Care and Use of Laboratory Animalsof China.
2.3. Preparation of HQT-medicated serum
The 30 rats were randomly divided into five groups. Rats in thevehicle control serum group were given 1mL/100 g of saline; rats inthe three HQT-medicated serum groups were orally administered low(2.7 g/kg/day), moderate (5.4 g/kg/day) and high (10.8 g/kg/day) dosesof HQT; rats in the fenofibrate-medicated serum group were orallyadministered 0.4 g/kg/day fenofibrate. The medication lasted sevenconsecutive days. Blood was collected aseptically via the abdominalaorta 1 h after the last treatment and then centrifuged. The sera fromthe same group were pooled, filtered through 0.22 μm filters, inacti-vated at 56 1C for 30 min, split, and stored at �70 1C for future use(Zhang et al., 2007).
2.4. Serum sample preparation for HPLC analysis
HPLC was used to analyze the quality of the HQT-medicatedserum by monitoring the three components in it. The standardswere quercetin, 23-O-acetylalisol B, and ursolic acid. These
Table 1Different components in the formula of HQT.
Pharmaceutical name English name Botanical plant name Family Part used Chinese name Ratio
Rhizoma Alismatis Rhizome of oriental water plantain Alisma orientale (Sam.) Juzep Alismataceae Rhizome Ze Xie 6Fructus Crataegi Hawthorn fruit Crataegus pinnatifida Bunge Rosaceae Fruit Shan Zha 6Pollen Typhae Cattail pollen Typha orientalis Presl Typhaceae Pollen Pu Huang 3Folium Nelumbinis Lotus leaf Nelumbo nucifera Gaertn Nymphaeaceae Leaf He Ye 4Radix Notoginseng Sanchi Panax pseudoginseng var Araliaceae Root San Qi 1
J. Yin et al. / Journal of Ethnopharmacology 154 (2014) 229–239230

standards were obtained from the National Institute for theControl of Pharmaceutical and Biology Products (Beijing, China).
Acetonitrile (1 mL) was added to the serum (500 μL) to pre-cipitate the proteins and the serum sample was vortexed for 30 s,then centrifuged for 10 min at 10000� g. The supernatant wastransferred to a clean tube and then evaporated to dryness undernitrogen gas (Yin et al., 2013). The residues were re-dissolved in100 μL of acetonitrile and filtered through a 0.2 μm filter. A 20 μLaliquot was injected onto the HPLC column.
The analytes were measured with a Shimadzu HPLC systemconsisting of two LC-20AT pumps, a Prominence SPD-M20A diodearray detector, an SCL-10A VP system controller, a six-port valve(Valco Instruments, Houston, TX) with a 100 μL sample loop, andan LC Solution workstation (Shimadzu, Japan). The mobile phaseconsisted of 100% acetonitrile (A) and 0.005% phosphoric acid inwater (B), and was programmed as follows: 0–40 min, 85–12% B;40–60 min, 12% B. The flow rate was 0.8 mL/min and the ambienttemperature was set at 30 1C. UV detection was at a wavelength of210 nm.
2.5. Cell culture and free fatty acid (FFA) treatment
Human hepatocyte cell line L02 and hepatocellular carcinomacell line HepG2 were obtained from China Cell Culture Center(Shanghai, China). L02 and HepG2 cells were routinely cultured inRPMI 1640 (Hyclone, Logan, USA) supplemented with 10% fetalbovine serum, 100 U/mL penicillin, and 100 μg/mL streptomycin ina humidified incubator under 95% air and 5% CO2 at 37 1C. The cellswere grown to 80% confluence and then starved in serum-freemedium overnight before treatment. To induce an overload offatty acids, the L02 and HepG2 cells were exposed to a mixtureof FFA (oleic acid:palmitic acid at 2:1) at a final concentration of1 mM for 24 h (Gomez-Lechon et al., 2007).
2.6. Cytotoxicity assay of HQT-medicated serum
The L02 and HepG2 cells were cultured with 10% vehiclecontrol serum, fenofibrate-medicated serum, or, low-, moderate-,or high-dose HQT-medicated serum. After 24 h, the lactate dehy-drogenase (LDH) activity in the supernatant was assayed witha commercial kit (Nanjing Jiancheng Bioengineering Institute,Nanjing, China).
2.7. Experimental design
The L02 and HepG2 cells were seeded into culture plates,incubated for 24 h, and then starved for 24 h in serum-freemedium. They were then divided into six groups: control group,FFA group, FFAþ10% low-dose HQT-medicated serum group(HL), FFAþ10% moderate-dose HQT-medicated serum group(HM), FFAþ10% high-dose HQT-medicated serum group (HH),and FFAþ10% fenofibrate-medicated serum group (FF, the positivecontrol). To ensure that the differences in the effects of the HQT-medicated sera were attributable to HQT and not to the sera, 10%vehicle control serum was added to control group and FFA group.The cells were pre-incubated with the corresponding medicatedsera for 24 h, after which 1 mM FFA was added to the culturesystem except the control group, which was incubated for afurther 24 h. Observations were made with methods appropriateto our different purposes.
2.8. Oil Red O staining
The cells in the different groups were pre-incubated with RMPI1640 containing 10% of the corresponding medicated serum for24 h. They were then exposed to 1 mM FFA for 24 h, fixed with 4%
paraformaldehyde and stained with a freshly prepared workingsolution of Oil Red O for 30 min at room temperature followed bybeing counterstained with hematoxylin before microscopic exam-ination (Olympus Corporation, Tokyo, Japan). To quantify the OilRed O content, isopropanol was added to each sample, which wasshaken at room temperature for 5 min, and the optical density ofeach 100 μL isopropanol-extracted sample was read with a spec-trophotometer at 510 nm.
2.9. Transmission electron microscopy
The cells were collected and fixed with 10% glutaraldehyde in0.1 M sodium cacodylate buffer (pH 7.4) for 1 h at room temperatureand then for 4 h at 4 1C. After fixation, the cells were dehydrated in agraded series of acetone and embedded in epoxy resin. Ultrathinsections (60–70 nm) were cut, stained with uranyl acetate and leadcitrate, and the sections were examined under a transmission electronmicroscope (JEM 1010; Japan).
2.10. Measurement of biochemical parameters and oxidative stress
The biochemical parameters, including triglyceride (TG), aspartateaminotransferase (AST), alanine aminotransferase (ALT) and LDH,were measured with enzymatic assays from Nanjing JiangchengBioengineering Institute (Nanjing, China), according to the manufac-turer's instructions. Absorbances were measured with a microplatespectrophotometric system at 500 nm, 510 nm, 510 nm, and 450 nm,respectively. The result of TG was expressed as μg TG per mg protein(μg/mg protein), and the results were expressed as unit per liter (U/L) for AST, ALT, and LDH. Markers of oxidative stress, including thetotal antioxidative capacity (T-AOC), malondialdehyde (MDA), super-oxidase dismutase (SOD), and glutathione (GSH), were measuredwith spectrophotometry using commercial assay kits from theJiancheng Bioengineering Institute. Absorbances were measuredwith a spectrophotometric system at 520 nm, 532 nm, 450 nm, and420 nm, respectively. The results were expressed as unit per milliliter(U/mL), nmol per milliliter (nmol/mL), unit per mg protein (U/mgprotein), and nmol per mg protein (nmol/mg protein), respectively.The cell protein contents were determined with a BCA Protein AssayKit (Pierce Biotechnology, Rockford, USA).
2.11. Western blot analysis
Cell lysates were prepared in RIPA buffer on ice. The proteinconcentrations were determined with the BCA Protein Assay Kit(Pierce Biotechnology). Equal amounts of protein were separatedby 6–15% SDS-PAGE, and transferred to polyvinylidene difluoride(PVDF) membranes (Millipore, Billerica, USA). The membraneswere blocked with 5% nonfat skim milk for 3 h at room tempera-ture and immunoblotted with polyclonal primary antibodiesdirected against adiponectin, SREBP-1, PPARα, CPT-1 and ACOX1(Abcam, Cambridge, MA), p-AMPK (Cell Signaling Technology, CA,USA) and glyceraldehyde 3-phosphate dehydrogenase (GADPH,the internal control). After the membranes had been washed, theimmunoblots were incubated with peroxidase-conjugated second-ary antibodies (diluted 1:5000) at 25 1C for 1 h. The proteins weredetected with a chemiluminescent substrate (ECL, Amersham,USA). Image J software was used for the densitometric analysisof the bands.
2.12. Real-time reverse transcription–polymerase chain reaction(RT–PCR)
Total RNA was extracted with Trizol Reagent (Invitrogen,Carlsbad, USA), according to the manufacturer's instructions. TotalRNA (1 μg) was reverse transcribed using an oligo (dT)16 primer to
J. Yin et al. / Journal of Ethnopharmacology 154 (2014) 229–239 231

generate cDNA. Real-time PCR was used to measure the mRNAexpression levels of SREBP-1, PPARα, CPT-1, ACOX1 and β-actin. Forreal-time PCR, the cDNA and primers were prepared with a SYBRGreen qPCR SuperMix Kit (Invitrogen), according to the instruction
manual. The sequences of the primers are shown in Table 2. Theamplification conditions were: 95 1C for 5 min for initial denatura-tion, followed by 40 amplification cycles of denaturation at 95 1Cfor 15 s and combined annealing and extension at 60 1C for 30 s.
Table 2Gene-specific primers used for Real-time RT–PCR.
Gene Forward Reverse
Human SREBP-1 50-CCTGTCTCTAGCCACTTTG-30 50-CGGTCTTAGGGTCAAGATC-30
Human PPARα 50-TGGCTCTTGACCCTATTGG-30 50-GGGAACAGATTTCCACATTG-30
Human CPT-1 50-CCTCCGTAGCTGACTCGGTA-30 50-GGAGTGACCGTGAACTGAAAG-30
Human ACOX1 50-GCGGACTACACTTCATAAATGC-30 50-CCACAGGACACCATTAAGC-30
Human β-actin 50-TGGATCAGCAAGCAGGAGTA-30 50-TCGGCCACATTGTGAACTTT-30
Fig. 1. HPLC pattern of HQT-medicated serum. (A) Standard reference material. quercetin (16.883 min, peak (1)), 23-O-acetylalisol B (42.762 min, peak (2)) and ursolic acid(45.243 min, peak (3)). (B) Blank serum. (C) HQT-medicated serum.
J. Yin et al. / Journal of Ethnopharmacology 154 (2014) 229–239232

2.13. Statistical analysis
All experimental data are expressed as means7S.D. The groupeddata were analyzed with SPSS 13.0. Statistical analysis was per-formed by analysis of variance (ANOVA) followed by Bonferroni or
Dunnett's T3 post hoc analysis (parametric). Po0.05 or Po0.01 wasconsidered to indicate statistical significance.
3. Results
3.1. HPLC analysis of HQT-medicated serum
HPLC showed that the HQT-medicated serum contained com-plex components, with several peaks at various retention times(Fig. 1C), those components including prototype componentsin crude drugs and their metabolites. The retention times of quer-cetin, 23-O-acetylalisol B, and ursolic acid were approximately16.883 min, 42.762 min, and 45.243 min, respectively.
3.2. Cytotoxic effects of HQT-medicated sera on L02 and HepG2 cells
The cytotoxic effects of the HQT-medicated sera on L02 and HepG2cells were presented in Table 3. Low-, moderate-, and high-dose
Table 3LDH release for cytotoxicity assay.
Treatment LDH Release (U/L)
L02 HepG2
Vehicle control serum group 106.6778.73 121.3372.52Low-dose HQT-medicated serum group 116.0073.00 a 119.3376.11a
Moderate-dose HQT-medicated serum group 108.3374.04 a 119.0072.00a
High-dose HQT-medicated serum group 104.6777.37 a 122.6774.51a
Fenofibrate-medicated serum group 111.6775.50 a 125.3373.51a
Results are expressed as means7S.D.a P40.05 compared with vehicle control serum group.
Fig. 2. Effects of HQT-medicated serum on cellular steatosis in L02 and HepG2 cells stimulated with FFA. (A) Oil Red O staining (magnification �400). (B) Oil Red Ocolorimetric assay. Results are expressed as means7S.D. nnPo0.01 compared with the control; ##Po0.01 compared with the FFA group. (C) Transmission electronmicroscope (arrows denote lipid droplets. Scale bar: 2 μm, magnification �10000). Con, control group; FFA, FFA group; HL, FFAþ low-dose HQT-medicated serum group;HM, FFAþmoderate-dose HQT-medicated serum group; HH, FFAþhigh-dose HQT-medicated serum group; and FF, FFAþ fenofibrate-medicated serum group. (Forinterpretation of the references to color in this figure legend, the reader is referred to the web version of this article.)
J. Yin et al. / Journal of Ethnopharmacology 154 (2014) 229–239 233

HQT-medicated and fenofibrate-medicated sera had no effect on LDHrelease compared with the vehicle control serum (P40.05).
3.3. HQT-medicated serum reduces FFA-induced cellular steatosis inL02 and HepG2 cells
As shown in Fig. 2A and C, there were marked differencesbetween the control groups and FFA groups. The L02 and HepG2cells in the control group had almost no steatosis, whereas severesteatosis and macro-lipid droplets were observed in the FFA group.The numbers and sizes of the lipid droplets were significantlyreduced in the HQT-medicated serum groups compared with thosein the FFA group. Furthermore, as shown in Fig. 2B, the lipid contentwas significantly elevated in the FFA groups compared with thecontrol groups (Po0.01), and significantly reduced in the HM, HH,and FF groups compared with the FFA groups (Po0.01) of bothcell lines.
3.4. HQT-medicated serum inhibits FFA-induced intracellulartriglyceride accumulation
As shown in Fig. 3, the intracellular TG content increasedsignificantly in the FFA group (Po0.01 versus the control group),and decreased significantly in the HM, HH and FF groups of bothcell lines (Po0.05 or Po0.01 versus the FFA group).
3.5. Effects of HQT-medicated serum on AST, ALT, and LDH
As shown in Fig. 4, the activities of the three biochemicalenzymes, including AST, ALT and LDH, were significantly increasedin the FFA-treated L02 and HepG2 cells compared with those inthe control group. However, pretreatment of the cells withmoderate- and high-dose HQT-medicated serum reduced thesethree enzyme activities (Po0.01 versus the FFA group), in addi-tion, the activities of these three enzymes were also reduced in theFF group.
3.6. Effects of HQT-medicated serum on oxidative stress
The levels of T-AOC and GSH, and SOD activities in the FFA groupswere all lower than those in the control groups (Po0.05 or Po0.01),whereas the levels of MDA were dramatically increased in both celllines compared with those in the control groups (Po0.01), suggest-ing that L02 and HepG2 cells treated with FFA exhibited oxidativestress. Moderate- and high-dose HQT-mediated serum increased thelevels of T-AOC and GSH, and SOD activities (Po0.05 or Po0.01),
and reduced the levels of MDA (Po0.01) compared with those inthe FFA group. Moreover, the levels of T-AOC and GSH, and SODactivities also increased and the levels of MDA decreased in the HLgroups of both cell lines, but the differences were not significantcompared with those in the FFA groups (Fig. 5).
3.7. Effects of HQT-medicated serum on FFA-inhibited adiponectinexpression in HepG2 cells
As shown in Fig. 6, concomitant with the improvement insteatosis, the expression of adiponectin in the FFA group was muchlower than that in the control group (Po0.01). However, pretreat-ment of the cells with one of the three different doses of HQT-medicated serum caused the expression of adiponectin protein toincrease dramatically (Po0.05 or Po0.01 versus the FFA group).
Fig. 3. Effect of HQT-medicated serum on the TG content in L02 and HepG2 cellsstimulated with FFA. Con, control group; FFA, FFA group; HL, FFAþ low-dose HQT-medicated serum group; HM, FFAþmoderate-dose HQT-medicated serum group;HH, FFAþhigh-dose HQT-medicated serum group; and FF, FFAþfenofibrate-medi-cated serum group. Results are expressed as means7S.D. nnPo0.01 compared withthe control; #Po0.05, ##Po0.01 compared with the FFA group.
Fig. 4. Effects of HQT-medicated serum on activities of AST (A) and ALT (B) and LDH(C) of L02 and HepG2 cells stimulated with FFA. Con, control group; FFA, FFA group;HL, FFAþ low-dose HQT-medicated serum group; HM, FFAþmoderate-dose HQT-medicated serum group; HH, FFAþhigh-dose HQT-medicated serum group; and FF,FFAþfenofibrate-medicated serum group. Results are expressed as means7S.D.nnPo0.01 compared with the control group; #Po0.05, ##Po0.01 compared withthe FFA group.
J. Yin et al. / Journal of Ethnopharmacology 154 (2014) 229–239234
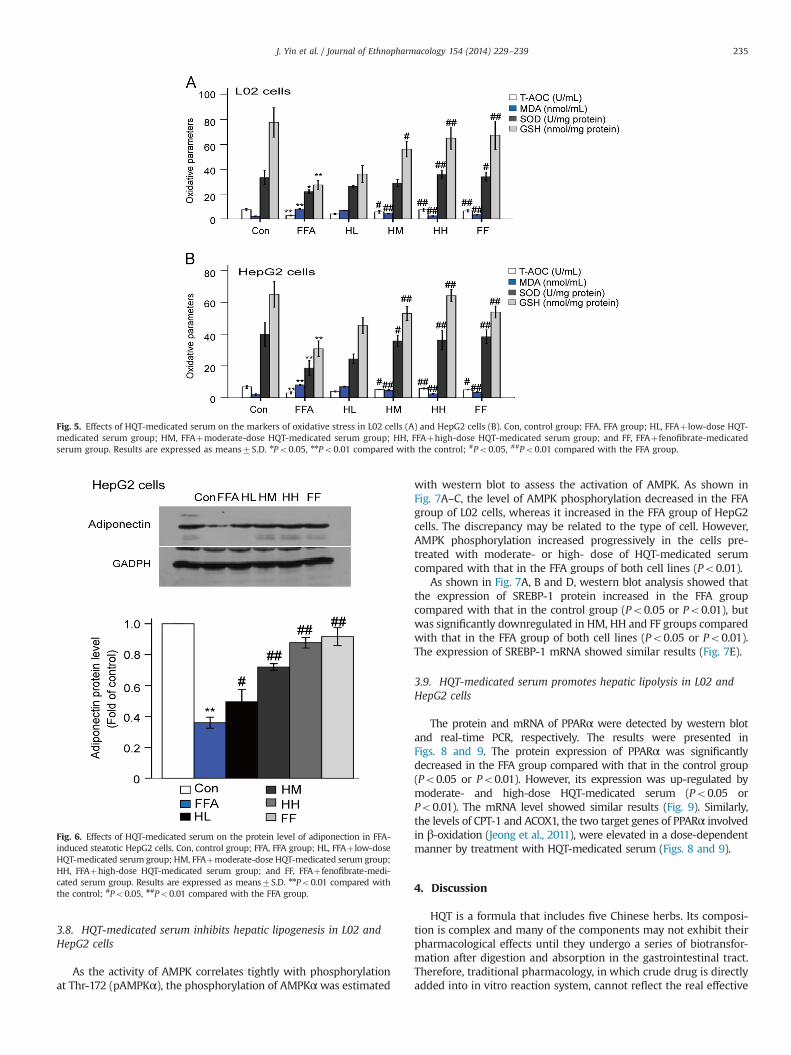
3.8. HQT-medicated serum inhibits hepatic lipogenesis in L02 andHepG2 cells
As the activity of AMPK correlates tightly with phosphorylationat Thr-172 (pAMPKα), the phosphorylation of AMPKαwas estimated
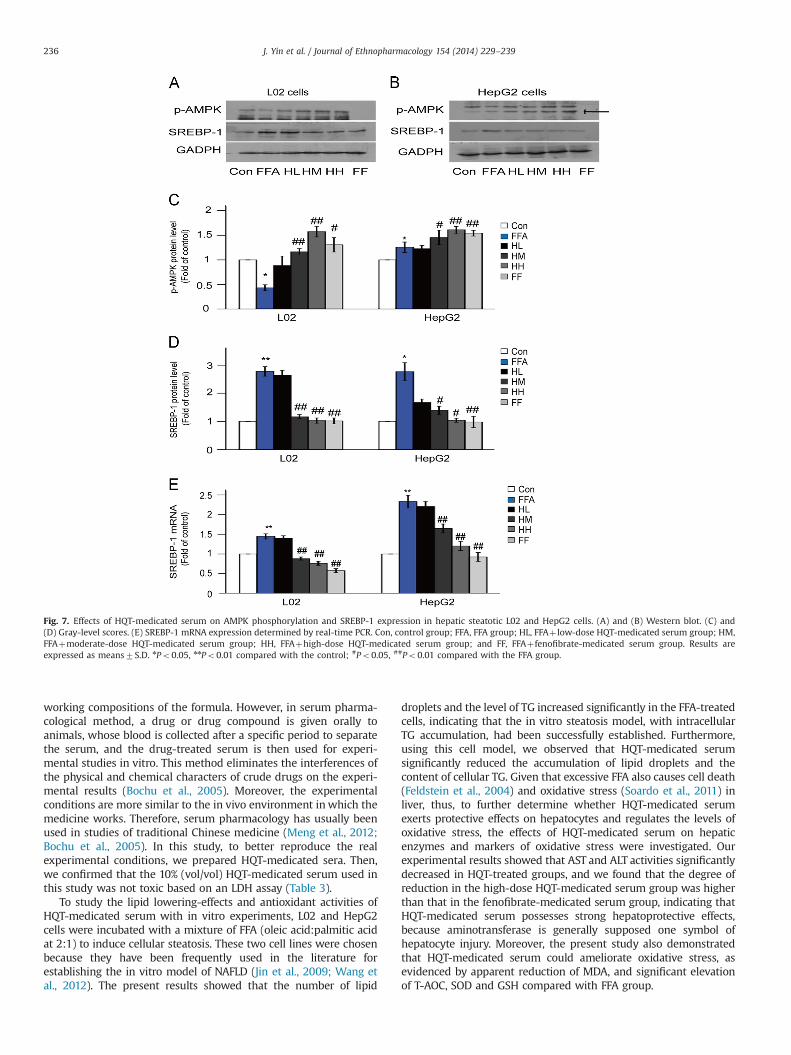
with western blot to assess the activation of AMPK. As shown inFig. 7A–C, the level of AMPK phosphorylation decreased in the FFAgroup of L02 cells, whereas it increased in the FFA group of HepG2cells. The discrepancy may be related to the type of cell. However,AMPK phosphorylation increased progressively in the cells pre-treated with moderate- or high- dose of HQT-medicated serumcompared with that in the FFA groups of both cell lines (Po0.01).
As shown in Fig. 7A, B and D, western blot analysis showed thatthe expression of SREBP-1 protein increased in the FFA groupcompared with that in the control group (Po0.05 or Po0.01), butwas significantly downregulated in HM, HH and FF groups comparedwith that in the FFA group of both cell lines (Po0.05 or Po0.01).The expression of SREBP-1 mRNA showed similar results (Fig. 7E).
3.9. HQT-medicated serum promotes hepatic lipolysis in L02 andHepG2 cells
The protein and mRNA of PPARα were detected by western blotand real-time PCR, respectively. The results were presented inFigs. 8 and 9. The protein expression of PPARα was significantlydecreased in the FFA group compared with that in the control group(Po0.05 or Po0.01). However, its expression was up-regulated bymoderate- and high-dose HQT-medicated serum (Po0.05 orPo0.01). The mRNA level showed similar results (Fig. 9). Similarly,the levels of CPT-1 and ACOX1, the two target genes of PPARα involvedin β-oxidation (Jeong et al., 2011), were elevated in a dose-dependentmanner by treatment with HQT-medicated serum (Figs. 8 and 9).
4. Discussion
HQT is a formula that includes five Chinese herbs. Its composi-tion is complex and many of the components may not exhibit theirpharmacological effects until they undergo a series of biotransfor-mation after digestion and absorption in the gastrointestinal tract.Therefore, traditional pharmacology, in which crude drug is directlyadded into in vitro reaction system, cannot reflect the real effective
Fig. 5. Effects of HQT-medicated serum on the markers of oxidative stress in L02 cells (A) and HepG2 cells (B). Con, control group; FFA, FFA group; HL, FFAþ low-dose HQT-medicated serum group; HM, FFAþmoderate-dose HQT-medicated serum group; HH, FFAþhigh-dose HQT-medicated serum group; and FF, FFAþ fenofibrate-medicatedserum group. Results are expressed as means7S.D. nPo0.05, nnPo0.01 compared with the control; #Po0.05, ##Po0.01 compared with the FFA group.
Fig. 6. Effects of HQT-medicated serum on the protein level of adiponection in FFA-induced steatotic HepG2 cells. Con, control group; FFA, FFA group; HL, FFAþ low-doseHQT-medicated serum group; HM, FFAþmoderate-dose HQT-medicated serum group;HH, FFAþhigh-dose HQT-medicated serum group; and FF, FFAþ fenofibrate-medi-cated serum group. Results are expressed as means7S.D. nnPo0.01 compared withthe control; #Po0.05, ##Po0.01 compared with the FFA group.
J. Yin et al. / Journal of Ethnopharmacology 154 (2014) 229–239 235
working compositions of the formula. However, in serum pharma-cological method, a drug or drug compound is given orally toanimals, whose blood is collected after a specific period to separatethe serum, and the drug-treated serum is then used for experi-mental studies in vitro. This method eliminates the interferences ofthe physical and chemical characters of crude drugs on the experi-mental results (Bochu et al., 2005). Moreover, the experimentalconditions are more similar to the in vivo environment in which themedicine works. Therefore, serum pharmacology has usually beenused in studies of traditional Chinese medicine (Meng et al., 2012;Bochu et al., 2005). In this study, to better reproduce the realexperimental conditions, we prepared HQT-medicated sera. Then,we confirmed that the 10% (vol/vol) HQT-medicated serum used inthis study was not toxic based on an LDH assay (Table 3).
To study the lipid lowering-effects and antioxidant activities ofHQT-medicated serum with in vitro experiments, L02 and HepG2cells were incubated with a mixture of FFA (oleic acid:palmitic acidat 2:1) to induce cellular steatosis. These two cell lines were chosenbecause they have been frequently used in the literature forestablishing the in vitro model of NAFLD (Jin et al., 2009; Wang etal., 2012). The present results showed that the number of lipid
droplets and the level of TG increased significantly in the FFA-treatedcells, indicating that the in vitro steatosis model, with intracellularTG accumulation, had been successfully established. Furthermore,using this cell model, we observed that HQT-medicated serumsignificantly reduced the accumulation of lipid droplets and thecontent of cellular TG. Given that excessive FFA also causes cell death(Feldstein et al., 2004) and oxidative stress (Soardo et al., 2011) inliver, thus, to further determine whether HQT-medicated serumexerts protective effects on hepatocytes and regulates the levels ofoxidative stress, the effects of HQT-medicated serum on hepaticenzymes and markers of oxidative stress were investigated. Ourexperimental results showed that AST and ALT activities significantlydecreased in HQT-treated groups, and we found that the degree ofreduction in the high-dose HQT-medicated serum group was higherthan that in the fenofibrate-medicated serum group, indicating thatHQT-medicated serum possesses strong hepatoprotective effects,because aminotransferase is generally supposed one symbol ofhepatocyte injury. Moreover, the present study also demonstratedthat HQT-medicated serum could ameliorate oxidative stress, asevidenced by apparent reduction of MDA, and significant elevationof T-AOC, SOD and GSH compared with FFA group.
Fig. 7. Effects of HQT-medicated serum on AMPK phosphorylation and SREBP-1 expression in hepatic steatotic L02 and HepG2 cells. (A) and (B) Western blot. (C) and(D) Gray-level scores. (E) SREBP-1 mRNA expression determined by real-time PCR. Con, control group; FFA, FFA group; HL, FFAþ low-dose HQT-medicated serum group; HM,FFAþmoderate-dose HQT-medicated serum group; HH, FFAþhigh-dose HQT-medicated serum group; and FF, FFAþfenofibrate-medicated serum group. Results areexpressed as means7S.D. nPo0.05, nnPo0.01 compared with the control; #Po0.05, ##Po0.01 compared with the FFA group.
J. Yin et al. / Journal of Ethnopharmacology 154 (2014) 229–239236

Adiponectin, an important adipocytokine, is reported to stimu-late glucose utilization and fat oxidation (Yamauchi et al., 2002).Several reports have found that serum level of adiponectin isdecreased in NAFLD (Sargin et al., 2005; Zheng et al., 2011), andtreatment with adiponectin or drugs that promote the expressionof adiponectin could ameliorate NAFLD (Xu et al., 2003; Jung et al.,2013). In this study, we found that HQT-medicated serum sig-nificantly increased the expression of adiponectin protein, and theresult indicates that the therapeutic effect of HQT-medicatedserum on hepatic steatosis might be partly attributable to theup-regulated expression of adiponectin.
In the liver, adiponectin acts by activating the AMPK and PPARαpathways (Kadowaki et al., 2006; Chen et al., 2013). As mentioned inthe introduction, AMPK and PPARα are two critical signaling mole-cules that control the pathways of lipid synthesis and hepatic fattyacid oxidation (You et al., 2004). Activation of AMPK leads to theinhibition of lipogenesis via the suppression of SREBP-1 in the liver.And SREBP-1 is the most important transcription factor regulationgenes involved in de novo lipogenesis (Shimano et al., 1999). PPARα isa nuclear receptor expressed primarily in the liver and regulates thetranscription of genes involved in hepatic fatty acid uptake and
oxidation (Lefebvre et al., 2006; Xu et al., 2009). Activation of PPARαincreases the β-oxidation of fatty acids and reduces cellular lipids(Pyper et al., 2010). In present study, HQT-medicated serum activatedAMPK phosphorylation and PPARα expression, down-regulatedSREBP-1 expression, and up-regulated CPT-1 and ACOX1 expression.These results suggest that HQT-medicated serum could decrease lipidsynthesis and increase fatty acid oxidation through activation ofAMPK and PPARα, which further inhibit the expression of SREBP-1and increase the expression of CPT-1 and ACOX1.
As shown with HPLC, quercetin, 23-O-acetylalisol B and ursolicacid are major constituents of HQT-medicated serum. Quercetin is animportant dietary flavonoid found in apples, berries, citrus fruits, tea,red wine and many medicinal plants, such as Fructus Crataegi andPollen Typhae. It has been reported to ameliorate insulin resistanceand increase cellular antioxidants (Vidyashankar et al., 2013). Quer-cetin also up-regulates the expression of PPARα and CPT-1, tworegulators of fatty acid oxidation, and down-regulates the expressionof SREBP-1 in the liver (Panchal et al., 2012). These changes lead to theattenuation of steatosis through increased fatty acid oxidation in theliver. 23-O-acetylalisol B is a major constituent of Rhizoma Alismatis,which is widely used in traditional Chinese medicine formulas to treat
Fig. 8. Effects of HQT-medicated serum on PPARα, CPT-1 and ACOX1 protein expression in hepatic steatotic L02 and HepG2 cells. Con, control group; FFA, FFA group; HL,FFAþ low-dose HQT-medicated serum group; HM, FFAþmoderate-dose HQT-medicated serum group; HH, FFAþhigh-dose HQT-medicated serum group; and FF,FFAþ fenofibrate-medicated serum group. Results are expressed as means7S.D. nPo0.05, nnPo0.01 compared with the control; #Po0.05, ##Po0.01 compared with theFFA group.
J. Yin et al. / Journal of Ethnopharmacology 154 (2014) 229–239 237

hyperlipidemic patients. It has been reported that treatment withRhizome Aismatis results in clear reductions in serum and livercholesterol and TG, and an increase in serum high-density lipoproteincholesterol in hyperlipidemic mice (Dan et al., 2011). Ursolic acid is atriterpene compound that is present in many fruits and plants, and isthe major component of several traditional medicine herbs, includingFructus Crataegi. It has a wide range of biological functions, includingincreasing the phosphorylation of AMPK and acetyl-CoA carboxylase(ACC) and the expression of CPT-1 and Sirt1 proteins. Besides, ursolicacid has recently been reported to be a PPARα agonist, increasing thebinding of activated PPARα to PPRE, thereby regulating the transcrip-tion of the PPARα target genes involved in cellular lipid metabolism(Jia et al., 2011). From these, we presume that quercetin, RhizomaAlismatis, and ursolic acid are major contributors to the beneficialeffects of HQT on hepatic lipid accumulation by upregulating theadiponectin expression via the activation of AMPK and PPARα, whichfurther inhibit the expression of SREBP-1 and increase the expressionof CPT-1 and ACOX1, and finally lead to a reduction in lipid synthesisand an increase in fatty acid oxidation (Fig. 10).
5. Conclusion
In conclusion, we have demonstrated that moderate and highdoses of HQT not only reduce lipid accumulation but also exert anantioxidant effect. We propose that moderate and high doses of HQTprevent hepatic steatosis by upregulating adiponectin expression,which activates the AMPK and PPARα signaling pathways, therebyinhibiting the expression of SREBP-1 and increasing the expressionof CPT-1 and ACOX1. Thus, hepatic lipogenesis is suppressed and
Fig. 9. Effects of HQT-medicated serum on PPARα, CPT-1 and ACOX1 mRNA levels in hepatic steatotic L02 and HepG2 cells. Con, control group; FFA, FFA group; HL, FFAþ low-dose HQT-medicated serum group; HM, FFAþmoderate-dose HQT-medicated serum group; HH, FFAþhigh-dose HQT-medicated serum group; and FF, FFAþ fenofibrate-medicated serum group. Results are expressed as means7S.D. *Po0.05, **Po0.01 compared with the control; #Po0.05, ##Po0.01 compared with the FFA group.
Fig. 10. HQT-medicated serum inhibits lipid accumulation and oxidative stressthrough the activation of the AMPK and PPARα pathways. HQT-medicated serumincreases the expression of adiponectin and induces the AMPK phosphorylation andPPARα activation, down-regulates SREBP-1 expression and up-regulates CPT-1 andACOX1 expression, thus reducing lipid synthesis and increasing fatty acid oxidation,which ultimately leads to a reduction in lipids ↑: up-regulation; ↓: down-regulation.
J. Yin et al. / Journal of Ethnopharmacology 154 (2014) 229–239238

hepatic lipolysis promoted. Consequently, HQT suppresses lipidaccumulation in the liver and could be developed as a potentialtherapeutic treatment for NAFLD.
Acknowledgments
This study was supported by grants from the Natural ScienceFoundation of China (No. 81274160), the Natural Science Foundationof Guangdong Province (No. S2012010009380) and the Science andTechnology Development project of Guangdong province (No.2010B060900056)
References
Bochu, W., Liancai, Z., Qi, C., 2005. Primary study on the application of SerumPharmacology in Chinese traditional medicine. Colloids and Surfaces B:Biointerfaces 43, 194–197.
Brown, M.S., Goldstein, J.L., 1997. The SREBP pathway: regulation of cholesterolmetabolism by proteolysis of a membrane-bound transcription factor. Cell 89,331–340.
Chen, H., Zhang, L., Li, X., Li, X., Sun, G., Yuan, X., Lei, L., Liu, J., Yin, L., Deng, Q.,Wang, J., Liu, Z., Yang, W., Wang, Z., Zhang, H., Liu, G., 2013. Adiponectinactivates the AMPK signaling pathway to regulate lipid metabolism in bovinehepatocytes. Journal of Steroid Biochemistry and Molecular Biology 138C,445–454.
Chen, J., Zhao, H., Yang, Y., Liu, B., Ni, J., Wang, W., 2011. Lipid-lowering andantioxidant activities of Jiang-Zhi-Ning in Traditional Chinese Medicine. Journalof Ethnopharmacology 134, 919–930.
Contos, M.J., Choudhury, J., Mills, A.S., Sanyal, A.J., 2004. The histologic spectrum ofnonalcoholic fatty liver disease. Clinic in Liver Disease 8, 481–500.
Dan, H., Wu, J., Peng, M., Hu, X., Song, C., Zhou, Z., Yu, S., Fang, N., 2011.Hypolipidemic effects of Alismatis rhizome on lipid profile in mice fed high-fat diet. Saudi Medical Journal 32, 701–707.
Feldstein, A.E., Werneburg, N.W., Canbay, A., Guicciardi, M.E., Bronk, S.F., Rydzewski, R.,Burgart, L.J., Gores, G.J., 2004. Free fatty acids promote hepatic lipotoxicity bystimulating TNF-alpha expression via a lysosomal pathway. Hepatology 40,185–194.
Feng, Q., Gou, X.J., Meng, S.X., Huang, C., Zhang, Y.Q., Tang, Y.J., Wang, W.J., Xu, L.,Peng, J.H., Hu, Y.Y., 2013. Qushi Huayu decoction inhibits hepatic lipidaccumulation by activating AMP-activated protein kinase in vivo and in vitro.Evidence-Based Complementary and Alternative Medicine 2013, 184358.
Gomez-Lechon, M.J., Donato, M.T., Martinez-Romero, A., Jimenez, N., Castell, J.V.,O'Connor, J.E., 2007. A human hepatocellular in vitro model to investigatesteatosis. Chemico Biological Interactions 165, 106–116.
Hesham, A.H., 2009. Nonalcoholic fatty liver disease in children living in theobeseogenic society. World Journal of Pediatrics 5, 245–254.
Jeong, H.W., Lee, J.W., Kim, W.S., Choe, S.S., Kim, K.H., Park, H.S., Shin, H.J., Lee, G.Y.,Shin, D., Lee, H., Lee, J.H., Choi, E.B., Lee, H.K., Chung, H., Park, S.B., Park, K.S.,Kim, H.S., Ro, S., Kim, J.B., 2011. A newly identified CG301269 improves lipidand glucose metabolism without body weight gain through activation ofperoxisome proliferator-activated receptor alpha and gamma. Diabetes 60,496–506.
Jia, Y., Bhuiyan, M.J., Jun, H.J., Lee, J.H., Hoang, M.H., Lee, H.J., Kim, N., Lee, D., Hwang, K.Y.,Hwang, B.Y., Choi, D.W., Lee, S.J., 2011. Ursolic acid is a PPAR-alpha agonist thatregulates hepatic lipid metabolism. Bioorganic & Medicinal Chemistry Letters 21,5876–5880.
Jin, X., Yang, Y.D., Chen, K., Lv, Z.Y., Zheng, L., Liu, Y.P., Chen, S.H., Yu, C.H., Jiang, X.Y.,Zhang, C.Y., Li, Y.M., 2009. HDMCP uncouples yeast mitochondrial respirationand alleviates steatosis in L02 and hepG2 cells by decreasing ATP and H2O2levels: a novel mechanism for NAFLD. Journal of Hepatology 50, 1019–1028.
Jung, T.W., Youn, B.S., Choi, H.Y., Lee, S.Y., Hong, H.C., Yang, S.J., Yoo, H.J., Kim, B.H.,Baik, S.H., Choi, K.M., 2013. Salsalate and adiponectin ameliorate hepaticsteatosis by inhibition of the hepatokine fetuin-A. Biochemical Pharmacology86, 960–969.
Kadowaki, T., Yamauchi, T., Kubota, N., Hara, K., Ueki, K., Tobe, K., 2006. Adiponectinand adiponectin receptors in insulin resistance, diabetes, and the metabolicsyndrome. Journal of Clinical Investigation 116, 1784–1792.
Lee, M.S., Kim, D., Jo, K., Hwang, J.K., 2010. Nordihydroguaiaretic acid protectsagainst high-fat diet-induced fatty liver by activating AMP-activated proteinkinase in obese mice. Biochemical and Biophysical Research Communications401, 92–97.
Lefebvre, P., Chinetti, G., Fruchart, J.C., Staels, B., 2006. Sorting out the roles of PPARalpha in energy metabolism and vascular homeostasis. Journal of ClinicalInvestigation 116, 571–580.
Long, Y.C., Zierath, J.R., 2006. AMP-activated protein kinase signaling in metabolicregulation. Journal of Clinical Investigation 116, 1776–1783.
Meng, Z.Z., Hu, J.H., Chen, J.X., Yue, G.X., 2012. Xiaoyaosan decoction, a traditionalchinese medicine, inhibits oxidative-stress-induced hippocampus neuronapoptosis in vitro. Evidence-Based Complementary and Alternative Medicine2012, 489254.
Niu, C., Chen, C., Chen, L., Cheng, K., Yeh, C., Cheng, J., 2011. Decrease of blood lipidsinduced by Shan-Zha (fruit of Crataegus pinnatifida) is mainly related to anincrease of PPARalpha in liver of mice fed high-fat diet. Hormone and MetabolicResearch 43, 625–630.
Panchal, S.K., Poudyal, H., Brown, L., 2012. Quercetin ameliorates cardiovascular,hepatic, and metabolic changes in diet-induced metabolic syndrome in rats.Journal of Nutrition 142, 1026–1032.
Pyper, S.R., Viswakarma, N., Yu, S., Reddy, J.K., 2010. PPARalpha: energy combustion,hypolipidemia, inflammation and cancer. Nuclear Receptor Signaling 8, e2.
Sargin, H., Sargin, M., Gozu, H., Orcun, A., Baloglu, G., Ozisik, M., Seker, M.,Uygur-Bayramicli, O., 2005. Is adiponectin level a predictor of nonalcoholicfatty liver disease in nondiabetic male patients. World Journal of Gastroenter-ology 11, 5874–5877.
Seo, Y.S., Kim, J.H., Jo, N.Y., Choi, K.M., Baik, S.H., Park, J.J., Kim, J.S., Byun, K.S., Bak, Y.T.,Lee, C.H., Kim, A., Yeon, J.E., 2008. PPAR agonists treatment is effective in anonalcoholic fatty liver disease animal model by modulating fatty-acid metabolicenzymes. Journal of Gastroenterology and Hepatology 23, 102–109.
Shimano, H., Yahagi, N., Amemiya-Kudo, M., Hasty, A.H., Osuga, J., Tamura, Y.,Shionoiri, F., Iizuka, Y., Ohashi, K., Harada, K., Gotoda, T., Ishibashi, S., Yamada,N., 1999. Sterol regulatory element-binding protein-1 as a key transcriptionfactor for nutritional induction of lipogenic enzyme genes. Journal of BiologicalChemistry 274, 35832–35839.
Soardo, G., Donnini, D., Domenis, L., Catena, C., De Silvestri, D., Cappello, D., Dibenedetto,A., Carnelutti, A., Bonasia, V., Pagano, C., Sechi, L.A., 2011. Oxidative stress is activatedby free fatty acids in cultured human hepatocytes. Metabolic Syndrome and RelatedDisorders 9, 397–401.
Tailleux, A., Wouters, K., Staels, B., 2012. Roles of PPARs in NAFLD: potentialtherapeutic targets. Biochimica et Biophysica Acta—Molecular and Cell Biologyof Lipids 1821, 809–818.
Tang, D., Zhang, Z., Gao, Y., Wei, Y., Han, L., 2009. Protective effects of serumcontaining Ginkgo biloba extract on glomerulosclerosis in rat mesangial cells.Journal of Ethnopharmacology 124, 26–33.
Tang, W.J., Zhou, B.J., Zhou, H., 2013. Pharmacodynamics research of Huganqingzhitablet on rats with non-alcoholic fatty liver model. Pharmacology and Clinics ofChinese Materia Medica 29, 169–172.
Torres, D.M., Williams, C.D., Harrison, S.A., 2012. Features, diagnosis, and treatmentof nonalcoholic fatty liver disease. Clinical Gastroenterology and Hepatology 10,837–858.
Trauner, M., Arrese, M., Wagner, M., 2010. Fatty liver and lipotoxicity. Biochimica etBiophysica Acta—Molecular and Cell Biology of Lipids 1801, 299–310.
Vidyashankar, S., Sandeep, V.R., Patki, P.S., 2013. Quercetin ameliorate insulinresistance and up-regulates cellular antioxidants during oleic acid inducedhepatic steatosis in HepG2 cells. Toxicology in vitro 27, 945–953.
Viollet, B., Guigas, B., Leclerc, J., Hebrard, S., Lantier, L., Mounier, R., Andreelli, F.,Foretz, M., 2009. AMP-activated protein kinase in the regulation of hepaticenergy metabolism: from physiology to therapeutic perspectives. Acta Physio-logica 196, 81–98.
Wang, M., Zhao, R., Wang, W., Mao, X., Yu, J., 2012. Lipid regulation effects ofPolygoni Multiflori Radix, its processed products and its major substances onsteatosis human liver cell line L02. Journal of Ethnopharmacology 139,287–293.
Xie, W., Zhao, Y., Du, L., 2012. Emerging approaches of traditional Chinese medicineformulas for the treatment of hyperlipidemia. Journal of Ethnopharmacology140, 345–367.
Xu, A., Wang, Y., Keshaw, H., Xu, L.Y., Lam, K.S., Cooper, G.J., 2003. The fat-derivedhormone adiponectin alleviates alcoholic and nonalcoholic fatty liver diseasesin mice. Journal of Clinical Investigation 112, 91–100.
Xu, K.Z., Zhu, C., Kim, M.S., Yamahara, J., Li, Y., 2009. Pomegranate flowerameliorates fatty liver in an animal model of type 2 diabetes and obesity.Journal of Ethnopharmacology 123, 280–287.
Yamauchi, T., Kamon, J., Minokoshi, Y., Ito, Y., Waki, H., Uchida, S., Yamashita, S.,Noda, M., Kita, S., Ueki, K., Eto, K., Akanuma, Y., Froguel, P., Foufelle, F., Ferre, P.,Carling, D., Kimura, S., Nagai, R., Kahn, B.B., Kadowaki, T., 2002. Adiponectinstimulates glucose utilization and fatty-acid oxidation by activating AMP-activated protein kinase. Nature Medicine 8, 1288–1295.
Yin, D., Liu, Z., Peng, D., Yang, Y., Gao, X., Xu, F., Han, L., 2013. Serum containing Tao-Hong-Si-Wu decoction induces human endothelial cell VEGF production viaPI3K/Akt-eNOS signaling. Evidence-Based Complementary and AlternativeMedicine 2013, 195158.
You, M., Matsumoto, M., Pacold, C.M., Cho, W.K., Crabb, D.W., 2004. The role ofAMP-activated protein kinase in the action of ethanol in the liver. Gastroenter-ology 127, 1798–1808.
Zhang, Y.H., Liu, J.T., Wen, B.Y., Xiao, X.H., 2007. in vitro inhibition of proliferation ofvascular smooth muscle cells by serum of rats treated with Dahuang Zhechongpill. Journal of Ethnopharmacology 112, 375–379.
Zheng, H., Li, S., Ma, L., Cheng, L., Deng, C., Chen, Z., Xie, C., Xiang, M., Jiang, W.,Chen, L., 2011. A novel agonist of PPAR-gamma based on barbituric acidalleviates the development of non-alcoholic fatty liver disease by regulatingadipocytokine expression and preventing insulin resistance. European Journalof Pharmacology 659, 244–251.
Zhou, B., Yan, Y., Huang, S., Yao, Y., Zhang, S., 2012. The quality standard study onHugan Qingzhi tablets. Journal of Chinese Medicinal Materials 35, 644–647.
Zhou, G., Myers, R., Li, Y., Chen, Y., Shen, X., Fenyk-Melody, J., Wu, M., Ventre, J.,Doebber, T., Fujii, N., Musi, N., Hirshman, M.F., Goodyear, L.J., Moller, D.E., 2001.Role of AMP-activated protein kinase in mechanism of metformin action.Journal of Clinical Investigation 108, 1167–1174.
J. Yin et al. / Journal of Ethnopharmacology 154 (2014) 229–239 239












![1 Green Wireless Power Transfer NetworksarXiv:1504.00639v1 [cs.NI] 2 Apr 2015 1 Green Wireless Power Transfer Networks Qingzhi Liu, Michał Golin´ski, Przemysław Pawełczak, and](https://img.dokumen.tips/doc/110x75/5e552312669e957aff4a2913/1-green-wireless-power-transfer-networks-arxiv150400639v1-csni-2-apr-2015-1.jpg)














