Embed Size (px)
Citation preview

http://creativecommons.org/licenses/by-sa/2.0/

Mathematical Modelling of Biological Networks
Prof:Rui [email protected]
973702406Dept Ciencies Mediques Basiques,
1st Floor, Room 1.08Website of the
Course:http://web.udl.es/usuaris/pg193845/Courses/Bioinformatics_2007/ Course: http://10.100.14.36/Student_Server/

Organization of the talk
• Network representations
• From networks to physiological behavior
• Types of models
• Types of problems
• Mathematical formalisms
• Creating and studying a mathematical model

Predicting protein networks using protein interaction data
Database of protein
interactions
Server/
Program
Your Sequence (A)
A
BC
D E
FContinue until you are satisfied
You do not need to go beyond this type of representation in Task 5!!!

What does this means?

Clear network representation is fundamental for clarity of analysis
A B
What does this mean?
Possibilities:
AB
Function
BA
Function
AB
Function
A B
Function
BA
Function

Having precise network representations is important
• We need to know exactly what is being represented, not just that A and B sort of interact in some way.
• This means that it is important to develop or use network representations that are accurate and in which a given element has a very specific meaning.
• Accurate computer representations and human readable representations are not necessarily the same.

Computer readable representations
• SBML (1999)
• CELLML (1999)
• BIOPAX (2002)
• Etc.There must be representations that are easier for humans to use.Let us take a look at one that chemist have been using for a century.

Defining network conventions
A B
C
Full arrow represents a flux of material between A and B
Dashed arrow represents modulation of a flux
+
Dashed arrow with a plus sign represents positive modulation of a flux
-
Dashed arrow with a minus sign represents negative modulation of a flux
A and B – Dependent Variables
(Change over time)
C – Independent variable
(constant value)

Defining network conventions
A B
C
Stoichiometric information needs to be included
Dashed arrow represents modulation of a flux
+
Dashed arrow with a plus sign represents positive modulation of a flux
Dashed arrow with a minus sign represents negative modulation of a flux
23 D+
Reversible Reaction

Defining network conventions
B
C
Stoichiometric information needs to be included
Dashed arrow represents modulation of a flux
+
Dashed arrow with a plus sign represents positive modulation of a flux
Dashed arrow with a minus sign represents negative modulation of a flux
2 A3 D

Renaming Conventions
C
Having too many names or names that are closely related may complicate interpretation and set up
of the model.
Therefore, using a structured nomenclature is important for book keeping
Let us call Xi to variable i
AB
DX3
X1X2
X4

New Network Representation
X2
X3
+2 X13 X4
C
AB
DX3
X1X2
X4

Production and sink reactions
X2X0
Independent Variable
Production Reaction
Sink Reaction

Test Cases: Metabolic Pathway
1 – Metabolite 1 is produced from metabolite 0 by enzyme 1
2 – Metabolite 2 is produced from metabolite 1 by enzyme 2
3 – Metabolite 3 is produced from metabolite 2 by enzyme 3
4 – Metabolite 4 is produced from metabolite 3 by enzyme 4
5 – Metabolite 5 is produced from metabolite 3 by enzyme 5
6 – Metabolites 4 and 5 are consumed outside the system
7 – Metabolite 3 inhibits action of enzyme 1
8 – Metabolite 4 inhibits enzyme 4 and activates enzyme 5
9 – Metabolite 5 inhibits enzyme 5 and activates enzyme 4

Test Cases: Gene Circuit1 – mRNA is synthesized from nucleotides
2 – mRNA is degraded
3 – Protein is produced from amino acids
4 – Protein is degraded
5 – DNA is needed for mRNA synthesis and it transmits information for that synthesis
6 – mRNA is needed for protein synthesis it transmits information for that synthesis
7 – Protein is a transcription factor that negatively regulates expression of the mRNA
7 – Lactose binds the protein reversibly, with a stoichiometry of 1 and creates a form of the protein that does not bind DNA.

Test Cases: Signal transduction pathway
1 – 2 step phosphorylation cascade
2 – Receptor protein can be in one of two forms depending on a signal S (S activates R)
3 – Receptor in active form can phosphorylate a MAPKKK.
4 – MAPKKK can be phosphorylated in two different residues; both can be phosphorylated simultaneously
5 – MAPKK can be phosphorylated in two different residues; both can be phosphorylated simultaneously
6 – Residue 1 of MAPKK can only be phosphorylated if both residues of MAPKKK are phosphorylated
7 – Residue 2 of MAPKKK can be phosphorylated if one and only one of the residues of MAPKKK are phosphorylated.
8 – All phosphorylated residues can loose phosphate spontaneously
9 – Active R inactivates over time spontaneously

Organization of the talk
• Network representations
• From networks to physiological behavior
• Types of models
• Types of problems
• Mathematical formalisms
• Creating and studying a mathematical model

In silico networks are limited as predictors of physiological behavior
What happens?
Probably a very sick mutant?

Dynamic behavior unpredictable in non-linear systems
X0 X1 X2 X3
X0
X1
X2
X3
t0 t1 t2 t3
t
X3

How to predict behavior of network or pathway?
• Build mathematical models!!!!

Organization of the talk
• Network representations
• From networks to physiological behavior
• Types of models
• Types of problems
• Mathematical formalisms
• Creating and studying a mathematical model

Types of Model
• Finite State Models– Bolean Network Models
• Stoichiometric Models– Flux balance analysis models
• Deterministic Models– Homogeneous– Spatial Detail
• Stochastic Models– Homogeneous– Spatial Detail

Finite State models
• A Finite state model is composed of – a set of nodes that are connected by – a set of edges. – Each node can have a finite number of
states and the – rules for changing these states with time
are transmitted through the edges and based on the state of the neighbors.

Boolean Networks
• A Boolean network model is composed of – a set of nodes that are connected by – a set of edges. – each node can have TWO states – the rules for changing these states with
time are transmitted through the edges and based on the state of the neighbors.

Boolean Networks are usefull
• They can give you information about the connectivity of your metabolism or gene circuit
• What you organism can or can not do may also depend on the connectivity of the regulation

Simple Finite State Gene Circuit
A B C
A – Positively regulates itself and b
Negatively regulates C
B – Positively regulates itself and c
C – Positively regulates itself and b
Negatively regulates A
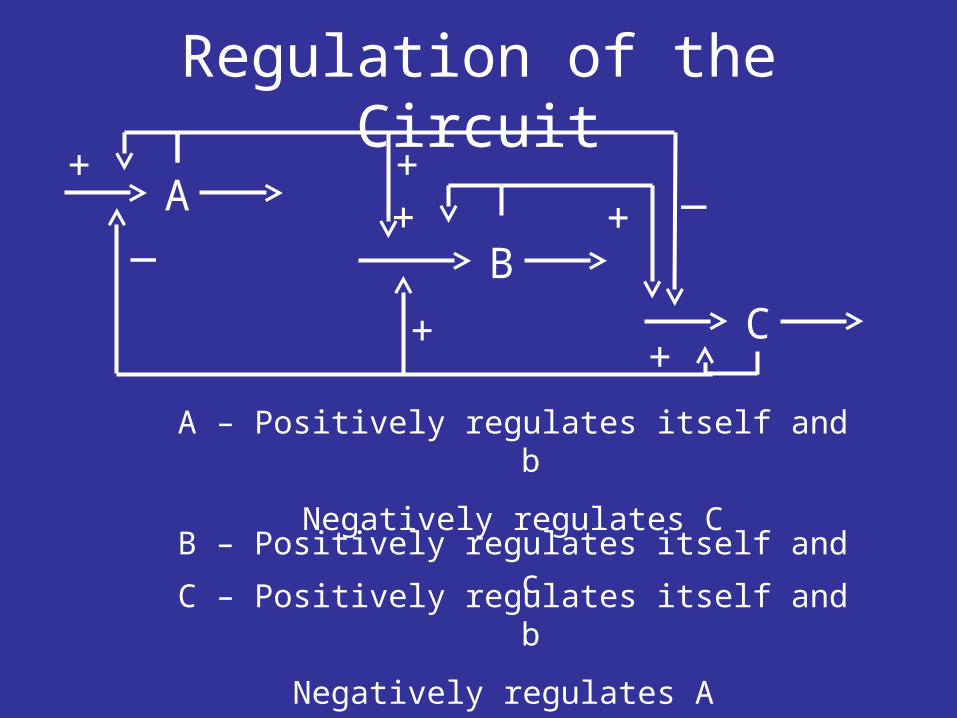
Regulation of the Circuit
A
B
C
A – Positively regulates itself and b
Negatively regulates C
B – Positively regulates itself and c
C – Positively regulates itself and b
Negatively regulates A
+ + _+ +
+
_
+

Threshold of expression in circuit
• A,B, C can have three levels of expression (0,1,2)
• Regulation of A or C occurs whenever a gene is in or above level 1
• Regulation of B occurs whenever a gene is in or above level 1

Logical rules for time change
Level of expression in the absence of all regulators
Level of expression in the presence of A, B or C
Level of expression of A,B, C in the presence of A and B,
A and C or B and C
Level of expression of A,B, C in the presence of A, B and C

Possible states
5 Steady States
2 Oscilatory states
Everything else a transient state

Stoichiometric Models
• A Stoichiometric model is composed of – a stoichiometric matrix that informs on the
number of molecules that are transformed – a flux vector that describes the rates of
change in the system

A simple stoichiometric model
A model system comprising three metabolites (A, B and C) with three reactions (internal fluxes, vi
including one reversible reaction) and three exchange fluxes (bi).

Mass balance
• Stoichiometric matrix S
• Flux matrix v
• S · v = 0 in steady state. Mass balance equations
accounting for all reactions and
transport mechanisms are written for each species. These
equations are then rewritten in matrix
form. At steady state, this reduces
to S · V=0.

Things to do
• Apply graph theory (or other) to derive information from the model
No effect
No Flux

Deterministic Models
• A deterministic model is an extension of a stoichiometric model. It is formed of – a stoichiometric matrix that informs on the
number of molecules that are transformed – a flux vector that describes the rates of
change in the system. The functions are continuous: dX/dt= S · v

A Bifunctional two component system
Bifunctional Sensor
S
S*
R*
R
Q1 Q2

X3
X1
X2
X4
X5 X6
A model with a bifunctional sensor
3/ 1/
4 / 2 /
dX dt dX dt
dX dt dX dt
13 151
11 121
1/ 3 5
1 2
g g
h h
dX dt X X
X X
Bifunctional Sensor O(rdinary)D(iferential)E(quation)s
21 26 224 22
2322 / 1 6 4 2 3g h hg g XdX dt X X X X

Homogenous Deterministic Models
• Assume system is spatially homogeneous
• If not use P(artial)D(ifferential)E(quations)s

Partial Differential Equations - Diffusion
∂
accumulation of [S] due to
transport
dAnJSJ
Rate of change of [S]
local production
of [S]
dVSdt
d
dVfS
f
dVJ
]S[Dft
]S[
]S[DJ From the definition of flux
For one-dimensional space,
x
uD
xu,tf
t
u
X
XX2
2
VVgx
Ddt
dVC
E.g. Extending the Hodgkin-Huxley model to voltage spread:

Non-Homogenous Deterministic Models
• Numerical solution is effectively done by coupling many systems of ODEs
• Much heavier computationally

What do deterministic models assume?
• That the number of particle changes in a continuum
• Is this true? Can we have 1 and half molecules of A?– NO!!!!!
• Now what?!?!?!

What do deterministic models assume?
• Law of large numbers (statistics)– The larger the population, the better the mean
value is as a representation of the population – What if there are 10 TFs in a cell?
– Stochastic models, either homogeneous or non-homogeneous!!!

Stochastic Models
• Replace continuous assumptions by discrete events
• Use rate constants as measures of probability
• Assume that at any give sufficiently small time interval only one event occurs

Organization of the talk
• Network representations
• From networks to physiological behavior
• Types of models
• Types of problems
• Mathematical formalisms
• Creating and studying a mathematical model

Goals of the model (I)
• Large scale modeling– Reconstructing the full network of the genome– Red Blood Cell Metabolism
• Modeling Specific Pathways/Circuits– Non-catalytic lipid peroxidation– MAPK Pathways
• Generating alternative hypothesys for the topology of the model.– ISC Reconstruction– Phosphate metabolism reconstruction

Goals of the model (II)
• Estimating parameter values– Estimating parameter values in the purine
metabolism
• Identifying Design Principles– Latter

Organization of the talk
• Network representations
• From networks to physiological behavior
• Types of models
• Types of problems
• Mathematical formalism
• Creating and studying a mathematical model

Representing the time behavior of your system
/dA dt
/ ,dA
dA dt A f A Cdt
/
dAdA dt
dt
A B
C
+
/dA
dA dt Adt

What is the form of the function?
A B
C
+
A or C
Flux1 2k A k CLinear 1 2
1 2 3 4
k A k C
K K A K C K AC
Saturating
4 41 2
44 41 2 3 4
k A k C
K K A K C K AC
Sigmoid

What if the form of the function is unknown?
A B
C
+
int intintint int
2 2
2int int
int
, ,, ,
, ,
operating operatingoperatingpo popo operating operating
po po
operating operatingpo pooperating
po
df A C df A CdAf A C f A C A A C C
dt dA dC
d f A C d f A CA A C C
dAdC d C
2
int
int
2 2
2int
int
,...
operatingpooperating
po
operatingpooperating
po
C C
d f A CA A
d A
Taylor Theorem:
f(A,C) can be written as a polynomial function of A and C using the function’s
mathematical derivatives with respect to the variables (A,C)

Are all terms needed?
A B
C
+
int
intint
intint
, ,
,
,
operatingpo
operatingpooperating
po
operatingpooperating
po
dAf A C f A C
dt
df A CA A
dA
df A CC C
dC
f(A,C) can be approximated by considering only a few of its mathematical derivatives
with respect to the variables (A,C)

Linear approximation
A B
C
+
1 20, ,
dAf A C f A C k A k C
dt
Taylor Theorem:
f(A,C) is approximated with a linear function by its first order derivatives with respect to
the variables (A,C)
1 2k A k CLinear

What if system is non-linear?
• Use a first order approximation in a non-linear space.

Logarithmic space is non-linear
A B
C
+
1 2, g gdAf A C A C
dt
g<0 inhibits flux
g=0 no influence on flux
g>0 activates flux
Use Taylor theorem in Log space

Why log space?
• Intuitive parameters• Simple, yet non-linear• Convex representation in cartesian
space• Linearizes exponential space
–Many biological processes are close to exponential → Linearizes mathematics

Why is formalism important?
• Reproduction of observed behavior– For example, inverse space may be better for
some models.
• Tayloring of numerical methods to specific forms of mathematical equations

Test Cases: Metabolic Pathway
X0 X1 X2
X3
X4
_
_
_
_
++

Test Cases: Gene circuit
X0 X1
X2 X3
X4 X5
+
++
X6
_
+

Test Cases: Signal transduction
X0
X1
X2
X4
X3
X5
X6
+ +
+
+
X4
X3
X5
X6
+
+
+
+

Inverse space is non-linear
A B
C
+
K<0 inhibits flux
K=0 no influence on flux
K>0 activates flux
Use Taylor theorem in inverse space

Organization of the talk
• Network representations
• From networks to physiological behavior
• Types of models
• Types of problems
• Mathematical formalism
• Creating and studying a mathematical model

A model of a biosynthetic pathway
10 13 111 1 0 3 1 1/ g g hdX dt X X X
11 222 1 1 2 2/ h hdX dt X X
X0 X1
_
+
X2 X3
X4
22 33 343 2 2 3 3 4/ h h hdX dt X X X
Constant
Protein using X3

What can you learn?
• Steady state response
• Long term or homeostatic systemic behavior of the network
• Transient response
• Transient or adaptive systemic behavior of the network

What else can you learn?
• Sensitivity of the system to perturbations in parameters or conditions in the medium
• Stability of the homeostatic behavior of the system
• Understand design principles in the network as a consequence of evolution

Steady state response analysis
10 13 111 1 0 3 1 1/ 0g g hdX dt X X X
11 222 1 1 2 2/ 0h hdX dt X X
22 33 343 2 2 3 3 4/ 0h h hdX dt X X X

How is homeostasis of the flux affected by changes in X0?
0 3 0 10 33 13( , ) ( , ) /L V X L X X g h g
Log[X0]
Log[V]
Low g10
Medium g10
Large g10
Increases in X0 always increase the homeostatic values of the flux through the pathway

How is flux affected by changes in demand for X3?
4 13 34 13 33( , ) / 0L V X g h g h
Log[X4]
Log[V]Large g13
Medium g13
Low g13

How is homeostasis affected by changes in demand for X3?
3 4 4 13 34 13 33( , ) ( , ) / / 0L X X L V X g h g h
Log[X4]
Log[X3]
Low g13
Medium g13
Large g13

What to look for in the analysis?
• Steady state response
•Long term or homeostatic systemic behavior of the network
• Transient response
•Transient of adaptive systemic behavior of the network

Transient response analysis
10 13 111 1 0 3 1 1/ g g hdX dt X X X
11 222 1 1 2 2/ h hdX dt X X
22 33 343 2 2 3 3 4/ h h hdX dt X X X
Solve numerically

Specific adaptive response10 13 11
1 1 0 3 1 1/ g g hdX dt X X X 11 22
2 1 1 2 2/ h hdX dt X X 22 33 34
3 2 2 3 3 4/ h h hdX dt X X X
Get parameter valuesGet concentration
valuesSubstitution
Solve numerically
Time
[X3]
Change in X4

General adaptive response10 13 11
1 1 0 3 1 1/ g g hdX dt X X X 11 22
2 1 1 2 2/ h hdX dt X X 22 33 34
3 2 2 3 3 4/ h h hdX dt X X X Normalize
Solve numerically with comprehensive scan of parameter values
Time
[X3]
Increase in X4
Low g13
Increasing g13
Threshold g13
High g13
Unstable system, uncapable of homeostasis if feedback is strong!!

Sensitivity analysis
• Sensitivity of the system to changes in environment–Increase in demand for product causes increase in flux through pathway
–Increase in strength of feedback increases response of flux to demand
–Increase in strength of feedback decreases homeostasis margin of the system

Stability analysis
• Stability of the homeostatic behavior
–Increase in strength of feedback decreases homeostasis margin of the system

How to do it
• Download programs/algorithms and do it– PLAS, GEPASI, COPASI SBML suites,
MatLab, Mathematica, etc.
• Use an on-line server to build the model and do the simulation– V-Cell, Basis

Design principles
•Why are there alternative designs of the same pathway?
•Dual modes of gene control
•Why is a given pathway design prefered over others?
•Overall feedback in biosynthetic pathways

Why regulation by overall feedback?
X0 X1
_
+
X2 X3
X4
X0 X1
_
+
X2 X3
X4
__
Overall feedback
Cascade feedback

Overall feedback improves functionality of the system
TimeSpurious stimulation
[C]Overall
Cascade
Proper stimulus
Overall
Cascade
[C]
StimulusOverall
Cascade
Alves & Savageau, 2000, Biophys. J.

Dual Modes of gene control

Demand theory of gene control
Wall et al, 2004, Nature Genetics Reviews
• High demand for gene expression→ Positive Regulation
• Low demand for gene expression → Negative mode of regulation

Summary
• From networks to physiological behavior
• Network representations
• Types of Models
• Types of Problems
• Mathematical formalism
• Studying a mathematical model













