Embed Size (px)
Citation preview

Francesco D’ErricoUMR 9933 du CNRS, Institut duQuaternaire, Avenue des Facultés,33405 Talence, France
Paola VillaUniversité Bordeaux 1, UMR 9933 duCNRS, Institut du Quaternaire, Avenuedes Facultés, 33405 Talence, France,& University of Colorado Museum,Boulder, Colorado 80309-0315,U.S.A.
Received 12 November 1996Revision received 11 February1997 and accepted 14 February1997
Keywords: Early symbolism, SEM,taphonomy, Lower/MiddlePaleolithic, carnivoremodifications, vascular grooves.
Holes and grooves: the contribution ofmicroscopy and taphonomy to theproblem of art origins
Optical and scanning electron microscopy, comparative anatomy, data frommodern and Pleistocene carnivore accumulations, and analysis of archaeologi-cal materials show that some of the pieces interpreted by various scholars asengraved or perforated bones from European Lower and Middle Paleolithicsites (such as Pech de l’Azé II, Stránska Skála, Kulna, Bois Roche and CuevaMorin) are not early manifestations of non-utilitarian behavior. Putativeengravings are in fact vascular grooves, while perforated pieces are partially-digested bones regurgitated by hyenas. The current debate on art origins hasoften been centered on the symbolic value and cognitive implications of theseand similar pieces without a first-hand analysis of the objects to provideconvincing demonstration of the human origins of the marks. Such demon-stration is a necessary prerequisite to any discussion of their significance for theevolution of symbolic behavior. ? 1997 Academic Press Limited
Journal of Human Evolution (1997) 33, 1–31
Introduction
Two opposing hypotheses have been proposed to account for the origins of symbolic behavior:on one side, there are the partisans of a symbolic explosion corresponding, in Europe, with theMiddle to Upper Paleolithic transition; on the other side, there are those who believe thatacquisition of modern cognitive abilities took place gradually and that evidence of symbolicbehavior is already present in the Lower and Middle Paleolithic.Proponents of gradualism find support in such evidence as ochre or hematite use, collection
and transport of crystals and fossils, and perforated and engraved portable objects of stone andbone found in several Lower and Middle Paleolithic sites in Europe, Africa and Asia. There is,however, no consensus on the nature and the significance of this evidence. Some scholars notonly believe that humans made, collected, or used most of these objects, but are also ready tolend a symbolic or non-utilitarian value to them (e.g., Marshack, 1976, 1988, 1991; Bednarik,1992, 1995; Bahn, 1996; cf. also Simek, 1992; Hayden, 1993; Wolpoff & Caspari, 1996).Others do not systematically question the artefactural nature of the objects, but reject theirsupposed symbolic meaning, proposing instead a utilitarian function, or deny their value asexpressions of a symbolic or artistic tradition by stressing the sparsity and heterogeneity of thesample compared with the artefactual richness of the Upper Paleolithic (Chase & Dibble, 1987,1992; Bar Yosef, 1988; Davidson & Noble, 1989; White, 1992, 1995; Stringer & Gamble,1993; Mellars, 1989, 1996; Byers, 1994). A few reject the idea of human manufacturefor several of these objects, and proposed natural causes, such as carnivore gnawing, rootmark-ing, erosion or chemical deterioration (Binford 1981; Davidson 1990; Chase 1990; d’Errico,1991). In sum, this topic has attracted a great deal of attention and has inspired heated debates.In spite of the variety of views expressed, many of these claims have remained unexamined.Although various alternative interpretations have been proposed, criteria to differentiate
between human-made engravings and perforations and those produced by other causes havenever been established; interpretations have remained at the level of conjecture (for anexception concerning perforations produced by carnivore puncture, see Chase, 1990). The
0047–2484/97/070001+31 $25.00/0/hu970141 ? 1997 Academic Press Limited

2 . ’ .
purpose of this paper is to develop criteria for recognizing naturally-produced perforations andengravings by combining optical and scanning electron microscopy, comparative anatomy,and data from modern and Pleistocene bone accumulations. We have found these criteriauseful for interpreting putative engravings and perforated bones from a number of Lower andMiddle Paleolithic sites.
Lower and Middle Paleolithic engraved and perforated bones
GroovesEngraved bones have been claimed for at least 16 Lower and Middle Paleolithic sites inEurope (Table 1). This sample, by no means complete, is composed of a variety of objects:several long bone and rib fragments, one ivory tusk (at Bilzingsleben), one vertebra (at StránskaSkála) one scapula (at La Quina) and one horse canine (at Prolom II). The ‘‘engraved motifs’’are also variable, consisting of notches (e.g., Suard), rectilinear grooves in parallel (e.g., LaQuina) or radial sets (e.g., Bilzingsleben, Stránska Skála) and curvilinear grooves (e.g., Pech del’Azé, Cueva Morin). In the following sections we review a number of purported engravingswith a particular arrangement or groove morphology for which we have been able to developspecific explanatory hypotheses.
Pech de l’Azé II (Dordogne, France). François Bordes (1969, 1972: p. 62) described an ox ribcarrying on its internal face ‘‘a series of lines and incisions which are clearly intentional, not therandom lines left by a flint cutting off the meat. . . . It seems to be by far the oldest engravingknown.’’ The bone (Figure 1) comes from the top of level 8, considered by Bordes as
Table 1 Purported engraved bones from Lower and Middle Paleolithic sites in Europe
Site Age or industry Bone objects Reference
Stránska Skála(Czech Republic)
Early Middle Pleistocene 2 Valoch, 1987, 1995; Patou-Mathis, 1995
Bilzingsleben (Germany) Isotope stage 11 or 9 6 Mania & Mania, 1988Grotte Vaufrey (France) Early Middle Paleolithic 1 Vincent, 1988a,bSuard (France) Middle Paleolithic,
isotope stage 63 Debenath & Duport, 1971
Pech de l’Azé II (France) ‘‘Acheulian’’ isotopestage 6
1 Bordes, 1969; Marshack, 1977
Schulen (Belgium) Last Glacial,Middle Paleolithic
1 Huyge, 1990
Bacho Kiro (Bulgaria) Mousterian 1 Marshack, 1976Prolom II (Ukraine) Last Glacial,
Middle Paleolithic4 Stepanchuk, 1993
La Quina (France) Mousterian 1 Martin, 1907–1910, 1910; Marshack, 1991La Ferrassie (France) Mousterian 2 Capitan & Peyrony, 1921; Marshack, 1976Marillac (France) Mousterian 1 Duport, 1973Petit Puymoyen (France) Mousterian 1 Debenath & Duport, 1971Abri Lartet, Montgaudier(France)
Mousterian 2 Debenath & Duport, 1971
Tagliente (Italy) Mousterian Unspecifiednumber
Leonardi, 1983, 1988
Cueva Morin (Spain) Mousterian 13 Echegaray & Freeman, 1971, 1973;Freeman & Echegaray, 1983
Cueva Beneito (Spain) Mousterian 1 Iturbe et al., 1993

3
Acheulian, and dated to the end of ‘‘Riss I’’; level 8 has now been assigned an estimated age,using electron spin resonance (ESR), of about 150,000 BP (corresponding to isotope stage 6;Grün et al., 1991). Marshack (1976: p. 278) accepted the intentionality of the markings andafter microscopic analysis asserted that ‘‘the image had been made sequentially as a seriesof connected, festooned double arcs, beginning at left and ending at right with a multiplemarking’’. He suggested (1977: p. 292) that this ‘‘meandering or serpentine image’’ involved‘‘cognitive strategies’’ comparable with those on the Romanelli and Parpalló engraved slabs ofUpper Paleolithic age.The intentional nature of the Pech de l’Azé II engraving is accepted by various authors (e.g.,
Delporte, 1990; Bednarik, 1992, 1994, 1995; Valoch, 1994; Cremadés et al., 1995; Wolpoff &Caspari, 1996); the object is cited as a precursor of symbolic representations (Donald, 1991).Other authors are ready to accept the human-made origin of the markings, but express doubtsas to the representational or symbolic value of the artefact (Chase & Dibble, 1987; Bar Yosef,1988; Stringer & Gamble, 1993). Binford (1981) questioned the artefactual nature of theobject, suggesting that the engravings were root marks. Later, Marshack (1991) suggested aparallel between the Pech de l’Azé and Cueva Morin meander (see below). These markingsshould, he says, be interpreted as the result of ‘‘partial and limited deterioration of some of thebone surface under particular conditions of moisture, drainage, acidity . . . chemistry andperhaps bacterial action’’. However, he did not provide a definite interpretation nor diagnosticfeatures supporting his new ideas. More recently, d’Errico (1995) has proposed that markingssuch as those from Pech de l’Azé, Cueva Morin and the Tagliente rock shelter (Leonardi,1988) should be interpreted as blood vessel impressions. These preliminary observations andcomparisons are more fully developed in the present paper.
Figure 1. Drawing of the Pech de l’Azé II rib: (top) depiction of all marks present on the surface; (bottom)marks interpreted as intentional by Bordes and Marshack (after Bordes, 1969).

4 . ’ .
Cueva Morin (Santander, Spain). Echegaray & Freeman (1971) reported 11 bones from theMousterian level 17 of Cueva Morin with meandering marks (Figure 2). They interpretedthese as intentional engravings, pushing back the origins of artistic behavior to the MiddlePaleolithic. Later Freeman (Echegaray & Freeman, 1973; Freeman, 1978) suggested thatportions of these marks may be natural, perhaps blood vessel impressions, that were laterintentionally modified and overmarked by humans. Echegaray (1988) stated that the naturalcauses of most of these marks have not yet been identified, although ‘‘production by humanagency is undeniable’’ for some pieces, which he states ‘‘seem most likely to be intentionallydecorative’’ (Echegaray & Freeman, 1971: Figure 74 and 1973: Plate D, here reproduced inFigure 2).
Stránska Skála. Valoch (1972, 1987) described as deliberately engraved a juvenile elephantidvertebral body found in Cave 8 of the Stránska Skála site. According to his description, the flatventral side of this sacral vertebra bears one set of seven radial grooves and two large U-shapedgrooves running obliquely from the upper to the lower edge of the bone (Figure 3). Valochnotes that none of the grooves can be considered a result of recent (i.e., excavation orpost-excavation) damage:
‘‘their hue and the rounded edges are identical with the rest of the bone surface. . . . Neither can theybe explained by the activities of predators . . . since any other vestiges documenting their gnawing orotherwise affecting by animals are missing’’ (Valoch, 1987: p. 141).
Valoch believes that the radial grooves were made by stone tools. Since the radial groovescannot be explained by any utilitarian function, he suggests that they might be interpreted asa ‘‘first attempt of aesthetic-symbolical manifestations’’. The object was recently examined byPatou-Mathis (1995), who found visual examination insufficient to diagnose the origin of the
Figure 2. Drawing of a Cueva Morin bones with meandering marks (after Echegaray & Freeman, 1971) andmacrophoto (after Marshack, 1991; reproduced with the publisher’s permission, scale absent in the original).

5
radial pattern and suggested scanning electron microscopy (SEM) analysis to solve theproblem.Bednarik (1992, 1995) accepts a non-utilitarian interpretation of the markings. Comparing
them with other putative engravings from Bilzingsleben and Prolom, he suggests that theStránska Skála radial grooves may be the early expression of a conceptual evolution of thegraphic motif of convergence from sets of unconnected lines to a more succinct pattern ofjoined lines. For him, the Stránska Skála engravings, like those from Bilzingsleben and Prolom,are recognizable as phosphene motifs (i.e., representations of luminous visual sensationsexperienced by pressing the eye-balls), comparable with those found in Australian petroglyphs,and supporting his theory that the earliest art ‘‘consists entirely of phosphene motifs’’(Bednarik, 1995: p. 614).
HolesHumanly perforated bones, often interpreted as pendants, have been claimed for at least sixMiddle Paleolithic sites: Pech de l’Azé II and Bois Roche (France), Kulna (Moravia, CzechRepublic), Repolusthöle (Austria), Bocksteinschmiede (Germany). Long bone shafts apparentlywith double or multiple perforations from Divje babe I (Slovenia) and Haua Fteah (Libya) havebeen interpreted as musical instruments. Numerous perforated animal phalanges, sometimesinterpreted as whistles, have also been reported from a number of Middle Paleolithic sites(e.g., La Quina, Combe Grenal, Bocksteinschmiede, Prolom II; Martin, 1907–1910; Wetzel &Bosinski, 1969; Stepanchuk, 1993). These have been discussed recently by Chase (1990), whoused actualistic data to support the interpretation as carnivore punctures, a hypothesispreviously put forward by Martin (1907–1910) for the majority of perforated phalanges atLa Quina. However, according to Martin, at least one reindeer phalanx, presenting twosymmetrical perforations on the posterior and anterior faces, was humanly-made.According to Taborin (1990), who has re-examined this latter object, the edges of the
perforations are sharp and angular, as in carnivore punctures, and cannot be due to drilling.Davidson (1990), also rejects the idea of human manufacture. We will discuss below the groupof perforated bones interpreted as pendants, to the exclusion of perforated phalanges (alreadydiscussed by Chase) and perforated teeth (for which we do not have adequate documentation).
Figure 3. The Stránska Skála elephant vertebra (after Valoch, 1987).

6 . ’ .
Pech de l’Azé II. This long bone fragment, found in level 4a, dated to isotope stage 5 (Grün etal., 1991) presents an incomplete hole (diameter 8·5 mm; Figure 4). It was considered ashuman-made by Bordes (1969), who stressed the regular shape of the perforation, the presenceof a flared facet on the medullar side of the hole and the apparent presence of striations dueto the drilling tool. According to Bordes (1969), the hole is incomplete because the piece wasbroken and polished by postdepositional abrasion. His diagnosis is accepted by Vincent (1993),who examined the piece and interprets the facet as a possible trace of opposite drilling. Vincentalso noticed that the cortical face of the fragment is exfoliated. Thus, she says, it is impossible
Figure 4. (a) and (b): perforated bone from layer 4(a) of Pech de l’Azé II, scale bar=1 cm; (c) and (d) SEMmicrographs showing detail of the hole surface and of the bone outer surface. Note the similarity between thetwo areas, and their parallel bone fibers revealed by etching and their resemblance to the surface ofhyena-regurgitated bones of Figure 15(b).

7
to establish if the perforation was actually done by opposite drilling since the original medullarface is not preserved.The artefactual nature of the Pech de l’Azé perforation is accepted by Harrold (1989);
Marshack (1988); Bednarik (1992) and Stringer & Gamble (1993) among others. Mellarsproposes chemical erosion as an alternative natural cause, and attributes this suggestion toChase (1990), although this latter author discusses only carnivore punctures on animalphalanges.
Kulna. This long bone fragment with a complete perforation (diameter 3·6 mm; Figure 5) wasfound in layer 7a containing a Micoquian stone industry and dated to about 45,000 BP(Valoch, 1988). Also studied by Vincent (1993), the hole is thought to be the result of oppositedrilling on the basis of the flaring of both edges.
Bois Roche. This bone fragment with a complete perforation (diameter 6 mm; Figure 5), wasdiscovered during excavation of this mall cave1 by B. Vandermeersch and was consideredanthropic by Vincent (1987, 1988a, 1993). The site has yielded a rich faunal assemblage, a fewMiddle Paleolithic artefacts, and some putative ‘‘bone tools’’, also studied by Vincent (1988a,1993). According to Delpech (in Vincent, 1993), the piece is a femoral shaft fragment of a largeungulate. Vincent observed the presence of an oblique facet on the cortical side of the holewhich she interpreted as a trace of opposite drilling. The absence of tool marks is attributed to1
Figure 5. Perforated bones from Kulna (left) and Bois Roche (right). Scale=1 cm. After Vincent, 1993.
Bois Roche, near Cognac, Charente, France is a small cave discovered in 1978, and under current excavation byP. Villa & L. Bartram (1996). At the time of the discovery the main chamber was filled by deposits almost to theceiling, its floor covered with bones and hyena coprolites. The known part of the deposit, dug to a depth of 60–70 cm,contains a large faunal assemblage, about 4500 specimens with coordinates, and several thousand small bonefragments recovered through fine-mesh water-screening. A few Levallois flakes date the top part of the deposits to theMiddle Paleolithic. Contextual evidence, i.e., very low ceiling height, abundance of coprolites and deciduous hyenateeth, combined with high frequency of gnaw marks, complete bone shafts and hundreds of small bone pieces bearingthe distinctive features of partial digestion by hyenas (circular holes, scalloped surfaces, sharp eroded edges), show thatthe excavated macrofaunal assemblage represents accumulation by hyenas using the small cave as a den.

8 . ’ .
the alteration of the bone surfaces. Both Taborin (1990) and Davidson (1990) are skeptical ofthe anthropic origin of the perforation, but in the absence of direct examination they do notsuggest alternative interpretations.
Bocksteinschmiede. Two pierced bones from the Micoquian layers of this cave (with anapproximate date within isotope stage 5) were published by Wetzel & Bosinski (1969). One ofthe two bones is a wolf metapodial, 6 cm long, with an irregular hole, approximately 3 mm indiameter, in its distal epiphysis (Figure 6). Davidson (1990) thinks that the hole is a product ofgnawing (or perhaps erosion of the bone surfaces), but Marshack (1988, 1991) considers ithuman-made. According to Marshack, the gouged area clearly visible on the distal edge of thehole on the posterior face of the bone is proof of the anthropic origin of the perforation. Heargues that gnawing produces planar, flat surfaces on bones. By contrast, the gouged area isconcave in shape. The text and the photo of rodent gnaw marks on a roe deer antler providedby him in support of his ideas indicates that Marshack (1991) does not distinguish carnivoregnaw marks (punctures, as implied by Davidson in this case) from rodent gnaw marks (parallelflat grooves).The second bone, a wolf caudal vertebra, bears a perforation near one of the epiphyses; the
object is now lost. Two more perforated bone fragments (Figure 6) were published byMarshack (1991), who interprets them as natural holes ‘‘perhaps bored by larvae’’ butnevertheless suggesting ‘‘the possibility of their use as bead or pendant’’.
Repolusthöle. The Middle Paleolithic levels of this cave (Mottl, 1951; Bednarik, 1992) haveyielded two perforated objects, interpreted as anthropic: a long bone fragment with a completeperforation near the edge of the fragment and a wolf incisor with a perforation near the apexof the root.
Divje babe I. This femur of a young cave bear with two holes on the posterior face was foundin layer 8 containing Mousterian artefact and dated to 45,100+1500–1800 (RIDDLLaboratory 745; Turk et al., 1995). Another possible hole, aligned with the first two on the longaxis of the bone, is present with only half its circumference preserved due to breakage. Turket al. (1995) suggest that it may be a musical instrument, but caution that human manufactureof the perforations is not yet proven, and that the alternative interpretation of these features ascarnivore punctures remain to be refuted.
Haua Fteah. A long bone shaft with a single perforation (about 7 mm in diameter) was found inthe Middle Paleolithic levels of Haua Fteah, Libya (McBurney, 1969). One of the shaft’sbroken edges is concave and has been interpreted as a remnant of a second hole, aligned withthe first. According to Davidson (1991), the hole is a carnivore puncture.
Analytical procedures
To check the various conflicting interpretations we have used comparative data obtained frommodern reference collections. Complementary information was provided by analog materialsfrom archaeological sites. Both kinds of materials have been submitted to morphometricand/or optical and SEM analysis. The same procedures have been followed for the putativelyengraved and perforated bone objects.

9
Figure 6. Top: distal epiphysis of a perforated wolf metapodial from Bocksteinschmiede, Germany. Bottom:two perforated bone fragments from Bockstein. After Marshack, 1991 (no scale in the original publication;reproduced with the publisher’s permission).

10 . ’ .
GroovesWe examined bones of large and medium-sized mammals in the modern osteologicalcollection of the Quaternary Institute in Bordeaux in search of surface features mimickinganthropic engravings. Grooves of this nature have been observed on some bovid bones, suchas ribs, a sacrum, and a humerus.In search of analogous archaeological materials with natural grooves we have examined all
faunal remains from two French Paleolithic sites. The large faunal assemblage of Bois Roche,an Upper Pleistocene hyena den (see above) provided four fragments of long bones (two bovidhumeral shafts and two shaft fragments) with grooves. Four more bones with similar grooveshave been found by us in the fauna from Pech de l’Azé II, the site which has yielded thewell-known ‘‘engraved’’ rib studied by Bordes (1969) and Marshack (1977). Our Pech de l’AzéII sample consists of three bovid ribs from layer 9 and one bovid sacrum from layer 7. Anotherpiece, a bovid humeral shaft fragment from Roc de Marcamps, a Magdalenian rock shelter(Gironde, France; Lenoir, 1983), was kindly given to us by Michel Lenoir.Bones from the osteological collection of the Quaternary Institute and from the Paleolithic
sites were examined with a reflected light microscope; selected areas were replicated withProvil L elastomer (Bayer, Leverkusen, Germany). Positive casts, made in RBS resin (T2LChimie, Chalabre, France), were mounted on metal supports, coated with gold-palladium andobserved with a SEM 840A Jeol (Bromage, 1987; d’Errico, 1988). The ‘‘engraved’’ rib fromPech de l’Azé II, kept in the Prehistory Museum at Les Eyzies, France, was submitted to thesame treatment; three areas of the rib internal face were replicated and observed withthe SEM.
HolesOur reference collection consists of 172 partially-digested bones collected by Sutcliffe (1970)from spotted hyena dens in Africa, now kept in the Natural History Museum in London(Table 2). As indicated by Sutcliffe (1970) (see also Kitching, 1963; Kruuk, 1972; Brain, 1981;Horwitz, 1990), adult hyenas frequently regurgitate indigestible materials; piles of bone, toothand hoof fragments packed with masses of hair were found by him at den entrances. Accordingto Sutcliffe (1970) distinctive features of regurgitated bones are: scalloped bone surfaces,knife-sharp edges between two eroded faces and small circular holes. Not all these features arepresent on all bones.Complementary information is provided by materials found in Pleistocene hyena dens. The
richest assemblage is provided by the site of Bois Roche where recent excavations have yielded
Hyena-regurgitated bones collected by Sutcliffe (collection kept inthe Natural History Museum in London)
Den locationNo. of bones
collected in densNo. of perforated
bones
Ngorongoro Crater (Tanzania) 89 17Alemeya (Ethiopia) 36 8Hwange Natural Park (Zimbabwe) 24 4Lake Abiata (Ethiopia) 18 0Queen Elizabeth Park (Uganda) 3 2Mt Elgon (Kenya) 2 1Total 172 32
Table 2

11
more than 3500 bones and bone fragments recorded with Cartesian coordinates and about30,000 bone splinters recovered through fine-mesh water screening. We have examined theentire assemblage and identified more than 400 fragments showing perforations and a varietyof surface modifications suggestive of partial digestion and regurgitation by hyenas (for otherkinds of bone modifications by hyenas see Sutcliffe, 1970; Klein, 1975; Richardson, 1980;Brain, 1981; Horwitz & Smith, 1988; Cruz-Uribe, 1991; Lyman, 1994).The following variables were recorded for both modern and Pleistocene samples: maximum
length and thickness of the bone fragments, type of bone tissue (compact or spongy), numbersof holes per fragment, and hole morphology (with complete or incomplete circumference).Hole diameters were recorded with a digital caliper to a precision of 0·01 mm. For comparisonwe measured diameters of nutrient foramina on Bois Roche bones of large size and no tracesof digestion.Characteristics of the Pleistocene bone surfaces were studied with optical and scanning
electron microscopes, using replicas. SEM analysis of bones regurgitated by hyenas fromSutcliffe’s collection was carried out at a low voltage (15 kV) on the uncoated originals. Moldsof two specimens clean enough to be replicated without removing adhering hair were madeusing the same procedures described above and observed under SEM.The ‘‘perforated’’ bone from Pech de l’Azé II (Bordes, 1969), kept in the National Museum
of Prehistory at Les Eyzies, France, and the Bois Roche specimen (Vincent, 1988a), kept in theQuaternary Institute in Bordeaux, have also been submitted to microscopic analysis. Publishedphotos and descriptions of the ‘‘perforated’’ piece from Kulna, the specimen diagnosed ashuman-made by Vincent (1993) have been used to record variables such as length, holediameter, hole morphology, and presence of diagnostic features of hyena-regurgitated bones.
Results
GroovesBones from the sites of Bois Roche and Roc de Marcamps (Figure 7) show the samemeandering and branching pattern described by Marshack (1977) and Freeman (1978) fromPech de l’Azé and Cueva Morin (Figures 1–2). These patterns are formed by double and triple(occasionally up to five) parallel grooves between 500 µm and 1 mm apart from the main one;the central groove is always deeper and wider. Groove segments can be rectilinear orcurvilinear and join either obliquely or perpendicularly.When observed with a SEM [Figure 8(b), (c)] these grooves show rounded edges and
U-shaped sections; their walls show a number of funnel-like openings of different sizes. In somecases the grooves appear to tunnel under the surface of the bone, as also observed by Marshack(1991) on the Morin bones. Our photos indicate that the grooves merge perfectly into the bonesurfaces and that they are natural features. It is clear that grooves should be interpreted asblood-vessel impressions; the funnel-like openings correspond to capillaries. This interpretationis confirmed by the findings of similar grooves on the surface of modern bones of variousspecies from the osteological reference collection in the Quaternary Institute at Bordeaux.Figure 8(a) shows the impression of a vascular groove on the surface of a modern bovidhumerus displaying the same features (rounded edges, U-shaped section, funnel-like openings)observed on the archaeological specimens. The same kind of grooves (occurring singly or ingroups) have been observed on a large sample of human tibiae from Anglo-Saxon cemeteriesby Wells (1963). He noted that 35% of his sample presented single channels, 40% had doublechannels, and the rest had three or more channels. He explains the single channels as marks

12 . ’ .
Figure 7. Meandering grooves on bovid humeral shafts from the archeological sites of Bois Roche [(a), (c)]and Roc de Marcamps (b). Scale=1 cm.

13
Figure 8. SEM micrographs of blood-vessel impressions observed on the surface of bovid humeri from theosteological reference collection of the Quaternary Institute of Bordeaux (a) and from the archeological siteof Bois Roche [(b), (c)]. (a) Scale bar=100 µm; (b) scale bar=1 mm; (c) scale bar=100 µm).

14 . ’ .
caused by a single vessel, while multiple channels would be due to an artery accompanied byone or two veins.Microscopic analysis of the internal face of the Pech de l’Azé rib reveals two different classes
of marks. The first group is composed of subparallel, curvilinear grooves [Figures 9(a) and10(a)] forming an interconnected pattern. In close-up view, these grooves possess roundededges, a U-shaped section and several small openings [Figure 9(b)], quite similar to the naturalfeatures in Figure 8 described as vascular grooves.The second group comprises straight or slightly-curved sharp striations, of different widths,
lengths and orientation, isolated or organized in parallel sets [Figure 10(b)]. These striations,which cover the entire surface of the bone, are traces of heavy mechanical abrasion which hasobliterated most microscopic anatomical features of the bone surface. In Figure 9(c) a sharpdouble horizontal striation crosses parallel smooth grooves; the intersection of the twomarkings clearly indicates that the thin slicing or trampling mark overlays the vascular groove,partially obliterating its walls.Note also that none of the grooves of Pech de l’Azé show the characteristics of experimental
engravings produced by a flint point, such as internal striations, sharp edges and angularsections (Shipman, 1981; d’Errico & Cacho, 1994). Three other bovid ribs from layer 9 of thesite show, on their internal face, vascular grooves identical to those on the ‘‘engraved’’ rib.These bones were not noticed by Bordes (1969).In the faunal collection from Pech de l’Azé we have found a bovid sacrum showing three
sets of grooves on the ventral face of the first and second sacral vertebrae (Figure 11). Two ofthe sets have grooves arranged in a radial pattern, converging toward the fusion line betweenthe first and second sacral vertebrae. This pattern is clearly analogous to the one described onthe Stránska Skála sacral vertebra, in terms of radial disposition of the grooves, their lengthrelative to bone size, and anatomical position on the ventral face of the bone. SEM inspectionof the Pech de l’Azé sacrum reveals the same feature described on the Bois Roche and modernbone sample: grooves with rounded, U-shaped sections, smooth edges, branching and capillaryopenings (Figure 12).In conclusion, it is clear that the engravings on the Pech de l’Azé II rib, and probably those
of Morin and Stránska Skála, are vascular grooves. They are not the result of taphonomicprocesses, as recently suggested by Marshack (1991), since they are present both in modernand in well-preserved prehistoric materials.
HolesAmong the hyena-regurgitated bones collected by Sutcliffe (1970), 32 of 172 (18·6%) showeither single or multiple holes with regular edges and circular or elliptical shapes; some of theholes have incomplete circumferences (Figure 13). On thinner specimens borders aretransparent, due to the action of the gastric hydrochloric acid which results in loss of mineralmatter [Figure 14(c)]. Recent experiments carried out on rodent bones, simulating digestion bypredators, suggest that hole formation is due to the combined action of acids and enzymespresent in the stomach (Denys et al., 1995).The bone fragment surfaces are generally corroded and pitted. At microscopic scale the
surfaces show patches of bowl-shaped microconcavities [Figure 15(a)]; their diameters can beas much as 2 mm. Etching caused by the hyena’s gastric acids reveals the fibrous bonestructure; microscopic fissure lines can be seen parallel to the bone fibers [Figure 15(b)].At Bois Roche, 431 small bone fragments show single and multiple perforations, either
complete or incomplete; hole shapes and edge morphology are almost identical to those in

15
Figure 9. Pech de l’Azé II rib. (a) detail showing an interconnected pattern of curvilinear grooves; (b)macrophoto of a groove with U-shaped section and openings of capillaries indicated by arrows; (c) heavilyabraded vascular grooves (vertical) overlain by a double striation (horizontal) probably produced bytrampling. (a) Scale=1 cm; (b) scale=500 µm; (c) scale=1 mm.

16 . ’ .
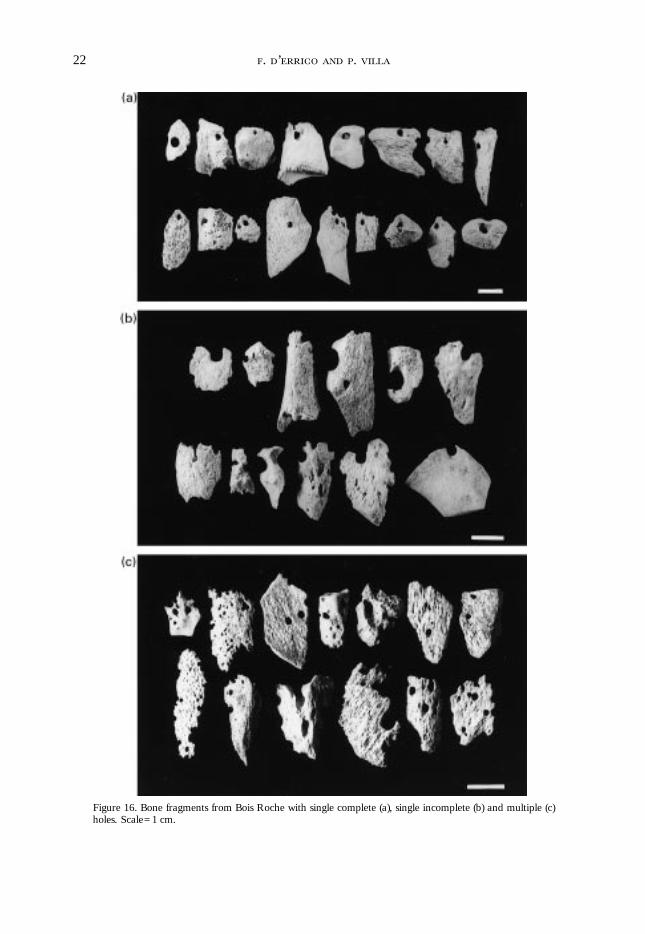
Sutcliffe’s sample [Figure 14(a), (b)]. Most of these pieces show the same corroded and pittedsurfaces (Figure 16); in several specimens these surfaces appear smoothed, perhaps bydepositional or postdepositional abrasion [Figure 15(c)]. In the Bois Roche sample, as inSutcliffe’s hyena den material, pieces have thin, sharp edges, meeting at acute angles.Many of the larger holes appear to be enlargements of pre-existing foramina attacked by
gastric acids. In fact, in a few pieces, identifiable to anatomical region, the hole is locatedwhere a foramen is expected. Furthermore, in several cases the perforation is oblique withrespect to the long-axis of the bone, a common feature of long bone nutrient foramina(Brookes, 1971).Quantitative analysis of the Bois Roche and modern regurgitated bones confirms the strong
similarity of the two samples. Perforated fragments with single holes are most common; thetwo samples show the same frequency distributions [Figure 17(a), (b)]. Hole diameters
Figure 10. SEM micrographs of the Pech de l’Azé rib, showing (a) two ‘‘engraved’’ curved lines; in (b) the‘‘engraved’’ lines are crossed by sets of short and sharp striations indicating mechanical abrasion.

17
vary between 0·3 and 9·5 mm in Sutcliffe’s sample, and between 0·4 and 9·1 mm at BoisRoche [Figure 17(c), (d)]. In both samples the large majority of the values range between 0·5and 4 mm. Figure 17(e), (f) shows that complete holes generally have smaller diameters thanincomplete holes. This seems to be due to the fact that incomplete holes are formed near theedge of the piece where the border is thinner and less resistant to the attack of acid. Note thatthe great majority of the holes in the Bois Roche sample have diameters smaller than those ofnutrient foramina measured on bones of the same assemblage (Figure 18). This suggests that,with the exception of those of larger size, most holes are not altered nutrient foramina.All fragments are small and lengths generally between 10 and 60 mm [Figure 17(g), (h)];
in this diagram Sutcliffe’s sample includes all regurgitated bones, whether perforated or not.The largest specimen collected by Sutcliffe is 11·8 mm; this piece is not included in ourdiagram to facilitate visual comparison with the Bois Roche length diagram. Thickness isgenerally less than 2 cm in both samples, with most examples falling in the 0·5–10 mm class[Figure 19(a), (b)].At Bois Roche most holes occur in compact bone [Figure 19(d)]. The same predominance
of compact bone can be observed among modern regurgitated bones, whether perforated ornot [Figure 19(c)]. Compact bone fragments are generally thinner than those whose spongybone is present [Figure 19(e), (f)]. The lower proportions of spongy bone fragments and theirrelatively greater thickness appear to be due to differential preservation because thin spongyfragments have a lower probability of survival. In both samples single holes occur mostfrequently in compact bone, which must be a consequence of greater resistance of compactbone to chemical action [Figure 19(g), (h)].On the basis of comparisons with Sutcliffe’s hyena-regurgitated samples we conclude that
the Bois Roche perforated bones are also hyena-regurgitated fragments. That these bones havebeen regurgitated by Pleistocene hyenas, that they have been submitted to postdepositionaland other long-term processes (such as sediment compaction and collagen loss), and that they
Figure 11. Bovid sacrum from layer 7 of Pech de l’Azé II with sets of vascular grooves arranged in a radialpattern. The arrow points to the area shown in Figure 12.

18 . ’ .
were deposited in a limestone cave makes the Bois Roche sample particularly useful forcomparisons with archaeological materials from limestone caves frequented by Pleistocenehyenas.It is now possible to say that features, typical of hyena-regurgitated bones are present on
archeological specimens previously interpreted as humanly perforated. Corroded surfaces canbe seen on the Kulna, Bois Roche (Vincent, 1988a) (Figure 5), Pech de l’Azé II [Figure 4(c),(d)] and Bocksteinschmiede (Figure 6 top); pits and/or bowl-shaped microconcavities are alsovisible on the Bois Roche and Pech de l’Azé specimens. Thin, sharp edges are present on allthe specimens. Holes on the Bois Roche and Pech de l’Azé bones have cylindrical sections withstraight edges, like bones digested by hyenas, and unlike Upper Paleolithic and experimentalperforations produced by rotation of a flint point. The latter pieces have conical or biconicalsections. None of the putatively perforated pieces show the striations and scraping marks
Figure 12. SEM micrographs of a bovid sacrum from layer 7 of Pech de l’Azé II. Notice the branchingpattern in (a); the arrow points to the area enlarged in (b).

19
associated with the use of stone tools, as documented experimentally and by Upper Paleolithicperforated objects (Stordeur, 1979; d’Errico et al., 1983). The parallel striations purportedlyproduced by a drilling tool (‘‘stries parallèles creusées par le silex tournant’’) noted by Bordeson the Pech de l’Azé specimen are the result of chemical alteration of the bone fibers, asdemonstrated by the fact that the bone outer surface and the hole inner surface have identicalmorphology [Figure 4(c), (d)]. The same morphology can be observed on modern and fossilhyena-regurgitated bones (Figure 15).The oblique flared facet on the external face of the Bois Roche specimen is clearly indicative
of a nutrient foramen entrance point. The incomplete hole of Pech de l’Azé was describedby Bordes as broken after manufacture and utilization. However, the bone edges adjacentto the hole meet at an acute angle, with no evidence of fracture, bearing instead a very
Figure 13. (a) Bones of juvenile wildebeest from Ngorongoro Crater (Tanzania), partly digested andregurgitated by spotted hyenas (courtesy of A. Sutcliffe), scale=1 cm. Notice the hair inside the largerspecimen. The specimen at the top has two adjacent incomplete holes. (b) Bone fragments regurgitated byhyenas from Queen Elizabeth Park (Uganda), with single, multiple complete and incomplete (at left) holes;scale bar in cm.

20 . ’ .
close resemblance to the sharp edges of incomplete holes on hyena-regurgitated fragments[Figure 16(b)].The length and hole diameters of all these specimens fall in the size range of hyena-
regurgitated bones [Figure 17(d), (h)]. The single large perforations observed in thesespecimens is not proof of their human manufacture, since single holes are very commonamong hyena-regurgitated bones [Figure 17(a), (b)]. The position of the hole at one end of thebone, the regular morphology of the hole, and the symmetrical shape of the piece, are allcharacters that can be found together in some of the hyena-regurgitated specimens, such as thepiece depicted in Figure 20 and several in Figure 16(a).The Bocksteinschmiede perforated wolf metapodial requires a separate discussion. The hole
morphology is irregular and does not resemble our reference material. The opposingperforations on the posterior and anterior face of the bone and their position at the articularend of the bone are clearly suggestive of carnivore punctures (as noted by Davidson, 1990). Incontrast to Davidson, we believe that the erosion of the bone’s distal end, described by him aspostdepositional, is, at least in part, the effect of partial digestion of a previously puncturedbone. Chase (1990: Figure 5) illustrates a cervid phalanx punctured near the distal end andsubsequently digested by a canid. The action of the gastric acid has resulted in markedalteration of the compact bone around the hole exposing the spongy tissue below. The holeand erosion observed on the Bocksteinschmiede piece closely resemble those on the objectillustrated by Chase. The gouged area near the hole edge, proof of anthropic origin for
Figure 14. (a) SEM micrograph of a marginal hole on a bone fragment regurgitated by a spotted hyena,Sutcliffe’s sample. (b) Marginal hole on a regurgitated bone fragment from Bois Roche, (c) edge of a thinbone fragment regurgitated by spotted hyena showing a transparent border, due to the action of thehydrochloric acid, Sutcliffe’s sample [(a)–(b), scale=1 mm, (c) scale=200 µm].

21
Figure 15. (a) Corroded surface on a regurgitated bone fragment from a hyena den near Alemeya (Ethiopia);macrophoto of a resin replica seen in transmitted light. (b) corroded surface on a hyena-regurgitated bonefragment from Sutcliffe’s collection, seen with a SEM; scale=500 µm. (c) SEM view of the corroded surfaceof a bone fragment with multiple holes from Bois Roche.
22 . ’ .
Figure 16. Bone fragments from Bois Roche with single complete (a), single incomplete (b) and multiple (c)holes. Scale=1 cm.

23
0
Hole diameter (mm)
Nu
mbe
r of
hol
es
1
10
2 3 4 5 6 7
Spotted hyaenas(Sutcliffe's sample)
n = 50
(e)8
6
4
2
0
Hole diameter (mm)
Nu
mbe
r of
hol
es
1 2 3 4 5 6 7
Bois Rochen = 937
(f)200
100
0
Specimen length (mm)
Nu
mbe
r of
spe
cim
ens
10
40
20 30 40 50 60 70 80 90
Spotted hyaena(Sutcliffe's sample)
n = 172
(g)
30
20
10
0
Specimen length (mm)
Nu
mbe
r of
spe
cim
ens
10
120
20 30 40 50 60 70 80 90
Bois Rochen = 430
(h)
80
20
100
60
40Bois Roche(Vincent 1988)
KulnaPech de l'Azé II
0
Hole diameter (mm)
Nu
mbe
r of
hol
es
1
12
2 3 4 5 6 7 8 9
Spotted hyaena(Sutcliffe's sample)
n = 50
(c)10
8
6
4
2
0
Hole diameter (mm)
Nu
mbe
r of
hol
es
1
400
2 3 4 5 6 7 8 9
Bois Rochen = 937
(d)
300
200
100Bois Roche
(Vincent 1988)
KulnaPech de l'Azé II
0
Number of holes
Nu
mbe
r of
spe
cim
ens
1
10
2 3 4 5 6 7 8 9 10
Spotted hyaena(Sutcliffe's sample)
n = 32
(a)
0
Number of holes
Nu
mbe
r of
spe
cim
ens
1
100
2 3 4 5 6 7 8 9 10
Bois Rochen = 429
(b)200
Figure 17. Frequency distributions of selected variables in the Bois Roche and Sutcliffe’s samples. (e) and (f)(.) Complete holes; (/) incomplete holes.

24 . ’ .
Marshack (1991), is also present on Chase’s object. Thus, the Bocksteinschmiede morphologyis better interpreted as a consequence of the combined effects of gnawing, partial digestion andpostdepositional erosion (which has played no role in modifying Chase’s specimen). It isimportant to note that carnivore activity is documented at the German site by two otherperforated bone fragments, described by Marshack as ‘‘perhaps bored by larvae’’, and possiblyused as pendants. These two fragments actually display all of the features of regurgitatedbones: small size, corroded surfaces, pits, and thin sharp edges (Figure 6 bottom).The Repolulsthöle perforated long bone fragment is known to us only through the drawing
published by Bednarik (1992) and a short description, with no details on possible manufacturetraces. We are, therefore, not really in a position to suggest an alternative interpretation ofthis object.The Divje babe I perforated bear femur cannot easily be interpreted as a hyena-regurgitated
bone due to its large size (11 cm), and to the hole diameters (about 8 mm), which put thisspecimen at the upper limit of known distributions for hyena-regurgitated bones. Thehypothesis of carnivore punctures was not excluded by the authors (Turq et al., 1995).Although they did not recognize gnaw marks on this piece, the crenulated morphology of onebroken edge of the bone suggests to us carnivore gnawing. The Haua Fteah bone has beenillustrated with a photo by Davidson (1991: Figure 1). No traces of stone tool marks can bedetected in this photo. The bone surface, however, is heavily exfoliated. Davidson’sinterpretation that this is a carnivore puncture is supported by the morphology of the internalwalls of the hole, which exhibits splintered and depressed bone lamellae. This suggests to himthe action of a carnivore tooth. Although this morphology could also be the result ofpostdepositional exfoliation of a pre-existing hole, we agree with Davidson that there is noevidence of human intervention.In summary, the size of the bone fragments, the presence of corroded surfaces associated
with pits and bowl-shaped microconcavities, the occurrence of thin edges meeting at acuteangles, and the diameters and general morphology of the holes, demonstrate that pieces like
0
Hole diameter (mm)
Nu
mbe
r of
hol
es
12
1
Bois RocheAnatomical holes
n = 40
10
8
6
4
2
92 3 4 5 6 7 8
Figure 18. Range of diameters in anatomical holes in the Bois Roche faunal assemblage.

25
0
Number of holes
Nu
mbe
r of
spe
cim
ens
1
12Spotted hyaenas
(Sutcliffe's sample)n = 172
(g)10
8
6
4
2
2 3 4 5 6 7 8 90
Number of holes
Nu
mbe
r of
spe
cim
ens
1
100
Bois Rochen = 432
(h)
2 3 4 5 6 7 8 9
0
Specimen thickness (mm)
Nu
mbe
r of
spe
cim
ens
1
30
2 3 4 5 6 7
Spotted hyaenas(Sutcliffe's sample)
n = 172
(e)
20
10
8 9 10 11 120
Specimen thickness (mm)
Nu
mbe
r of
spe
cim
ens
1
100
2 3 4 5 6 7
Bois Rochen = 431
(f)
8 9 10 11 12
80
60
40
20
0
Bone type
Nu
mbe
r of
spe
cim
ens
150
Compact
Spotted hyaenas(Sutcliffe's sample)
n = 172
(c)
100
50
Compact and spongy
Spongy0
Bone type
Nu
mbe
r of
spe
cim
ens
400
Compact
Bois Rochen = 432
(d)
300
200
Compact and spongy
Spongy
100
0
Specimen thickness (mm)
Nu
mbe
r of
spe
cim
ens
120
5 10 15 20
Bois Rochen = 431
(b)100
80
60
40
20
0
Specimen thickness (mm)
Nu
mbe
r of
spe
cim
ens
40
5 10 15 20
Spotted hyaena(Sutcliffe's sample)
n = 172
(a)
30
20
10
Figure 19. Frequency distributions of selected variables in the Bois Roche and Sutcliffe’s samples. (e)–(f) (.)Compact; ( ) compact and spongy; (/) spongy.

26 . ’ .
Kulna, Pech de l’Azé and Bois Roche are not intentionally perforated, but are bone fragmentspartially digested by hyenas.
Discussion
Natural causes have already been suggested for grooves in bones such as those from Pech del’Azé II and Morin. As noted above, Binford (1981) thought that they could have been caused
Figure 20. Pseudo-pendant from Bois Roche. Top: scale=1 cm; bottom: SEM micrograph of the hole edgeshowing regular morphology without depressed margins.

27
by plant roots etching the bone surfaces. Chaix (in Baud, 1986: p. 143) and Morel (1986)report grooves with rounded sections produced by the action of carnivorous mollusks on bonesurfaces. None of the grooves on the specimens discussed here can be attributed to thesecauses. Figure 9(b) shows capillary openings in the groove which are not consistent with rootmarks or marks produced by gastropods. SEM photos published by Andrews (1990) of rootmarks on a cow’s rib clearly show smooth, uniform walls with no openings inside the channel.In the Natural History Museum in London we have observed modern bone pieces collected bySutcliffe with roots still adhering to the bone surface. These grooves also have smooth wallswithout openings.Shipman & Rose (1984) have shown that vascular grooves on bovid distal humeri can mimic
cut marks. On human tibiae these grooves have sometimes been interpreted as a pathologicalphenomenon or as traces of surgical operation done with stone tools (Møller-Christensen, 1955and Wakefield & Dellinger, 1937 in Wells, 1963). Our work shows that vascular grooves, thatcan be found in other anatomical regions of humeri, on other kinds of bone (i.e., ribs,vertebrae), and with much more complex patterns, have been confused with deliberateengravings.Freeman (1978) has suggested that the Cueva Morin grooves might, in fact, be vascular
grooves intentionally modified and overmarked by humans. This statement can be acceptedonly when a microscopic analysis documenting the occurrence of tool marks overmarking thegrooves is undertaken.In addition to those already discussed above, a number of natural processes may produce
holes on bones. Perforations by mushrooms and bacteria, experimentally reproduced byseveral authors, and identified on fossil bone (Baud, 1986), are too small (between 1 and10 µm) to be of relevance for this paper. Beetles of the genus Dermestes, in their larval state, boreinto a variety of organic materials (Hinton, 1945; Jodry & Stanford, 1992) producing holes6 mm in diameter. Even larger holes (8–10 mm) are reported by Kitching (1980) andattributed to carrion beetles. These kinds of hole are produced in long bones in order to usethe inner cavity as a pupation chamber. Only bones thus perforated and postdepositionallybroken into flat fragments could mimic pieces studied in this paper. More importantly, thearcheological pieces show features such as corroded and pitted surfaces and thin edges whichclearly point to the action of hyenas.
Final remarks
Although we have not been able to examine all putative engraved and perforated bones ofLower and Middle Paleolithic age, our survey seems to indicate a difference between these twocategories of objects. Some of the bones with either notches or single stroke lines illustrated byphotos appear to be strong candidates for being true human-made artefacts, while none of thepublished so-called perforated bones appear to possess convincing attributes of humanmanufacture. We are not denying the possibility of non-utilitarian or symbolic behavior in theLower and Middle Paleolithic. However, we have shown that some of these pieces have beenmisinterpreted, and cannot be used as evidence in favor of the emergence of symbolic behaviorin early times.We should underline the fact that our survey is limited to only one category of possible
non-utilitarian behavior. Several stone objects, reported from Lower and Middle Paleolithicsites (e.g., from Bacho Kiro, Temnata, Quneitra: Marshack, 1976, 1995; Cremades et al.,1995) appear, on the basis of published photos, to bear deliberate engravings. Criteria we have

28 . ’ .
developed for the interpretation of marked bones cannot be effectively employed to verifyengravings on stone. However, the general procedures followed here using actualistic andexperimental data, can provide firmer foundations for arguments concerning: (1) the humanorigins of the marks, (2) the technical behavior documented (e.g., tool type, stroke directions,chronological order of markings), and (3) the significance of the behavior with respect to theproblem of the origin of symboling.We have gained important insights by systematic examination of the assemblages from Pech
de l’Azé II and Bois Roche, two sites that have provided some of the objects discussed here.Visual inspection of the bone pieces in search of similar features, and knowledge of theassemblage depositional context are essential elements to consider in understanding the natureand the significance of apparently exceptional pieces. In contrast, the usual proceduresfollowed by scholars in describing ‘‘art’’ objects has been to concentrate attention on theseunique findings, without systematic analysis of the rest of the assemblage. A comprehensiveoverview of the assemblage is necessary to eliminate the possibility that the piece, far frombeing an idiosyncratic example of a rare behavior, is only an individual within a population ofsimilar but less striking phenomena.A clear understanding of the problem of art origins has been made difficult by the idea that
proof will come from the accumulation of individual cases. Since most of the cases consist ofobjects of unverified human manufacture or of unverified behavioral significance, theirquantity should not be considered as an element of proof (Bednarik, 1992, 1995).An aspect we have not treated, but one that should always be carefully considered prior to
acceptance of a piece as documenting early non-utilitarian behavior, is the stratigraphicprovenience and cultural attribution of the object. The small size of many of these objects andthe fact that sites often contain overlying Upper Paleolithic levels, are factors not to beoverlooked. Trampling experiments (Villa & Courtin, 1983; Gifford-Gonzalez et al., 1985)suggests that vertical displacement of small objects is a likely occurrence in sedimentsunconsolidated at the time of occupation, causing mixing of materials belonging to twoseparate levels. At several stratified sites refitting has demonstrated vertical displacement ofmaterials up to 1 m, and across distinct excavation levels (e.g., Terra Amata, Lazaret, Hortus,Grotte Vaufrey, Lunel Viel; Villa, 1982, 1983; Patou, 1984; Geneste, 1988; Le Grand, 1994).In sum, we believe that a detailed, validating analysis of each object interpreted as an early
manifestation of non-utilitarian behavior is a necessary prerequisite to any discussion of theirsignificance for the evolution of human cognitive abilities.
Acknowledgements
This research was made possible in part by grants from the L. S. B. Leakey Foundation andthe French Ministry of Culture to Paola Villa for the excavation and analysis of the Bois Rochematerials. We owe a special debt of gratitude to Anthony Sutcliffe for his permission to studyhis collection of hyena-regurgitated bones, and for very helpful discussions during thepreparation of this work. Our thanks also go to Jerry Hooker, Curator of VertebratePaleontology in the Natural History, Museum of London. We thank Jean-JacquesCleyet-Merle, Chief Curator of the Les Eyzies Museum, for facilitating the study of the Pechde l’Azé II rib and perforated bone. The Pech de l’Azé II faunal assemblage was studiedthanks to the kindness of Françoise Delpech. The Roc de Marcamps and Camiac material wasgenerously given to us by Michel Lenoir. We thank three JHE referees and Terry Harrison forvery useful, critical comments.

29
References
Andrews, P. (1990). Owls, Caves and Fossils. Chicago: University of Chicago Press.Bahn, P. (1996). New developments in Pleistocene art. Evol. Anthrop. 4, 204–215.Bar Yosef, O. (1988). Evidence for Middle Paleolithic symbolic behaviour: a cautionary note. In (O. Bar Yosef, Ed.)L’Homme de Neandertal, Vol. 5, pp. 11–16. Liège: ERAUL.
Baud, Ch. A. (1986). Altérations osseuses post-mortem d’origine fongique ou bacterienne. In (H. Duday & Cl. Masset,Eds) Anthropologie Physique et Archéologique, pp. 135–144. Paris: CNRS.
Bednarik, R. G. (1992). Palaeoart and archaeological myths. Cambridge Archaeol. J. 2, 27–57.Bednarik, R. G. (1994). Art origins. Anthropos 89, 169–180.Bednarik, R. G. (1995). Concept-mediated marking in the Lower Palaeolithic. Curr. Anthropol. 36, 605–616.Binford, L. R. (1981). Bones, Ancient Men and Modern Myths. New York: Academic Press.Bordes, F. (1969). Os percé moustérien et os gravé acheuléen du Pech de l’Azé II. Quaternaria 11, 1–5.Bordes, F. (1972). A Tale of Two Caves. New York: Harper & Row.Brain, C. K. (1981). The Hunters or the Hunted? Chicago: University of Chicago Press.Bromage, T. G. (1987). The scanning microscopy replica technique and the recent applications to the study of fossilbone. Scan. Microsc. 1, 607–613.
Brookes, M. (1971). The Bloody Supply of Bone. London: Butterworths.Byers, A. M. (1994). Symboling and the Middle-Upper Palaeolithic transition: a theoretical and methodologicalcritique. Curr. Anthropol. 35, 369–399.
Capitan, L. & Peyrony, D. (1921). Les origines de l’art à l’Aurignacien moyen-nouvelles fouilles à La Ferrassie.Rev. Anthropol. 1921, 92–112.
Chase, P. G. (1990). Sifflets du Paléolithique moyen (?). Les implications d’un coprolithe de coyote actuel. Bull. Soc.Préhist. Française 87, 165–167.
Chase, P. G. & Dibble, H. L. (1987). Middle Paleolithic: a review of current evidence and interpretations. J. Anthropol.Archaeol. 6, 263–296.
Chase, P. G. & Dibble, H. L. (1992). Scientific archaeology and the origins of symbolism: a reply to Bednarik.Cambridge Archaeol. J. 2, 43–51.
Cremadés, M., Laville, H., Sirakov, N. & Kozlowski, J. K. (1995). Une pierre gravée de 50 000 ans B.P. dans lesBalkans. Paléo 7, 201–209.
Cruz-Uribe, K. (1991). Distinguishing hyena from hominid bone accumulations. J. Field Arch. 18, 476–486.Davidson, I. (1990). Bilzingsleben and early marking. Rock Art Res. 7, 52–56.Davidson, I. (1991). The archaeology of language origins-a review. Antiquity 65, 39–48.Davidson, I. & Noble, W. (1989). The archaeology of perception. Curr. Anthropol. 30, 125–155.D’Errico, F. (1988). The use of resin cast for the study of use-wear. In (S. L. Olsen, Ed.) Scanning Electron Microscopy inArchaeology, pp. 155–167. British Archaeological Reports, International Series 452.
D’Errico, F. (1991). Carnivore traces or mousterian skiffle? Rock Art Res. 8, 61–63.D’Errico, F. (1995). Comment to Bednarik. Curr. Anthropol. 36, 618–620.D’Errico, F. & Cacho, C. (1994). Notation versus decoration in the Upper Paleolithic: a case study from Tossal de laRoca, Alicante, Spain. J. Archaeol. Sci. 21, 185–200.
D’Errico, F., Giacobini, G. & Puech, F. (1983). Varnish replicas: a new method for the study of worked bone surfaces.Ossa 9–11, 29–51.
Debenath, A. & Duport, L. (1971). Os travaillés et os utilisés de quelques gisements préhistoriques charentais.Mém. Soc. Archéol. Hist. Charante, pp. 183–201.
Delporte, H. (1990). L’Image des Animaux dans l’Art Préhistorique. Cahors: Picard.Denys, C., Fernandez-Jalvo, Y. & Dauphin, Y. (1995). Experimental taphonomy: preliminary results of digestion ofmicromammal bones in the laboratory. C. R. Acad. Sci. Paris, Sér. IIa 321, 803–809.
Donald, M. (1991). Origins of Modern Mind. Cambridge: Harvard University Press.Duport, L. (1973). Catalogue de l’exposition de Préhistoire de la Charente. Poitiers.Echegaray, J. G. (1988). Decorative patterns in the Mousterian of Cueva Morín. In (O. Bar Yosef, Ed.) L’Homme deNeandertal, Vol. 5, pp. 37–42. Liège: ERAUL.
Echegaray, J. G. & Freeman, L. G. (1971). Cueva Morin. Excavaciones 1966–1968. Santander: Publicationes delPatronato de la Cuevas Prehistoricas de la Provincia de Santander. n. 6.
Echegaray, J. G. & Freeman, L. G. (1973). Cueva Morin. Excavaciones 1969. Santander: Patronato de las CuevasPrehistoricas de la Provincia de Santander.
Freeman, L. G. (1978). Mousterian worked bones from Cueva Morin (Santander, Spain). A preliminary description.In (L. G. Freeman, Ed.) Views of the Past, pp. 29–51. The Hague: Mouton.
Freeman, L. G. & Echegaray, J. G. (1983). Tally-marked bone from Mousterian levels at Cueva Morín (Santander,Spain). In Homenaje al Prof. M. Almagro Basch. Vol. 1, pp. 143–147. Madrid: Ministerio de Cultura.
Geneste, J. M. (1988). Les industries de la grotte Vaufrey: technologie du débitage, economie et circulation de lamatière première lithique. In (J. Ph. Rigaud, Ed.) La Grotte Vaufrey à Cenac et Sain-Julien (Dordogne). Paleoenvironnement,Chronologie et Activités Humaines, pp. 529–533. Paris: Mémoires Soc. Préhist. Française 19.

30 . ’ .
Gifford-Gonzalez, D. P., Damrosch, D. B., Damrosch, D. R., Pryor, J. & Thunen, R. L. (1985). The third dimensionin site structure: an experiment in trampling and vertical dispersal. Am. Antiq. 50, 803–818.
Grün, R., Mellars, P. & Laville, H. (1991). ESR chronology of a 100,000-year at Pech de l’Azé II, France. Antiquity65, 544–551.
Harrold, F. B. (1989). Mousterian, Chatelperronian and Early Aurignacian in Western Europe: Continuity orDiscontinuity? In (P. Mellars & C. Stringer, Eds) The Human Revolution: Behavioural and Biological Perspectives on theOrigins of Modern Humans, pp. 677–713. Edinburgh: Edinburgh University Press.
Hayden, B. (1993). The cultural capacities of Neandertals: a review and re-evaluation. J. hum. Evol. 24, 113–146.Hinton, H. E. (1945). Monograph of the Beetles Associated with Stored Products. Vol. 1. London: British Museum.Horwitz, L. K. (1990). The origin of partially digested bones recovered from archaeological contexts in Israel.Paléorient. 16, 97–106.
Horwitz, L. K. & Smith, P. (1988). The effects of striped hyaena activity on human remains. J. Arch. Sci. 15, 471–481.Huyge, D. (1990). Mousterian skiffle? Note on a middle palaeolithic engraved bone from Schulen, Belgium. Rock ArtRes. 7, 125–132.
Iturbe, G., Fumanal, M. P., Carrion, J. S., Cortell, E., Martinez, R., Guillem, M. P., Garralda, M. D. &Vandermeersch, B. (1993). Cova Beneito (Muro, Alicante): una perspectiva interdisciplinare. Recerques del Museod’Alcoi 2, 23–88.
Jodry, M. A. & Stanford, D. (1992). Stewart’s Cattle Guard Site: an analysis of bison remains in a Folsom kill-butcherycampsite. In (D. Stanford & J. Day, Eds) Ice Age Hunters of the Rocky Mountains, pp. 101–168. Niwot: University Pressof Colorado.
Kitching, J. M. (1980). On some fossil arthropoda from Limeworks, Makapansgat. Palaeont. Afr. 23, 63–68.Kitching, J. W. (1963). Bone, Tooth and Horn Tools of Palaeolithic Man. Manchester: Manchester University Press.Klein, R. G. (1975). Paleoanthropological implications of the nonarcheological bone assemblage from Swartklip I,South-Western Cape Province, South Africa. Quat. Res. 5, 275–288.
Kruuk, H. (1972). The Spotted Hyena. Chicago: University of Chicago Press.Le Grand, Y. (1994). Approche méthodologique et technologique d’un site d’habitat du Pléistocène moyen: la grotte n. 1 du Mas de Caves(Lunel Viel, Hérault). PhD Dissertation. Université de Provence, 269 p.
Lenoir, M. (1983). La Paléolithique des Basses Vallées de la Dordogne et de la Garonne. PhD Dissertation, University ofBordeaux 1.
Leonardi, P. (1983). Incisioni musteriane del Riparo Tagliente in Valpantena nei Monti Lessini presso Verona (Italia).In Homenaje al Prof. Martin Almagro Basch, pp. 149–156. Madrid: Ministerio de Cultura.
Leonardi, P. (1988). Art paléolithique mobilier et pariétal en Italie. L’Anthropologie 92, 139–202.Lyman, L. R. (1994). Vertebrate Taphonomy. Cambridge: Cambridge University Press.Mania, D. & Mania, U. (1988). Deliberate engravings on bone artifacts of Homo erectus. Rock Art Res. 5, 91–107.Marshack, A. (1976). Some implications of paleolithic symbolic evidence for the origin of language. Curr. Anthropol. 17,276–282.
Marshack, A. (1977). The meander as a system: the analysis and recognition of iconographic units in UpperPalaeolithic compositions. In (J. Ucko, Ed.) Form in Indigenous Art: Schematization in the Art of Aboriginal Australia andPrehistoric Europe, pp. 286–317. London: Duckworth.
Marshack, A. (1988). The Neanderthals and the human capacity for symbolic thought: cognitive and problem-solvingaspects of Mousterian symbol. In (O. Bar Yosef, Ed.) L’Homme de Neandertal, Vol. 5, pp. 57–91. Liège: ERAUL.
Marshack, A. (1991). A reply to Davidson on Mania & Mania. Rock Art Res. 8, 47–58.Marshack, A. (1995). A middle Paleolithic symbolic composition from the Golan Heights: the earliest known depictiveimage. Curr. Anthropol. 37, 356–365.
Martin, H. (1907–1910). Recherches sur l’Évolution du Moustérien dans le Gisement de la Quina (Charente) Industrie Osseuse,Vol. 1. Paris: Schleicher Frères.
Martin, H. (1910). Fragment d’omoplate de bovidé avec traits gravés intentonnels. Bull. Soc. Préhist. Française 7,40–42.
McBurney, C. B. M. (1969). The Haua Fteah (Cyrenaica) and the Stone Age of the southeast Mediterranean. Cambridge:Cambridge University Press.
Mellars, P. (1989). Major issues in the emergence of modern humans. Curr. Anthropol. 30, 349–385.Mellars, P. (1996). The Neanderthal Legacy. Princeton: Princeton University Press.Morel, Ph. (1986). Quelques polis naturels d’apparence trompeuse. Artefacts 3, 43–45.Mottl, M. (1951). Die Repolust-Höle bei Peggau (Steiermark) und ihre eiszeitlichen Bewohner. Archaeologica Austriaca8, 1–78.
Patou, M. (1984). Contribution à l’étude des mammifères des couches supérieures du Lazaret (Nice): méthodes et résultats paléontologiqueset palethnographiques. PhD Dissertation, Université Paris VI.
Patou-Mathis, M. (1995). Etude préliminaire de certaines pièces osseuses de Stránska Skála (Moravie) présentantdes stigmates d’intervention humaine. In (R. Musil, Ed.) Stránska Skála Hill, pp. 169–176. Brno: MoravianMuseum.
Richardson, P. R. K. (1980). Carnivore damage to antelope bones and its archaeological implications. Paleont. Afr. 23,109–125.

31
Shipman, P. (1981). Application of scanning electron microscopy to taphonomic problems. In (A. M. Cantwell, J. B.Griffin & N. Rothschild, Eds) The Research Potential of Anthropological Museum Collections, pp. 357–385. Ann. N. Y. Acad.Sci. 276, New York.
Shipman, P. & Rose, J. (1984). Cutmark mimics on modern and fossil bovid bones. Curr. Anthropol. 25, 116–117.Simek, J. F. (1992). Neanderthal cognition and the Middle to Upper Palaeolithic transition. In (G. Braur & F. H.Smith, Eds) Continuity or Replacement. Controversy in Homo sapiens Evolution, pp. 231–245. Rotterdam: Balkema.
Stepanchuk, V. N. (1993). Prolom II, a Middle Paleolithic site in the Eastern Crimea with non-utilitarian boneartefacts. Proc. Prehist. Soc. 59, 17–37.
Stordeur, D. (1979). Les aiguilles à Chas au Paléolithique. Paris: CNRS.Stringer, C. & Gamble, C. (1993). In Search of the Neanderthals. New York: Thames & Hudson.Sutcliffe, J. A. (1970). Spotted hyena: crusher, gnawer, digester and collector of bones. Nature 227, 1110–1113.Taborin, Y. (1990). Les prémices de la parure. In (C. Farizy, Ed.) Paléolithique Moyen Recent et Paléolithique Supérieur Ancienen Europe, pp. 335–224. Nemours: Musée de Préhistoire d’Ile de France.
Turk, I., Dirjec, J. & Kavur, B. (1995). Ali so v sloveniji nasli najstarejse glasbilo v europi? (The oldest musicalinstrument in Europe discovered in Slovenia?). Razprave IV. razreda SAZU (Ljubliana) 36, 287–293.
Valoch, K. (1972). Gab es eine altpaläolithishe Besiedlung der Stránská skála? In (R. Musil, Ed.) Stránská Skála I,pp. 199–204. Brno: Anthropos 20.
Valoch, K. (1987). The early Palaeolithic site Stránská skála I. Near Brno (Czechoslovakia). Anthropologie 25, 125–142.Valoch, K. (1988). Le Taubachien et le Micoquien de la grotte de Kulna en Moravie (Tchecoslovaquie). In (M. Otte,Ed.) L’Homme de Néandertal, Vol. 4. La Technique. pp. 205–217. Liège: ERAUL.
Valoch, K. (1994). Archaeology of the Neanderthalers and their contemporaries. In (S. J. De Laet, Ed.) History ofHumanity, Vol. 1, ch. 10, pp. 107–116. Paris and London: Routledge and Unesco.
Valoch, K. (1995). The earliest occupation of Europe: Eastern Central and Southeastern Europe. In (W. Roebroeks& T. van Kolfschoten, Eds) The Earliest Occupation of Europe, pp. 67–84. Lieden: University of Leiden.
Villa, P. (1982). Conjoinable pieces and site formation processes. Am. Antiq. 47, 276–290.Villa, P. (1983). Terra Amata and the Middle Pleistocene Archaeological Record of Southern France. Berkeley and Los Angeles:University of California Press.
Villa, P. & Bartram, L. (1996). Flaked bone from a hyena den. Paléo 8, 143–159.Villa, P. & Courtin, J. (1983). The interpretation of stratified sites: a view from underground. J. Archeol. Sci. 10,267–281.
Vincent, A. (1987). Outillage osseux du Paléolithique moyen à Bois Roche (Cherves-Richemont, Charente)? Etudepréliminaire. In Préhistoire de Poitou-Charente, Problèmes Actuels, pp. 201–206. Paris: C.T.H.S.
Vincent, A. (1988a). L’os comme artefact au Paléolithique moyen: principes d’étude et premiers résultats. In (M. Otte,Ed.) L’Homme de Néandertal, Vol. 4. La Technique, pp. 185–196. ERAUL: Liège.
Vincent, A. (1988b). Rémarques préliminaires concernant l’outillage osseux de la Grotte Vaufrey. In (J. Ph. Rigaud,Ed.) La Grotte Vaufrey à Cenac et Sain-Julien (Dordogne). Paleoenvironnement, Chronologie et Activités Humaines, pp. 529–533.Paris: Mém. Soc. Préhist. Française 19.
Vincent, A. (1993). L’outillage osseux au Paléolithique moyen: une nouvelle approche. PhD Dissertation, University of Paris.Wells, C. (1963). Cortical grooves on the tibia. Man 63, 112–114.Wetzel, R. & Bosinski, G. (1969). Die Bocksteinschmiede im Lonetal. Stuttgart: Veröffentichungen des Staatlichen Amtesfür Denkmalpflege.
White, R. (1992). Beyond art: toward an understanding of the origins of material representations in Europe. Ann. Rev.Anthropol. 21, 537–564.
White, R. (1995). Comment on R. Bednarik’s article ‘‘Concept-mediated marking in the Lower Paleolithic’’. Curr.Anthropol. 36, 623–625.
Wolpoff, M. & Caspari, R. (1996). Why aren’t Neandertals modern humans? In (O. Bar-Yousef, L. Cavalli-Sforza,R. J. March, M. Piperno, Eds) The Lower and Middle Palaeolithic, Colloquium IX and X, XIII UISPP Congress,pp. 133–156. Forli: A.B.A.C.O.




























