Embed Size (px)
Citation preview

[CANCER RESEARCH 41. 1751-1756. May 1981]0008-5472/81 /0041-OOOOS02.00
Heterogeneity of Malignant Cells from a Human Colonie Carcinoma1
Michael G. Brattain,2 W. David Fine, F. Mahnaz Khaled, Jerry Thompson, and Diane E. Brattain
Department of Biochemistry, University of Alabama in Birmingham, Birmingham, Alabama 35294
ABSTRACT
Three subpopulations of malignant cells were isolated froma primary cell culture of a single human colonie carcinoma. Thevariant cells were established as cell lines designated HCT116, HCT 116a, and HCT 116b, respectively. In vitro characterizations of the variant lines included growth in 0.5% agaroseand growth on confluent layers of mouse fibroblasts. HCT 116ashowed the highest colony formation in agarose and on confluent fibroblasts, while colony formation by HCT 116 washigher than that of HCT 116b in both of these systems. All ofthe variant lines were tumorigenic in athymic nude mice giveninjections of 10 x 106 cells, but the time between inoculation
and tumor development (latency period) was approximately 10times longer for HCT 116b as for HCT 116a and 8 times longerthan for HCT 116. HCT 116b was not tumorigenic at aninoculum of 5 X 106 cells, while both HCT 116 and 116a were
tumorigenic at this level. However, HCT 116a was clearly moretumorigenic than was HCT 116 on the basis of the number ofanimals developing tumors at inocula of both 10 x 106 and 5x 106 cells and on the basis of their differences in latency
periods. While all the cell lines had near diploid numbers ofchromosomes, each line showed a distinct histological patternwhen grown as xenografts in athymic nude mice.
INTRODUCTION
Recent investigations have indicated morphological andfunctional heterogeneity among the malignant cells of individualcolonie carcinomas. Studies from our laboratory demonstratedmorphological heterogeneity among the malignant cells purified from individual primary human colonie carcinomas (5).Dexter et al. (8) described the isolation of 2 clones of malignantcells from a single colonie carcinoma. These 2 clones weredifferent with regard to several in vitro properties, includingmorphology, cloning efficiency in soft agar, and saturationdensity. However, differences between the in vivo tumorigeniccapabilities of the 2 clones were not described. Recently, wedescribed the isolation of 2 subpopulations of malignant cellsfrom an established cell line of human colonie carcinoma(HT29) with different tumorigenic capabilities (14). In this study,we describe the characterization of 3 subpopulations of malignant cells with different tumorigenic capabilities isolated froma primary culture of a human colonie carcinoma.
MATERIALS AND METHODS
Tissue Culture. A primary human colonie carcinoma, designated HCT 116, was obtained from surgery through theauspices of the DAB Tissue Procurement Service. The tumor
' Supported by Grant CA 21520 from the National Cancer Institute through
the National Large Bowel Cancer Project. American Cancer Society Grant PDT-109A, and Grant CA 13148 from the NIH.
2 To whom requests for reprints should be addressed.
Received June 16, 1980; accepted February 3, 1981.
was prepared for tissue culture as described previously (2, 6).Briefly, the tumor was minced into 1- to 2-cu mm pieces inMcCoy's tissue culture medium containing 20% HFBS3 and
antibiotics [gentamicin 4.3 ¿ig/ml), streptomycin (90 ¿ig/ml),and penicillin (90 units/ml)], washed extensively in the samemedium, and disaggregated with 0.25% trypsin (Grand IslandBiological Co., Grand Island, N. Y.) for 8 periods of 20 min asdescribed previously for human colonie carcinoma (4). Cellsobtained by this procedure were greater than 90% viable bytrypan blue exclusion. Cells (107) or trypsinized chunks of
tissue remaining after the eighth trypsin treatment were platedinto 75-sq cm flasks in the medium described above andmaintained at 37°in a humidified atmosphere of 5% CO2.
Once they were established, cultures were maintained in thesame medium described above except that HFBS levels werereduced to 10%. Confluent cultures were subcultured with0.25% trypsin in Joklik's tissue culture medium containing
0.1% EDTA.Separation of Subpopulations. Primary cultures of HCT 116
contained 3 morphological variants. These variants (designatedHCT 116, HCT 116a, and HCT 116b) were separated andisolated by sedimentation of cells from primary cultures in alinear density gradient of Percoli and tissue culture medium(1.02 to 1.13 g/ml). Procedures similar to those describedpreviously were utilized for centrifugation, collection of fractions containing separated cells, and tissue culture (3, 4, 17).Briefly, separation of 10 x 106 cells from primary cultures wasachieved after centrifugation at 1000 x g for 30 min at 4°in
a CRU 5000 centrifuge. The sterile density gradient was collected by displacement with 60% sucrose, and the cells wereharvested by centrifugation at 97 x g for 7.5 min after dilutionof the fractions with tissue culture medium. Pelleted cells wereresuspended in 5 ml of the growth medium containing 10%HFBS (described above) and inoculated into 25-sq cm tissueculture flasks until sufficient numbers of cells had grown forsubculture and subsequent inoculation of 75-sq cm flasks.
Determination of Growth Characteristics in Vitro. Saturation density and doubling times were determined from growthcurves as we have described previously (6). Cells (105) wereinoculated into 25-sq cm dishes and counted every other dayafter trypsinization until cell numbers were constant for a 6-dayperiod. Cell counts on duplicate cultures were performed withhemocytometer chambers.
Determination of growth in semisolid medium was performedas we have described previously (14,15). Briefly, 5x10" cells
were suspended in the growth medium containing 0.5% aga-rose and inoculated onto previously formed underlayers of 1%agarose containing growth medium in 9.6-sq cm tissue culturedishes. The dishes were equilibrated overnight in 5% CO2 at37° and then sealed with Parafilm to prevent drying. After 3
weeks of incubation, colony formation was determined bymicroscopically counting colonies of 20 or more cells. Prior to
3 The abbreviations used are: HFBS. heat-inactivated fetal bovine serum; CEA.
carcinoembryonic antigen.
MAY 1981 1751
on June 7, 2018. © 1981 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

M. G. Brattain et al.
counting, agarose cultures were fixed with glutaraldehyde.Control dishes were fixed with glutaraldehyde immediately aftersolidification of the 0.5% agarose mixture containing cells.
The ability to grow on confluent layers of C3H10T1/2 mousefibroblasts in 25-sq cm flasks was determined by plating 5 X10" cells and microscopically counting the resultant colonies
after 7 days. Cocultures of tumor cells and fibroblasts weregiven a complete medium change 4 days after seeding andimmediately prior to scoring colonies on the seventh day.
CEA was determined with the commercially available ra-dioimmunoassay as we have described previously (13). CEAassays were performed on cells from confluent cultures and onspent media which had been in contact with confluent culturesof cells for 72 hr. In the case of HCT 116a cells, which will notform completely confluent cultures, assays were performed oncells and spent media from cultures containing the highest celldensity obtained with this line.
Modal chromosome numbers were determined as describedpreviously after blocking cultured cells at metaphase by a 24-hr Colcemid treatment (14). Briefly, treated cells were resus-pended in water for 10 min at 37°,harvested by centrifugaron,
fixed in methanohacetic acid (3:1, v/v) for 1.5 hr, applied toslides, and stained with 20% Giemsa for 7 min.
Tumorigenicity. Athymic nude mice were given s.c. injections with trypsinized or scraped cells in tissue culture mediumwithout HFBS. Animals were observed daily for the formationof tumors, and the period between injection and tumor development (at least 1 cm in diameter) was referred to as the"latency period" in this study. Tumors were removed from
animals, fixed in formalin, and processed for histological examination by the routine techniques.
RESULTS
Establishment of Cell Lines. Primary cultures of HCT 116contained 3 morphological varieties of cells referred to as HCT116 (Fig. 1), HCT 116a (Fig. 2), and HCT 116b (Fig. 3),respectively. By phase-contrast microscopy, HCT 116 grew asclosely packed polygonal cells. HCT 116a cells were roundedand grew in tightly packed clusters which would not formmonolayers or become completely confluent. HCT 116b cellswere polygonal but would not become tightly packed until thecells became confluent. HCT 116 cells were present in primarycultures with far greater frequency than HCT 116a or HCT116b cells (75 to 90% of the cells were HCT 116 as estimatedby phase-contrast microscopy).
The morphological variation observed in these cultures suggested the possibility of functional variation among the celltypes. In order to determine functional variability, it was necessary to separate the various cell types and then grow sufficient numbers in cell culture for characterization. We utilizedcentrifugation in a linear gradient of Percoli in tissue culturemedium to obtain separation of the 3 cell types. We had usedsimilar methods for the separation of malignant colon cellspreviously (3-5). All 3 cell types showed wide ranges of density(Table 1), but the lower density range (1.026 g/ml) for HCT116a allowed for the separation of a fraction of these cells fromthe other 2 cell types, while the very high density of some HCT116b cells (1.092 g/ml) allowed for the isolation of a fractionof these cells. Separated HCT 116a and HCT 116b cells fromthese density ranges were plated and grown in cell culture.
Table 1
Separation of variant subpopulations by centrifugation
Cells from each individual fraction were recovered and cultured as describedin "Materials and Methods." Density ranges were determined by refractometry
of the individual fractions as described by Pertoft and Laurent (17). Fractionswere determined to contain a given cell type on the basis of whether colonies ofthe cell type were observed in cultures derived from the cells recovered from thatfraction.
Density range
CelllineHCT
116aHCT 116HCT 116bFractions
in which celltype wasrecovered1.026-1.068
1.037-1.0801.044-1.092Fractions
utilized forpure cultures of the cell
type1.026-1.0321.050-1.074
1.086-1.092
The cells isolated from the gradient at a density of 1.026 g/mlcontained a mixture of fibroblasts and HCT 116a cells. Thenumber of cells recovered at this density amounted to approximately 6% of the cells recovered from the gradient. Aftergrowth in culture, HCT 116a cells were separated from contaminating fibroblasts by a second density gradient centrifugation.No fibroblasts were observed in cultures initiated with cellsfrom a density of 1.092 g/ml (HCT 116b). Cells recovered atthis density amounted to less than 3% of the cells recoveredfrom the density gradient. When cells grown from culturesinitiated with low-density HCT 116a cells or high-density HCT
116b cells were applied to a second density gradient centrifugation, they exhibited the full range of densities shown by theoriginal cells from the primary cultures.
Since HCT 116 was present in considerably greater concentrations than the other subpopulations, it was isolated by repeated differential trypsinization and reseeding at low cellconcentrations in a manner similar to that used by Dexter era/.(9) for the isolation of a subpopulation of cells from a mammarytumor and by Tan et al. (23) for the removal of fibroblasts fromcultures of mouse colonie carcinoma cells. Our procedure isbriefly described below. Cultures of cells obtained from thedensity range of 1.050 to 1.074 g/ml were allowed to grow tonear confluency. These cultures contained HCT 116a cells,HCT 116b cells, and fibroblasts, but crude estimates by phase-
contrast microscopy indicated that on the order of 75 to 90%of the surface area of the flask was covered with HCT 116cells. In order to obtain pure HCT 116 cells, flasks weresubjected to our normal subculture procedure involving 0.25%trypsin:0.1% EDTA except that instead of allowing a 10-min
exposure, the trypsin:EDTA mixture was removed after 3 minat 37°. The cells were then harvested by centrifugation and
inoculated into a 25-sq cm flask. Fresh trypsin:EDTA was
added to the original culture flasks, and the above procedurewas repeated for 3 additional periods. Cells harvested after thefourth period of trypsin:EDTA exposure were seeded into flasksat a concentration of 10" cells/flask and allowed to grow to
near confluency (approximately 10 days). Examination of thecultures by phase-contrast microscopy indicated that no other
cell types except HCT 116 were present. However, the cultureswere again subjected to the same differential trypsinizationprocedure described above. Cultures of HCT 116 cells obtained from the fourth trypsin:EDTA treatment were utilized asparent cultures for the remainder of the studies describedbelow. The characterizations described below were performedat passage 10 or above for each of the "purified" cell types.
1752 CANCER RESEARCH VOL. 41
on June 7, 2018. © 1981 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

Heterogeneity of Cells from a Colonie Carcinoma
In Vitro Growth Characterization. The growth characteristics of the 3 variant populations are presented in Table 2.Doubling times for the 3 variants were not significantly different.The saturation densities of HCT 116 and HCT 116a wereapproximately twice that of HCT 116b. It should be noted,however, that HCT 116a would never grow to confluency (i.e.,completely cover the surface area of the flask). All of thevariants formed colonies when grown in semisolid medium oron confluent layers of mouse fibroblasts. HCT 116a cellsformed the highest number of colonies in soft agarose cultureand on mouse fibroblasts, while HCT 116 cells showed higherlevels of colony formation than did HCT 116b under thoseconditions. It is interesting to note that the efficiency of colonyformation was generally higher on confluent fibroblasts than insemisolid medium for all 3 variants. HCT 116a cells had veryhigh levels of CEA relative to HCT 116 cells, while HCT 116bcells contained negligible levels of this antigen.
Tumorigenicity. The 3 subpopulations showed variation withrespect to their tumorigenic abilities when injected s.c. intoathymic nude mice (Table 3). The relative tumorigenic abilitiesof the variants were consistent with their in vitro abilities forgrowth in semisolid medium and on confluent fibroblasts. HCT116a cells formed tumors at both high and low inocula of miceand had the shortest latency periods relative to the other variantsubpopulations (5 and 9 days for inocula of 10 x 106 and 5x 106 cells, respectively). HCT 116 cells showed intermediate
tumorigenic ability as approximately 50% of the animals giveninjections at the lower inocula developed tumors with an average latency period of 16 days. Injection of 10 x 106 HCT 116b
cells resulted in xenograft formation in 50% of the recipientanimals; however, latency periods were much higher than forHCT 116a or HCT 116. Inoculation of 5 x 106 HCT 116b cells
failed to produce tumors in 6 mice observed for periods rangingfrom 2 to 4 months (depending upon the survival of the individ-
Table 2
Growth characteristics of the variant subpopulations in vitro
Colony formation(no. of colonies/no, ofcells plated x 100%)
CelllineHCT
116aHCT 116HCT 116bTD
<hr)22.26a
21, 2024, 20üaiuration
density(10s cells/
sqcm)3.9,
4.23.9. 4.31.8,2.4On
confluentmouse fibroblasts18.8,
14.39.0. 6.73.6, 2.9In
0.5%agarose9.2,
8.32.9, 3.92.1, 2.3CEA(ng/106
cells)2360
76Negligible
The 2 sets of numbers under each heading represent results obtained from2 different experiments. Numbers on the left represent experiments performedwith cells obtained between passages 15 and 20, while the second set ofexperiments was performed with cells obtained between passages 30 and 40(depending upon the cell line).
Table 3
Tumorigenicity of variant subpopulations in athymic nude mice
CelllineHCT
116aHCT
116HCT
116bInoculum
(x 106cells)5
105
10510No.
of animals withtumors/no, of ani
mals giveninjection4/5
6/64/9
5/80/6
3/6Latency
period(days)5,
9. 10, 133, 3, 4, 5. 7,814,
11, 19, 215, 6, 6, 6, 5.846,
57, 61
ual mouse). In order to determine the stability of the variantlines, tumorigenicity experiments were conducted at 9 and 13months after the establishment of pure cultures of the subpop-ulations. No differences in their tumorigenicity were observedat these times.
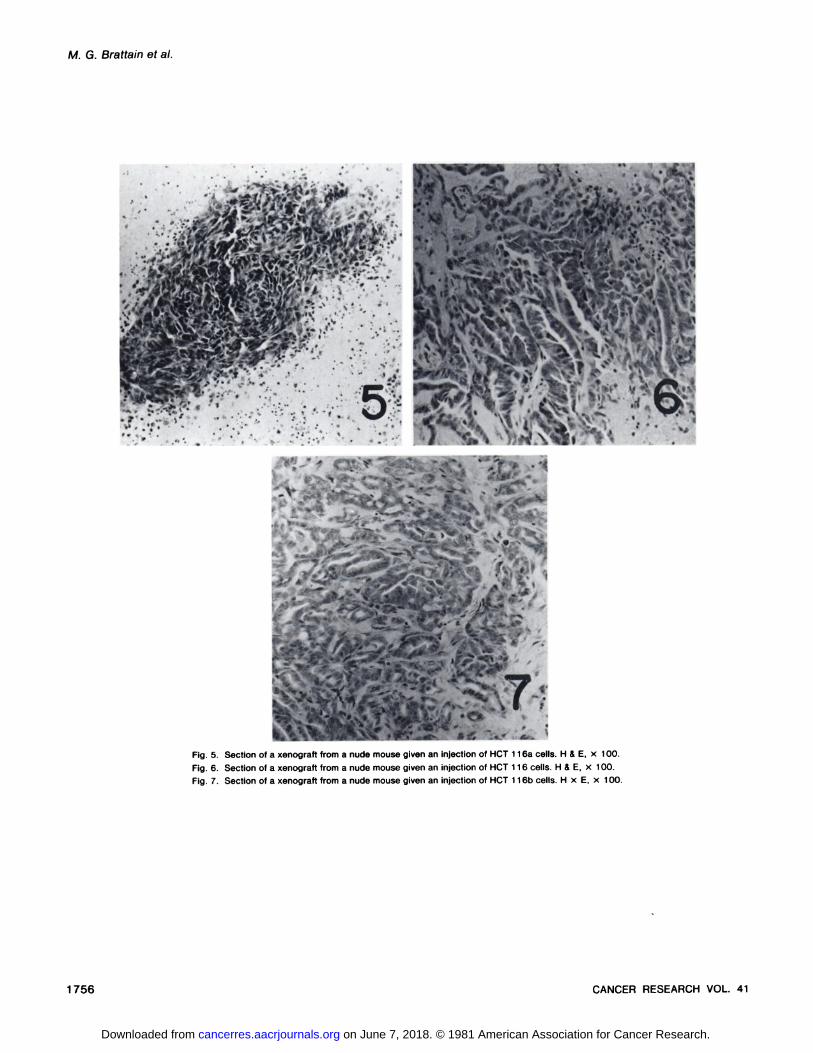
Histological examination of the tumors grown in nude miceindicated that all 3 types of cells gave rise to epithelioid tumors.Tumors from HCT 116a cells are less differentiated than aretumors resulting from the injection of HCT 116 or HCT 116bcells (Figs. 5 to 7). Tumors from HCT 116b cells show aconsistently different histological pattern from those obtainedfrom HCT 116 cells. All 3 variants showed a modal chromosome number of 46 (counting at least 50 mitoses). Less than10% of the mitoses contained 45 chromosomes in each of thevariants, and when 45 chromosomes were present, the missingchromosome appeared to be a random loss.
DISCUSSION
Heterogeneity of the malignant cells comprising individualtumors has been described for a number of cellular and functional properties. Among others, these include morphology andstate of differentiation (18, 19), metastatic and invasive ability(9-11, 16), karyotype (1), and pharmacological response to
drugs (7, 12). More complete summaries of the identificationand isolation of heterogeneous subpopulations of malignantcells have recently been published (7, 20). The studies described in this report demonstrate the potential complexity ofhuman colonie carcinoma with regard to the presence of diverse cell types with variable functional capabilities.
Several investigators (7-9, 20) have summarized some im
plications of tumor heterogeneity in relation to therapeuticapproaches. The existence of discrete subpopulations raisesseveral questions regarding both the biological expression ofcancer and its treatment. From the standpoint of the chemotherapy of cancer, heterogeneous subpopulations of malignantcells suggest that some subpopulations in an individual tumormay respond well to particular therapeutic regimens whileothers may not respond at all. Current evidence would seem toindicate that this is indeed the case in some systems (12). Itfollows then that the identification and characterization of thesubpopulations of cells comprising an individual tumor mightbe of paramount importance in its treatment. In vitro techniques, such as the stem cell soft agar assay (22), for theprediction of drug sensitivity may be quite lacking for manytumors in that subpopulations comprising a small percentageof the cells could be insensitive to the particular drug testedand yet go undetected because of their small contribution tothe total numbers of colonies observed in untreated controls(14). The currently available evidence does suggest that stemcell assays are more effective for predicting which drugs willnot be effective for treating a patient's tumor, rather than
predicting drugs to which a particular tumor would be sensitive(24).
Morphological heterogeneity of purified cells from coloniecancer has been described (3, 5, 21). Dexter et al. (8) described the isolation of 2 clones of cells derived from a singlehuman colonie cancer. These clones showed some variation intheir in vitro growth properties, but differences in their tumorigenic capability in athymic nude mice were not described. Wehave described the isolation of a subpopulation of cells from
MAY 1981 1753
on June 7, 2018. © 1981 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

M. G. Brattain et al.
an established human colon carcinoma (HT29) which formsmetastasis in nude mice, whereas the parental cells will formtumors only at the primary site of injection (14). However, thissubpopulation of cells could have developed as a result ofprogression during the long period in which the cells have beenin culture or as a response to the treatment of the cells with 5-fluorouracil. In this study, we have characterized variant cellsubpopulations from a single primary colon carcinoma whichhave different degrees of tumorigenic potential as judged bytheir abilities to form tumors in nude mice. An important question arising from the identification of heterogeneous subpopulations concerns the nature of their origin. Are these subpop-ulations the result of different stages of differentiation, or dothey represent different levels of progressive mutation? Wecannot answer this question at present, but it would be relevantto point out that most of the cells of all the subpopulationsdescribed in this study contain normal chromosome numbers.This is in contrast to the subpopulations isolated from humancolonie carcinomas which have been described in previousreports (8, 9). While chromosome data and the state of differentiation observed on histological sections of nude mousetumors arising from the injection of the subpopulations arecertainly not proof that these subpopulations are related by acommon path of differentiation, the evidence presently available suggests that they might be a good model for studies ofdifferentiation in human colonie carcinoma.
The utility of in vitro markers for the prediction of tumorigenicpotential has been of considerable interest in recent years.There are, however, relatively few systems for which cells ofdiffering tumorigenic and aggressive abilities derived from thesame tumor are available for comparison and evaluation ofmarkers. HCT 116 and its variants were characterized withrespect to their abilities to grow on confluent fibroblasts and insemisolid medium. It is interesting to note that, in this system,increasing tumorigenic potential of the individual subpopula-tions is associated with high colony formation on fibroblastsand in agarose. On the other hand, the characterization ofmetastatic and nonmetastatic cells from the established cellline HT29 indicated that the more aggressive metastatic cellsshowed lower colony formation in semisolid medium than didtheir nonmetastatic counterparts (14). Studies performed withsubpopulations of cells isolated from a murine mammary tumoralso indicated that the more aggressive cell types in vivo didnot necessarily show a high degree of tumorigenic potential bythe criteria of in vitro markers of tumorigenicity (9). Thus, itdoes not appear to be a general rule that in vitro markersreflect the in vivo tumorigenic potential of cell lines.
While the variant cell lines established in this study haveretained the morphologies seen in the primary expiant of theoriginal colon tumor, they have been in culture for an extendedperiod of time. As with all long-term cultures, one must be
aware of the possibility of changes occurring in the cell lines.Thus, the properties of the cells carried in vitro may not atpresent be reflective of properties of the cells comprising the
original tumor.
REFERENCES
1. Becker, F. F., Klein, K. M., Wolman, S. R., Asofsky, R., and Sell, S.Characterization of primary hepatocellular carcinomas and initial transplantgenerations. Cancer Res., 38: 3758-3763, 1978.
2. Brattain, M. G. Tissue dissaggregation. In: M. Melamed, P. Mullaney, andM. Mendelsohn (eds.). Flow Cytometry and Sorting, p. 193. New York: JohnWiley & Sons, 1979.
3. Brattain, M. G., Green, C.. Kimball, P. M., Marks, M.. and Khaled, M.Isoenzymes of ß-hexosaminidase from normal rat colon and colonie carcinoma. Cancer Res., 39: 4083-4090, 1979.
4. Brattain. M. G.. Kimball, P. M.. Pretlow, T. G., and Pitts, A. M. Partialpurification of human colonie carcinoma cells by sedimentation. Br. J.Cancer. 35: 850-857, 1977.
5. Brattain. M. G., Pretlow, T. P., and Pretlow, T. G. Cell fractionation of largebowel cancer. Cancer (Phila.), 40: 2479-2486, 1977.
6. Brattain. M. G., Strobel-Stevens, J., Fine, D., Webb. M.. and Sarrif, A. M.Establishment of mouse colonie carcinoma cell lines with different metastaticproperties. Cancer Res., 40: 2142-2146, 1980.
7. Calabresi. P.. Dexter, D. L., and Heppner, G. H. Clinical and pharmacologicalimplications of cancer cell differentiation and heterogeneity. Biochem. Phar-macol., 28: 1933-1941, 1979.
8. Dexter. D. L., Barbosa, J. A., and Calabresi, P. N,N-Dimethylformamide-induced alteration of cell culture characteristics and loss of tumorigenicityin cultures of human colon carcinoma cells. Cancer Res., 39: 1020-1025,1979.
9. Dexter, D. L., Kowalski. H. M., Blazar, B. A., Fligiel. Z.. Vogel. R., andHeppner, G. H. Heterogeneity of tumor cells from a single mouse mammarytumor. Cancer Res., 38: 3174-3181, 1978.
10. Fidler, I. J. Tumor heterogeneity and the biology of cancer invasion andmetastasis. Cancer Res., 38. 2651-2660. 1978.
11. Fidler, I. J., and Kripke, M. L. Metastasis results from preexisting variantcells within a malignant tumor. Science (Wash. D. C.), 797: 893-895, 1977.
12. Heppner, G. H., Dexter, D. L., DeNucci, T., Miller. F. R., and Calabresi, P.Heterogeneity in drug sensitivity among tumor cell subpopulations of a singlemammary tumor. Cancer Res., 38: 3758-3763, 1978.
13. Kimball. P.M., and Brattain, M. G. A comparison of methods for the isolationof carcinoembryonic antigen. Cancer Res.. 38: 619-623, 1978.
14. Kimball, P. M., and Brattain, M. G. Isolation of a cellular subpopulation froma human colonie carcinoma cell line. Cancer Res., 40: 1574-1579, 1980.
15. Kimball, P. M., Brattain, M. G., and Pitts, A. M. A soft-agar procedure formeasuring growth of human colonie carcinomas. Br. J. Cancer, 37: 1015-1019. 1978.
16. Kripke, M. L., Gruys, E.. and Fidler, I. J. Metastatic heterogeneity of cellsfrom an ultraviolet light-induced murine fibrosarcoma of recent origin. Cancer Res., 38: 2962-2967, 1978.
17. Pertoft, H., and Laurent, C. Isopycnic separation of cells and organelles bycentrifugation in modified silica gradients. In: N. Catsimpoolas (ed.), Methodsof Cell Separation, p. 25. New York: Plenum Publishing Corp., 1977.
18. Pierce, G. B., Nakane, P. K., Martinez-Hernandez, A., and Ward, J. M.Ultrastructural comparison of differentiation of stem cells of murine adeno-carcinomas of colon and breast with their normal counterparts. J. Nati.Cancer Inst.. 58: 1329-1345, 1977.
19. Pierce, G. B., Stevens, L. C., and Nakane, P. K. Ultrastructural analysis ofthe early stages of development of teratocarcinomas. J. Nati. Cancer Inst.,39. 755-773, 1967.
20. Poste, G., and Fidler, I. J. The pathogenesis of cancer metastasis. Nature(Lond.), 283. 139-146, 1980.
21. Pretlow, T. P., Glover, G. L.. and Pretlow, T. G., II. Purification of malignantcells and lymphocytes from a rat transplantable mucinous adenocarcinomaof the colon by isokinetic sedimentation in gradients of Ficoll. J. Nati. CancerInst., 59:981-987, 1977.
22. Salmon. S. E.. Hamburger, A. W., Schonlen. B., Durie, B. G. M., Alberts, D.S., and Moon, T. E. Quantitation of differential sensitivity of human tumorcells to anticancer drugs. N. Engl. J. Med., 298: 1321-1327, 1978.
23. Tan, M. H., Holyoke, E. D., and Goldrosen, M. H. Brief communications.Murine colon adenocarcinomas: methods for selective culture in vitro. J.Nati. Cancer Inst., 56: 871-874, 1976.
24. Von Hoff, D. Clinical correlation of drug sensitivity in tumor stem cell assay.Proc. Am. Assoc. Cancer Res., 27: 134, 1980.
1754 CANCER RESEARCH VOL. 41
on June 7, 2018. © 1981 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

Heterogeneity of Cells from a Colonie Carcinoma
Fig. 1. Phase-contrast microscopy of HCT 116 cells in culture, x 100.Fig. 2. Phase-contrast microscopy of HCT 116a cells in culture, x 100.Fig. 3. Phase-contrast microscopy of HCT 116b cells in culture, x 100.Fig. 4. Phase-contrast microscopy of HCT 116 colony on confluent mouse fibroblasts. x 100.
MAY 1981 1755
on June 7, 2018. © 1981 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
M. G. Brattain et al.
s •
KW •••y »'C,Ãjr:¿SB# v-'"''>: '.«
^ - •Õ.-Ü
Fig. 5. Section of a xenograft from a nude mouse given an injection of HCT 116a cells. H & E, x 100.
Fig. 6. Section of a xenograft from a nude mouse given an injection of HCT 116 cells. H & E, x 100.
Fig. 7. Section of a xenograft from a nude mouse given an injection of HCT 116b cells. H x E, x 100.
1756 CANCER RESEARCH VOL. 41
on June 7, 2018. © 1981 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

1981;41:1751-1756. Cancer Res Michael G. Brattain, W. David Fine, F. Mahnaz Khaled, et al. CarcinomaHeterogeneity of Malignant Cells from a Human Colonic
Updated version
http://cancerres.aacrjournals.org/content/41/5/1751
Access the most recent version of this article at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://cancerres.aacrjournals.org/content/41/5/1751To request permission to re-use all or part of this article, use this link
on June 7, 2018. © 1981 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from





























