Embed Size (px)
Citation preview

Heritability of Yield Components Under Irrigated and DroughtConditions in Andigenum Potatoes
Rolando Cabello & Philippe Monneveux &
Merideth Bonierbale & M. Awais Khan
# Potato Association of America 2014
Abstract Drought is one of the major abiotic factors thataffect potato production in the tropics and sub-tropics.Molecular plant breeding offers promising opportunities fordeveloping drought tolerant cultivars, but genetic gains to beachieved through breeding depends strongly on the geneticvariance and heritability of traits used in the selection process.In the present study, genetic variance and heritability of tuberyield and its components were estimated in the native potatocultivar group Andigenum, expected to include promisingdrought tolerant progenitors. Twenty-seven full sib familiesobtained from crossing 18 parents arranged in 3 mating setswere grown under irrigated and terminal drought conditions.Heritability for tuber yield, estimated from female to malevariance components was 0.68 and 0.41, respectively, in theirrigated treatment, and only 0.18 and 0.06 under droughtconditions. Tuber number was highly significantly correlatedto tuber yield but the heritability of this trait (estimated bothfrom female to male variance) was lower than yield heritabil-ity and its non-additive genetic variance was higher than itsadditive genetic variance, particularly when estimated fromthe female variance component. Heritability of average weightper tuber estimated for male variance component was veryhigh under both irrigated and drought conditions. Its additivegenetic variance was much higher than its non-additive genet-ic variance. However, average weight per tuber correlated totuber yield within parents but not within families. Theseresults indicate that selection for increased tuber yield underdrought conditions in the Andigenum cultivar group may beslow due to its low heritability and high value of non-additivegenetic variance. They also suggest that yield componentsmay not be useful as indirect selection criterion for yield,because of their low heritability and/or poor association with
yield. The identification of drought tolerance related traitswith high heritability and closely associated to tuber yieldunder drought is consequently required to improve adaptationof potato to drought-prone conditions and efficient utilizationof Andigenum potatoes for this purpose.
Resumen La sequía es uno de los factores abióticosprincipales que afectan la producción de papa en los trópicosy subtrópicos. El mejoramiento molecular de plantas ofreceoportunidades prometedoras para desarrollar variedadestolerantes a la sequía, pero las ganancias genéticas a lograr através del mejoramiento, dependen fuertemente de la varianzagenética y de la heredabilidad de los caracteres usados en elproceso de selección. En el presente estudio, se estimaron lavarianza genética y la heredabilidad del rendimiento detubérculo y sus componentes en variedades nativas de papadel grupo Andigenum, esperando que se incluyeranprogenitores prometedores tolerantes a la sequía. A 27familias completas obtenidas del cruzamiento de 18progenitores arreglados en tres juegos de apareamiento, seles cultivó bajo condiciones de riego y de sequía terminal.La heredabilidad para rendimiento de tubérculo, estimada delos componentes de varianza de machos y hembras, fue de0.68 y 0.41, respectivamente, en el tratamiento con riego, ysolo de 0.18 y 0.06 bajo condiciones de agobio hídrico. Elnúmero de tubérculos estuvo alta y significativamentecorrelacionado al rendimiento, pero la heredabilidad de estecarácter (estimada de la varianza, tanto de hembras como demachos) fue más baja que la heredabilidad de rendimiento, ysu varianza genética no aditiva fue más alta que su varianzagenética aditiva, particularmente cuando se estimó delcomponente de varianza femenino. La heredabilidad delpromedio del peso por tubérculo, estimada para elcomponente de varianza masculino, fue muy alta bajo ambascondiciones, de riego y de sequía. Su varianza genética aditivafue mucho más alta que la no aditiva. No obstante, el
R. Cabello : P. Monneveux :M. Bonierbale :M. A. Khan (*)International Potato Center (CIP), AP 1558, Lima 12, Perue-mail: [email protected]
Am. J. Potato Res.DOI 10.1007/s12230-014-9379-7

promedio de peso por tubérculo se correlacionó conrendimiento de tubérculo entre progenitores pero no dentrode las familias. Estos resultados indican que la selección paraaumento del rendimiento de tubérculo bajo condiciones desequía en las variedades del grupo Andigenum pudiera serlenta debido a su baja heredabilidad y al alto valor de lavarianza genética no aditiva. También sugieren que loscomponentes de rendimiento pudieran no ser de utilidad comocriterio indirecto de selección para rendimiento, debido a subaja heredabilidad y/o su pobre asociación con rendimiento.Se requiere, consecuentemente, la identificación de caracteresrelacionados a la tolerancia a la sequía con alta heredabilidad yestrechamente asociados al rendimiento de tubérculo bajoagobio hídrico, para mejorar la adaptación de la papa acondiciones propicias a la falta de agua y utilización eficientede las papas Andigenum para este propósito.
Keywords Additive genetic variance . Andigenum .
Drought . Heritability . Potato
Introduction
Potato is the fourth most important food crop in the world withan annual production of 330 million tons (FAO 2009).However, this crop regularly suffers drought stress in mostgrowing areas due to erratic rainfall or inadequate irrigationtechniques. Drought reduces leaf area (Jefferies 1993), rate ofphotosynthesis per unit of leaf area (Ta et al. 2003), andharvest index (DeBlonde et al. 1999) with the magnitude ofthe response depending on the intensity and timing of waterlimitation (Spitters and Schapendonk 1990).
Potatoes exhibit broad variation for drought tolerance. Themost widely produced and consumed potato, the Tuberosumcultivar group, is considered highly susceptible to droughtstress (Weisz et al. 1994). Due to their evolution and cultiva-tion in the cold and dry Andean Altiplano, Andean potatoesare postulated to be more drought tolerant than Tuberosumpotatoes (Ritter et al. 2008). A recent large-scale screening ofdifferent cultivar groups confirmed the potential of Andeanpotatoes for improving drought tolerance and identified ac-cessions for use in breeding programs (Cabello et al. 2012).The Andigenum group in particular appeared to be an excel-lent candidate for introgressing drought tolerance traits intothe world-wide cultivated Tuberosum. Andigenum potatopresents large variation and adaptation to harsh conditions,being cultivated throughout the Andes from Venezuela toNorthern Argentina (Huaman and Ross 1985) at altitudesbetween 1,950 and 4,050 m (Ochoa 2003). Its tetraploidgenome allows for hybrid generation with Tuberosum(Kumar and Kang 2006). Vasquez-Robinet et al. (2008) re-ported that photosynthesis was less affected by drought inAndigenum than in Tuberosum. Breeding for drought
tolerance can be done either by selecting directly for yieldunder drought conditions or by indirect selection. It is gener-ally recognized that genetic variance and broad sense herita-bility of yield decline with increasing moisture stress (Rosielleand Hamblin 1981; Blum 1988; Bolaños and Edmeades1996). Under drought conditions it is consequently difficultto make genetic progress by selecting for yield per se and theuse of secondary traits is then recommended (Edmeades et al.1997). Secondary traits are plant characteristics other thanprimary edible yield that provide additional information abouthow the plant performs in a given environment (Lafitte et al.2003). The relative usefulness of a secondary trait as indirectselection criteria for grain yield is determined by the magni-tudes of its genetic variance, heritability and genetic correla-tion with the primary trait (Falconer 1981). A secondary traitshould also be easy to measure and not associated with yieldloss under unstressed conditions. In the present study, weinvestigated the possibility of using yield components assecondary traits for improving drought tolerance in theAndigenum cultivar group. For this purpose, we examinedtheir heritability and relationship with yield under both irri-gated and drought conditions.
Materials and Methods
Eighteen native potato cultivars from the Andigenum cultivargroup were used in the experiments (Table 1). Selection ofthese cultivars was based on previous drought tolerance eval-uation (Cabello et al. 2012). The 18 cultivars were used asparents in a North Carolina genetic design II (Ortiz andGolmirzaie 2002) and randomly grouped into three sets eachcontaining three female and three male parents. The male andfemale parents were crossed (Table 2), resulting in 27 full sibfamilies that were evaluated in LaMolina experimental stationof CIP (International Potato Center) located at the coastal partof Peru (latitude −12.08°, longitude −76.91° and altitude240 m above sea level).
The soil at the experimental site is an Entisol, containing46% sand, 32% silt, and 22% clay. Bulk density ranged from1.3 to 1.2 and field capacity from 24 % to 21 % moisture byweight down the soil profile to 100 cm. Soil organic mattercontent was 3.1 % and pH was 7.1. Electrical conductivity(EC) and cation exchange capacity were 2.7 dS/m and10.7 Meq 100 g−1, respectively.
Climatic conditions in La Molina are mainly characterizedby the absence of rainfall, a high relative humidity (between71.3 and 92.0 %), and a low vapor deficit pressure (from 3810−6 MPa in August to 58 10−6 MPa in November). Annualaverage minimum and maximum temperature were 14.9 °Cand 18.0 °C, respectively. Monthly maximum temperatureranged from 17.1 °C (in August) to 19.1 °C (in November).
Am. J. Potato Res.
Accumulated solar radiation increased toward the end of thecropping season and ranged from 1,671 to 4,808 W m−2.
The 27 families and corresponding parents were evaluatedunder irrigated and terminal drought conditions. For compar-ison, the hybrid potato cultivars Revolución, Yungay, andAchirana were included in the experiment. Revolución is animproved variety, well adapted to Peruvian Central Coast, but
susceptible to drought. ‘Yungay’ was developed in the 1950sand is adapted to highland conditions. It grows well in poorsoils and is tolerant to drought and mild frost and producestubers of excellent quality. ‘Achirana’ is also a drought toler-ant improved variety, grown in several countries world-wide.The experimental design was a randomized complete blocksplit plot design with 3 replications. The whole plot wasassigned to two treatments and water was applied to eachtreatment using furrow irrigation similar to methods describedin Cabello et al. (2012). Plants were irrigated once each weekin the irrigated treatment sufficient to provide 360 mm waterover the growing season. In the terminal drought treatment,irrigation was stopped 8 weeks after planting when irrigationreached the equivalent of 180 mm water. The subplot wasassigned to sets and the sub-subplot to families within a set. Inthe experimental design sets and families within a set wereassigned at random. Planting material was 30 seedling tubersper progeny over three replications (i.e., 10 plants/progeny perreplication). Seedling tubers were stored in a cold storage andthen moved to a room where temperature was controlled tobreak the dormancy and provide uniform sprouting prior toplanting them in field. Experiments were planted in Augustand harvested in December. Water flow in the experiment wasregulated by using siphon tubes. Soil moisture was monitoredusing soil moisture HOBO (Onset Computer Corporation,Bourne, MA, USA) sensors at depths of 45 cm and 60 cm.
Tuber yield (g plant−1), tuber number (per plant) and aver-age weight per tuber (g) of the 27 full sib families, 18 parents,and 3 cultivars were measured under irrigated and droughttreatments. Genetic variances and broad sense heritabilitywere calculated for the progeny plants for the irrigated anddrought treatments, according to Falconer (1981) using SASsoftware (SAS Institute Inc. USA).
Results
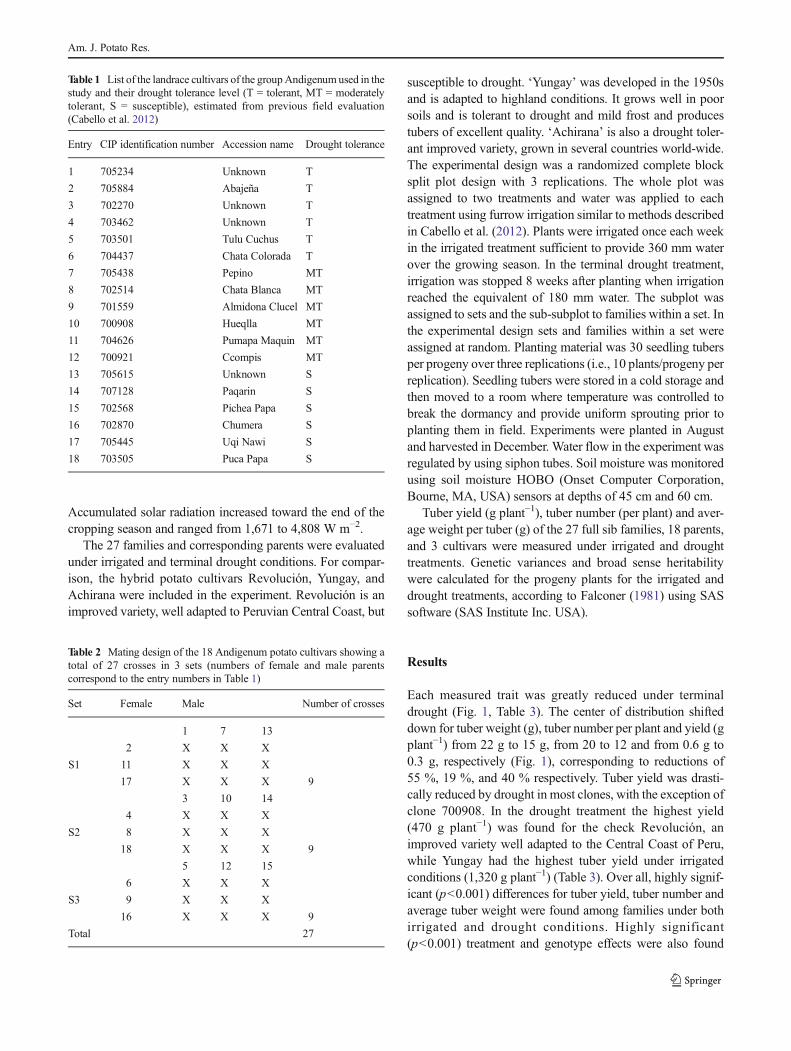
Each measured trait was greatly reduced under terminaldrought (Fig. 1, Table 3). The center of distribution shifteddown for tuber weight (g), tuber number per plant and yield (gplant−1) from 22 g to 15 g, from 20 to 12 and from 0.6 g to0.3 g, respectively (Fig. 1), corresponding to reductions of55 %, 19 %, and 40 % respectively. Tuber yield was drasti-cally reduced by drought in most clones, with the exception ofclone 700908. In the drought treatment the highest yield(470 g plant−1) was found for the check Revolución, animproved variety well adapted to the Central Coast of Peru,while Yungay had the highest tuber yield under irrigatedconditions (1,320 g plant−1) (Table 3). Over all, highly signif-icant (p<0.001) differences for tuber yield, tuber number andaverage tuber weight were found among families under bothirrigated and drought conditions. Highly significant(p<0.001) treatment and genotype effects were also found
Table 2 Mating design of the 18 Andigenum potato cultivars showing atotal of 27 crosses in 3 sets (numbers of female and male parentscorrespond to the entry numbers in Table 1)
Set Female Male Number of crosses
1 7 13
2 X X X
S1 11 X X X
17 X X X 9
3 10 14
4 X X X
S2 8 X X X
18 X X X 9
5 12 15
6 X X X
S3 9 X X X
16 X X X 9
Total 27
Table 1 List of the landrace cultivars of the groupAndigenum used in thestudy and their drought tolerance level (T = tolerant, MT = moderatelytolerant, S = susceptible), estimated from previous field evaluation(Cabello et al. 2012)
Entry CIP identification number Accession name Drought tolerance
1 705234 Unknown T
2 705884 Abajeña T
3 702270 Unknown T
4 703462 Unknown T
5 703501 Tulu Cuchus T
6 704437 Chata Colorada T
7 705438 Pepino MT
8 702514 Chata Blanca MT
9 701559 Almidona Clucel MT
10 700908 Hueqlla MT
11 704626 Pumapa Maquin MT
12 700921 Ccompis MT
13 705615 Unknown S
14 707128 Paqarin S
15 702568 Pichea Papa S
16 702870 Chumera S
17 705445 Uqi Nawi S
18 703505 Puca Papa S
Am. J. Potato Res.
for these traits. The treatment by genotype interaction wassignificant (p<0.05).
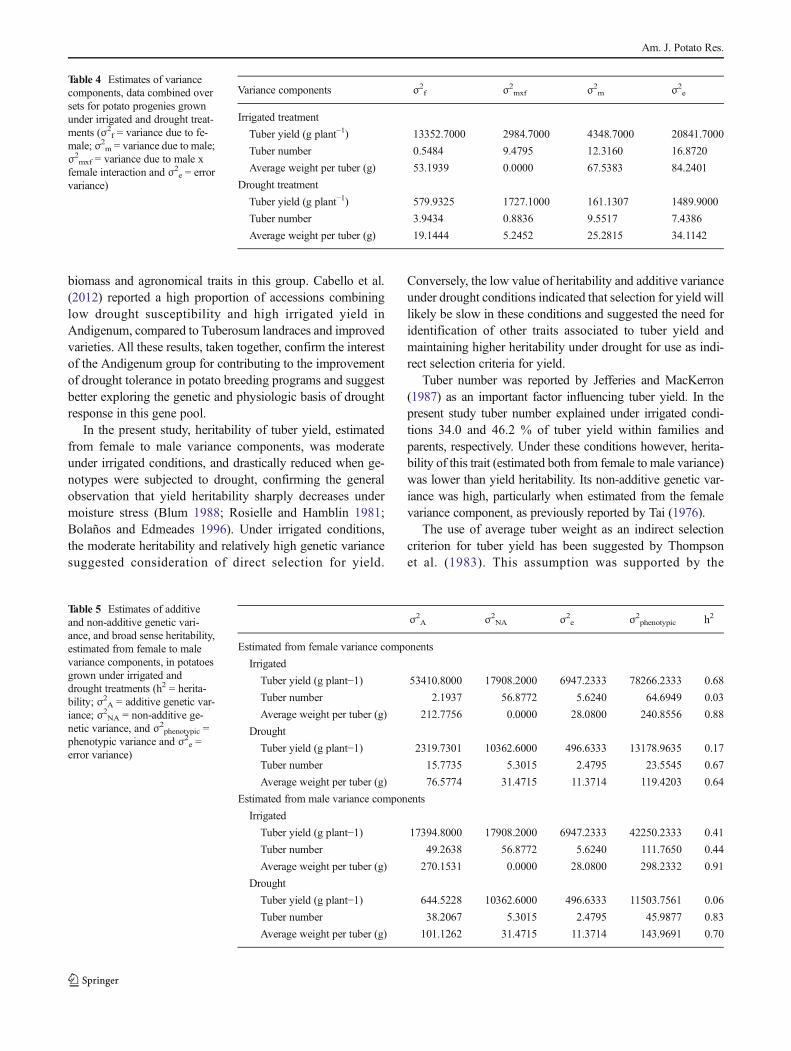
For tuber yield, female variance component was higherthan male variance component in both irrigated and droughtconditions (Table 4). Conversely, for tuber number and aver-age weight per tuber, male variance component was higherthan female variance component. In the irrigated treatment,heritability of tuber yield, estimated from female to malevariance components, was moderate (Table 5). Additive andnon-additive genetic variances had similar magnitudes. Yieldheritability was drastically reduced when genotypes weresubjected to drought. Additive genetic variance was alsolower than under irrigated conditions.
Heritability of tuber number estimated for female variancecomponent was low in the irrigated treatment. Non-additivegenetic variance was higher than additive genetic variance,particularly when estimated from the female variance compo-nent. Tuber number showed a strong positive correlation totuber yield within both parents and families (Table 6).Heritability of average weight per tuber estimated for malevariance component was very high under irrigated conditions.Additive genetic variance was much higher than non-additivegenetic variance. Average weight per tuber correlated to tuberyield within parents but not within families (Table 6). Linearregression showed interaction between all variables amongparents and families in both treatments, with the exception oftuber yield (g plant−1) and tuber number in drought treatment(Fig. 2). Tuber number predicted total tuber yield (g plant−1) inirrigated treatment for parents (r2=0.47) and for families (r2=0.34). Also for parents in the irrigated treatment, averageweight per tuber (g) predicted tuber yield (g plant−1) moder-ately (r2=0.44) (Fig. 2a and b). However, in the droughttreatment, tuber number was a poor predictor of tuber yield(g plant−1) for both parents and families (r2=0.18 and 0.10,respectively) (Fig. 2c and d).
Heritability remained high for both tuber number and av-erage weight per tuber under drought conditions and additive
genetic variance was higher than non-additive genetic vari-ance for both components. No strong correlation was found,however, between these two components and tuber yieldunder drought conditions (Table 6). A highly significant neg-ative correlation was noted among families between tubernumber and average weight per tuber (r=0.726, p<0.001and r=0.854, p<0.001, under irrigated and drought condi-tions, respectively). Crosses of the Andigenum group alsoshowed great variation for drought tolerance, for tuber numberper plant, yield (g plant−1), and average weight per tuber (g).For example, plants resulting from crosses using 702270 asthe male parent had more than 30 tubers per plant, i.e., morethan all crosses and controls, and no reduction in yield (gplant−1) and average weight per tuber (g) under terminaldrought.
Discussion
The tested Andigenum accessions showed diverse responsesto terminal drought stress, one of many drought scenarios thatpotato production faces globally. Interestingly, the ranges ofyield components were greatly narrowed under drought com-pared to irrigated conditions, an indication of similar basicsurvival mechanisms in varieties with great diversity underoptimal conditions. A broad variation for gene expression andphysiological traits under drought was previously noted in theAndigenum group. For example, Watkinson et al. (2006)reported substantial variation for the constitutive expressionlevels of genes encoding enzymes of the flavonoid pathway.Vasquez-Robinet et al. (2008) observed significant differencesbetween two Andigena landraces, ‘Sullu’ and ‘Negra Ojosa’,grown under drought conditions, for photosynthetic response,mitochondrial metabolic activity, induction of chloroplast-localized anti-oxidants, chaperone genes, and ABA inducedtranscription factors. This variation at the genetic and physio-logical level might be related to the plant architectural,
Fig. 1 Distribution of values for tuber number per plant, yield (g plant−1) and average weight per tuber (g) under normal irrigation and terminal drought
Am. J. Potato Res.

Table 3 Mean tuber yield (g plant−1), tuber number and average weightper tuber (g) of 18 parental clones, 27 full sib potato progenies, and 3checks under irrigated and drought conditions in La Molina, Lima, Peru.
Within each group, accessions are ranked by decreasing order of yieldunder drought conditions
CIP identification number Yield (g plant−1) Tuber number Average weight per tuber (g)
Irrigated Drought Irrigated Drought Irrigated Drought
Parental clones
29 705884 1,025±170 375±59 37±7 28±3 29±9 13±1
44 705445 1,247±89 371±114 29±6 26±3 44±9 14±3
28 705234 867±131 289±67 31±4 14±2 28±2 20±3
42 702568 939±181 285±64 17±2 11±2 55±8 25±2
43 702870 991±18 274±34 25±1 18±2 39±2 15±0
40 705615 569±89 267±34 19±1 16±2 31±6 17±4
33 704437 451±89 262±33 13±1 16±2 36±10 17±0
30 702270 147±21 239±19 14±4 35±1 11±1 7±1
32 703501 708±100 232±23 22±3 22±1 32±2 11±2
39 700921 360±82 218±23 17±1 15±1 22±6 14±1
35 702514 665±63 217±97 24±4 20±1 28±3 11±6
37 700908 185±80 213±59 14±4 19±4 13±3 11±1
34 705438 453±90 212±41 23±2 18±3 19±3 12±1
38 704626 486±186 205±19 19±4 22±3 25±5 9±1
45 703505 610±62 204±56 20±3 13±2 30±3 15±3
36 701559 703±72 202±8 39±4 24±5 18±0 9±2
31 703462 241±63 165±57 15±2 13±3 17±6 12±4
41 707128 473±174 128±14 12±8 8±1 46±13 16±1
Crossesa
10 703462/702270 749±34 446±52 22±4 23±3 34±7 20±1
9 705445/705615 816±179 434±94 22±5 13±2 38±8 33±6
26 702870/700921 692±45 393±67 10±3 8±2 71±20 48±10
1 705884/705234 768±85 385±29 29±7 17±3 27±8 23±3
16 703505/702270 930±154 384±50 25±2 20±1 37±4 19±3
20 704437/700921 637±233 382±47 11±4 9±3 58±13 43±7
25 702870/703501 583±209 377±53 11±5 11±3 55±9 35±5
2 705884/705438 722±152 363±38 22±7 18±2 36±18 20±2
21 704437/702568 634±99 347±31 15±2 13±1 41±5 28±4
13 702514/702270 602±86 340±47 17±4 14±1 36±4 24±4
17 703505/700908 561±137 337±55 12±3 11±2 47±10 31±4
8 705445/705438 999±164 336±42 31±9 17±4 33±5 20±2
18 703505/707128 671±109 335±53 11±2 9±2 59±4 37±6
12 703462/707128 679±80 331±9 12±2 12±2 56±10 29±4
24 701559/702568 602±54 329±39 18±5 16±8 35±12 23±12
27 702870/702568 735±51 326±21 15±5 8±2 53±12 39±5
19 704437/703501 771±287 314±24 21±10 11±1 41±11 28±1
4 704626/705234 523±139 312±80 26±5 18±3 21±7 17±2
11 703462/700908 519±76 307±47 16±3 14±0 32±3 23±4
23 701559/700921 731±236 302±26 18±2 10±4 40±9 33±9
22 701559/703501 511±184 283±27 14±5 10±2 37±6 29±7
3 705884/705615 482±141 282±39 17±6 17±2 29±5 17±1
7 705445/705234 741±180 274±36 20±4 12±1 38±9 23±2
6 704626/705615 493±167 273±19 15±8 14±5 35±8 20±7
14 702514/700908 410±110 268±23 11±1 7±1 36±10 38±8
5 704626/705438 447±111 259±21 22±2 16±5 21±5 17±4
15 702514/707128 422±122 256±52 7±2 7±2 57±11 37±9
Checks
46 Revolución 1,144±305 470±5 12±3 10±0 91±8 45±0
48 Yungay 1,320±226 357±66 23±3 17±1 58±12 21±5
47 Achirana 611±160 327±43 9±4 8±1 69±23 42±3
aMean of a cross is based on 30 plants (3 replications of 10 plants per replications)
Am. J. Potato Res.
biomass and agronomical traits in this group. Cabello et al.(2012) reported a high proportion of accessions combininglow drought susceptibility and high irrigated yield inAndigenum, compared to Tuberosum landraces and improvedvarieties. All these results, taken together, confirm the interestof the Andigenum group for contributing to the improvementof drought tolerance in potato breeding programs and suggestbetter exploring the genetic and physiologic basis of droughtresponse in this gene pool.
In the present study, heritability of tuber yield, estimatedfrom female to male variance components, was moderateunder irrigated conditions, and drastically reduced when ge-notypes were subjected to drought, confirming the generalobservation that yield heritability sharply decreases undermoisture stress (Blum 1988; Rosielle and Hamblin 1981;Bolaños and Edmeades 1996). Under irrigated conditions,the moderate heritability and relatively high genetic variancesuggested consideration of direct selection for yield.
Conversely, the low value of heritability and additive varianceunder drought conditions indicated that selection for yield willlikely be slow in these conditions and suggested the need foridentification of other traits associated to tuber yield andmaintaining higher heritability under drought for use as indi-rect selection criteria for yield.
Tuber number was reported by Jefferies and MacKerron(1987) as an important factor influencing tuber yield. In thepresent study tuber number explained under irrigated condi-tions 34.0 and 46.2 % of tuber yield within families andparents, respectively. Under these conditions however, herita-bility of this trait (estimated both from female to male variance)was lower than yield heritability. Its non-additive genetic var-iance was high, particularly when estimated from the femalevariance component, as previously reported by Tai (1976).
The use of average tuber weight as an indirect selectioncriterion for tuber yield has been suggested by Thompsonet al. (1983). This assumption was supported by the
Table 4 Estimates of variancecomponents, data combined oversets for potato progenies grownunder irrigated and drought treat-ments (σ2
f = variance due to fe-male; σ2m = variance due to male;σ2
mxf = variance due to male xfemale interaction and σ2e = errorvariance)
Variance components σ2f σ2mxf σ2m σ2e
Irrigated treatment
Tuber yield (g plant−1) 13352.7000 2984.7000 4348.7000 20841.7000
Tuber number 0.5484 9.4795 12.3160 16.8720
Average weight per tuber (g) 53.1939 0.0000 67.5383 84.2401
Drought treatment
Tuber yield (g plant−1) 579.9325 1727.1000 161.1307 1489.9000
Tuber number 3.9434 0.8836 9.5517 7.4386
Average weight per tuber (g) 19.1444 5.2452 25.2815 34.1142
Table 5 Estimates of additiveand non-additive genetic vari-ance, and broad sense heritability,estimated from female to malevariance components, in potatoesgrown under irrigated anddrought treatments (h2 = herita-bility; σ2A = additive genetic var-iance; σ2
NA = non-additive ge-netic variance, and σ2
phenotypic =phenotypic variance and σ2
e =error variance)
σ2A σ2NA σ2e σ2phenotypic h2
Estimated from female variance components
Irrigated
Tuber yield (g plant−1) 53410.8000 17908.2000 6947.2333 78266.2333 0.68
Tuber number 2.1937 56.8772 5.6240 64.6949 0.03
Average weight per tuber (g) 212.7756 0.0000 28.0800 240.8556 0.88
Drought
Tuber yield (g plant−1) 2319.7301 10362.6000 496.6333 13178.9635 0.17
Tuber number 15.7735 5.3015 2.4795 23.5545 0.67
Average weight per tuber (g) 76.5774 31.4715 11.3714 119.4203 0.64
Estimated from male variance components
Irrigated
Tuber yield (g plant−1) 17394.8000 17908.2000 6947.2333 42250.2333 0.41
Tuber number 49.2638 56.8772 5.6240 111.7650 0.44
Average weight per tuber (g) 270.1531 0.0000 28.0800 298.2332 0.91
Drought
Tuber yield (g plant−1) 644.5228 10362.6000 496.6333 11503.7561 0.06
Tuber number 38.2067 5.3015 2.4795 45.9877 0.83
Average weight per tuber (g) 101.1262 31.4715 11.3714 143.9691 0.70
Am. J. Potato Res.
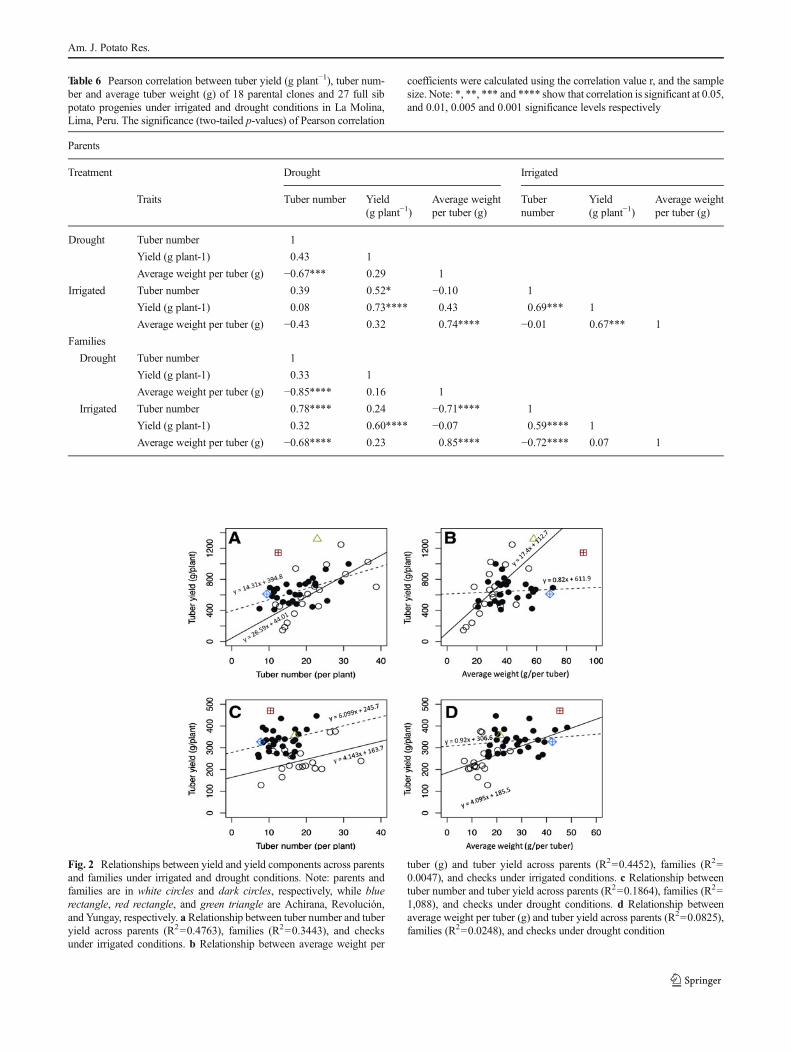
Table 6 Pearson correlation between tuber yield (g plant−1), tuber num-ber and average tuber weight (g) of 18 parental clones and 27 full sibpotato progenies under irrigated and drought conditions in La Molina,Lima, Peru. The significance (two-tailed p-values) of Pearson correlation
coefficients were calculated using the correlation value r, and the samplesize. Note: *, **, *** and **** show that correlation is significant at 0.05,and 0.01, 0.005 and 0.001 significance levels respectively
Parents
Treatment Drought Irrigated
Traits Tuber number Yield(g plant−1)
Average weightper tuber (g)
Tubernumber
Yield(g plant−1)
Average weightper tuber (g)
Drought Tuber number 1
Yield (g plant-1) 0.43 1
Average weight per tuber (g) −0.67*** 0.29 1
Irrigated Tuber number 0.39 0.52* −0.10 1
Yield (g plant-1) 0.08 0.73**** 0.43 0.69*** 1
Average weight per tuber (g) −0.43 0.32 0.74**** −0.01 0.67*** 1
Families
Drought Tuber number 1
Yield (g plant-1) 0.33 1
Average weight per tuber (g) −0.85**** 0.16 1
Irrigated Tuber number 0.78**** 0.24 −0.71**** 1
Yield (g plant-1) 0.32 0.60**** −0.07 0.59**** 1
Average weight per tuber (g) −0.68**** 0.23 0.85**** −0.72**** 0.07 1
Fig. 2 Relationships between yield and yield components across parentsand families under irrigated and drought conditions. Note: parents andfamilies are in white circles and dark circles, respectively, while bluerectangle, red rectangle, and green triangle are Achirana, Revolución,and Yungay, respectively. aRelationship between tuber number and tuberyield across parents (R2=0.4763), families (R2=0.3443), and checksunder irrigated conditions. b Relationship between average weight per
tuber (g) and tuber yield across parents (R2=0.4452), families (R2=0.0047), and checks under irrigated conditions. c Relationship betweentuber number and tuber yield across parents (R2=0.1864), families (R2=1,088), and checks under drought conditions. d Relationship betweenaverage weight per tuber (g) and tuber yield across parents (R2=0.0825),families (R2=0.0248), and checks under drought condition
Am. J. Potato Res.

association of this trait with tuber yield and by its highheritability and additive genetic variance suggesting a highresponse to selection for this component. In our study, however,average weight per tuber correlated poorly with tuber yield,particularly across families. This is likely due to the highlysignificant negative genetic correlation between tuber numberand weight already reported by Thompson et al. (1983) andconfirmed in our study, suggesting that any selection for improv-ing one will decrease the other, with a reduced impact on finaltuber yield. In commercial potato production, tuber weight andsize contribute to potato quality traits (like shape, appearance,absence of diseases or defects, flavor, and cooked texture) andcan be considered as breeding objectives per se, largely deter-mining the quality for export, sorting, grading, and packaging.The high heritability of this component observed in this study inthe cultivar group Andigenum, both under irrigated and droughtconditions, suggest that this trait can be genetically improved.
In conclusion, yield components (Tuber number, andAverage weight per tuber (g)) do not appear as good candi-dates for indirect selection in potato, because of their lowheritability and/or lack of association with tuber yield. Thisresult suggests active exploration of the possibility of usingmorpho-physiological traits implicated in drought tolerance asindirect selection criterion for yield in this species.
Acknowledgments The authors acknowledge support from KarinaValverde, and Enrique Chujoy during the experimental design, fieldevaluations and analysis of data.
References
Blum, A. 1988. Plant breeding for stress environments. Boca Raton: CRCPress.
Bolaños, J., and G.O. Edmeades. 1996. The importance of the anthesis-silking interval in breeding for drought tolerance in tropical maize.Field Crops Research 48: 269–286.
Cabello, R., E. Chujoy, F. Mendiburu, M. Bonierbale, P. Monneveux, W.Roca, and E. Chujoy. 2012. Large-scale evaluation of potatoesimproved varieties, pre-breeding material and landraces for droughttolerance. American Journal of Potato Research 89(5): 400–410.
Deblonde, P.M.K., A.J. Haverkort, and J.F. Ledent. 1999. Responses ofearly and late potato cultivars to moderate drought conditions:agronomic parameters and carbon isotope discrimination.European Journal of Agronomy 11: 91–105.
Edmeades, G.O., M. Bänziger, D. Beck, J. Bolaños, and A. Ortega. 1997.Development and per se performance of CIMMYT maize popula-tions as drought-tolerant sources. In Developing drought and low-Ntolerant maize, ed. G.O. Edmeades, M. Bänziger, H.R. Mickelson,and C.B. Peña-Valdivia, 254–262. Mexico: CIMMYT.
Falconer, D.S. 1981. Introduction to quantitative genetics, 2nd ed.London: Longmans Green.
FAO. 2009. http://faostat.fao.org/site/567/default.aspx#ancor.Huaman, Z., and R.W. Ross. 1985. Updated listing of potato species
names, abbreviations and taxonomic status. American PotatoJournal 62: 629–641.
Jefferies, R.A. 1993. Responses of potato genotypes to drought. I.Expansion of individual leaves and osmotic adjustment. Annals ofApplied Biology 122: 93–104.
Jefferies, R.A., and D.K.L. MacKerron. 1987. Aspects of the physiolog-ical basis of cultivar differences in yield of potato under droughtedand irrigated conditions. Potato Research 30: 201–217.
Kumar, R., and G.S. Kang. 2006. Usefulness of Andigena. Solanumtuberosum ssp. andigena. genotypes as parents in breeding earlybulking potato cultivars. Euphytica 150: 107–115.
Lafitte, R., A. Blum, and G. Courtois. 2003. Secondary traits to help identifydrought-tolerant genotypes. In Breeding rice for drought-proneenvironments, ed. K.S. Fischer, R. Lafitte, S. Fukai, Q. Atlin, and B.Hardy, 37–48. Los Baños: International Rice Research Institute.
Ochoa, C. 2003. Las papas del Perú. Base de datos 1947–1997, Lima.Perú. Centro Internacional de la Papa. CIP. - Universidad NacionalAgraria La Molina. UNALM. - Agencia Suiza para el Desarrollo yla Cooperación. COSUDE. 185p.
Ortiz, R., and A.M. Golmirzaie. 2002. Genetic parameters for agronomiccharacteristics. II. Intermediate and advanced stages in a true potatoseed breeding population. Hereditas 139: 217–222.
Ritter, E., L. Barandella, R. López, and J.I. Ruiz de Galaretta. 2008.Exploitation of exotic, cultivated Solanum germplasm for breedingand commercial purposes. Potato Research 51: 301–311.
Rosielle, A.A., and J. Hamblin. 1981. Theoretical aspects of selection foryield in stress and non-stress environments.Crop Science 21: 943–946.
Spitters, C.J.T., and A.H.C.M. Schapendonk. 1990. Evaluation of breed-ing strategies for drought tolerance in potato by means of cropgrowth simulation. Plant and Soil 123: 193–203.
Ta, J., S.K. Kaushik, J.S. Minhas, and V. Bhardwaj. 2003. The Potato:breeding for biotic and abiotic stress. New Delhi: Mehta. 446 p.
Tai, G.C.C. 1976. Estimation of general and specific combining abilitiesin potato. Canadian Journal of Genetics and Cytology 18:463–470.
Thompson, P.G., H.A. Mendoza, and R.L. Plaisted. 1983. Estimation ofgenetic parameters for characters selected for potato propagation bytrue seed in an andigena population. American Potato Journal 60:393–401.
Vasquez-Robinet, C., S.P. Mane, A.V. Ulanov, J.I. Watkinson, V.K.Stromberg, D. De Koeyer, R. Schafleitner, D.B. Willmot, M.Bonierbale, and H.J. Bohnert. 2008. Physiological and molecularadaptations to drought in Andean potato genotypes. Journal ofExperimental Botany 59: 2109–2123.
Watkinson, J.I., L. Hendricks, A.A. Sioson, C. Vasquez-Robinet, V.Stromberg, L.S. Heath, M. Schuler, H.J. Bohnert, M. Bonierbale,and R. Grene. 2006. Accessions of Solanum tuberosum ssp.andigena show differences in photosynthetic recovery after droughtstress as reflected in gene expression profiles. Plant Science 171:745–758.
Weisz, R., J. Kaminski, and Z. Smilowitz. 1994. Water-deficit effects onpotato leaf growth and transpiration: utilizing fraction extractablesoil-water for comparison with other crops. American PotatoJournal 71: 829–840.
Am. J. Potato Res.





























