Embed Size (px)
Citation preview

Molecular Cloning and Characterization of Human Trabeculin-a, aGiant Protein Defining a New Family of Actin-binding Proteins*
(Received for publication, August 20, 1999)
Yaping Sun, Jinyang Zhang, Stine-Kathrein Kraeft, Daniel Auclair, Mau-Sun Chang, Yuan Liu,Rebecca Sutherland, Ravi Salgia, James D. Griffin, Louis H. Ferland, and Lan Bo Chen‡
From the Dana Farber Cancer Institute, Harvard Medical School, Boston, Massachusetts 02115
We describe the molecular cloning and characteriza-tion of a novel giant human cytoplasmic protein, trabec-ulin-a (Mr 5 614,000). Analysis of the deduced amino acidsequence reveals homologies with several putative func-tional domains, including a pair of a-actinin-like actinbinding domains; regions of homology to plakins at ei-ther end of the giant polypeptide; 29 copies of a spectrin-like motif in the central region of the protein; two po-tential Ca21-binding EF-hand motifs; and a Ser-richregion containing a repeated GSRX motif. With similar-ities to both plakins and spectrins, trabeculin-a appearsto have evolved as a hybrid of these two families ofproteins. The functionality of the actin binding domainslocated near the N terminus was confirmed with anF-actin binding assay using glutathione S-transferasefusion proteins comprising amino acids 9–486 of thededuced peptide. Northern and Western blotting andimmunofluorescence studies suggest that trabeculin isubiquitously expressed and is distributed throughoutthe cytoplasm, though the protein was found to begreatly up-regulated upon differentiation of myoblastsinto myotubes. Finally, the presence of cDNAs similar to,yet distinct from, trabeculin-a in both human and mousesuggests that trabeculins may form a new subfamily ofgiant actin-binding/cytoskeletal cross-linking proteins.
The cellular cytoskeleton is a highly organized structure com-posed of three types of filaments: microfilaments, whose principalpolymeric component is actin; intermediate filaments, whichcome in vimentin, keratin, and other “flavors”; and microtubulesmade of polymeric tubulin. A large number of support proteinsare associated with these filaments, which together define cellsize and shape, both during development and once cells haveattained their terminally differentiated state. The cytoskeleton isboth static and adaptive, providing a dynamic cellular architec-ture that is capable of receiving and responding to informationfrom the environment. This adaptability is determined by theability of cytoskeletal proteins that form the supramoleculararrays, such as plakins, spectrins, and many others, to act astargets or intermediates in signal transduction pathways (1–3).Although the complexity of the cytoskeleton is not fully de-scribed and novel components certainly remain to be identified, adetailed understanding of the architecture of supramolecularcomplexes and the structure-function relationships of the compo-
nent molecules has been achieved in some cases. For example,the organization of the muscle sarcomere, with each unit com-prised of over 30 individual polypeptides, has been elucidated (4).More variant structures such as focal contacts are also wellunderstood, both at the level of their basic molecular componentsand their recruitment of signaling and structural proteins, whichvaries depending on the status of the cell (5–7). The regulationand assembly of actin filament bundles during growth, develop-ment, and adaptive responses to extracellular stimuli (e.g. stressfiber formation) has also been studied extensively (8–11).
Plakins are a family of large cytoskeletal proteins (.200kDa) that serve as cross-linkers between cytoskeletal filamentsand in some cases as filament attachment points with theplasma membrane (12). The family currently comprises fourprincipal members: desmoplakin, plectin, bullous pemphigoidantigen 1 (BPAG1), and envoplakin. All plakins share generalstructural features, with globular N- and C-terminal regionsseparated by a central rod domain. There is also extensiveconservation of sequence and function between the members ofthe family. The C terminus of all plakins binds intermediatefilaments and has conserved “helix/b-turn” motifs. The N ter-minus is responsible for the localization to plasma membranesites and/or mediates interaction with other cytoskeletal fila-ments. All plakins share a N-terminal microtubule bindingdomain (though not all plakins have been observed to associatewith microtubules) (12, 13), and plectin and BPAG1 have anactin-binding domain in this region (12). The central rod do-main, finally, mediates homodimerization by forming an anti-parallel a-helical coiled-coil structure with two plakin mole-cules (14). Thus, plakins appear to be asymmetric cross-linkingproteins but may be able to achieve functional symmetrythrough dimerization. In addition, at least plectin and desmo-plakin are phosphorylated, and in the latter case this seems toaffect keratin binding (15, 16).
The spectrin family, which also includes fodrin, dystrophin,utrophin, and protein 4.1, is a group of actin-binding proteinsthat appear to have evolved from an a-actinin ancestral genethrough a series of elongation and duplication events on thebasic repeat (17, 18). As such, the presence of an actin bindingdomain (calponin homology) at the N terminus and of a cal-modulin-like Ca21-binding site near the C terminus, both pres-ent on a-actinin, is a hallmark of this protein family. Thespectrin repeat motif folds into a triple-helix coiled-coil forminga flexible rod (19), which allows spectrin and fodrin (non-eryth-roid spectrin) to provide the plasma membrane with supportand elasticity (20–22).
In this paper, we describe the molecular cloning and charac-terization of a novel human cytoplasmic giant protein, trabec-ulin-a. This protein binds F-actin, is ubiquitously expressed,and is distributed throughout the cytoplasm. It also showsstrong similarities to both plakins and spectrins and appears tohave evolved as a hybrid from the two families. Furthermore,
* The costs of publication of this article were defrayed in part by thepayment of page charges. This article must therefore be hereby marked“advertisement” in accordance with 18 U.S.C. Section 1734 solely toindicate this fact.
The nucleotide sequence(s) reported in this paper has been submittedto the GenBankTM/EBI Data Bank with accession number(s) AF141968.
‡ To whom correspondence should be addressed: Dana Farber CancerInstitute, Harvard Medical School, 44 Binney St., Boston, MA 02115Tel.: 617-632-3386; Fax: 617-632-4470; E-mail: [email protected].
THE JOURNAL OF BIOLOGICAL CHEMISTRY Vol. 274, No. 47, Issue of November 19, pp. 33522–33530, 1999© 1999 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in U.S.A.
This paper is available on line at http://www.jbc.org33522
by guest on April 4, 2019
http://ww
w.jbc.org/
Dow
nloaded from

the presence of similar yet distinct cDNAs in both humans andmice suggests that trabeculins may form a new subfamily ofgiant actin-binding/cytoskeletal cross-linking proteins.
EXPERIMENTAL PROCEDURES
Cell Culture—The MRC-5 (human lung fibroblast), CCL-105 (humanadrenal cortex carcinoma), and BSC-1 (African green monkey kidneyepithelium) cell lines were purchased from ATCC, and the FS-2 (humanforeskin fibroblasts) and JMN (human mesothelioma) cells were kindlyprovided by the late Dr. Ruth Sager (Dana-Farber Cancer Institute)and Dr. Jim Rheinwald (Brigham and Women’s Hospital), respectively.These cell lines were maintained in Dulbecco’s modified essential me-dium (DMEM)1 supplemented with 10% calf serum, 1 mM glutamine, 10units/ml penicillin, and 0.1 mg/ml streptomycin. The C2C12 (murinemyoblast) line was from ATCC and cultured in DMEM supplementedwith 20% fetal calf serum, 1 mM glutamine, 10 units/ml penicillin, and0.1 mg/ml streptomycin. Preparation of myoblasts and differentiationinto myotubes was carried out as described (23, 24); to induce myoblastfusion, the medium was changed to DMEM with 2% horse serum(differentiation medium, DM).
Northern Blot Analysis—Multiple tissue Northern blots were pur-chased from CLONTECH and used according to the manufacturer’sinstructions. Total RNA from primary myoblasts and differentiatedmyotubes was prepared using RNazol. Northern blots were preparedand probed as described previously (23). The trabeculin probe used wasabout 3 kilobase pairs in length and spanned the region from 996 to4082 bp of the trabeculin-a cDNA, corresponding to amino acids9–1036.
Generation of Trabeculin Antibodies—GST fusion proteins of tra-beculin fragments 4HP6 (amino acids 2933–3558) and 4HP12 (aminoacids 1868–2230) were generated by subcloning of the relevant frag-ment into vector pGEX3T (CLONTECH). Purification of GST fusionproteins from Escherichia coli was carried out as described previously(25). Briefly, the transformed bacterial cell culture (DH5a) was grownovernight in L broth supplemented with 50 mg/ml ampicillin (LB-Amp),diluted 1:10 in fresh LB-Amp, and further grown for 1 h at 37 °C.Protein production was induced by adding 1 mM isopropyl-1-thio-b-D-galactopyranoside for 2 h at 37 °C. Cells were collected and resus-pended in PBS before sonication and centrifugation at 10,000 3 g for 10min. The supernatant was incubated with glutathione-agarose beads tocapture fusion proteins. The glutathione-agarose beads were washedwith PBS, and fusion proteins were eluted with free glutathione (5 mM
glutathione in 50 mM Tris, pH 8.0). Cleavage of trabeculin fragmentsfrom GST was carried out using either thrombin or factor Xa, and thepurified fragments were mixed and used to raise an antiserum inrabbits (polyclonal B13 antiserum).
Western Blot Analysis—Post-nuclear lysates were prepared from celllines or mouse brain homogenates, as follows. Cells were lysed in abuffer containing 50 mM Hepes, pH 7.4, 100 mM NaCl, 10 mM NaF, 10mM iodoacetamide, 1% Triton X-100, and the “complete” protease in-hibitor mixture from Roche Molecular Biochemicals. Nuclei and celldebris were removed by centrifugation, and either total supernatantproteins were precipitated using acetone or immunoprecipitation wasperformed. In the latter case cell lysates were precleared with proteinA-Sepharose (Amersham Pharmacia Biotech) for 30 min before immu-noprecipitation of specific proteins with immune serum. Immunopre-cipitates made with preimmune serum were used as control. Proteinswere resolved by 6% SDS-PAGE and transferred to polyvinylidenedifluoride membranes (Millipore). Membranes were blocked using 5%BSA before probing with the relevant antibody. The secondary sheepanti-rabbit antibody conjugated to horseradish peroxidase (AmershamPharmacia Biotech) was used at 1:25,000 dilution, and the blots weredeveloped by chemiluminescence using the ECL kit from AmershamPharmacia Biotech.
Immunofluorescence—Cells were grown on glass coverslips and fixedin 220 °C methanol for 5 min before washing in PBS. Staining withrabbit anti-trabeculin antiserum (polyclonal B13) was carried out at37 °C for 1 h. The secondary rhodamine-conjugated anti-rabbit antibody(Jackson ImmunoResearch, West Grove, PA) was used at 100 ng/ml andapplied for 30 min at 37 °C, before washing and mounting in glycerol/
gelatin (Sigma). For actin staining, cells were fixed in 2% paraformal-dehyde at room temperature for 20 min, followed by washing in PBSand 10 min incubation in sucrose buffer (10 mM Hepes, 3 mM MgCl2, 50mM NaCl, 300 mM sucrose, 0.5% Triton X-100). Actin fibers were thenlabeled with Texas Red-conjugated phalloidin (Molecular Probes, Eu-gene, OR) at 37 °C for 20 min. For microtubule visualization, cells wereincubated with the cross-linking agent, dithiobis(succinimidyl propio-nate) at 0.2 mg/ml for 5 min at room temperature, and washed in PBSand microtubule stabilizing buffer (MTSB: 100 mM PIPES, 1 mM EGTA,4% polyethylene glycol 8000). Cells were then permeabilized with 0.5%Triton X-100, washed twice in PBS, and fixed in ice-cold methanol for 5min. The anti-a-tubulin monoclonal antibody (Sigma) was applied at37 °C for 1 h, followed by incubation with secondary rhodamine-conju-gated anti-mouse antibody (Jackson ImmunoResearch), 100 ng/ml for30 min at 37 °C, and then washing and mounting in glycerol/gelatin(Sigma).
Cells were examined using an LSM410 confocal laser scanning mi-croscope (Carl Zeiss, Germany) equipped with an external argon-kryp-ton laser (568 nm). Optical sections of 512 3 512 pixels were digitallyrecorded in the 23 line-averaging mode. Images were processed forreproduction using the Photoshop software (Adobe Systems, Moun-tainview, CA).
Actin Filaments Co-sedimentation Assay—An actin binding assaysystem (Cytoskeleton, Inc.) was used to examine the ability of GST-TrABD, a GST fusion protein comprising trabeculin’s a-actinin-likeactin binding domains (amino acids 9–486), to bind to F-actin. TheGST-TrABD fusion protein was generated as described above. a-Actininand BSA (supplied with the assay kit) were used as positive andnegative controls, respectively, and the assay was carried out essen-tially as described (24). Recombinant proteins were centrifuged (20 min,28 pounds/square inch; ;100,000 3 g) in a Beckman Airfuge to removeaggregates. F-actin was prepared by resuspension in General ActinBuffer (supplied with the kit) and polymerization at room temperaturefor 1 h. The test proteins (.20 mM) were mixed with F-actin (23 mM) andincubated at room temperature for 30 min. Filaments were pelleted bycentrifugation at 150,000 3 g for 1.5 h at 24 °C. Supernatants andpellets were separated, and the proteins present in each were analyzedby 10% SDS-PAGE and Coomassie Blue staining. Actin-binding pro-teins and F-actin were present in the pellet, whereas proteins that didnot associate with actin remained in the supernatant.
Cytoskeletal Disruption—Disruption of microtubules was induced inCCL-105 cells, with 2 mg/ml nocodazole (Sigma) applied for 2 h at 37 °C,and disruption of actin fibers with 1 mM cytochalasin D (Sigma) underthe same conditions. Cells were then washed, fixed, and labeled asdescribed before.
RESULTS
Molecular Cloning of Trabeculin-a—Human trabeculin-awas cloned from the HL1131b human prostate cDNA library(23, 26). Sixteen contiguous partial cDNAs were identified thatenabled assembly of the full-length trabeculin-a cDNA struc-ture (Fig. 1A); the composite trabeculin-a cDNA sequence hasbeen deposited in GenBankTM (accession number AF141968).The total sequence identified in our screening was 17,749 bp,including a 16,122-bp open reading frame, a 973-bp 59-untrans-lated region, and a 654-bp 39-untranslated region. The deducedamino acid sequence of the open reading frame reveals a pro-tein of 5373 amino acids, with a predicted Mr of 614,000. Fig.1B shows the predicted amino acid sequence of human tra-beculin-a. Fluorescence in situ hybridization was performed tomap the human chromosomal location of the trabeculin-a gene.A 6112-bp fragment of trabeculin-a (bp 8161–14272, aminoacids 2397–4433) hybridized to a single locus, on chromosome1 region p33-p34.2 (not shown).
Since trabeculin is a novel protein, we performed a Northernblot analysis to determine the tissue distribution of its tran-scripts in humans. Fig. 1C shows the Northern blot for severalhuman tissues. The probe used in this analysis was a 3-kilo-base pair fragment of the trabeculin-a cDNA (from 996 to 4082bp). All tissues shown had detectable levels of trabeculinmRNAs, with the highest expression in muscle (heart andskeletal), prostate, gastrointestinal tract (intestine and colon),and gonads, and lowest expression in brain, spleen, thymus,
1 The abbreviations used are: DMEM, Dulbecco’s modified essentialmedium; bp, base pair(s); PIPES, 1,4-piperazinediethanesulfonic acid;GST, glutathione S-transferase; PAGE, polyacrylamide gel electro-phoresis; BSA, bovine serum albumin; PBS, phosphate-buffered saline;ABD, actin binding domain; EST, expressed sequence tag; BPAG1,bullous pemphigoid antigen 1; MTBD, microtubule binding domain.
Trabeculin-a, New Actin Cross-linking Protein 33523
by guest on April 4, 2019
http://ww
w.jbc.org/
Dow
nloaded from
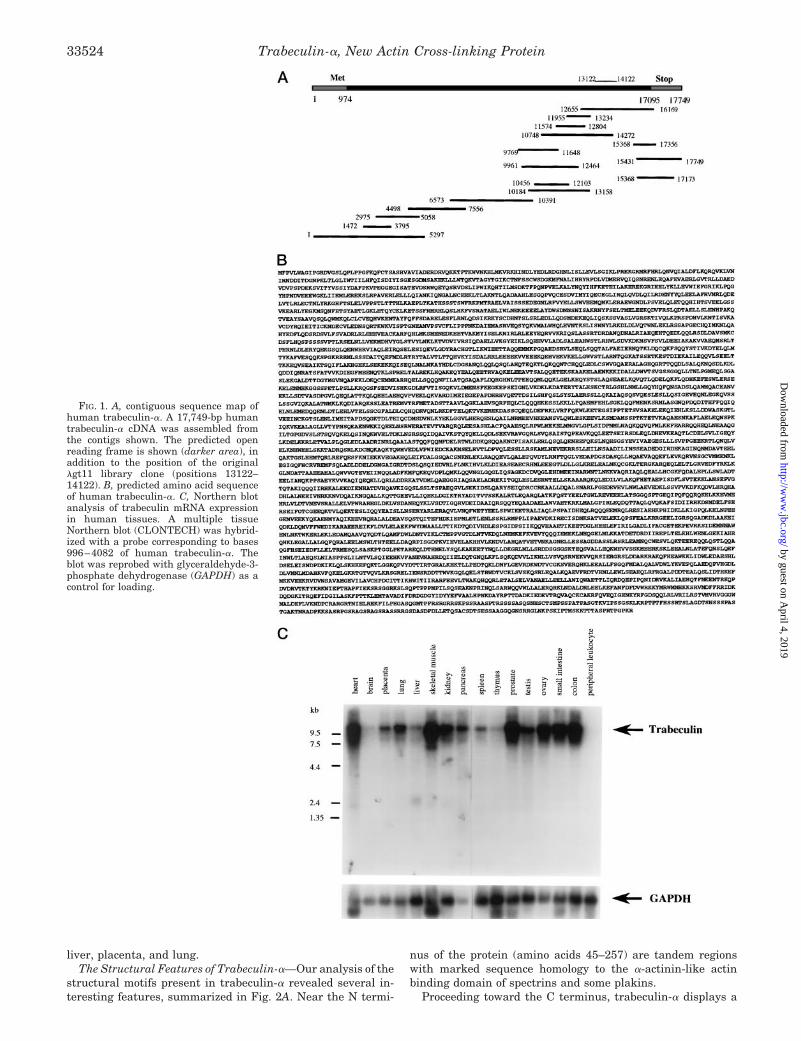
liver, placenta, and lung.The Structural Features of Trabeculin-a—Our analysis of the
structural motifs present in trabeculin-a revealed several in-teresting features, summarized in Fig. 2A. Near the N termi-
nus of the protein (amino acids 45–257) are tandem regionswith marked sequence homology to the a-actinin-like actinbinding domain of spectrins and some plakins.
Proceeding toward the C terminus, trabeculin-a displays a
FIG. 1. A, contiguous sequence map ofhuman trabeculin-a. A 17,749-bp humantrabeculin-a cDNA was assembled fromthe contigs shown. The predicted openreading frame is shown (darker area), inaddition to the position of the originallgt11 library clone (positions 13122–14122). B, predicted amino acid sequenceof human trabeculin-a. C, Northern blotanalysis of trabeculin mRNA expressionin human tissues. A multiple tissueNorthern blot (CLONTECH) was hybrid-ized with a probe corresponding to bases996–4082 of human trabeculin-a. Theblot was reprobed with glyceraldehyde-3-phosphate dehydrogenase (GAPDH) as acontrol for loading.
Trabeculin-a, New Actin Cross-linking Protein33524
by guest on April 4, 2019
http://ww
w.jbc.org/
Dow
nloaded from

region of approximately 900 amino acids that strongly resem-bles a region common to all plakins and found to be the micro-tubule binding domain (MTBD) of BPAG1 (13); and the C-terminal region of trabeculin-a contains a 76 amino acid regionwith strong homology (52% identity, 69% homology) to anotherplakin, GAR22 (27). These two plakin-like motifs, together withthe presence of an actin binding domain, suggest that trabecu-lin-a may interact with various cytoskeletal proteins.
Perhaps the most striking feature of trabeculin-a resides inits central region: a 29-fold repeat of a 110–120 amino acidstretch, bearing considerable identity to the spectrin-like re-peats found in human dystrophin and other spectrins (17, 28)(Fig. 2B). Secondary structure prediction based on the se-quence information suggest that each repeat is made up ofthree a-helices, as for the spectrin-type repeat. Similarities
with the spectrin-like repeat also include a very conservedtryptophan residue at position 18 in the alignment followed bya hydrophobic residue, conservation of positively and nega-tively charged residues (Fig. 2B, in red and purple) at givenpositions throughout the alignment, a leucine at the end ofeach repeat, and abundant prolines and glycines (Fig. 2B, inyellow) in the loop regions.
There are also multiple regions in trabeculin-a that may beable to transduce information for intracellular signaling path-ways as follows: two tandem calcium-binding EF-hand motifs(amino acids 5000–5100), similar in sequence and relativeposition to those in the C terminus of dystrophin and otherspectrins (17, 29); and several tyrosine residues that havesurrounding sequences consistent with their being tyrosinekinase substrates. Finally, the extreme C terminus of trabecu-
FIG. 2. A, schematic representation of the putative structural features human trabeculin-a. aa, amino acid. B, alignment of the 29 spectrinrepeats of human trabeculin-a. The multiple alignment was made using the CLUSTAL W program (EMBL), and the figure was prepared with theJalview editor (Michele Clamp, European Bioinformatics Institute). C and D, alignment of trabeculin homologues from human, mouse, D.melanogaster, and C. elegans, in the regions homologous to the actin binding domain of a-actinin (C) and to the plakin GAR22 (D).
Trabeculin-a, New Actin Cross-linking Protein 33525
by guest on April 4, 2019
http://ww
w.jbc.org/
Dow
nloaded from

lin-a contains a serine-rich region (amino acids 5350–5500 are28% serine), including a GSRX repeat motif also present in thesame relative position (40–50 amino acids to the C terminus) inplectin and desmoplakin.
Trabeculin Homologues and Family—Several structural ho-mologues of human trabeculin-a have been reported, including,notably, Kakapo from Drosophila melanogaster (30–32) and itsCaenorhabditis elegans counterpart, CeKak (30); and themouse ACF7 gene (GenBankTM accession number AF150755),2
which shares 88% overall identity with the human trabeculin-aclone reported here. Alignments of the deduced protein se-quences for two of the best conserved regions, the putativeactin binding domains and the GAR22 homology, are shown inFig. 2, C and D. A 1157-bp partial human ACF7 cDNA has alsobeen reported (33) which ends with an 886-bp stretch identicalto a region near the N terminus of our trabeculin-a cDNA,starting right around the beginning of the putative actin bind-ing domain.
Another partial human cDNA clone was reported recently(34) that displays 77% identity with a region near the C termi-nus of trabeculin-a, over its 1054 deduced amino acid sequence.This region includes conserved EF-hand motifs, GAR22 homol-ogy region, and GSRX repeats (Fig. 3). Correspondingly, wealso found over a dozen mouse ESTs with strong similarities tomouse ACF7, although clearly of non-identical sequence. Forexample, the longest of these ESTs (GenBankTM accessionnumber AA014416) encodes the last 134 amino acids of amouse trabeculin homologue with 75% overall identity withmouse ACF7. It is notable that these clones display a conservedGSRX repeat but not the general serine-rich C terminus (Fig. 3for human; not shown for mouse). These clones would seem tocorrespond to a separate trabeculin gene in human and mousewhich, along with trabeculin-a/ACF7, would define a new pro-tein family.
Trabeculin Is Distributed Throughout the Cytoplasm andDoes Not Form Distinct or Focal Structures—We raised anantibody (B13) to the spectrin homology region of trabeculin toperform immunofluorescence studies of the subcellular local-ization of the protein. Since this region of trabeculin revealsclose homology to many other proteins, it was critical to estab-lish the specificity of this antibody, before the subsequent anal-yses. Fig. 4A shows that in Western analysis, the B13 anti-trabeculin antibody recognizes the trabeculin portions of thebacterially expressed GST-trabeculin fusion proteins to whichit was raised. The B13 antiserum recognizes two high molecu-lar weight proteins on Western blots of cultured CCL-105 orMRC-5 cells (Fig. 4B) or of mouse brain lysate (not shown).Moreover, B13 immunoprecipitates these proteins specificallyfrom cell lysates. These data suggest that the high molecularweight proteins are in fact trabeculins and that the B13 anti-trabeculin antibody does not significantly cross-react withother proteins.
We performed an immunofluorescence analysis of variouscell types using the B13 anti-trabeculin antibody. A represent-ative selection of these data are shown in Fig. 4, C—F. Thesefigures show the staining pattern of anti-trabeculin in theCCL-105, BSC-1, JMN, and FS-2 cell lines. As a negativecontrol, staining with the preimmune serum was performed,and only minimal background staining was observed (data notshown). In these and the more than 20 other cell types weexamined (not shown), the subcellular distribution of anti-trabeculin staining had the following salient features: 1) dis-tribution throughout the cytoplasm, 2) exclusion from the nu-
cleus, and 3) appearance as a filamentous network, in somecells displaying a fiber-like staining pattern reminiscent ofactin stress fibers. Occasionally, we observed a punctate orglobular staining that might be associated with localization tovesicular structures like the Golgi apparatus or endoplasmicreticulum (not shown).
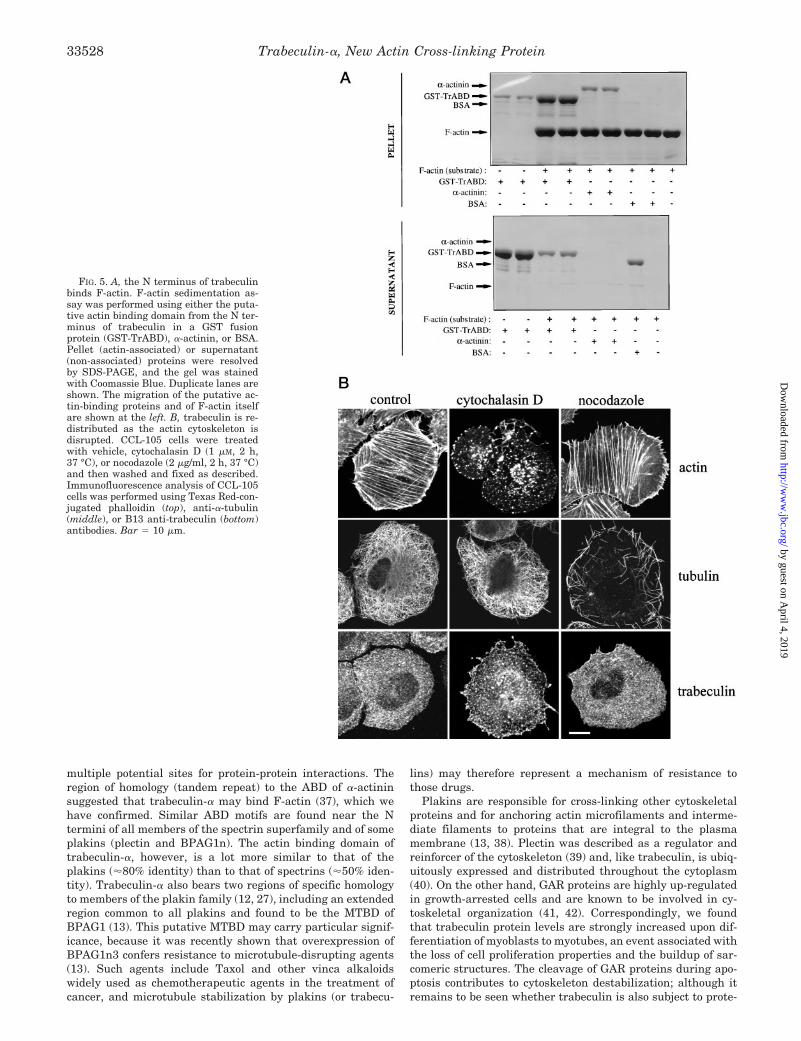
Trabeculin-a Binds F-actin through Its N-terminal a-Acti-nin-like Domains—Since the N terminus of trabeculin-a con-tains a domain that has significant homology to the a-actinin-like actin binding domain, we constructed a GST fusion proteinof the putative N-terminal actin binding domain (ABD) of tra-beculin-a (amino acids 9–486), GST-TrABD (82 kDa by SDS-PAGE). We then tested the ability of GST-TrABD to bindF-actin in a sedimentation binding assay (24).
Fig. 5A shows that GST-TrABD efficiently binds F-actinfilaments. In this experiment, GST-TrABD, positive (a-actinin)or negative (BSA) control proteins were incubated with F-actin,
2 C. Leung, D. Sun, M. Zheng, D. Knowles, and R. K. H. Liem, EMBLaccession number AF150755.
FIG. 3. Alignment of the cDNA sequences for human trabecu-lin-a and -b, limited to the region for which trabeculin-b se-quence is available (1054 amino acids of the C terminus). Con-served residues are boxed, and conserved domains are identified.
Trabeculin-a, New Actin Cross-linking Protein33526
by guest on April 4, 2019
http://ww
w.jbc.org/
Dow
nloaded from

and the reactions were centrifuged to separate binding fromnon-binding proteins. Both GST-TrABD and a-actinin, but notBSA, co-precipitated with F-actin. We verified separately thatGST does not bind actin (not shown).
Effects of Nocodazole and Cytochalasin D—We also testedwhether an association of GST-TrABD with actin could bedetected in a cellular context. Due to the large size and modularnature of trabeculin, an approach where association is detectedby co-immunoprecipitation was not feasible. Therefore, we ex-amined the pattern of trabeculin distribution in the absenceand presence of drugs that disrupt the cytoskeleton. We stainedfor actin, tubulin, and trabeculin in CCL-105 cells treated witheither cytochalasin D, which disrupts microfilaments (35), ornocodazole, a disrupter of microtubules (36). Fig. 5B shows thatcytochalasin D effectively disrupts actin-containing microfila-ments, causing a transition from prominently filamentous topunctate actin staining, with no remaining evidence of linearstructures. Cytochalasin D has no effect on the organization ofmicrotubules, as observed with tubulin staining. In contrast,nocodazole disrupts tubulin-containing microtubules but doesnot change actin filament organization. We stained cells fortrabeculin in the absence or presence of either of these drugs.In untreated cells, the ubiquitous, fine filament structure oftrabeculin was evident. In cells treated with nocodazole, nochange in the distribution of trabeculin staining was observed.However, cytochalasin D treatment resulted in a marked (al-though not complete) disruption of trabeculin fine filamentstructures and the appearance of punctate aggregates of tra-beculin. These results suggest that trabeculin is at least in part
associated with actin filaments.Myoblast to Myotube Transition Involves an Up-regulation in
Trabeculin Levels—Since trabeculin appears to be an actin-binding protein and myogenesis involves a major increase inactin expression, we examined the levels of trabeculin in myo-blasts undergoing differentiation to myotubes. C2C12 cellswere placed in differentiation medium (DM) and monitoredover a 6-day period for morphology (not shown) and trabeculinexpression. Fig. 6A shows that trabeculin mRNA levels in-crease steadily from Day 0 (100% myoblasts) to Day 5–6 (fusedmyotubes). In contrast, myosin levels, reflecting the differenti-ation to functional muscle, only increase toward the end of thefusion process, when the cells develop their contractileapparatus.
Fig. 6B illustrates the difference in trabeculin expressionlevels between myoblasts and myotubes. A mixture of myo-blasts and myotubes were stained with the B13 anti-trabeculinantibody. As above, trabeculin is distributed throughout thecytoplasm and is excluded from the nuclei in all cells. There isa striking difference, however, between the levels of staining inthe fused myotubes compared with the individual myoblasts.Taken together with the Northern blot data of Fig. 6A, theseresults suggest that myoblast fusion is accompanied by anup-regulation of trabeculin levels.
DISCUSSION
The data presented above describe the molecular cloning andinitial analysis of a novel human cytoskeletal protein, trabecu-lin-a. The domain structure of trabeculin-a is complex, with
FIG. 4. A, the B13 anti-trabeculin anti-body is reactive against the fusion pro-teins to which it was raised. Two GSTfusion proteins (4HP6 and 4HP12) of tra-beculin were generated, mixed together,and used as immunogens in rabbits. Theresulting immune serum (B13) was thenused at 1 mg/ml in Western blot analysis.Samples of the GST-trabeculins were ei-ther left intact (Int) or cleaved using Fac-tor Xa (Xa) or thrombin (Th) to separateGST from the trabeculin fragment. Thesamples were resolved by SDS-PAGE,transferred to polyvinylidene difluoridemembrane, and probed with B13 anti-tra-beculin. B13 anti-trabeculin recognizesboth fusion proteins before and aftercleavage, thus the relevant epitope is notin the GST portion of the fusion. B, theB13 anti-trabeculin antibody immunopre-cipitates (IP) and recognizes two high mo-lecular weight proteins from cell lysates.Cell lysates were prepared from CCL-105or MRC-5 tumor cells. Immunoprecipita-tion was performed with either preim-mune serum (PI) or immune serum B13(I). Proteins from immunoprecipitationsor post-nuclear lysate (lys) were resolvedby SDS-PAGE and transferred to polyvi-nylidene difluoride membranes for West-ern analysis with 1 mg/ml B13 anti-trabec-ulin. C—F, immunofluorescence analysisof trabeculin subcellular localization.Cells were prepared as described under“Experimental Procedures.” Staining wascarried out using the B13 anti-trabeculinantibody, with appropriate preimmuneserum-staining controls (not shown). C,CCL-105, human adrenal cortex carcino-ma; D, BSC-1, African green monkey kid-ney epithelia; E, JMN, human mesotheli-oma; F, FS-2, human foreskin fibroblasts.Bar in E 5 10 mm; bar in F 5 25 mm.
Trabeculin-a, New Actin Cross-linking Protein 33527
by guest on April 4, 2019
http://ww
w.jbc.org/
Dow
nloaded from
multiple potential sites for protein-protein interactions. Theregion of homology (tandem repeat) to the ABD of a-actininsuggested that trabeculin-a may bind F-actin (37), which wehave confirmed. Similar ABD motifs are found near the Ntermini of all members of the spectrin superfamily and of someplakins (plectin and BPAG1n). The actin binding domain oftrabeculin-a, however, is a lot more similar to that of theplakins ('80% identity) than to that of spectrins ('50% iden-tity). Trabeculin-a also bears two regions of specific homologyto members of the plakin family (12, 27), including an extendedregion common to all plakins and found to be the MTBD ofBPAG1 (13). This putative MTBD may carry particular signif-icance, because it was recently shown that overexpression ofBPAG1n3 confers resistance to microtubule-disrupting agents(13). Such agents include Taxol and other vinca alkaloidswidely used as chemotherapeutic agents in the treatment ofcancer, and microtubule stabilization by plakins (or trabecu-
lins) may therefore represent a mechanism of resistance tothose drugs.
Plakins are responsible for cross-linking other cytoskeletalproteins and for anchoring actin microfilaments and interme-diate filaments to proteins that are integral to the plasmamembrane (13, 38). Plectin was described as a regulator andreinforcer of the cytoskeleton (39) and, like trabeculin, is ubiq-uitously expressed and distributed throughout the cytoplasm(40). On the other hand, GAR proteins are highly up-regulatedin growth-arrested cells and are known to be involved in cy-toskeletal organization (41, 42). Correspondingly, we foundthat trabeculin protein levels are strongly increased upon dif-ferentiation of myoblasts to myotubes, an event associated withthe loss of cell proliferation properties and the buildup of sar-comeric structures. The cleavage of GAR proteins during apo-ptosis contributes to cytoskeleton destabilization; although itremains to be seen whether trabeculin is also subject to prote-
FIG. 5. A, the N terminus of trabeculinbinds F-actin. F-actin sedimentation as-say was performed using either the puta-tive actin binding domain from the N ter-minus of trabeculin in a GST fusionprotein (GST-TrABD), a-actinin, or BSA.Pellet (actin-associated) or supernatant(non-associated) proteins were resolvedby SDS-PAGE, and the gel was stainedwith Coomassie Blue. Duplicate lanes areshown. The migration of the putative ac-tin-binding proteins and of F-actin itselfare shown at the left. B, trabeculin is re-distributed as the actin cytoskeleton isdisrupted. CCL-105 cells were treatedwith vehicle, cytochalasin D (1 mM, 2 h,37 °C), or nocodazole (2 mg/ml, 2 h, 37 °C)and then washed and fixed as described.Immunofluorescence analysis of CCL-105cells was performed using Texas Red-con-jugated phalloidin (top), anti-a-tubulin(middle), or B13 anti-trabeculin (bottom)antibodies. Bar 5 10 mm.
Trabeculin-a, New Actin Cross-linking Protein33528
by guest on April 4, 2019
http://ww
w.jbc.org/
Dow
nloaded from

olysis in these circumstances, it may be noteworthy that theGAR homology region of trabeculin-a does cover the GAR con-sensus region for the activity of aspartate-directed cysteineproteases, which mediate apoptosis (42).
The most striking structural feature of trabeculin-a is per-haps the 29-repeat of the spectrin motif, which occupies about60% of the primary sequence of the whole protein. The spectrin-type repeat motif folds into a triple a-helical coiled-coil (28)forming a semi-rigid rod-like structure (19, 43, 44). Thus, tra-beculin molecules may bind at intervals along filaments ofactin or other cytoskeletal targets, such that homo- or hetero-filament bridges would be formed by the interactions of cy-toskeletal proteins with cognate binding sites on trabeculinnear its N and C termini. The separation of these target mol-ecules (or filaments) would thus be determined by the rigidstructure formed by the spectrin-like repeats in the centralregion of trabeculin.
The general structure of trabeculin-a globular N and C ter-mini separated by a rod domain and the plakin homologyregions might suggest that trabeculin-a is in fact a new mem-ber of the plakin family. However, the rod domain of trabecu-lin-a is not akin to that of plakins, but is spectrin-like. Also, allfour classical plakins bind intermediate filament through ana-helix/b-turn motif (often repeated) located at their C termi-nus (12), which trabeculin-a lacks. With some respects, there-fore, trabeculin-a appears to be more similar to members of thespectrin protein family. All these proteins have the generalstructure N terminus/ABD/spectrin repeat/Ca21-binding do-main/C-terminus (17), although as mentioned above, the actin-
binding domain of trabeculin-a is more similar to that of theplakins that carry it (plectin and BPAG1n) than to that ofspectrins. Taken together, these structural observations sug-gest that this novel protein may be a “hybrid” with plakin-likeN (ABD and MTBD) and C termini (GAR22 and Ser-rich/GSRXrepeat) and a spectrin-like rod domain plus adjacent sequence(including the calmodulin homology) in the middle.
During our cloning and analysis of human trabeculin-a, theD. melanogaster protein Kakapo was described in three studiesreporting its cloning and functional analysis (30–32). Kakapo(45) is a 5437-amino acid protein with sequence and structuralsimilarities to trabeculin-a. Similarities include putative actin-binding motifs near its N terminus, two plakin homology re-gions (common MTBD and Gas2/GAR22), and Ca21-bindingEF-hand motifs. Both proteins also have the critical spectrin-type a-helical repeats (22 in Kakapo, 29 in trabeculin) formingthe major part of their sequence. Functional analysis of Kakapoin Drosophila embryos (30–32) suggested a role for Kakapo inanchoring muscle cells to the epidermis. The model proposed inthese studies places Kakapo at the termini of microtubules inthe epidermal cells, linking these and the cortical actin cy-toskeleton to integrins in the epidermal basolateral membrane(the well conserved Gas2/GAR22 homology domain seems im-portant for this interaction, as mutants in this region fail todifferentiate properly). In turn, these integrins are suggestedto interact with extracellular matrix proteins that form bridgeswith the muscle cells.
Despite their very similar domain structures, trabeculin-adoes not appear to be the functional homologue of Drosophila’sKakapo in the human system; trabeculin does not significantlylocalize to focal complexes and is distributed throughout thecytoplasm. In contrast, Kakapo is found exclusively at the cellperiphery. Moreover, trabeculin is apparently not associatedwith the microtubule system since the disruption of these fila-ments does not cause a redistribution of trabeculin. Rather, theidentification of two distinct trabeculin/ACF7 genes in humanand mouse, and of alternatively spliced isoforms at least inmouse, points to the existence of a family of trabeculins whosemembers may have characteristic functions and distribution.We propose the nomenclature trabeculin-a for the human genereported here and its mouse homologue, ACF7 (GenBankTM
accession number AF150755)2 with its splice variants (46), andtrabeculin-b for the human gene cloned by Nagase et al. (34)and its mouse counterpart defined by EST AA014416 and oth-ers (GenBankTM). It is not clear from simple sequence analysiswhether Kakapo from Drosophila and CeKak from C. elegans(30) may truly be assigned to either of these classes, and itshould be noted that our searches revealed only one putativetrabeculin homologue in the completely sequenced genome ofC. elegans.
The existence of different trabeculin isoforms may also ex-plain some of the unexpected observations reported herein.First, Western blotting of CCL-105 and MRC-5 cells post-nu-clear protein preparations revealed two distinct anti-trabecu-lin-reactive bands, and these may be related to trabeculins a
and b. Second, some of the anti-trabeculin-reactive filamentousstructures seemed to resist cytochalasin D treatment, eventhough the actin-containing filaments were completely de-stroyed; it is possible that some trabeculin isoforms are notassociated with actin microfilaments. Indeed, some of themouse ACF7 isoforms reported by Bernier et al. (46) lack theN-terminal a-actinin-like actin binding domains; similarly, anisoform of the plakin BPAG1, BPAG1n3 (13), and an alterna-tive transcript of Kakapo, “Kakapo form B” (30), have beendescribed which lack part of this domain and have impairedactin binding ability. (It should also be noted that our anti-
FIG. 6. A, Northern analysis of trabeculin expression during myoblastdifferentiation. Myoblasts were differentiated into multinucleated myo-tubes over 6 days in differentiation medium (DM). mRNA was preparedfrom cells harvested at daily intervals, and Northern analysis using theindicated probes was performed. Trabeculin protein is more abundantin differentiated myotubes. B, immunofluorescent staining of mixedmyoblasts and myotubes (cultured in DM for 6 days) using anti-trabe-culin antibody. Trabeculin protein is more abundant in differentiatedmyotubes. Undifferentiated myoblasts are faintly labeled, whereasmultinucleated myotubes stain very brightly with anti-trabeculin anti-bodies. Bar 5 25 mm.
Trabeculin-a, New Actin Cross-linking Protein 33529
by guest on April 4, 2019
http://ww
w.jbc.org/
Dow
nloaded from

trabeculin polyclonal B13 antiserum was raised against thespectrin repeat region. Since such spectrin repeats are likely tobe present in both trabeculin-a and -b subfamilies, our anti-serum would likely label both gene products and their vari-ants.) Third, the divergence in the 59-ends of human trabecu-lin-a and ACF7 cDNAs may result from the alternative splicingof a common primary transcript or by the activity of alternativepromoters, with the N terminus of trabeculin-a being replacedby another sequence in the ACF7 transcript.
These possibilities were also evoked by Byers et al. (33), andboth a multiplicity of alternatively spliced transcripts and thepresence of multiple promoters appear to be commonplaceamong plakins (12, 13). As mentioned earlier, three isoforms ofthe murine ACF7 have also been described, which apparentlyresult from differential splicing of exons near the N terminus,encompassing part of the a-actinin-like actin-binding motifs(46).
Similarly, the common sequence between the two humanclones of trabeculin-a (the full-length sequence reported hereand the partial, N-terminal human ACF7 clone of Byers et al.(33)) starts at an Asp residue near the beginning of the a-acti-nin-like actin binding domain homology. It may be noteworthythat both cDNAs exhibit an in-frame ATG not far upstream ofthat point: 114 bp/38 amino acids for our trabeculin-a clone,and 219 bp/73 amino acids for human ACF7.
It is thus possible that different isoforms of trabeculin existthat bind different targets and perhaps serve different func-tions. Alternatively, the specificity/affinity of the terminal in-teractions of trabeculin may also be modified by regulatorydomains within the trabeculin molecule or by associated adapt-ers. Trabeculin indeed has multiple putative sites for the re-ceipt or transduction of extracellular signals, including puta-tive tyrosine phosphorylation sites and calcium-bindingdomains. The serine-rich region at the C terminus of trabecu-lin-a is also a likely target for phosphorylation. Phosphoryla-tion of a similar serine-rich domain in the C-terminal region ofthe b-chain of spectrin was reported to regulate the mechanicalstability of erythrocyte membranes (47). In metabolic labelingexperiments using [g-32P]ATP followed by immunoprecipita-tion with the B13 antiserum, we observed that trabeculin wasindeed heavily phosphorylated (data not shown). The non-con-servation of the C-terminal Ser-rich region among trabeculinsa and b in both human and mouse may point to a site offunctional differentiation between the two trabeculin isoforms;conversely, the conservation of the GSRX repeat within thisSer-rich region suggests the possibility of a yet undiscoveredfunctionality for this motif.
It will be important to determine the type(s), if any, of signaltransduction that may be carried out by trabeculin, sincechanges in the behavior of such proteins in response to extra-cellular stimuli may have profound morphological and evendevelopmental consequences for the cell. If this is the case, anygenetic alteration of trabeculin function could have seriousconsequences in vivo. Partial or total loss of function mutationsmay result in profound defects in the organization and main-tenance of the actin cytoskeleton. In this light, it is noteworthythat chromosome 1 region p33-p34.2, where the trabeculinlocus is found, is often deleted in human dystrophies. (Byers etal. (33) also localized ACF7/trabeculin-a to human chromosome1.) With only a portion of dystrophies directly attributable todystrophin defects, trabeculin may be viewed as a candidategene for these disorders.
In summary, we have described the novel human cytoskel-etal giant protein, trabeculin-a, a member of a new family ofactin-binding proteins. Trabeculins are widely expressed andare distributed throughout the cytoplasm of all cells examined.
Trabeculin-a can bind actin filaments and possibly other cy-toskeletal target molecules, possibly contributing to their spa-tial separation by virtue of the rigid structure formed by themultiple spectrin-type repeats in its center. In addition, trabe-culin-a contains several putative sites that may be involved inthe transduction of signals to intracellular pathways. Expres-sion of trabeculin may also be linked to the differentiation/growth status of muscle cells, as its expression levels aregreatly increased upon myogenic differentiation. Further studyof this new giant actin-binding protein family may lend insightinto novel aspects of cellular architecture.
Acknowledgment—We thank Dr. Helen Turner for valuable assist-ance in the preparation of this manuscript.
REFERENCES
1. Schmidt, A., and Hall, M. N. (1998) Annu. Rev. Cell Dev. Biol. 14, 305–3382. Hall, A. (1994) Annu. Rev. Cell Biol. 10, 31–543. Hall, A. (1998) Science 279, 509–5144. Squire, J. M. (1997) Curr. Opin. Struct. Biol. 7, 247–2575. Burridge, K., Fath, K., Kelly, T., Nuckolls, G., and Turner, C. (1988) Annu.
Rev. Cell Biol. 4, 487–5256. Lo, S. H., and Chen, L. B. (1994) Cancer Metastasis Rev. 13, 9–247. Yamada, K. M., and Geiger, B. (1997) Curr. Opin. Cell Biol. 9, 76–858. Lamarche, N., Tapon, N., Stowers, L., Burbelo, P. D., Aspenstrom, P., Bridges,
T., Chant, J., and Hall, A. (1996) Cell 87, 519–5299. Nobes, C. D., and Hall, A. (1995) Cell 81, 53–62
10. Ridley, A. J., and Hall, A. (1992) Cell 70, 389–39911. Ridley, A. J., Paterson, H. F., Johnston, C. L., Diekmann, D., and Hall, A.
(1992) Cell 70, 401–41012. Ruhrberg, C., and Watt, F. M. (1997) Curr. Opin. Genet. & Dev. 7, 392–39713. Yang, Y., Bauer, C., Strasser, G., Wollman, R., Julien, J. P., and Fuchs, E.
(1999) Cell 98, 229–23814. Green, K. J., Virata, M. L., Elgart, G. W., Stanley, J. R., and Parry, D. A. (1992)
Int. J. Biol. Macromol. 14, 145–15315. Foisner, R., Malecz, N., Dressel, N., Stadler, C., and Wiche, G. (1996) Mol. Biol.
Cell 7, 273–28816. Stappenbeck, T. S., Lamb, J. A., Corcoran, C. M., and Green, K. J. (1994) J.
Biol. Chem. 269, 29351–2935417. Pascual, J., Castresana, J., and Saraste, M. (1997) Bioessays 19, 811–81718. Hartwig, J. H. (1994) Protein Profile 1, 706–77819. Grum, V. L., Li, D., MacDonald, R. I., and Mondragon, A. (1999) Cell 98,
523–53520. Svoboda, K., Schmidt, C. F., Branton, D., and Block, S. M. (1992) Biophys. J.
63, 784–79321. Bennett, V., and Gilligan, D. M. (1993) Annu. Rev. Cell Biol. 9, 27–6622. Dahl, S. C., Geib, R. W., Fox, M. T., Edidin, M., and Branton, D. (1994) J. Cell
Biol. 125, 1057–106523. Lo, S. H., An, Q., Bao, S., Wong, W. K., Liu, Y., Janmey, P. A., Hartwig, J. H.,
and Chen, L. B. (1994) J. Biol. Chem. 269, 22310–2231924. Lo, S. H., Janmey, P. A., Hartwig, J. H., and Chen, L. B. (1994) J. Cell. Biol.
125, 1067–107525. Smith, D. B., and Johnson, K. S. (1988) Gene (Amst.) 67, 31–4026. Davis, S., Lu, M. L., Lo, S. H., Lin, S., Butler, J. A., Druker, B. J., Roberts,
T. M., An, Q., and Chen, L. B. (1991) Science 252, 712–71527. Zucman-Rossi, J., Legoix, P., and Thomas, G. (1996) Genomics 38, 247–25428. Koenig, M., and Kunkel, L. M. (1990) J. Biol. Chem. 265, 4560–456629. Kawasaki, H., and Kretsinger, R. H. (1995) Protein Profile 2, 297–49030. Gregory, S. L., and Brown, N. H. (1998) J. Cell Biol. 143, 1271–128231. Prokop, A., Uhler, J., Roote, J., and Bate, M. (1998) J. Cell Biol. 143,
1283–129432. Strumpf, D., and Volk, T. (1998) J. Cell Biol. 143, 1259–127033. Byers, T. J., Beggs, A. H., McNally, E. M., and Kunkel, L. M. (1995) FEBS Lett.
368, 500–50434. Nagase, T., Ishikawa, K., Suyama, M., Kikuno, R., Miyajima, N., Tanaka, A.,
Kotani, H., Nomura, N., and Ohara, O. (1998) DNA Res. 5, 277–28635. Cooper, J. A. (1987) J. Cell Biol. 105, 1473–147836. Presley, J. F., Cole, N. B., Schroer, T. A., Hirschberg, K., Zaal, K. J., and
Lippincott-Schwartz, J. (1997) Nature 389, 81–8537. Dubreuil, R. R. (1991) BioEssays 13, 219–22638. Fuchs, E., and Cleveland, D. W. (1998) Science 279, 514–51939. Allen, P. G., and Shah, J. V. (1999) Bioessays 21, 451–45440. Foisner, R., and Wiche, G. (1991) Curr. Opin. Cell Biol. 3, 75–8141. Schneider, C., King, R. M., and Philipson, L. (1988) Cell 54, 787–79342. Brancolini, C., Benedetti, M., and Schneider, C. (1995) EMBO J. 14,
5179–519043. Speicher, D. W., and Marchesi, V. T. (1984) Nature 311, 177–18044. Tang, H. Y., Chaffotte, A. F., and Thacher, S. M. (1996) J. Biol. Chem. 271,
9716–972245. Prout, M., Damania, Z., Soong, J., Fristrom, D., and Fristrom, J. W. (1997)
Genetics 146, 275–28546. Bernier, G., Mathieu, M., De Repentigny, Y., Vidal, S. M., and Kothary, R.
(1996) Genomics 38, 19–2947. Manno, S., Takakuwa, Y., Nagao, K., and Mohandas, N. (1995) J. Biol. Chem.
270, 5659–5665
Trabeculin-a, New Actin Cross-linking Protein33530
by guest on April 4, 2019
http://ww
w.jbc.org/
Dow
nloaded from

Bo ChenYuan Liu, Rebecca Sutherland, Ravi Salgia, James D. Griffin, Louis H. Ferland and Lan
Yaping Sun, Jinyang Zhang, Stine-Kathrein Kraeft, Daniel Auclair, Mau-Sun Chang,Defining a New Family of Actin-binding Proteins
, a Giant ProteinαMolecular Cloning and Characterization of Human Trabeculin-
doi: 10.1074/jbc.274.47.335221999, 274:33522-33530.J. Biol. Chem.
http://www.jbc.org/content/274/47/33522Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/274/47/33522.full.html#ref-list-1
This article cites 47 references, 16 of which can be accessed free at
by guest on April 4, 2019
http://ww
w.jbc.org/
Dow
nloaded from





























