Embed Size (px)
Citation preview

GroEL array systemPaul Voziyan Bryan TiemanMary JohnstonGreg BomhoffAngela Chao
Cryo-EM Chaperonin-GS structuresScott Falke, graduate student (KUMC)
Florence Tama Charles Brooks Scripps
Edward Gogol collaborator - UMKC
KUMC Lied Research Fund
Designing a Chaperonin/Osmolyte Folding Array System

The need for a broad folding array system
Expressed proteins membrane proteins,truncated proteins, mutant proteins, affinity tagged proteins
Soluble and properly folded
Proper products or Crystallized proteins (low hanging fruit)
purification
large, smallAggregates orinclusion bodiesare usually shelved
Soluble butmisfolded,kineticallytrapped

+
ribosome
Active protein
misfoldingpathway IX
aggregation (mass action)
aggregate growth
aggregate growth
Cellular Folding
unfolding
folding
MOLECULARCHAPERONES
ATP
ADP
Chaperonins

142 Ao
155 Ao
33 Ao
GroEL (14 subunits)no nucleotide
GroES (7 subunits)
140Ao
140Ao
14 ATP binding sites
Chaperonins from E. coli (homologous to Hsp60 in mitochondria)
GroEL-GroES-ADP

Diagram courtesy ofHelen Saibil, Birbeck College

The chaperonin cycle
Step 3
ATP ATP
180o
ATP ATP ADP ADP
ATP ATP
ADP
ADP ADP
ATPase
ATP +
ATP ATP
ATP
Step 1 Step 2
Step 4
folded

GroEL
GS monomer51 kDa
Glutamine synthetasedodecamer (Class II(?))

Domains fit inside 10 A EM density map
GroEL no nucleotides
highest affinityfor folding polypeptides
polypeptidebinding sites
Ranson et al., 2001 Cell, 107, 869-879.
o

GroEL + ATP
low affinityfor folding polypeptides
polypeptidebinding sitesbecome lesssolvent exposed in ATP boundform.
Domains fit inside 10 A Cryo-EM density map
Ranson et al., 2001 Cell, 107, 869-879.
o

flipped180o
GroEL cryo images 13 A collected by S. Falke
o
Data collected at Scripps in collaboration with R. Milligan

flipped180o
substrate GS
GroEL cryo images 13 A collected by S. Falke
o
Data collected at Scripps in collaboration with R. Milligan

cis
Flexible fitting of crystal structure to EM mapNormal mode analysis fitting – F. Tama, C. Brooks

cis
Trans ring
3 mer fit fromNormal modeanalysisview from inside cavity
Substrate free

cis (Protein bound ring)
Trans ring
3 mer fit fromNormal modeanalysisview from inside cavity
GS bound

spontaneous foldingof Glutamine synthetase
Stable GroEL-GS complex
addition of GroESand ATP
50
% o
rigi
nal
act
ivit
y
Time (min)30 60 90 120 150 180
100
75
25
Typical protein folding experiment with chaperonins Fisher, 1992, Biochemistry

Up-down dimer contacts of Glutamine synthetase consist of swapped secondary elements.
Valine 468(c-terminal amino acid)
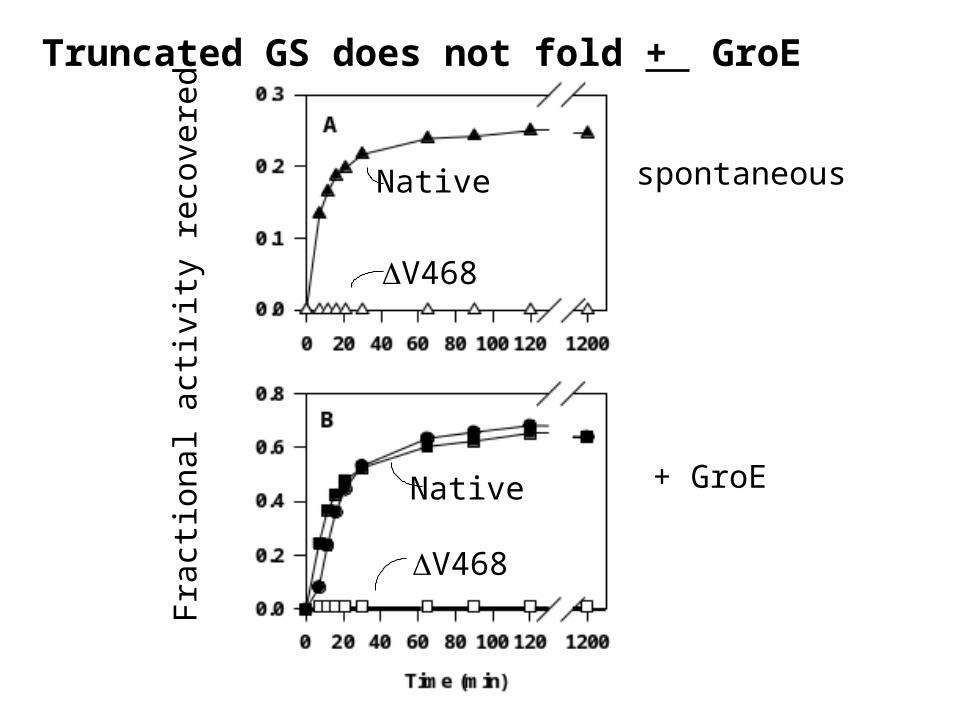
Truncated GS does not fold + GroE
spontaneousNative
Native
V468
V468
+ GroE
Fra
ctio
nal a
ctiv
ity
reco
vere
d

GroEL/ES-GS 468 complex
0 1 2 5 10 30 120 min
12-mer of native wt GS
misfolded aggregate
NB
Reassembled wt GS 12-mer
A0 1 2 5 10 30 120 min NGroEL/ES - wt GS complex
12-mer ofnative GS 468
Truncated GS does not assemble from GroE

Combination of other chaperone systemsand lower temperatures did not facilitate GS 468 assembly and reactivation
•DnaK (hsp 70), DnaJ (hsp40), GrpE
•small hsps
•GroE chaperonins + all of the above
•All of the above at 20C.
•All of the above at 15C.
NO ACTIVITY OBSERVED


Additive Activity recovered after 20 h (fraction of native) Additive alone with GroEL-ATP with GroEL-GroES-ATP
1 M betaine << 0.130.01 0.130.01
1 M sarcosine << 0.040.01 0.200.06
1 M sucrose 0.050.02 0.360.07 0.300.07
0.5 M KGlu << 0.090.01 0.350.06
1 M TMAO << 0.220.05 0.450.09
4M glycerol 0.180.04 0.480.08 0.470.09
<<, activity was below the detection limit of the assay.
Numerous osmolytes can facilitate folding of 468 GSfrom GroE (Voziyan et al., 2000, J. Pharm. Sci. Voziyan and Fisher, 2000, Protein. Sci.

Additive Activity recovered after 20 h (fraction of native) Additive alone with GroEL-ATP with GroEL-GroES-ATP
1 M betaine << 0.130.01 0.130.01
1 M sarcosine << 0.040.01 0.200.06
1 M sucrose 0.050.02 0.360.07 0.300.07
0.5 M KGlu << 0.090.01 0.350.06
1 M TMAO << 0.220.05 0.450.09
4M glycerol 0.180.04 0.480.08 0.470.09
<<, activity was below the detection limit of the assay.
Numerous osmolytes can facilitate folding of 468 GSfrom GroE (Voziyan et al., 2000, J. Pharm. Sci. Voziyan and Fisher, 2000, Protein. Sci.

What is the highest protein concentration we can fold with the chaperonin system?

Concentrate complex
+GroE
•With this method, V468 GS and GS can be refolded with >75% activity at 2 mg/ml (no limit found yet)
• Rhodanese - 5 mg/ml (no limit found as of yet).
Protein concentration
Act
ivit
y re
gain
GroEL-protein complexes can be concentrated(Fisher, J. Biol. Chem., 1993, Smith and Fisher, J. Biol. Chem. 1995
Voziyan et al., J. Pharm. Sci. 2000)
Activity regain no longerconcentration dependent
Protein concentration
Optimal GroEL-substrate formation

Assembly and reactivation of dodecameric GSoptimal vs. concentrated (37oC)
active dodecamer
0.25 mg/ml
0.025 mg/ml
20
40
60
80
10 20 30
Time (minutes)
Rea
ctiv
atio
n (
% o
f or
igin
al)

Another Advantage of this system - test for conditions that remove the GroES and ATP requirement from refolding assay. Typical chaperonin substrates...
•Mitochondrial malate dehydrogenase - folds with ATP/and all osmolytes tested at 20-37 C, ADP alone works in place of ATP
•Rhodanese - folds with ATP/glycerol even at 37 C
•Citrate synthase- ATP and polyols (no GroES needed), ADP alone works in place of ATP
•Alcohol dehydrogenase - ATP and most osmolytes

-0.1
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1
0 10 20 30 40 50 60
Porcine Mitochondrial Malate Dehydrogenase Chaperonin Requirements
Spontaneous
GroEL Alone
GroEL and ATP
GroEL, GroES, and ATP
PmMDH
Time (minutes)
37°C

PmMDH Renaturation with GroE Chaperonins in 35% Glycerol
Time (minutes)
0 20 40 60 80 100 120
Fra
ctio
n of
Rec
over
ed P
mM
DH
Act
ivity
0.0
0.2
0.4
0.6
0.8
1.0
Spontaneous
GroEL Alone
GroEL and ATP
GroEL, GroES, and ATP
In the Presence of 35% Glycerol, the Chaperonin Requirements for folding of PmMDH become less stringent
Tieman et al., 2001, J. Biol. Chem.
35% glycerol alone

Glutamine synthetase can fold from the chaperonin with glycerol (or sucrose) alone. No longer requires ATPVoziyan and Fisher, 2002, Arch. Biochem. Biophys.

Fra
ctio
n R
ecov
ered
Act
ivit
y
Time (hr)
0 1 2 3
0.8
0.6
0.4
0.2
Reused beads
+ATP
Renaturation of Glutamine synthetase from immobilized EL-Beads(attached through succinimide linkage)
Glycerol 2M (no ATP)
Beads alone (no EL attached)

MDH +Beads alone
MDH+EL beads
MDH +EL-beadsGlycerolATP
MDH +EL-beadsGlycerolATP2nd trial-same beads
Fra
cti
on
al acti
vit
y
reg
ain
aft
er
two
hou
rs
0.15
0.30
0.45
Minimal requirements with osmolytes preserved using the immobilized system –with a more stringentGroE substrate - Malate dehydrogenase.

Folding from Inclusion bodies
isolate inclusion body - sucrose cushion
wash and unfold (GnHCl)
refold with chaperonin-osmolyte array
test for correct folding
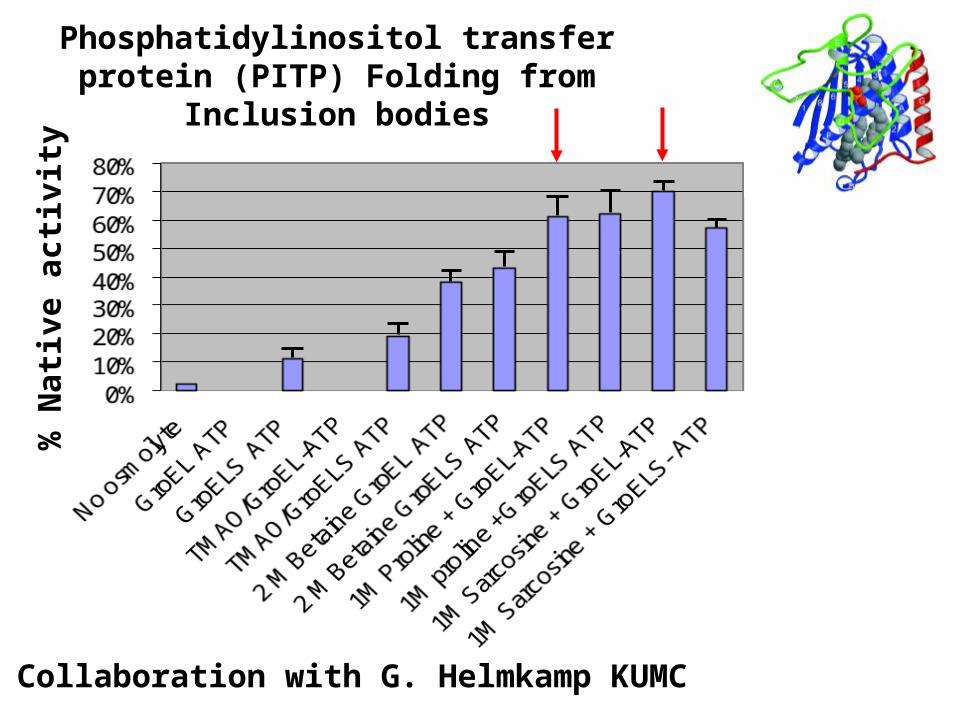
Phosphatidylinositol transfer protein (PITP) Folding from Inclusion bodies
% N
ativ
e ac
tivi
ty
Collaboration with G. Helmkamp KUMC

Folding from inclusion bodies –Protein X in vitro (Arginine best osmolyte)
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 218M urea +Buffer+BSA +Buffer +GroEL + + + + + + + + + + + + + +GroES + + + + + + +ARG + + + + + + + + + +GS/GSSG + + + + + + + +PDI + + + + + + + + + +
28.4 kDa
21.7 kDa
Standard,10 ng
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21
68.8 kDa
52.5 kDa
Western Blot of soluble fractions with GP-antibody detection
Additives

IX
inactive protein
GroELGroEL-substratefolding platform
concentrate
multiple wellfolding array
additive 1
additive 2
additive 3
additive i
ATP(ADP)
0%
10%
90%
0%
BEST FOLDING
CONDITIONS
Dilute into wells
Free or immobilizedGroEL
(patent pending)

More applications:
GroE-osmolyte system– testing for ligands or osmolytes to correct a structural defect using an array!
Known –some Osmolyte systems can reverse assembly or folding defects

•If the protein binds a ligand (L)
release I
I N N+L N-L
excess ligand shifts equilibrium to foldedform.
GroEL array system may allow us to screen for small molecules to prevent aggregation.
Advantage of GroEL – all intermediates are initially bound.
Aggregate
GroEL-I

Failures can be reversed with the correct ligand
collaboration with R. Middaugh at KU Lawrence
Acid Fibroblast growth factoraggregates easily
CHAPERONINS enhancedaggregation!!! `
•The ligand Heparin shifts the foldingequilibrium to the folded soluble state.

Folding mutant Protein
Disease Osmolyte or ligand
Reference
Cystic Fibrosis transmembrane conductance regulator protein (chloride transport)
Cystic Fibrosis Glycerol, CPX ? ?(SciClone)
Brown et al., 1996 Cell stress Chaperones, 1: 117-125
aquaporin Nephrogenic Diabetes Insipidus
Glycerol Tamarappoo et al., 1999, JBC 274: 34825
Alpha 1 trypsin inhibitor Emphysema, liverinjury
Glycerol Burrows et al., (2000) PNAS 97: 1796
Beta glucosidase Gaucher Disease N-n-nonyl)deoxynojirimycin – NN-DNJ
Sawkar et al., 2002, PNAS, 99: 15428
Transthyretin Familial AmyloidCardiomyophathy
L-thyroxine, Flurbiprofien, resveratrol etc.
Klabunde et al., 2000, Nat. Struc. Biol. 7:312.
V2 vasopression receptor
Nephrogenic Diabetes Insipidus
Nonpeptidic V2R antagonists
Morello et al., 2000, J. Clin. Invest. 105: 887

Jiu-Li Song and David Chuang ( 2001, J. Biol. Chem., 276, 40241-46 )
Branched chain - ketoacid decarboxylase normally a tetramer, - mutant (Maple syrup urine disease) an inactive soluble dimer.
With complete GroE system –in combination with TMAO corrects defect. Once identified, TMAO ALONE corrects assembly defects in branched chain - ketoacid decarboxylase (not as well as GroE-TMAO combination)
FOLDED ACTIVE Tetramer STATE REMAINED ACTIVE WHEN THE OSMOLYTE IS REMOVED!!
Using the GroEL-ES-osmolyte system

•Potential to rapidly identify the best folding solution
•When used as a folding platform, the chaperonins capture folding intermediates and can maintain thesemetastable states in aggregation free states.
•Substrates released from GroEL at very highconcentrations.
•Requirement for GroES (and ATP) can be eliminated.
•Works from an immobilized support and is reusable.
•May be used to search for therapeutic ligands to preventaggregation
Implications of this research:



2.5 A atomic model of E coli Glutamine synthetase dodecamer
Top view(one hexamer) Side view
Almassy et al., 1987
Mg ions

Time (min)
Truncated GS folds in the presence of Glycerol(20-35%) and GroE
V468 GS
Native GS
+ ATP
Fra
ctio
nal
Act
ivit
y re
cove
red






























