Embed Size (px)
Citation preview

6 • Great ape cognitive systemsANNE E. RUSSONPsychology Department, Glendon College of York University, Toronto
INTRODUCTION
This chapter considers cognition in great apes as inte-grated systems that orchestrate the many abilities thatgreat apes express, systems for which satisfactory char-acterizations remain elusive. In part, difficulties owe toresearch trends. Empirical studies have been guided bydiverse and sometimes contradictory models, questions,measures, tasks, and living conditions. Performancelevels have proven inconsistent across individuals, rear-ing conditions, and testing conditions, and evidence ispatchy across species for virtually any facet of cogni-tion. Evidence on wild great apes, the most importantfrom an evolutionary perspective, is especially patchybecause research has favored captives; much of whatis available was collected for other purposes, so it wasneither described nor analyzed with cognition in mind.The issues at stake are also hard-felt ones that touch onthe human–nonhuman boundary, so entrenched beliefsinfect how the literature is interpreted and even what ofit is read.
Attempts have none the less been made to developan integrated model of great ape cognition using avail-able evidence. They include both edited survey vol-umes (Matsuzawa 2001a; Parker, Mitchell & Miles 1999;Russon, Bard & Parker 1996) and integrative reviews,three of the latter as major books (Byrne 1995 (RWB),Parker & McKinney 1999 (P&M); Tomasello & Call1997 (T&C)) and others as articles (e.g., Byrne 1997;Suddendorf & Whiten 2001; Thompson & Oden 2000;Whiten & Byrne 1991). My aim is not to analyze thisterrain, yet again, in detail, but to offer a compact misea date to ground evolutionary reconstruction. Guidingquestions are “what, if anything, about great ape cog-nition requires evolutionary explanations beyond thosedeveloped for other nonhuman primates?”, and “how isgreat ape cognition best characterized with respect toevolutionary questions?”
CONCEPTS AND MODELS OFCOGNITION
Situating great ape cognition comparatively hinges onmental processes that support symbolism, notably rep-resentation, metarepresentation, and hierarchization.Weaker and stronger conceptualizations exist for eachand which is used affects assessments of great apes’ capa-bilities.
Weak meanings of symbolism include reference byarbitrary convention (Peirce 1932/1960), using inter-nal signs like mental images to stand for referentsrather than using direct sensations or motor actions,and solving problems mentally versus experientially. Inthe strong sense, symbolism refers to self-referring sys-tems wherein phenomena owe their significance andeven existence to other symbols in the system ratherthan to sensorimotor entities (e.g., Deacon 1997; Donald2000; Langer 2000). Representation can refer to anyform of mental coding that stands for entities, perceptualincluded (Perner 1991; Whiten 2000) or, more strongly,to recalling to mind or “re-presenting” mental codesfor entities and simple object relations in the absenceof normal sensorimotor cues (P&M; Whiten 2000).Meanings of metarepresentation range from represent-ing other representations (e.g., Leslie 1987; Matsuzawa1991; Whiten & Byrne 1991) to representing a represen-tation as a representation, i.e., an interpretation of a sit-uation (Perner 1991). Meanings of hierarchization spancreating new, higher-order cognitive structures fromlower-level ones (i.e., structures with superordinate–subordinate features: Byrne & Byrne 1991; Case 1985;Langer 1998) to generating cognitive structures thatshow embedding (e.g., classification showing nesting ofclasses: Langer 1998).
Developmentalists commonly consider weak andstrong forms to be related in humans, as basic andadvanced ontogenetic achievements of early and later
Copyright Anne Russon and David Begun 2004.
76

Great ape cognitive systems 77
childhood respectively (Table 6.1). Comparative pri-mate cognition often shares this view (P&M; Whiten2000). Insofar as symbols must be grounded in realworld referents at some point (Donald 1991) and weaksymbolism is the more likely in great apes, I considergreat ape cognition relative to weak symbolism and itsassociated processes (strong representation, weak hier-archization, weak metarepresentation). The terms sym-bolic, representation, hierarchization, and metarepre-sentation henceforth refer to these meanings.
The models guiding empirical studies of great apecognition also contribute to disparities because of theways in which they shape the generation of evidenceand the interpretive frameworks they impose. Severalimportant models are sketched below to suggest theirstrengths and limitations for understanding great apecognition.
Animal models designed for nonhuman mentalityhave been frequent frameworks for studies of great apecognition. They concentrate on the non-symbolic, asso-ciative processes presumed to govern nonhuman cog-nition, for example trial-and-error experiential learn-ing or behavior chains. This leaves them conceptuallyand methodologically impoverished concerning sym-bolic cognition (Anderson 1996; RWB; Rumbaugh 1970;T&C), quantification and logic being important excep-tions (e.g., Boysen & Hallberg 2000; Thompson & Oden2000), so relatively little of the evidence they have gener-ated helps determine whether great apes, or any species,are capable of symbolic cognition.
Generality–modularity models are potentiallyimportant because they concern cognitive architecture.In this view, favored by evolutionary psychologists andneo-nativists, there exist cognitive “modules,” problem-specific cognitive structures that represent innatelyspecified neurological systems and operate with rela-tive autonomy, as well as general purpose or centralprocesses that apply across problem types and affectsystem-wide properties (e.g., representation, executivecontrol structures, working memory). These modelshave influenced understandings of great ape cognitionwith their assumption that modular architecture char-acterizes nonhuman cognition and generality evolveduniquely in humans (e.g., Mithen 1996; T&C; Tooby& Cosmides 1992). Little if any empirical study hasexamined cognitive architecture in great apes, however.Studies of great ape cognition have typically assumedmodularity and have aimed for clean tests of individual
problem-specific structures – effectively eliminatingchances for detecting use of multiple or general purposeprocesses. Given the lack of relevant empirical evidence,these models remain speculative concerning great apes.
Cognitive science models portray the mind as adevice for processing, storing, integrating, and trans-forming information. Some of their concepts have beenincorporated into models of cognitive development (e.g.,Case 1985; Leslie 1987; Pascual-Leone 1987), othershave aided in detecting hierarchization in great ape cog-nition (Byrne & Byrne 1991; Byrne, Corp & Byrne 2001;Byrne & Russon 1998; P&M; Russon 1998). Limita-tions concern portraying cognition in static, mechanisticterms that may not apply to living beings.
Models of human cognitive development haveproven valuable for assessing primate cognition com-paratively because they provide conceptual and method-ological tools for assessing non-symbolic and symboliccognition within one unified framework and the gen-eration and structure of cognition. Piaget’s model sup-ported the first developmental studies of great ape cog-nition; among its greatest contributions is its portrayal ofcognition as constructed progressively during ontogenyand directly affected by interaction with the environ-ment. Early piagetian studies focused on sensorimotor(human infant) cognition, which relies on pre-symbolicprocesses similar to those portrayed in animal models, sosimilar limits apply. Recently, neo-piagetian models havesupported studies of the rudimentary symbolic range(for an overview, see P&M). Models inspired by Vygot-sky, which portray socio-cultural forces like apprentice-ship or enculturation as fundamental to cognitive devel-opment (e.g., Donald 2000; T&C), have spawned manystudies on social cognition and cognitive developmentin great apes. Given how richly primate lives are sociallyembedded, their merits are obvious. Among these mod-els may be included models of understanding others’mental states, or theory of mind, which some propose tounderpin much cognitive progress in early human child-hood (e.g., Carruthers & Smith 1996). Two such modelshave been applied to great apes, both proposing thatgeneral-purpose cognitive processes in the rudimen-tary symbolic range underwrite this progress (second-order representation – Leslie 1987; secondary represen-tation – Perner 1991). While both offer useful tools forassessing rudimentary symbolic processes, their focuson one ability series in the social domain risks under-representing the breadth of great apes’ achievements.

Tab
le6.
1.P
roce
ssesa
ndstr
uctu
resp
osite
din
rudi
men
tary
sybo
licle
velc
ogni
tion,
inhu
man
deve
lopm
enta
lper
spec
tive
Age
Piag
etC
ase1
Lan
ger2
Les
lie3
Pern
er4
(yr)
(gen
eral
)(c
ausa
lity)
(logi
c-m
ath)
(The
ory
ofM
ind)
(The
ory
ofM
ind)
0Se
nsor
imot
orst
age
Sche
mat
aSe
nsor
imot
orst
age
1st-
orde
rco
gniti
on1s
t-or
der
repr
esen
tatio
nPr
imar
yre
pres
enta
tion
1.5
2
Ope
ratio
nalc
onso
lidat
ion
(Int
er-)
rela
tiona
lcog
nitio
nun
ifoca
lrel
atio
nalo
pera
tions
2nd-
orde
rco
gniti
on2n
d-or
der
repr
esen
tatio
nm
etar
epre
sent
atio
nSe
cond
ary
repr
esen
tatio
nre
-rep
rese
ntpr
imar
yre
pres
enta
tions
mul
tiple
repr
esen
tatio
nsst
age
6,sy
mbo
ls,
repr
esen
tati
on3.
5Pr
e-op
erat
iona
lsta
geSy
mbo
licsu
bper
iod
Bifo
calr
elat
iona
lope
ratio
ns(1
st-o
rder
sym
bolic
)5
Intu
itive
subp
erio
dE
labo
rate
dre
latio
nalo
pera
tions
(2nd
-ord
ersy
mbo
lic)
3rd-
orde
rco
gniti
onR
easo
nac
ross
met
arep
rese
ntat
iona
lst
ruct
ures
Met
arep
rese
ntat
ion
sym
bols
Not
atio
n:Sy
mbo
l,re
pres
enta
tion,
met
arep
rese
ntat
ion
defin
edin
text
;no
rmal
/bol
dty
pein
dica
tes
wea
k/st
rong
mea
ning
sre
spec
tivel
y;m
ajor
cogn
itive
peri
ods
are
unde
rlin
ed;s
igni
fican
tpro
cess
esor
stru
ctur
esw
ithin
peri
ods
are
inde
nted
.1
Cas
e(1
985,
1996
)m
odel
sca
usal
cogn
ition
atth
ele
vel
ofop
erat
ing
onob
ject
–obj
ect
rela
tions
,i.e
.,re
latio
nal
(pro
perl
y,in
ter-
rela
tiona
l)co
gniti
on.
Chi
ldre
nde
velo
pst
ruct
ures
first
fors
ingl
e,si
mpl
ere
latio
ns,t
hen
rela
tions
-bet
wee
n-re
latio
ns,t
hen
coor
dina
ting
incr
easi
ngnu
mbe
rsof
rela
tiona
lstr
uctu
res:
12–2
0m
o–
repr
esen
tone
rela
tions
hip
betw
een
two
item
s(o
pera
tiona
lcon
solid
atio
n);2
0–27
mo
–re
pres
ento
nein
ter-
rela
tiona
lstr
uctu
re(u
nifo
calo
pera
-tio
ns);
27–4
0m
o–
repr
esen
ttw
oin
ter-
rela
tiona
lstr
uctu
res(
bifo
calr
elat
iona
lope
ratio
ns,fi
rsto
rder
sym
bolic
);40
–60
mo
–in
ter-
rela
tem
ore
inte
r-re
latio
nal
stru
ctur
es(e
labo
rate
dre
latio
nalo
pera
tions
,sec
ond
orde
rsy
mbo
lic).
2L
ange
r(1
998,
2000
)m
odel
slo
gica
lope
ratio
nson
subj
ects
’spo
ntan
eous
obje
ctgr
oupi
ngs.
0–12
mo
–m
ake
one
set
ofob
ject
sw
ithon
ecl
ass
prop
erty
,m
ap1s
t-or
der
oper
atio
nson
toit
(1st
-ord
erco
gniti
on);
18–3
6m
o–
mak
etw
oco
ntem
pora
neou
sse
ts,m
ap2n
d-or
der
oper
atio
nson
toth
ese
ts(2
nd-o
rder
cogn
ition
);>
36m
o–
mak
eth
ree
cont
empo
rane
ous
sets
,map
3rd-
orde
rop
erat
ions
onto
them
(e.g
.,co
nstr
uctc
orre
spon
denc
es)(
3rd-
orde
rco
gniti
on).
3L
eslie
(198
7).F
irst
-ord
er(p
rim
ary)
repr
esen
tatio
nsen
code
entit
iesi
nan
accu
rate
,lite
ralw
ay;t
hey
are
perc
eptu
ally
base
dan
dde
fined
inse
nsor
imot
orco
des
bydi
rect
sem
antic
rela
tion
with
the
wor
ld;m
ultip
lepr
imar
yre
pres
enta
tions
ofa
situ
atio
nca
nex
ist.
Seco
nd-o
rder
repr
esen
tatio
ncr
eate
sa
deco
uple
dco
pyof
apr
imar
yre
pres
enta
tion
then
reco
nstr
ucts
orre
desc
ribe
sit;
mak
ing
ade
coup
led
copy
enta
ilsm
etar
epre
sent
atio
n;se
cond
orde
rre
fers
tobe
ing
deri
ved
from
apr
imar
yre
pres
enta
tion;
seco
nd-o
rder
repr
esen
tatio
nsty
pica
llyre
mai
nan
chor
edto
part
sof
the
prim
ary
repr
esen
tatio
n.4
Pern
er(1
991)
.0–1
yr–
repr
esen
tatio
nspo
rtra
yth
ecu
rren
tsitu
atio
nre
alis
tical
ly(p
rim
ary
repr
esen
tatio
ns);
only
one
prim
ary
repr
esen
tatio
nof
the
situ
atio
nex
ists
;1–4
yr–
othe
rre
pres
enta
tion(
s)of
the
situ
atio
n(p
ast,
futu
re)
(sec
onda
ryre
pres
enta
tions
)ar
een
tert
aine
dsi
mul
tane
ousl
yw
ithth
epr
imar
yre
pre-
sent
atio
n;>
4yr
–re
pres
enta
tions
ofot
herr
epre
sent
atio
nsar
ecre
ated
and
unde
rsto
odas
repr
esen
tatio
ns(i.
e.,a
sint
erpr
etat
ions
)(st
rong
met
arep
rese
ntat
ion)
.

Great ape cognitive systems 79
More broadly, reservations are that Vygotsky-basedmodels tend to emphasize socio-cultural factors to theneglect of individual and biological ones, while Piaget-based models suffer the opposite bias. Together, thesemodels offer rich portrayals of cognitive developmentand have spawned comparative models situating primatecognition in developmental and evolutionary perspec-tive (e.g., P&M).
I favor development frameworks because they allowassessment of symbolic processes, their constitution,and continuities as well as discontinuities betweenhuman and nonhuman primates. I adopt them here asthe basis for interpreting evidence.
EVIDENCE
For evidence, I relied on recent integrative reviews(RWB, P&M, T&C) more than edited volumes, to priv-ilege syntheses over the breadth of current views, plusfindings appearing since their publication (post 1998).I concentrated on achievements linked with symbolismas the critical cognitive threshold and feral great apes1 asmost relevant to evolutionary questions. Table 6.2 sum-marizes this evidence, arranged by the cognitive struc-tures inferred in terms of cognitive domain (broad areasof knowledge, typically physical, logico-mathematical,social, linguistic), problem-specific structures (abilityseries), and complexity (level). This arrangementderives from models of human cognitive developmentnear the rudimentary symbolic range (Table 6.1).
My coverage of the evidence is inevitably incom-plete but sufficient to establish broad patterns. Evidencefor complex achievements is substantial, for instance,and the relevance of complex skills to feral life is clear inall cognitive domains even though little evidence derivesfrom feral subjects. Equally clear and needing explana-tion are the repertoire’s impressive breadth and “open-ness” (i.e., including apparently “atypical” language andmathematical abilities). Disputes in any case lie less withwhat great apes achieve than with cognitive inferences,so more important cautions are that inferences are con-troversial, numerous factors complicate interpretation,and I inevitably glossed over subtleties and debates inworking towards an overall picture.
A long-standing concern is variability in achieve-ments across problem types, individuals, species,and contexts. Some report great apes outperforming5- to 6-year-old humans (e.g., Call & Rochat 1996),
others report them failing at simple levels of under-standing (e.g., Povinelli 2000). While this variabil-ity may be meaningful (e.g., cognitive differencesbetween species, significant features of cognitive devel-opment, module-like cognitive architecture), it alsoreflects confounding factors extensive enough to under-mine interpretation.2 Because quantitative breakdownsremain un-interpretable, I have not provided them.Most experts in any case consider that all great apes shareroughly equivalent cognitive capacities (RWB, P&M,T&C) and it is these similarities that are of primaryinterest here.
COGNITIVE LEVELS: THEHIGH-MINDED
An important consideration in analyzing the cognitiongoverning great apes’ complex achievements is that itmay involve higher levels of cognitive abstraction, notjust very rapid processing, extended working memory,or new types of abilities (Roberts & Mazmanian 1988).Humans, great apes, and some monkeys can master mak-ing and using tools, for instance, so all share the means–end type of cognition; great apes and humans differ inachieving higher levels of means–end cognition that sup-port more complex tool use (e.g., Visalberghi & Limon-gelli 1996). What levels great apes attain is a major focusof current debate. Three levels recognized in humandevelopment beyond pre-symbolic, sensorimotor cogni-tion (with its schemata, i.e., first-order or primary rep-resentations) are probably important to resolving thisdebate (see Table 6.2). These are:
(1) Emergence of rudimentary symbols.Around 1.5 years of age, humans begin creating andusing simple symbols, like mental images, to stand forreferents instead of having to use direct sensorimotorinformation. A classic example is inferring where anitem is hidden after watching it be displaced “invisibly,”along a trajectory that passes behind barriers; thisshows that the actor can mentally reconstruct eventsit did not directly perceive (de Blois, Novak & Bond1998). Early symbols have been attributed to strongrepresentation (Piaget 1952, 1954; P&M), understand-ing relational categories between entities external tothe actor (Herrnstein 1990; Rumbaugh & Pate 1984;Spearman 1927; Thompson & Oden 2000; T&C), orrepresenting single object–object relations (Case 1985).This level is usefully viewed as a transition, i.e., a phase

Tab
le6.
2.G
reat
apes
’cog
nitiv
eac
hiev
emen
tsan
dco
gniti
veab
ilitie
s
Phys
ical
Dom
ain
Cog
nitio
nSe
ries
/lev
elA
chie
vem
ent
Com
men
ts–
exam
ples
Sour
cesa
Obj
ectc
once
pt:d
evel
opin
gth
eco
ncep
tof“
obje
ct”
inth
een
viro
nmen
t(ex
tend
sonl
yto
sens
orim
otor
stage
6T
rans
ition
al(1
.5–2
yrs)
Tra
ckin
visi
ble
disp
lace
men
ts1
(2)3
416
Cau
sali
ty:d
ynam
icre
latio
nsbe
twee
nob
ject
swhe
nex
tern
alfo
rces
affe
ctth
emT
rans
ition
alIn
cons
iste
ntbu
tins
ight
ful
(Inc
onsi
sten
tsuc
cess
)1
23
12(1
.5–2
yrs)
mak
e&
use
rake
tool
Sing
leob
ject
–obj
ectr
elat
ions
Ter
tiary
rela
tions
betw
een
obje
cts
12
3R
udim
enta
ryC
onsi
sten
trak
ing
Rak
ew
ithco
nsis
tent
succ
ess
1(2
)8sy
mbo
licA
dvan
ceto
olpr
epar
atio
nE
mer
ges
inch
ildre
n>
2yr
sol
d(T
&C
)1
23
1112
(2–3
.5yr
s)H
iera
rchi
calt
echn
ique
sM
anua
land
tool
(set
s,se
ries
,met
a-)
1(2
)35
78
1017
1820
Com
posi
teto
ols
i.e.,
mul
ti-to
olas
sem
blag
es1
(2)3
615
17In
ter-
rela
tiona
lobj
ectu
sei.e
.,re
latio
ns-b
etw
een-
rela
tions
1(2
)39
1720
Coo
pera
tive
hunt
ing
Arb
orea
lity–
prey
–hun
ter
rela
tions
13
12
Spa
ce:s
patia
lund
ersta
ndin
g(k
now
ledg
e,re
latio
ns)
and
reas
onin
gT
rans
ition
alD
etou
rre
barr
ier,
chec
kfo
odin
adva
nce
(1)2
3N
avig
ate
2-di
men
sion
alm
aze
221
Arb
orea
l“cl
ambe
r”tr
avel
3St
ack
bloc
ksPu
tobj
ects
inco
ntai
ners
,sta
ck1
Rud
imen
tary
Blo
ckas
sem
bly
Tw
oor
mor
ebl
ocks
,var
ious
lyre
late
d1
sym
bolic
Dra
wci
rcle
orcr
oss
1T
iesi
mpl
ekn
otW
indi
ngan
din
sert
ing
113
“Map
”re
adU
sesc
ale
mod
els,
TV
,pho
tos
1(2
)14
19E
uclid
ean
men
talm
aps
Min
imiz
esi
te–s
itetr
avel
dist
ance
1(2
)312
22b
Plan
trav
elro
utes
Lea
stdi
stan
ce,a
rbor
ealr
oute
s1
(2)3
Not
es:
aC
ogni
tive
attr
ibut
ions
base
don
crite
ria
disc
usse
din
text
;sou
rce
brac
kete
dw
hen
my
attr
ibut
ion
diff
ers
from
that
ofth
eau
thor
sci
ted.
bSy
mbo
l-tr
aine
dgr
eata
pes
test
ed.
Sour
ces:
1,Pa
rker
&M
cKin
ney
1999
;2,T
omas
ello
&C
all1
997;
3,B
yrne
1995
;4,d
eB
lois
,Nov
ak&
Bon
d19
98;5
,Cor
p&
Byr
ne20
02;6
,Sug
iyam
a19
97;7
,R
usso
n19
98;8
,Byr
ne&
Rus
son
1998
,9,R
usso
n&
Gal
dika
s19
93;1
0,R
usso
n19
99a;
11,F
ox,S
itom
pul&
van
Scha
ik19
99;1
2,B
oesc
h&
Boe
sch-
Ach
erm
ann
2000
;13,
Map
le19
80;1
4,K
uhlm
eier
etal
.199
9;15
,Ber
mej
o&
Ille
ra19
99;1
6,C
all2
001a
;17,
Yam
akos
hi,C
hapt
er9,
this
volu
me;
18,S
toke
s&
Byr
ne20
01;1
9,K
uhlm
eier
&B
oyse
n20
01;2
0,M
atsu
zaw
a20
01b;
21,I
vers
on&
Mat
suza
wa
2001
;22,
Men
zel,
Sava
ge-R
umba
ugh
&M
enze
l200
2.
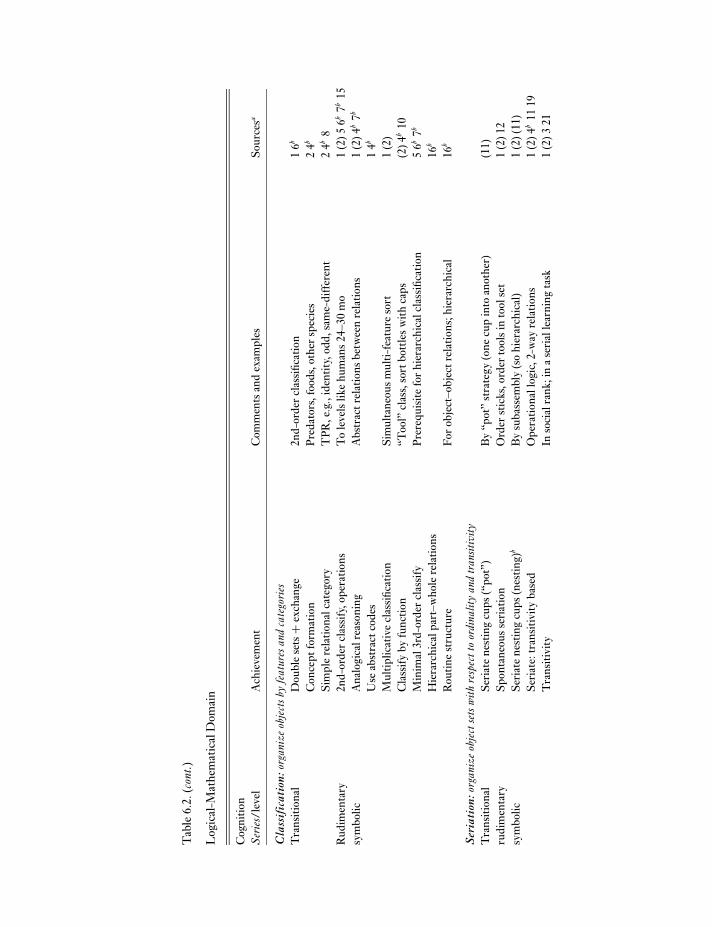
Tab
le6.
2.(c
ont.)
Log
ical
-Mat
hem
atic
alD
omai
n
Cog
nitio
nSe
ries
/lev
elA
chie
vem
ent
Com
men
tsan
dex
ampl
esSo
urce
sa
Cla
ssif
icat
ion:
orga
nize
obje
ctsb
yfe
atur
esan
dca
tego
ries
Tra
nsiti
onal
Dou
ble
sets
+ex
chan
geC
once
ptfo
rmat
ion
Sim
ple
rela
tiona
lcat
egor
y
2nd-
orde
rcl
assi
ficat
ion
Pred
ator
s,fo
ods,
othe
rsp
ecie
sT
PR,e
.g.,
iden
tity,
odd,
sam
e-di
ffer
ent
16b
24b
24b
8R
udim
enta
rysy
mbo
lic2n
d-or
der
clas
sify
,ope
ratio
nsA
nalo
gica
lrea
soni
ngU
seab
stra
ctco
des
Mul
tiplic
ativ
ecl
assi
ficat
ion
To
leve
lslik
ehu
man
s24
–30
mo
Abs
trac
trel
atio
nsbe
twee
nre
latio
ns
Sim
ulta
neou
sm
ulti-
feat
ure
sort
1(2
)56b
7b15
1(2
)4b
7b
14b
1(2
)C
lass
ifyby
func
tion
Min
imal
3rd-
orde
rcl
assi
fyH
iera
rchi
calp
art–
who
lere
latio
nsR
outin
est
ruct
ure
“Too
l”cl
ass,
sort
bott
les
with
caps
Prer
equi
site
for
hier
arch
ical
clas
sific
atio
n
For
obje
ct–o
bjec
trel
atio
ns;h
iera
rchi
cal
(2)4
b10
56b
7b
16b
16b
Ser
iati
on:o
rgan
ize
obje
ctse
tsw
ithre
spec
tto
ordi
nalit
yan
dtr
ansit
ivity
Tra
nsiti
onal
Seri
ate
nest
ing
cups
(“po
t”)
By
“pot
”st
rate
gy(o
necu
pin
toan
othe
r)(1
1)ru
dim
enta
rysy
mbo
licSp
onta
neou
sse
riat
ion
Seri
ate
nest
ing
cups
(nes
ting)
b
Seri
ate:
tran
sitiv
ityba
sed
Tra
nsiti
vity
Ord
erst
icks
,ord
erto
ols
into
olse
tB
ysu
bass
embl
y(s
ohi
erar
chic
al)
Ope
ratio
nall
ogic
,2-w
ayre
latio
nsIn
soci
alra
nk;i
na
seri
alle
arni
ngta
sk
1(2
)12
1(2
)(11
)1
(2)4
b11
191
(2)3
21

Tab
le6.
2.(c
ont.)
Log
ical
-Mat
hem
atic
alD
omai
n
Cog
nitio
nSe
ries
/lev
elA
chie
vem
ent
Com
men
tsan
dex
ampl
esSo
urce
sa
Num
ber/
Qua
ntit
y:as
sess
obje
ctse
tsw
ithre
spec
tto
num
bero
rqua
ntity
Tra
nsiti
onal
Sequ
entia
llyta
gse
vera
lite
ms
Sequ
entia
llyta
g+
labe
lnum
ber
12
12
Rud
imen
tary
sym
bolic
bC
ount
Com
pare
prop
ortio
nsC
onse
rve
num
ber
(1:1
)Su
mm
atio
nQ
uant
ified
(soc
ial)
reci
proc
ityPl
anne
dnu
mer
ical
orde
ring
Rev
erse
cont
inge
ncy
task
Sym
bolic
quan
tity
judg
men
t
Exa
ctnu
mbe
rof
item
sin
arra
ysF
ract
ion,
quan
tity-
base
dan
alog
yU
nder
stan
d1:
1co
rres
pond
ence
Add
ing
item
sin
crea
ses
quan
tity
Mea
tsha
reru
les,
exch
ange
groo
m/f
avor
sSe
quen
ceal
lite
ms
befo
reac
ting
Cho
ose
smal
ler
of2
arra
ysto
getm
ore
Sele
ctar
ray
for
quan
tity
usin
gsy
mbo
ls
1(2
)14
1(2
)1
(2)
(2)9
1314
1(2
)39
2022
15 1718
17
Con
serv
atio
n:co
nser
vepr
oper
tieso
fobj
ects
that
unde
rgo
tran
sform
atio
nsR
udim
enta
rysy
mbo
licb
Con
serv
equ
antit
y(c
once
ptua
l)Ph
ysic
ally
tran
sfor
med
(sol
id&
liqui
d)1
(2)
Not
es:
aco
gniti
veat
trib
utio
nsba
sed
oncr
iteri
adi
scus
sed
inte
xt;s
ourc
ebr
acke
ted
whe
nm
yat
trib
utio
ndi
ffer
sfr
omth
atof
the
auth
ors
cite
d.b
sym
bol-
trai
ned
grea
tape
ste
sted
.So
urce
s:1,
Park
er&
McK
inne
y19
99;2
,Tom
asel
lo&
Cal
l199
7;3,
Byr
ne19
95;4
,Tho
mps
on&
Ode
n20
00;5
,Lan
ger
2000
;6,P
otıe
tal.
1999
;7,S
pino
zzie
tal.
1999
;8,T
anak
a20
01;9
,Sou
sa&
Mat
suza
wa
2001
;10,
Rus
son
1999
a;11
,Joh
nson
-Pyn
net
al.1
999;
John
son-
Pynn
&F
raga
szy
2001
;12,
Ber
mej
o&
Ille
ra19
99;
13,C
all2
000;
14,B
eran
2001
;15,
Bir
o&
Mat
suza
wa
1999
;16,
Spin
ozzi
&L
ange
r199
9;17
,Boy
sen
&B
ernt
son
1995
;Boy
sen
etal
.199
6;18
,Shu
mak
eret
al.2
001;
19,K
awai
&M
atsu
zaw
a20
00;2
0,B
oesc
h&
Boe
sch-
Ach
erm
ann
2000
;21,
Tom
onag
a&
Mat
suza
wa
2000
;22,
Mita
ni&
Wat
ts20
01;M
itani
,Wat
ts&
Mul
ler
2002
.(N
ote:
rela
ted
stud
ies
are
grou
ped.
)

Tab
le6.
2.(c
ont.)
Soci
aldo
mai
n
Cog
nitio
nSe
ries
/lev
elA
chie
vem
ent
Com
men
tsan
dex
ampl
esSo
urce
sa
Soc
iall
earn
ing
&im
itat
ion:
soci
ally
influ
ence
dle
arni
ng;
imita
tion
isle
arni
ngto
done
wac
tsby
seei
ngth
emdo
neT
rans
ition
alD
efer
red
imita
tion
Act
ion-
leve
lim
itatio
n•g
estu
res
•act
ions
onob
ject
sIm
itate
actio
nse
quen
ce
Del
ayed
imita
tion
ofno
vela
ctio
ns“I
mpe
rson
atio
n,”
toso
me
Spon
tane
ous
gest
ures
,ges
ture
sign
sSi
mpl
eto
olus
e,ob
ject
man
ipul
atio
n2-
actio
nse
quen
ceor
long
er
124
b
13
12
315
221
23
414
1824
b29
3437
23
624
b
Rud
imen
tary
Sym
bolic
Prog
ram
-lev
elim
itatio
nM
ime
inte
nt,r
eque
st,t
each
Rou
tine
stru
ctur
e,re
latio
ns-b
etw
een-
rela
tions
Act
out(
for
othe
r),e
xpre
ssin
tent
13
45
1423
24b
1(2
)37
33b
Pre
tens
e:re
-ena
ctac
tions
outsi
deth
eiru
sual
cont
exta
ndw
ithou
tthe
irus
ualo
bjec
tives
Tra
nsiti
onal
Re-
enac
teve
nts
(scr
ipts
)B
asic
sym
bolic
play
“Fee
d”do
ll,“t
ake”
phot
ow
ithca
mer
a1
27
81
Rud
imen
tary
Sym
bolic
Sym
bolic
obje
ctus
eA
dvan
ced
sym
bolic
play
Rol
epl
ayD
emon
stra
tion
teac
h
With
subs
titut
eob
ject
(e.g
.,lo
gba
by)
Play
mot
her’
sor
anot
her’
sro
le
1(2
)1
(2)3
1 13
Soc
ialk
now
ledg
e&
theo
ryof
min
d:un
ders
tand
ing
othe
rs:
beha
vior
s,ro
les,
and
men
tals
tate
sT
rans
ition
alM
irro
rse
lf-re
cogn
ition
Gai
not
her’
sat
tent
ion
Inte
rpre
tvis
ualp
ersp
ectiv
eT
hird
-par
tyre
latio
ns(T
&C
)Pr
e-se
lect
allie
sC
onve
rsat
iona
lcon
tinge
ncy
Impu
tein
tent
ions
Wai
tfor
,voc
aliz
e/ge
stur
eto
gain
atte
ntio
nT
rack
othe
r’s
gaze
(e.g
.,to
getf
ood)
Cur
ryfa
vor
with
pote
ntia
lhel
pers
Con
text
-app
ropr
iate
resp
onse
sU
nfini
shed
,del
iber
ate
(vs.
acci
dent
al)a
cts
12
31
23
3032
33b
391
23
1127
2835
362 2 31
b
1(2)
311
(17)
3439

Tab
le6.
2.(c
ont.)
Soci
aldo
mai
n
Cog
nitio
nSe
ries
/lev
elA
chie
vem
ent
Com
men
tsan
dex
ampl
esSo
urce
sa
Rud
imen
tary
Sym
bolic
Impu
tekn
owle
dge,
com
pete
nce
Em
path
yT
ake
com
plem
enta
ryro
leR
ole
reve
rsal
Coo
pera
tion
(ena
ct,p
lan)
Qua
ntifi
edre
cipr
ocity
2nd-
orde
rin
tent
iona
lity
Tra
nsiti
vity
inso
cial
rank
Com
plex
coal
ition
s/al
lianc
es
Kno
wle
dge-
sens
itive
soci
alac
tivity
Con
sole
(nb.
post
deat
h),m
edia
tere
conc
iliat
ion
Coo
pera
tive
hunt
,rol
e-ba
sed
team
wor
k
Bal
ance
dre
veng
e,sh
are,
lose
rhe
lp2n
d-or
der
tact
ical
dece
ptio
n(w
ithho
ldite
m,
mis
lead
,cou
nter
-dec
eive
),te
ach
Soci
alse
riat
ion
1(2
)311
1921
2829
391
1011
1(2
)31
(2)3
281
(2)3
111
(2)3
1238
1(2
)328
1(2
)33
9
Sen
seof
self
:se
lfaw
aren
essa
ndse
lfun
ders
tand
ing
(cog
nitio
nsab
outt
hese
lf)T
rans
ition
alM
irro
rse
lfre
cogn
ition
Self
labe
lSe
lfco
nsci
ous
beha
vior
Self
conc
ept
Und
erst
and
see-
know
inse
lf
Pers
onal
pron
ouns
As
aca
usal
agen
tK
now
ifyo
ukn
ow,b
ased
onw
haty
ousa
w
12
31
31 1
23
26R
udim
enta
rySy
mbo
licIn
dire
ctse
lfre
cogn
ition
Sens
eof
poss
essi
onSe
lfev
alua
tive
emot
ions
Pict
ure,
shad
ow
Sham
e,gu
ilt,p
ride
112
13 1
Not
es:
aco
gniti
veat
trib
utio
nsba
sed
oncr
iteri
adi
scus
sed
inte
xt;s
ourc
ebr
acke
ted
whe
nm
yat
trib
utio
ndi
ffer
sfr
omth
atof
the
auth
ors
cite
d.b
sym
bol-
trai
ned
grea
tape
ste
sted
.So
urce
s:1,
Park
er&
McK
inne
y19
99;2
,Tom
asel
lo&
Cal
l199
7;3,
Byr
ne19
95;4
,Myo
wa-
Yam
akos
hi&
Mat
suza
wa
1999
;5,B
yrne
&R
usso
n19
98;6
,Whi
ten
1998
b;7,
Rus
son
2002
b;8,
Sudd
endo
rf&
Whi
ten
2001
;9,P
arke
r,C
hapt
er4,
this
volu
me;
10,d
eW
aal&
Aur
eli1
996;
11,B
oesc
h&
Boe
sch-
Ach
erm
ann
2000
;12,
Patt
erso
n&
Lin
den
1981
;13,
Noe
,de
Waa
l&va
nH
ooff
1980
;14,
Rus
son
1999
a;15
,Tan
ner
&B
yrne
unpu
blis
hed;
16,H
are
etal
.200
0;17
,Cal
l&T
omas
ello
1998
;18,
Stoi
nski
etal
.200
1;19
,Whi
ten
2000
;20,
Whi
ten
1998
a;21
,Boy
sen
1998
;22,
Cal
l200
1b;2
3,St
okes
&B
yrne
2001
;24,
Ber
ing,
Bjo
rkla
nd&
Rag
an20
00;
Bjo
rkla
ndet
al.
2002
;25
,C
all,
Agn
etta
&T
omas
ello
2000
;26
,C
all
&C
arpe
nter
2001
;27
,C
all,
Har
e&
Tom
asel
lo19
98,
2000
;H
are
etal
.20
00,
2001
;T
omas
ello
,Cal
l&H
are
1998
;Tom
asel
lo,H
are
&A
gnet
ta19
99;T
omas
ello
,Har
e&
Fogl
eman
2001
;28,
Hir
ata
&M
atsu
zaw
a20
01;2
9,H
irat
a&
Mat
suza
wa
2000
;30
,Hos
tett
er,C
ante
ra&
Hop
kins
2001
;31,
Jens
vold
&G
ardn
er20
00;3
2,L
eave
ns&
Hop
kins
1999
;33,
Men
zel1
999;
34,M
yow
a-Ya
mak
oshi
&M
atsu
zaw
a20
00;
35,I
taku
ra&
Tan
aka
1998
;36,
Peig
not&
And
erso
n19
99;3
7,C
usta
nce
etal
.200
1;38
,Mita
ni&
Wat
ts20
01;M
itani
,Wat
ts&
Mul
ler
2002
;39,
Bla
ke,C
hapt
er5,
this
volu
me.
(Not
e:re
late
dst
udie
sar
egr
oupe
d.)
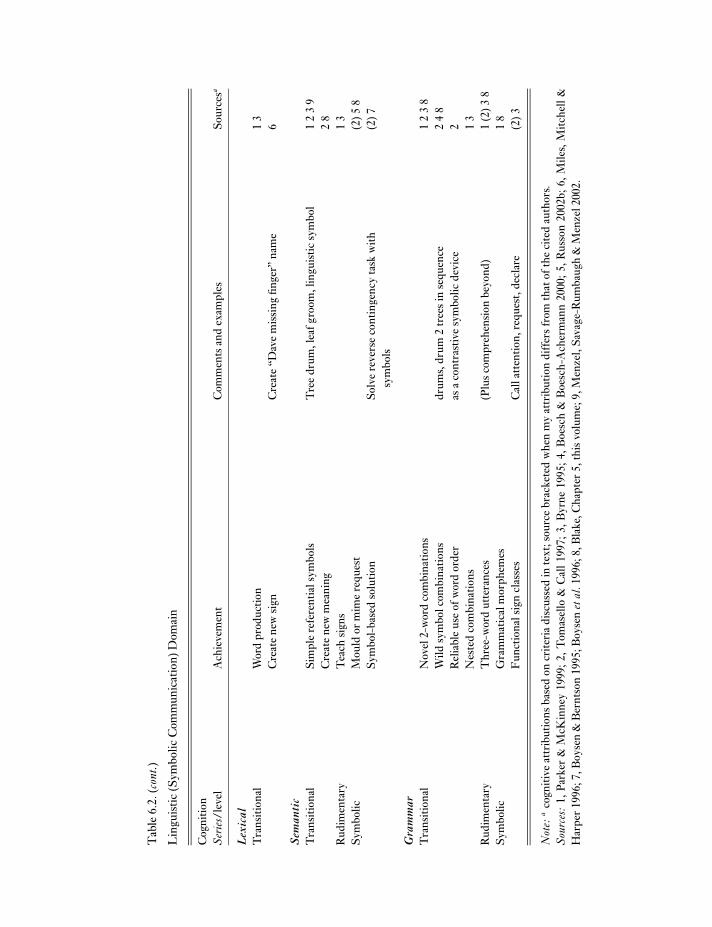
Tab
le6.
2.(c
ont.)
Lin
guis
tic(S
ymbo
licC
omm
unic
atio
n)D
omai
n
Cog
nitio
nSe
ries
/lev
elA
chie
vem
ent
Com
men
tsan
dex
ampl
esSo
urce
sa
Lex
ical
Tra
nsiti
onal
Wor
dpr
oduc
tion
Cre
ate
new
sign
Cre
ate
“Dav
em
issi
ngfin
ger”
nam
e1
36
Sem
anti
cT
rans
ition
alSi
mpl
ere
fere
ntia
lsym
bols
Cre
ate
new
mea
ning
Tre
edr
um,l
eafg
room
,lin
guis
ticsy
mbo
l1
23
92
8R
udim
enta
rySy
mbo
licT
each
sign
sM
ould
orm
ime
requ
est
Sym
bol-
base
dso
lutio
nSo
lve
reve
rse
cont
inge
ncy
task
with
sym
bols
13
(2)5
8(2
)7
Gra
mm
arT
rans
ition
alN
ovel
2-w
ord
com
bina
tions
Wild
sym
bolc
ombi
natio
nsR
elia
ble
use
ofw
ord
orde
rN
este
dco
mbi
natio
ns
drum
s,dr
um2
tree
sin
sequ
ence
asa
cont
rast
ive
sym
bolic
devi
ce
12
38
24
82 1
3R
udim
enta
rySy
mbo
licT
hree
-wor
dut
tera
nces
Gra
mm
atic
alm
orph
emes
Fun
ctio
nals
ign
clas
ses
(Plu
sco
mpr
ehen
sion
beyo
nd)
Cal
latt
entio
n,re
ques
t,de
clar
e
1(2
)38
18
(2)3
Not
e:a
cogn
itive
attr
ibut
ions
base
don
crite
ria
disc
usse
din
text
;sou
rce
brac
kete
dw
hen
my
attr
ibut
ion
diff
ers
from
that
ofth
eci
ted
auth
ors.
Sour
ces:
1,Pa
rker
&M
cKin
ney
1999
;2,T
omas
ello
&C
all1
997;
3,B
yrne
1995
;4,B
oesc
h&
Boe
sch-
Ach
erm
ann
2000
;5,R
usso
n20
02b;
6,M
iles,
Mitc
hell
&H
arpe
r19
96;7
,Boy
sen
&B
ernt
son
1995
;Boy
sen
etal
.199
6;8,
Bla
ke,C
hapt
er5,
this
volu
me;
9,M
enze
l,Sa
vage
-Rum
baug
h&
Men
zel2
002.

86 A. E. RUSSON
of reorganizing or transforming lower-level structuresinto new, higher-level ones (Case 1985, 1992). Eatingwith a spoon, for instance, can be achieved either usinga complex action strategy governed by a combinationof sensorimotor-level motor action schemata or using asimple higher-level strategy that consolidates this com-bination of schemata into one operation on a relation-ship. Importantly, behavior in transitional periods mayowe to cognitive structures at either the lower or higherlevel – here, sensorimotor schemata or simple symbols.
(2) Rudimentary (first-order) symbolic-levelstructures. From about 1.5–2 to 3.5–4 years of age,human children create cognitive structures that repre-sent simple events and relationships among them (Case1985; P&M). Behavioral examples are simple word com-binations, using two tools in interrelated fashion, andsymbolic pretend play.
Several models portray cognitive development inthis phase in terms of creating higher-level cognitivestructures derived from sensorimotor ones, i.e., theyrepresent, in the sense of recoding or redescribing, exist-ing representations. Case (1985) construes this as oper-ating on relations-between-relations, where one rela-tionship is subordinated to another or used as a wayto effect change in another. Included are coordinatingtwo different relationships into one “inter-relational”cognitive structure (e.g., hammer-hit-nut with nut-on-anvil) and coordinating two inter-relational structures.Other models are second-order cognition (Langer 2000)and second-order representations (Leslie 1987). Second-order cognition is exemplified by creating two sets con-currently, so that items are similar within each setand different between sets (e.g., red balls, blue balls);this involves simultaneously managing the relationshipwithin each set (same item class) and a higher-order rela-tionship between two sets (different classes). Second-order representations are derivatives of realistic (first-order) representations, for example using a banana as atelephone. To avoid confusion, Leslie argued, “banana-as-telephone,” must remain linked to its first-order rep-resentation, “banana-as-banana,” yet decoupled from it(i.e., marked as an imagined copy). Making a decoupledcopy requires re-representing an existing representa-tion, so second-order representations are higher-levelstructures.
A competing model of cognition in this rangeis secondary representations (Perner 1991), where re-presentations are subsequent presentations of something
previously present in the mind. Examples are entertain-ing past or future representations of a situation or bring-ing schemata to mind without their normal sensorimo-tor cues. Secondary representation may be what allowscoordinating multiple models of a situation, which mayenable tracking where an object went after it moved alongan invisible trajectory, pretending that an empty cup isfull, or interpreting external representations of a situ-ation. Children in the secondary representation phasecan represent how things might be as well as how theyactually are; previously, they could only represent thelatter (Whiten 1996). Secondary representations, likesecond-order ones, are representations of a situationentertained concurrently with the situation’s realisticor current representation and they represent somethingabout the relations among multiple representations of asituation; differently, secondary representations are nothigher-level structures. They remain pre-symbolic inPerner’s view; strong metarepresentation, which followsthem, is the simplest symbolic process.
(3) Strong (second-order) symbolic-levelstructures. Strong symbolism emerges around 4 yearsof age. Understanding that people can hold false beliefsabout the world is the accepted benchmark (Whiten1996). To Perner, this requires appreciating that othersmay have different thoughts about reality than oneself,i.e., understanding re-presentations as re-presentations(interpretations) or strong metarepresentation. Analternative model is third-order cognition, where third-order structures are structures that encompass multiplesecond-order ones in superordinate–subordinatefashion (Langer 1998, 2000). An example is composingthree matching sets of items, which creates hierarchicalcorrespondences between the sets, i.e., a superordinatecategory subsuming two subordinate, second-orderones. Three sets is the minimum needed for hierarchicalclassification, which enables truly hierarchical cognition(Langer 1998).
Levels in great ape cognition
I attributed cognitive levels to great apes’ complexachievements, per Table 6.2, using recognized indi-cators of early symbolic processes in humans. Indica-tors of rudimentary symbolic-level cognition includedweak hierarchization (e.g., routines that subsume sub-routines), tasks first solved by children between 2 and3.5 years of age, tests of abilities accepted as higher-order

Great ape cognitive systems 87
ones (e.g., analogies), and manipulating relations-between-relations. I interpreted achievements emergingin 1.5- to 2-year-old humans and taken to mark thethreshold of weak symbolism as transitional. I consid-ered levels that original authors attributed but privilegedthe indicators noted above. T&C did not analyze greatape achievements individually, for example, so some oftheir cognitive attributions lack substance. On this basis,I consider the four positions currently entertained on thecognitive levels great apes attain.
(1) Great ape cognition operates with same low-level associative processes attributed to all nonhumanspecies (e.g., Balda, Pepperberg & Kamil 1998). Allthree reviews reject this position because of substan-tial evidence for higher-level cognitive processes in greatapes. Recent informed opinion concurs (e.g., Matsuzawa2001b; Suddendorf & Whiten 2001; Thompson & Oden2000; Table 6.2). Low-level associative processes liketrial-and-error learning and sequential chaining are nec-essary but not sufficient to account for great apes’achievements.
(2) Great apes share with all anthropoid primates acognitive level beyond other mammals, understandingthird-party relations (TPRs) (T&C). T&C define TPRsas interactions among third parties in which the actordoes not participate, for example separating interven-tions and mediating reconciliations. Tomasello’s groupadvocates this position but most other experts disagree(Matsuzawa 2001b, P&M, Russon 1999b, RWB, Sud-dendorf & Whiten 2001, Thompson & Oden 2000).T&C consistently interpret great apes’ achievementswith undue skepticism and monkeys’ with undue gen-erosity; for instance, no evidence supports their claimthat monkeys can perceive, let alone judge, relations-between-relations (Parker 1998, Chapter 4, this volume;Rumbaugh 2000; Russon 1999b; Thompson & Oden2000).
T&C’s relational cognition model is itself prob-lematic (Russon 1999b), although many agree withthem that understanding relational categories andrelations-between-relations is among great apes’ crown-ing achievements. T&C characterize great apes’ rela-tional achievements as understanding TPRs, construedas a generalized ability governed by advanced sen-sorimotor cognition (stages 5 and 6). This cognitionreaches into a transitional range where either sensorimo-tor or symbolic structures can generate achievements.Stage 6 also supports understanding single relational
categories but not relations-between-relations; the lat-ter requires rudimentary symbolic cognition becauseit concerns relations between abstract entities (Case1985). T&C rely exclusively on sensorimotor measures,so they fail to assess whether early symbolic or sen-sorimotor processes generate achievements and theyunderrate achievements involving relations-between-relations, such as great apes’ meta-tool and tool set use.Their TPR model also conflates transitional with rudi-mentary symbolic achievements, confounding two levelsof probable significance in distinguishing great ape frommonkey cognition.
(3) Great apes surpass other nonhuman primates inattaining secondary representation, which may charac-terize the 1.5- to 3.5-year phase in human cognitivedevelopment, but fall short of strong symbolic lev-els (e.g., Suddendorf & Whiten 2001; Whiten 1996).Suddendorf and Whiten’s (2001) review of great apes’achievements on invisible displacements, means–endreasoning, pretense, mirror self-recognition, mentalstate attribution, and understanding imitation supportstheir conclusion that great apes achieve secondary rep-resentation up to the level of 2-year-old humans. This isconsistent with the common characterization that greatapes acquire language abilities up to the level of human2 year olds (e.g., Blake, Chapter 5, this volume).
This review neglects to consider great apes’ highestlevel achievements in pretense and means–ends reason-ing, however, or any of their achievements in logico-mathematical or spatial reasoning (e.g., Langer 1996;Mitchell 2002; Table 6.2) so it does not provide athorough test of position 3. It also emphasizes humanachievements in the second year and underplays thethird, situating it closer to position 2 than position 3.Scale model use and minimal third-order classifying,which humans master in their third year, have beenshown in great apes (Kuhlmeier, Boysen & Mukobi1999; Poti et al. 1999; Spinozzi et al. 1999). Scale modeluse in particular may involve using models as repre-sentations, putting great apes on the brink of strongmetarepresentation.
Secondary representation also fails to account forthe higher level structures that can enrich cognitionbeyond sensorimotor levels, especially those concern-ing relations-between-relations. Great apes’ complexfeeding techniques and their logical and quantitativeachievements offer prime evidence of such higher-levelcognitive structures (Byrne & Byrne 1991; Byrne et al.

88 A. E. RUSSON
2001; Langer 2000; Matsuzawa 1996, 2001b; P&M;Russon 1998; Spinozzi & Langer 1999). The secondaryrepresentation concept fails to address the structure thatindividuals may add to a representation by re-presentingit or precisely how multiple representations of a situa-tion are related; models proposing higher-order struc-tures fill this gap (Case 1985; Leslie 1994; Whiten 2000).Actors may not only recall alternative realistic repre-sentations of a situation (e.g., past, future), for instance,they may also re-represent the situation differently fromany reality they have experienced (e.g., a banana as atelephone) and/or at a higher level (one relationshipvs. multiple schemata). While the secondary represen-tation concept is valuable in suggesting where higher-level cognitive structures are not used to entertain mul-tiple representations of a situation, it fails to considercircumstances in which they are.
(4) Great apes surpass other nonhuman primatesin attaining rudimentary symbolic-level cognition (e.g.,RWB; Langer 1996; Matsuzawa, 2001b; P&M; Russon1998, 1999a). P&M, RWB, and many recent studies(Table 6.2) support this position for all great ape species,in all cognitive domains, based on recognized indicesof weak symbolism (weak hierarchization, abilities rec-ognized to involve higher-order processes, relations-between-relations). Comparable achievements claimedfor monkeys have been shown to involve performancesbased on response rules generated by simpler processes,probably associative ones (Parker, Chapter 4, this vol-ume; Thompson & Oden 2000).
Many current disagreements stem from whatassessment tools are used and what meanings of sym-bolism, metarepresentation, and hierarchization areapplied (Whiten 1996). With the meanings and assess-ments used here, the best interpretation of current evi-dence is that great apes attain rudimentary symbolic-level cognition and in this, they surpass other nonhumanprimates.
The levels that great apes achieve within therudimentary symbolic range are relatively uncharted.Assessment remains an impediment because many cur-rent tests for symbolism use threshold criteria (e.g.,metarepresentation, hierarchization). Indices of earlysymbolic levels have been used in a few cases, e.g., num-ber of relational operators, complexity of classification,depth of hierarchies, or human age norms (Byrne &Russon 1998; Kuhlmeier et al. 1999; Matsuzawa 2001b;P&M; Poti’ et al. 1999; Russon & Galdikas unpublished;
Spinozzi et al. 1999; Thompson & Oden 2000). Thesesuggest great apes’ cognitive ceiling at a hierarchicaldepth of about three levels (e.g., use a hammer stone tohit (a nut placed on (an anvil stone placed on a wedge,to level it)) – Matsuzawa 2001b; and see Yamakoshi,Chapter 9, this volume), coordinating three object–object relations in one inter-relational structure (e.g.,coordinate anvil-on-wedge, nut-on-anvil, and hammer-hit-nut – P&M; Russon & Galdikas unpublished), orminimal third-order classification (e.g., create three con-temporaneous sets with similar items within sets anddifferences between sets – Langer 2000; Poti’ et al.1999; Spinozzi et al. 1999). All remain consistent withPremack’s (1988) rule of thumb, that great apes reachlevels like 3.5-year-old children but not beyond.
COGNITIVE INTERCONNECTION:THE ORCHESTRALLY MINDED
Cognitive facilitation refers to achievements madethrough interplay among different types of cognition.It is an important source of an actor’s cognitive power:tasks that require interconnecting several abilities can besolved, and individual abilities can advance by exploitingother abilities (Langer 1996). Cracking a nut, forexample, might require using a stone hammer–anviltool set (means–end reasoning), identifying a substitutewhen the best hammer is unavailable (logical reason-ing), and obtaining the substitute from a companion(social cognition), or classification abilities might beextended by categorizing according to causal utility.Cognitive facilitation almost certainly occurs in greatapes. Chimpanzees skilled in symbol use solved anal-ogy problems better than chimpanzees without symbolskills, for example (Premack 1983; Thompson, Oden &Boysen 1997).
Facilitation has received little attention in greatapes despite its implications for cognitive architecture.If it occurs, especially across domains, then qualitativelydifferent cognitive structures can operate and inter-act beyond the bounds of the problem types for whichthey were designed: that is, the cognitive system cannotsimply comprise a collection of independent, special-purpose modules. Facilitation is also important com-paratively because it has been claimed to be uniquelyor at least characteristically human, for whom it hasbeen likened to fluidity of thought, multiple intelligencesfunctioning seamlessly together, a passion for the

Great ape cognitive systems 89
analogical, and mapping across knowledge systems (e.g.,Gardner 1983; Mithen 1996). What enables facilitationis unresolved. Hypotheses include analogical reasoning,which transfers knowledge from one problem type toanother (Thompson & Oden 2000), or synchronizingdevelopmental progress across distinct types of cogni-tion so that their structures build upon common expe-riences, which promote interplay by serving as bridgesbetween cognitive structures (Langer 1996, 2000). Pos-sibilities typically require hierarchization; analogicalreasoning, for instance, involves judging if one relation-ship is the same as another, i.e., logical equivalences atabstract levels, which is founded on the ability to judgerelations-between-relations.
In part, systematic evidence on facilitation in greatapes is meager because studies of nonhuman cognitionhave tended to control against using multiple abilitiesin aiming for clean tests of single abilities. Among thefew sources of systematic evidence are studies of logic,which show that analogical reasoning is within the nor-mal reach of great apes but not other nonhuman primates(Oden, Thompson & Premack 1990; Thompson et al.1997; Thompson & Oden 2000). For feral great apes,P&M is the only review to have systematically consid-ered achievements that may involve facilitation. I con-sider evidence for facilitation across physical, logical,and social domains as the most important in compara-tive perspective.
Logical–PhysicalGreat apes interconnect logical with physical cognitionwhen they classify items by function or functional rela-tions, for example sort items into sets of toys and toolsor sort bottles with caps (Savage-Rumbaugh et al. 1980;Tanaka 1995; Thompson & Oden 2000), use substi-tute tools (Figure 6.1), or classify foods on the basis ofthe technique for obtaining them (Russon 1996, 1999a,2002a). A rehabilitant orangutan stored termite nestfragments on specific parts of his body, in the order inwhich he planned to open them, to streamline his termiteforaging (Russon 2002a) and a rehabilitant chimpanzeemade and used a seriated set of stick tools (ordered fromsmallest to largest) to extract honey from a bee’s nest(Brewer & McGrew 1990).
Social–physicalGreat apes use socially mediated learning in acquiringfood processing skills (Boesch 1991; Byrne & Byrne
Figure 6.1. Princess, an adult female rehabilitant orangutan, blowson the burning tip of a mosquito coil. A paper marked with two dotsis at her feet. She had drawn the dots by touching the coil’s burningtip to the paper, i.e., substituting the coil for a pen. She oftenscribbled in notebooks with pens, so she used a functional similaritybetween pens and the coil, that both have tips that can mark paper.She did not simply confuse the two tools. She drew differentlywith the coil (touch vs. scribble) and she fixed it differently (if apen did not mark when she scribbled, she fixed it by biting at itstip or by clicking the pen’s switch to advance the tip; to fix her coil,she blew on its tip).
1993; Inoue-Nakamura & Matsuzawa 1997; Matsuzawa& Yamakoshi 1996; Russon 1999a, 2003a,b). When theyuse imitation or demonstration to advance complexfood processing skills, social cognition contributes tophysical cognition at rudimentary symbolic levels. Themost complex cases known concern stone nut-crackingin west African chimpanzees: mothers demonstrate totheir offspring how to use stone hammers, and offspringreplicate the techniques they were shown (Boesch 1991,1993). Mithen (1996) argued that food sharing, used as

90 A. E. RUSSON
a medium for social interaction with formalized sharingrules, uses “natural history” cognition to enhance socialproblem solving. If so, chimpanzees show this capability:they share meat in rule-governed fashion to serve socialfunctions and social relationships are important in dis-tribution (Boesch & Boesch-Achermann 2000; Goodall,1986; Mitani & Watts 2001; Mitani, Watts & Muller2002).
Logical–Social–SymbolicBoysen et al. (1996) used a reverse contingency taskto test if chimpanzees could select the smaller of twoarrays to gain greater rewards against a social competi-tor. Boysen showed two dishes of candies to a dyad ofsymbol-trained chimpanzees, had one choose a dish bypointing, and then gave the chosen dish to the otherchimpanzee and the leftover dish to the chooser. Shownreal candies, choosers consistently picked the dish withmore – to their disadvantage. When number symbolsreplaced candies, choosers consistently picked the dishwith fewer – to their advantage. Symbols improved thesechimpanzees’ ability to solve a quantification (logical)problem. Orangutans also solve this task, without sym-bol skills and using real candies (Shumaker et al. 2001).If subjects interpreted this as a competitive social task, asintended, their quantification (logical) abilities assistedtheir social problem solving.
Complex facilitationsSome expertise taps all three domains interactively. Themost complex is chimpanzee cooperative hunting inthe Taı forest (Boesch & Boesch-Achermann 2000).Once a hunting group detects a red colobus group,the ideal hunt has four phases involving four roles(driver, chaser, ambusher, captor). Participants must beable to alter their actions flexibly and rapidly to trackcolobus’ attempts to escape; they also take differentroles and accommodate their actions to chimpanzeesin other roles. If successful, they share the meat for-mally according to each participant’s role in the hunt,age, and dominance. Successful cooperative hunting inthe forest, a three-dimensional space with low visibility,requires hunters to “perceive other hunters as indepen-dent agents with their own intentions and competence,attribute abilities to the prey that differ from those ofconspecifics, and understand the causality of the exter-nal relation between prey and other hunters” (Boesch& Boesch-Achermann 2000: 242). It requires cognitiveabilities in the physical domain (space – arboreal loco-
motion and routes; causality – predicting how chasing,blocking, or driving will affect colobus’ flight path andthe canopy), the social domain (self-manipulating thepresentation of oneself to the colobus; figuring one’sweight into arboreal travel; enacting complementaryroles), and the logical domain (quantifying how to dis-tribute meat sharing). Hunters can change roles repeat-edly over the course of a hunt, so some must have allor most of these cognitive capabilities and use them ininterconnected fashion.
Evidence for cognitive facilitation jibes with thecomplex, varying, and multifaceted challenges facinggreat apes in their natural habitat (Boesch & Boesch-Achermann 2000; Russon in 2003b). Their foragingoffers a prime example: it calls for a wide spectrumof abilities to organize biological knowledge, constructforaging techniques, acquire alternative strategies, andnegotiate cooperative and competitive social foragingsituations (Russon 2002a; Stokes 1999). The multi-faceted nature of complex foraging tasks calls for com-bining high-level abilities, and interactions among taskcomponents call for interconnecting them. Evidence forcognitive facilitation also jibes with evidence that greatapes spontaneously transfer expertise from one domainto another (Thompson & Oden 2000), with Parker’s(1996) apprenticeship model of interconnected physicaland social abilities, and with arguments that intercon-necting mechanisms of some sort are essential to cogni-tive systems that handle different types of information inparallel using distinct modules (Mithen 1996). It clearlyrefutes strictly modular models of cognition in greatapes.
GENERATING GREAT APECOGNITION
The variability and flexibility of great apes’ cognitiveabilities, including the capacity to generate unusual abili-ties as needed and the roughly consistent cognitive ceil-ing across abilities, domains, and species, suggest thattheir cognitive systems may be better characterized bythe processes that generate them than by specific abilitiessuch as tool use or self-concept. Generative processes areconsidered below.
Development and culture
Developmental models of human cognition have prob-ably been fruitful in studying great apes because their

Great ape cognitive systems 91
cognitive structures develop in similar fashion (Langer1996, 2000; P&M). Like humans, great apes experi-ence extensive and lengthy sensory, motor, socio-sexual,brain, and cognitive development that is affected byage and experience and is concentrated in immaturity(Boesch & Boesch-Achermann 2000; Inoue-Nakamura& Matsuzawa 1997; Langer 1996; Matsuzawa 2001b;P&M; Poti et al. 1999; Russon in 2003b; Spinozzi et al.1999). Their complex structures develop on the basisof simpler ones and emerge late in immaturity (Langer1996, 2000; Matsuzawa 2001b; P&M). Their complexforaging techniques, for example, develop piecemealover many years with youngsters first acquiring basicelements, next assembling them into a basic strat-egy, then gradually elaborating it (Fox, Sitompul &van Schaik 1999; Inoue-Nakamura & Matsuzawa 1997;Russon 2002a; 2003a).
In life history perspective, developmental mod-els are also consistent with evidence that: (1) cogni-tive capacity peaks in juveniles and levels off after ado-lescence; (2) parents contribute to acquiring advancedjuvenile as well as basic infant skills; (3) rudimentarysymbolic level abilities emerge post-infancy, around themove to semi-independent life; (4) most adult-levelexpertise is mastered by adolescence, around the moveto fully independent life; and (5) post-adolescent learn-ing seems less flexible (Boesch 1991; Boesch & Boesch-Achermann 2000; Ingmanson 1996; Inoue-Nakamura& Matsuzawa 1997; King 1994; Parker 1996; P&M). Allcorrelate with the slower pace and disproportionatelyprolonged immaturity that distinguish great ape devel-opment from that of other nonhuman primates (P&M;Kelley, Chapter 15, Ross, Chapter 8, this volume). Com-pared with humans, great apes’ cognitive developmentis faster in the first year of life but subsequently slower(P&M; Poti et al. 1999; Spinozzi et al. 1999), whichexplains why some of the distinctive abilities they sharewith humans develop later and persist longer.
Social–cultural influences, interwoven with indi-vidual experience, also contribute to cognitively gov-erned achievements in great apes, as they do in humans(e.g., P&M; Tomasello 1999; T&C; van Schaik et al.2003; Whiten et al. 1999). The distribution of “atypical”abilities and some complex skills in the wild, for instance,shows that great apes may not realize some com-plex achievements without appropriate socio-culturalsupport despite appropriate individual opportunities(van Schaik et al. 2003; Whiten et al. 1999). If theirachievements are products of combining socio-cultural
with individual experience during development, thenenculturation should be primarily responsible. In greatapes enculturation probably resembles apprenticeship(guided participation in shared activities of a routinenature; Rogoff 1992) and supports and perhaps extendstheir natural behavioral repertoires (Boesch & Boesch-Achermann 2000; Matsuzawa et al. 2001; Parker 1996;P&M; Russon 1999b, 2003b; Suddendorf & Whiten2001). It has been assigned responsibility for achieve-ment variability across wild, captive-reared, and human-enculturated great apes (e.g., Donald 2000; T&C).
Great apes’ cultural and cognitive processes aremore tightly interwoven than this scenario suggests.Cultural processes depend on what information canbe shared and how, which depend on information pro-cessing capabilities, i.e., cognition. Great apes’ culturalprocesses may be exceptionally powerful among non-human primates because they access high-level cogni-tive capabilities unique to great apes and humans (e.g.,imitation, self-awareness, demonstration; Parker 1996).Conversely, great apes’ cognitive achievements are prob-ably boosted by cultural processes. Chimpanzee cul-tures show ratcheting, for instance, the accumulationof cultural variants over time, in the form of cumulativemodifications to complex techniques (McGrew 1998;Yamakoshi & Sugiyama 1995). This probably allowslearners to acquire more complex techniques than theywould have constructed independently. If enculturationhas a special role to play in cognitive development, itmay primarily affect high levels, as it typically does inhumans (P&M; Tomasello 1999). No convincing evi-dence exists, however, for claims that human encul-turation induces higher-level cognitive structures ingreat apes than species-normal enculturation (Boesch &Boesch-Achermann 2000; Langer 2000; P&M; Russon1999b; Spinozzi et al. 1999; Suddendorf & Whiten2001).
Generating cognitive structures
A final issue is what mental processes generate great apecognitive development and how, especially their distinc-tive cognitive structures. Great apes consistently attainthe same cognitive level across cognitive domains, rudi-mentary symbolism, which suggests that centralizedgenerative processes that operate across the whole cog-nitive system govern their cognitive development, ratherthan processes specific to a single cognitive domain orproblem type. That the level achieved supports simple

92 A. E. RUSSON
symbols suggests hierarchization as a good candidatefor that centralized generative process: It is consideredessential to the cognitive abilities and achievements thatdistinguish the great apes among nonhuman primates(e.g., simple language, abstract level problem solving,complex tool and manual foraging techniques) (Case1985; Piaget 1954) and, among nonhuman primates,only great apes show evidence of hierarchization (e.g.,Byrne 1997; Langer 1996; Matsuzawa 2001b; Russonet al. 1998).
Great apes’ flexible range of high-level cognitiveabilities could be generated by hierarchization usedin conjunction with combinatorial mechanisms, in theform of hierarchical mental construction (e.g., Byrne1997; Gibson 1990, 1993; Langer 1994, 1996). Com-binatorial mechanisms are centralized generative pro-cesses that combine, decompose, and recombine mul-tiple mental units at a time, as in combining actions orobjects in sequence; they probably generate cognitivestructures in all primates (Langer 2000). The patternin which great apes acquire food processing techniquesis consistent with a hierarchical mental constructionmodel of cognitive development (e.g., Inoue-Nakamura& Matsuzawa 1997; Russon 2002a; Stokes & Byrne2001). Infant chimpanzees acquiring stone nut-crackingskills, for instance, first learn the individual basic actionsneeded to crack nuts and apply single actions to singleobjects (only stone, only nut); next, they apply mul-tiple actions to multiple objects (some stones, some nuts,stones and nuts) combined in sequence (some are inef-fective, e.g., put a nut on a stone but hit the nut witha hand then pick up a piece of kernel from a brokenshell on the ground and eat that); finally, they integrateappropriate combinations into more complex, hier-archically organized techniques showing understandingof action–object relationships (Inoue-Nakamura &Matsuzawa 1997). To date, other nonhuman primateshave not shown hierarchically organized techniques(Harrison 1996). Great apes reach only rudimentarysymbolic levels, however, and their achievements arerougher-grained than humans’, i.e., focused primarilyon general problem features and less able to incorpo-rate fine ones (Langer 1996; P&M; Spinozzi & Langer1999). Their low symbolic ceiling may reflect limitedhierarchization relative to humans, described as shal-low (Byrne 1997; Matsuzawa 2001b) or protohierarchi-cal (Langer 2000). The rougher grain may reflect lowerlimits on the number of units they can combine at once.
Cognitive facilitation may take great ape cognitionbeyond modularity, and it may hinge on hierarchization(RWB; Case 1985; Karmiloff-Smith 1992; Thompson& Oden 2000; see Langer 1996, 2000 for alternatives).3
This link is supported by evidence that cognitive facili-tation in great apes is limited, because this is consistentwith shallow hierarchization. Shallow hierarchizationgenerates only rudimentary hierarchical cognitive struc-tures, which remain more isolated than the higher-levelcognitive structures that human hierarchization gener-ates (Case 1985).
This sort of model, which characterizes greatape cognition in terms of central generative processes,may help explain several features that have puzzledscholars. The “atypical” abilities that emerge in greatapes under highly nurturing human rearing conditions(e.g., linguistic and mathematical abilities: Gardner,Gardner & van Cantfort 1989; Tomasello 1999) maysimply be customized abilities of the sort expected fromgenerative cognitive systems that build structures to suitthe specific challenges encountered during development(Boesch & Boesch-Achermann 2000; P&M; Rumbaugh2000; Swartz, Sarauw & Evans 1999). Marked individ-ual differences in achievements may similarly be normalfeatures of generative cognitive systems. “Atypical” abil-ities may also have feral counterparts, making them lessunusual than suggested. Feral communication sugges-tive of symbolism has been reported, for example treedrumming, leaf clipping, knuckle-knocking, demonstra-tion teaching (Boesch 1991, 1993, 1996), symbolic eat-ing (Schaller 1963, Russon 2002b), miming requests(Russon 2002b), and placing leaves to indicate traveldirection (Savage-Rumbaugh et al. 1996) (see also Blake,Chapter 5, this volume), as have complex quantita-tive abilities such as seriation (arranging items in agraded series: Brewer & McGrew 1990) and body-part counting (using body parts to order items: Russon2002a).
Generative models also suggest how modularity–generality may play out in great apes. In humans,module-like structures may be products of genera-tive processes operating in the context of problem-specific constraints and innately founded structures(e.g., Elman et al. 1996; Greenfield 1991; Karmiloff-Smith 1992; Langer 2000; P&M). Human cognitivestructures change with development: they have beencharacterized as relatively undifferentiated at their earl-iest (i.e., applicable generally, across problem types),

Great ape cognitive systems 93
subsequently differentiated into domain- and problem-specific structures with module-like features (applicableto specific problem types), and finally interconnected(applicable across problem types when used in combi-nation) (e.g., Case 1996; Greenfield 1991; Langer 1996,2000; P&M). Generalized capabilities may then existin undifferentiated and interconnected forms, module-like structures may be developmental products, anddevelopment may affect the qualities of modularity andgenerality and the balance between them. Great apes’cognitive development shows similar patterns althoughcomparatively, differentiation and interconnection pro-ceed more slowly after the first year and are ultimatelyless powerful (e.g., P&M). Great ape cognition theninvolves both modularity and generality, and charac-terizations in terms of generality–modularity are likelyflawed if they fail to consider developmental change or todistinguish undifferentiated from interconnected formsof generality (e.g., Mithen 1996).
DISCUSSION
Evidence consistently supports conclusions that greatapes differ cognitively from other nonhuman primates.Virtually all experts agree, there is no longer any justi-fication for reducing great ape cognition to associativeprocesses or lumping great apes with other nonhumanprimates. What sets great ape cognitive achievementsapart is not specific problem-specific abilities such as tooluse, imitation, or self-concept. It is rather the broaderand more open repertoire of abilities, rudimentary sym-bolic levels achieved across domains, and limited inter-connectedness among them. What underpins this suiteof cognitive structures may be centralized generativeprocesses that operate ontogenetically, limited hier-archization and perhaps facilitation being the bestcurrent candidates. This characterization is not new.Revisiting it, however, helps articulate what needs evolu-tionary explanation: more powerful generative processesthat produce rudimentary symbolism and limited fluid-ity of thought.
This characterization helps explain why it has beendifficult to get a handle on great ape mentality. First,if variable achievement is intrinsic to great ape cogni-tion, then studies that have tested great apes as imma-tures or reared in non-stimulating environments havefailed to tap their full potential. Second, achievementsduring the transition from sensorimotor to rudimentary
symbolic cognition may be governed by either advancedsensorimotor or primitive symbolic-level structures(Case 1985). It is possible to distinguish the two behav-iorally, and studies that failed to do probably under-estimated subjects’ level of cognitive functioning (e.g.,Byrne & Russon 1998; Russon 1998). Third, if enter-taining multiple representations of a situation under-pins rudimentary symbolic-level cognition then greatapes, like 2- to 3.5-year-old children, should be able toentertain symbolic and perceptual representations con-currently. In such children, when the two representa-tions conflict, perceptual representations tend to over-ride symbolic ones for control of behavior; they havebeen described as perception bound because they areeasily swayed by perceptual cues (Case 1996; P&M).Chimpanzees have shown similar tendencies. Theysolved a reverse contingency task (what you pick goesto your partner) when it was presented with symbolsbut failed when it was presented with real candies,so they can function symbolically but not when per-ceptual cues are salient (Boysen et al. 1996, 1999).This suggests that their symbol use is unstable andthey, like young children, may fail symbolic tasks notbecause they lack the capability but because percep-tual cues activate this bias. Orangutans without sym-bol skills solved the reverse contingency task with realcandies (Shumaker et al. 2001), so even great apes cansometimes privilege symbolic over perceptual solutions.These difficulties do not render it futile to study rudi-mentary symbolic cognition in great apes: many diffi-culties are assessment related and have been resolvedfor humans. What is needed is greater attention to thequalities of rudimentary symbolic cognition and fac-tors that contribute to variability in its development andapplication.
The characteristics of great ape cognition thatrequire evolutionary explanation are among those cur-rently treated as evolutionary achievements of thehuman lineage. That these qualities appeared earlier inprimate evolutionary history does not alter their signif-icance but it does change their role, from innovationsof the human lineage to foundations for its elaborations.This affects evolutionary reconstructions of human cog-nition that use great apes to represent the ancestral cog-nition from which human cognition evolved becausethey typically assume great apes to be incapable of sym-bolic cognition (see Russon, Chapter 1, this volume). Wenow know that this assumption is incorrect, in at least

94 A. E. RUSSON
one form. Accurate models of great ape cognition arethen important next steps towards better understand-ing of great ape and human cognitive evolution.
ACKNOWLEDGMENTS
My thanks to David Begun and Sue T. Parker for valu-able comments on earlier drafts of this paper. Theresearch on which this chapter was based was sup-ported by funding from the Natural Sciences and Engi-neering Council of Canada, Glendon College, and YorkUniversity.
1 Feral, here, includes wild and reintroduced individuals liv-
ing free in natural habitat. I grouped reintroduced with wild
great apes because both face species-typical rather than human-
devised problems. Their achievements may differ in their speci-
fic nature (e.g., reintroduced orangutans often show complex
tool use but wild ones rarely do) but not in cognitive complexity,
which is the major concern here (Russon 1999b).
2 (1) This body of evidence is expected to be small because complex
achievements should be rare relative to average ones. (2) If evi-
dence on great ape cognition is notoriously patchy, evidence on
complex cognition should be even more so. (3) On tasks tapping
an actor’s highest-level capabilities, high performance variability
is expected (Spinozzi et al. 1999; Swartz et al. 1999). (4) Method-
ological confounds can cause performance variation, especially
misleading cues that undermine performance and scaffolding
that boosts it. The number of items that must be held concur-
rently in working memory to solve a task affects success for
example, and how a task is presented can increase or decrease
that number (Pascual-Leone & Johnson 1999). If threshold tests
are used, such confounds can affect assessments of cognitive
levels. (5) Few studies have verified that their tests for great
apes are commensurate with human benchmarks; close scrutiny
often shows they are not (e.g., P&M). (6) In children at rudi-
mentary symbolic levels, perceptual processes readily dominate
symbolic ones and unstable achievement is common. If great
apes function at this level, comparable instabilities are prob-
able (e.g., Boysen et al. 1996; Case 1985; Boysen, Mukobi &
Berntson 1999).
3 Similar suggestions use terms like representational redescription
(Karmiloff-Smith 1992), abstract level generalization (RWB),
higher levels of abstraction (Case 1985), and analogical reason-
ing (Thompson & Oden 2000). Langer’s (1996, 2000) alternative
is that facilitation may owe to developmental synchronization,
i.e., yoking developmental progress across distinct types of
cognition so they develop together rather than independently;
this offers the best timing pattern possible for interconnecting
them.
Anderson, J. R. (1996). Chimpanzees and capuchin monkeys:comparative cognition. In Reaching into Thought: TheMinds of the Great Apes, ed. A. E. Russon, K. A. Bard &S. T. Parker, pp. 23–56. Cambridge, UK: CambridgeUniversity Press.
Balda, R. P., Pepperberg, I. M. & Kamil, A. C. (1998).Preface. In Animal Cognition in Nature: The Convergenceof Psychology and Biology in Laboratory and Field, ed.R. Balda, I. Pepperberg & A. Kamil, pp. vii–ix. NewYork: Academic Press.
Beran, M. J. (2001). Summation and numerousnessjudgments of sequentially presented sets of items bychimpanzees (Pan troglodytes). Journal of ComparativePsychology, 115(2), 181–91.
Bering, J. M., Bjorkland, D. F. & Ragan, P. (2000). Deferredimitation of object-related actions in human-rearedjuvenile chimpanzees and orangutans. DevelopmentalPsychobiology, 36, 218–32.
Bermejo, M. & Illera, G. (1999). Tool-set for termite-fishingand honey extraction by wild chimpanzees in the LossiForest, Congo. Primates, 40(4), 619–27.
Biro, D. & Matsuzawa, T. (1999). Numerical ordering in achimpanzee (Pan troglodytes): planning, executing, andmonitoring. Journal of Comparative Psychology, 113(2),178–85.
Bjorkland, D. F., Yunger, J. L., Bering, J. M. & Ragan, P.(2002). The generalization of deferred imitation inenculturated chimpanzees (Pan troglodytes). AnimalCognition, 5, 49–58.
Boesch, C. (1991). Symbolic communication in wildchimpanzees. Human Evolution, 6(1), 81–90.
(1993). Aspects of transmission of tool-use in wildchimpanzees. In Tools, Language, and Cognition inHuman Evolution, ed. K. R. Gibson & T. Ingold,pp. 171–83. Cambridge, UK: Cambridge UniversityPress.
(1996). Three approaches for understanding chimpanzeeculture. In Reaching into Thought: The Minds of theGreat Apes, ed. A. E. Russon, K. A. Bard, & S. T.Parker, pp. 404–29. Cambridge, UK: CambridgeUniversity Press.

Great ape cognitive systems 95
Boesch, C. & Boesch-Achermann, H. (2000). TheChimpanzees of the Taı Forest: Behavioural Ecology andEvolution. Oxford: Oxford University Press.
Boysen, S. T. (1998). Attribution processes in chimpanzees:Heresy, hearsay, or heuristic? Paper presented at the17th Congress of the International PrimatologicalSociety, Antananarivo, Madagascar.
Boysen, S. T. & Berntson, G. G. (1995). Responses toquantity: Perceptual versus cognitive mechanisms inchimpanzees (Pan troglodytes). Journal of ExperimentalPsychology, Animal Behavior Processes, A21 (1), 82–6.
Boysen, S. T. & Hallberg, K. I. (2000). Primate numericalcompetence: contributions toward understandingnonhuman cognition. Cognitive Science, 24(3), 423–43.
Boysen, S. T., Berntson, G. G., Hannan, M. B. & Cacioppo,J. T. (1996). Quantity-based inference and symbolicrepresentations in chimpanzees (Pan troglodytes).Journal of Experimental Psychology: Animal BehaviorProcesses, 22, 76–86.
Boysen, S. T., Berntson, G. G. & Mukobi, K. L. (2001). Sizematters: impact of size and quantity on array choice bychimpanzees (Pan troglodytes). Journal of ComparativePsychology, 115, 106–10.
Boysen, S. T., Mukobi, K. L. & Berntson, G. G. (1999).Overcoming response bias using symbol representationof number by chimpanzees (Pan troglodytes). AnimalLearning and Behavior, 27, 229–35.
Brewer, S. & McGrew, W. C. (1990). Chimpanzee use of atool-set to get honey. Folia Primatologica, 54, 100–4.
Byrne, R. W. (1995). The Thinking Ape. Oxford: OxfordUniversity Press.
(1997). The technical intelligence hypothesis: analternative evolutionary stimulus to intelligence? InMachiavellian Intelligence II: Extensions and Evaluations,ed. R. W. Byrne & A. Whiten, pp. 289–311. Cambridge,UK: Cambridge University Press.
Byrne, R. W. & Byrne, J. M. E. (1991). Hand preferences inthe skilled gathering tasks of mountain gorillas (Gorillagorilla berengei). Cortex, 27, 521–46.
(1993). Complex leaf-gathering skills of mountain gorillas(Gorilla g. beringei): variability and standardization.American Journal of Primatology, 31, 241–61.
Byrne, R. W. & Russon, A. E. (1998). Learning by imitation:a hierarchical approach. Behavioural and Brain Sciences,21, 667–721.
Byrne, R. W., Corp, N. & Byrne, J. M. E. (2001). Estimatingthe complexity of animal behaviour: how mountaingorillas eat thistles. Behaviour, 138, 525–57.
(2000). Estimating and operating on discrete quantities inorangutans (Pongo pygmaeus). Journal of ComparativePsychology, 114, 136–47.
(2001a). Object permanence in orangutans (Pongopygmaeus), chimpanzees (Pan troglodytes), and children(Homo sapiens). Journal of Comparative Psychology,115(2), 159–71.
(2001b). Body imitation in an enculturated orangutan(Pongo pygmaeus). Cybernetics and Systems, 32, 97–119.
Call, J. & Carpenter, M. (2001). Do apes and children knowwhat they have seen? Animal Cognition, 4, 207–20.
Call, J. & Rochat, P. (1996). Liquid conservation inorangutans (Pongo pygmaeus) and humans (Homosapiens): individual differences and perceptual strategies.Journal of Comparative Psychology, 110(3), 219–32.
Call, J. & Tomasello, M. (1998). Distinguishing intentionalfrom accidental actions in orangutans (Pongo pygmaeus),chimpanzees (Pan troglodytes), and human children(Homo sapiens). Journal of Comparative Psychology,112(2), 192–206.
Call, J., Agnetta, B. & Tomasello, M. (2000). Cues thatchimpanzees do and do not use to find hidden objects.Animal Cognition, 3, 23–4.
Call, J., Hare, B. A. & Tomasello, M. (1998). Chimpanzeegaze following in an object-choice task. AnimalCognition, 1, 89–99.
Carruthers, P. & Smith, P. K. (ed.) (1996). Theories of Theoriesof Mind. Cambridge, UK: Cambridge University Press.
Case, R. (1985). Intellectual Development from Birth toAdulthood. Orlando, FL: Academic Press.
(1992). The Mind’s Staircase: Exploring the ConceptualUnderpinnings of Children’s Thought and Knowledge.Hillsdale, NJ: Lawrence Erlbaum Associates.
(1996). Introduction: reconceptualizing the nature ofchildren’s conceptual structures and their developmentin middle childhood. In The Role of Central ConceptualStructures in the Development of Children’s Thought(Series No. 246, Vol. 61), ed. R. Case & Y. Okamoto,pp. 1–26. Chicago, IL: University of Chicago Press.
Corp, N. & Byrne, R. W. (2002). The ontogeny of manualskill in wild chimpanzees: evidence from feeding on thefruit of Saba florida. Behaviour, 139(1), 137–68.
Custance D. M., Whiten, A., Sambrook, T. & Galdikas, B.(2001). Testing for social learning in the “artificial fruit”processing of wildborn orangutans (Pongo pygmaeus),Tanjung Puting, Indonesia. Animal Cognition, 4, 305–13.
Deacon, T. W. (1997). The Symbolic Species: The Coevolutionof Language and the Brain. New York: W. W. Norton.

96 A. E. RUSSON
de Blois, S. T., Novak, M. A. & Bond, M. (1998). Objectpermanence in orangutans (Pongo pygmaeus) andsquirrel monkeys (Saimiri sciureus). Journal ofComparative Psychology, 112(2), 137–53.
de Waal, F. B. M. & Aureli, F. (1996). Consolation,reconciliation, and a possible cognitive differencebetween macaques and chimpanzees. In Reaching intoThought: The Minds of the Great Apes, ed. A. E. Russon,K. A. Bard & S. T. Parker, pp. 80–110. Cambridge, UK:Cambridge University Press.
Donald, M. (1991). Origins of the Modern Mind: Three Stagesin the Evolution of Cognition and Culture. Cambridge,MA: Harvard University Press.
(2000). The central role of culture in cognitive evolution:a reflection on the myth of the “isolated mind.” InCulture, Thought, and Development, ed. L. P. Nucci,G. B. Saxe & E. Turiel, pp. 19–38. Mahwah, NJ:Lawrence Erlbaum Associates.
Elman, J., Bates, E., Johnson, M., Karmiloff-Smith, A.,Parisi, D. & Plunkett, K. (1996). Rethinking Innateness.Cambridge, MA: MIT Press.
Fox, E. A., Sitompul, A. F. & van Schaik, C. P. (1999).Intelligent tool use in wild Sumatran orangutans. In TheMentalities of Gorillas and Orangutans: ComparativePerspectives, ed. S. T. Parker, R. W. Mitchell & H. L.Miles, pp. 99–116. Cambridge, UK: CambridgeUniversity Press.
Gardner, H. (1983). Frames of Mind. New York: Basic.Gardner, R. A., Gardner B. T. & van Cantfort, T. E. (ed.)
(1989). Teaching Sign Language to Chimpanzees. Albany,NY: SUNY Press.
Gibson, K. R. (1990). New perspectives on instincts andintelligence: brain size and the emergence ofhierarchical mental construction skills. In “Language”and Intelligence in Monkeys and Apes: ComparativeDevelopmental Perspectives, ed. S. T. Parker & K. R.Gibson, pp. 97–128. New York: Cambridge UniversityPress.
(1993). Beyond neoteny and recapitulation: newapproaches to the evolution of cognitive development.In Tools, Language and Cognition in Human Evolution,ed. K. R. Gibson & T. Ingold, pp. 273–8. Cambridge,UK: Cambridge University Press.
Goodall, J. (1986). The Chimpanzees of Gombe: Patterns ofBehavior. Cambridge, MA: The Belknap Press ofHarvard University Press.
Greenfield, P. M. (1991). Language, tools and brain: theontogeny and phylogeny of hierarchically organized
sequential behavior. Behavioral and Brain Sciences, 14,531–95.
Hare, B., Call, J., Agnetta, B. & Tomasello, M. (2000).Chimpanzees know what conspecifics do and do not see.Animal Behaviour, 59, 771–85.
Hare, B., Call, J. & Tomasello, M. (2001). Do chimpanzeesknow what conspecifics know? Animal Behaviour, 61,139–51.
Harrison, K. E. (1996). Skills used in food processing byvervet monkeys, Cercopithecus aethiops. Unpublisheddoctoral dissertation, University of St. Andrews, St.Andrews, Scotland.
Herrnstein, R. J. (1990). Levels of stimulus control: afunctional approach. Cognition, 37(1–2), 133–66.
Hirata, S. & Matsuzawa, T. (2000). Naive chimpanzees’(Pan troglodytes) observation of experienced conspecificsin a tool task. Journal of Comparative Psychology, 114(3),291–6.
Hirata, S. & Matsuzawa, T. (2001). Tactics to obtain a hiddenfood item in chimpanzee pairs (Pan troglodytes). AnimalCognition, 4, 285–95.
Hostetter, A. B., Cantera, M. & Hopkins, W. D. (2001).Differential use of vocal and gestural communicationby chimpanzees (Pan troglodytes) in response to theattentional state of a human (Homo sapiens). Journal ofComparative Psychology, 115(4), 337–43.
Ingmanson, E. (1996). Tool-using behavior in wild Panpaniscus: social and ecological considerations. InReaching into Thought: The Minds of the Great Apes, ed.A. E. Russon, K. A. Bard & S. T. Parker, pp. 190–210.Cambridge, UK: Cambridge University Press.
Inoue-Nakamura, N. & Matsuzawa, T. (1997). Developmentof stone tool use by wild chimpanzees (Pan troglodytes).Journal of Comparative Psychology, 111, 159–73
Itakura, S. & Tanaka, M. (1998). Use of experimenter-givencues during object-choice tasks by chimpanzees (Pantroglodytes), an orangutan (Pongo pygmaeus), and humaninfants (Homo sapiens). Journal of ComparativePsychology, 112, 119–26.
Jensvold, M. L. A. & Gardner, R. A. (2000). Interactive useof sign language by cross-fostered chimpanzees (Pantroglodytes). Journal of Comparative Psychology, 114(4),335–46.
Johnson-Pynn, J. & Fragaszy, D. M. (2001). Do apes andmonkeys rely upon conceptual reversibility? A reviewof studies using seriated nesting cups in children andnonhuman primates. Animal Cognition, 4,315–24.

Great ape cognitive systems 97
Johnson-Pynn, J., Fragaszy, D. M., Hirsch, E. M., Brakke,K. E. & Greenfield, P. M. (1999). Strategies used tocombine seriated cups by chimpanzees (Pan troglodytes),bonobos (Pan paniscus), and capuchins (Cebus apella).Journal of Comparative Psychology, 113(2),137–48.
Karmiloff-Smith, A. (1992). Beyond Modularity: ADevelopmental Perspective on Cognitive Science.Cambridge, MA: MIT Press.
Kawai, N. & Matsuzawa, T. (2000). Numerical memory spanin a chimpanzee. Nature, 403, 39–40.
King, B. J. (1994). The Information Continuum. Santa Fe,NM: School of American Research.
Kuhlmeier, V. A. & Boysen, S. T. (2001). The effect ofresponse contingencies on scale model task performanceby chimpanzees (Pan troglodytes). Journal ofComparative Psychology, 115(3), 300–6.
Kuhlmeier, V. A., Boysen, S. T. & Mukobi, K. L. (1999).Scale-model comprehension by chimpanzees (Pantroglodytes). Journal of Comparative Psychology, 113(4),396–402.
Langer, J. (1994). From acting to understanding: thecomparative development of meaning. In The Natureand Ontogenesis of Meaning, ed. W. F. Overton & D.Palermo, pp. 191–214. Hillsdale, NJ: Erlbaum.
(1996). Heterochrony and the evolution of primatecognitive development. In Reaching into Thought: TheMinds of the Great Apes, ed. A. E. Russon, K. A. Bard &S. T. Parker, pp. 257–77. Cambridge, UK: CambridgeUniversity Press.
(1998). Phylogenetic and ontogenetic origins of cognition:Classification. In Piaget, Evolution, and Development,ed. J. Langer & M. Killen, pp. 33–54. Mahwah, NJ:Lawrence Erlbaum Associates.
(2000). The descent of cognitive development (withpeer commentaries). Developmental Science, 3(4),361–88.
Leavens, D. & Hopkins, W. (1998). Intentionalcommunication by chimpanzees: a cross-sectional studyof the use of referential gestures. DevelopmentalPsychology, 34(5), 813–22.
Leavens, D. A. & Hopkins, W. D. (1999). The whole-handpoint: the structure and function of pointing from acomparative perspective. Journal of ComparativePsychology, 113(4), 417–25.
Leslie, A. M. (1987). Pretense and representation in infancy:the origins of “theory of mind.” Psychological Review,94, 412–26.
(1994). Pretending and believing: issues in the theory ofToMM. Cognition, 50, 211–38.
Maple, T. L. (1980). Orang-utan Behavior. New York: VanNostrand Reinhold.
Matsuzawa, T. (1991). Nesting cups and metatools inchimpanzees. Behavioral and Brain Sciences, 14(4),570–1.
(1996). Chimpanzee intelligence in nature and in captivity:isomorphism in symbol use and tool use. In Great ApeSocieties, ed. W. C. McGrew, L. F. Marchant & T.Nishida, pp. 196–209. Cambridge, UK: CambridgeUniversity Press.
(ed.) (2001a). Primate Origins of Human Cognition andBehavior. Tokyo: Springer-Verlag.
(2001b). Primate foundations of human intelligence: a viewof tool use in nonhuman primates and fossil hominids.In Primate Origins of Human Cognition and Behavior,ed. T. Matsuzawa, pp. 3–25. Tokyo: Springer-Verlag.
Matsuzawa, T. & Yamakoshi, G. (1996). Comparison ofchimpanzee material culture between Bossou andNimba, West Africa. In Reaching into Thought: TheMinds of the Great Apes, ed. A. E. Russon, K. A. Bard &S. T. Parker, pp. 211–32. Cambridge, UK: CambridgeUniversity Press.
Matsuzawa, T., Biro, D., Humle, T., Inoue-Nakamura, N.,Tonooka, R. & Yamakoshi, G. (2001). Emergence ofculture in wild chimpanzees: education bymaster-apprenticeship. In Primate Origins of HumanCognition and Behavior, ed. T. Matsuzawa, pp. 557–74.Tokyo: Springer-Verlag.
McGrew, W. C. (1998). Culture in nonhuman primates?Annual Review of Anthropology, 27, 301–28.
Menzel, C. R. (1999). Unprompted recall and reporting ofhidden objects by a chimpanzee (Pan troglodytes) afterextended delays. Journal of Comparative Psychology,113(4), 426–34.
Menzel, C. R., Savage-Rumbaugh, E. S. & Menzel, E. W.Jr. (2002). Bonobo (Pan paniscus) spatial memory andcommunication in a 20-hectare forest. InternationalJournal of Primatology, 23(3), 601–19.
Miles, H. L., Mitchell, R. W. & Harper, S. (1996). Simonsays: the development of imitation in an enculturatedorangutan. In Reaching into Thought: The Minds of theGreat Apes, ed. A. E. Russon, S. T. Parker & K. A. Bard,pp. 278–99. Cambridge, UK: Cambridge UniversityPress.
Mitani, J. C. & Watts, D. P. (2001). Why do chimpanzeeshunt and share meat? Animal Behaviour, 61, 1–10.

98 A. E. RUSSON
Mitani, J. C. Watts, D. P. & Muller, M. N. (2002). Recentdevelopments in the study of wild chimpanzee behavior.Evolutionary Anthropology, 11, 9–25.
Mitchell, R. W. (ed.) (2002). Pretending and Imagination inAnimals and Children. Cambridge, UK: CambridgeUniversity Press.
Mithen, S. (1996). The Prehistory of the Mind: The CognitiveOrigins of Art and Science. London: Thames andHudson.
Myowa-Yamakoshi, M. & Matsuzawa, T. (1999). Factorsinfluencing imitation of manipulatory actions inchimpanzees (Pan troglodytes). Journal of ComparativePsychology, 113(2), 128–36.
(2000). Imitation of intentional manipulatory actions bychimpanzees (Pan troglodytes). Journal of ComparativePsychology, 114(4), 381–91.
Noe, R., de Waal, F. B. M. & van Hooff, J. A. R. A. M.(1980). Types of dominance in a chimpanzee colony.Folia Primatologica, 34, 90–110.
Oden, D. L., Thompson, R. K. R. & Premack, D.(1990). Infant chimpanzees (Pan troglodytes)spontaneously perceive both concrete and abstractsame/different relations. Child Development, 61, 621–31.
Parker, S. T. (1996). Apprenticeship in tool-mediatedextractive foraging: imitation, teaching andself-awareness in great apes. In Reaching into Thought:The Minds of the Great Apes, ed. A. E. Russon, K. A.Bard & S. T. Parker, pp. 348–70. Cambridge, UK:Cambridge University Press.
(1998). Review of Primate Cognition. Quarterly Review ofBiology, 73, 540–1.
Parker, S. T. & McKinney, M. (1999). Origins of Intelligence:The Evolution of Cognitive Development in Monkeys,Apes, and Humans. Baltimore, MD: The Johns HopkinsPress.
Parker, S. T., Mitchell, R. W. & Miles, H. L. (ed.) (1999).The Mentalities of Gorillas and Orangutans: ComparativePerspectives. Cambridge, UK: Cambridge UniversityPress.
Pascual-Leone, J. (1987). Organismic processes forneo-Piagetian theories: a dialectical causal accountof cognitive development. International Journal ofPsychology, 22, 531–70.
Pascual-Leone, J. & Johnson, J. (1999). A dialecticalconstructivist view of representation: role of mentalattention, executives, and symbols. In Development ofMental Representation: Theories and Applications, ed.I. E. Sigel, pp. 169–200. Mahwah, NJ: Erlbaum.
Patterson, F. G. & Linden, H. (1981). The Education of Koko.New York: Holt, Rinehart and Winston.
Peirce, C. S. (1932/1960). The icon, index, and symbol. InCollected Papers of Charles Sanders Peirce, Vol. II, ed.C. Hartshorne & P. Weiss, pp. 156–73. Cambridge, MA:Harvard University Press.
Peignot, R. & Anderson, J. R. (1999). Use ofexperimenter-given manual and facial cues by gorillas(Gorilla gorilla) in an object-choice task. Journal ofComparative Psychology, 113(3), 253–60.
Perner, J. (1991). Understanding the Representational Mind.Cambridge, MA: Bradford/MIT.
Piaget, J. (1952). The Origins of Intelligence in Children. NewYork: International Universities Press.
(1954). The Construction of Reality in the Child. New York:Basic Books.
Poti’, P., Langer, J., Savage-Rumbaugh, S. & Brakke, K. E.(1999). Spontaneous logico-mathematical constructionsby chimpanzees (Pan troglodytes, P. paniscus). AnimalCognition, 2, 147–56.
Povinelli, D. J. (2000). Folk Physics for Apes. New York:Oxford University Press.
Premack, D. (1983). The codes of man and beast. Behavioraland Brain Sciences, 6, 125–37.
(1988). “Does the chimpanzee have a theory of mind?”revisited. In Machiavellian Intelligence: Social Expertiseand the Evolution of Intellect in Monkeys, Apes andHumans, ed. R. W. Byrne & A. Whiten, pp. 160–79.Oxford: Clarendon Press.
Roberts, W. A. & Mazmanian, D. S. (1988). Concept learningat different levels of abstraction by pigeons, monkeys,and people. Animal Behavior Processes, 14, 247–60.
Rogoff, B. (1992). Apprenticeship in Thinking. New York:Oxford University Press.
Rumbaugh, D. M. (1970). Learning skills in anthropoids. InPrimate Behavior: Developments in Field and Laboratory,ed. L. A. Rosenblum, pp. 1–70. New York: AcademicPress.
(2000). Review of The Mentalities of Gorillas andOrangutans. International Journal of Primatology, 21(4),769–71.
Rumbaugh, D. M. & Pate, J. L. (1984). Primates’ learning bylevels. In Behavioral Evolution and Integrative Levels, ed.G. Greenberg & E. Tobach, pp. 221–40. Hillsdale, NJ:Erlbaum.
Russon, A. E. (1996). Imitation in everyday use: matchingand rehearsal in the spontaneous imitation ofrehabilitant orangutans (Pongo pygmaeus). In Reaching

Great ape cognitive systems 99
into Thought: The Minds of the Great Apes, ed. A. E.Russon, K. A. Bard & S. T. Parker, pp. 152–76.Cambridge, UK: Cambridge University Press.
(1998). The nature and evolution of orangutanintelligence. Primates, 39, 485–503.
(1999a). Orangutans’ imitation of tool use: a cognitiveinterpretation. In The Mentalities of Gorillas andOrangutans: Comparative Perspectives, ed. S. T. Parker,R. W. Mitchell & H. L. Miles, pp. 117–46. Cambridge,UK: Cambridge University Press.
(1999b). Naturalistic approaches to orangutan intelligenceand the question of enculturation. International Journalof Comparative Psychology, 12(4), 181–202.
(2002a). Return of the native: cognition and site specificexpertise in orangutan rehabilitation. InternationalJournal of Primatology, 23(3), 461–78.
(2002b). Pretending in free-ranging rehabilitantorangutans. In Pretending and Imagination in Animalsand Children, ed. R. W. Mitchell, pp. 229–40.Cambridge, UK: Cambridge University Press.
(2003a). Comparative developmental perspectives onculture: the great apes. In Between Biology and Culture:Perspectives on Ontogenetic Development, ed. H. Keller,Y. Poortinga & A. Schoelmerich, pp. 30–56. Cambridge,UK: Cambridge University Press.
(2003b). Developmental perspectives on great apetraditions. In Towards a Biology of Traditions:Models and Evidence, ed. D. Fragaszy & S. Perry,pp. 187–212. Cambridge, UK: Cambridge UniversityPress.
Russon, A. E., Bard, K. A. & Parker, S. T. (ed.) (1996).Reaching into Thought: The Minds of the Great Apes.Cambridge, UK: Cambridge University Press.
Russon, A.E., Mitchell, R.W., Lefebvre, L. & Abravanel, E.(1998). The comparative evolution of imitation. InPiaget, Evolution, and Development, ed. J. Langer & M.Killen, pp. 103–43. Hillsdale, NJ: Lawrence ErlbaumAssociates.
Savage-Rumbaugh, E. S., Williams, S. L., Furuichi, T. &Kano, T. (1996). Language perceived: paniscus branchesout. In Great Ape Societies, ed. W. C. McGrew, L. F.Marchant & T. Nishida, pp. 173–84. Cambridge, UK:Cambridge University Press.
Savage-Rumbaugh, E. S., Rumbaugh, D. M., Smith, S. T. &Lawson, J. (1980). Reference: the linguistic essential.Science, 210, 922–5.
Schaller, G. B. (1963). The Mountain Gorilla: Ecologyand Behavior. Chicago, IL: University of Chicago Press.
Shumaker, R. W., Palkovich, A. M., Beck, B. B., Guagnano,G. A. & Morowitz, H. (2001). Spontaneous use ofmagnitude discrimination and ordination by theorangutan (Pongo pygmaeus). Journal of ComparativePsychology, 115(4), 385–91.
Sousa, C. & Matsuzawa, T. (2001). The use of tokens asrewards and tools by chimpanzees (Pan troglodytes).Animal Cognition, 4, 213–21.
Spearman, C. (1927). The Abilities of Man: Their Nature andMeasurement. New York: Macmillan.
Spinozzi, G. & Langer, J. (1999). Spontaneous classificationin action by a human-enculturated and language-rearedbonobo (Pan paniscus) and common chimpanzees (Pantroglodytes). Journal of Comparative Psychology, 113(3),286–96.
Spinozzi, G., Natale, F., Langer, J. & Brakke, K. E. (1999).Spontaneous class grouping behavior by bonobos (Panpaniscus) and common chimpanzees (P. troglodytes).Animal Cognition, 2, 157–70.
Stoinski, T. S., Wrate, J. L., Ure, N. & Whiten, A. (2001).Imitative learning by captive western lowland gorillas(Gorilla gorilla gorilla) in a simulated food processingtask. Journal of Comparative Psychology, 115(3),272–81.
Stokes, E. J. (1999). Feeding Skills and the Effect of Injuryon Wild Chimpanzees. Unpublished PhD dissertation,University of St. Andrews, St. Andrews, Scotland.
Stokes, E. J. & Byrne, R. W. (2001). Cognitive abilities forbehavioural flexibility in wild chimpanzees (Pantroglodytes): the effect of snare injury on complexmanual food processing. Animal Cognition, 4, 11–28.
Suddendorf, T. & Whiten, A. (2001). Mental evolution anddevelopment: evidence for secondary representation inchildren, great apes, and other animals. PsychologicalBulletin, 127(5), 629–50.
Sugiyama, Y. (1997). Social tradition and the use oftool-composites by wild chimpanzees. EvolutionaryAnthropology, 6, 23–27.
Swartz, K. B., Sarauw, D. & Evans, S. (1999). Comparativeaspects of mirror self-recognition in great apes. In TheMentalities of Gorillas and Orangutans: ComparativePerspectives, ed. S. T. Parker, R. W. Mitchell & H. L.Miles, pp. 283–294. Cambridge, UK: CambridgeUniversity Press.
Tanaka, M. (1995). Object sorting in chimpanzees (Pantroglodytes): classification based on physical identity,complementarity, and familiarity. Journal ofComparative Psychology, 109, 151–61.

100 A. E. RUSSON
(2001). Discrimination and categorization of photographsof natural objects by chimpanzees (Pan troglodytes).Animal Cognition, 4, 201–11.
Thompson, R. K. R. & Oden, D. L. (2000). Categoricalperception and conceptual judgments by nonhumanprimates: the paleological monkey and the analogicalape. Cognitive Science, 24(3), 363–96.
Thompson, R. K. R., Oden, D. L. & Boysen, S. T.(1997). Language-naive chimpanzees (Pan troglodytes)judge relations between relations in a conceptualmatch-to-sample task. Journal of ExperimentalPsychology: Animal Behavior Processes, 23, 31–43.
Tomasello, M. (1999). The Cultural Origins of HumanCognition. Cambridge, MA: Harvard UniversityPress.
Tomasello, M. & Call, J. (1997). Primate Cognition. NewYork: Oxford University Press.
Tomasello, M., Call, J. & Hare, B. (1998). Five primatespecies follow the visual gaze of conspecifics. AnimalBehaviour, 55, 1063–9.
Tomasello, M., Hare, B. & Agnetta, B. (1999). Chimpanzees,Pan troglodytes, follow gaze geometrically. AnimalBehaviour, 58, 769–77.
Tomasello, M., Hare, B. & Fogleman, T. (2001). Theontogeny of gaze following in chimpanzees, Pantroglodytes, and rhesus macaques, Macaca mulatta.Animal Behaviour, 61, 335–43.
Tomonaga, M. & Matsuzawa, T. (2000). Sequentialresponding to Arabic numerals with wild cards by thechimpanzee (Pan troglodytes). Animal Cognition, 3,1–11.
Tooby, J. & Cosmides, L. (1992). The psychologicalfoundations of culture. In The Adapted Mind, ed. J. H.Barkow, L. Cosmides & J. Tooby, pp. 19–136. NewYork: Oxford University Press.
van Schaik, C. P., Ancrenaz, M., Borgen, G., Galdikas,B., Knott, C. D., Singleton, I., Suzuki, A., Utami,S. & Merrill, M. (2003). Orangutan cultures andthe evolution of material culture. Science, 299,102–5.
van Schaik, C. P., Deaner, R. O. & Merrill, M. Y. (1999). Theconditions for tool use in primates: implications for theevolution of material culture. Journal of HumanEvolution, 36, 719–41.
Visalberghi, E. & Limongelli, L. (1996). Acting andunderstanding: tool use revisited through the minds ofcapuchin monkeys. In Reaching into Thought: The Mindsof the Great Apes, ed. A. E. Russon, K. A. Bard & S. T.Parker, pp. 57–79. Cambridge, UK: CambridgeUniversity Press.
Whiten, A. (1996). Imitation, pretence, and mindreading:secondary representation in comparative primatologyand developmental psychology? In Reaching intoThought: The Minds of the Great Apes, ed. A. E. Russon,K. A. Bard & S. T. Parker, pp. 300–24. Cambridge, UK:Cambridge University Press.
(1998a). Evolutionary and developmental origins of themindreading system. In Piaget, Evolution, andDevelopment, ed. J. Langer & M. Killen, pp. 73–99.Hillsdale, NJ: Lawrence Erlbaum Associates.
(1998b). Imitation of the sequential structure of actions bychimpanzees (Pan troglodytes). Journal of ComparativePsychology, 112, 270–81.
(2000). Chimpanzee cognition and the question of mentalre-representation. In Metarepresentation: AMultidisciplinary Perspective, Vancouver Studies inCognitive Science, ed. D. Sperber, pp. 139–67. Oxford:Oxford University Press.
Whiten, A. & Byrne, R. W. (1991). The emergence ofmetarepresentation in human ontogeny and primatephylogeny. In Natural Theories of Mind, ed. A. Whiten,pp. 267–82. Oxford: Basil Blackwell.
Whiten, A., Goodall, J., McGrew, W. C., Nishida, T.,Reynolds, V., Sugiyama, Y., Tutin, C. E. G., Wrangham,R. W. & Boesch, C. (1999). Culture in chimpanzees.Nature, 399, 682–5.
Yamakoshi, G. & Sugiyama, Y. (1995). Pestle-poundingbehaviour of wild chimpanzees at Bossou, Guinea: anewly observed tool-using behaviour. Primates, 36(4),489–500.





























