Embed Size (px)
Citation preview

Plant Physiol. (1992) 100, 1994-20010032-0889/92/100/1 994/08/$01 .00
Received for publication March 26, 1992Accepted August 5, 1992
Gibberellins and the Legume-Rhizobium Symbiosis'
Ill. Quantification of Gibberellins from Stems and Nodules of Lima Bean and Cowpea
Raymond C. Dobert2, Stewart B. Rood, Karen Zanewich, and Dale G. Blevins*Interdisciplinary Plant Group, Agronomy Department, University of Missouri, Columbia, Missouri 6521 1
(R.C.D., D.G.B.); and Department of Biology, The University of Lethbridge, Lethbridge, Alberta, Canada TI K 3M4(S.B.R., K.Z.)
ABSTRACT
Lima bean (Phaseolus lunatus L.) plants inoculated with Brady-rhizobium sp. strain 127E14 displayed a period of marked inter-node elongation that was not observed in plants inoculated withother compatible bradyrhizobia, including strain 127E15. Whenstrain 127E14 nodulated an alternate host, cowpea (Vigna ungui-culata L. Walp), a similar, although less dramatic growth responseinduced by the bacteria was observed. It has been speculated thatthe elongative growth promotion brought about by inoculationwith strain 127E14 is mediated by gibberellins (GAs). Using deu-terated internal standards and gas chromatography-mass spectros-copy analysis, we have quantified the levels of GA,, GA20, GA19,and GA44 in nodules and stems of two varieties of lima bean (bushand pole) and one variety of cowpea that were inoculated witheither strain 127E14 or 127E15. In nodules formed by strain 127E14on lima bean, endogenous levels of GA20 and GA1, were 10 to 40times higher (35-88 ng/g dry weight) than amounts found innodules formed by strain 127E15 (2.2-3.9 ng/g dry weight). Rela-tive amounts of GA44 were also higher (4- to 11-fold) in 127E14nodules, but this increase was less pronounced. The rhizobial-induced increase of these GAs in the nodule occurred in both poleand bush varieties and seemed to be independent of host mor-phology. Regardless of rhizobial inoculum, levels of the "bioactive"GA1 in the nodule (0.3-1.1 ng/g dry weight) were similar. Incowpea nodules, a similar, although smaller, difference in GAcontent due to rhizobial strain was observed. The concentration ofGA1 in lima bean stems was generally higher than that observed inthe nodule, whereas concentrations of the other GAs measuredwere lower. In contrast with the nodule, GA concentrations in limabean stems were not greater in plants inoculated with strain127E14, and in some cases the slower growing plants inoculatedwith strain 127E15 actually had higher levels of GA20, GA19, andGA". Thus, there were major differences in concentrations of theprecursors to GA1 in nodules formed by the two bacterial strains,which were positively correlated with the observed elongationgrowth. These results support the hypothesis that the rhizobial
'This work was supported through National Sciences and Engi-neering Council of Canada grant GP 38682 to S.B.R. and a grantfrom the Missouri Fertilizer & Lime Council to D.G.B. R.C.D. wassupported by Food for the 21st Century, and Interdisciplinary PlantGroup through the U.S. Department of Agriculture. National NeedsFellowship program. Contribution No. 11,647 to Missouri Agricul-tural Experiment Station Journal Series.
2 Present address: Department of Agronomy, 1575 Linden Drive,University of Wisconsin-Madison, Madison, WI 53706.
1994
strain modifies the endogenous GA status of the symbiotic system.This alteration in GA balance within the plant, presumably, under-lies the observed growth response.
The process of nodule initiation on roots of leguminousplants by the soil bacterium Bradyrhizobium has been widelystudied. Although a signal-exchange mechanism utilized dur-ing the early stages of nodule development has been identi-fied (21), little is known regarding the means by which thesetwo distinct organisms communicate with each other oncethe symbiosis is established. Phytohormones have long beenconsidered potential mediators in the process of nodule ini-tiation (14, 24-26, 31). However, the possibility that phyto-hormones are utilized by either partner to control and main-tain the symbiotic association once it is formed remainsspeculative.
GA3-like substances are present in nodules formed on arange of leguminous (1, 4, 7, 9, 26, 35) and nonleguminous(16) hosts. In numerous bioassay systems, which vary in theirsensitivity to different GAs, levels of GA-like substances innodule tissue have been found to range from 9.8 ng/g freshweight in Phaseolus vulgaris (26) to extraordinarily high levelsof 1000 ng/g fresh weight in Phaseolus aureus (4) and 3340ng/g fresh weight in Lupinus (7). Generally, the concentrationof GA-like substances has been found to be higher in nodulesthan in nearby root tissue (7, 26, 35), and, in one instance,differences in the spectrum of GA-like substances were de-tected between root and nodule tissue (35). This has led tothe conclusion that the rhizobial microsymbiont can contrib-ute to the quantity and perhaps variety of GAs present in thenodule.
Conversely, Atzom et al. (1), using immunoassay tech-niques to characterize and quantify GA-like substances in P.vulgaris nodules and roots, found that the presumed GAcontent and composition of nodules were similar to that ofuninoculated root tissue. However, the root-to-root nodulecomparison made in this study (1) may not be equivalent to
3Abbreviations: GA., gibberellin A.; amu, atomic mass unit; DAP,days after planting; EtOAc, ethyl acetate; MeOH, methanol; SiO2,silica gel; SIM, selected ion monitoring.
www.plantphysiol.orgon July 15, 2018 - Published by Downloaded from Copyright © 1992 American Society of Plant Biologists. All rights reserved.

GIBBERELLIN QUANTIFICATION IN LIMA BEAN-RHIZOBIUM SYMBIOSIS
those in previous reports (6, 35) where roots from nodulatedplants were analyzed. The noninoculated roots that Atzornet al. (1) analyzed were grown on ammonium nitrate, and ithas been speculated that nitrogen nutrition (both type andamount) can have a profound effect on the status of GAswithin the plant (20). Thus, GAs in root tissue of plantsgrown on a combined N source might be expected to differfrom roots dependent on symbiotically fixed nitrogen. Thefinding that free-living Rhizobium phaseoli cultures produceGA,, GA20, and GA4, the same compounds detected in rootand nodule tissue, led Atzom et al. (1) to conclude that thebacteria did not elicit the production of additional types ofGA within the nodule. In addition to the definitive GC-MSidentifications of GAs in R. phaseoli cultures (1), other re-searchers have demonstrated that free-living rhizobia (in-cluding members of both the Rhizobium [fast-growing] andBradyrhizobium [slow-growing] genera) have the capacity toproduce GA-like substances (18, 22, 35). This evidence sup-ports the hypothesis that at least some rhizobial strainspossess the genetic and enzymic capacity to synthesize GAs.
It was previously shown (9) that nodules formed on Hen-derson lima bean (Phaseolus lunatus L.) by Bradyrhizobium sp.strain 127E14 contain relatively high levels (500 ng/g freshweight) of GA-like substances relative to similar nodulesformed by strain 127E15 (38 ng/g fresh weight) or to nearbyroot tissue (<2.2 ng/g fresh weight). Thus, in contrast to thefindings of Atzom et al. (1), it appears that, in the lima beansymbiosis, the rhizobial strain can influence the GA contentof the nodule. Plants inoculated with strain 127E14 alsoexhibited a marked period of rapid internode elongation thatwas not observed in plants inoculated with strain 127E15 (9,33). Similarly, stem and petiole length of cowpea (Vignaunguiculata L. Walp) was greater when plants were inoculatedwith strain 127E14 (6), although the magnitude of growthpromotion was not as large as in lima bean. The rhizobium-induced elongation response could also be mimicked byexogenous GA3 application and suppressed by treating plantswith inhibitors of GA biosynthesis (6, 9). This preliminarywork supports the basic hypothesis that the root nodule cancontribute GAs to the host plant.We have identified GA,, GA3, GA19, GA20, GA29, and GA44
from root nodules formed on lima bean by Bradyrhizobiumsp. strain 127E14 (5), and GA8 and these same GAs wereidentified in cowpea nodules (6). GA,, GA3, GA19, GA20, andGA44 have also been identified in lima bean stem tissue (5),and, in addition to these compounds, GA29 was detected incowpea stems (6).The unequivocal identification of these GAs, whose pres-
ence is indicative of the early 13-hydroxylation biosyntheticpathway, provides the background needed to assess the roleof rhizobia in determining the GA status of both the noduleand the host plant. Using deuterated intemal standards andGC-SIM analysis, we now quantify the levels of GA1, GA19,GA20, and GA44 in nodule and stem tissue from a series ofsymbiotic pairings. By utilizing two distinct rhizobial strainsdiffering in their capacity to promote the accumulation ofGA-like substances within the nodule and elicit stem elon-gation, we investigated the possibility that the bacterial mi-crosymbiont can alter the amounts of specific GAs within thenodule and, consequently, host morphology.
MATERIALS AND METHODS
Plant Materials
Lima bean (Phaseolus lunatus L.) seeds (cv Henderson orcv King of the Garden, Burpee Seed Co., Warminster, PA)were treated with a 10% (v/v) ethanol solution for 3 min,rinsed with deionized water, and germinated for 5 d on papertowels. Cowpea (Vigna unguiculata L. Walp) seeds (cv PurpleHull) were similarly treated with ethanol and germinated for4 d. Five seeds, with radicles approximately 3 cm in length,were planted in 20-cm pots filled with a 50:50 (v/v) per-lite:sand mix. Sand was disinfected with steam for 4 to 6 hprior to use. At the time of planting, each seedling was dipinoculated in an aqueous suspension containing Bradyrhizo-bium sp. strains 127E14 or 127E15 (Nitragin, Milwaukee, WI)that were derived from cultures maintained on solid yeast-mannitol medium (34). Plants received a standard minus-Nnutrient solution (5) via a wick connecting the top pot withthe lower nutrient reservoir. Plants were grown in a green-house during the months of September and October withoutsupplemental lighting (photoperiod approximately 12 h).After 3 to 5 weeks of growth, stem tissue above the cotyle-donary node and root nodules were harvested from eachplant. Total height of Henderson lima beans was also meas-ured at the time of harvest.
Extraction of GAs
Nodule and stem tissue were frozen in liquid N2 at thetime of harvest and freeze-dried prior to analysis. Extractionof tissue followed a procedure similar to that outlined earlier(5). Triplicate samples of freeze-dried tissue (0.5 g dry weight/replicate) were homogenized in cold 80% MeOH in a blenderfor stems or in a tissue homogenizer for nodules. Deuteratedinternal standards (2 ng of [2H2]GA1, 10 ng of [2H2]GA19, and10 ng of [2H2]GA20), provided by Professors L. Mander (Aus-tralian National University, Australia) and N. Murofushi(University of Tokyo, Japan) were added to each sample atthis time. Particulate debris was removed from nodule ho-mogenates by centrifuging samples at 3000g for 15 min. Theresulting nodule supernatant and stem homogenates werevacuum filtered, and 0.1 M Na2HPO4 buffer, pH 8.0 (1 mL/3 mL filtrate), was added. MeOH was removed from samplesin vacuo to yield an aqueous sample. The pH of stem extractswas adjusted to pH 9.0 and partitioned twice against one-third volume of water-saturated diethyl ether to removepigments. Insoluble polyvinylpolypyrrolidone was added tonodule and stem aqueous fractions, and the resulting slurrywas vacuum filtered as described (12). Samples were acidified(pH 2.8) and partitioned three times against water-saturatedEtOAc (one-third aqueous volume) to extract the free (non-conjugated) GAs. Water in the acidic EtOAc fraction wasremoved by freezing followed by filtration of the sample toremove the ice. The EtOAc was removed in vacuo, and theresulting residue was dissolved in MeOH:EtOAc (1:1) andfurther purified by SiO2 column chromatography prior toGC-MS analysis or directly analyzed by ion-suppressionHPLC.
1 995
www.plantphysiol.orgon July 15, 2018 - Published by Downloaded from Copyright © 1992 American Society of Plant Biologists. All rights reserved.

Plant Physiol. Vol. 100, 1992
Ion-Suppression Reverse-Phase HPLC Separation
In initial attempts to purify and quantify the endogenousGAs from nodule tissue, a gradient profile that employed anaqueous phosphoric acid/MeOH solvent system (17) wasused. This method provided a high degree of chromato-graphic resolution and allowed for detection of GAs bymonitoring absorbance at 208 nm. The acidic EtOAc residuewas dissolved in MeOH and passed through a prewetted C18cartridge Sep-Pak (Millipore, Bedford, MA). Samples wereredissolved in 100 ,uL 60:40 MeOH:0.01 M H3PO4 and injectedonto a reverse-phase C18 column (,gBondapak C18, 3.9 x 30mm, Waters, Milford, MA) eluted with a gradient of MeOHand 0.01 M H3PO4 (pH 3.0). The gradient profile was asfollows: from 0 to 18 min, isocratic 40% MeOH; from 18 to39 min, linear 40 to 75% MeOH; from 39 to 54 min, linearfrom 75 to 80% MeOH; and a 6-min rinse with 100% MeOHprior to the next injection. Flow rate through the column was2.5 mL/min with fractions collected every minute. Resultingfractions were dried and redissolved in 95% ethanol andaliquots (1/200) were tested for GA-like activity using theTan-ginbozu dwarf rice (Oryza sativa L.) bioassay (23). Trip-licate samples were analyzed with similar results.
SiO2 Partition Column Chromatography
In samples to be further analyzed by GC-MS, the acidicEtOAc residue was dissolved in MeOH:EtOAc (1:1) andloaded onto a stack of glass-fiber filter discs. Prior to sampleapplication, discs were preloaded with [3H]GA standards(approximately 60 KBq each of [3H]GA1 [1.2 TBq/mmol] and[3H]GA4 [approximately 35 GBq/mmol], Amersham, Arling-ton Heights, IL) to allow for appropriate groupings of frac-tions resulting from subsequent chromatographic separations.Glass fiber discs were placed on a SiO2 column bed (1.0 g ofWoelm 32-100, Universal Scientific, Atlanta, GA) and stepeluted with solvents varying in their proportion of formicacid-saturated hexane and EtOAc (5, 8). Aliquots of theresulting fractions were assayed for radioactivity. A singlefraction for further analysis was produced by grouping allSi02 fractions between, and inclusive of, the peaks of labeledGA4 and GA,.
Reverse-Phase HPLC Separation
The grouped SiO2 fractions were dissolved in MeOH,passed through a C18 cartridge Sep-Pak, and then filtered(Sep-Pak and 0.45 ,um Millex-HV, Millipore). Samples wereredissolved in 80% MeOH and subjected to reverse-phaseHPLC separation on a ,uBondapak C18 column (3.9 x 30 mm,Waters) eluted with an anhydrous MeOH:1% (v/v) aqueousacetic acid gradient as described (5, 19). With flow throughthe column at 1 mL/min, 42 2-mL fractions were collectedand aliquots were assayed for radioactivity. The retentiontimes of [3H]GA1 and [3H]GA4 were used to group fractionsthat would contain GAI/3 (fractions 21-24), GA20 (fractions25-28), and GA19/44 (fractions 29-33). Dried samples weregrouped and levels of these GAs were determined byGC-SIM.
GC-MS Analysis and Endogenous GA Quantification
Grouped HPLC fractions were methylated with etherealdiazomethane, and trimethylsilyl derivatives were producedby incubating samples for 30 min at 600C in 50:50 (v/v)bistrimethyl-silyltrifluoroacetamide with 1% trimethyl chlo-rosilane and pyridine (5, 27). Derivatized samples were in-jected onto a 15-m DB-5 microcapillary column (&W Scien-tific Inc., Folson, CA) in a Hewlett-Packard 5890 Series II gaschromatograph and run as previously described (5). The GCwas coupled to a Hewlett-Packard 5970A mass selectivedetector operating at 1800 electron volts (27). The identity ofeluted GAs, possessing an appropriate retention time, wasverified by monitoring the abundance of two or three diag-nostic ions of both endogenous and deuterated GAs. Levelsof endogenous GAs were determined by measuring the abun-dance of ion pairs 506/508 amu for GA1, 434/436 amu forGA19, and 418/420 amu for GA20. The ratio of integratedpeak areas between protonated (endogenous 1H) and deuter-ated (2H) GAs, numerically corrected for mutual contributionfrom each other, was used to calculate the amount of endog-enous GA (David Pearce, University of Calgary, personalcommunication; 10). The relative amount of endogenousGA44 was determined by comparing the measured intensityof GA44 at 432 amu to the corrected abundance of [2H2]GA19.Greatest accuracy was obtained with this method when theratio of endogenous GA to deuterated standard fell between1:20 and 5:1. With the exception of the GA20 analysis in stemsamples from cowpea and lima bean inoculated with strain127E15, the ratios of endogenous to deuterated GA werewithin this range. Variability within the data for nodulesamples was routinely low, especially in nodule extracts fromlima bean, whereas values for stem samples were slightlymore variable.
RESULTS AND DISCUSSION
In previous studies, a number of GAs characteristic of theearly 13-hydroxylation pathway were identified in both limabean and cowpea stems and nodules (5, 6, 11). In this report,using deuterated internal standards, we have been able toquantify the levels of 'bioactive' GA1 and its presumedprecursors GA20, GA19, and GA44 from stem and nodule tissuefrom several symbiotic pairings.Henderson lima bean and cowpea plants inoculated with
either strain 127E14 or 127E15 were both harvested at twodifferent time points to account for the possibility that levelsof GAs in the stem and nodule might be affected by devel-opmental changes. Harvests of Henderson lima beans weretimed so that, at the time of the first harvest, plant growthwas primarily vegetative, and plant height differences due torhizobial inoculum were small. At the second harvest, mostplants had begun to flower, and differences in plant heightwere significant. The pole lima bean cultivar King of theGarden was harvested at a single time, because this cultivardoes not show a discernible difference in plant height due torhizobial inoculant.
In the experiments of Triplett et al. (33), Henderson limabean plants inoculated with strain 127E14 began to displaythe 'bolting response' between 25 and 35 DAP. However, it
1 996 DOBERT ET AL.
www.plantphysiol.orgon July 15, 2018 - Published by Downloaded from Copyright © 1992 American Society of Plant Biologists. All rights reserved.

GIBBERELLIN QUANTIFICATION IN LIMA BEAN-RHIZOBIUM SYMBIOSIS
has been observed that the timing of this rapid internodeelongation is linked more to a developmental schedule thanto a chronological one. Developmental cues such as the onsetof reproductive growth or release of apical dominance (i.e.lateral branch development), which vary depending on en-vironmental conditions, may be more important in regulatingthis growth response. Nonetheless, even at 19 DAP totalheight was significantly different between plants inoculatedwith strain 127E14 (14.9 cm) and those inoculated with strain127E15 (10.0 cm). At the second harvest (28 DAP), theseheight differences had become even more dramatic (68.4 cmversus 41.4 cm).
Dwarf-Rice Bioassay
GA-like substances from nodule tissue formed on limabean roots by the rhizobial strains 127E14 and 127E15 havepreviously been purified by TLC and analyzed by the lettucehypocotyl bioassay (9). In an attempt to further characterizeand quantify the GAs in this tissue, nodule extracts from 35-d-old lima beans were subjected to ion-suppression reverse-phase HPLC (17), and resultant fractions were bioassayed inthe Tan-ginbozu dwarf rice test. By this technique, severalregions that exhibited strong GA-like activity were detectedin extracts of 127E14 nodules (Fig. 1), substantiating thefindings of Evensen and Blevins (9) that this tissue containedsignificant levels of GA-like substances when compared to127E15 nodules. These regions of bioactivity often coincidedwith absorbance peaks at 208 nm, presumably due to thepresence of GAs. However, co-chromatography with GAstandards (GA3, GA4+7) did not provide conclusive evidenceregarding the identity of these bioactive peaks. Fractionsresulting from this gradient were not suitable for derivatiza-tion due to a phosphate residue in dried samples, and, thus,
8 80
a Eg 60e
c 20
o
275 -
S 225 -
c E1
iCS 175 -
_c _
I 125 -
75
10 20 30 40
BioassayI Response
AI I
0 10 20 30Retention Time (min)
GA-like substances could not be identified. To alleviate thisproblem, samples were eluted with a volatile acid (acetic acid)in all subsequent analyses.
GA Quantification in Lima Bean Nodules
The most accurate and precise means of quantifying GAsis through the use of deuterated internal standards and GC-MS analysis (15). Without the benefit of prior studies detailingthe levels of particular GAs within root nodules, direct com-parisons with our present data cannot be made. In ouranalysis, the range in GA, concentration in nodules was 0.25to 4.7 ng/g dry weight (Figs. 2 and 3). Using radioimmuno-assay to quantify apparent GA, levels in bean nodules,Atzorn et al. (1) reported a concentration of 2.1 ng GA1/g,similar to the values observed in this study. Levels of GA20,GA19, and GA44 in lima bean and cowpea nodules variedgreatly with the rhizobial inhabitant (see below) but were atleast two times greater than earlier radioimmunoassay datafor GA20 in P. vulgaris nodules (1).When the concentrations of GA1, GA20, and GA19 were
summed (excluding GA44, which has not been absolutelyquantified), total GA content ranged from 5 to 132 ng/g dryweight for lima bean nodules, and from 29 to 104 ng/g dryweight for cowpea. On a fresh weight basis, assuming 6 gfresh weight = 1 g dry weight, the "total' GA concentrationranged from <1.0 to 22 ng/g fresh weight. These values arelower than the numerous bioassay-based quantifications ofnodular GA levels (4, 7, 26, 35), including earlier findings inthis same symbiotic system (9). Although the quantities ofGA-like substances reported by Evensen and Blevins (9) wereas much as 23-fold higher than the levels reported here, therelative difference in total GA concentration between 127E14and 127E15 nodules remains the same. The total amount of
1nnt -. ME
80
60
40
20
50
40 50
01 .0 10 20 30 40 50
200Bioassay
175 - Response
150 -
125 -
- - - * ~~~~~~~Control75
0 10 20 30Retention Time (min)
40 50
Figure 1. UV trace and Tan-ginbozu dwarf rice bioassay response of HPLC eluate from purified lima bean nodule extracts formed byBradyrhizobium sp. strain 127E14 or 127E15. Acidic EtOAc extracts of 35-d-old nodules were purified by reverse-phase ion suppressionHPLC, and the GA-like activity in resulting fractions was detected by the Tan-ginbozu dwarf rice bioassay.
127E14 NodulesUV Trace
He
127E15 NodulesUV Trace
inn) . in
-4l
1997
D
www.plantphysiol.orgon July 15, 2018 - Published by Downloaded from Copyright © 1992 American Society of Plant Biologists. All rights reserved.

Plant Physiol. Vol. 100, 1992
Figure 2. Quantification of endogenous GAs innodules and stems of lima bean inoculated withBradyrhizobium sp. strain 127E14 or 127E15.Quantification based on the abundance ratiobetween the major ion of endogenous GA andthe [2H2]GA internal standard as determinedby GC-SIM. GA44 levels are relative to [2H2]-GA,9 abundance. Values represent means (±SE)of three replicate samples harvested at the in-dicated times from either bush (Henderson) orpole (King of the Garden) lima beans. Asterisks(*) indicate data sets where only a single quan-tification was possible, and thus SE values couldnot be calculated. Plants inoculated with127E14 are represented by hatched bars; plantsinoculated with 127E15 are represented byopen bars.
LIMA BEANNODULE
LIMA BEANSTEM
.C.U
GA, GA, 1.3*127E14
1.5- L1127E15 - 1.5
3.1 ~~~~~~1.1 1.0 1.1 .
01 1.0 0.9r 0.7 0.9 1.0 01
cm i E |0.60.5 04 . 0.3 04 0.5
84.8 88.1 GA20 7.7 GA2080 --8
~:60 6a 0
-P ~~~~~~~~~37.101 40 4 01
20 1.2 2
2.2 383.5 0.7 0.9 1. T.0 A
a
Cp
60-
50-
40
30
20
10
0)01
01,a
50 -
40
3 30-
CC 20
10 -
Henderson Henderson KOG Henderson Henderson KOG19 DAP 28 DAP 30 DAP 19 DAP 28 DAP 30 DAP
GA-like substances detected in HPLC fractions of 127E14nodule extracts by the dwarf rice bioassay was 90 ng/g freshweight (Fig. 1), which, although lower than the value re-ported with the lettuce hypocotyl bioassay (9), was still higherthan levels determined by GC-MS.
In all legume hosts examined, the levels of GAs, with theexception of GA,, were much greater in nodule tissue inhab-ited by strain 127E14 than 127E15 nodules (Figs. 2 and 3).However, when comparing levels of GAs from differentsamples, one should consider that physiological responses toexogenously applied phytohormones follow an exponentialscale (32), and thus small differences in concentrationbrought about by variability in the extraction and recoveryefficiencies are probably biologically insignificant. In in-stances where concentration differences are less than 2-fold,a high degree of statistical significance should be observedbefore differences are considered to be of physiological im-portance.
In Henderson lima beans harvested at either 19 or 28 DAP,
levels of GA20 were elevated 20- to nearly 40-fold in 127E14nodules compared to 127E15 nodules. Similar, although lessdramatic differences were observed in the levels of GA,9 andGA44. The accumulation of these GAs was in sharp contrastto the observed levels of GA,, which were very similar inlima bean nodules inoculated with these two rhizobial strains(Fig. 2). The similarity in nodule GA, levels, coupled with anapparent accumulation of its immediate precursors in 127E14nodules, suggests that metabolism in these nodules may berestricted at the 3f-hydroxylation step converting GA20 toGA,. Alternatively, GA, precursor accumulation in 127E14nodules could be explained by an accelerated synthesis ofthe GA intermediates preceding those measured, without aconcomitant increase in the conversion of GA20 to GA,.
In this study, the origin of the GAs within the nodule hasnot been determined. However, the concentration of thesephytohormones is likely to be determined by the relativerates of GA import to and export from the nodule as well assynthesis and degradation of GAs within the nodule. In initial
2-0 I I
100 I
1 998 DOBERT ET AL.
2.0
10
www.plantphysiol.orgon July 15, 2018 - Published by Downloaded from Copyright © 1992 American Society of Plant Biologists. All rights reserved.
GIBBERELLIN QUANTIFICATION IN LIMA BEAN-RHIZOBIUM SYMBIOSIS
host plant. Thus, contrary to earlier work with Rhizobiumphaseoli (1), these data demonstrate that the bacterial micro-symbiont can have a major influence on the GA content ofthe nodule either directly through de novo bacterial GAsynthesis or by stimulating GA biosynthesis by the plant.
cb0
42.9 GA2 GA20 5.0
40' 4.0
30 26.5 215 0 T 2.
2.2 3.0
F 20 2.014.2 12
1.010 071.0
0 1 0.0
60I 58. GA19 I GA19 1 6.0
50'
3: 40-
- 30^ 1.1
20 15.817
3.9
3.6
2. 12.3
0:a
5.0
4.0
3.0
2.0
1.0
0
0
P6c
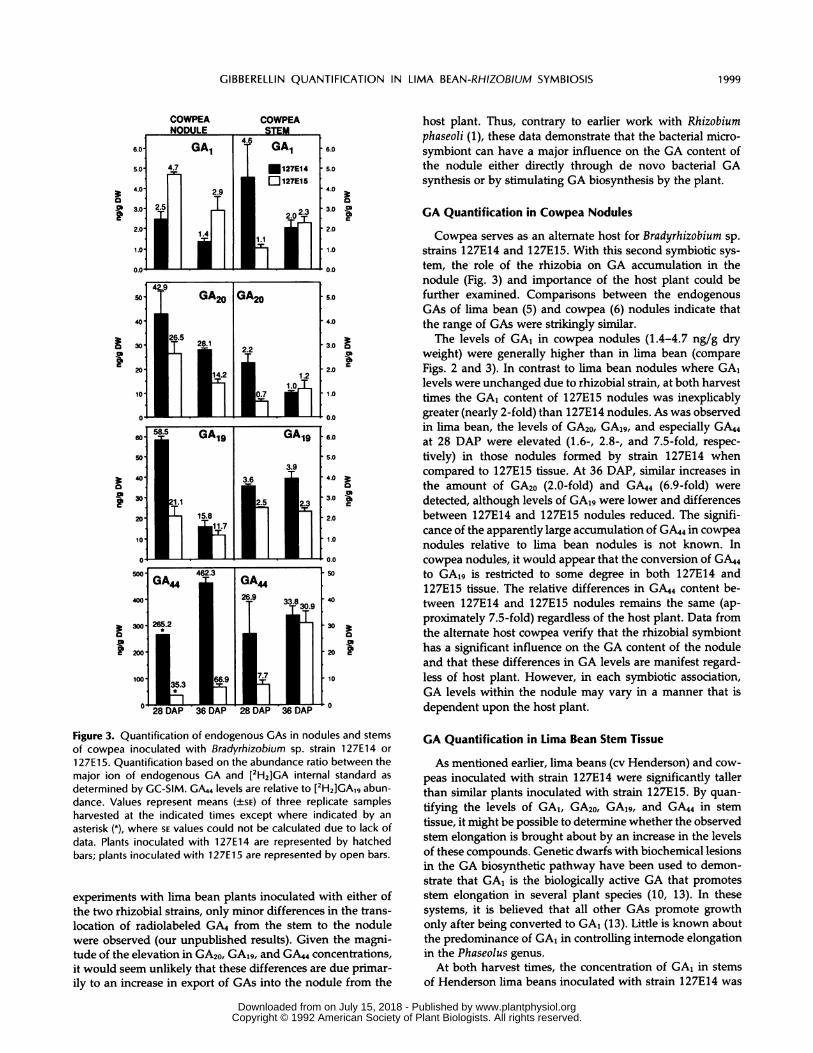
GA Quantification in Cowpea Nodules
Cowpea serves as an alternate host for Bradyrhizobium sp.
strains 127E14 and 127E15. With this second symbiotic sys-
tem, the role of the rhizobia on GA accumulation in thenodule (Fig. 3) and importance of the host plant could befurther examined. Comparisons between the endogenousGAs of lima bean (5) and cowpea (6) nodules indicate thatthe range of GAs were strikingly similar.The levels of GA, in cowpea nodules (1.4-4.7 ng/g dry
weight) were generally higher than in lima bean (compareFigs. 2 and 3). In contrast to lima bean nodules where GA,levels were unchanged due to rhizobial strain, at both harvesttimes the GA, content of 127E15 nodules was inexplicablygreater (nearly 2-fold) than 127E14 nodules. As was observedin lima bean, the levels of GA20, GA19, and especially GA44at 28 DAP were elevated (1.6-, 2.8-, and 7.5-fold, respec-
tively) in those nodules formed by strain 127E14 whencompared to 127E15 tissue. At 36 DAP, similar increases inthe amount of GA20 (2.0-fold) and GA44 (6.9-fold) were
detected, although levels of GA19 were lower and differencesbetween 127E14 and 127E15 nodules reduced. The signifi-cance of the apparently large accumulation of GA44 in cowpeanodules relative to lima bean nodules is not known. Incowpea nodules, it would appear that the conversion of GA44to GA19 is restricted to some degree in both 127E14 and127E15 tissue. The relative differences in GA44 content be-tween 127E14 and 127E15 nodules remains the same (ap-proximately 7.5-fold) regardless of the host plant. Data fromthe alternate host cowpea verify that the rhizobial symbionthas a significant influence on the GA content of the noduleand that these differences in GA levels are manifest regard-less of host plant. However, in each symbiotic association,GA levels within the nodule may vary in a manner that isdependent upon the host plant.
c
50
40
30 3
20
I10
AI I36 228BDAP 36DAP 28DAP 36DAP
Figure 3. Quantification of endogenous GAs in nodules and stemsof cowpea inoculated with Bradyrhizobium sp. strain 127E14 or
127E15. Quantification based on the abundance ratio between themajor ion of endogenous GA and [2H2]GA internal standard as
determined by GC-SIM. GA44 levels are relative to [2H2]GA19 abun-dance. Values represent means (±SE) of three replicate samplesharvested at the indicated times except where indicated by an
asterisk (*), where SE values could not be calculated due to lack ofdata. Plants inoculated with 127E14 are represented by hatchedbars; plants inoculated with 127E15 are represented by open bars.
experiments with lima bean plants inoculated with either ofthe two rhizobial strains, only minor differences in the trans-location of radiolabeled GA4 from the stem to the nodulewere observed (our unpublished results). Given the magni-tude of the elevation in GA20, GA19, and GA44 concentrations,it would seem unlikely that these differences are due primar-ily to an increase in export of GAs into the nodule from the
GA Quantification in Lima Bean Stem Tissue
As mentioned earlier, lima beans (cv Henderson) and cow-
peas inoculated with strain 127E14 were significantly tallerthan similar plants inoculated with strain 127E15. By quan-
tifying the levels of GA1, GA20, GA19, and GA44 in stemtissue, it might be possible to determine whether the observedstem elongation is brought about by an increase in the levelsof these compounds. Genetic dwarfs with biochemical lesionsin the GA biosynthetic pathway have been used to demon-strate that GA1 is the biologically active GA that promotesstem elongation in several plant species (10, 13). In thesesystems, it is believed that all other GAs promote growthonly after being converted to GA1 (13). Little is known aboutthe predominance of GA1 in controlling internode elongationin the Phaseolus genus.At both harvest times, the concentration of GA1 in stems
of Henderson lima beans inoculated with strain 127E14 was
COWPEA COWPEA
n _ . Aoo
1 999
www.plantphysiol.orgon July 15, 2018 - Published by Downloaded from Copyright © 1992 American Society of Plant Biologists. All rights reserved.

Plant Physiol. Vol. 100, 1992
the same as in those inoculated with strain 127E15 (Fig. 2).Likewise, for plants harvested at 19 DAP, the levels of GA20,GA19, and GA44 in the stem were unaffected by rhizobialinoculum. At 28 DAP, plants inoculated with 127E14 were65% taller than 127E15-inoculated plants. Unexpectedly, thelevels of GA20, GA19, and GA44 were all higher (7.8x, 1.6X,5 0x, respectively) in stems of the shorter 127E15-inoculatedplants. The relative accumulation of the precursors to GA, in127E15 plants might indicate that, in this stem tissue, the 3,B-hydroxylation step proceeds at a slower pace than in 127E14tissue. Regardless of differences in plant height, the stem dryweight per plant at both harvest times was unaffected byrhizobial inoculation (data not shown). Thus, the observeddifferences in GA20, GA19, and GA44 levels could not beattributed to a dilution effect due to greater biomass produc-tion in 127E14 plants.
After 30 d of growth, the lima bean pole variety King ofthe Garden usually attained a height of over 1.5 m, with alarge portion of the stem still actively elongating. A highdegree of variability in plant height was observed with theseplants. Therefore, differences in height between plants inoc-ulated with strain 127E14 and 127E15 were not significant.When GA levels in the stem tissue of plants inoculated witheither rhizobial strain were compared, differences were notapparent (Fig. 2). Although the level of GA, in 127E15 stemswas over three times greater than in 127E14 stems, thisdifference may, in part, be attributed to a greater variabilityin the tissue. In pole beans, as in the bush cultivar Henderson,an elevation in the level of GAs in the nodule was not directlytranslated into a widespread increase in the concentration offree GAs in the stem. Localized increases in GA levels, whichwould have gone undetected in our analyses, cannot bediscounted.
GA Quantification in Cowpea Stem Tissue
In the cowpea cultivar Purple Hull, stems were thick andintemodes remained relatively short but were still responsiveto exogenous GA application (6). Cowpeas inoculated withstrain 127E14 were 23% taller than those inoculated withstrain 127E15 after 42 d (6). Additionally, the effects of127E14-induced growth and GA treatment appeared to beadditive. Thus, as in lima bean it was speculated that strain127E14 promoted growth in cowpea through a mechanismthat was mediated by GAs (6).When the levels of GA1, GA19, GA20, and GA44 were
determined in this species, some striking differences fromlima bean were observed (Fig. 3). In all cowpea stem samples,GA1 concentration was greater than the levels found in limabean. Plants inoculated with strain 127E14 and harvested 28DAP contained a greater (4.3-fold) concentration of GA1 than127E15 plants. However, by 36 DAP, this difference in GA1levels was no longer apparent. The concentration of GA20 in28-d-old stems was also greater (3.4-fold) in 127E14 plantsthan in 127E15 plants, and again this difference was notobserved by 36 DAP. Although differences in cowpea heightat 28 DAP in a previous experiment (6) were found to beinsignificant, plant height of the tissue harvested for theseGA analyses was not determined.
CONCLUSIONS
The observation that the levels of GA1 are similar betweenthe rapidly expanding stems of Henderson lima beans inoc-ulated with strain 127E14 and the slow-growing stems from127E15 plants agrees with previous findings that GA1 con-centration is not always correlated with growth rate (28, 29).Graebe (13) has argued that the 'turnover' or metabolism ofGA1 (i.e. GA20 -- GA1 -- GA8) may be of greater importancethan the steady-state concentration of GA1 in controllingtissue elongation. In elongating cells, rapid metabolism ofGA1 to GA8 could prevent the accumulation of GA1, althoughlocalized levels of GA1 could reach some threshold concen-tration needed to promote growth (13). However, in a rangeof internode-length mutants of pea, Steane et al. (30) foundlittle evidence to support the theory that GA1 reception andaction is dependent on metabolism of GA1 to GA8.By 28 DAP, Henderson plants inoculated with strain
127E15 had the greatest proportion of nongrowing to grow-ing stem tissue (our unpublished data). Likewise, only theseplants, and not those harvested at 19 DAP, accumulate GA1precursors in stem tissue. Thus, in the transition from arapidly-growing to a slow-growing state, it is possible thatthe turnover rate of GA, decreased, allowing GA20 accumu-lation. In plants inoculated with Bradyrhizobium sp. strain127E14, the transition in stem tissue from a rapidly growingto a slow-growing state may be altered due to an elevationin the flux of 13-hydroxylated GA intermediates from noduleto shoot. The outward manifestation of this change in GAbalance would be the observed period of rapid internodeelongation.
In the experiments of Evensen and Blevins (9), lima beansinoculated with strain 127E15, when compared with thoseinoculated with strain 127E14, became less responsive to GA3treatment as plants matured. This finding also supports thenotion that 127E14-nodulated plants are maintained in aresponsive growth state, whereas plants inoculated withstrain 127E15 show a decline in competence for growth.
Analysis of GA content and quality in the legume symbioticsystem can be complicated by the possibility that two orga-nisms are contributing to the overall pool of GAs within thenodule and perhaps even in the shoot. Appropriate controlsare difficult to obtain because, as mentioned earlier, a non-nodulated legume nourished by an extemal N source isdifferent from one that is nodulated. Similarly, differences inGA production between free-living Bradyrhizobium bacteriaand bacteroids should be expected considering that each formhas a distinct genetic expression and metabolism. Thus, ashas been previously proposed (1), use of both plant andbacterial mutants may be the sole means to determine theimportance of the rhizobial symbiont to the GA status of theplant.From the data presented here, it is not possible to elucidate
the means by which an elevation of GA levels in the noduleis translated into a growth response in the stem. However,our results provide clear evidence that the rhizobial micro-symbiont can have a profound influence on the levels ofspecific GAs within the nodule. Our findings lend furthersupport to the hypothesis that a rhizobium-induced alterationin the GA balance within lima bean plants leads to an increase
DOBERT ET AL.2000
www.plantphysiol.orgon July 15, 2018 - Published by Downloaded from Copyright © 1992 American Society of Plant Biologists. All rights reserved.

GIBBERELLIN QUANTIFICATION IN LIMA BEAN-RHIZOBIUM SYMBIOSIS
in stem elongation. Together with other experimental evi-dence (2, 3), these data suggest that factors other than N are
exported by the rhizobial symbiont to the host plant.
LITERATURE CITED
1. Atzorn R, Crozier A, Wheeler CT, Sandberg G (1988) Produc-tion of gibberellins and indole-3-acetic acid by Rhizobiumphaseoli in relation to nodulation of Phaseolus vulgaris roots.Planta 175: 532-538
2. Badenoch-Jones J, Summons RE, Rolfe BG, Letham DS (1984)Phytohormones, Rhizobium mutants and nodulation in leg-umes. IV. Auxin metabolites in pea root nodules. J PlantGrowth Regul 3: 23-29
3. Badenoch-Jones J, Summons RE, Rolfe BG, Letham DS (1984)Phytohormones, Rhizobium mutants and nodulation in leg-umes. V. Cytokinin metabolism in effective and ineffectivepea root nodules. Plant Physiol 74: 239-246
4. Dangar TK, Basu PS (1987) Studies on plant growth substances,IAA metabolism and nitrogenase activity in root nodules ofPhaseolus aureus Roxb. var mungo. Biol Plant 29: 350-354
5. Dobert RC, Rood SB, Blevins DG (1992) Gibberellins and theRhizobium-legume symbiosis. I. Endogenous gibberellins oflima bean (Phaseolus lunatus L) stems and nodules. PlantPhysiol 98: 221-224
6. Dobert RC, Rood SB, Blevins DG (1992) Gibberellins and theRhizobium-legume symbiosis. II. Rhizobial-induced increase ininternode length and identification of endogenous gibberellinsof cowpea (Vigna unguiculata [L]. Walp) stems and nodules. JPlant Growth Regul (in press)
7. Dullaart J, Duba LI (1970) Presence of gibberellin-like sub-stances and their possible role in auxin bioproduction in rootnodules and roots of Lupinus luteus L. Acta Bot Neerl 19:877-883
8. Durley RC, Crozier A, Pharis RP, McLaughlin GE (1972)Chromatography of 33 gibberellins on a gradient eluted silicagel partition column. Phytochemistry 11: 3029-3033
9. Evensen K, Blevins DG (1981) Differences in endogenous levelsof gibberellin-like substances in nodules of Phaseolus lunatusL. plants inoculated with two Rhizobium strains. Plant Physiol68: 195-198
10. Fujioka S, Yamane H, Spray CR, Gaskin P, MacMillan J,Phinney BO, Takahashi N (1988) Qualitative and quantita-tive analyses of gibberellins in vegetative shoots of normal,dwarf-1, dwarf-2, dwarf-3, and dwarf-5 seedlings of Zea maysL. Plant Physiol 88: 1367-1372
11. Garcia-Martinez JL, Keith B, Bonner BA, Stafford AE, Rap-paport L (1987) Phytochrome regulation of the response toexogenous gibberellins by epicotyls of Vigna sinensis. PlantPhysiol 85: 212-216
12. Glenn JL, Kuo CC, Durley RC, Pharis RP (1972) Use on
insoluble polyvinyl-pyrrolidone for purification of plant ex-
tracts and chromatography of plant hormones. Phytochemis-try 11: 345-351
13. Graebe JE (1987) Gibberellin biosynthesis and control. AnnuRev Plant Physiol 38: 419-465
14. Grobbelaar N, Clarke B, Hough MC (1971) The nodulationand nitrogen fixation of isolated roots of Phaseolus vulgaris L.III The effect of carbon monoxide and ethylene. In TA Lie, EGMulder, eds, Plant and Soil Special Volume BiologicalNitrogen Fixation in Natural and Agricultural Habitats. Mar-tinus Nijhoof, The Hague, pp 215-221
15. Hedden P (1987) Gibberellins. In L Rivier, A Crozier, eds,
Principles and Practice of Plant Hormone Analysis. AcademicPress, New York, pp 9-99
16. Henson IE, Wheeler CT (1977) Hormones in plants bearingnitrogen-fixing root nodules: gibberellin-like substances inAlnus glutinosa (L.) Gaertn. New Phytol 78: 373-381
17. Jensen E, Crozier A, Monteiro AM (1986) Analysis of gibber-ellins and gibberellin conjugates by ion-suppression reversed-phase high-performance liquid chromatography. J Chroma-togr 367: 377-384
18. Katznelson H, Cole SE (1965) Production of gibberellin-likesubstances by bacteria and actinomycetes. Can J Microbiol 11:733-741
19. Koshioka M, Harada J, Takeno K, Noma M, Sassa T, OgiyamaK, Taylor JS, Rood SB, Legge RL, Pharis RP (1983) Reversed-phase C18 high-performance liquid chromatography of acidicand conjugated gibberellins. J Chromatogr 256: 101-115
20. Krauss A, Marschner H (1982) Influence of nitrogen nutrition,daylength and temperature on contents of gibberellic acid andabscisic acid and on tuberization in potato plants. Potato Res25: 13-21
21. Lerouge P, Roche P, Faucher C, Maillet F, Truchet G, PromeJC, Denarie J (1990) Symbiotic host-specificity of Rhizobiummeliloti is determined by a sulphated and acylated glucosamineoligosaccharide signal. Nature 344: 781-784
22. Lluch C, Ligero F, Olivares J (1983) Efecto de las gibberlinasproducidas por Rhizobium meliloti en la nodulacion de alfalfa(Medicago sativa L.). Anal Edaf Agrobiol 42: 793-805
23. Murakami Y (1968) A new rice seedling bioassay for gibberel-lins, 'microdrop method,' and its uses for testing extracts ofrice and morning glory. Bot Mag Tokyo 81: 33-43
24. Phillips DA (1971) Cytokinin production by Rhizobium japoni-cum. Physiol Plant 23: 1057-1063
25. Phillips DA (1971) Abscisic acid inhibition of root nodule initi-ation in Pisum sativum. Planta 100: 181-190
26. Radley M (1961) Gibberellin-like substances in plants. Nature191: 684-685
27. Rood SB, Blake TJ, Pharis RP (1983) Gibberellins and heterosisin maize. II. Response to gibberellic acid and metabolism of[3H] gibberellin A20. Plant Physiol 71: 645-651
28. Rood SB, Larsen KM, Mander LN, Abe H, Pharis RP (1986)Identification of endogenous gibberellins from Sorghum. PlantPhysiol 82: 330-332
29. Sponsel VM (1986) Gibberellins in dark- and red-light-grownshoots of dwarf and tall cultivars of Pisum sativum: the quan-tification, metabolism and biological activity of gibberellins inProgress No. 9 and Alaska. Planta 168: 119-129
30. Steane DA, Ross nl, Reid JB (1989) Metabolism of [3H] gibber-ellins Al in a range of internode length mutants of Pisum. JPlant Physiol 135: 70-74
31. Thimann KV (1936) On the physiology of the formation ofnodules on legume roots. Proc Natl Acad Sci USA 22:511-514
32. Trewavas A (1981) How do plant growth substances work?Plant Cell Environ 4: 203-228
33. Triplett EW, Heitholt JJ, Evensen KB, Blevins DG (1981)Increase in internode length of Phaseolus lunatus L. caused byinoculation with a nitrate reductase-deficient strain of Rhizo-bium sp. Plant Physiol 67: 1-4
34. Vincent JM (1970) A Manual for the Practical Study of Root-Nodule Bacteria. IBM Handbook No. 15. Blackwell ScientificPublications, Oxford, p 145
35. Williams PM, Sicardi De Mallorca M (1982) Abscisic acid andgibberellin-like substances in roots and root nodules of Glycinemax. Plant Soil 65: 19-26
2001
www.plantphysiol.orgon July 15, 2018 - Published by Downloaded from Copyright © 1992 American Society of Plant Biologists. All rights reserved.
![P-ISSN: 2394-0514 Antiplasmodial properties of plants ... · signaling symbiotic bacteria in the legume rhizobium symbiosis and are important in root and shoot development [14]. This](https://img.dokumen.tips/doc/110x75/5ffe13ac80f1ab6812653a9e/p-issn-2394-0514-antiplasmodial-properties-of-plants-signaling-symbiotic-bacteria.jpg)




























