Embed Size (px)
Citation preview

ORIGINAL ARTICLE
IL-7 downregulates IL-7Ra expression in human CD8T cells by two independent mechanisms
Feras M Ghazawi1,2,5, Elliott M Faller1,2,5, Scott M Sugden1,2, Juzer A Kakal1,2 and Paul A MacPherson1,2,3,4
Interleukin (IL)-7 is an essential nonredundant cytokine, and throughout the lifespan of a T-cell signaling via the IL-7 receptor
influences cell survival, proliferation and differentiation. It is therefore no surprise that expression of the IL-7 receptor
alpha-chain (CD127) is tightly regulated. We have previously shown that IL-7 downregulates expression of CD127 at the cell
surface and now elucidate the kinetics of that suppression and demonstrate that IL-7 downregulates CD127 transcripts and
surface protein in primary human CD8 T cells by two separate pathways. We show that IL-7 induces the initial reduction in cell-
surface CD127 protein independent of transcriptional suppression, which is delayed by 40–60 min. Although IL-7-mediated
downregulation of CD127 transcripts is dependent on Janus kinase (JAK)/STAT5, the early downregulation of surface CD127
protein is independent of JAK activity. The data further illustrate that low levels of IL-7 induce smaller and transient decreases
in CD127 transcripts and surface protein, whereas higher concentrations induce more profound and sustained suppression.
Such flexibility in receptor expression likely allows for fine-tuned immune responses in human CD8 T cells in different
microenvironments and in response to different immunological challenges.
Immunology and Cell Biology (2013) 91, 149–158; doi:10.1038/icb.2012.69; published online 4 December 2012
Keywords: interleukin-7; interleukin-7 receptor; CD127; cytotoxic T lymphocytes
Interleukin (IL)-7 has essential, nonredundant roles in the develop-ment, maintenance and function of T cells.1,2 It is a master regulatorof human and murine T-cell homeostasis and is required for survivalof naive and mature peripheral T cells3 and for the establishment andmaintenance of T-cell memory.4–7 IL-7 also has a pivotal role in theactivation and proliferation of cytotoxic CD8 T cells.8–10 IL-7enhances TCR-dependent T-cell activation and proliferationin vivo11 and significantly augments the cytotoxic activity of effectorT cells.12–16 IL-7 signaling is initiated upon binding of IL-7 to itshigh-affinity site on the IL-7 receptor alpha-chain (CD127), whichthen dimerizes with the second component of the IL-7 receptor, thecommon g-chain (CD132), which is shared with the receptors forIL-2, -4, -9, -15 and -21.1,17 The interaction of the two receptorsubunits brings the associated Janus kinases (JAKs)-1 and -3 intoproximity and allows their subsequent trans-phosphorylation andactivation. This leads to phosphorylation of CD127 at a criticaltyrosine residue (Y449) by JAK1, allowing docking and subsequentphosphorylation and activation of signal transducer and activatorof transcription (STAT) proteins and the p85 subunit ofphosphoinositide 3 (PI3) kinase, both of which subsequentlyregulate the expression and activity of a number of downstreamgenes and gene products.1,18 Despite the central importance of IL-7and its receptor in the development, maintenance and function of
T cells, the mechanisms regulating expression of the IL-7 receptor inresponse to IL-7 signaling have been only partially characterized.19–22
We and others have shown that IL-7 downregulates expression ofthe CD127 receptor subunit on the surface of T cells.19–25 Althoughsome groups have shown transcriptional suppression of the CD127gene in response to IL-7,19,22 others have found that IL-7 does notaffect CD127 mRNA levels in human T cells.21 Further, where IL-7has been shown to suppress CD127 gene transcription, it has not beenestablished whether the loss of CD127 protein from the cell surface isthe result of transcriptional downregulation and reduced proteinsynthesis or the result of a second mechanism acting at the cellmembrane.19,22 Indeed, it was recently reported IL-7 downregulatessurface CD127 protein in the T-leukemia cell lines HPB-ALL andTAIL7 through receptor internalization.24 Whether this applies toprimary human CD8 T cells has yet to be determined. In addition, thedetailed kinetics and roles that PI3K and JAK and downstreamphosphorylation of STAT proteins have in regulating the amount ofCD127 on the cell membrane or CD127 gene transcription or bothhave never been delineated.
In view of this, we set out to clearly and systematically define themechanisms by which IL-7 downregulates CD127 expression on thesurface of resting human CD8 T cells and to determine to what extentthe early downregulation of surface CD127 protein is dependent on
1Ottawa Hospital Research Institute, Ottawa, Ontario, Canada; 2Department of Biochemistry, Microbiology and Immunology, Faculty of Medicine, University of Ottawa, Ottawa,Ontario, Canada; 3Department of Medicine, Faculty of Medicine, University of Ottawa, Ottawa, Ontario, Canada and 4Division of Infectious Diseases, Ottawa Hospital GeneralCampus, Ottawa, Ontario, Canada
Correspondence: Dr PA MacPherson, Division of Infectious Diseases, The Ottawa Hospital, General Campus, 501 Smyth Road, Ottawa, Ontario, Canada K1H 8L6.E-mail: [email protected]
5These authors contributed equally to this study.
Received 20 August 2012; revised 16 September 2012; accepted 29 October 2012; published online 4 December 2012
Immunology and Cell Biology (2013) 91, 149–158& 2013 Australasian Society for Immunology Inc. All rights reserved 0818-9641/13
www.nature.com/icb

transcriptional suppression. We demonstrate for the first time thatIL-7 suppresses expression of its own receptor in primary humanCD8 T cells by two independent mechanisms, one transcriptional andone at the level of surface protein. Through these pathways, IL-7provides negative feedback on its own signaling cascades, allowing forfine-tuned immune responses in human CD8 T cells in differentmicroenvironments and in response to different immunologicalchallenges.
RESULTS
IL-7 downregulates CD127 mRNA transcripts and surface proteinin a time- and dose-dependent mannerWe first set out to elucidate the kinetics of IL-7-mediated suppressionof CD127 surface protein and mRNA transcripts in primary human
CD8 T cells. To do this, purified CD8 T cells were incubated withincreasing concentrations of IL-7 (100–10 000 pg ml�1), and levels ofCD127 mRNA transcripts were measured by quantitative PCR(qPCR) and compared with untreated cells, normalizing to RPS18expression. Surface CD127 protein expression was monitored by flowcytometry. As shown in Figure 1a, IL-7 suppresses the level of CD127mRNA transcripts in CD8 T cells in a dose-dependent manner. Atleast 100 pg ml�1 IL-7 was required to see a decrease in the level ofCD127 mRNA (34±4% reduction), with 10 000 pg ml�1 inducing a78±8% decline in transcripts. Maximal suppression at all concentra-tions of IL-7 occurred at 3 h. At lower concentrations of IL-7 (100–1000 pg ml�1), the reduction in CD127 transcripts was transient witha dose response in time to recovery, whereas 10 000 pg ml�1 IL-7maintain transcriptional suppression for over 72 h. Similarly, the
Figure 1 IL-7 downregulates CD127 mRNA transcripts and surface protein in CD8 T cells in a time- and dose-dependent manner. Purified CD8 T cells were
incubated in media alone or in the presence of increasing concentrations of IL-7 (100–10 000pgml�1) for up to 72h. (a) CD127 mRNA transcript levels
were measured by qPCR normalizing to RPS18. (b) Surface CD127 protein expression was analyzed by flow cytometry. Values represent relative CD127
mRNA or surface protein expression (percent positive cells) compared with media controls. Error bars represent s.e. of the mean (s.e.m.) of four
independent experiments. (c) Representative flow-cytometry histograms from one individual in (b) showing CD127 expression on CD8 T cells incubated in
media alone or with IL-7 (250 and 10000pg ml�1) for 3–12 h.
IL-7 downregulates IL-7Ra expressionFM Ghazawi et al
150
Immunology and Cell Biology

effects of IL-7 on CD127 surface protein are also dose dependent bothin the extent of the downregulation and in the duration ofsuppression (Figure 1b). A transient (17±3%) decrease in surfaceCD127 protein could be detected on CD8 T cells with as little as100 pg ml�1 of IL-7. This effect became more marked with increasingconcentrations of IL-7, where 5000 pg ml�1 induced a 41±2%reduction in CD127 surface protein within 3 h, with a maximalsuppression of 90±2% at 12 h. Recovery of CD127 on the cell surfacetook progressively longer with increasing doses of IL-7, but concen-trations of 5000–10 000 pg ml�1 maintained suppression for over72 h. Thus, we show that the effects of IL-7 on suppressing CD127mRNA transcripts and surface protein are both dose- and time-dependent, with transient reductions in expression at lower IL-7concentrations (100–1000 pg ml�1) and sustained suppression athigher concentrations (10 000 pg ml�1). The maximal reductions inCD127 mRNA and surface protein occur at 3 and 12 h, respectively, atall concentrations of IL-7.
IL-7 downregulates CD127 protein at the cell surface independentof transcriptional suppressionAs shown in Figure 1, the maximal effect of IL-7 on surface CD127protein expression was observed several hours following maximaltranscriptional suppression. This could suggest that loss of CD127protein from the cell membrane is entirely dependent on transcrip-tional suppression of the CD127 gene and natural turnover of surfaceprotein. We felt this was unlikely, however, because IL-7 induced amaximal reduction in surface CD127 protein within 12 h, whereas thenatural half-life of CD127 at the cell surface is approximately 55 h.26
To determine to what extent the downregulation of surface CD127protein is dependent on transcriptional suppression, we treated CD8T cells with IL-7 (10 ng ml�1) and measured CD127 mRNAtranscripts and surface protein at 20 min intervals by qPCR andflow cytometry, respectively. Interestingly, the downregulation ofCD127 protein at the cell surface occurred within 20 min, followingaddition of IL-7, and leveled off at 54±4% pretreatment levels overthe next 2 h (Figure 2a). In contrast, IL-7-induced suppression ofCD127 mRNA transcripts was delayed by 40–60 min post treatment(Figure 2b). These data then demonstrate that the initial down-regulation of CD127 protein at the cell surface actually precedes and istherefore likely independent of transcriptional suppression.
To further distinguish downregulation of surface CD127 proteinfrom transcriptional suppression, we treated CD8 T cells simulta-neously with IL-7 and dexamethasone (Dex), a glucocorticoidpreviously shown to upregulate CD127 transcripts in murine B andT cells.27,28 We reasoned if IL-7 is able to downregulate CD127 surfaceprotein independent of transcriptional suppression, we would see thisreduction at the cell surface, even when levels of CD127 transcripts areaugmented by the simultaneous addition of Dex. As expected(Figure 3a), treatment with Dex alone (20mM) increased CD127mRNA transcript levels in human CD8 T cells some 2.6-fold within1 h, whereas IL-7 alone (10 ng ml�1) reduced CD127 transcripts by64±5% at 4 h, reaching 80±4% suppression at 24 h. In contrast, thecombination of IL-7 plus Dex together maintained CD127 mRNAtranscripts at levels comparable to that in untreated cells. Interestingly,when added in combination with IL-7, Dex appeared to induce aninitial increase in CD127 transcripts within 1 h presumably due to therapid effect of Dex binding to the glucocorticoid receptor in thecytoplasm and its translocation to the cell nucleus. This was followedby an apparent decline in CD127 transcripts by 4 h likely due to thedelayed suppressive effects of IL-7. Although these variations wereconsistent, the changes were not statistically significant compared
with cells maintained in medium alone, and, ultimately, CD127mRNA leveled off to pretreatment levels at 24 h. Despite the initialincrease and overall lack of change in the level of CD127 transcripts,IL-7 still downregulated CD127 protein at the cell surface by 55±8%in the presence of Dex (Figure 3b). These data then confirm that IL-7downregulates surface CD127 protein independent of transcription.Notably, at 24 h, the level of surface CD127 protein was significantlyhigher in the presence of Dex plus IL-7 over IL-7 alone. This is mostlikely due to synthesis and replenishment of new CD127 protein inthe presence of normal transcript levels and highlights the contribu-tion of IL-7-induced transcriptional suppression in further limitingCD127 surface expression at later time points.
Finally, to fully separate IL-7-induced downregulation of CD127protein at the cell surface from transcriptional suppression, weutilized a Jurkat-CD127 cell line engineered to expresses the CD127cDNA from the cytomegalovirus promoter.22 Because IL-7 is notknown to attenuate transcription from the cytomegalovirus promoter,we expected CD127 transcripts would remain unchanged in thepresence of IL-7, whereas surface CD127 protein would bedownregulated. As shown in Figure 4, this is exactly what we found.Although treatment with IL-7 did not suppress CD127 mRNAtranscripts in the Jurkat-CD127 cell line (Figure 4a) over 6 h, IL-7did significantly downregulate CD127 protein at the cell surface
Figure 2 IL-7 induces the downregulation of surface CD127 protein within
20 min while suppression of CD127 mRNA is delayed by 40–60 min.
Purified CD8 T cells were incubated in media alone or in the presence of
IL-7 (10 ngml�1) and analyzed at 20 min intervals. (a) Surface CD127expression was measured by flow cytometry. (*Po0.05 compared with
media). (b) CD127 mRNA transcript levels were measured by qPCR
normalizing to RPS18 (*Po0.05 compared with media). Values represent
relative CD127 mRNA or surface protein (mean fluorescence intensities)
compared with media controls. Error bars represent s.e.m of at least four
independent experiments.
IL-7 downregulates IL-7Ra expressionFM Ghazawi et al
151
Immunology and Cell Biology

(Figure 4b). This downregulation of surface CD127 protein in theabsence of changes in CD127 mRNA levels confirms that IL-7 inducesthe loss of CD127 protein from the cell surface independent of mRNAsuppression.
IL-7-mediated downregulation of CD127 mRNA transcripts isdependent on JAK/pSTAT5JAK/STAT and PI3K are considered the major IL-7-mediated signalingpathways in T cells,1,2 and we therefore set out to determine which, ifeither, of these pathways is involved in IL-7-induced downregulationof CD127 transcripts and surface protein. The role of IL-7 inactivating JAK3/STAT5 signaling is well established,20,22,23,29 and weconfirm here that IL-7 induces phosphorylation of STAT5 in primaryhuman CD8 T cells (Figure 5a). Phosphorylation of STAT3 inresponse to other g chain cytokines, such as IL-2,30 IL-4,30 IL-9,31
IL-1530 and IL-21,32 has been previously reported, but to ourknowledge, induction of STAT3 phosphorylation by IL-7 in humanCD8 T cells has not been investigated. To examine this, we treatedCD8 T cells with IL-7 (10 ng ml�1) or IL-10 (10 ng ml�1) as a positivecontrol33–35 and measured intracellular phospho-STAT3 by flowcytometry. As shown in Figure 5b, IL-10 induces STAT3 phosphor-ylation, whereas IL-7 does not. Finally, we used a synthetic STAT5
inhibitor that specifically targets the SH2 domain of STAT5 to blockits phosphorylation in the presence of IL-7. Figure 5c shows a dosetitration and complete block of STAT5 phosphorylation in thepresence of IL-7 plus 500mM inhibitor.
To examine the role of the JAK/STAT5 and PI3K signaling pathwaysin IL-7-induced downregulation of CD127 transcripts, we pretreatedCD8 T cells with inhibitors of either JAK or PI3K for 1 h followed byIL-7 (10 ng ml�1) for 80 min. The PI3K inhibitor (LY294002; 10mM)successfully blocked phosphorylation of Akt following IL-7 stimula-tion in primary human CD8 T cells, whereas the JAK inhibitor(1.5mM) blocked IL-7-induced STAT5 phosphorylation (data notshown). As shown in Figure 6a, preincubating cells with JAK inhibitorcompletely blocked IL-7-mediated downregulation of CD127 tran-scripts, whereas inhibition of PI3K signaling had no effect. Theseresults clearly indicate a requirement for JAK activity but not PI3K inthe downregulation of CD127 mRNA by IL-7. We next questionedwhether STAT5 phosphorylation had a role in the suppression ofCD127 transcripts by IL-7. To investigate this, CD8 T cells werepreincubated for 1 h with 500mM STAT5 inhibitor followed by IL-7(10 ng ml�1) for 60 min, and levels of CD127 mRNA transcripts weremeasured by qPCR. As shown in Figure 6b, inhibition of STAT5phosphorylation blocked the effect of IL-7, confirming that transcrip-tional suppression of CD127 by IL-7 is dependent on the JAK/STAT5signaling pathway.
Figure 3 IL-7 downregulates the expression of surface CD127 protein, even
when CD127 mRNA levels are augmented by Dex. Purified CD8 T cells were
incubated in media alone or in the presence of IL-7 (10 ngml�1), Dex
(20mM) or IL-7 plus Dex for 1, 4 and 24h. (a) CD127 mRNA transcript
levels were measured by qPCR normalizing to RPS18. (b) Surface CD127
expression was analyzed by flow cytometry (*Po0.05 and #P40.05
compared with media). Values represent relative CD127 mRNA or surface
protein expression (percent positive cells) compared with media control.
Error bars represent s.e.m of four independent experiments.
Figure 4 IL-7 downregulates the expression of surface CD127 protein in theJurkat-CD127 cell line without affecting CD127 mRNA levels. Jurkat-
CD127 cells were incubated in media alone or with IL-7 (10 ngml�1) for 1,
3 and 6 h. (a) CD127 mRNA transcript levels were measured by qPCR
normalizing to RPS18 (P¼0.126 at 1 h, 0.821 at 3 h and 0.328 at 6 h
relative to media). (b) Surface CD127 expression was analyzed by flow
cytometry (*Po0.006 compared with media). Data are represented as
relative changes compared with media controls. Error bars represent s.e.m
of three independent experiments.
IL-7 downregulates IL-7Ra expressionFM Ghazawi et al
152
Immunology and Cell Biology

Although IL-7-mediated downregulation of CD127 protein at thecell surface occurs independently of JAK/STAT5 and PI3K,IL-7-induced degradation of CD127 is JAK dependentTo examine the requirement of the JAK/STAT5 and PI3K signalingpathways in IL-7-stimulated downregulation of CD127 protein at thecell surface, CD8 T cells were we pretreated with inhibitors of eitherJAK, PI3K or STAT5 for 1 h followed by IL-7 (10 ng ml�1) for 80 min
in the case of JAK and PI3K inhibitors or for 60 min in the case of theSTAT5 inhibitor. Levels of surface CD127 protein were then measuredby flow cytometry. As shown in Figures 7a and b, inhibition of JAK,PI3K and STAT5 phosphorylation had no effect on IL-7-mediated
Figure 6 IL-7-mediated suppression of CD127 transcripts is dependent on
JAK activity and STAT5 phosphorylation but not on PI3K. Purified CD8
T cells were maintained in media alone or preincubated with inhibitors of
JAK (1.5mM), PI3K (10mM) or STAT5 phosphorylation (500mM) for 1 h
followed by treatment with IL-7 (10 ngml�1) for 80min in the case of the
JAK and PI3K inhibitors (a) and for 60min in the case of the STAT5
inhibitor (b). CD127 transcript levels were measured by qPCR normalizingto RPS18. Data are represented as relative changes compared with media
control. (a) *Po0.005 relative to media. (b) P¼0.02 IL-7 compared with
media, P¼0.37 STAT5iþ IL-7 compared with STAT5i and P¼0.01
STAT5iþ IL-7 compared with IL-7. Error bars represent s.e.m of four
independent experiments.
Figure 5 IL-7 induces the phosphorylation of STAT5 but not STAT3 in
human CD8 T cells. Purified CD8 T cells were maintained in media alone or
treated with IL-7 (10ng ml�1) or IL-10 (10 ngml�1) for 45min and
then fixed, permeabilized and stained with fluorescein isothiocyanate
(FITC)-labeled anti-phospho-STAT3 or anti-phospho-STAT5 antibodies.
(a) Representative flow-cytometry histogram showing phospho-STAT5 in
untreated cells (gray fill) and cells stimulated with IL-7 (black line).
(b) Representative flow-cytometry histogram showing phospho-STAT3 in
untreated cells (gray fill) and cells stimulated with IL-7 (black line) or IL-10
(dashed line). (c) Purified CD8 T cells were maintained in media alone,
treated with IL-7 (10ng ml�1) for 8 h or preincubated with increasing
concentrations of STAT5 inhibitor (100–500mM) for 1 h and then treatedwith IL-7 (10 ngml�1) for an additional 8 h. Cells were then fixed,
permeabilized and stained with FITC-labeled anti-phospho-STAT5 antibodies
and analyzed by flow cytometry. Data show percent positive cells for each
condition (*Po0.03 and #P¼0.49 compared with media). Error bars
represent s.e.m of four independent experiments.
IL-7 downregulates IL-7Ra expressionFM Ghazawi et al
153
Immunology and Cell Biology
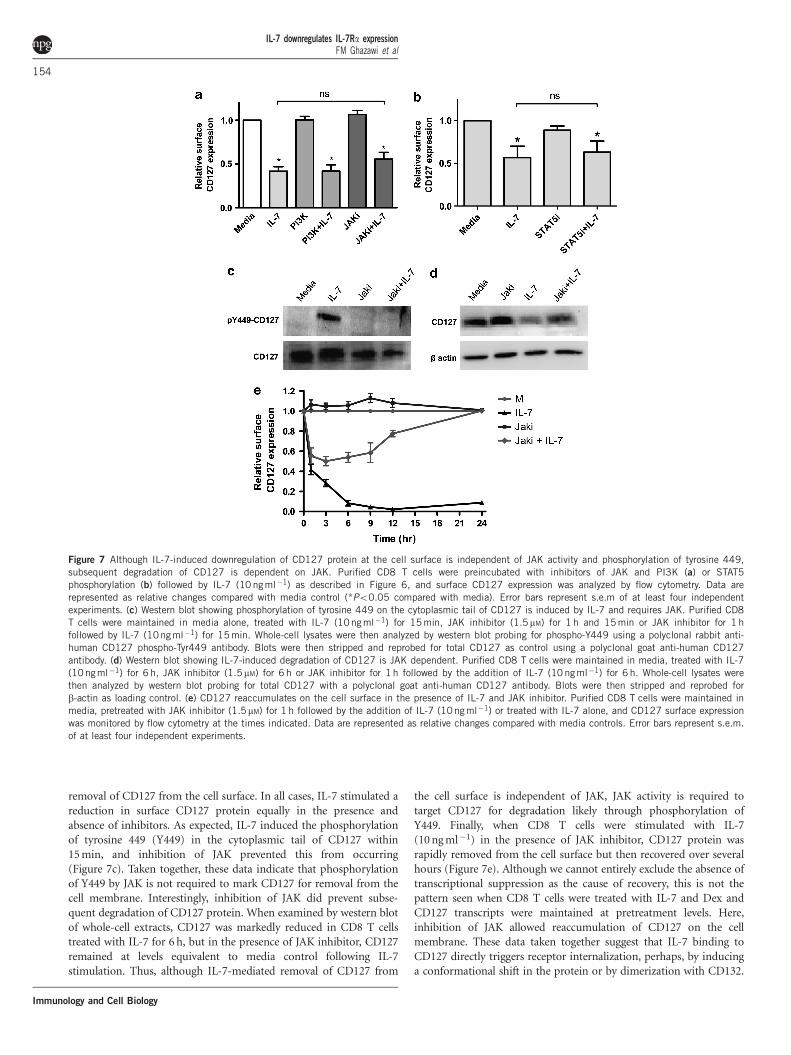
removal of CD127 from the cell surface. In all cases, IL-7 stimulated areduction in surface CD127 protein equally in the presence andabsence of inhibitors. As expected, IL-7 induced the phosphorylationof tyrosine 449 (Y449) in the cytoplasmic tail of CD127 within15 min, and inhibition of JAK prevented this from occurring(Figure 7c). Taken together, these data indicate that phosphorylationof Y449 by JAK is not required to mark CD127 for removal from thecell membrane. Interestingly, inhibition of JAK did prevent subse-quent degradation of CD127 protein. When examined by western blotof whole-cell extracts, CD127 was markedly reduced in CD8 T cellstreated with IL-7 for 6 h, but in the presence of JAK inhibitor, CD127remained at levels equivalent to media control following IL-7stimulation. Thus, although IL-7-mediated removal of CD127 from
the cell surface is independent of JAK, JAK activity is required totarget CD127 for degradation likely through phosphorylation ofY449. Finally, when CD8 T cells were stimulated with IL-7(10 ng ml�1) in the presence of JAK inhibitor, CD127 protein wasrapidly removed from the cell surface but then recovered over severalhours (Figure 7e). Although we cannot entirely exclude the absence oftranscriptional suppression as the cause of recovery, this is not thepattern seen when CD8 T cells were treated with IL-7 and Dex andCD127 transcripts were maintained at pretreatment levels. Here,inhibition of JAK allowed reaccumulation of CD127 on the cellmembrane. These data taken together suggest that IL-7 binding toCD127 directly triggers receptor internalization, perhaps, by inducinga conformational shift in the protein or by dimerization with CD132.
Figure 7 Although IL-7-induced downregulation of CD127 protein at the cell surface is independent of JAK activity and phosphorylation of tyrosine 449,
subsequent degradation of CD127 is dependent on JAK. Purified CD8 T cells were preincubated with inhibitors of JAK and PI3K (a) or STAT5
phosphorylation (b) followed by IL-7 (10 ngml�1) as described in Figure 6, and surface CD127 expression was analyzed by flow cytometry. Data are
represented as relative changes compared with media control (*Po0.05 compared with media). Error bars represent s.e.m of at least four independent
experiments. (c) Western blot showing phosphorylation of tyrosine 449 on the cytoplasmic tail of CD127 is induced by IL-7 and requires JAK. Purified CD8T cells were maintained in media alone, treated with IL-7 (10ng ml�1) for 15 min, JAK inhibitor (1.5mM) for 1 h and 15 min or JAK inhibitor for 1 h
followed by IL-7 (10ngml�1) for 15 min. Whole-cell lysates were then analyzed by western blot probing for phospho-Y449 using a polyclonal rabbit anti-
human CD127 phospho-Tyr449 antibody. Blots were then stripped and reprobed for total CD127 as control using a polyclonal goat anti-human CD127
antibody. (d) Western blot showing IL-7-induced degradation of CD127 is JAK dependent. Purified CD8 T cells were maintained in media, treated with IL-7
(10ngml�1) for 6 h, JAK inhibitor (1.5mM) for 6 h or JAK inhibitor for 1 h followed by the addition of IL-7 (10 ngml�1) for 6 h. Whole-cell lysates were
then analyzed by western blot probing for total CD127 with a polyclonal goat anti-human CD127 antibody. Blots were then stripped and reprobed for
b-actin as loading control. (e) CD127 reaccumulates on the cell surface in the presence of IL-7 and JAK inhibitor. Purified CD8 T cells were maintained in
media, pretreated with JAK inhibitor (1.5mM) for 1 h followed by the addition of IL-7 (10ngml�1) or treated with IL-7 alone, and CD127 surface expression
was monitored by flow cytometry at the times indicated. Data are represented as relative changes compared with media controls. Error bars represent s.e.m.
of at least four independent experiments.
IL-7 downregulates IL-7Ra expressionFM Ghazawi et al
154
Immunology and Cell Biology

Once in the early endosome, CD127 may then be sorted, such thatCD127 phosphorylated at Y449 by JAK is targeted for degradation,whereas unphosphorylated CD127 is recycled back to the cell surface.
DISCUSSION
Although IL-7 signaling has pivotal roles in the development andfunction of T cells, the mechanisms regulating expression of the IL-7receptor have only been partially characterized. Although severalstudies have already reported that IL-7 downregulates the expressionof CD127 mRNA transcripts and surface protein in human andmurine T cells,19,22 conflicting data have been provided suggestingthat IL-7 does not affect CD127 gene transcription.21 Whether thesediscordant results are due to the source of T cells used in each study(for example, umbilical cord versus adult peripheral blood), cultureconditions or the single time point of analysis is unclear. Further, ithas not been determined whether IL-7-mediated downregulation ofCD127 protein at the cell surface is the result of transcriptionalsuppression and reduced protein synthesis or due to a secondpathway acting at the cell surface as has been reported forT-leukemic cell lines.24 Here we address a number of thesequestions, delineating the detailed kinetics and separatecontributions of IL-7-mediated suppression of CD127 genetranscription and the initial downregulation of CD127 surfaceprotein in primary human CD8 T cells, and provide evidence withrespect to the mechanisms and pathways by which these occur.
The effects of IL-7 on reducing CD127 transcripts and surfaceprotein are dose dependent both in the extent of the downregulationand in the duration of suppression with maximal effects of IL-7 onCD127 mRNA and surface protein at 3 and 12 h, respectively. Lowconcentrations of IL-7 (100–1000 pg ml�1) induced transient declinesin CD127 mRNA and protein, whereas higher concentrations(10 000 pg ml�1) sustained suppression for over 72 h. Although thismay be due to either a bona fide concentration effect or sustainedsignaling at higher IL-7 concentrations, either way, this allowsflexibility in regulating CD127 expression in different microenviron-ments with different levels of IL-7. Although decreased transcriptionof the CD127 gene was viewed as the primary mechanism by whichIL-7 downregulates surface CD127 protein,19,20,22,36 we now showthat transcriptional suppression contributes only at later time pointsand serves to maintain low CD127 levels after the receptor has beenremoved from the cell membrane by IL-7. We demonstrate by severalmethods that IL-7 reduces CD127 transcripts and surface proteinthrough separate pathways. In both the Jurkat-CD127 cell line andwhen added to primary human CD8 T cells in combination with Dex,IL-7 removed CD127 protein from the cell surface in the absence ofchanges in CD127 mRNA levels. We also show that IL-7-mediateddownregulation of CD127 protein at the cell surface is rapid andoccurs within 20 min before there is any reduction in CD127transcripts. This results in an initial new equilibrium in proteinturnover, maintaining lower but relatively constant CD127 expressionon the cell surface over the next 2 h until CD127 transcriptionalsuppression kicks in to prevent protein replenishment and furtherdownregulate CD127 expression. The contribution of transcriptionalsuppression in reducing CD127 expression at later time points isevident in CD8 T cells treated with IL-7 and Dex, where CD127mRNA and surface protein are higher at 24 h compared with cellstreated with IL-7 alone. This effect is less evident in the Jurkat-CD127cells because protein turnover on the surface of this cell line is muchmore rapid compared with primary cells (data not shown). Thesedata together suggest that brief exposure to IL-7 may allow rapidtransient decreases in surface CD127 protein, whereas sustained
exposure to IL-7 results in transcriptional suppression and furtherreductions in receptor expression over time. This would provide afinely tuned mechanism by which CD8 T cells could respond to smallchanges in IL-7 concentrations in the microenvironment and todifferent immunological challenges.
Suppression of CD127 mRNA transcripts by IL-7 is dependent onthe JAK/STAT5 signaling pathway. It is already established that IL-7stimulation leads to phosphorylation of CD127 at Y449 by JAK3,1
allowing docking via their SH2 domains and subsequentphosphorylation of STAT5 proteins.1,18,37 Phospho-STAT5 thendimerizes and translocates to the nucleus where it may eitherdirectly or indirectly suppress CD127 transcription. Given the delayin transcriptional suppression by 40–60 min following stimulationwith IL-7, we hypothesize that IL-7 signaling via STAT5 results in theupregulation of a transcriptional repressor or an epigenetic modifierprotein that in turn suppresses CD127 gene expression. Consistentwith this hypothesis, the translational inhibitor cycloheximide blocksthe ability of IL-7 to downregulate CD127 transcripts in human CD8T cells (Ghazawi et al., manuscript in preparation), confirming thatnew protein synthesis is in fact required for the transcriptionalsuppression. Park et al.19 reported a similar effect with cycloheximideand implicated the transcriptional repressor Gfi-1 in thedownregulation of CD127 gene expression in CD8 T cells in mice.However, to date, the repressor induced by IL-7 and responsible forsuppressing CD127 gene transcription in human CD8 T cells has yetto be identified.
The IL-7-mediated downregulation of CD127 protein on thesurface of human CD8 T cells is rapid and occurs within 20 min.Similarly, Swainson et al.20 reported that surface CD127 protein isreduced on human CD4 T cells within 30 min of IL-7 stimulation(10 ng ml�1), and Henriques et al.24 documented the rapidinternalization of CD127 protein from the surface of the leukemiccell line HPB-ALL following addition of IL-7 (50 ng ml�1). We showhere that, as expected, IL-7 induces the phosphorylation of tyrosine449 within the cytoplasmic tail of CD127 by JAK and that thisphosphorylation is not required for the initial removal of CD127protein from the cell membrane. Indeed, IL-7 still downregulatessurface CD127 protein in the presence of JAK inhibitor. However,inhibition of JAK does prevent subsequent degradation of CD127 andallows the reaccumulation of receptor on the cell surface over 12–24 h.Taken together, the data suggest that IL-7 binding to CD127 directlytriggers receptor internalization, perhaps, through dimerization withCD132 or by inducing a conformational shift in the protein. Once inthe early endosome, CD127 may then be sorted, such that CD127phosphorylated at Y449 by JAK is targeted for degradation, whereasunphosphorylated CD127 is recycled back to the cell surface. Thesepathways are currently being further delineated in our laboratory.
The rapid downregulation of surface CD127 protein by IL-7 beforeCD127 transcriptional suppression and the dose response in extentand duration of suppression highlights the importance of a fine-tunedresponse to IL-7 in CD8 T cells. IL-7 has several pivotal roles in CD8T-cell survival and function including but not limited to theupregulation of Bcl-2,23 induction of perforin expression,15
enhanced glucose metabolism38 as well as the establishment andmaintenance of memory.6,39,40 The fine-tuned modulation of surfaceCD127 protein may allow CD8 T cells to regulate these functionsin response to varying IL-7 concentrations in differentmicroenvironments and in response to different immunologicalchallenges. A disruption in this dynamicity could also lead to pooror inappropriate CD8 T-cell responses. Indeed, dysregulation of IL-7signaling has been implicated in a number of diseases, such as
IL-7 downregulates IL-7Ra expressionFM Ghazawi et al
155
Immunology and Cell Biology

multiple sclerosis,41,42 rheumatoid arthritis and juvenile idiopathicarthritis, as well as in breast cancer.43,44 The expression of CD127 isalso dysregulated in Epstein–Barr virus,45–47 cytomegalovirus45,47 andhepatitis C virus48 infections. HIV infection in particular is associatedwith decreased expression of CD127 on CD4 and CD8 T cells.25,49–55
In fact, we have previously shown that soluble HIV Tat proteindownregulates CD127 on the surface of CD8 T cells isolated fromhealthy donors and targets the receptor for degradation by theproteasome.15,26 Understanding the detailed mechanisms by whichIL-7 regulates expression of its own receptor may well lead to noveltherapeutic strategies aimed at improving T-cell number and functionin lymphopenic conditions, as well as in chronic viral infections andseveral autoimmune diseases.
MATERIALS AND METHODS
Jurkat-CD127 cell line and primary human CD8 T cellsThe Jurkat-CD127 cell line was a kind gift from Dr Rene van Lier (Academic
Medical Centre, Amsterdam, The Netherlands). These cells contain the
retrovirally transduced CD127-IRES-GFP sequence and express the human
CD127 cDNA from the cytomegalovirus promoter.22
Primary human CD8 T cells were isolated as previously described.15,23,26
Peripheral blood mononuclear cells were isolated from the blood of healthy
volunteers by Ficoll-Paque density centrifugation followed by CD8 T-cell
purification using magnetic-activated cell sorting human CD8 microbeads and
the AutoMACS Isolation System (Miltenyi Biotech, Auburn, CA, USA).
Purified CD8 T cells and Jurkat-CD127 cells were cultured at a density of
1� 106 cells per ml at 37 1C in media comprised of RPMI 1640 (Hyclone,
Logan, UT, USA) supplemented with 100 U ml�1 penicillin, 100mg ml�1
streptomycin and 0.2 M L-glutamine (Gibco, Grand Island, NY, USA) and
either 20% fetal calf serum (Cansera, Rexdale, Ontario, Canada) for primary
CD8 T cells or 10% fetal calf serum for the Jurkat-CD127 cells. All research
conducted using human blood was reviewed and approved by the Ottawa
Hospital Research Ethics Board.
ReagentsIL-7 and IL-10, obtained from Invitrogen Biosource (Carlsbad, CA, USA), were
resuspended in phosphate-buffered saline plus 0.1% bovine serum albumin at
a stock concentration of 10 ngml�1 and stored at �80 1C. Dex was obtained
from BioVision (San Francisco, CA, USA) and was resuspended in dimethyl
sulfoxide at a stock concentration of 10 mM and stored at �20 1C. STAT5
inhibitor (573108; 0.1 M), JAK inhibitor 1 (420097; 10 mM) and PI3K inhibitor
(LY294002; 10 mM) were purchased from EMD Biosciences (San Diego, CA,
USA) and were dissolved in dimethyl sulfoxide at the indicated stock
concentrations and stored at �20 1C. CD8 T cells were incubated with these
inhibitors at the indicated concentrations for 1 h before the addition of IL-7.
Quantitative real-time PCRTotal RNA was harvested from 5� 105 Jurkat-CD127 or CD8 T cells using the
RNAqueous-4PCR kit for isolation of DNA-free RNA (Ambion, Austin, TX,
USA) according to the manufacturer’s instructions. RNA was quantified using
a Nanodrop ND-1000 instrument (Nanodrop, Wilmington, DE, USA) and
reverse transcribed using the iScript cDNA Synthesis Kit from Bio-Rad
(Hercules, CA, USA). Equal volumes of cDNA from each sample were then
mixed with IQ SYBR Green Supermix (Bio-Rad) and used to quantify CD127
transcripts relative to the RPS18 reference gene. Both reverse transcription and
qPCR were carried out in the iCycler thermal cycler from Bio-Rad. Samples
were incubated at 95 1C for 3 min and then amplified over 40 cycles as follows:
95 1C for 30 s, 59 1C for 20 s and 72 1C for 20 s. At the end of each cycle,
samples were held at 85 1C for 5 s for fluorescence acquisition. Following
amplification, samples were denatured for 1 min at 95 1C followed by final
extension at 72 1C for 1 min. CD127 transcripts were amplified using a forward
primer spanning the exon 4/5 boundary (50-ATGGACGCATGTGAATTTA
TC-30) and a reverse primer in exon 5 (50-GGGAGATGGATCCTATC-30). The
forward primer for RPS18 spanned the exon 2/3 boundary (50-CTGCCATTAA
GGGTGTGG-30) with the reverse primer in exon 3 (50-TCCATCCTTT
ACATCCTTCTG-30). Data analysis was carried out using the 2�DDCt principle
with the Bio-Rad Gene expression analysis macro program. The resulting
expression values were plotted using GraphPad Prism software (La Jolla, CA,
USA), and statistical analysis was done using a two-tailed, paired Student’s
t-test with 95% confidence intervals.
Western blot analysisCD8 T cells (1� 106 cells per condition) were collected by centrifugation at
6000 g for 10 min, and whole-cell extracts were prepared by resuspending the
cell pellets in 50ml RIPA buffer (50 mM Tris–HCl pH 7.4, 150 mM NaCl, 2 mM
EDTA, 1% NP-40 and 0.1% SDS with 1� Halt Protease Inhibitor Cocktail
(Thermo Scientific, Rockford, IL, USA)) and incubating at 4 1C for 45 min.
Protein concentration was determined by the Bradford protein assay. Equal
amounts of total cell protein were then resolved by SDS–PAGE and transferred
to polyvinylidene difluoride membranes (Millipore, Bedford, MA, USA) using
a semi-dry transfer apparatus (Bio-Rad) for 30 min at 15 V. Following transfer,
membranes were blocked overnight in Tris-buffered saline with Tween-20
(TBST, 50 mM Tris–HCl pH7.6, 150 mM NaCl and 0.1% Tween-20), containing
5% nonfat milk. Total CD127 protein was detected using a polyclonal goat
anti-human CD127 antibody (R&D Systems, Minneapolis, MN, USA),
whereas CD127 phosphorylated at Tyr449 was detected using a polyclonal
rabbit anti-human CD127 phospho-Tyr449 antibody (Rockland, Biomol,
Hamburg, Germany) at a concentration of 1mg ml�1 in TBST blocking
solution. Horseradish peroxidase-conjugated Donkey anti-goat (1:5000) and
Donkey anti-rabbit (1:5000) from R&D Systems were used as secondary
antibodies, and proteins were visualized by chemiluminescence using the ECL
Advance Western Blotting Detection Kit (GE Healthcare, Chalfont St Giles,
UK) according to the manufacturer’s instructions. For reprobing for loading
controls, membranes were stripped for 30 min at 50 1C in stripping buffer (2%
SDS, 62.5 mM Tris–HCl pH 6.8 and 100 mM b-mercaptoethanol), washed five
times with TBST, blocked with 5% nonfat milk in TBST for 1 h and then
reprobed with a monoclonal mouse anti-human b-actin antibody (C4; Santa
Cruz Biotechnology, Santa Cruz, CA, USA) diluted (1:200, 1mg ml�1) in TBST
blocking solution followed by an horseradish peroxidase-conjugated Donkey
anti-mouse secondary antibody (R&D Systems) at 1:5000 dilution.
Flow cytometryAnti-CD8-phycoerythrin-Cy5 (PC5) (B9.11) and anti-CD127-phycoerythrin
(PE) (R34.34) fluorochrome-labeled monoclonal antibodies were purchased
from Immunotech Beckman Coulter (Marseille, France). Monoclonal anti-
phospho-STAT5-fluorescein isothiocyanate (FITC) (Y694) and anti-phospho-
STAT3-(Y705) antibodies were purchased from BD Biosciences (San Jose, CA,
USA). All fluorochrome-labeled antibodies used for flow cytometry were
titrated and used at saturating concentrations.
Surface CD127 protein expression was analyzed by incubating cells with
anti-CD127-phycoerythrin antibodies for 30 min in the dark at room
temperature followed by analysis on a Coulter Epics ALTRA flow cytometer
(Fullerton, CA, USA). The protocol was modified slightly in Figure 2a. To
prevent further internalization of CD127 while staining with the antibody,
CD8 T cells were treated with IL-7 for the times indicated and then incubated
with anti-CD127-phycoerythrin antibodies for 30 min on ice. Intracellular
phospho-STAT3 and phospho-STAT5 staining was performed as follows:
isolated CD8 T cells were fixed in 2% paraformaldehyde for 10 min at room
temperature and then permeabilized by incubating in cold 100% methanol for
an additional 10 min at 4 1C. After washing in phosphate-buffered saline, cells
were incubated with anti-phospho-STAT3-fluorescein isothiocyanate or anti-
phospho-STAT5-fluorescein isothiocyanate antibodies in the dark for 30 min
and then analyzed by flow cytometry. Live cells were gated on the basis of side
and forward scatter, and at least 10 000 events were recorded for each sample.
Isotype controls were performed for each fluorochrome-conjugated antibody.
Resulting profiles were analyzed with FCS Express 2 software (De Novo, Los
Angeles, CA, USA), and statistical analysis was done using a two-tailed, paired
Student’s t-test with 95% confidence intervals.
CONFLICT OF INTERESTThe authors declare no conflict of interest.
IL-7 downregulates IL-7Ra expressionFM Ghazawi et al
156
Immunology and Cell Biology

ACKNOWLEDGEMENTSThis work was supported by a grant from the Canadian Institutes of Health
Research (CIHR). PAM is the principal recipient of the research grant. FMG
and SMS are recipients of CIHR doctoral scholarships and EMF is a recipient
of a postdoctoral fellowship from the Ontario HIV Treatment Network. PAM
is supported in part by a mid-career salary award from the Department of
Medicine, University of Ottawa. We thank the volunteers who donated blood
samples for this study.
Author contributions: FMG carried out the qPCR in CD8 T cells and Jurkat-
CD127 cells and flow cytometry in CD8 T cells (Figures 1a, 2b, 3a, 4a, 6, 7b
and e), wrote and revised the manuscript. EMF carried out flow cytometry in
CD8 T cells (Figures 1b and c, 2a, 3b and 5). SMS performed flow cytometry
in Jurkat-CD127 cells and CD8 T cells and carried out western blot analysis
(Figures 4b and 7a, c–e). JAK participated in the ground work of the project.
PAM designed the research, analyzed and interpreted data, and co-wrote the
manuscript.
1 Jiang Q, Li WQ, Aiello FB, Mazzucchelli R, Asefa B, Khaled AR et al. Cell biologyof IL-7, a key lymphotrophin. Cytokine Growth Factor Rev 2005; 16:513–533.
2 Fry TJ, Mackall CL. Interleukin-7: from bench to clinic. Blood 2002; 99: 3892–3904.3 Khaled AR, Durum SK. Lymphocide: cytokines and the control of lymphoid home-
ostasis. Nat Rev Immunol 2002; 2: 817–830.4 Becker TC, Wherry EJ, Boone D, Murali-Krishna K, Antia R, Ma A et al. Interleukin 15
is required for proliferative renewal of virus-specific memory CD8 T cells. J Exp Med2002; 195: 1541–1548.
5 Goldrath AW, Sivakumar PV, Glaccum M, Kennedy MK, Bevan MJ, Benoist C et al.Cytokine requirements for acute and Basal homeostatic proliferation of naive andmemory CD8þ T cells. J Exp Med 2002; 195: 1515–1522.
6 Kieper WC, Tan JT, Bondi-Boyd B, Gapin L, Sprent J, Ceredig R et al. Overexpression ofinterleukin (IL)-7 leads to IL-15-independent generation of memory phenotype CD8þT cells. J Exp Med 2002; 195: 1533–1539.
7 Tan JT, Ernst B, Kieper WC, LeRoy E, Sprent J, Surh CD. Interleukin (IL)-15and IL-7 jointly regulate homeostatic proliferation of memory phenotype CD8þ cellsbut are not required for memory phenotype CD4þ cells. J Exp Med 2002; 195:1523–1532.
8 Maraskovsky E, Teepe M, Morrissey PJ, Braddy S, Miller RE, Lynch DH et al. Impairedsurvival and proliferation in IL-7 receptor-deficient peripheral T cells. J Immunol 1996;157: 5315–5323.
9 Jicha DL, Mule JJ, Rosenberg SA. Interleukin 7 generates antitumor cytotoxicT lymphocytes against murine sarcomas with efficacy in cellular adoptive immunother-apy. J Exp Med 1991; 174: 1511–1515.
10 Sun JC, Lehar SM, Bevan MJ. Augmented IL-7 signaling during viral infection drivesgreater expansion of effector T cells but does not enhance memory. J Immunol 2006;177: 4458–4463.
11 Saini M, Pearson C, Seddon B. Regulation of T cell-dendritic cell interactions by IL-7governs T-cell activation and homeostasis. Blood 2009; 113: 5793–5800.
12 Sin JI, Kim J, Pachuk C, Weiner DB. Interleukin 7 can enhance antigen-specificcytotoxic-T-lymphocyte and/or Th2-type immune responses in vivo. Clin Diagn LabImmunol 2000; 7: 751–758.
13 Kos FJ, Mullbacher A. Induction of primary anti-viral cytotoxic T cells by in vitrostimulation with short synthetic peptide and interleukin-7. Eur J Immunol 1992; 22:3183–3185.
14 Carini C, Essex M. Interleukin 2-independent interleukin 7 activity enhances cytotoxicimmune response of HIV-1-infected individuals. AIDS Res Hum Retroviruses 1994;10: 121–130.
15 Faller EM, McVey MJ, Kakal JA, MacPherson PA. Interleukin-7 receptor expression onCD8 T-cells is downregulated by the HIV Tat protein. J Acquir Immune Defic Syndr2006; 43: 257–269.
16 Catalfamo M, Henkart PA. Perforin and the granule exocytosis cytotoxicity pathway.Curr Opin Immunol 2003; 15: 522–527.
17 Goodwin RG, Friend D, Ziegler SF, Jerzy R, Falk BA, Gimpel S et al. Cloning of thehuman and murine interleukin-7 receptors: demonstration of a soluble form andhomology to a new receptor superfamily. Cell 1990; 60: 941–951.
18 Kang J, Der SD. Cytokine functions in the formative stages of a lymphocyte’s life. CurrOpin Immunol 2004; 16: 180–190.
19 Park JH, Yu Q, Erman B, Appelbaum JS, Montoya-Durango D, Grimes HL et al.Suppression of IL7Ralpha transcription by IL-7 and other prosurvival cytokines: a novelmechanism for maximizing IL-7-dependent T cell survival. Immunity 2004; 21:289–302.
20 Swainson L, Verhoeyen E, Cosset FL, Taylor N. IL-7R alpha gene expression is inverselycorrelated with cell cycle progression in IL-7-stimulated T lymphocytes. J Immunol2006; 176: 6702–6708.
21 Vranjkovic A, Crawley AM, Gee K, Kumar A, Angel JB. IL-7 decreases IL-7 receptoralpha (CD127) expression and induces the shedding of CD127 by human CD8þT cells. Int Immunol 2007; 19: 1329–1339.
22 Alves NL, van Leeuwen EM, Derks IA, van Lier RA. Differential regulation of humanIL-7 receptor alpha expression by IL-7 and TCR signaling. J Immunol 2008; 180:5201–5210.
23 Faller E, Kakal J, Kumar R, Macpherson P. IL-7 and the HIV Tat protein actsynergistically to down-regulate CD127 expression on CD8 T cells. Int Immunol2009; 21: 203–216.
24 Henriques CM, Rino J, Nibbs RJ, Graham GJ, Barata JT. IL-7 induces rapid clathrin-mediated internalization and JAK3-dependent degradation of IL-7Ralpha in T cells.Blood 2010; 115: 3269–3277.
25 Sasson SC, Zaunders JJ, Zanetti G, King EM, Merlin KM, Smith DE et al. Increasedplasma interleukin-7 level correlates with decreased CD127 and Increased CD132extracellular expression on T cell subsets in patients with HIV-1 infection. J Infect Dis2006; 193: 505–514.
26 Faller EM, Sugden SM, McVey MJ, Kakal JA, MacPherson PA. Soluble HIV Tat proteinremoves the IL-7 receptor alpha-chain from the surface of resting CD8 T cells andtargets it for degradation. J Immunol 2010; 185: 2854–2866.
27 Lee HC, Shibata H, Ogawa S, Maki K, Ikuta K. Transcriptional regulation of themouse IL-7 receptor alpha promoter by glucocorticoid receptor. J Immunol 2005; 174:7800–7806.
28 Mittelstadt PR, Ashwell JD. Disruption of glucocorticoid receptor exon 2 yields aligand-responsive C-terminal fragment that regulates gene expression. Mol Endocrinol2003; 17: 1534–1542.
29 Crawley AM, Vranjkovic A, Young C, Angel JB. Interleukin-4 downregulates CD127expression and activity on human thymocytes and mature CD8þ T cells. Eur JImmunol 2010; 40: 1396–1407.
30 Johnston JA, Bacon CM, Finbloom DS, Rees RC, Kaplan D, Shibuya K et al. Tyrosinephosphorylation and activation of STAT5, STAT3, and Janus kinases by interleukins 2and 15. Proc Natl Acad Sci USA 1995; 92: 8705–8709.
31 Demoulin JB, Van Snick J, Renauld JC. Interleukin-9 (IL-9) induces cell growth arrestassociated with sustained signal transducer and activator of transcription activationin lymphoma cells overexpressing the IL-9 receptor. Cell Growth Differ 2001; 12:169–174.
32 Goodman WA, Young AB, McCormick TS, Cooper KD, Levine AD. Stat3 phosphorylationmediates resistance of primary human T cells to regulatory T cell suppression.J Immunol 2011; 186: 3336–3345.
33 Murray PJ. The JAK-STAT signaling pathway: input and output integration. J Immunol2007; 178: 2623–2629.
34 Zhang B, Zhang Y, Niu L, Vella AT, Mittler RS. Dendritic cells and Stat3 are essentialfor CD137-induced CD8 T cell activation-induced cell death. J Immunol 2010; 184:4770–4778.
35 Hirata N, Yanagawa Y, Iwabuchi K, Onoe K. Selective regulation of interleukin-10production via Janus kinase pathway in murine conventional dendritic cells. CellImmunol 2009; 258: 9–17.
36 Kim HR, Hwang KA, Kim KC, Kang I. Down-regulation of IL-7Ralpha expression inhuman T cells via DNA methylation. J Immunol 2007; 178: 5473–5479.
37 Kim K, Khaled AR, Reynolds D, Young HA, Lee CK, Durum SK. Characterization of aninterleukin-7-dependent thymic cell line derived from a p53(�/�) mouse. J ImmunolMethods 2003; 274: 177–184.
38 Wofford JA, Wieman HL, Jacobs SR, Zhao Y, Rathmell JC. IL-7 promotes Glut1trafficking and glucose uptake via STAT5-mediated activation of Akt to support T-cellsurvival. Blood 2008; 111: 2101–2111.
39 Kaech SM, Tan JT, Wherry EJ, Konieczny BT, Surh CD, Ahmed R. Selective expressionof the interleukin 7 receptor identifies effector CD8 T cells that give rise to long-livedmemory cells. Nat Immunol 2003; 4: 1191–1198.
40 Prlic M, Lefrancois L, Jameson SC. Multiple choices: regulation of memory CD8 T cellgeneration and homeostasis by interleukin (IL)-7 and IL-15. J Exp Med 2002; 195:F49–F52.
41 Gregory SG, Schmidt S, Seth P, Oksenberg JR, Hart J, Prokop A et al. Interleukin 7receptor alpha chain (IL7R) shows allelic and functional association with multiplesclerosis. Nat Genet 2007; 39: 1083–1091.
42 Kreft KL, Verbraak E, Wierenga-Wolf AF, van Meurs M, Oostra BA, Laman JD et al. TheIL-7Ralpha pathway is quantitatively and functionally altered in cd8 t cells in multiplesclerosis. J Immunol 2012; 188: 1874–1883.
43 O’Doherty C, Alloza I, Rooney M, Vandenbroeck K. IL7RA polymorphisms and chronicinflammatory arthropathies. Tissue Antigen 2009; 74: 429–431.
44 Vudattu NK, Magalhaes I, Schmidt M, Seyfert-Margolis V, Maeurer MJ. Reducednumbers of IL-7 receptor (CD127) expressing immune cells and IL-7-signalingdefects in peripheral blood from patients with breast cancer. Int J Cancer 2007;121: 1512–1519.
45 van Leeuwen EM, de Bree GJ, Remmerswaal EB, Yong SL, Tesselaar K, ten Berge IJet al. IL-7 receptor alpha chain expression distinguishes functional subsets of virus-specific human CD8þ T cells. Blood 2005; 106: 2091–2098.
46 Wherry EJ, Day CL, Draenert R, Miller JD, Kiepiela P, Woodberry T et al. HIV-specificCD8 T cells express low levels of IL-7Ralpha: implications for HIV-specific T cellmemory. Virology 2006; 353: 366–373.
47 Sauce D, Larsen M, Leese AM, Millar D, Khan N, Hislop AD et al. IL-7R alphaversus CCR7 and CD45 as markers of virus-specific CD8þ T cell differentiation:contrasting pictures in blood and tonsillar lymphoid tissue. J Infect Dis 2007; 195:268–278.
48 Golden-Mason L, Burton Jr JR, Castelblanco N, Klarquist J, Benlloch S,Wang C et al. Loss of IL-7 receptor alpha-chain (CD127) expression inacute HCV infection associated with viral persistence. Hepatology 2006; 44:1098–1109.
IL-7 downregulates IL-7Ra expressionFM Ghazawi et al
157
Immunology and Cell Biology

49 Boutboul F, Puthier D, Appay V, Pelle O, Ait-Mohand H, Combadiere B et al.Modulation of interleukin-7 receptor expression characterizes differentiation of CD8T cells specific for HIV, EBV and CMV. Aids 2005; 19: 1981–1986.
50 Colle JH, Moreau JL, Fontanet A, Lambotte O, Joussemet M, Delfraissy JF et al. CD127expression and regulation are altered in the memory CD8 T cells of HIV-infectedpatients–reversal by highly active anti-retroviral therapy (HAART). Clin Exp Immunol2006; 143: 398–403.
51 Koesters SA, Alimonti JB, Wachihi C, Matu L, Anzala O, Kimani J et al. IL-7Ralphaexpression on CD4(þ ) T lymphocytes decreases with HIV disease progression andinversely correlates with immune activation. Eur J Immunol 2006; 36: 336–344.
52 MacPherson PA, Fex C, Sanchez-Dardon J, Hawley-Foss N, Angel JB. Interleukin-7receptor expression on CD8(þ ) T cells is reduced in HIV infection and partially
restored with effective antiretroviral therapy. J Acquir Immune Defic Syndr 2001; 28:454–457.
53 Mercier F, Boulassel MR, Yassine-Diab B, Tremblay C, Bernard NF, Sekaly RP et al.Persistent human immunodeficiency virus-1 antigenaemia affects the expression ofinterleukin-7Ralpha on central and effector memory CD4þ and CD8þ T cell subsets.Clin Exp Immunol 2008; 152: 72–80.
54 Paiardini M, Cervasi B, Albrecht H, Muthukumar A, Dunham R, Gordon S et al. Loss ofCD127 expression defines an expansion of effector CD8þ T cells in HIV-infectedindividuals. J Immunol 2005; 174: 2900–2909.
55 Rethi B, Fluur C, Atlas A, Krzyzowska M, Mowafi F, Grutzmeier S et al. Loss ofIL-7Ralpha is associated with CD4 T-cell depletion, high interleukin-7 levels and CD28down-regulation in HIV infected patients. Aids 2005; 19: 2077–2086.
IL-7 downregulates IL-7Ra expressionFM Ghazawi et al
158
Immunology and Cell Biology





























