Embed Size (px)
Citation preview

Genetics of barley grain size
Arnis Druka
IAEA Regional Training Course Phenotypical Characterization of Mutants 19-23 May 2013 Amman, Jordan

T h e o u t l i n e
1) Introduction – modelling grain yield from a geneticists point of view
2) Generation and analysis of the phenotype data matrix
3) Generation and genotyping of Recombinant Inbred Line (RIL) population
4) Linkage mapping using J/QTL
5) Known rice grain size genes – how good candidates are they?
6) What Bowman-backcross derived lines are good for?

Today
Enhancing gene prediction using Bowman backcross-derived lines
and barley-rice synteny
Thursday, May 23

Germplasm identification
STW trait
Changing gene action
Gene identification
How it works?
Genetic linkage mapping
Molecular genetics golden
promise

Germplasm – what’s available?
• ‘Immortal’ bi-parental populations (eg GPMx) • Multi-parental crosses • Random collections • Association panels • Original ‘off-type’ lines (induced and
spontaneous variants) • Backcross-derived lines • F2 populations • F1 seeds

Udda Lundqvist
• Generated large collection of induced barley mutant lines (~10,000)
• Performed extensive allelism testing

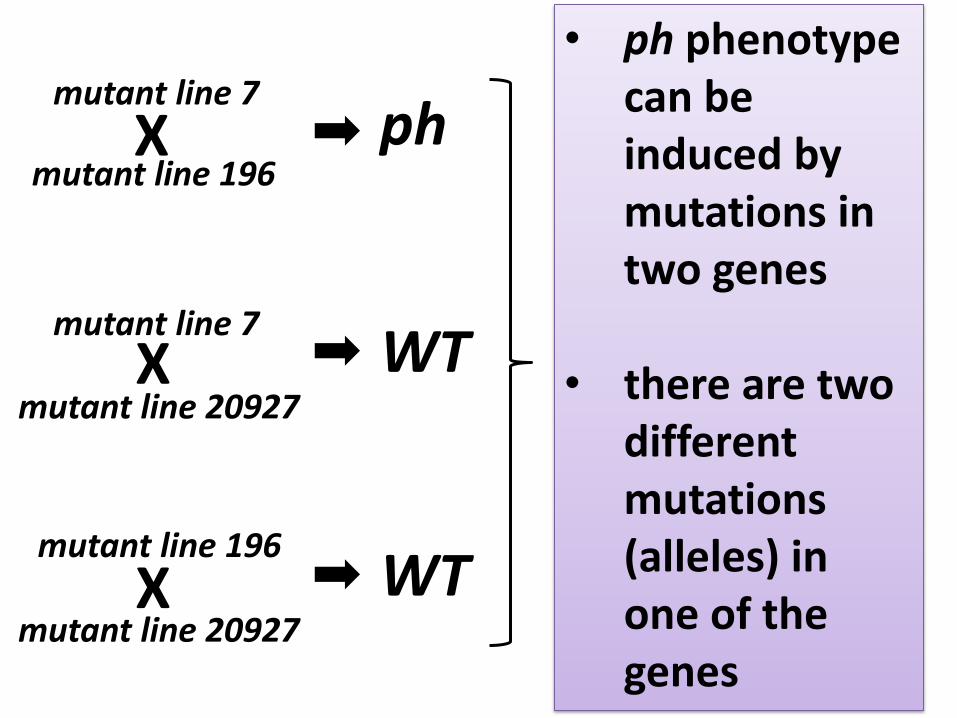
Mutagenesis experiment WT mutant line 196
mutant line 7, mutant line 196 and mutant line 20927 have the same phenotype ph
mutant line 7
mutant line 20927
What happened? • Three independent mutations in one gene? • Or, three different genes determine
phenotype ph?

To know which is which:
1. Cross each mutant line to every other
2. Plant F1 seed
3. Observe F1 plant Ph phenotype
4. Make sure that crosses worked by
genotyping F1 plants
Essential, but may not be straightforward
mutant line 196
mutant line 7
mutant line 20927
X
mutant line 7
mutant line 196
mutant line 20927
X
X
ph
WT
WT
• ph phenotype can be induced by mutations in two genes
• there are two different mutations (alleles) in one of the genes

• This is called allelism test
• Udda Lundqvist performed such allelism tests for
many phenotypes in many cases identifying 20-30
alleles (called allelic series)
• Induced allelic series are critical resource for barley
gene validation because
• Probability of occurring multiple deleterious
mutations in the same gene in natural
conditions is extremely low
• Phenotypes generally are stronger

Barley Genetics Newsletter
V26

Graingenes
http://wheat.pw.usda.gov/

Bowman backcross-derived
lines

Jerome Franckowiak • Collected and generated large number of mutant lines
including many Udda’s mutants
• Author of the cultivar Bowman
• Performed monumental backcrossing program
generating ~1000 Bowman backcross-derived lines
Hermitage Research Station QAAFI
Jerry Franckowiak North Dakota State
University, Fargo USA
Jerry Franckowiak Hermitage Research Station
Warwick, Queensland AUSTRALIA
J e r r y F r a n c k o w i a k , B o w m a n & B C L s
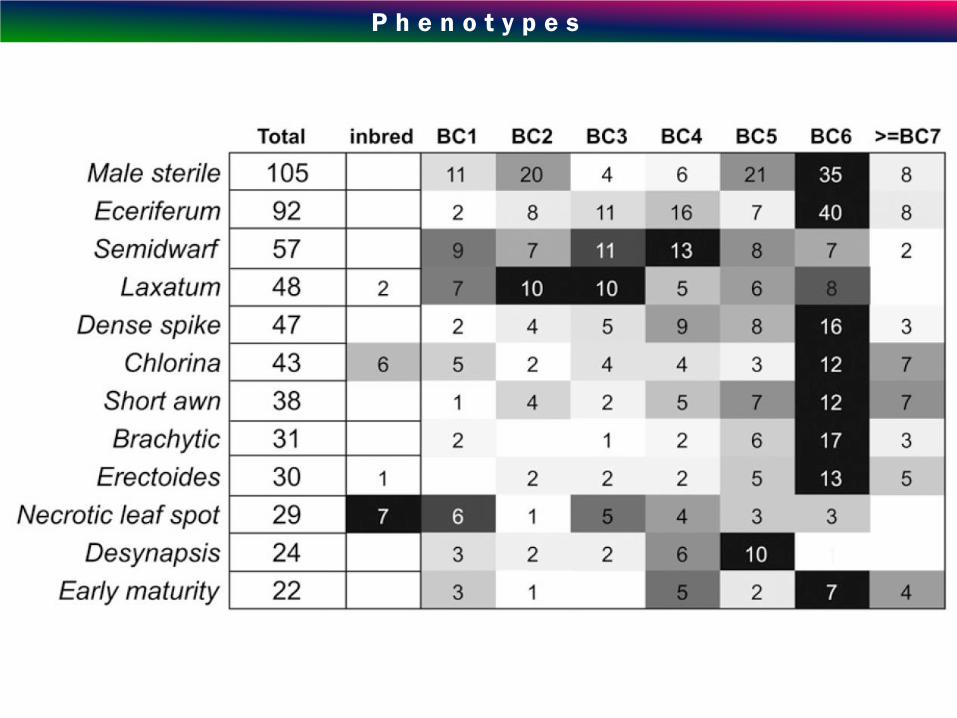
P h e n o t y p e s

OREGON WOLFE BARLEYS DHL population
Pat Hayes, Oregon State University, Corvallis, USA
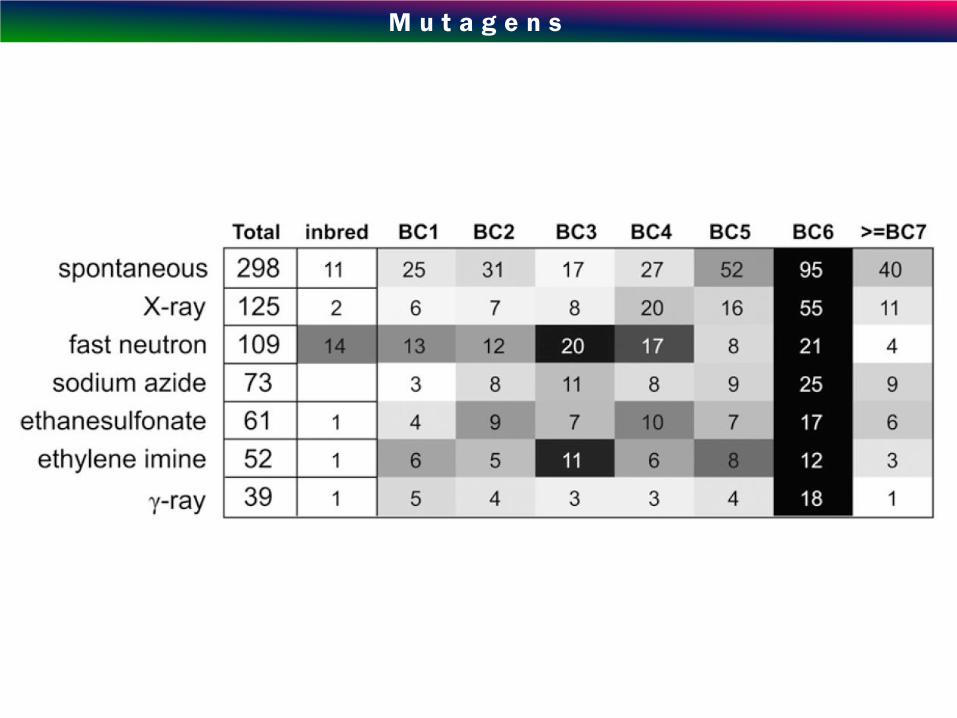
M u t a g e n s
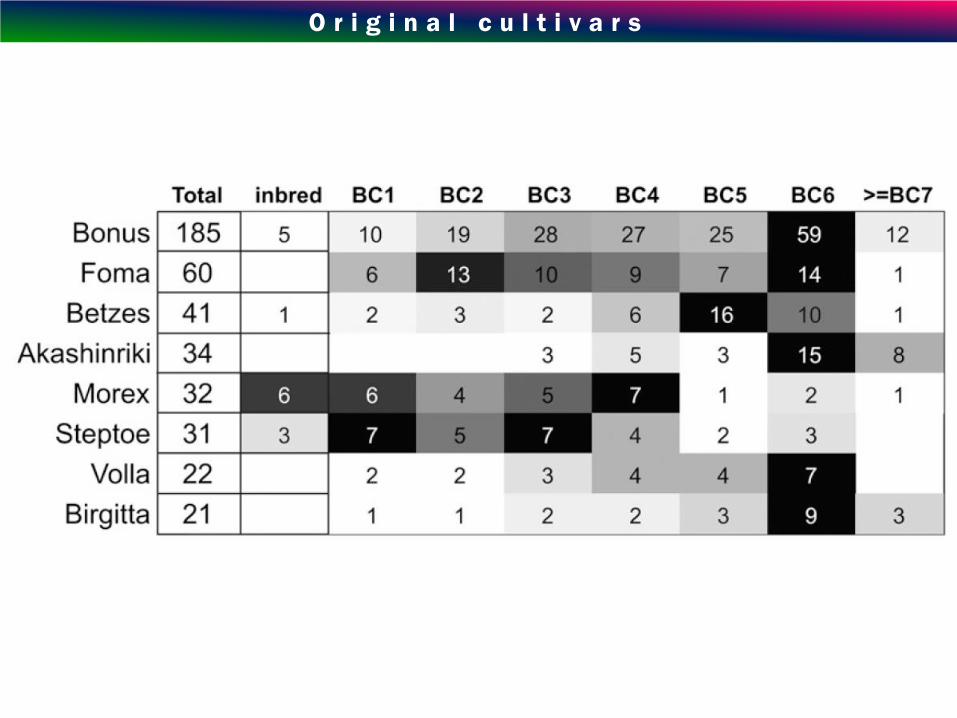
O r i g i n a l c u l t i v a r s
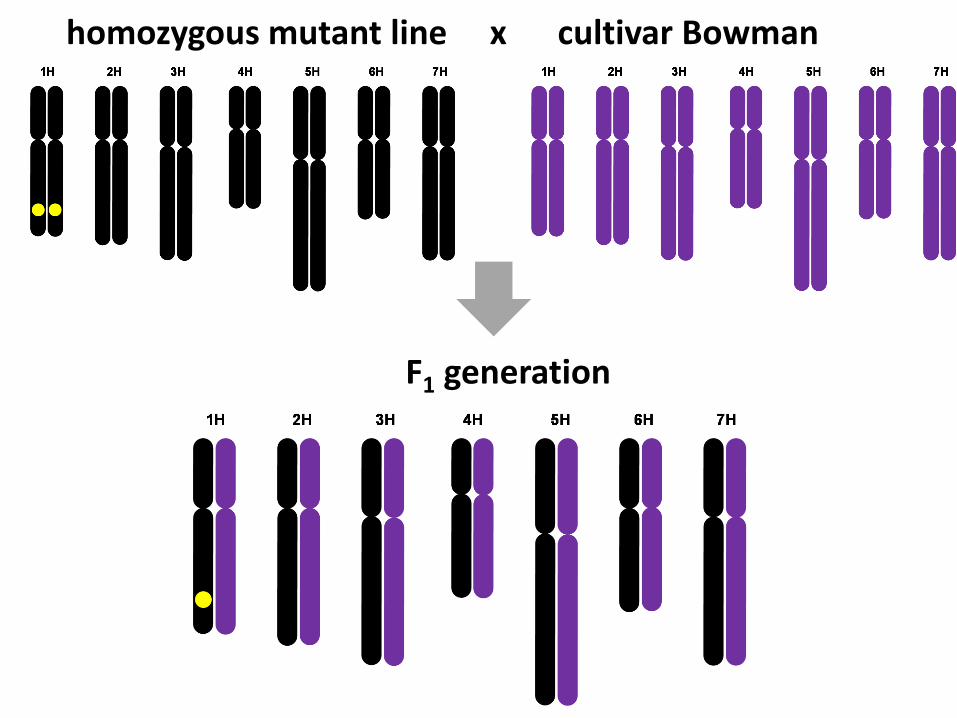
homozygous mutant line x cultivar Bowman
F1 generation
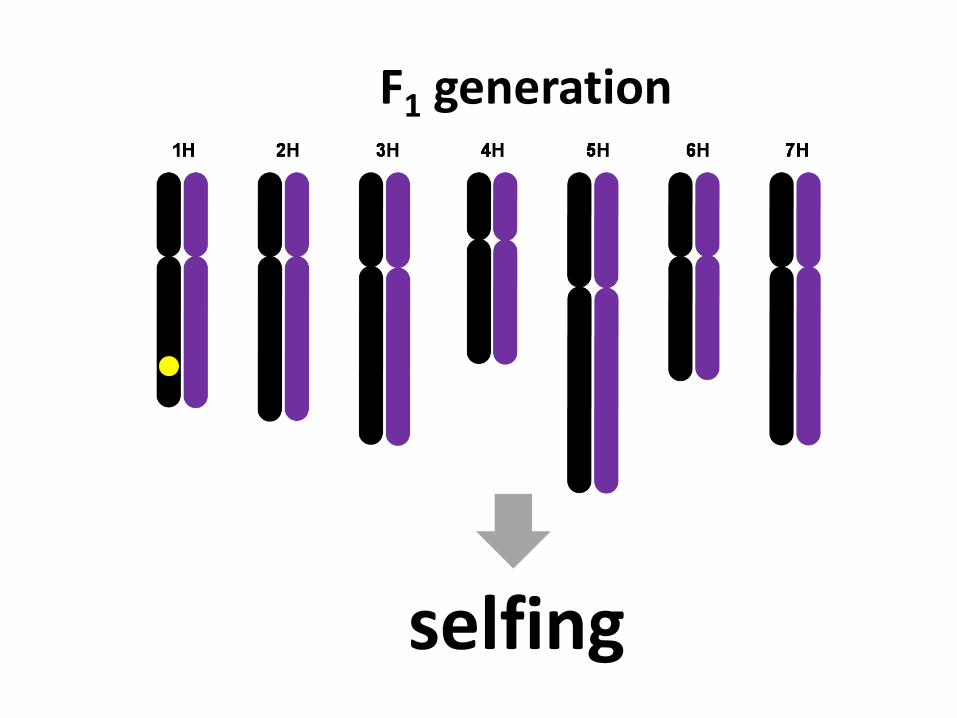
F1 generation
selfing

recombinant mutant line X
cultivar Bowman
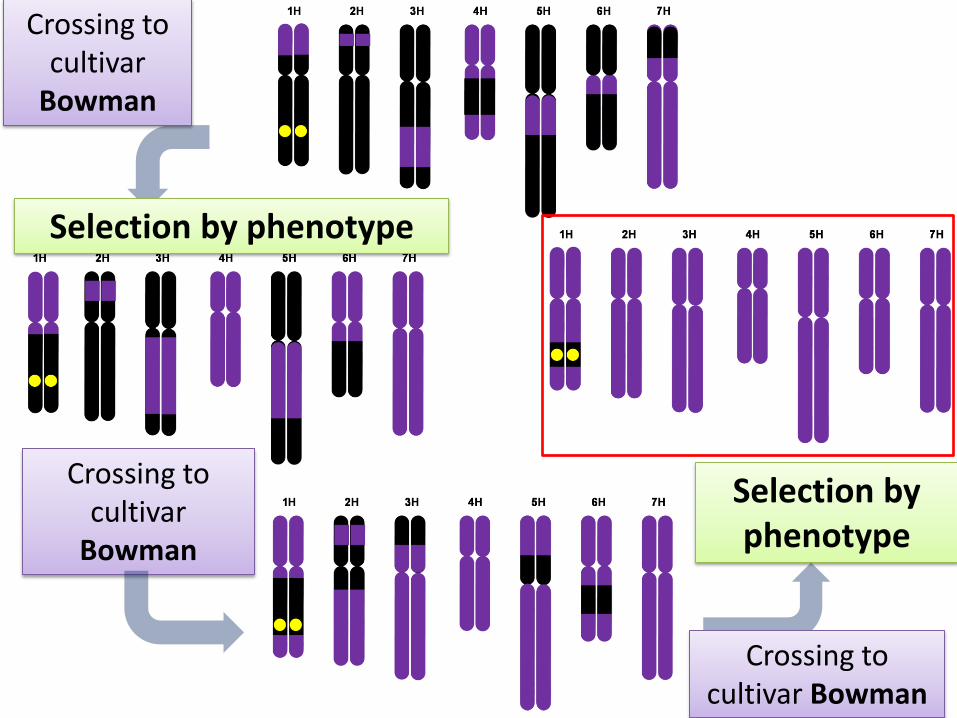
Crossing to cultivar
Bowman
Crossing to cultivar
Bowman
Crossing to cultivar Bowman
Selection by phenotype
Selection by phenotype
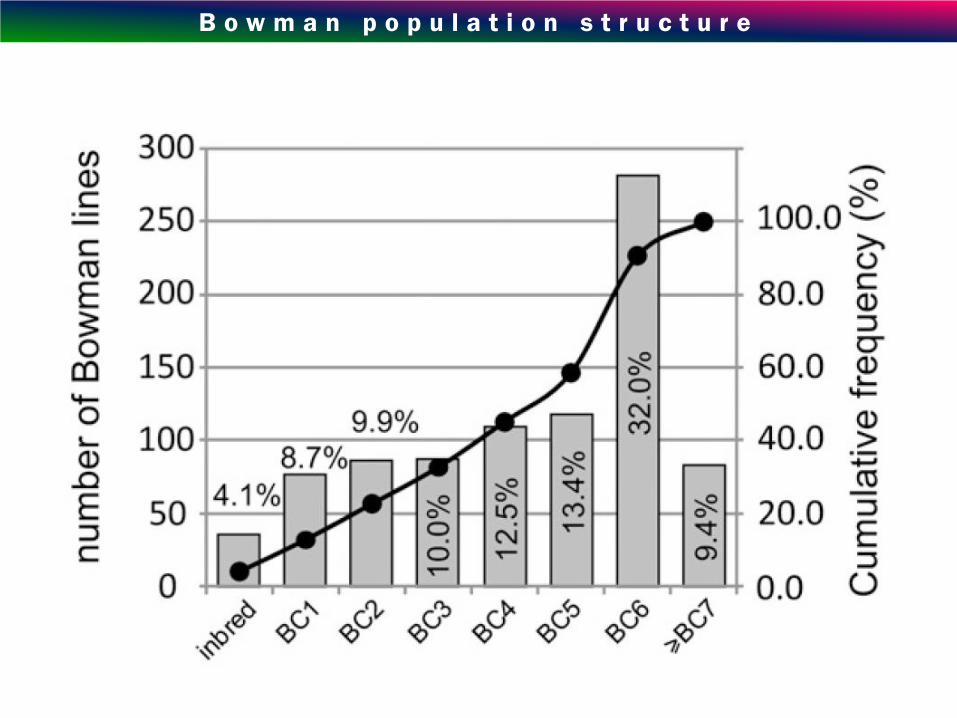
B o w m a n p o p u l a t i o n s t r u c t u r e

How do we know that after > 4 rounds of BC there is a single, small
introgression left?
?
BW999
Beadarray technology and the data sets
Bowman Cy3 signal
BW
99
9 C
y3 s
ign
al
Bowman Cy5 signal
BW
99
9 C
y5 s
ign
al
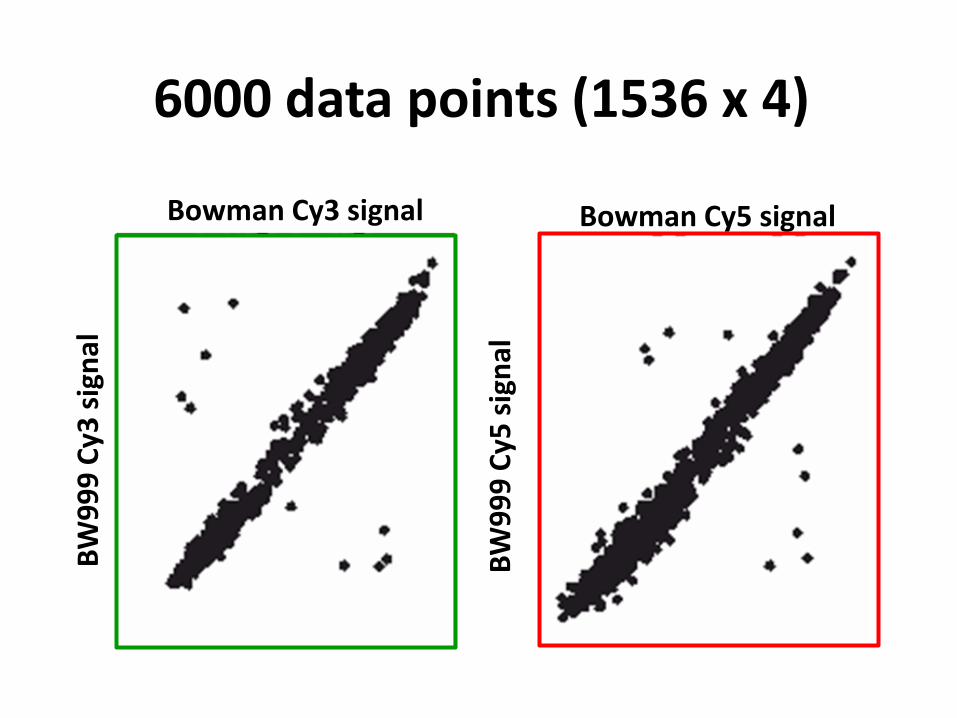
6000 data points (1536 x 4)
Bowman Cy3 signal
BW
99
9 C
y3 s
ign
al
Bowman Cy5 signal
BW
99
9 C
y5 s
ign
al
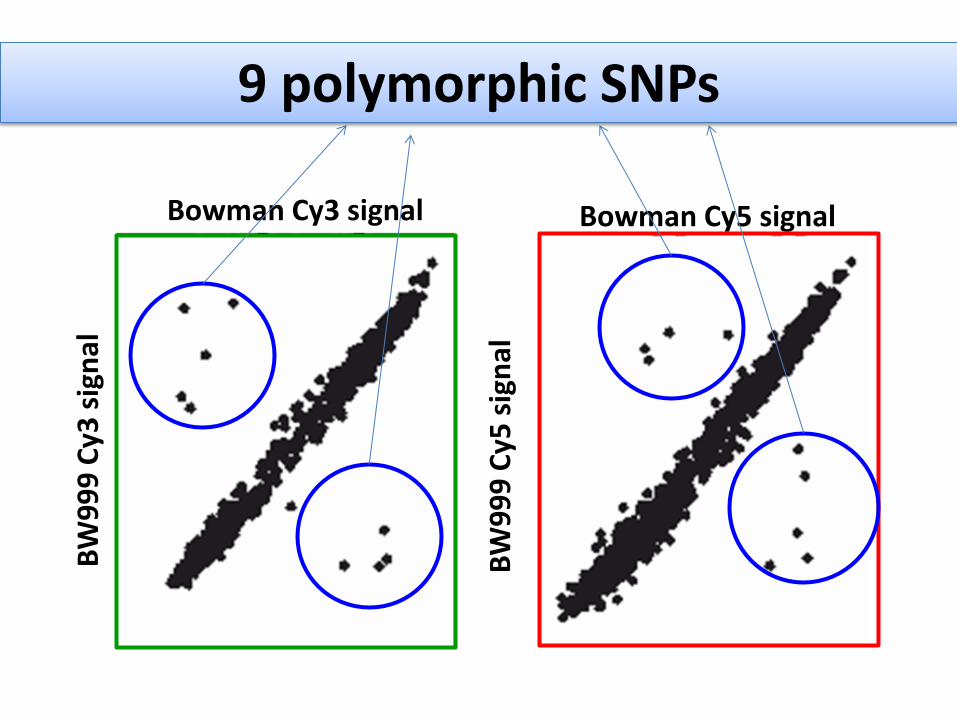
9 polymorphic SNPs

9 polymorphic SNPs
How they are spread across the genome?
SNP 47 SNP169 SNP 589 SNP1675 SNP 469 SNP 800 SNP BEE SNP 504
SNP 9
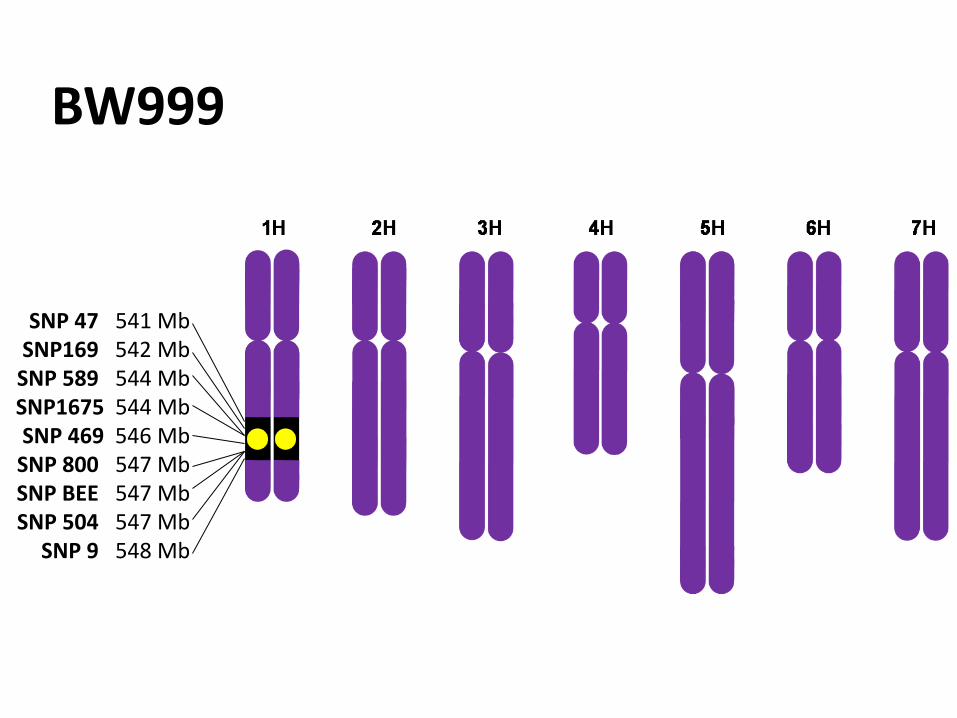
BW999
SNP 47 541 Mb SNP169 542 Mb
SNP 589 544 Mb SNP1675 544 Mb SNP 469 546 Mb
SNP 800 547 Mb SNP BEE 547 Mb SNP 504 547 Mb
SNP 9 548 Mb

SNP 47 541 Mb SNP169 542 Mb
SNP 589 544 Mb SNP1675 544 Mb SNP 469 546 Mb
SNP 800 547 Mb SNP BEE 547 Mb SNP 504 547 Mb
SNP 9 548 Mb
Introgression size
548 – 541 = 7 Mb
SNP positions

cM Mb 1H 133 581 2H 149 628 3H 155 565 4H 115 543 5H 169 560 6H 127 539 7H 140 601
total 988 4,016
number of genes 30,400 genome size (Mb) 5,300
genes/cM 30.76 genes/Mb 5.7 Mb/cM 5.4
chr

SNP 47 541 Mb SNP169 542 Mb
SNP 589 544 Mb SNP1675 544 Mb SNP 469 546 Mb
SNP 800 547 Mb SNP BEE 547 Mb SNP 504 547 Mb
SNP 9 548 Mb
Introgression size
548 – 541 = 7 Mb
Number of genes
7 x 5.7= 40
SNP positions

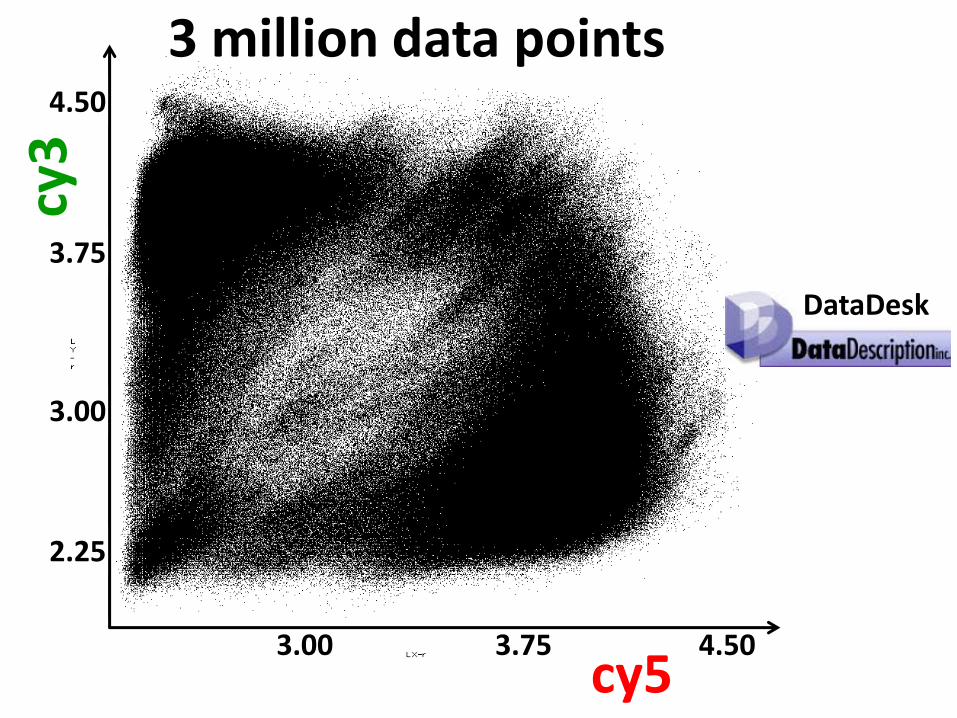
Bowman backcross-derived line genotyping experiment
976 Bowman lines 1536 SNPs each
2 signals each SNP
2,998,272 data points
4.50
3.75
3.00
2.25
3.00 3.75 4.50 cy5
cy3
3 million data points
DataDesk
Based on genotyping of 976 Bowman backcross-derived lines with 1536 known SNPs
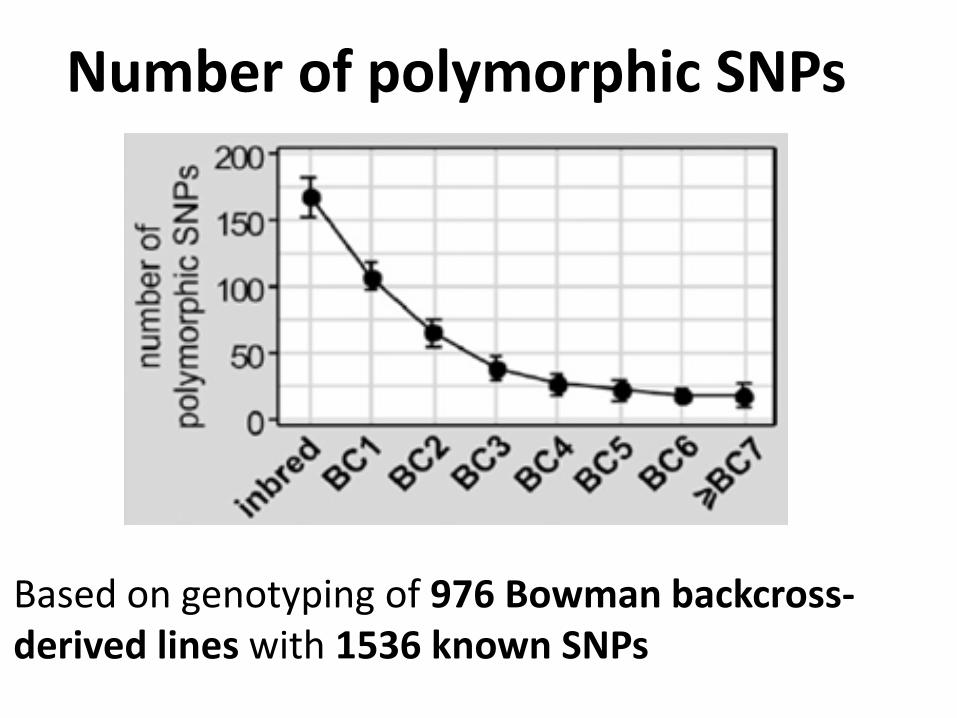
Number of polymorphic SNPs
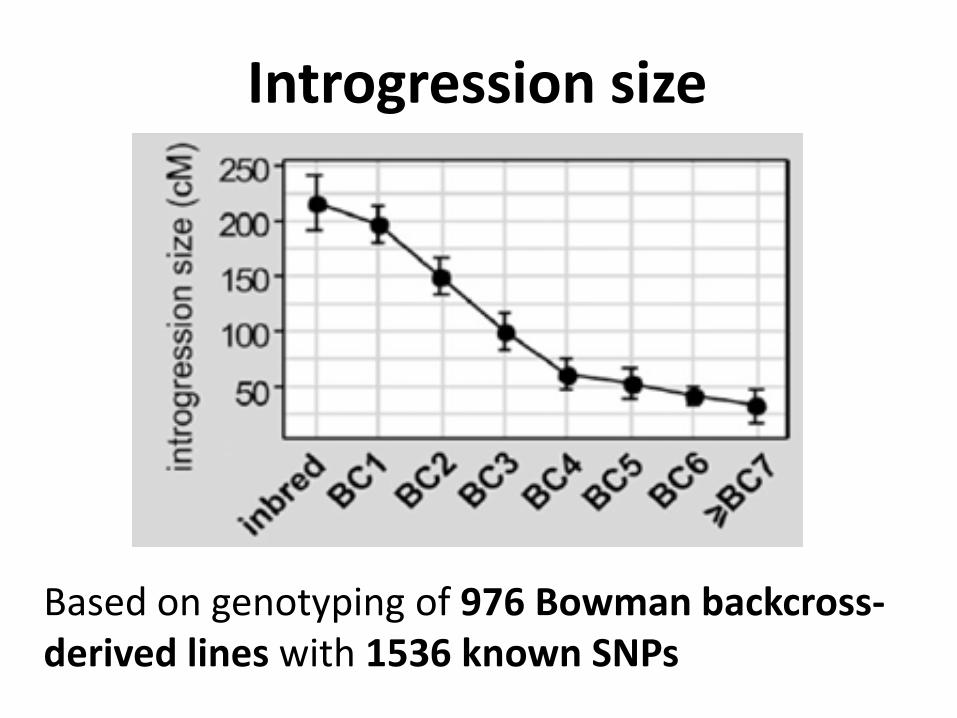
Based on genotyping of 976 Bowman backcross-derived lines with 1536 known SNPs
Introgression size

Based on genotyping of 976 Bowman backcross-derived lines with 1536 known SNPs
Number of introgressions

Barley – rice synteny

• Nucleotide sequences around SNPs are known
• SNPs come from barley gene sequences • Rice genome sequence and identities of
many genes are known • Barley and rice proteins have similar
amino acid sequences • Multiple chromosomal regions have
similar gene order in barley and rice
Why can synteny-based gene predictions work?
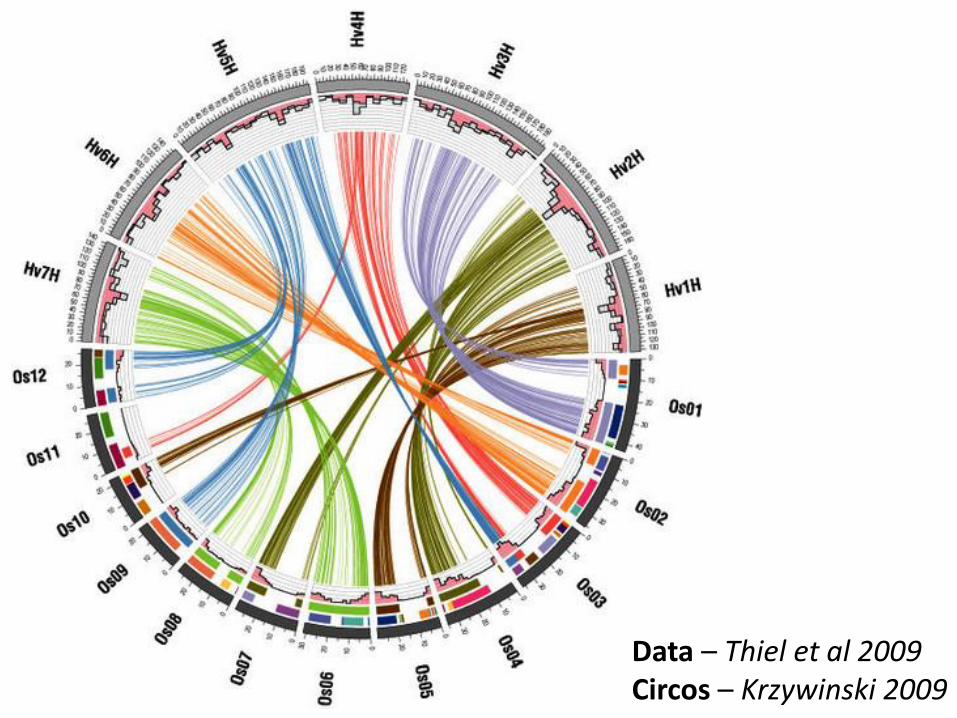
Data – Thiel et al 2009 Circos – Krzywinski 2009

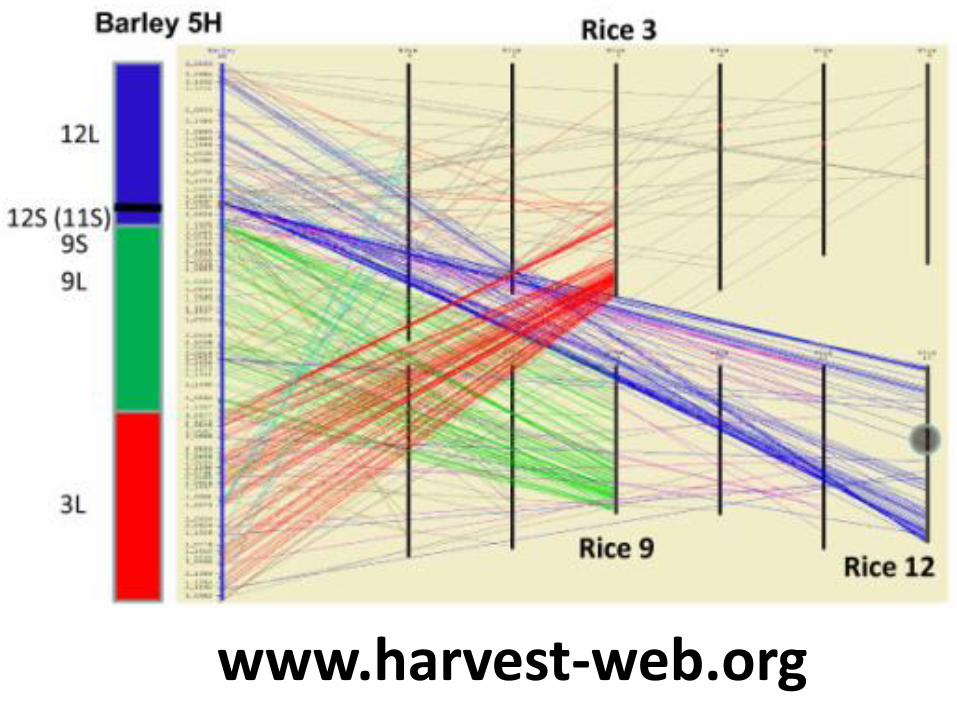
www.harvest-web.org

HARVEST
www.harvest-web.org
Tim Close University of California Riverside
USA
www.harvest-web.org

SNP 47 541 Mb SNP169 542 Mb
SNP 589 544 Mb SNP1675 544 Mb SNP 469 546 Mb
SNP 800 547 Mb SNP BEE 547 Mb SNP 504 547 Mb
SNP 9 548 Mb
SNP positions • SNPs come from
barley gene sequences
Use these sequences to identify rice gene
homologs
See if synteny can be established
If yes, see what’s interesting is in rice
The National Center for Biotechnology Information
NCBI Set up by US Government
http://www.ncbi.nlm.nih.gov/

Altschul SF, Gish W, Miller W, Myers EW, Lipman DJ (1990) Basic local alignment search tool. J Mol Biol 215: 403–410
BLAST - Basic Local Alignment Search Tool
David Lipman

http://rice.plantbiology.msu.edu

Kawahara, Y., de la Bastide, M., Hamilton J. P., Kanamori, H., McCombie, W. R., Ouyang, S., Schwartz, D. C., Tanaka, T., Wu, J., Zhou, S., Childs, K. L., Davidson, R. M., Lin, H., Quesada-Ocampo, L., Vaillancourt, B., Sakai, H., Lee, S. S., Kim, J., Numa, H., Itoh, T., Buell, C. R., Matsumoto, T. 2013 Improvement of the Oryza sativa Nipponbare reference genome using next generation sequence and optical map data. Rice 6:4.
Robin Buell

Considerations using synteny approach
• Not all genes are in syntenuous
positions
• Phenotype equivalency problem
• Missing and/or diverged genes

(Educated) guessing approach to clone gene underlying
mat-a phenotype
Mats Hansson & Team Barley
Carlsberg Laboratory
Application: cloning eam8 (or mat-a) gene

difference in heading date (5-10 days)
m a t p h e n o t y p e i n b a r l e y
wild type mutant
Image from: Nat Genet. 2012 Dec;44(12):1388-92.

Cloning eam8 (or mat-a) gene using backcross-derived lines
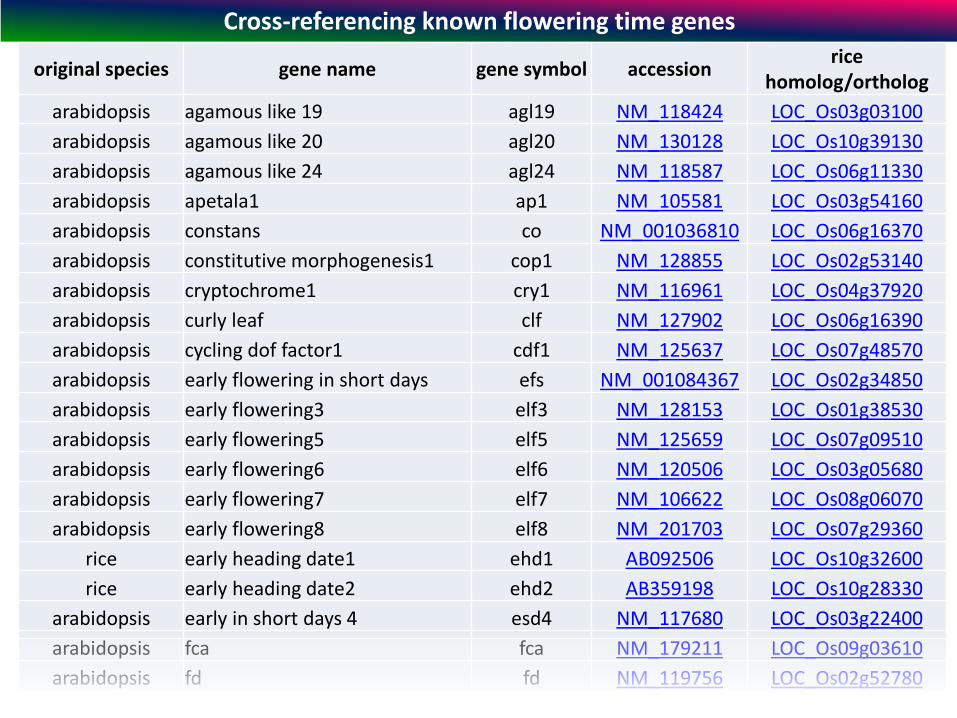
original species gene name gene symbol accession rice
homolog/ortholog
arabidopsis agamous like 19 agl19 NM_118424 LOC_Os03g03100
arabidopsis agamous like 20 agl20 NM_130128 LOC_Os10g39130
arabidopsis agamous like 24 agl24 NM_118587 LOC_Os06g11330
arabidopsis apetala1 ap1 NM_105581 LOC_Os03g54160
arabidopsis constans co NM_001036810 LOC_Os06g16370
arabidopsis constitutive morphogenesis1 cop1 NM_128855 LOC_Os02g53140
arabidopsis cryptochrome1 cry1 NM_116961 LOC_Os04g37920
arabidopsis curly leaf clf NM_127902 LOC_Os06g16390
arabidopsis cycling dof factor1 cdf1 NM_125637 LOC_Os07g48570
arabidopsis early flowering in short days efs NM_001084367 LOC_Os02g34850
arabidopsis early flowering3 elf3 NM_128153 LOC_Os01g38530
arabidopsis early flowering5 elf5 NM_125659 LOC_Os07g09510
arabidopsis early flowering6 elf6 NM_120506 LOC_Os03g05680
arabidopsis early flowering7 elf7 NM_106622 LOC_Os08g06070
arabidopsis early flowering8 elf8 NM_201703 LOC_Os07g29360
rice early heading date1 ehd1 AB092506 LOC_Os10g32600
rice early heading date2 ehd2 AB359198 LOC_Os10g28330
arabidopsis early in short days 4 esd4 NM_117680 LOC_Os03g22400
arabidopsis fca fca NM_179211 LOC_Os09g03610
arabidopsis fd fd NM_119756 LOC_Os02g52780
Cross-referencing known flowering time genes

marker id chr pos (cM) LOC_rice chr rice 5'-end (Mb) annotation_rice
ABC11085-1-1-168 1H 132.5 LOC_Os05g50360 Os05 28,771,533 anaphase-promoting complex subunit 10
3_0803 1H 132.5 LOC_Os07g04220 Os07 1,849,699 wound and phytochrome signaling receptor like kinase
6473-811 1H 134.0 LOC_Os05g50480 Os05 28,856,869 expressed protein
ConsensusGBS0554-4 1H 135.6 LOC_Os01g14590 Os01 8,174,691 pathogen-related protein
11454-414 1H 135.6 LOC_Os01g42960 Os01 24,787,885 electron transporter
ConsensusGBS0450-1 1H 135.6 LOC_Os01g69970 Os01 40,794,608 periodic tryptophan protein 1
9022-543 1H 135.6 LOC_Os05g50800 Os05 29,054,191 protein ABIL1
ABC05061-1-1-159 1H 135.6 LOC_Os05g50840 Os05 29,078,396 Grave disease carrier
3_0277 1H 135.6 LOC_Os05g50930 Os05 29,141,766 RNA polymerase sigma factor rpoD
3_0517 1H 135.6 LOC_Os07g08430 Os07 4,331,942 indole-3-glycerol phosphate lyase
3653-519 1H 135.6 LOC_Os09g36680 Os09 21,157,464 ribonuclease 3 precursor
3671-59 1H 135.6 LOC_Os09g36700 Os09 21,167,169 extracellular ribonuclease LE precursor
3639-969 1H 136.3 LOC_Os05g51180 Os05 29,272,704 plasminogen activator inhibitor 1 RNA-binding protein
4927-1340 1H 137.8 LOC_Os05g51450 Os05 29,416,207 endopeptidase Clp
5316-739 1H 138.3 LOC_Os05g51470 Os05 29,423,748 cupin, RmlC-type
4057-2114 1H 138.3 LOC_Os05g51480 Os05 29,428,583 DNA damage binding protein 1a
5222-919 1H 138.3 LOC_Os05g51500 Os05 29,451,410 eukaryotic translation initiation factor 5B
3_0231 1H 138.3 LOC_Os05g51540 Os05 29,470,870 expressed protein
3246-1135 1H 138.9 LOC_Os05g51530 Os05 29,468,418 vacuolar ATP synthase subunit C
elf3
ft
spa1
elf3 LOC_Os01g38530
21,638,883
spa1 LOC_Os05g49590
28,382,352
ft LOC_Os01g11940
6,493,516
Identifying eam8 (or mat-a) candidate genes using barley-rice synteny

Causal gene – the one with multiple
deleterious sequence variants
In this case it turned
out to be elf3
elf3 LOC_Os01g38530 21,638,883
spa1 LOC_Os05g49590 28,382,352
ft LOC_Os01g11940 6,493,516
Re-sequence each candidate gene from
some of the
87 available mat-a mutant
lines
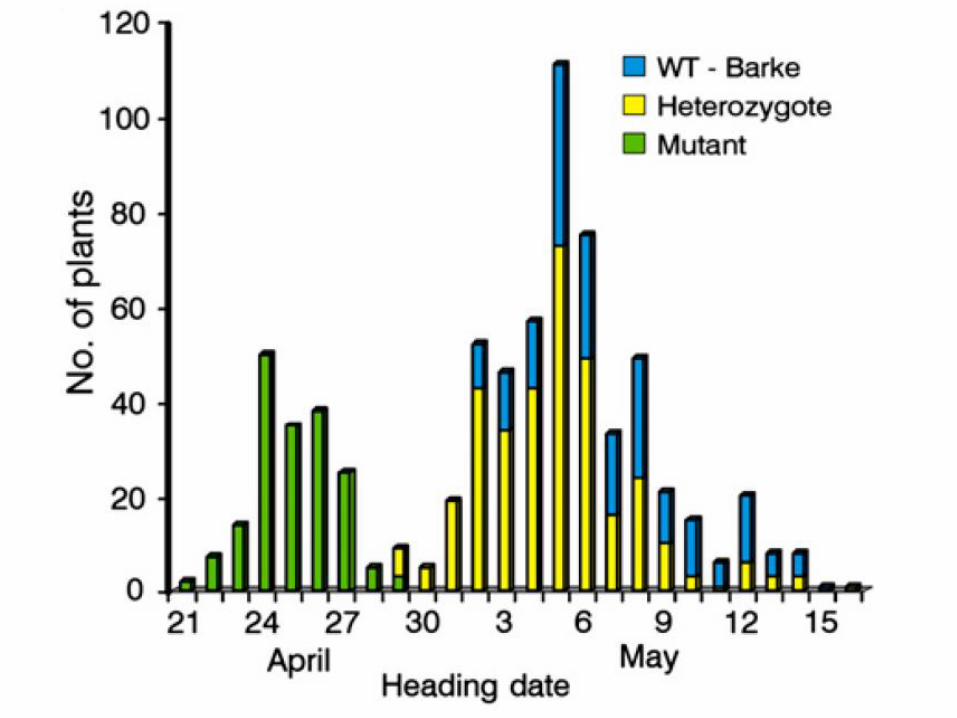
Validation of elf3 as eam8 (or mat-a) gene

Udda’s allelic series
Validation of elf3 as eam8 (or mat-a) gene using allelic series

BA R C O D E c r o s s e s
David Harrap KWS UK Ltd
F1 seeds &
F2 families

R e f e r e n c e

THANK YOU!
All presentations and data sets are available for downloading from here:
http://www.ilze-arnis.net/jordan_2013


![Genetic Dissection of Barley MorphologyGenetic Dissection of Barley Morphology and Development1[W][OA] Arnis Druka, Jerome Franckowiak, Udda Lundqvist,Nicola Bonar, Jill Alexander,](https://img.dokumen.tips/doc/110x75/5e3ea50ec4f01c37bf32b85e/genetic-dissection-of-barley-genetic-dissection-of-barley-morphology-and-development1woa.jpg)


























