Embed Size (px)
DESCRIPTION
U TADEO LOZANO
Citation preview

1
2
3
4Q14Q15
5
6789
1 0
1112131415
1617181920
37
3839
40
41
42
43
44
45
46
Gene xxx (2014) xxx–xxx
Q16
GENE-40122; No. of pages: 10; 4C:
Contents lists available at ScienceDirect
Gene
j ourna l homepage: www.e lsev ie r .com/ locate /gene
Molecular characterization and developmental expression of vitellogeninin the oriental river prawn Macrobrachium nipponense and the effects ofRNA interference and eyestalk ablation on ovarian maturation
OO
F
Hongkun Bai a, Hui Qiao b, Fajun Li a,c, Hongtuo Fu a,b,⁎, Shengming Sun b, Wenyi Zhang b, Shubo Jin a,b,Yongsheng Gong a, Sufei Jiang b, Yiwei Xiong b
a Wuxi Fisheries College, Nanjing Agricultural University, Wuxi 214081, PR Chinab Key Laboratory of Freshwater Fisheries and Germplasm Resources Utilization, Ministry of Agriculture, Freshwater Fisheries Research Center, Chinese Academy of Fishery Sciences, Wuxi 214081,PR Chinac Weifang University of Science and Technology, Shouguang 262700, PR China
UN
R
Abbreviations:M. nipponense,MacrobrachiumnipponenVgR, vitellogenin receptor; GIH, gonad-inhibiting hormnipponense vitellogenin; qPCR, quantitative real-time PdsRNA, double-strandedRNA; ELISA, enzyme-linked immuterference;O, ovary; B, brain; L, hepatopancreas; Fx, hemoabdominal ganglion; CS, cleavage stage; BS, blastula stage;stage; PS, protozoea stage; ZS, zoea stage; L1, the first dayday larva after hatching; L10, the tenth day larva after hatafter hatching; P10, the tenth day post-larvae aftermetampost-larvae aftermetamorphosis; P30, the thirty day post-⁎ Corresponding author at: Wuxi Fisheries College, N
Wuxi 214081, Jiangsu Province, PR China.E-mail address: [email protected] (H. Fu).
http://dx.doi.org/10.1016/j.gene.2014.12.0080378-1119/© 2014 Published by Elsevier B.V.
Please cite this article as: Bai, H., et al., MolMacrobrachium nipponense and the effects o
Pa b s t r a c t
a r t i c l e i n f o21
22
23
24
25
26
27
28
29
30
Article history:Received 2 August 2014Received in revised form 17 November 2014Accepted 5 December 2014Available online xxxx
Keywords:Macrobrachium nipponenseVitellogeninEyestalk ablationRNA interference
31
32
33
34
35
36
RECTED Vitellogenin (Vg) is the precursor of yolk protein, which functions as a nutritive resource that is important forembryonic growth and gonad development. In this study, the cDNA encoding the Vg gene from the orientalriver prawn Macrobrachium nipponense was cloned using expressed sequence tag (EST) analysis and the rapidamplification of cDNA ends (RACE) approach. The transcript encoded 2536 amino acids with an estimated mo-lecular mass of 286.810 kDa. Quantitative real-time PCR indicated high expression of Mn-Vg in the femaleovary, hemocytes, and hepatopancreas. As ovaries developed, the expression level of Mn-Vg increased in boththe hepatopancreas and ovary. In the hepatopancreas, the expression level rose more slowly at the early stageof vitellogenesis and reached the peak more rapidly compared to the expression pattern in ovary. The observedchanges inMn-Vg expression level at different development stages suggest the role of nutrient source in embry-onic and larval development. Eyestalk ablation caused the Mn-Vg expression level to increase significantly com-pared to eyestalk-intact groups during the ovary development stages. Ablation accelerated ovary maturation byremoving hormone inhibition of Mn-Vg in the hepatopancreas and ovary. In adult females, Mn-Vg dsRNA injec-tion resulted in decreased expression of Mn-Vg in both the hepatopancreas and ovary, and two injection treat-ment dramatically delayed ovary maturation. Vg RNA interference down-regulated the vitellogenin receptor(VgR) expression level in the ovary, which illustrates the close relationship between Vg and VgR in the processof vitellogenesis.
© 2014 Published by Elsevier B.V.
R
47
48
49
50
51
52
CO1. Introduction
The oriental river prawn Macrobrachium nipponense (Crustacea;Decapoda; Palaemonidae) is an important commercial prawn speciesthat is widely distributed in freshwater areas of China and other Asiancountries. The total fishing production reaches 230,248 t per year in
53
54
55
56
57
58
59
60
61
62
63
64
se; Vg, vitellogenin;Vn, vitellin;one; Mn-Vg, Macrobrachium
CR; GSI, gonadosomatic index;nosorbent assay; RNAi, RNA in-lymph; H, heart; E, eyestalk; AG,GS, gastrula stage; NS, naupliuslarva after hatching; L5, the fifthching; L15, the fifteen day larvaorphosis; P20, the twenty day
larvae aftermetamorphosisanjing Agricultural University,
ecular characterization and df RNA ..., Gene (2014), http://
China (Bureau of Fishery, 2011), with an annual production value ofmore than 100 million RMB. As the scale of production expanded, “sex-ual precocity” began to appear. This term refers to earlymale and femalegonad development, which leads to excessive propagation and over-population; in theM. nipponense population this occurs especially in au-tumn. Precocity results in the coexistence of multiple generations,intensive breeding density, and lack of oxygen, which can lead to shortlife span and low market value of the product. Thus, this phenomenonis restricting the sustainable development ofM. nipponense. Understand-ing the reproductive process and the mechanisms that regulate ovarianmaturation in M. nipponense is crucial to improving production of thiscommercially important species.
Vitellogenin (Vg), which is the precursor of vitellin (Vn), is synthe-sized by female shrimp during gonad maturation. In the mature femaleprawn, gonad maturity depends on the rapid synthesis and accumula-tion of Vg in the oocytes during the breeding season (Wilder et al.,2010). In many oviparous vertebrate and invertebrate animals, Vn pro-vides the substrate and energy for embryonic and ovarian development
evelopmental expression of vitellogenin in the oriental river prawndx.doi.org/10.1016/j.gene.2014.12.008

T
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
134
135
136
137
138
139
140
141
142
143
144
145
146
147
148
149
150
151
152
153
154
155
156
157
158
159
160
161
162
163
164
165
166
t1:1Table 1t1:2Q9Primers used for cDNA clone.
t1:3Q10Primer name Sequence (5′ → 3′) Purpose
t1:4Mn-Vg-R1 ATTACAGGTGTGCAGAGTTCCCTC Primer for 5′-RACEt1:5Mn-Vg-R2 AAGTACCCTACCTGAACCACCT primer for 5′-RACEt1:65′-RACE outer CATGGCTACATGCTGACAGCCTA Primer for 5′-RACEt1:75′-RACE inner CGCGGATCCACAGCCTACTGATGA
TCAGTCGATGPrimer for 5′-RACE
t1:83′-RACE outer TACCGTCGTTCCACTAGTGATTT Primer for 3′-RACEt1:93′-RACE inner CGCGGATCCTCCACTAGTGATTTCAC
TATAGGPrimer for 3′-RACE
t1:10Mn-Vg-F1 TCTTGTTAACTGGATCGTCCACG Primer for 3′-RACEt1:11Mn-Vg-F2 AAGCTCTCTTCGTACCTGTTCAG Primer for 3′-RACEt1:12VG-A-F1 TCTGGCGACAGCCTCAGCTGGT Middle segment At1:13VG-A-R1 TTGATGACAGTGAACGTTCCTGA Middle segment At1:14VG-B-F1 CTGGAGCAGTCAAGGTTATGGT Middle segment Bt1:15VG-B-R1 TACAGAGCACACGATTCCAGAC Middle segment Bt1:16VG-C-F1 GCCAGAGAAAATGGAGTTGGTG Middle segment Ct1:17VG-C-R2 TTGGTACTGAGAGCTTCCTTGG Middle segment Ct1:18VG-D-F1 GTCAGGCGAAACATCACAAGTC Middle segment Dt1:19VG-D-R1 CCTGTGACCTTCTGTTCCTCTC Middle segment Dt1:20VG-E-F1 CTGTTGCTTGATGTCACCCTCTC Middle segment Et1:21VG-E-R1 ACCGTGCATTATGGTGGCTTGA Middle segment E
2 H. Bai et al. / Gene xxx (2014) xxx–xxx
UNCO
RREC
(e.g., carbohydrates, amino acids, lipids, vitamins, phosphorus, sulfur,and trace elements) (Matozzo et al., 2008). The complete cDNA se-quence encoding Vg has been cloned for many species of decapod crus-tacean. In addition, the molecular characteristics and the regulatorymechanism of Vg have been widely studied (Tsutsui et al., 2000; Yanget al., 2000; Raviv et al., 2006; Okumura et al., 2007; Jia et al., 2013).However, molecular studies of M. nipponense Vg (Mn-Vg) are neededto better understand themechanisms involved in the reproductive pro-cess of this species.
In crustaceans, the site and the process of Vg synthesis are still con-troversial. Mainly have two kinds: extra-ovarian sources, namely by theorgan beyond ovary synthesis precursor, Vg was considered to be takeninto the developing oocytes from the hemolymph by the vitellogeninreceptor (VgR) via receptor-mediated endocytosis. The mechanismsfor endocytotic internalization of Vg have been well studied in certainoviparous vertebrates (Schneider, 1992) and insects (Sappington andRaikhel, 1998), but such studies in crustaceans are limited. Vg synthesisalsomay be endogenous (i.e., auto-synthesis), whereby the oocyte itselfproduces Vgwith participation from relevant organelles. For example, theovary was found to be the site of Vg synthesis in Penaeus semisulcatus(Browdy et al., 1990) and Callinectes sapidus (Lee and Watson, 1995).However, the Vg gene was uniquely expressed in the hepatopancreas ofMacrobrachium rosenbergii (Yang et al., 2000) and Pandalus hypsinotus(Tsutsui et al., 2004). Further studies indicated that both the ovary andhepatopancreas were the Vg synthesis sites in Marsupenaeus japonicus(Okumura et al., 2007), Fenneropenaeus merguiensis (Phiriyangkul et al.,2007), Litopenaeus vannamei (Raviv et al., 2006), Penaeus japonicus(Tsutsui et al., 2000), Metapenaeus ensis and Penaeus monodon (Tiuet al., 2006a, 2006b).
In decapods, vitellogenesis is hormonally regulated, and it can beinhibited by the occurrence of hormones in the neurosecretory cells ofthe X-organ/sinus gland. For example, gonad-inhibiting hormone(GIH), which is synthesized in the X-organ/sinus complex, is thoughtto play an inhibitory role for the initiation of vitellogenesis in theovary (De Kleijn et al., 1994; De Kleijn et al., 1998; Gu et al., 2002).Adiyodi and Adiyodi (1970) reported that eyestalk ablation removedthe inhibition of neuropeptides and accelerated the accumulation ofVg. In addition, Jayasankar et al. (2002) found that eyestalk ablationincreased Vg synthesis in the hepatopancreas of the giant freshwaterprawnM. rosenbergii. In P. japonicus, VgmRNA transcriptsweremeasuredboth in the hepatopancreas and the ovary in normal and eyestalk-ablatedadult shrimp. An obvious increment of mRNA levels was revealed in theovary, whereas mRNA levels were negligible in the hepatopancreas(Tsutsui et al., 2005). Overall, existing data indicate that the mechanismsfor hormonal regulation of Vg synthesis vary among crustaceans. Thus,molecular characterization and functional studies of Vg are critical to un-derstand the reproductive mechanisms in M. nipponense. Such informa-tion can be used to improve aquaculture production in practice.
In this study, we cloned the cDNA encoding the Vg gene fromM. nipponense (Mn-Vg) and conducted structural and phylogeneticanalyses. The expression profiles of different tissues and developmentstages (embryo and larvae) were determined using quantitative real-time PCR (qPCR). qPCR was also used to evaluate the effects of eyestalkablation to gain a better understanding of the hormonal regulationmechanism involved in Vg synthesis. RNAi technology was firstly ap-plied to investigate the expression pattern of Vg in ovary cycles. The re-sults of this study should be helpful for developing methods to copewith the problem of sexual precocity in the aquaculture setting.
2. Materials and methods
2.1. Experiment animal
Adult healthy M. nipponense were obtained from Tai lake in Wuxi,China (120°13′44″E, 31°28′22″N). The body weight of the female/maleprawns ranged from 1.26 to 4.25 g. Individuals, feed with paludina
Please cite this article as: Bai, H., et al., Molecular characterization and dMacrobrachium nipponense and the effects of RNA ..., Gene (2014), http://
ED P
RO
OF
twice per day, were acclimatized in a recirculating water aquariumsystem filled with aerated freshwater (25–28 °C) before tissues andembryos were collected. A variety of tissues including: ovary, heart, he-patopancreas, muscle, hemocytes, gill, eyestalk, gut and brain were dis-sected out from mature female/male prawn. As to the collection ofhemocytes, we drawed hemolymph from the body cavity, centrifugedit at 12000 rpm for 10min, removed the supernatant, and then collectedthe hemocyte precipitate to extract RNA. Ovarian maturity was classi-fied into six stages according to gonadosomatic index (GSI), color, exter-nal morphology and histological feature: Proliferation (stage I, GSI =0.85 ± 0.46), fusion nucleolus (stage II, GSI = 2.54 ± 1.28), oil globules(stage III, GSI=4.67±0.98), yolk granule (stage IV, GSI=7.55±0.40),maturation (stage V, GSI = 9.85 ± 2.74), and paracmasis (stage VI,GSI = 1.24 ± 0.62) (Wu et al., 2009). Each developmental stage of lar-vae was assessed by the criteria of Chen et al. (2012) and Kang et al.(1996). These samples were maintained in RNA protect liquid (Takara)until total RNA extraction.
2.2. The isolation of RNA and the synthesis of cDNA
Total RNA was extracted from mixed tissues using RNAiso Plus Re-agent (TaKaRa, Japan). RNA samples were treated with DNase I toremove contaminated genomic DNA for further experiments. Firststrand cDNA was synthesized from 3 μg total RNA using Reverse Tran-scriptase M-MLV Kit (TaKaRa, Japan).
2.3. Cloning and sequence analysis of Mn-Vg
Initially, the partial fragment of Mn-Vg cDNA was obtained fromnormalized cDNA library of ovary and testis (article is in press) in ourlab. Vg complete sequence was cloned by the method of rapid amplifi-cation of the cDNA ends (RACE). The 3′-RACE and 5′-RACE terminalfragments were extended using 3′-full RACE Core Set Ver. 2.0 Kit and5′-full RACE Kit (TaKaRa, Japan). In addition, we examined the middlesequence by dividing it into five fragments. All the primers used in theclone were listed in Table 1. The PCR products were purified using GelExtraction kit (Sangon, China), and sequenced by ABI3730 Biosystem,USA analyzer after insertion into PMD-20T vector (Takara, Japan). Toconfirm the validity of the sequence data that was obtained, each frag-ment was sequenced at least three times. According to the sequencesacquired above, 3′-RACE and 5′-RACE products and the fragment ofmiddle sequence, the full length ofMn-Vg cDNAwas spliced. Sequenceswere analyzed based on the nucleotide and protein databases using the
evelopmental expression of vitellogenin in the oriental river prawndx.doi.org/10.1016/j.gene.2014.12.008

T
167
168
169
170
171
172
173
174
175
176
177
178
179
180
181
182
183
184
185
186
187
188
189
190
191
192
193
194
195
196
197
198
199Q17
200
201
202
203
204
205
206
207
208
209
210
211
212
213
214
215
216
217
218
219
220
221
222
223
224
225
226
227Q18
228
229
230
231
232
233
234
235
236
237
238
239
240
241
242
243
244
245
247247
248
249
250
t2:1
t2:2Q11
t2:3
t2:4
t2:5Q12t2:6
t2:7
t2:8
t2:9
t2:10
t2:11Q13t2:12
t2:13
t2:14
3H. Bai et al. / Gene xxx (2014) xxx–xxx
ORREC
BLASTX and BLASTN search program (http://www.ncbi.nlm.nih.gov/BLAST/) of GenBank. The homology search for the nucleotide andprotein sequences was carried out by the BLAST algorithm at NCBI(http://www.ncbi.nlm.nih.gov/). Deduced amino acid sequences wereobtained using an ORF finder program (http://www.ncbi.nlm.nih.gov/gorf/gorf.html). The signal sequence was predicted using programSignalP (http://www.cbs.dtu.dk/services/SignalP/). The NetNGlyc 1.0Server (http://www.cbs.dtu.dk/services/NetNGlyc/) (N-X-S/T) wasused to identify glycosylation site. The phosphorylation site was foundby the NetPhos 2.0 Server (http://www.cbs.dtu.dk/services/NetPhos/).Phylogenetic analysis of M. nipponense Vg was constructed using theneighbor-joining method of MEGA 5.0.
2.4. Expression of Vg in different development stage and tissue
The expression level of Mn-Vg in different development stages ofembryo, post-larvae, juvenile prawns and different tissues was demon-strated by qPCR. Total RNA treated with RNase-free DNase I (Sangon,China) to eliminate possible genomic DNA contamination. The concen-tration of RNA was quantified by BioPhotometer (Eppendorf). 1 μg oftotal RNA was reverse-transcribed by iScript™ cDNA Synthesis Kit per-fect Real Time (BIO-RAD,USA) following themanufacturer's instruction.The qPCR amplificationswere carried out in a total volumeof 25 μL, con-taining 1 μL cDNA (50 ng), 10 μl SsoFast™ EvaGreen Supermix (Bio-Rad,USA), 0.5 μL 10 μM of genes specific forward and reverse primer(Table 2), and 13 μL of DEPC-water. The reaction mixture was initiallyincubated at 95 °C for 30 s to activate theHot Start TaqDNApolymerase,followed by 40 cycles of 95 °C for 10 s and 60 °C for 10 s, melting curewas performed at the end of qPCR reaction at 65–95 °C (in 0.5 °C inc)for 10 s. At least three replicate qPCRs were performed per sampleand three prawns were analyzed for each sample, and amplification ofβ-actin was used as the internal control (Zhang et al., 2013). The signif-icant differences of expressions were showed at p b 0.05. The relativecopy number of Vg mRNAwas calculated according to the 2−ΔΔCT com-parative CT method (Livak and Schmittgen, 2001).
2.5. Expression of Mn-Vg gene mRNAs after eyestalk ablation
Adult femaleM. nipponense prawns used in this study were selectedfrom Tai lake Wuxi, approximately 1.50 g–3.0 g in wet weight. Theovary development stage of each prawn was identified in the prolifera-tion (stage I). For eyestalk ablation experiment, about 80 prawns weredivided into two groups, one group was served as intact (un-eyestalkablated) and another one was eyestalk ablated (experimental group).The experiments were performed in duplicate and maintained for a pe-riod of 15 days at 28 °C. Female prawn eyestalks were removed by cau-terization with red hot tweezer experiment. Samples from differenttissues were collected from prawns at 1, 3, 5, 7, 9, 11 and 13 days after
UNC
251
252
253
254
255
256Q19
257Q20
258
259
260
261
262
263
264
265
266
Table 2Primers used for quantitative PCR and RNA inference.
Primername
Sequence (5′ → 3′) Purpose
VG-Q-F GAAGTTAGCGGAGATCTGAGGT Primer for MnVG expressionVG-Q-R CCTCGTTGACCAATCTTGAGAG Primer for MnVG expressionVG-I-F TAATACGACTCACTATAGGTGC
CAAGAAAAAGCTCCTGTPrimer for MnVG dsRNA preparation
VG-I-R TAATACGACTCACTATAGGGCCAAAGGTTGGTGCATAGT
Primer for MnVG dsRNA preparation
VG-QI-F TGCTCTTGCTCTACTCAAGTCC Primer for MnVG dsRNA detectionVG-QI-R CTGATGACAGTGAACGTTCCTG Primer for MnVG dsRNA detectionVGR-QI-F TACCACTTCGTCACAGATGCAG Primer for MnVGR dsRNA detectionVGR-QI-R CTTGTCGCACCAGTAGATCCTC Primer for MnVGR dsRNA detectionβ-Actin F TATGCACTTCCTCATGCCATC Primer for β-actin expressionβ-Actin R AGGAGGCGGCAGTGGTCAT primer for β-actin expression
Please cite this article as: Bai, H., et al., Molecular characterization and dMacrobrachium nipponense and the effects of RNA ..., Gene (2014), http://
ED P
RO
OF
eyestalk ablation (N≥ 3). All collect sampleswere stored at RNA sampleprotect liquid. qPCRwas used to detect the expression pattern after eye-stalk ablation. The specific method of transcription and qPCR process isthe same with different development stages and tissues.
2.6. RNA interference (RNAi)
The specific primers containing T7 promoter site of RNA interferencewere designed using Snap Dragon tools (http://www.flyrnai.org/cgi-bin/RNAifind_primers.pl). Vg dsRNA synthesis primers were shown inTable 2. The PCR products were purified with a gel extraction kit(Sangon, Shanghai, China). According to the Transcript Aid™ T7 HighYield Transcription kit (Fermentas, Inc, USA) manufacturer's instruc-tions, gene-specific dsRNAwas synthesized in vitro. Purity and integrityof the dsRNAwere examined by standard agarose gel electrophoresis. Theconcentration of dsRNA was measured at 260 nm by using a BioPhotom-eter (Eppendorf, Hamburg, Germany), and then save at −20 °C. Allprimers usedwere listed in Table 2. VgR detection primerswere designedaccording to the sequence of NCBI under accession (GenBank KJ768658).
For the short-term in vivo dsRNA injection experiment, 150 healthmature female M. nipponense (each weighing 1.6–2.3 g) were selectedto inject into pericardial cavity. The female prawns, selected in the pro-liferation stage (stage I), were assigned to the three treatment groups:Vg-dsRNA injection (N = 50), two Vg-dsRNA injection (N = 50) andvehicle injection (N = 50). Each prawn was injected with 4 μg/g Vg-dsRNA or 4 μg vehicles. For two injection group, supplemented another4 μg/g Vg-dsRNA in the seventh day after the first injection. The VgmRNA expression of the ovary and hepatopancreas were investigatedto detect the interference efficiency by qPCR after injection for 1, 3, 5,7, 9, 11, 13, 15 and 17 days (N≥ 3). The body weight and ovary weightof each prawn was recorded to measure the GSI.
For two injection group, samples were captured respectively in the1, 5, 9, 13, 17, 21, and 25 days after the injection (N ≥ 3). All tissueswere saved in the RNA protect liquid.
2.7. Statistical analysis
The prawns were weighed and the ovaries were dissected out andindexes were determined using the following formula:
gonadsomatic index GSIð Þ ¼ wet weight of the ovarywet weight of the prawn
� 100%:
Quantitative data were expressed as means ± SD.Statistical differences were estimated by one-way ANOVA followed
by Duncan's multiple range test.
3. Results
3.1. Molecular cloning and structural analysis of the Mn-Vg gene
The full-length Mn-Vg gene is 7804 base pairs (bps) long and in-cludes a 5′-terminal untranslated region (UTR) of 34 bp, a 7611 bpopen reading frame (ORF) encoding 2536 amino acid (aa) residues,and a 159 bp 3′-terminal UTR (excluding the poly(A) + tail). The Mn-Vg cDNA sequence was submitted to GenBank under the accessionnumber KJ768657. Fig. 1 shows the sketch map of the deduced aminoacid sequences, and the specific sequence information forMn-Vg is pro-vided in Fig. 1s.Mn-Vg has an estimatedmolecularmass of 286.810 kDaand a theoretical isoelectric point of 9.08. SignalP software analysis re-vealed that the deduced peptides contained a putative 20 aa signal pep-tide and a cleavage site between Pro20 and Ser21. The deduced aminoacid sequences included six N-glycosylation sites (N-X-S/T) and twopu-tative subtilisin cleavage sites (RQKR). In addition, 151 phosphorylationsites (90 Ser, 40 Thr, and 21 Try) were identified by the NetPhos 2.0Server. Alignment with other species revealed that the Mn-Vg protein
evelopmental expression of vitellogenin in the oriental river prawndx.doi.org/10.1016/j.gene.2014.12.008

T
267
268
269
270
271
272
273
274
275
276
277
278
279
280
281
282
283
284
285
286
287
288
289
290
291
292
293
294
295
296
297
298
299
300
301
302
303
304
305
306
307
308
309
310
311
312
313
314
315
316
317
318
319
320
321
322
323
324
325
326
327
328
329
330
331
332
333
334
335
336
337
338
339
340
341
342
343
344
345
346
347
348
349
350
351
352
353
354
355
356
357
358
359
360
361
362
363
364
365
366
RERR
RERRSignal peptide
GLLG
Fig. 1. The sketchmap of the deduced amino acid sequences ofM. nipponenseVg. The conserved domain (Vitellogenin_N, DUF 1943, VWFD) of VGwas colored. PutativeQ1 signal peptide andsubtilisin cleavage sites (RQKR) are highlighted.
4 H. Bai et al. / Gene xxx (2014) xxx–xxx
UNCO
RREC
contains conserved domains, including Vitellogenin_N, VWFD, and thedomain of unknown function (DUF) 1943 super family. The first twoare highly conserved in other insect and crustacean species. A BLASTsearch revealed thatMn-Vgwas highly similar to theVg of the followingcrustaceans: M. rosenbergii (91% identity), Exopalaemon carinicauda(76% identity), P. hypsinotus (61% identity), P. japonica (60% identi-ty), Cherax quadricarinatus (39% identity), Homarus americanus(38% identity), M. japonicus (38% identity), F. chinensis (38% identity),and P. merguiensis (37% identity).
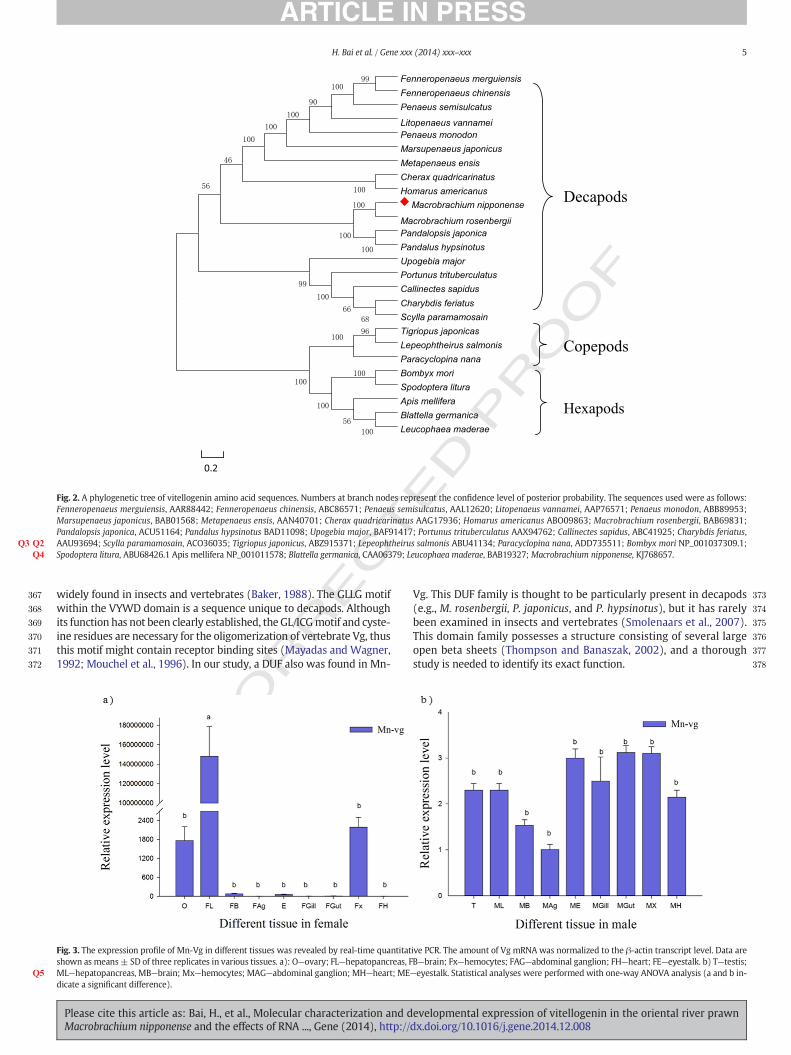
A neighbor-joining phylogenic tree (Fig. 2) was constructed basedon the Vg amino acid sequences of all reported decapods, three repre-sentative copepods, and five hexapods. The branches of the phylogenictree revealed that Mn-Vg was most closely related to M. rosenbergii,followed by P. japonica and P. hypsinotus. Vgs of all crustaceans clusteredtogether as a group. However, copepods and hexapods formed a distinc-tive cluster separated from the other crustacean Vg proteins, and Vgsfrom decapods were more closely related to those of copepods than tothose of hexapods.
3.2. Tissue distribution of the Mn-Vg gene
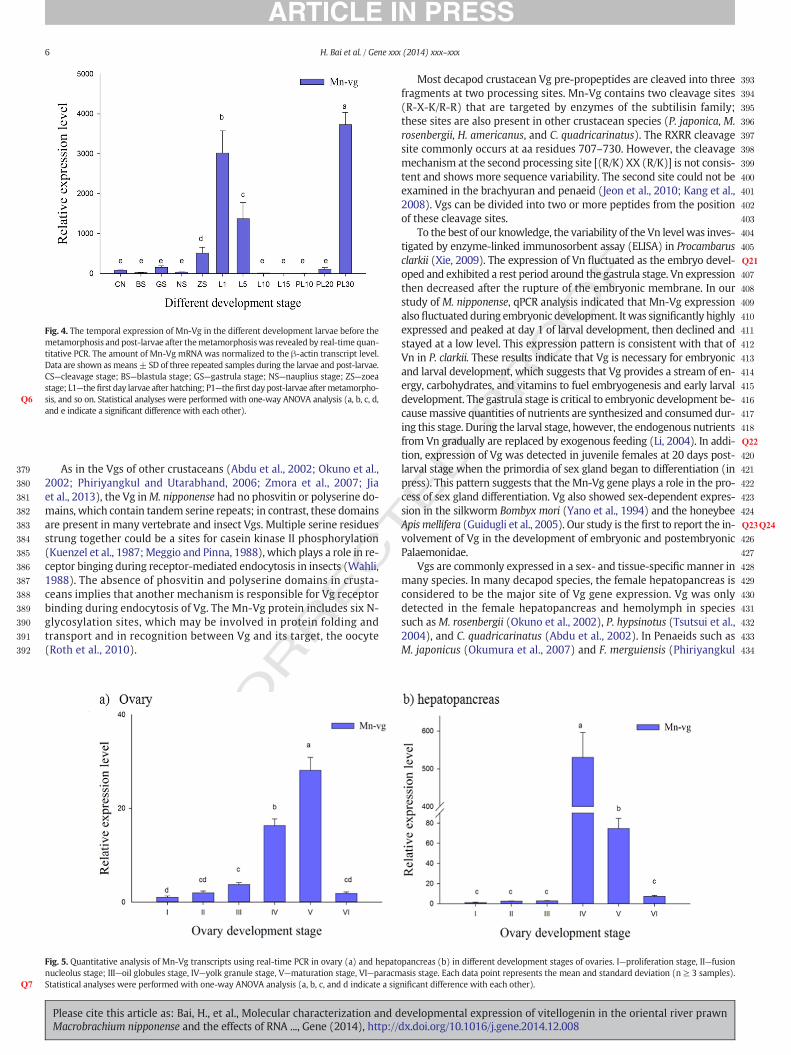
The distribution and expression of theMn-Vg genewere analyzed byqPCR in selected female prawn tissues. Mn-Vg was highly expressed inthe hepatopancreas (Fig. 3) (P b 0.001), followed by the ovary and hemo-cytes. Expression was significantly lower in the other tissues (i.e., brain,gill, heart, and abdominal ganglion). In male prawn tissues, few mRNAtranscripts were detected, and they were present at extremely low levels(i.e., testis, brain, gill, heart, and abdominal ganglion). The expressionlevel in male tissues is less than one per few million when comparedwith the expression level of female hepatopancreas.
3.3. Expression of the Mn-Vg gene in larvae and post-larvae
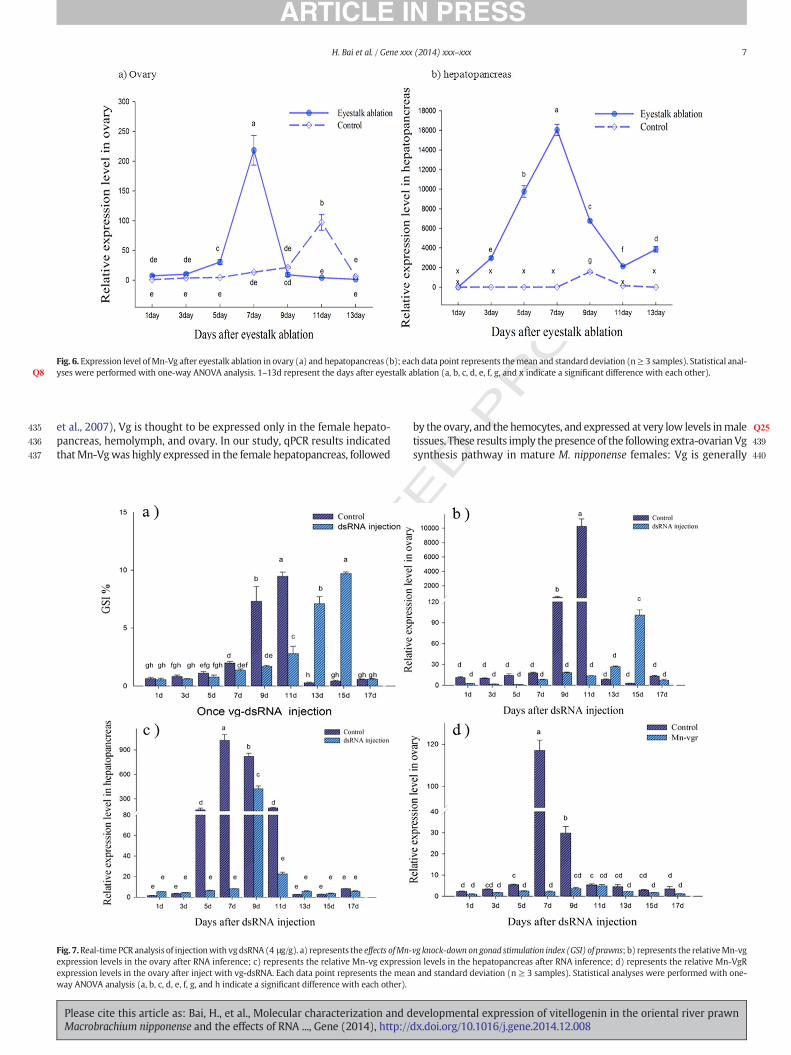
The expression pattern of Mn-Vg at different developmental stages(embryo, larvae, and post-larvae) was evaluated by qPCR. Mn-Vg washighly expressed in the cleavage, gastrula, and zoea stages, but expres-sion declined slightly in the blastula and nauplius stages (Fig. 4). Oncelarvae ruptured the embryonicmembrane, Vg expression decreased sig-nificantly and was maintained at relatively low level. However, a sharpincrease in expression occurred beginning 20 days after metamorphosis,and the highest expression level was measured 30 days after metamor-phosis. The expression level of 30 days post-larval showed significant dif-ference with other development stages.
3.4. Expression of the Mn-Vg gene in different stages of ovarian development
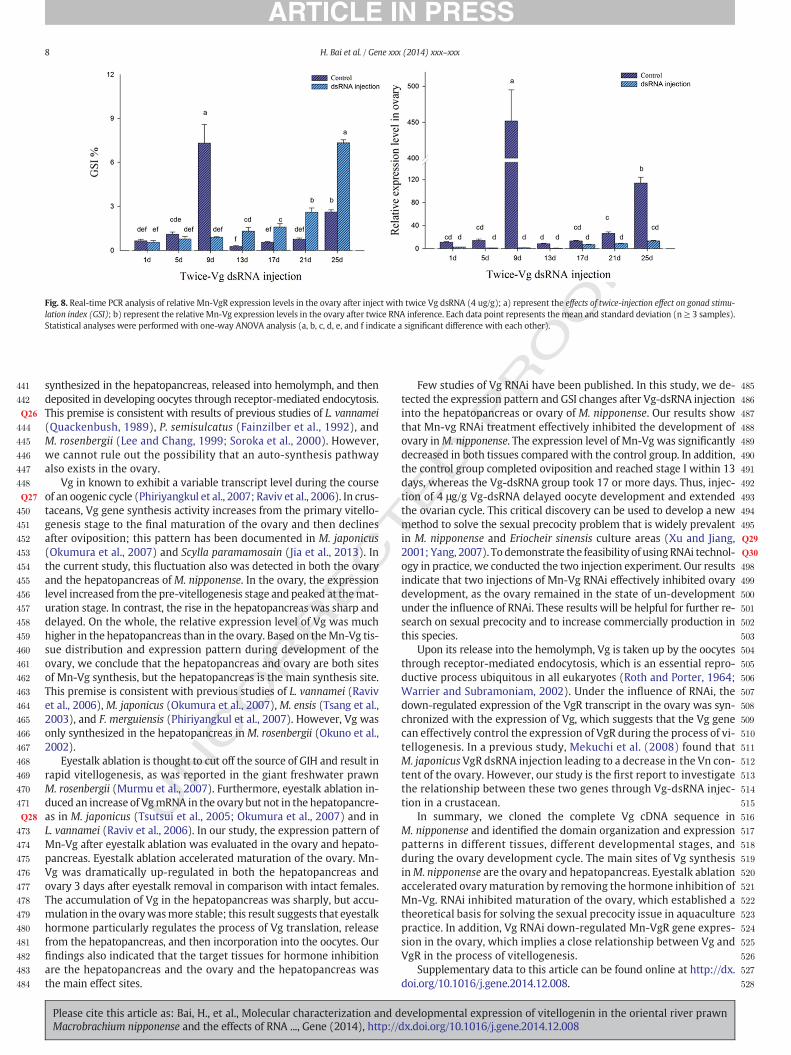
Fig. 5 shows the expression pattern of Mn-Vg in the ovary and hepa-topancreas during the reproductive cycle. The qPCR results show thatthe Mn-Vg level increased in both organs as ovarian developmentprogressed. In the hepatopancreas, the expression level gradually in-creased from stages I to III and then abruptly increased and peaked atstage IV. After reproductive molting, the expression of Mn-Vg in stageVI decreased rapidly to reach the same level of stage I. In the ovary,Mn-Vg gene expression increased gradually from stage I to stage Vand reached the peaked. In the ovary, the expression level rose morerapidly at the early stage of vitellogenesis and reached the peak more
Please cite this article as: Bai, H., et al., Molecular characterization and dMacrobrachium nipponense and the effects of RNA ..., Gene (2014), http://
ED P
RO
OF
slowly compared to the expression pattern in hepatopancreas. Howev-er, the relative expression level in the hepatopancreaswasmuch higherthan that in the ovary.
3.5. Expression of Mn-Vg mRNAs after eyestalk ablation
The expression level of the Mn-Vg after eyestalk removal was mon-itored by qPCR in the ovary and hepatopancreas (Fig. 6). When com-pared with the control female prawn group, the reproductive moltcycle in the eyestalk ablation group was dramatically shortened bothin two tissues. In the hepatopancreas, Mn-Vg expression was up-regulated 3 days after eyestalk ablation and reached a level of 3000-fold higher than that of the control group (P b 0.01). Overall, themagni-tude of gene expression level detected in the experimental group wasmuch higher than that in the control group. As the ovary developed,Mn-Vg expression in the hepatopancreas peaked 7 days after eyestalkablation in the treatment group, whereas it peaked at 9 days in the con-trol group. A similar expression pattern was found in the ovary, but theexpression level in the eyestalk ablation grouppeaked4 days in advanceof the control group.
3.6. Expression of Vg and VgR after RNAi
Many researchers have reported that injection of double strandedRNA (dsRNA) leads to gene silencing. When RNA interference (RNAi)was used in this study, the Vg-dsRNA clearly inhibited development ofthe ovary based on the gonadosomatic index (GSI) (Fig. 7a) and expres-sion pattern data. In the ovary, injection of Vg-dsRNA resulted in an 80%decrease of Vg expression after 3 days (Fig. 7b). Although the expressionlevel increased in both the test and control groups as the ovarymatured,different patternswere observed.When the control group had complet-ed a normal reproductive cycle, the experimental group was still in theyolk accumulation phase (stage III). The same expression pattern wasdetected in the hepatopancreas (Fig. 7c). These results illustrate thatRNAi was able to delay the development cycle of the ovary.
In order to detect the effectiveness of Vg-dsRNA in inhibiting matura-tion of the ovary,we injected another 4 μg/g of dsRNA in the two injectiongroup after oneweek. The delay inmaturation in the experimental groupwas even more pronounced compared to that in the one injection andcontrol groups. When the control prawns had begun a new reproductivecycle, the two injection group was still in the pre-vitellogenesis stage ac-cumulating yolk protein (Fig. 8).
qPCR analysis indicated that injection of Vg-dsRNA resulted in down-regulation of VgR. The Mn-VgR gene transcript level declined by 90% inthe ovary, presumably due to decreased Vg expression (Fig. 7d). In theprocess of ovarianmaturation, the level of Mn-VgR fluctuated in the con-trol group, whereas it stayed low in the Vg RNAi group.
4. Discussion
In this study,we identified the completeMn-Vg transcript sequence,which is approximately 8 kb in size with 2536 aas encoded by its ORF.The deduced amino acid sequences revealed the common characteristicsequence of insect Vg (Chen et al., 1997; Sappington and Raikhel, 1998;Sappington et al., 2002), with considerable conservation, particularly inthe N-terminus. The vitellogenin N-terminal and VYWD domains are
evelopmental expression of vitellogenin in the oriental river prawndx.doi.org/10.1016/j.gene.2014.12.008
CTED P
RO
OF
367
368
369
370
371
372
373
374
375
376
377
378
0.2
Fenneropenaeus merguiensisFenneropenaeus chinensisPenaeus semisulcatus
Litopenaeus vannameiPenaeus monodonMarsupenaeus japonicusMetapenaeus ensisCherax quadricarinatusHomarus americanus
Macrobrachium nipponense
Macrobrachium rosenbergiiPandalopsis japonicaPandalus hypsinotusUpogebia majorPortunus trituberculatusCallinectes sapidusCharybdis feriatusScylla paramamosainTigriopus japonicasLepeophtheirus salmonisParacyclopina nanaBombyx moriSpodoptera lituraApis melliferaBlattella germanicaLeucophaea maderae
Decapods
Copepods
Hexapods
Fig. 2. A phylogenetic tree of vitellogenin amino acid sequences. Numbers at branch nodes represent the confidence level of posterior probability. The sequences used were as follows:Fenneropenaeus merguiensis, AAR88442; Fenneropenaeus chinensis, ABC86571; Penaeus semisulcatus, AAL12620; Litopenaeus vannamei, AAP76571; Penaeus monodon, ABB89953;Marsupenaeus japonicus, BAB01568; Metapenaeus ensis, AAN40701; Cherax quadricarinatus AAG17936; Homarus americanus ABO09863; Macrobrachium rosenbergii, BAB69831;Pandalopsis japonica, ACU51164; Pandalus hypsinotus BAD11098; Upogebia major, BAF91417; Portunus trituberculatus AAX94762; Callinectes sapidus, ABC41925; Charybdis feriatus,AAU93694; Scylla paramamosain, ACO36035; Tigriopus japonicusQ2 , ABZ915371; Lepeophtheirus salmonis ABU41134; Paracyclopina nanaQ3 , ADD735511; Bombyx mori NP_001037309.1;Spodoptera litura, ABU68426.1 Apis mellifera NP_001011578; Blattella germanica, CAA06379; Leucophaea maderae, BAB19327; Macrobrachium nipponense,Q4 KJ768657.
Q5
5H. Bai et al. / Gene xxx (2014) xxx–xxx
REwidely found in insects and vertebrates (Baker, 1988). The GLLG motif
within the VYWD domain is a sequence unique to decapods. Althoughits function has not been clearly established, the GL/ICGmotif and cyste-ine residues are necessary for the oligomerization of vertebrate Vg, thusthis motif might contain receptor binding sites (Mayadas and Wagner,1992; Mouchel et al., 1996). In our study, a DUF also was found in Mn-
UNCO
R
Fig. 3. The expression profile of Mn-Vg in different tissues was revealed by real-time quantitatshown as means± SD of three replicates in various tissues. a): O—ovary; FL—hepatopancreas, FML—hepatopancreas, MB—brain; Mx—hemocytes; MAG—abdominal ganglion; MH—heart; MEdicate a significant difference).
Please cite this article as: Bai, H., et al., Molecular characterization and dMacrobrachium nipponense and the effects of RNA ..., Gene (2014), http://
Vg. This DUF family is thought to be particularly present in decapods(e.g., M. rosenbergii, P. japonicus, and P. hypsinotus), but it has rarelybeen examined in insects and vertebrates (Smolenaars et al., 2007).This domain family possesses a structure consisting of several largeopen beta sheets (Thompson and Banaszak, 2002), and a thoroughstudy is needed to identify its exact function.
ive PCR. The amount of Vg mRNA was normalized to the β-actin transcript level. Data areB—brain; Fx—hemocytes; FAG—abdominal ganglion; FH—heart; FE—eyestalk. b) T—testis;—eyestalk. Statistical analyses were performed with one-way ANOVA analysis (a and b in-
evelopmental expression of vitellogenin in the oriental river prawndx.doi.org/10.1016/j.gene.2014.12.008
T
379
380
381
382
383
384
385
386
387
388
389
390
391
392
393
394
395
396
397
398
399
400
401
402
403
404
405
406Q21
407
408
409
410
411
412
413
414
415
416
417
418
419Q22
420
421
422
423
424
425Q23Q24
426
427
428
429
430
431
432
433
434
Fig. 4. The temporal expression of Mn-Vg in the different development larvae before themetamorphosis and post-larvae after themetamorphosiswas revealed by real-time quan-titative PCR. The amount of Mn-Vg mRNA was normalized to the β-actin transcript level.Data are shown asmeans± SD of three repeated samples during the larvae and post-larvae.CS—cleavage stage; BS—blastula stage; GS—gastrula stage; NS—nauplius stage; ZS—zoeastage; L1—thefirst day larvae after hatching; P1—thefirst day post-larvae aftermetamorpho-sis, and so on. Statistical analyses were performed with one-way ANOVA analysis (Q6 a, b, c, d,and e indicate a significant difference with each other).
Q7
6 H. Bai et al. / Gene xxx (2014) xxx–xxx
REC
As in the Vgs of other crustaceans (Abdu et al., 2002; Okuno et al.,2002; Phiriyangkul and Utarabhand, 2006; Zmora et al., 2007; Jiaet al., 2013), the Vg inM. nipponense had no phosvitin or polyserine do-mains, which contain tandem serine repeats; in contrast, these domainsare present in many vertebrate and insect Vgs. Multiple serine residuesstrung together could be a sites for casein kinase II phosphorylation(Kuenzel et al., 1987;Meggio and Pinna, 1988), which plays a role in re-ceptor binging during receptor-mediated endocytosis in insects (Wahli,1988). The absence of phosvitin and polyserine domains in crusta-ceans implies that another mechanism is responsible for Vg receptorbinding during endocytosis of Vg. The Mn-Vg protein includes six N-glycosylation sites, which may be involved in protein folding andtransport and in recognition between Vg and its target, the oocyte(Roth et al., 2010).
UNCO
R
Fig. 5. Quantitative analysis of Mn-Vg transcripts using real-time PCR in ovary (a) and hepatonucleolus stage; III—oil globules stage, IV—yolk granule stage, V—maturation stage, VI—paracmStatistical analyses were performed with one-way ANOVA analysis (a, b, c, and d indicate a sig
Please cite this article as: Bai, H., et al., Molecular characterization and dMacrobrachium nipponense and the effects of RNA ..., Gene (2014), http://
ED P
RO
OF
Most decapod crustacean Vg pre-propeptides are cleaved into threefragments at two processing sites. Mn-Vg contains two cleavage sites(R-X-K/R-R) that are targeted by enzymes of the subtilisin family;these sites are also present in other crustacean species (P. japonica, M.rosenbergii, H. americanus, and C. quadricarinatus). The RXRR cleavagesite commonly occurs at aa residues 707–730. However, the cleavagemechanism at the second processing site [(R/K) XX (R/K)] is not consis-tent and shows more sequence variability. The second site could not beexamined in the brachyuran and penaeid (Jeon et al., 2010; Kang et al.,2008). Vgs can be divided into two or more peptides from the positionof these cleavage sites.
To the best of our knowledge, the variability of the Vn levelwas inves-tigated by enzyme-linked immunosorbent assay (ELISA) in Procambarusclarkii (Xie, 2009). The expression of Vn fluctuated as the embryo devel-oped and exhibited a rest period around the gastrula stage. Vn expressionthen decreased after the rupture of the embryonic membrane. In ourstudy of M. nipponense, qPCR analysis indicated that Mn-Vg expressionalsofluctuated during embryonic development. It was significantly highlyexpressed and peaked at day 1 of larval development, then declined andstayed at a low level. This expression pattern is consistent with that ofVn in P. clarkii. These results indicate that Vg is necessary for embryonicand larval development, which suggests that Vg provides a stream of en-ergy, carbohydrates, and vitamins to fuel embryogenesis and early larvaldevelopment. The gastrula stage is critical to embryonic development be-causemassive quantities of nutrients are synthesized and consumed dur-ing this stage. During the larval stage, however, the endogenous nutrientsfrom Vn gradually are replaced by exogenous feeding (Li, 2004). In addi-tion, expression of Vg was detected in juvenile females at 20 days post-larval stage when the primordia of sex gland began to differentiation (inpress). This pattern suggests that the Mn-Vg gene plays a role in the pro-cess of sex gland differentiation. Vg also showed sex-dependent expres-sion in the silkworm Bombyx mori (Yano et al., 1994) and the honeybeeApis mellifera (Guidugli et al., 2005). Our study is the first to report the in-volvement of Vg in the development of embryonic and postembryonicPalaemonidae.
Vgs are commonly expressed in a sex- and tissue-specific manner inmany species. In many decapod species, the female hepatopancreas isconsidered to be the major site of Vg gene expression. Vg was onlydetected in the female hepatopancreas and hemolymph in speciessuch asM. rosenbergii (Okuno et al., 2002), P. hypsinotus (Tsutsui et al.,2004), and C. quadricarinatus (Abdu et al., 2002). In Penaeids such asM. japonicus (Okumura et al., 2007) and F. merguiensis (Phiriyangkul
pancreas (b) in different development stages of ovaries. I—proliferation stage, II—fusionasis stage. Each data point represents the mean and standard deviation (n ≥ 3 samples).nificant difference with each other).
evelopmental expression of vitellogenin in the oriental river prawndx.doi.org/10.1016/j.gene.2014.12.008
RO
OF
435
436
437
438Q25
439
440
Fig. 6. Expression level ofMn-Vg after eyestalk ablation in ovary (a) and hepatopancreas (b); each data point represents themean and standard deviation (n≥ 3 samples). Statistical anal-yses were performed with one-way ANOVA analysis.Q8 1–13d represent the days after eyestalk ablation (a, b, c, d, e, f, g, and x indicate a significant difference with each other).
7H. Bai et al. / Gene xxx (2014) xxx–xxx
et al., 2007), Vg is thought to be expressed only in the female hepato-pancreas, hemolymph, and ovary. In our study, qPCR results indicatedthatMn-Vgwas highly expressed in the female hepatopancreas, followed
UNCO
RRECT
Fig. 7.Real-time PCR analysis of injectionwith vgdsRNA (4 μg/g). a) represents the effects ofMn-expression levels in the ovary after RNA inference; c) represents the relative Mn-vg expressiexpression levels in the ovary after inject with vg-dsRNA. Each data point represents the meaway ANOVA analysis (a, b, c, d, e, f, g, and h indicate a significant difference with each other).
Please cite this article as: Bai, H., et al., Molecular characterization and dMacrobrachium nipponense and the effects of RNA ..., Gene (2014), http://
Pby the ovary, and the hemocytes, and expressed at very low levels inmaletissues. These results imply thepresence of the following extra-ovarianVgsynthesis pathway in mature M. nipponense females: Vg is generally
ED
vg knock-down ongonad stimulation index (GSI) of prawns; b) represents the relativeMn-vgon levels in the hepatopancreas after RNA inference; d) represents the relative Mn-VgRn and standard deviation (n ≥ 3 samples). Statistical analyses were performed with one-
evelopmental expression of vitellogenin in the oriental river prawndx.doi.org/10.1016/j.gene.2014.12.008
T
OO
F
441
442
443Q26
444
445
446
447
448
449Q27
450
451
452
453
454
455
456
457
458
459
460
461
462
463
464
465
466
467
468
469
470
471
472Q28
473
474
475
476
477
478
479
480
481
482
483
484
485
486
487
488
489
490
491
492
493
494
495
496Q29
497Q30
498
499
500
501
502
503
504
505
506
507
508
509
510
511
512
513
514
515
516
517
518
519
520
521
522
523
524
525
526
527
528
Fig. 8. Real-time PCR analysis of relative Mn-VgR expression levels in the ovary after inject with twice Vg dsRNA (4 ug/g); a) represent the effects of twice-injection effect on gonad stimu-lation index (GSI); b) represent the relative Mn-Vg expression levels in the ovary after twice RNA inference. Each data point represents themean and standard deviation (n≥ 3 samples).Statistical analyses were performed with one-way ANOVA analysis (a, b, c, d, e, and f indicate a significant difference with each other).
8 H. Bai et al. / Gene xxx (2014) xxx–xxx
UNCO
RREC
synthesized in the hepatopancreas, released into hemolymph, and thendeposited in developing oocytes through receptor-mediated endocytosis.This premise is consistent with results of previous studies of L. vannamei(Quackenbush, 1989), P. semisulcatus (Fainzilber et al., 1992), andM. rosenbergii (Lee and Chang, 1999; Soroka et al., 2000). However,we cannot rule out the possibility that an auto-synthesis pathwayalso exists in the ovary.
Vg in known to exhibit a variable transcript level during the courseof an oogenic cycle (Phiriyangkul et al., 2007; Raviv et al., 2006). In crus-taceans, Vg gene synthesis activity increases from the primary vitello-genesis stage to the final maturation of the ovary and then declinesafter oviposition; this pattern has been documented in M. japonicus(Okumura et al., 2007) and Scylla paramamosain (Jia et al., 2013). Inthe current study, this fluctuation also was detected in both the ovaryand the hepatopancreas of M. nipponense. In the ovary, the expressionlevel increased from the pre-vitellogenesis stage and peaked at themat-uration stage. In contrast, the rise in the hepatopancreas was sharp anddelayed. On the whole, the relative expression level of Vg was muchhigher in the hepatopancreas than in the ovary. Based on theMn-Vg tis-sue distribution and expression pattern during development of theovary, we conclude that the hepatopancreas and ovary are both sitesof Mn-Vg synthesis, but the hepatopancreas is the main synthesis site.This premise is consistent with previous studies of L. vannamei (Ravivet al., 2006), M. japonicus (Okumura et al., 2007), M. ensis (Tsang et al.,2003), and F. merguiensis (Phiriyangkul et al., 2007). However, Vg wasonly synthesized in the hepatopancreas in M. rosenbergii (Okuno et al.,2002).
Eyestalk ablation is thought to cut off the source of GIH and result inrapid vitellogenesis, as was reported in the giant freshwater prawnM. rosenbergii (Murmu et al., 2007). Furthermore, eyestalk ablation in-duced an increase of VgmRNA in the ovary but not in the hepatopancre-as in M. japonicus (Tsutsui et al., 2005; Okumura et al., 2007) and inL. vannamei (Raviv et al., 2006). In our study, the expression pattern ofMn-Vg after eyestalk ablation was evaluated in the ovary and hepato-pancreas. Eyestalk ablation accelerated maturation of the ovary. Mn-Vg was dramatically up-regulated in both the hepatopancreas andovary 3 days after eyestalk removal in comparison with intact females.The accumulation of Vg in the hepatopancreas was sharply, but accu-mulation in the ovarywasmore stable; this result suggests that eyestalkhormone particularly regulates the process of Vg translation, releasefrom the hepatopancreas, and then incorporation into the oocytes. Ourfindings also indicated that the target tissues for hormone inhibitionare the hepatopancreas and the ovary and the hepatopancreas wasthe main effect sites.
Please cite this article as: Bai, H., et al., Molecular characterization and dMacrobrachium nipponense and the effects of RNA ..., Gene (2014), http://
ED P
RFew studies of Vg RNAi have been published. In this study, we de-tected the expression pattern and GSI changes after Vg-dsRNA injectioninto the hepatopancreas or ovary of M. nipponense. Our results showthat Mn-vg RNAi treatment effectively inhibited the development ofovary inM. nipponense. The expression level of Mn-Vg was significantlydecreased in both tissues comparedwith the control group. In addition,the control group completed oviposition and reached stage I within 13days, whereas the Vg-dsRNA group took 17 or more days. Thus, injec-tion of 4 μg/g Vg-dsRNA delayed oocyte development and extendedthe ovarian cycle. This critical discovery can be used to develop a newmethod to solve the sexual precocity problem that is widely prevalentin M. nipponense and Eriocheir sinensis culture areas (Xu and Jiang,2001; Yang, 2007). To demonstrate the feasibility of using RNAi technol-ogy in practice, we conducted the two injection experiment. Our resultsindicate that two injections of Mn-Vg RNAi effectively inhibited ovarydevelopment, as the ovary remained in the state of un-developmentunder the influence of RNAi. These results will be helpful for further re-search on sexual precocity and to increase commercially production inthis species.
Upon its release into the hemolymph, Vg is taken up by the oocytesthrough receptor-mediated endocytosis, which is an essential repro-ductive process ubiquitous in all eukaryotes (Roth and Porter, 1964;Warrier and Subramoniam, 2002). Under the influence of RNAi, thedown-regulated expression of the VgR transcript in the ovary was syn-chronized with the expression of Vg, which suggests that the Vg genecan effectively control the expression of VgR during the process of vi-tellogenesis. In a previous study, Mekuchi et al. (2008) found thatM. japonicus VgR dsRNA injection leading to a decrease in the Vn con-tent of the ovary. However, our study is the first report to investigatethe relationship between these two genes through Vg-dsRNA injec-tion in a crustacean.
In summary, we cloned the complete Vg cDNA sequence inM. nipponense and identified the domain organization and expressionpatterns in different tissues, different developmental stages, andduring the ovary development cycle. The main sites of Vg synthesisinM. nipponense are the ovary and hepatopancreas. Eyestalk ablationaccelerated ovarymaturation by removing the hormone inhibition ofMn-Vg. RNAi inhibited maturation of the ovary, which established atheoretical basis for solving the sexual precocity issue in aquaculturepractice. In addition, Vg RNAi down-regulated Mn-VgR gene expres-sion in the ovary, which implies a close relationship between Vg andVgR in the process of vitellogenesis.
Supplementary data to this article can be found online at http://dx.doi.org/10.1016/j.gene.2014.12.008.
evelopmental expression of vitellogenin in the oriental river prawndx.doi.org/10.1016/j.gene.2014.12.008

T
529
530Q31Q32
531
532
533
534
535
536
537
538
539
540
541542543544545546547548549550551552Q33553554555556557558559560561562563564565566567568569570571572573574575576577578579580581582583584585586587588589590591592593594595596597598599600601602603604605606
607608609610611612613614615616617618619620621622623624625626627628629630631632633634635636637638639640641642643644645646647648649650651652653654655656657658659660661662663664665666667668669670671672673674675676677678679680681682683684685686687688689690691692
9H. Bai et al. / Gene xxx (2014) xxx–xxx
UNCO
RREC
Acknowledgments
The project was supported by the Freshwater Fisheries ResearchCenter, China Central Governmental Research Institutional Basic SpecialResearch Project from the Public Welfare Fund (2013JBFM15), the Na-tional Natural Science Foundation of China (Grant No. 31272654), theNational Science & Technology Supporting Program of the 12th Five-year Plan of China (Grant No. 2012BAD26B04), Jiangsu Provincial NaturalScience Foundation for Young Scholars of China (Grant No. BK2012091),the Science & Technology Supporting Program of Jiangsu Province(GrantNo. BE2012334), and the three aquatic projects of Jiangsu Province(D2013-6).
References
Abdu, U., Davis, C., Khalaila, I., Sagi, A., 2002. The vitellogenin cDNA of Cherax quadricarinatusencodes a lipoprotein with calcium binding ability, and its expression is induced follow-ing the removal of the androgenic gland in a sexually plastic system. Gen. Comp.Endocrinol. 127 (3), 263–272.
Adiyodi, K.G., Adiyodi, R.G., 1970. Endocrine control of reproduction in decapod Crusta-cea. Biol. Rev. 45 (2), 121–164.
Baker, M.E., 1988. Is vitellogenin an ancestor of apolipoprotein B-100 of human lipopro-tein and human lipoprotein lipase. Biochemistry 255, 1057–1060.
Browdy, C.L., Fainzilber, M., Tom, M., Loya, Y., Lubzens, E., 1990. Vitellin synthesis in rela-tion to oogenesis in in vitro-incubated ovaries of Penaeus semisulcatus (Crustaceas,Decapoda, Penaeidae). Exp. Zool. 255, 205–215.
Bureau of Fishery, Ministry of Agriculture, P.R.C., 2011. Fisheries economic statistics. ChinaFishery Yearbook. China Agricultural Press, Beijing, p. 236.
Chen, J.S., Sappington, T.W., Raikhel, A.S., 1997. Extensive sequence conservation amonginsect, nematode, and vertebrate vitellogenins reveals ancient common ancestry.Mol. Evol. 44 (4), 440–451.
Chen, Y., Zhu, Q., Chen, H., Zhu, X.L., Cui, Z., Qiu, G.F., 2012. The morphological and histolog-ical observation of embryonic development in the oriental river prawn Macrobrachiumnipponense. J. Shanghai Ocean Univ. 21 (1), 33–40.
De Kleijn, D.P., Janssen, K.P., Waddy, S.L., Hegeman, R., Lai, W.Y., Martens, G.J., Van Herp, F.,1998. Expression of the crustacean hyperglycaemic hormones and the Gonadinhibiting hormone during the reproductive cycle of the female American lobsterHomarus americanus. J. Endocrinol. 156 (2), 291–298.
Fainzilber, M., Tom, M., Shafir, M., Applebaum, S.W., Lubzens, E., 1992. Is there extraovariansynthesis of vitellogenin in penaeid shrimp. Biol. Bull. 183 (2), 233–241.
Gu, P.L., Tobe, S.S., Chow, B.K., Chu, K.H., He, J.G., Chan, S.M., 2002. Characterization of anadditional molt inhibiting hormone-like neuropeptide from the shrimp Metapenaeusensis. Peptides 23 (11), 1875–1883.
Guidugli, K.R., Piulachs, M.D., Bellés, X., Lourenço, A.P., Simões, Z.L., 2005. Vitellogenin ex-pression in queen ovaries and in larvae of both sexes of Apis mellifera. Arch. InsectBiochem. Physiol. 59 (4), 211–218.
Jayasankar, V., Tsutsui, N., Jasmani, S., Saido-Sakanaka, H., Yang, W.J., Okuno, A., Tran, T.T.,Aida, K., Wilder, M.N., 2002. Dynamics of vitellogenin mRNA expression and changesin hemolymph vitellogenin levels during ovarian maturation in the giant freshwaterprawn Macrobrachium rosenbergii. J. Exp. Zool. 293 (5), 675–682.
Jeon, J.M., et al., 2010. Characterization of two vitellogenin cDNAs from a Pandalus shrimp(Pandalopsis japonica): expression in hepatopancreas is down-regulated by endosul-fan exposure. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 157 (1), 102–112.
Jia, X., Chen, Y., Zou, Z., Lin, P., Wang, Y., Zhang, Z., 2013. Characterization and expressionprofile of Vitellogenin gene from Scylla paramamosain. Gene 520 (2), 119–130.
Kang, X.J., Sun, T.C., Wang, S.A., 1996. Preliminary study on the isolated culture of the fer-tilized egg and embryonic development of theMacrobrachium. nipponense (CrustaceaDecapoda). J. Hebei Univ. (Nat. Sci. Ed.) 16 (4), 41–45.
Kang, B.J., et al., 2008. Vitellogenesis in both sexes of gonochoristic mud shrimp, Upogebiamajor (Crustacea): analyses of vitellogenin gene expression and vitellogenin process-ing. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 149, 589–598.
Kleijn, D.P., Jansse, K.P., Martens, G.J., Herp, F., 1994. Cloning and expression of two crus-tacean hyperglycemic-hormone mRNAs in the eyestalk of the crayfish Orconecteslimosus. Eur. J. Biochem. 224 (2), 623–629.
Kuenzel, E.A., Mulligan, J.A., Sommercorn, J., Krebs, E.G., 1987. Substrate specificity deter-minants for casein kinase II as deduced from studies with synthetic peptides. J. Biol.Chem. 262 (19), 9136–9140.
Lee, F.Y., Chang, C.F., 1999. Hepatopancreas is the likely organ of vitellogenin synthe-sis in the freshwater prawn, Macrobrachium rosenbergii. J. Exp. Zool. 284 (7),798–806.
Lee, C.Y., Watson, R.D., 1995. In vitro study of vitellogenesis in the blue crab (Callinectessapidus): site and control of vitellin synthesis. J. Exp. Zool. 271 (5), 364–372.
Li, H.D., 2004. Nutrition on Embryonic Development of Redclaw Crayfish, Cheraxquadricarinatus. (Master degree thesis). Hua Dong Normal University of China,Shang Hai.
Livak, K.J., Schmittgen, T.D., 2001. Analysis of relative gene expression data usingreal-time quantitative PCR and the 2 (−Delta Delta C (T)). Method. Methods25, 402–408.
Matozzo, V., Gagne, F., Marin, M.G., Ricciardi, F., Blaise, C., 2008. Vitellogenin as a biomark-er of exposure to estrogenic compounds in aquatic invertebrates: a review. Environ.Int. 34 (4), 531–545.
Please cite this article as: Bai, H., et al., Molecular characterization and dMacrobrachium nipponense and the effects of RNA ..., Gene (2014), http://
ED P
RO
OF
Mayadas, T.N.,Wagner, D.D., 1992. Vicinal cysteines in the pro-sequence play a role in vonWillebrand factor multimer assembly. Proc. Natl. Acad. Sci. U. S. A. 89, 3531–3535.
Meggio, F., Pinna, L.A., 1988. Phosphorylation of phosvitin by casein kinase-2 provides theevidence that phosphoserines can replace carboxylic amino acids as specificity deter-minants. Biochim. Biophys. Acta 971 (2), 227–231.
Mekuchi, M., Ohira, T., Kawazoe, I., Jasmani, S., Suitoh, K., Kim, Y.K., Jayasankar, V.,Nagasawa, H., Wilder, M.N., 2008. Characterization and expression of the putativeovarian lipoprotein receptor in the kuruma prawn, Marsupenaeus japonicus. Zool.Sci. 25 (4), 428–437.
Mouchel, N., Trichet, V., Betz, A., Le Pennec, J.P., Wolff, J., 1996. Characterization of vitello-genin from rainbow trout (Oncorhynchus mykiss). Gene 174 (1), 59–64.
Murmu, K., Sahu, N.P., Mallik, S.K., Reddy, A.K., Kohli, M.P.S., 2007. Rematuration of spentMacrobrachium rosenbergii female broodstock through dietarymanipulation and eye-stalk ablation. Isr. J. Aquacult. - Bamidgeh 59, 104–110.
Okumura, T., Yamano, K., Sakiyama, K., 2007. Vitellogenin gene expression and hemo-lymph vitellogenin during vitellogenesis, final maturation, and oviposition in femalekuruma prawn,Marsupenaeus japonicus. Comp. Biochem. Physiol. A Mol. Integr. Phys-iol. 147 (4), 1028–1037.
Okuno, A., Yang, W.J., et al., 2002. Deduced primary structure of vitellogenin in the giantfreshwater prawn, Macrobrachium rosenbergii, and yolk processing during ovarianmaturation. J. Exp. Zool. 292 (5), 417–429.
Phiriyangkul, P., Utarabhand, P., 2006. Molecular characterization of a cDNA encoding vi-tellogenin in the banana shrimp, Penaeus (Litopenaeus) merguiensis and sites of vitel-logenin mRNA expression. Mol. Reprod. Dev. 73 (4), 410–423.
Phiriyangkul, P., Puengyam, P., Jakobsen, I.B., Utarabhand, P., 2007. Dynamics of vitello-genin mRNA expression during vitellogenesis in the banana shrimp, Penaeus(Fenneropenaeus) merguiensis using real-time PCR. Mol. Reprod. Dev. 74 (9),1198–1207.
Quackenbush, L.S., 1989. Vitellogenesis in the shrimp, Penaeus vannamei: in vitrostudies of the isolated hepatopancreas and ovary. Comp. Biochem. Physiol. B 94(2), 253–261.
Raviv, S., Parnes, S., Segall, C., Davis, C., Sagi, A., 2006. Complete sequence ofLitopenaeus vannamei (Crustacea: Decapoda) vitellogenin cDNA and its expres-sion in endocrinologically induced sub-adult females. Gen. Comp. Endocrinol.145 (1), 39–50.
Roth, T.F., Porter, K.R., 1964. Yolk protein uptake in the oocyte of the mosquito Aedesaegypti. J. Cell Biol. 20 (2), 313–332.
Roth, Z., et al., 2010. N-glycan moieties of the crustacean egg yolk protein and their glyco-sylation sites. J. Glycoconj. 27 (1), 159–169.
Sappington, T.W., Raikhel, A.S., 1998. Molecular characteristics of insect vitellogenins andvitellogenin receptors. Insect Biochem. Mol. Biol. 28 (5), 277–300.
Sappington, T.W., Oishi, K., Raikhel, A.S., 2002. Structural characteristics of insectvitellogenins. Prog. Vitellogenesis 12 (Part A), 69–101.
Schneider, W.J., 1992. Lipoprotein receptors in oocyte growth. Clin. Invest. 70 (5),385–390.
Smolenaars, M.M., Madsen, O., Rodenburg, K.W., et al., 2007. Molecular diversity and evo-lution of the large lipid transfer protein superfamily. J. Lipid Res. 48 (3), 489–502.
Soroka, Y., Milner, Y., Sagi, A., 2000. The hepatopancreas as a site of yolk protein synthesisin the prawn Macrobrachium rosenbergii. Invertebr. Reprod. Dev. 37 (1), 61–68.
Thompson, J.R., Banaszak, L.J., 2002. Lipid–protein interactions in lipovitellin. J. Biochem.41 (30), 9398–9409.
Tiu, S.H.K., Hui, J.H., He, J.G., Tobe, S.S., Chan, S.M., 2006a. Characterization of vitellogeninin the shrimp Metapenaeus ensis: expression studies and hormonal regulation ofMeVg1 transcription in vitro. Mol. Reprod. Dev. 73 (4), 424–436.
Tiu, S.H.K., Hui, J.H.L., Mak, A.S.C., He, J., Chan, S., 2006b. Equal contribution of hepatopan-creas and ovary to the production of vitellogenin (PmVg1) transcripts in the tigershrimp, Penaeus monodon. Aquaculture 254 (1), 666–674.
Tsang, W., Quackenbush, L.S., Chow, B.K.C., Tiu, S.H.K., He, J., Chan, S., 2003. Organizationof the shrimp vitellogenin gene: evidence of multiple genes and tissue specific ex-pression by the ovary and hepatopancreas. Gene 303, 99–109.
Tsutsui, N., et al., 2000. Molecular characterization of a cDNA encoding vitellogenin and itsexpression in the hepatopancreas and ovary during vitellogenesis in the kurumaprawn, Penaeus japonicus. Zool. Sci. 17 (5), 651–660.
Tsutsui, N., et al., 2004. Molecular characterization of a cDNA encoding vitellogenin in thecoonstriped shrimp, Pandalus hypsinotus and site of vitellogenin mRNA expression. J.Exp. Zool. A Ecol. Genet. Physiol. 301 (10), 802–814.
Tsutsui, N., Kim, Y.K., Jasmani, S., Ohira, T., Wilder, M.N., Aida, K., 2005. The dynamics ofvitellogenin gene expression differs between intact and eyestalk ablated kurumaprawn, Penaeus (Marsupenaeus) japonicus. Fish. Sci. 71 (2), 249–256.
Wahli, W., 1988. Evolution and expression of vitellogenin genes. Trends Genet. 4 (8),227–232.
Warrier, S., Subramoniam, T., 2002. Receptor mediated yolk protein uptake in the crabScylla serrata: crustacean vitellogenin receptor recognizes related mammalianserum lipoproteins. Mol. Reprod. Dev. 61 (4), 536–548.
Wilder, M.N., Okumura, T., Tsutsui, N., 2010. Reproductive mechanisms in Crustacea fo-cusing on selected prawn species: vitellogenin structure, processing and syntheticcontrol. Aqua Biosci. Monogr. 3, 73–110.
Wu, P., Qi, D., Chen, L.Q., et al., 2009. Gene discovery from an ovary cDNA library of orien-tal river prawn Macrobrachium nipponense by ESTs annotation. J. Comp. Biochem.Physiol. D 4 (2), 111–120.
Xie, S., 2009. The Early Development and Vitellin Levels of the Crayfish Procambarusclarkii. (Master degree thesis), Hebei University (June).
Xu, R.W., Jiang, J.P., 2001. A preliminary study on causes of sexual premature of Eriocheirsinensis. J. Zhejiang Ocean Univ. 3, 002.
Yang, H.J., 2007. Technical measures to control the precocious puberty in Macrobrachiumnipponense. J. Fish. 10, 48-48 (guide to be rich).
evelopmental expression of vitellogenin in the oriental river prawndx.doi.org/10.1016/j.gene.2014.12.008

693694695696697698699
700701702703704705706
707
10 H. Bai et al. / Gene xxx (2014) xxx–xxx
Yang, W.J., Ohira, T., Tsutsui, N., Subramoniam, T., Huong, D.T.T., Aida, K., Wilder, M.N.,2000. Determination of amino acid sequence and site of mRNA expression of fourvitellins in the giant freshwater prawn, Macrobrachium rosenbergii. J. Exp. Zool. 287(6), 413–422.
Yano, K., Sakurai, M.T., Izumi, S., Tomino, S., 1994. Vitellogenin gene of the silkworm,Bombyx mori: structure and sex-dependent expression. FEBS Lett. 356 (2),207–211.
UNCO
RRECT
Please cite this article as: Bai, H., et al., Molecular characterization and dMacrobrachium nipponense and the effects of RNA ..., Gene (2014), http://
Zhang, Y.P., Jiang, S.F., Xiong, Y.W., Sun, S.M., Qiao, H., Jin, S.B., Gong, Y.S., Fu, H.T.,2013. Molecular cloning and expression analysis of extra sex combs gene dur-ing development in Macrobrachium nipponense. Turk. J. Fish. Aquat. Sci. 13,331–340.
Zmora, N., Trant, J., Chan, S.M., Chung, J.S., 2007. Vitellogenin and its messenger RNA dur-ing ovarian development in the female blue crab, Callinectes sapidus: gene expression,synthesis, transport, and cleavage. Biol. Reprod. 77 (1), 138–146.
ED P
RO
OF
evelopmental expression of vitellogenin in the oriental river prawndx.doi.org/10.1016/j.gene.2014.12.008




























![Turbos de Geometría Variable (Vtg)[1][1]](https://img.dokumen.tips/doc/110x75/55cf8e9a550346703b93d857/turbos-de-geometria-variable-vtg11.jpg)
