Embed Size (px)
Citation preview

Ann. H i m . Cenet. (1994). 58. 275295 Printed in Great Britain
275
Genetic epidemiology of hereditary non-polyposis colorectal cancer syndromes in Modena, Italy: results of a complex segregation analysis
C. SCAPOLI', M. PONZ DE LEON2, R. SASSATELL12, P. BENATT12, L. RONCUCC12, A. COLLJNS3, N. E . MORTON3 AND I . BARRAI'
Department of Evolutionary Biology, University of Ferrara Zstituto di Patologia Medica, the Colorectal Cancer Study Group of the University of Modenu
and of Health Care District 16 CRC Research Group in Genetic Epidemiology, Department of Child Health,
Princess Anne Hospital, Southampton SO9 4HA
SUMMARY
Complex segregation analysis was conducted in a series of patients with hereditary non- polyposis colorectal cancer (HNPCC) ascertained through probands registered in the Cancer Registry of the Health Care District of Modena in Northern Italy. Altogether there were 71 nuclear families segregating for HNPCC in 28 pedigrees. The analysis favoured the two-loci model, in which the segregation a t the major locus is compatible with codominant transmission with a frequency of 0.0044 for the high-risk allele for HNPCC and a lifetime penetrance of 0.728 for heterozygotes.
INTRODUCTION
There is now considerable evidence from epidemiological, familial and molecular studies to support a contribution of genetic factors in the etiology of colorectal cancer (Bishop & Thomas, 1990). The inherited susceptibilities to colorectal cancer are usually studied separately for the polyposis and non-polyposis syndromes. Neoplasms may develop in Familial Adenomatous Polyposis (FAP), which is associated with the presence of large numbers of adenomatous polyps throughout the colon. FAP and related syndromes show features of autosomal dominant transmission (Graham et al. 1978, 1988; Naylor & Lebenthal, 1980; Lipkin et al. 1981 ; Vena et al. 1984). An autosomal dominant basis has also been described for Hereditary Non-Polyposis Colorectal Cancer (HNPCC) (Lynch et al. 1988a, b ) , a condition in which neoplasms develop in the absence of large numbers of polyps.
HNPCC, the object of the present study, can be divided into two subcategories: Lynch syndrome I, which predisposes specifically to colon cancer, and Lynch syndrome 11, in which there is a predisposition to colorectal, endometrial and stomach cancers (Lynch et al. 1985). The distinction between the two clinical entities has been questioned by some authors (Mecklin et al. 1986).
In addition to FAP and HNPCC, familial aggregates of colorectal (and other) tumours have frequently been reported and usually attributed to polygenic inheritance (Lovett, 1976 ; Rozen et al. 1987a, b) . Lynch and coworkers suggested that about 5 % of all colon cancers were due to hereditary monogenic factors, that approximately 10-15 % could be attributed to polygenic inheritance and that the remainder were truly sporadic (Lynch et al. 1985).
In a study of classic segregation analysis conducted on 605 families of probands with colorectal carcinoma, ascertained through the Cancer Registry of the Health Care District of

276 C. SCAPOLI AND OTHERS
Modena in Northern Italy, which included several families classified as suspected HNPCC, segregation was compatible with dominant transmission of susceptibility to carcinoma due to a major gene with greatly reduced penetrance in heterozygotes (Ponz de Leon et al. 1992).
In the present work we have analysed, with the method of complex segregation analysis (Morton ef al. 1983), 28 pedigrees with HNPCC which were selected during the period 1984-9 through the activity of the Colorectal Cancer Registry (CCR) of Modena (Ponz de Leon et al. 1992).
SUBJECTS AND METHODS
The study was carried out in the Health Care District of Modena in Northern Italy on patients listed or referred to the CCR of Local Health District 16 during the period 1984-9.
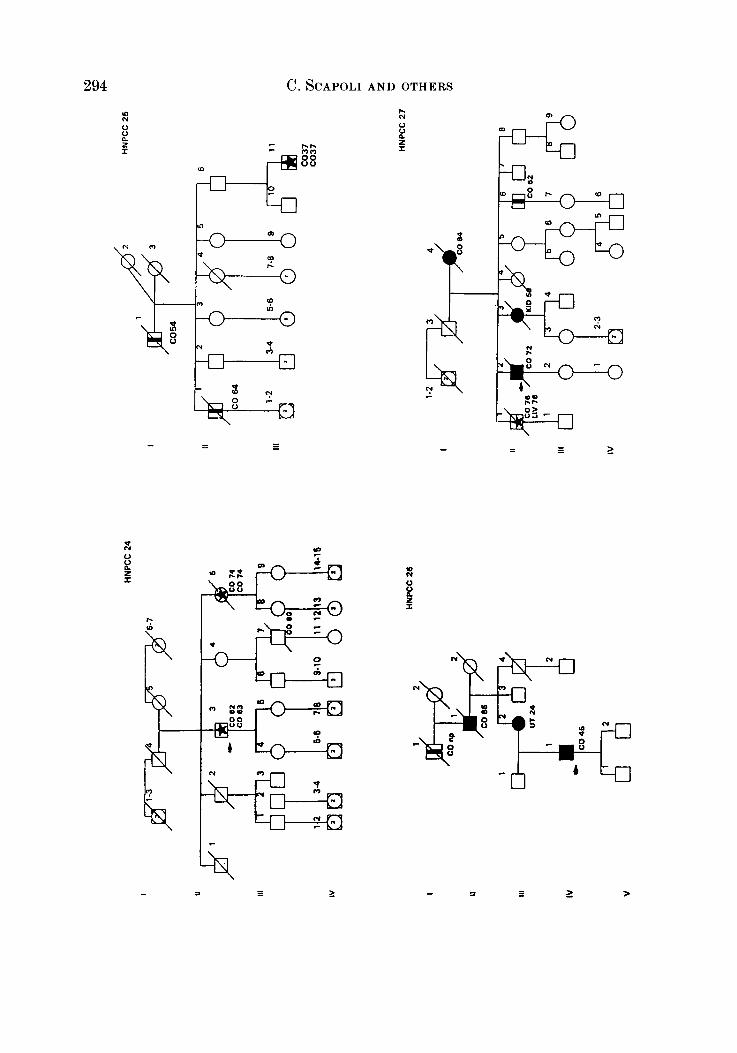
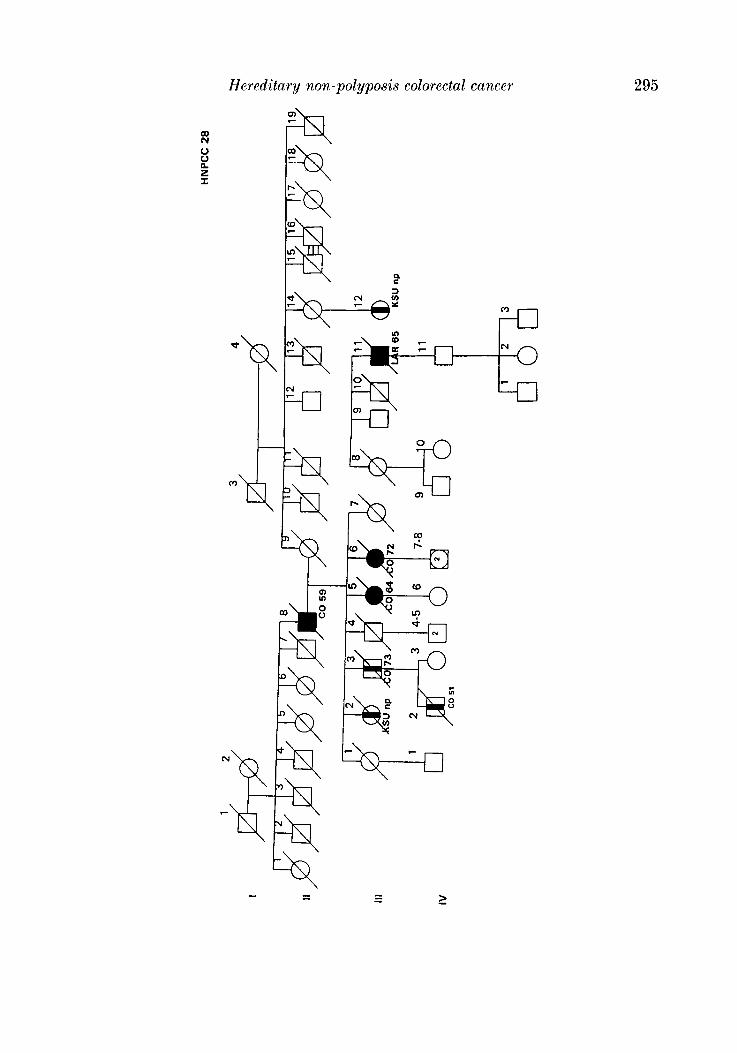
In the district there is only one Institute of Pathology to which all the biopsies from the various hospitals (and private clinics) are sent for histological diagnosis. In this way consecutive patients undergoing treatment for colorectal cancer (histologically proven in 93.5 % of cases) a t any centre of the district were admitted in the CCR and considered probands or index cases. During hospitalization, probands and/or their close relatives were interviewed, and besides personal, clinical and pathological data, an accurate genealogical tree was traced for each patient in which the main causes of morbidity and mortality among first-degree relatives were recorded. Colorectal cancer in relatives was verified from death certificate and/or hospital records (66.2 %). With this procedure we selected 28 pedigrees that satisfied criteria defined by the International Collaborative Group (Vasen et al. 1991) with slight modifications (Benatti et nE. 1993; Ponz de Leon et al. 1993), and appeared to segregate for suspected HNPCC (see Appendix). Overall, in these 28 segregating pedigrees there were 71 nuclear families and 553 individuals.
The criteria of inclusion were : ( 1 ) at least 3 relatives should have histologically verified colorectal cancer and one of them should be a first-degree relative to the others; (2) Familial Adenomatous Polyposis should be excluded; (3) a t least two successive generations should be affected; (4) in one of the relatives colorectal cancer should be diagnosed under 50 years of age.
Pedigree members for which the proband had no knowledge of the medical status were classified as unknown phenotype; all other individuals were taken to be unaffected. Age a t death of persons affected by tumours was taken, if not exactly known, to be a t the time of last observation.
Out of 553 individuals only 1 person was younger than 20 (he was 19) and he was added to the age group 20-34.
The HNPCC pedigrees were partitioned into nuclear families, and sibships admitted in the sample were of four types.
Type 1 : index sibships including the proband as an offspring (single selection). Type 2 : children of proband (complete selection). Type 3 : the sibships of the parents of proband (truncate selection). Type 4 : every other nuclear family with members affected by colorectal cancer and
ascertained in the pedigree of a proband (truncate selection). Kuclear families classified according to these rules were submitted to the computer programs
POINTER (Morton et al. 1983) and COMDS (Morton et al. 1991). The programs were transported to the CINECA in Italy and are accessible upon request to one of us (C.S.).

Hereditary non-polyposis colorectal cancer 277
Pedigrees were consecutively numbered for identification purposes. Since vertical trans- mission was one of the criteria of classification of colorectal cancer as suspected HNPCC, each family was conditioned on multiple affected. For nuclear families including the proband (types 1 and 2), when both a parent and a sib were classified as affected, we took as pointer ( a ) for odd- numbered pedigrees the affected parent and ( b ) for even-numbered pedigrees one affected sib. In this way we chose at random the pointer in the families, avoiding ascertainment bias. For the other families (types 3 and 4) pointers were chosen among the closest affected relatives (Morton et al. 1983).
Each individual was assigned, taking into account specific mortality, to one of eight liability classes based on sex and age. The morbid risk ( R ) of colorectal cancer attributed to t h e j t h liability class was defined as:
where I j is the cumulative incidence to the midpoint and Mj-l is the cumulative specific mortality to the end of the preceding class (Iselius et al. 1991 ; Morton, 1991 ; Shields et al. 1994).
Therefore Rj is the probability that a random individual observed in the j th class (dead or alive) be affected. The incidence ( I ) and mortality ( M ) of colorectal cancers were derived, for the period 1984-9, from the Colorectal Cancer Registry of Local Health District 16 of Modena (Ponz de Leon et al. 1991).
Besides the liability classes, we defined two new variables that were Severity, specified only for individuals affected by HNPCC, and Diathesis, specified only if the subject was not affected by this cancer (Morton et al. 1991, p. 310). Each individual, taking into account these subdivisions, was assigned to one of eight severity (or diathesis) classes. In the first four classes of severity, corresponding to four different age groups, were classified individuals that did not have a second cancer beside HNPCC; and in the second four classes the individuals having a second cancer, always in the same four age classes. Similarly, for diathesis, in the first four classes, corresponding to the same four age groups, were included unaffected individuals, and in the second four classes patients having a non-HNPCC cancer.
Thus each individual, taking into account these subdivisions, was assigned to one of eight severity (or diathesis) classes depending on the frequency of the age group of the subject, and on the risk of being affected or not affected by cancer.
In this calculation of the risk to develop cancer, independently from the site of the tumour, we used data from the nearby Parma province for 1978-82 (I.A.R.C., 1987) for the incidence and data referred to the whole of Italy in 1985 (W.H.O., 1988) for mortality and applied the same formula (Equation (1)) used for colorectal cancer risk. Since we know the risk of having a colorectal cancer Rc0,*, and the risk of having a generic cancer we obtained, for the age group j, the risk of having any, except colorectal, cancer namely R,,,,, as :
where Rco,* and Rall,j was the average risk between males and females belonging to the age groupj. In summary, the probability of an individual belonging to a j t h diathesis (or severity) class, irrespectively of sex, depends on the frequency of the age group the subject belongs to, and on the risk to be ( R ) or not to be ( 1 - R ) affected, where R = R,,,,, for diathesis and R = Rall,j for severity.

278 C. SCAPOLI AND OTHERS
In segregation analysis the parameters of the model, gene frequency (q ) , major locus dominance ( d ) , displacement between homozygotes ( t ) , polygenic heritability ( H ) , diathesis (B) and severity (8) were estimated by maximizing the likelihood of the phenotypes of the family. Both the probability density of offspring phenotypes conditional on parental phenotypes (conditional likelihood approach) and the joint probability density of parent-offspring phenotypes (joint likelihood approach) were investigated in POINTER. In COMIIS both probit and logit models are implemented (Morton et al. 1991), but for the two-loci model only the probit model (the same as POINTER) under conditional likelihood was explored.
The maximum, or general, likelihood was calculated by estimating all parameters simultaneously. This is called the full model. To test competing hypotheses, minus twice the log likelihood ( - 2 1nL + C) calculated under the full model is subtracted from the likelihood obtained when the values of one or more parameters are assigned under hypothesis. This difference is distributed as a x2 with degrees of freedom equal to the number of parameters assigned.
RESULTS
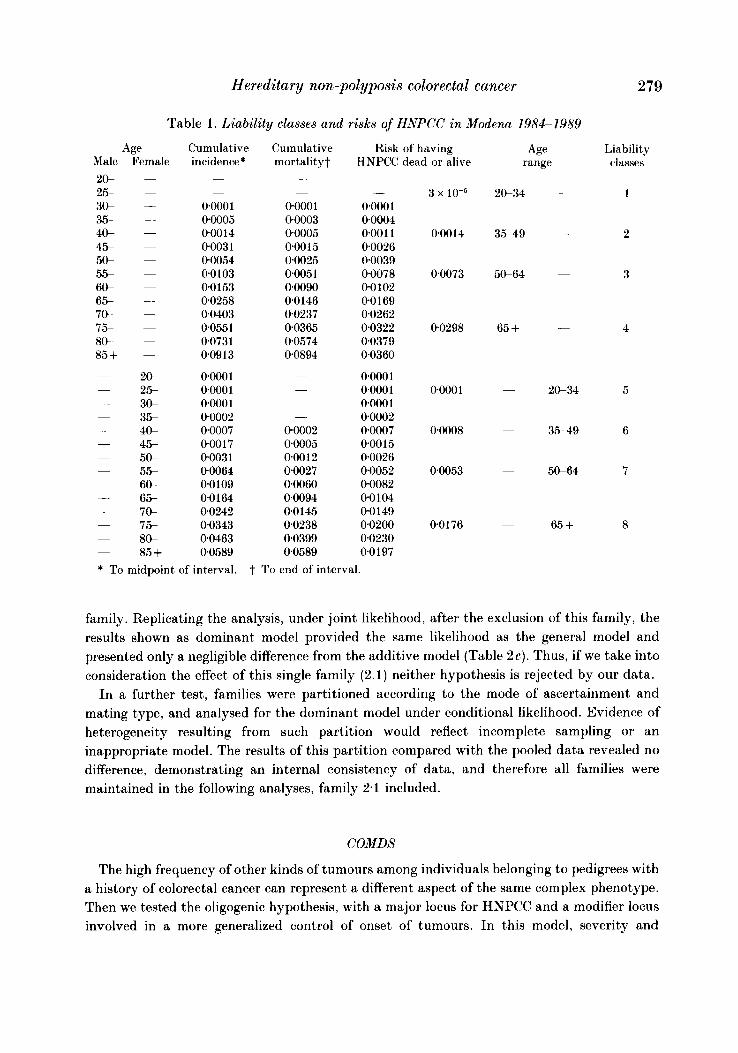
Table 1 shows the risk, subdivided by sex, of having colorectal cancer for 14 age groups. The risks have been grouped in 8 liability classes (4 for men and 4 for women) by averaging the values belonging to the different age classes. This risk and the risk of having any other tumour was subsequently used to estimate the probability for a given individual to belong to a specific diathesis (or severity) class, as defined in Subjects and Methods.
POINTER
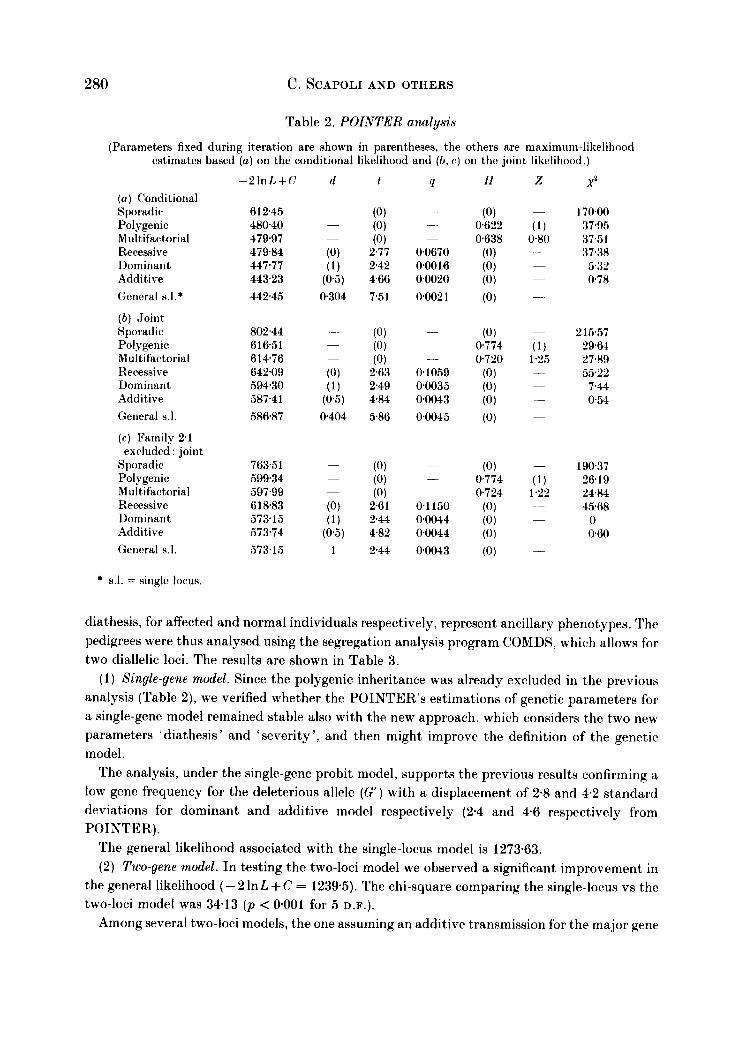
The results of the complex segregation analysis of 71 nuclear families using POINTER are given in Table 2. We recall that cancers simultaneously present with HNPCC are not considered in this procedure, but only in COMDS.
(1) Conditional likelihood (Table Za) . The model which does not make provision for family resemblance, in which the familial occurrence of colorectal cancer can be attributed to chance, i.e. sporadic, is strongly rejected (x2c31 = 170; p 4 0.001).
All models incorporating a major gene (q > 0) for genetic transmission of HNPCC show better likelihoods than either polygenic or multifactorial models. However, the model fitting a recessive major gene is always rejected.
Among the monogenic models, the additive one results in a likelihood not significantly different from the general single-gene model (x2L11 = 0.78).
( 2 ) Joint likelihood (Table 2b). The analysis under joint likelihood gives results which can be superimposed on those obtained under conditional likelihood.
The finding under POINTER indicates a better fit for the additive model. Several previous studies of the genetics of colorectal cancer were compatible with autosomal dominant transmission for HNPCC (Lynch et al. 1988a, b ) . This was suggested also by a recent investigation based on classical segregation analysis (Ponz de Leon et al. 1992). Thus, to investigate our results further we compared the two Mendelian hypotheses, of additivity and dominance, by analysing the likelihood ratio of each family. From the comparison it emerged that the difference between the two models could be ascribed almost completely to a single
Male 20- 25- 3 s 35- 40- 45- 50- 55- 60- 65- 70- 75- 80- 85 + -
-
-
~
-
-
-
~
-
-
-
-
~
-
Hereditary non-polyposis colorectal cancer
Table 1 . Liability classes and risks of HNPCC in Modena 1984-1989
Age Cumulative Cumulative Risk of having Female incidence* mortality? HNPCC dead or alive
~
-
0~000 1 00005 000 14 0003 1 00054 00103 00153 00258 00403 0.0551 00731 009 13
00001 00001 0~000 1 00002 00007 00017 0003 1 00064 00109 00164 00242 00343 00463 00589
-
-
0000 1 00003 00005 0.00 15 00025 00051 00090 00146 00237 00365 00574 0.0894 -
-
~
-
00002 0*0005 oao 12 00027 00060 00094 00145 0.0238 00399 00589
~
-
0.000 1 00004 0001 1 0.0026 0.0039 00078 00102 00169 0.0262 00322 00379 0.0360
0.000 1 0.000 1 0000 1 00002 00007 0.001 5 0.0026 0.0052 00082 00104 0 0 149 0.0200 0.0230 0.0197
3 x 10-5
000 14
00073
00298
00001
00008
00053
00176
Age range
20-34
3.549
50-64
65 +
~
-
-
~
-
-
~
-
20-34
3.549
5&64
65 +
279
Liability classes
1
2
3
4
5
6
7
8
* To midpoint of interval. t To end of interval.
family. Replicating the analysis, under joint likelihood, after the exclusion of this family, the results shown as dominant model provided the same likelihood as the general model and presented only a negligible difference from the additive model (Table 2c) . Thus, if we take into consideration the effect of this single family (2.1) neither hypothesis is rejected by our data.
In a further test, families were partitioned according to the mode of ascertainment and mating type, and analysed for the dominant model under conditional likelihood. Evidence of heterogeneity resulting from such partition would reflect incomplete sampling or an inappropriate model. The results of this partition compared with the pooled data revealed no difference, demonstrating an internal consistency of data, and therefore all families were maintained in the following analyses, family 2.1 included.
COMDS
The high frequency of other kinds of tumours among individuals belonging to pedigrees with a history of colorectal cancer can represent a different aspect of the same complex phenotype. Then we tested the oligogenic hypothesis, with a major locus for HNPCC and a modifier locus involved in a more generalized control of onset of tumours. In this model, severity and
280 C. SCAPOLI AND OTHERS
Table 2. POINTER analysis
(Parameters fixed during iteration are shown in parentheses, the others are maximum-likelihood estimates based ( a ) on the conditional likelihood and ( b , c) on the joint likelihood.)
( a ) Conditional Sporadic, Polygenic Multifactorial Recessive Dominant Additive General s.l.*
( b ) ,Joint Sporadic Polygenic Multifactorial Recessive Dominant Additive General 8.1.
(c) Family 2.1
Sporadic Polygenic. Multifactorial Recessive Dominant Additive General 5.1.
exrluded : joint
- 2 I n L + C d
6 1 2.45 48040 479.97 479.84 447.77 443.23 442.45
802.44 61651 61476 642.09 59430 587.41 586.87
763.5 1 599.34 597.99 618.83 57315 573.74 573.15
Q
-
.- .
-
0.0670 0.0016 0.0020 0002 1
-
-
-
01059 00035 00043 00045
-
-
-
0 1 150 0.0044 0.0044 00043
X2
170.00 37.95 37.51 37.38 5.32 078
2 15.57 29.64 27.89 55.22
7.44 0.54
19037 26.19 24.84 45.68
0 060
* 8.1. = single locus.
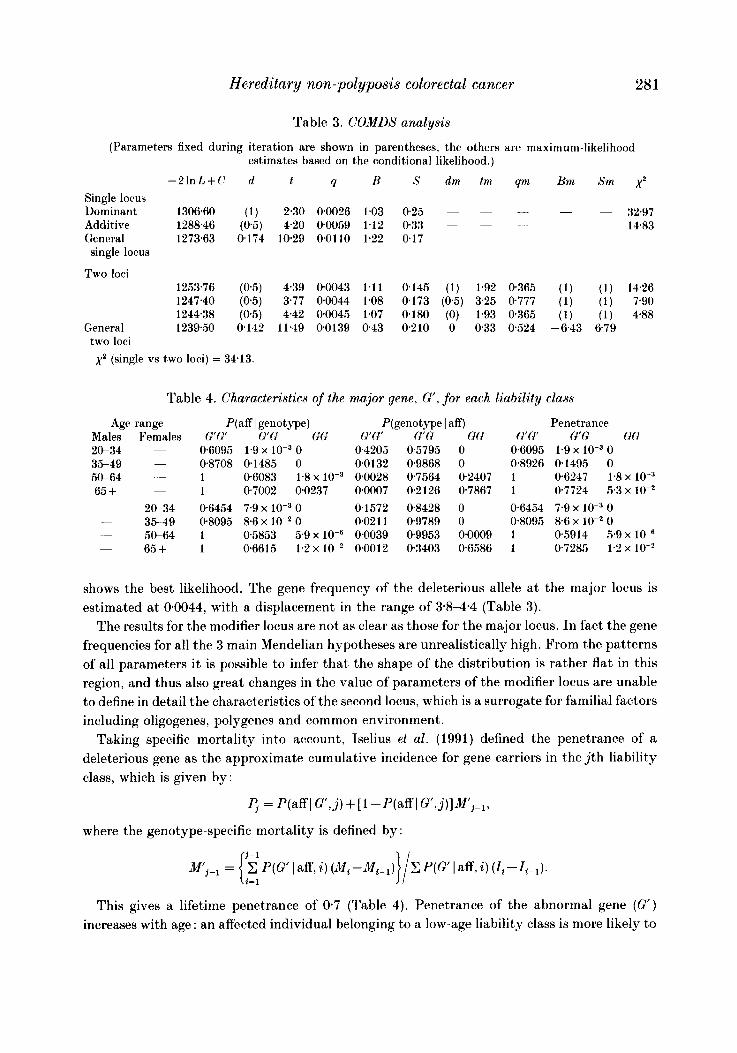
diathesis, for affected and normal individuals respectively, represent ancillary phenotypes. The pedigrees were thus analysed using the segregation analysis program COMDS, which allows for two diallelic loci. The results are shown in Table 3.
( 1 ) Single-gene model. Since the polygenic inheritance was already excluded in the previous analysis (Table 2), we verified whether the POINTER’S estimations of genetic parameters for a single-gene model remained stable also with the new approach, which considers the two new parameters ‘diathesis’ and ‘severity’, and then might improve the definition of the genetic model.
The analysis, under the single-gene probit model, supports the previous results confirming a low gene frequency for the deleterious allele (G‘) with a displacement of 2.8 and 4.2 standard deviations for dominant and additive model respectively (2.4 and 4.6 respectively from POINTER).
The general likelihood associated with the single-locus model is 1273-63. (2) Two-gene model. In testing the two-loci model we observed a significant improvement in
the general likelihood ( - 2 lnL+C = 1239.5). The chi-square comparing the single-locus vs the two-loci model was 34.13 ( p < 0.001 for 5 D.F.).
Among several two-loci models, the one assuming an additive transmission for the major gene
Hereditary non-polyposis colorectal cancer 281
Table 3. COMDS analysis
(Parameters fixed during iteration are shown in parentheses, the others are maximum-likelihood estimates based on the conditional likelihood.)
-2 lnL+C d t 9 B S dm tm qm Bni Sm x2 Single locus
- - Dominant 1306.60 (1) 2.30 00026 1.03 025 ~ ~ ~ 32.97 Additive 1288.46 (0.5) 4.20 00059 1.12 033 - - __ -
~ 14.83 General 1273.63 0174 10.29 00110 1.22 0.17 single locus
Two loci 1253.76 (0.5) 4.39 00043 1.11 0145 (1) 1.92 0365 (1) (1) 14.26 1247.40 (05) 3.77 0.0044 1.08 0.173 (0.5) 325 0777 (1) (1) 7.90 1244.38 (05) 442 00045 1.07 0.180 (0) 1.93 0365 (1) (1) 4.88
General 1239.50 0142 11.49 00139 043 0210 0 033 0524 -6.43 679 two loci x2 (single vs two loci) = 34.13.
Table 4. Characteristics of the
Age range Males Females 2G34 35-49 5&64 -
65 +
~
-
~
- 20-34 - 35-49 - 50-64 ~ 65 +
P(aff I genotype) G'G' G'G GG
0.6095 1.9 x 0 08708 01485 0 1 06083 1.8 x 1 07002 00237 06454 7.9 x 0 08095 8.6 x lo-' 0 1 0,5853 5.9 x lo-' 1 06615 1.2 x 10P
major gene, G', for each liability class
Qgenotype 14 G'G G G GG
0.4205 05795 0 00132 0.9868 0 00028 0,7564 02407 00007 0.2126 0,7867 0.1572 08428 0 0.0211 09789 0 0.0039 09953 0*0009 0.0012 03403 06586
Penetrance G'Q G'c: GO
0.6095 1.9 x 0 0.8926 01495 0 1 06247 1.8 x 1 0.7724 5.3 x lo-' 0.6454 7.9 x 0 0.8095 8.6 x lo-' 0 1 0.5914 5 9 x lo-' 1 0.7285 1.2 x lo-'
shows the best likelihood. The gene frequency of the deleterious allele a t the major locus is estimated at 00044, with a displacement in the range of 3.84 .4 (Table 3).
The results for the modifier locus are not as clear as those for the major locus. In fact the gene frequencies for all the 3 main Mendelian hypotheses are unrealistically high. From the patterns of all parameters i t is possible to infer that the shape of the distribution is rather flat in this region, and thus also great changes in the value of parameters of the modifier locus are unable to define in detail the characteristics of the second locus, which is a surrogate for familial factors including oligogenes, polygenes and common environment.
Taking specific mortality into account, Iselius et al. (1991) defined the penetrance of a deleterious gene as the approximate cumulative incidence for gene carriers in the j t h liability class, which is given by :
where the genotype-specific mortality is defined by :
This gives a lifetime penetrance of 0.7 (Table 4). Penetrance of the abnormal gene (G') increases with age : an affected individual belonging to a low-age liability class is more likely to

282 C. SCAPOLI AND OTHERS
have a putative colorectal cancer gene than an affected individual in a high-age liability class. For instance, an affected male aged between 35 and 49 has a 98% probability of carrying the deleterious allele (denoted by P(genotype I aff) in Table 4), compared with 21 for an aflect,ed male older than 65.
DISCUSSION
Several authors, and in particular Lynch and coworkers, made a substantial contribution to the definition of hereditary non-polyposis colon cancer (Lynch et al. 1988a, b ; Fitzgibbons et al. 1987).
In a recent investigation Ponz de Leon et al. (1992), using data derived from a population- based Registry, obtained further indications that colorectal carcinoma may be attributed to a t least two genetically heterogeneous entities : ( 1) an autosomal dominant gene with high penetrance, in a minor fraction of HNPCC families and (2) an autosomal dominant gene with very low penetrance, in a major fraction of families with individuals affected by cancers other than colon cancers. This result is in agreement with reports of Burt et al. (1985) and Cannon-Albright et al. (1988), who suggested the same genetic basis for susceptibility to apparently sporadic colorectal cancer and polyps.
In the present study the analysis of 28 HNPCC pedigrees with complex segregation analysis under the mixed model (POINTER), which tests whether the distribution of a complex phenotype is a consequence of a major locus or multifactorial inheritance, suggests the presence of a major gene that most likely is transmitted autosomally with intermediate dominance (Table 2 ) . This additive transmission is further supported by the results of the analysis of an oligogenic model from which, moreover, the presence of a t least a second gene with a modifying effect is indicated (xz = 3413 with five degrees of freedom; see Table 3).
The present set of data (see Appendix) is best fitted by an additive model for the major locus with an estimated gene frequency of 0.0044. This is close to the figure of 0.006 estimated by Houlston et al. (1992) and in surprising agreement with the estimation of 0.0052 obtained by Ponz de Leon et al. (1992). From Table 4 it appears that homozygote and heterozygote genotypes present a rather different lifetime penetrance, equal to 100% for G'G' and ranging between 72 YO and 77 YO for G G , and the difference becomes higher for the early age classes.
From the additive model, the estimation of the probability to be a carrier indicates that a t least 21% of the total burden of colorectal cancer in the general population is due to the heterozygous state of the gene in males. The percentage increases for females, where i t reaches 34%.
Recently Peltomiiki et al. (1993) and Aaltonen et al. (1993) provided evidence, in two large kindred with HNPCC, of a close linkage of predisposition for this disease with microsatellite markers of chromosome 2. The gene itself has recently been isolated. Previously, two genes tightly linked to FAP (APC and MCC) a t 5q21-22 were identified simultaneously by two groups (Groden et al. 1991 ; Joslyn et al. 1991 ; Kinsler et al. 1991 ; Nishisho et al. 1991), but causative germline mutations were seen only in APC.
Our results seem to support a two-loci model for the HNPCC syndrome. In addition to testing genetic transmission models, our results provide a more accurate estimate of risk for HNPCC for relatives of patients than the empirical risks, which ignore the etiology of the disorder. The genetic component of risk at different ages for relatives, obtained by using the estimated

Hereditary non-polyposis colorectal cancer 283
probability of inheriting the deleterious gene and the age-specific penetrance, indicates that for an early age of diagnosis the genetic risk to offspring is high, while it decreases with increasing age at diagnosis (Table 4).
This work was supported by grants of 60% and 40% from the Ministry of Universities and Scientific and Technological Research.
REFERENCES
AALTONEN, L. A., PELTOMAKI, P., LEACH, F. S. et al. (1993). Clues to the pathogenesis of familial colorectal cancer. Science 260, 812-816.
BENATTI, P., SASSATELLI, R., RONCUCCI, L. et al. (1993). Tumour spectrum in hereditary non-polyposis colorectal cancer (HNPCC) and in families with ‘suspected HNPCC’. A population-based study in Northern Italy. Int. J . Cancer 54, 371-377.
BISHOP, D. T. & THOMAS, H. J . W. (1990). The genetics of colorectal cancer. Cancer Surveys 9, 585-604. BURT. R. W . , BISHOP, D. T.. CANNON, L. A. el al. (1985). Dominant inheritance of adenomatous colonic polyps
CANNON-ALBRIGHT, L. A., SKOLNICK, M. H., BISHOP, T. et al. (1988). Common inheritance of susceptibility t>o
FITZGIBBONS, JR R. J.. LYNCH, H. T., STANISLAV, G. V. et al. (1987). Recognition and treatment of patients
GRAHAM, S., DAYAL, H., SWANSON, M. et al. (1978). Diet in the epidemiology of cancer of the colon and rectum.
GRAHAM, S., MARSHALL, *J., HAUQHEY, B. et al. (1988). Dietary epidemiology of cancer of the colon in western
GRODEN, J., THLIVERIS, A., SAMOWITZ, W. et a l . (1991). Identification and characterization of the familial
HOULSTON, R. S., COLLINS, A., SLACK, J. et al. (1992). Dominant genes for colorectal cancer are not rare. Ann.
INTERNATIONAL AGENCY FOR RESEARCH ON CANCER (1987). Cancer incidence in five continents, pp. 566-569.
ISELIUS, L., SLACK, J., LITTLER, M. et al. (1991). Genetic epidemiology of breast cancer in Britain. Ann. Hum.
JOSLYN, G., CARLSON, M., THLIVERIS, A. et al. (1991). Identification of deletion mutations and three new genes
KINSLER, K. W., NILBERT, M. C., LI-KUO, S. U. e f al. (1991). Identification of FAP genes from chromosome
LIPKIN, M., WINAWER, S. J. & SHERLOCK, P. (1981). Early identification of individuals a t increased risk for
LOVETT, E. (1976). Family studies in cancer of the colon and rectum. Br. J . Surg. 63, 13-18. LYNCH, H. T., WATSON, P., KRIEGLER, M. et al. (1988a). Differential diagnosis of hereditary nonpolyposis
LYNCH, H. T., WATSON, P., LANSPA, S. J . et al. (1988b). Natural history of colorectoral cancer in hereditary
LYNCH, H. T., KIMBERLING, W., ALBANO, W. A. et aJ. (1985). Hereditary nonpolyposis colorectal cancer (Lynch
MECKLIN, J. P., JARVINEN, H. J. & PELTOKALLIO, P. (1986). Cancer family syndrome. Genetic analysis of 22
MORTON, N. E. (1991). Pitfalls and prospects in genetic epidemiology of cancer. In Recent ProgresR in Genetic
MORTON, N. E., RAO, D. C. & LALOUEL, J . M. (1983). Methods in Genetic Epidemiology, ch. 5, pp. 62-98. Basel:
MORTON, N. E., SHIELDS, I). C. & COLLINS, A. (1991). Genetic epidemiology of complex phenotypes. Ann. Hum.
NAYLOR, E. W. & LEBENTHAL, E. (1980). Gardner’s syndrome: recent developments in research and
NISHISHO, I . , NAKAMURA, Y., MITOSHI, Y. et al. (1991). Mutations of chromosome 5q21 genes in FAP and
PELTOMAKI, P., AALTONEN, I,. A, , SISTONEN, P. et al. (1993). Genetic mapping of a locus predisposing to human
and colorectal cancer. N . Engl. J . Med. 312. 1540-1544.
colonic adenomatous polyps and associated colorectal cancers. N . Engl. J . Med. 319, 533-537.
with hereditary nonpolyposis colon cancer (Lynch syndromes I and 11). Ann. Surg. 206, 289-294.
J . Natn. Cancer Inst. 61, 70S714.
New York. A m . J . Epidemiol. 128, 490-503.
adenomatous polyposis coli gene. Cell 66, 589-600.
Hum. Genet. 56, 99-103.
Lyon, France.
Genet. 55 , 151-159.
a t the familial polyposis locus. Cell 66, 601-613.
5q21. Science 253, 661-665.
cancer of the large intestine. Part I : definition of high risk populations. Clin. Bull. 11, 13-21.
colorectal cancer (Lynch syndrome I and Lynch syndrome 11). Dis. Colon Rectum 31, 372-377.
nonpolyposis colorectal cancer (Lynch syndromes I and 11). Dis. Colon Rectum 31, 439-444.
syndromes I and 11). I . Clinical description of resource. Cancer 56, 934-938.
Finnish kindreds. Gastroenterology 90, 328-333.
Epidemiology of Cancer (ed. H . T. Lynch and P. Tautu), pp. 18-26. Heidelberg: Springer-Verlag.
Karger.
Genet. 55 , 301-314.
management. Dig. Dis . Sci. 25, 945-959.
colorectal cancer patients. Science 253, 665-669.
colorectal cancer. Science 260. 810-812.

284 C. SCAPOLI AND OTHERS
PONZ DE LEON, M., SASSATELLI, R., BENATTI, P. et al . (1993). Identification of hereditary nonpolysis colorectal cancer in the general population. Cancer 54, 3493-3501.
PONZ DE LEON, M., SASSATELLI, R., SCALMATI, A. et al. (1991). Epidemiology of Tumours of the Colon and Rectum: Incidence, Mortality and Survival in the Health Care District of Modena, 1984-1989, pp. 1-42. Modena : Artioli.
PONZ DE LEON, M., SCAPOLI, C., ZANGHIERI, G. et al. (1992). Genetic transmission of colorectal ('anoer: exploratory data analysis from a population-based registry. J . Med. Gen. 29, 531-538.
HOZEN, P . . FIREMAN, Z., FIGER, A. et al . ( 1 9 8 7 ~ ) . Family history of colorectal cancer as a marker of potential malignancy within a screening program. Cancer 60, 248-254.
ROZEN, P., LYNCH, H. T., FIGER, A. et al . (1987b). Familial colon cancer in the Tel-Aviv area and influence of ethnic origin. Cancer 60, 2355-2359.
SHIELDS, D. C., MARLOW, A. J., HOULSTON, R. S., ECCLES, D. M. & MORTON, N. E. (1994). Prediction of genetic risks from segregation analyses of morbid risks. Hum. Hered. 44, 52-55.
VASEN, H . F. A., MECKLIN, J. P. , MEERA KHAN, P. et al . (1991). The International Collaborative Group on Hereditary Non-Polyposis Colorectal Cancer (ICG-HNPCC). Dis. Colon Rectum 34, 424-425.
VENA, J. E., GRAHAM, S., ZIELEZNY, M. et al. (1984). Lifetime occupational exercise and colon cancer. Am. J . Epidemiol. 119, 1005-1014.
WORLD HEALTH ORGANIZATION (1988). World Health Statistics Annual, pp. 42&421. Geneva.
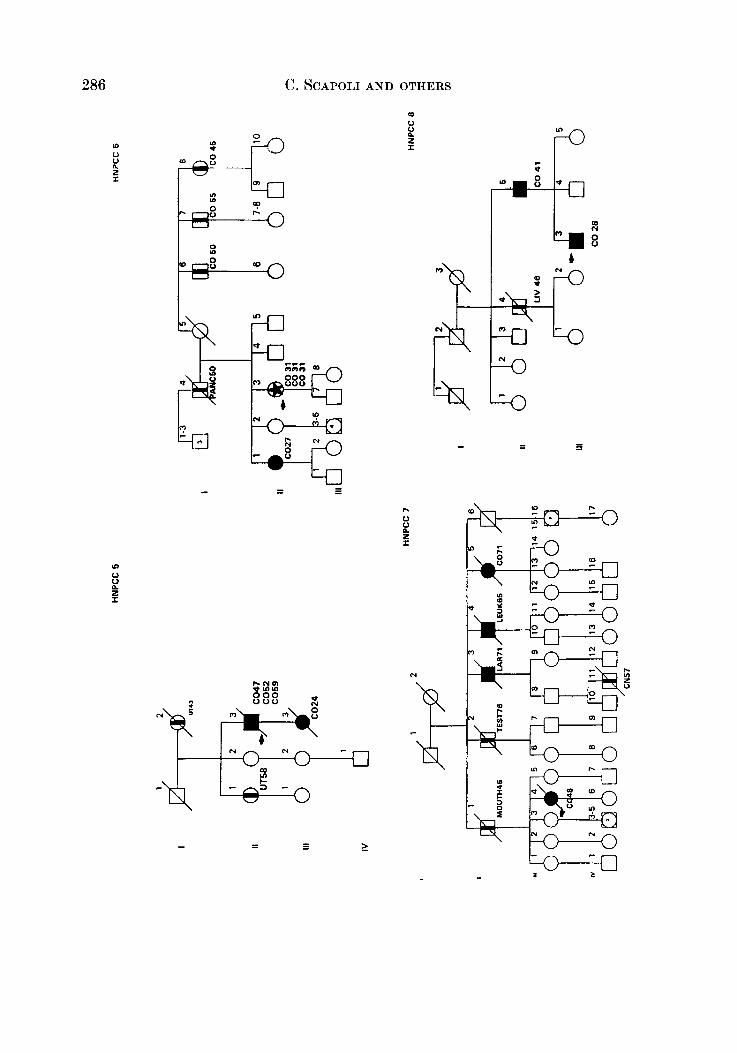
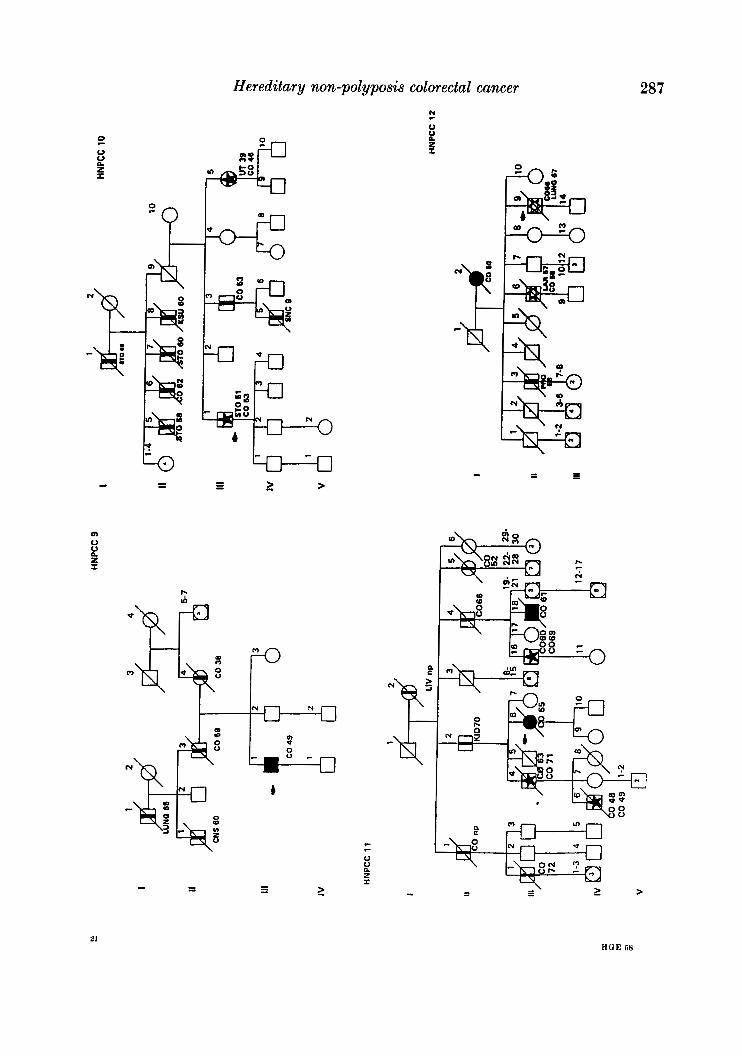
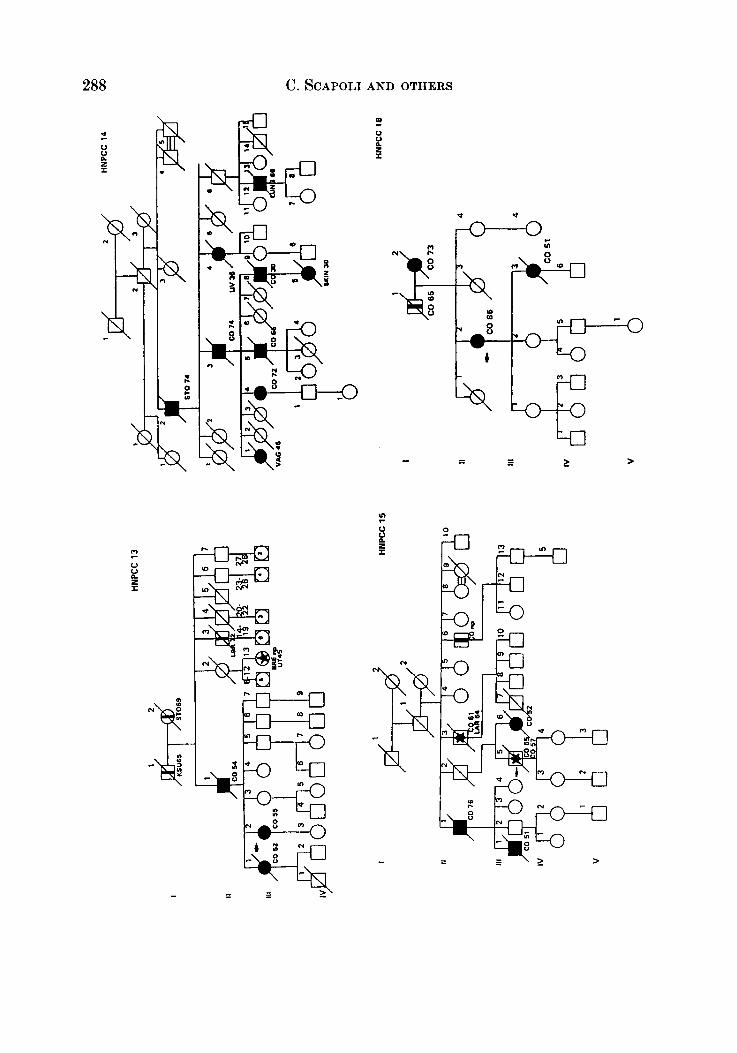
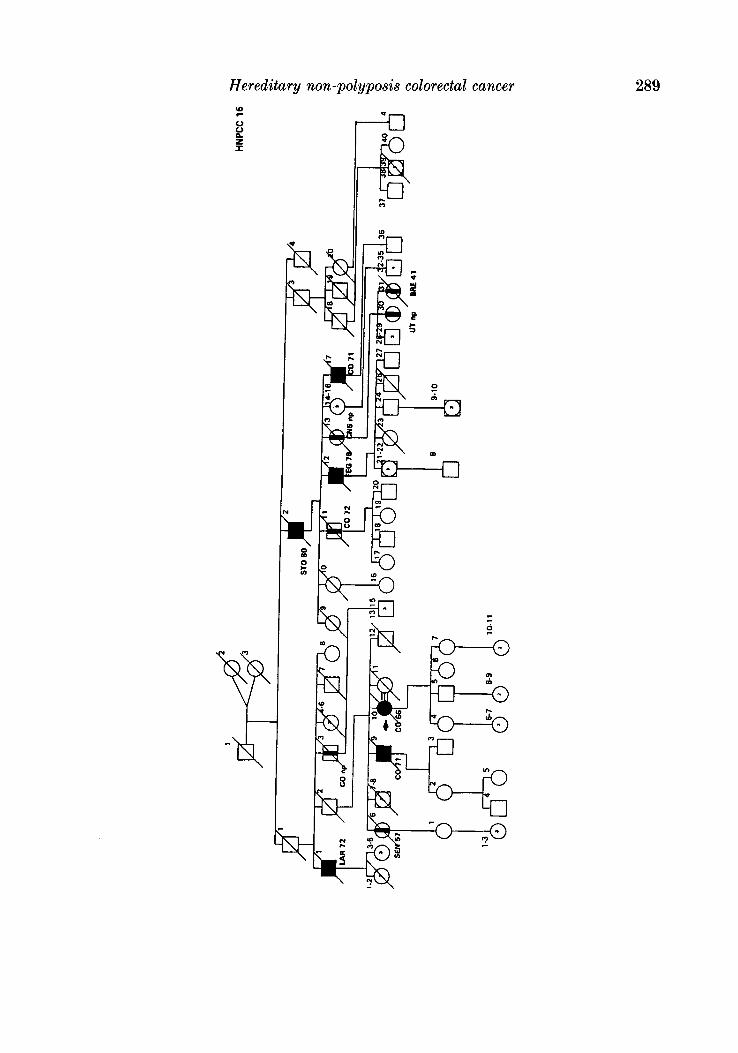
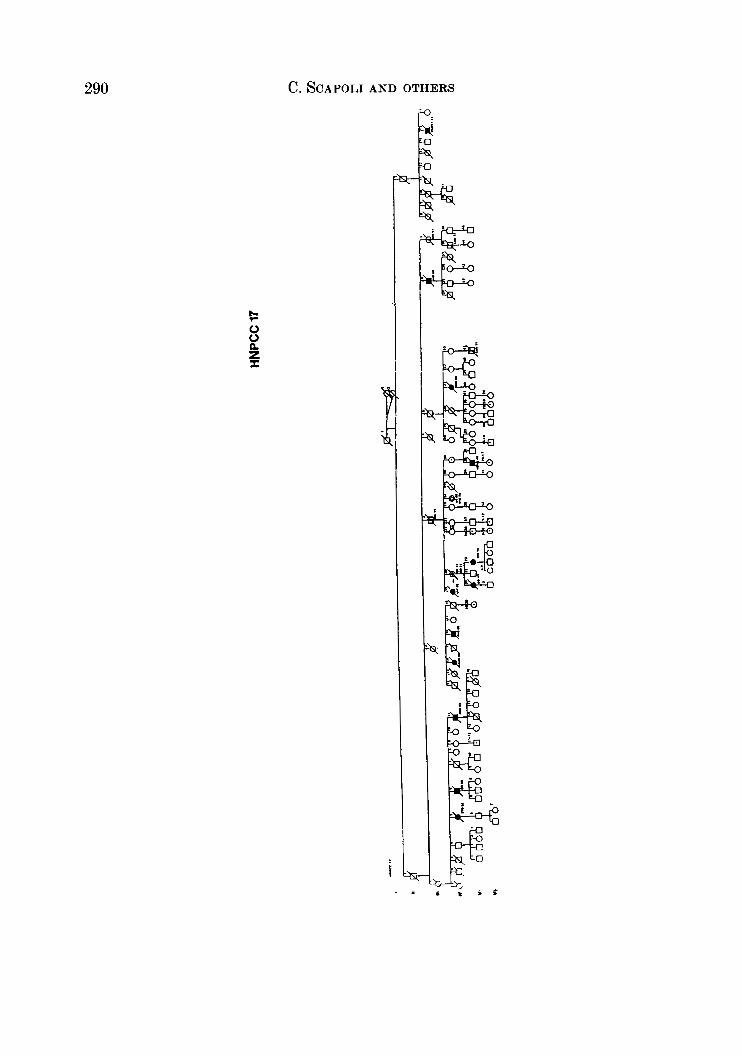
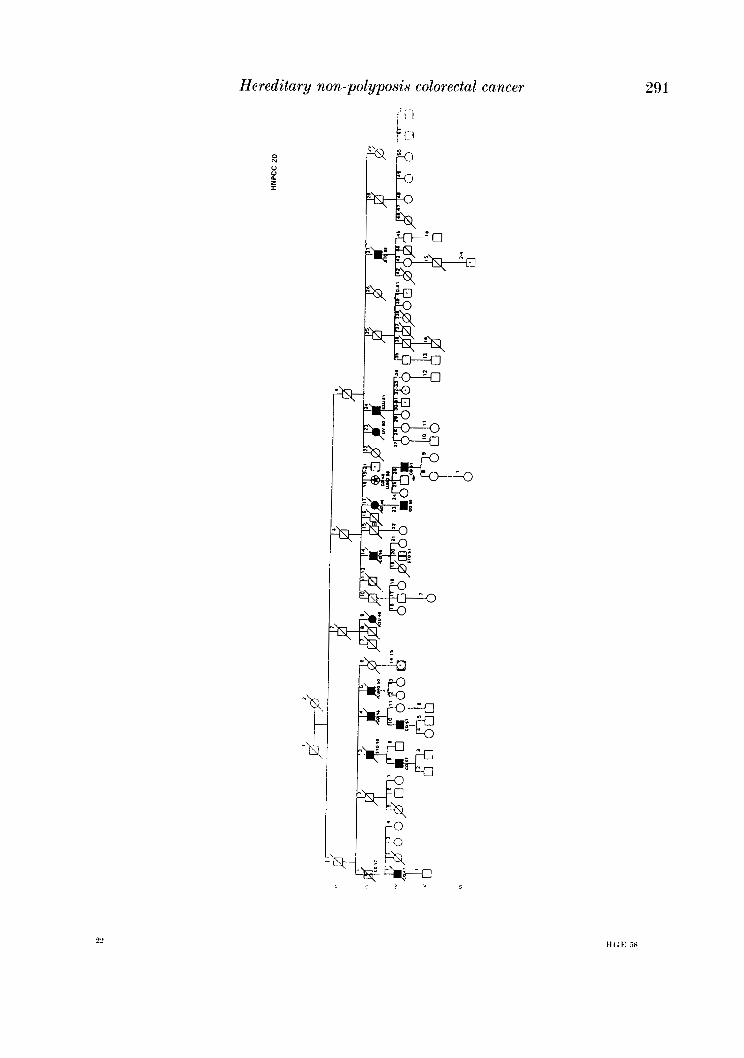
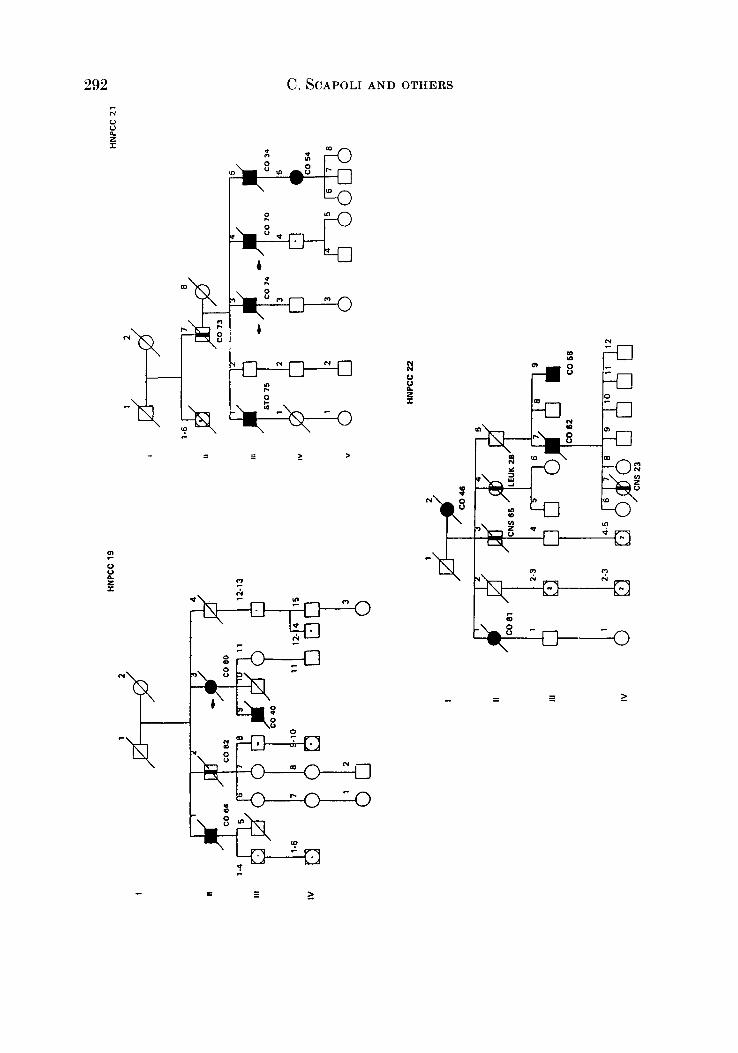
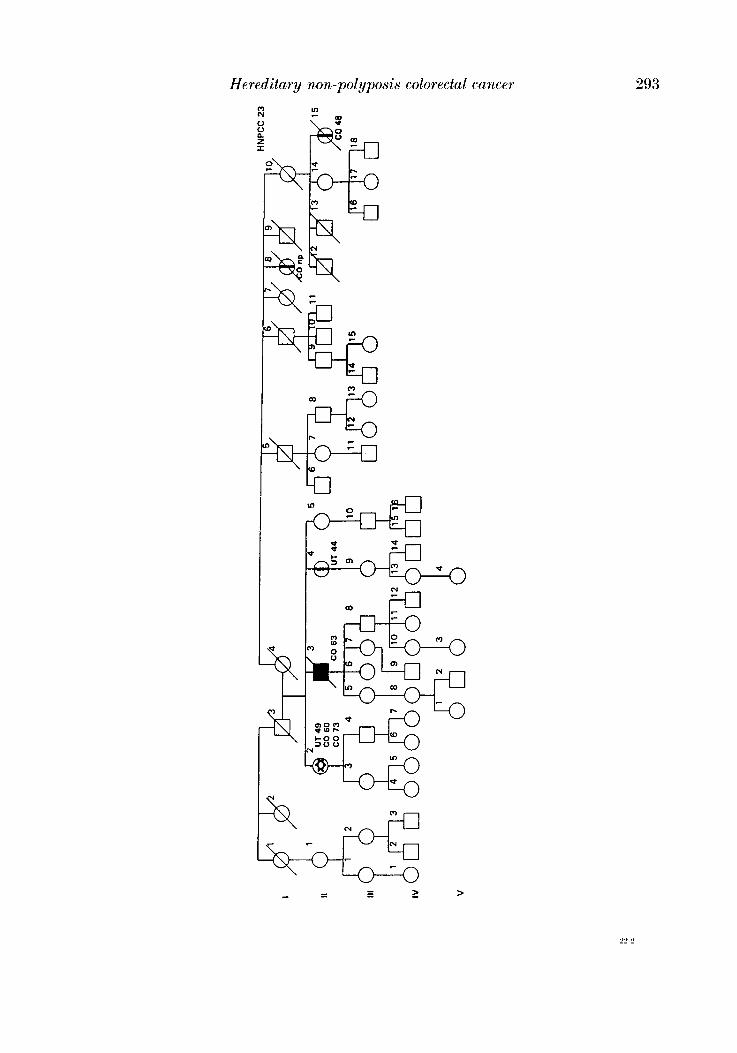
APPENDIX
LEQEND
MALE OR FEMALE 0 OR 0 UNAFFECTED BY CANCE R
MULTIPLE PRIMARY CANCERS VERIFIED & I @ 8 MALWPEMALE PROGENY
PROGENY EITHER SEX El * PRO BAND
TWINS: UNDEFINED ZYWSIS n
CANCER SITE
CO COLON RECTUM
8TO STOMACH
L A R L A R Y N X
UT UTERU8
BRE BREAST
THYR THYROID OLAND
KID KIDNBY AND
LIV LIVER
C N I CENTRALNERVOUI
URINARY TRACT
WlTEY
PRO PROITKTE
K I U I I T E UNKNOWN
MULTIPLE WIVES %
Hereditary non-polyposis colorectal cancer
0
285
c
&I- *
c N
D
286 C. SCAPOLI AND OTHERS
m
W
I : :
N
4 c
Hereditary non-polyposis colorectal cancer
i" N N
10
N
c
1 3
c r-
P
287
21 HCE 58
288
0 0
z I
.t d d
c In
0 n
N 8 -
4
\
c In
D
N
a 8 -
4
I
I 0
2 r P
2
I-- -*
Hereditary non-polyposis colorectal cancer In
0
z I
c
Y
v -&
289
290 C. SCAPOLI AND OTHERS
- .
Hereditary non-polyposis colorectal cancer 29 1
292 r N
C. SCAPOLI AND OTHERS
ol c u 2 I P
0
" * .
Hereditary non-polyposis colorectal cancer 293
m (D
’ 8
>
294 C. SCAPOLI AND OTHERS
b N
0 P I
0 N
0
2 I x
c-0; v v 0 0
>
Hereditary non-polyposis colorectal cancer
P
P' o n
I%-
%-
295





























