Embed Size (px)
Citation preview

GL
HGa
b
Gc
a
ARRA
KIHLO
1
stiect1w
Nc
1d
Journal of Clinical Virology 51 (2011) 170– 174
Contents lists available at ScienceDirect
Journal of Clinical Virology
jo u r n al hom epage: www.elsev ier .com/ locate / j cv
enetic diversity and antiviral drug resistance of pandemic H1N1 2009 inebanon
assan Zaraketa,∗, Hiroki Kondob, Carelle Tabetc, Rima Hanna-Wakimc, Yasushi Suzukib,hassan S. Dbaiboc, Reiko Saitob, Hiroshi Suzukib
Division of Virology, Department of Infectious Diseases, St Jude Children’s Research Hospital, 262 Danny Thomas Place, Mail Stop 330, Memphis, TN 38105, USADivision of Public Health, Department of Infectious Disease Control and International Medicine, Niigata University,raduate School of Medical and Dental Sciences, Niigata 951-8510, JapanDivision of Pediatric Infectious Diseases, Department of Pediatrics and Adolescent Medicine, American University of Beirut, Beirut, P.O. Box 113/6044/B21, Lebanon
r t i c l e i n f o
rticle history:eceived 11 January 2011eceived in revised form 3 April 2011ccepted 6 April 2011
eywords:nfluenza virus1N1 pdmebanonseltamivir resistance
a b s t r a c t
Background: In June 2009, the World Health Organization announced the 21st century’s first influenzapandemic caused by pandemic influenza H1N1 2009 (H1N1 pdm).Objectives: Our goal was to analyze antiviral drug resistance and the phylogenetic relationships amonghemagglutinin (HA) and neuraminidase (NA) genes of H1N1 pdm samples in Lebanon.Study design: Nasopharyngeal swabs were collected from 197 patients with influenza-like illness fromMay 2009 through January 2010. Of the 50 influenza A-positive samples, 30 were analyzed for antiviraldrug resistance by using in vitro susceptibility assays and cycling-probe real-time PCR. The HA and NAgenes were also analyzed.Results: The results of hemagglutination-inhibition assays confirmed that all 30 analyzed samples wereH1N1 pdm. In July 2009, community transmission of H1N1 pdm was detected in Lebanon, and an outbreakoccurred in October 2009. The outbreak cases were caused by a strain with 4 mutations in the NA gene(i.e., V42I, N68T, N248D, and E462K) and 2 mutations in the HA gene: 1 in the Ca1 antigenic site (i.e.,
S206T) and 1 in the Ca2 antigenic site (i.e., D225E). This strain was closely related to a major H1N1pdm cluster that was isolated worldwide. All 30 samples were amantadine-resistant, and none werezanamivir-resistant. The 1 oseltamivir-resistant sample appeared to be from a community-transmittedcase in an otherwise healthy 2-year-old child.Conclusion: Continuous monitoring of oseltamivir susceptibility among H1N1 pdm is essential to guidethe effective use of this drug.. Background
Influenza virus infection is a great burden on health, causingeasonal outbreaks that affect approximately 20% of the popula-ion each year.1 Pandemic viruses emerge periodically when annfluenza virus acquires genes to which humans have not beenxposed, resulting in increased morbidity and mortality. Pandemicsan result in high morbidity and mortality rates. For example,he 1918 Spanish influenza, the 1957 Asian influenza, and the968 Hong Kong influenza pandemics killed 25–55 million peopleorldwide.1,2
In April 2009, a new influenza virus emerged in Mexico andorth America3,4 and rapidly spread to more than 70 countries,ausing approximately 28,000 infections and 144 deaths before
∗ Corresponding author. Tel.: +1 901 595 8757; fax: +1 901 595 8559.E-mail address: [email protected] (H. Zaraket).
386-6532/$ – see front matter © 2011 Elsevier B.V. All rights reserved.oi:10.1016/j.jcv.2011.04.001
© 2011 Elsevier B.V. All rights reserved.
the World Health Organization announced it to be the 21st cen-tury’s first pandemic, known as pandemic influenza H1N1 2009(H1N1 pdm). This virus combines gene segments from North Amer-ican avian and classical swine influenza, human influenza, andEurasian swine influenza5; thus, it is distinct from the seasonalH1N1 influenza virus that was circulating in humans, enabling itto spread in the thus-far naive population.
The first 3 cases of H1N1 pdm in Lebanon were detected atBeirut International Airport on May 30, 2009.6 Although there is nonational influenza surveillance system in Lebanon, our group hasbeen conducting influenza surveillance in the country since 2007,which enabled us to collect samples during the first few months ofthe pandemic.
2. Objective
Our goal was to characterize antiviral drug susceptibility ofH1N1 pdm isolates in Lebanon and to determine the phylogenetic

linical Virology 51 (2011) 170– 174 171
r(o
3
3
sossautTaiHtto
3
tiaiAaRS
3
Tawn(saRSbA
3
ctflbSMwt
0
20
40
60
80
100
120
140
May Jun Jul Aug Sept Oct Nov Dec Jan
01029002
Num
ber o
f cas
es
Inf luen za AILI
Fig. 1. Graphical illustration of the influenza-like illness (ILI) cases and the influenza
H. Zaraket et al. / Journal of C
elationships among the hemagglutinin (HA) and neuraminidaseNA) genes of these isolates and their relationship to isolates fromther countries.
. Study design
.1. Sample collection
Between May 2009 and January 2010, two nasopharyngealwabs were collected from each patient at the infectious diseasesutpatient clinic at the American University of Beirut, who pre-ented with at least 1 of the following influenza-like illness (ILI)ymptoms: fever of at least 38 ◦C, cough, rhinorrhea, headache, orbdominal symptoms (i.e., diarrhea and vomiting). One swab wassed to screen for influenza A by using a rapid influenza detec-ion kit, Quick S-Influ A/B “Seiken” (Denka Seiken Tokyo, Japan).he second swab was placed in virus transport medium and storedt −80 ◦C until further analysis. A representative portion of thenfluenza A-positive swabs was shipped to the Division of Publicealth, Niigata University, Japan, for further analysis. The Institu-
ional Review Board of the American University of Beirut approvedhis study, and informed consent was obtained from each patientr patient’s guardian before sample collection.
.2. Virus isolation and characterization
Madin-Darby Canine Kidney (MDCK) cells were inoculated withhe swab suspension (100 �L) and incubated until the character-stic cytopathic effect could be observed. To determine the typend subtype of each isolate, we performed hemagglutination-nhibition (HI) assays using A/California/07/09 (H1N1 pdm),/Victoria/210/2009 (H3N2), B/Brisbane/60/2008 (influenza B)ntisera (Denka Seiken Tokyo, Japan), and guinea pig red blood cells.eference antigens for the antisera were purchased from Denkaeiken (Tokyo, Japan) and included as controls.
.3. RNA extraction, PCR, and sequencing
RNA isolation was performed by using Extragen II (Kainos,okyo, Japan). First-strand complementary DNA was then gener-ted by reverse transcription with U12 primers.7,8 HA and NA genesere amplified and sequenced as described previously.9 Phyloge-etic tree analyses were performed by using the MEGA programversion 4.0).10 Reference strains in the tree analyses includedome of the closest BLAST search hits for which both the HAnd NA genes’ sequences were available on the Influenza Virusesource Data Base (http://www.ncbi.nlm.nih.gov/genomes/FLU/).equences generated in this study were deposited in the Japan Gen-ank database (http://www.ddbj.nig.ac.jp/; accession numbers:B603583–AB603642).
.4. Antiviral drug susceptibility testing
In vitro amantadine susceptibility was tested by using 50% tissueulture inhibitory dose (TCID50) assays. The 50% inhibitory concen-rations (IC50) of oseltamivir and zanamivir were assayed by using auorescent-based neuraminidase inhibition assay with methylum-elliferone N-acetylneuraminic acid (MUNANA) as the substrate.11
ingle-nucleotide polymorphisms at codons encoding S31N in the2 gene fragment and H275Y (N1 numbering) in the NA gene,hich confer resistance to amantadine and oseltamivir, respec-
ively, were analyzed by using cycling-probe real-time PCR.9,12
A rapid test-positive cases detected in Lebanon from May 2009 through January2010.
4. Results
4.1. Virus isolation
From May 2009 through January 2010, a total of 197 cases of ILIsymptoms were screened in Beirut, Lebanon (Fig. 1). Of these, 50were influenza A-positive by rapid influenza detection kit; 7 wereisolated in July, 39 in October, and 4 in November 2009 (Fig. 1). Ofthese 50 influenza A-positive swabs, only 30 samples were shippedto Japan for further analysis due to the high shipping cost of thepandemic virus. All 30 samples were confirmed to be influenza AH1N1 pdm viruses by HI assay and HA-specific PCR and sequencingupon virus isolation in MDCK cells.
4.2. Phylogenetic analysis of the HA and NA genes
The HA and NA genes of the 30 virus isolates were sequencedto determine their phylogenetic relationships. In the HA phy-logeny, the Lebanese isolates and the reference strains could bedivided into 2 main clusters that could be distinguished by a Serto Thr mutation at residue 206 (H3 numbering) (Fig. 2).13 Cluster1 included viruses isolated in July and 1 virus each from Octoberand November. Cluster 2 harbored 25 of the 30 Lebanese isolates,and its subcluster possessed an additional D225E mutation thatwas detected only in the 21 Lebanese isolates obtained in Octo-ber and November. Isolates in this subcluster shared high sequencehomology (99.7–100%). The S206T and D225E mutations occur inthe HA receptor-binding domain and in antigenic sites Ca1 and Ca2,respectively (Fig. 3).
Three of the isolates belonging to cluster 1 in the HA tree, includ-ing one Lebanese isolate (09L-06), belonged to cluster 2 in the NAtree. Cluster 2’s lineage and a subset of cluster 1’s isolates (basedon the HA tree) had V42I and N248D mutations in their NA genes.Therefore, these 2 mutations in NA were selected before the S206Tmutation in HA. Of the 21 Lebanese isolates with the HA-D225Emutation, 18 clustered together in the NA gene tree and were nearlyidentical (>99% homology). These isolates shared 2 characteristicmutations, N68T and E462K, in their NA gene. A BLAST search ofthese isolates’ sequence showed that these mutations were unique(at least among the published sequences) to the Lebanese isolates,suggesting that this strain evolved locally or regionally and causeda local outbreak in October and November. Distribution of isolates
among clusters 1 and 2 (for either gene) did not relate to the monthof isolation, and isolates from different months intermingled inboth clusters in the HA and NA trees.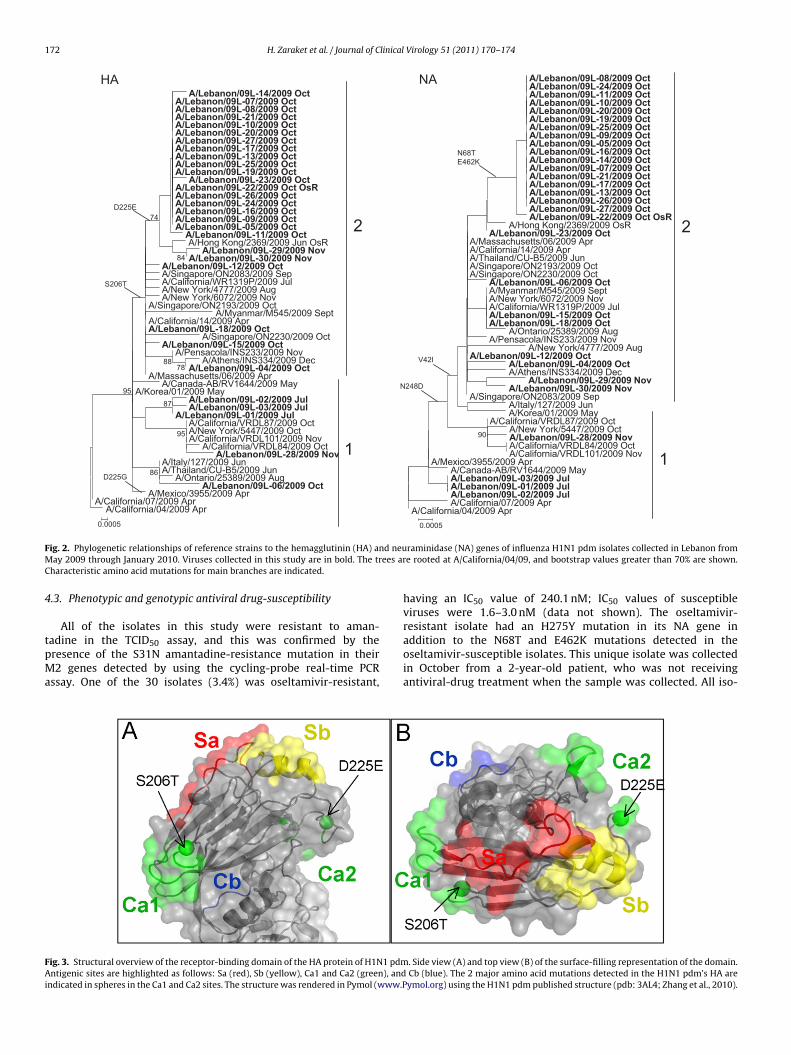
172 H. Zaraket et al. / Journal of Clinical Virology 51 (2011) 170– 174
A/Lebano n/09L- 08/2009 OctA/Lebano n/09L- 24/2009 OctA/Lebano n/09L- 11/2009 OctA/Lebano n/09L- 10/2009 OctA/Lebano n/09L- 20/2009 OctA/Lebano n/09L- 19/2009 OctA/Lebano n/09L- 25/2009 OctA/Lebano n/09L- 09/2009 OctA/Lebano n/09L- 05/2009 OctA/Lebano n/09L- 16/2009 OctA/Lebano n/09L- 14/2009 OctA/Lebano n/09L- 07/2009 OctA/Lebano n/09L- 21/2009 OctA/Lebano n/09L- 17/2009 OctA/Lebano n/09L- 13/2009 OctA/Lebano n/09L- 26/2009 OctA/Lebano n/09L- 27/2009 OctA/Lebano n/09L- 22/2009 Oct OsR
A/Hong Kon g/23 69/2009 OsRA/Lebano n/09L-23 /2009 Oc t
A/Massach usetts/06/200 9 AprA/Cali fornia/14 /200 9 AprA/Thailand/CU- B5/200 9 JunA/Singapore/ ON2193/20 09 Oc tA/Singapore/ ON2230/2 009 Oc t
A/Lebano n/09L-06 /2009 Oc tA/Myanmar/M545 /200 9 Sep tA/New York/6072 /20 09 NovA/Calif ornia/WR1319P/2 009 JulA/Lebano n/09L-15 /2009 Oc tA/Lebano n/09L-18 /2009 Oc t
A/O ntario/25389 /200 9 AugA/Pensacola/INS233 /200 9 Nov
A/New York/4777/20 09 Au gA/Lebano n/09L-12 /2009 Oc t
A/Lebano n/09L-04 /2009 Oc tA/Athens/INS334/2009 De c
A/Lebano n/09L- 29/2009 NovA/Lebano n/09L-30 /2009 Nov
A/Singapore/ ON2083/20 09 SepA/Italy/127/2009 JunA/Korea/01/ 2009 May
A/Calif ornia/VRDL87/2 009 OctA/New York/5447/20 09 OctA/Lebano n/09L-28 /2009 NovA/Calif ornia/VRDL84/2 009 OctA/Calif ornia/VRDL101/ 2009 Nov
A/Mexic o/3 955/200 9 AprA/Canada-AB/ RV16 44/2 009 MayA/Lebano n/09L-03 /2009 Ju lA/Lebano n/09L-01 /2009 Ju lA/Lebano n/09L-02 /2009 Ju lA/Cali fornia/07 /200 9 Apr
A/Cali fornia/04 /200 9 Apr
90
0.0005
HA NA
S206T
D225E
N68T E462K
N248D
2 2
11
V42I
A/Lebano n/09L-14 /2009 Oc tA/Lebano n/09L-07 /2009 Oc tA/Lebano n/09L-08 /2009 Oc tA/Lebano n/09L-21 /2009 Oc tA/Lebano n/09L-10 /2009 Oc tA/Lebano n/09L-20 /2009 Oc tA/Lebano n/09L-27 /2009 Oc tA/Lebano n/09L-17 /2009 Oc tA/Lebano n/09L-13 /2009 Oc tA/Lebano n/09L-25 /2009 Oc tA/Lebano n/09L-19 /2009 Oc t
A/Lebano n/09L-23 /2009 Oc tA/Lebano n/09L-22 /2009 Oct Os RA/Lebano n/09L-26 /2009 Oc tA/Lebano n/09L-24 /2009 Oc tA/Lebano n/09L-16 /2009 Oc tA/Lebano n/09L-09 /2009 Oc tA/Lebano n/09L-05 /2009 Oc t
A/Lebano n/09L-11 /2009 Oc tA/Hong Kon g/23 69/2009 Ju n Os R
A/Lebano n/09L-29 /2009 NovA/Lebano n/09L-30 /2009 Nov
A/Lebano n/09L-12 /2009 Oc tA/Singapore/ ON2083/20 09 SepA/Calif ornia/WR1319P/2 009 JulA/New York/4777/20 09 Au gA/New York/6072/20 09 Nov
A/Singapore/ ON2193/20 09 Oc tA/Myanmar/M545 /2009 Sep t
A/Cali fornia/14 /2009 AprA/Lebano n/09L-18 /2009 Oc t
A/Singa pore/ON2230 /20 09 OctA/Lebano n/09L-15 /2009 Oc t
A/Pensacol a/INS233 /2009 NovA/Athens/INS334/200 9 De c
A/Lebano n/09L-04 /2009 Oc tA/Massach usetts/06/200 9 Apr
A/Cana da-AB /RV16 44/2009 MayA/Korea/01/ 2009 May
A/Lebano n/09L-02 /2009 Ju lA/Lebano n/09L-03 /2009 Ju l
A/Lebano n/09L-01 /2009 Ju lA/Calif ornia/VRDL87/20 09 OctA/New York/5447/20 09 OctA/Calif ornia/VRDL101/2 009 Nov
A/Calif ornia/VRDL84/20 09 OctA/Lebano n/09L-28 /2009 Nov
A/Italy/127 /2009 JunA/Thailand/CU- B5/200 9 Jun
A/O ntario/25389 /200 9 AugA/Lebano n/09L-06 /2009 Oc t
A/Mexic o/3 955/2009 AprA/Cali fornia/07 /2009 Apr
A/Cali fornia/04 /2009 Apr
95
87
78
86
88
84
95
74
0.0005
D22 5G
F d neuM ees arC
4
tpMa
FAi
ig. 2. Phylogenetic relationships of reference strains to the hemagglutinin (HA) anay 2009 through January 2010. Viruses collected in this study are in bold. The tr
haracteristic amino acid mutations for main branches are indicated.
.3. Phenotypic and genotypic antiviral drug-susceptibility
All of the isolates in this study were resistant to aman-
adine in the TCID50 assay, and this was confirmed by theresence of the S31N amantadine-resistance mutation in their2 genes detected by using the cycling-probe real-time PCRssay. One of the 30 isolates (3.4%) was oseltamivir-resistant,
ig. 3. Structural overview of the receptor-binding domain of the HA protein of H1N1 pdmntigenic sites are highlighted as follows: Sa (red), Sb (yellow), Ca1 and Ca2 (green), and
ndicated in spheres in the Ca1 and Ca2 sites. The structure was rendered in Pymol (www.P
raminidase (NA) genes of influenza H1N1 pdm isolates collected in Lebanon frome rooted at A/California/04/09, and bootstrap values greater than 70% are shown.
having an IC50 value of 240.1 nM; IC50 values of susceptibleviruses were 1.6–3.0 nM (data not shown). The oseltamivir-resistant isolate had an H275Y mutation in its NA gene in
addition to the N68T and E462K mutations detected in theoseltamivir-susceptible isolates. This unique isolate was collectedin October from a 2-year-old patient, who was not receivingantiviral-drug treatment when the sample was collected. All iso-. Side view (A) and top view (B) of the surface-filling representation of the domain. Cb (blue). The 2 major amino acid mutations detected in the H1N1 pdm’s HA areymol.org) using the H1N1 pdm published structure (pdb: 3AL4; Zhang et al., 2010).

linical
ls
5
JiiasVgmgtsmwbabvd
m2gEaAfarl2awipacIy6mvoTeetu
aprpmtdwrcr
H. Zaraket et al. / Journal of C
ates were sensitive to zanamivir (IC50, 0.4–2.5 nM; data nothown).
. Discussion
We detected community-transmitted cases of H1N1 pdm inuly 2009, nearly 6 weeks after the first pandemic cases arrivedn Lebanon.6 The Ca1 antigenic site in the majority of the Lebanesesolates had an S206T mutation, which has been reported as a char-cteristic mutation of a widespread H1N1 pdm strain.13–15 Wehowed that this strain also had 2 amino acid mutations in NA:42I and N248D. The phylogenetic history of these viruses sug-ested that these NA mutations evolved prior to the HA S206Tutation. This strain was successful in continuous circulation, sug-
esting that it has a better transmission and infection efficiencyhan older viruses do, but the mechanisms responsible for this aretill unknown.15 Most of the Lebanese isolates also had a D225Eutation in their Ca2 antigenic sites. Viruses with this mutationere highly homologous, were all isolated in October and Novem-
er 2009, and had identical NA genes that shared 2 mutations (N68Tnd E462K), suggesting that this strain was the cause of a local out-reak in Lebanon during these months. It is noteworthy that olderiruses without the N68T and E462K mutations in NA were alsoetected in Lebanon during October but at a lower frequency.
A BLAST search of the HA sequences revealed that the D225Eutation was detected in a virus isolated in Hong Kong in June
009; however, that isolate did not possess N68T and E462K NAene mutations. The HA D225E mutation was also detected in angyptian isolate during October 2009, but no NA sequence wasvailable from this isolate to compare with those from our study.16
D225G mutation in the HA protein was associated with severe andatal H1N1 pdm cases.17,18 The increased disease severity could bettributed to enhanced binding of 225G HA to �2-3-linked sialyleceptors of a type expressed in the bronchial and lung epithe-ial cells, allowing deeper virus infection.18 However, 225D and25E variants of the HA protein have low-to-moderate bindingffinities to �2-3-linked sialyl receptors.18 None of the patientsith D225E-mutated viruses in this study had severe symptoms,
n agreement with previous reports.17,18 Additionally, none of ouratients had the D225G variant. The S206T and D225E mutationsre in receptor-binding and antigenic sites of the HA protein andould thus improve the adaptation of H1N1 pdm to human hosts.n a report of analyzed serum samples from pre-H1N1 pandemicears, 6–9% of those aged 18–64 years and 33% of those aged over0 years had cross-reactive antibodies.19 Therefore, selection ofutations in the antigenic sites of the HA protein of the pandemic
irus might also suggest immune selection during the early phasef the pandemic because of pre-existing cross-reactive antibodies.he N68T and E462K mutations in the NA are located far from thenzyme pocket’s active site or framework, and therefore are notxpected to affect the enzyme’s activity. However, whether or nothese mutations contribute to virus adaptation or pathogenicity isnknown.
All of the H1N1 pdm viruses isolated in this study weremantadine-resistant and zanamivir-sensitive, consistent withreviously published data.3 However, the only oseltamivir-esistant isolate was obtained from a child who was notreviously treated with oseltamivir, indicating community trans-ission of oseltamivir-resistant H1N1 pdm, which could be due
o the wide use of oseltamivir in Lebanon during the pan-emic. Most of the reported oseltamivir-resistant H1N1 pdm cases
ere isolated from immunocompromised or hospitalized patientseceiving prophylactic or treatment oseltamivir,20–22 and very fewases of community transmission of resistant viruses have beeneported.23,24 The results of in vivo studies in mice and ferret
Virology 51 (2011) 170– 174 173
models showed that oseltamivir-resistant viruses are pathogenicand can be efficiently transmitted through direct contact but notvia respiratory droplets.25–27 Inefficient droplet transmission ofoseltamivir-resistant viruses could explain the very low preva-lence of these viruses. Therefore, continuous monitoring of antiviraldrug-susceptibility of influenza is important to guide the effectiveuse of currently available anti-influenza drugs.
Conflict of interest
GD has received support to conduct clinical trials of oseltamivir.
Funding
U.S.-Japan Acute Respiratory Infection Panel (The Ministry ofHealth, Labor, and Welfare, Japan).
Ethical approval
The Institutional Review Board, American University of Beirut.
Acknowledgements
We are very thankful for the excellent technical assistance ofAkemi Watanabe and the extensive secretarial work of YoshikoKato. We also thank Richard Webby for reviewing the manuscriptand Cherise Guess for editing the manuscript.
References
1. Nicholson KG, Wood JM, Zambon M. Influenza. Lancet 2003;362:1733–45.2. Schnitzler SU, Schnitzler P. An update on swine-origin influenza virus A/H1N1:
a review. Virus Genes 2009;39:279–92.3. Dawood FS, Jain S, Finelli L, Shaw MW, Lindstrom S, Garten RJ, et al. Emer-
gence of a novel swine-origin influenza A (H1N1) virus in humans. N Engl J Med2009;360:2605–15.
4. Neumann G, Noda T, Kawaoka Y. Emergence and pandemic potential of swine-origin H1N1 influenza virus. Nature 2009;459:931–9.
5. Smith G, Vijaykrishna D, Bahl J, Lycett S, Worobey M, Pybus O, et al. Origins andevolutionary genomics of the 2009 swine-origin H1N1 influenza A epidemic.Nature 2009;25:1122–5.
6. Ministry of Public Health Lebanon. Press Release May 2009.http://www.moph.gov.lb/Media/Pages/SwineFluOutbreak.aspx.
7. Hoffmann E, Stech J, Guan Y, Webster RG, Perez DR. Universal primerset for the full-length amplification of all influenza A viruses. Arch Virol2001;146:2275–89.
8. Dapat C, Saito R, Kyaw Y, Naito M, Hasegawa G, Suzuki Y, et al. Epidemiology ofhuman influenza A and B Viruses in Myanmar from 2005 to 2007. Intervirology2009;52:310–20.
9. Suzuki Y, Saito R, Sato I, Zaraket H, Nishikawa M, Tamura T, et al. Identifica-tion of oseltamivir resistance among pandemic and seasonal influenza A(H1N1)viruses by His275Tyr genotyping assay using cycling probe method. J Clin Micro-biol 2011;49:125–30.
10. Tamura K, Dudley J, Nei M, Kumar S. MEGA4: Molecular Evolutionary GeneticsAnalysis (MEGA) software version 4.0. Mol Biol Evol 2007;24:1596–9.
11. Hurt AC, Barr IG, Hartel G, Hampson AW. Susceptibility of human influenzaviruses from Australasia and South East Asia to the neuraminidase inhibitorszanamivir and oseltamivir. Antiviral Res 2004;62:37–45.
12. Suzuki Y, Saito R, Zaraket H, Dapat C, Caperig-Dapat I, Suzuki H. Rapid andspecific detection of amantadine-resistant influenza A viruses with a Ser31Asnmutation by the cycling probe method. J Clin Microbiol 2010;48:57–63.
13. Fereidouni SR, Beer M, Vahlenkamp T, Starick E. Differentiation of twodistinct clusters among currently circulating influenza A(H1N1)v viruses,March–September 2009. Euro Surveill 2009:14.
14. Goni N, Moratorio G, Ramas V, Coppola L, Chiparelli H, Cristina J. Phyloge-netic analysis of pandemic 2009 influenza A virus circulating in the SouthAmerican region: genetic relationships and vaccine strain match. Arch Virol2010;156:87–94.
15. Shiino T, Okabe N, Yasui Y, Sunagawa T, Ujike M, Obuchi M, et al. Molec-ular evolutionary analysis of the influenza A(H1N1)pdm, May–September,2009: temporal and spatial spreading profile of the viruses in Japan. PLoS ONE
2010;5:e11057.16. Adwan G. Molecular characterization and phylogenetic analysis of Middle East2009 H1N1 pdm isolates. Asian Pacific J Trop Med 2010;3:624–8.
17. Chen H, Wen X, To KK, Wang P, Tse H, Chan JF, et al. Quasispecies of theD225G substitution in the hemagglutinin of pandemic influenza A(H1N1) 2009

1 linical
74 H. Zaraket et al. / Journal of Cvirus from patients with severe disease in Hong Kong, China. J Infect Dis2010;201:1517–21.
18. Liu Y, Childs RA, Matrosovich T, Wharton S, Palma AS, Chai W, et al. Alteredreceptor specificity and cell tropism of D222G hemagglutinin mutants iso-lated from fatal cases of pandemic A(H1N1) 2009 influenza virus. J Virol2010;84:12069–74.
19. Centers for Disease Control and Prevention. Serum cross-reactive anti-body response to a novel influenza A (H1N1) virus after vaccinationwith seasonal influenza vaccine. MMWR Morb Mortal Wkly Rep 2009;58:521–4.
20. Centers for Disease Control and Prevention. Oseltamivir-resistant novelinfluenza A (H1N1) virus infection in two immunosuppressed patients– Seattle, Washington, 2009. MMWR Morb Mortal Wkly Rep 2009;58:
893–6.21. Centers for Disease Control and Prevention. Oseltamivir-resistant 2009pandemic influenza A (H1N1) virus infection in two summer campers receiv-ing prophylaxis – North Carolina, 2009. MMWR Morb Mortal Wkly Rep2009;58:969–72.
Virology 51 (2011) 170– 174
22. World Health Organization. Oseltamivir resistance in immunocompro-mised hospital patients (2009 December). http://www.who.int/csr/disease/swineflu/notes/briefing 20091202/en/index.html.
23. Gulland A. First cases of spread of oseltamivir resistant swine flu betweenpatients are reported in Wales. Br Med J 2009;339:b4975.
24. Le QM, Wertheim HF, Tran ND, van Doorn HR, Nguyen TH, Horby P. A commu-nity cluster of oseltamivir-resistant cases of 2009 H1N1 influenza. N Engl J Med2009;362:86–7.
25. Duan S, Boltz DA, Seiler P, Li J, Bragstad K, Nielsen LP, et al. Oseltamivir-resistantpandemic H1N1/2009 influenza virus possesses lower transmissibility and fit-ness in ferrets. PLoS Pathog 2010;6:e1001022.
26. Kiso M, Shinya K, Shimojima M, Takano R, Takahashi K, Katsura H, et al. Char-acterization of oseltamivir-resistant 2009 H1N1 pandemic influenza A viruses.
PLoS Pathog 2010;6:e1001079.27. Seibert CW, Kaminski M, Philipp J, Rubbenstroth D, Albrecht RA, Schwalm F,et al. Oseltamivir-resistant variants of the 2009 pandemic H1N1 influenza Avirus are not attenuated in the guinea pig and ferret transmission models. JVirol 2010;84:11219–26.





























