Embed Size (px)
Citation preview

JOURNAL OF CLINICAL MICROBIOLOGY, Dec. 1993, p. 3109-31140095-1137/93/123109-06$02.00/0Copyright C 1993, American Society for Microbiology
Vol. 31, No. 12
Genetic Diversity among Borrelia burgdorfeni Isolates fromWood Rats and Kangaroo Rats in California
BARBARA C. ZINGG,1 RICHARD N. BROWN,2 ROBERT S. LANE,2 AND RANCE B. LEFEBVRE'*Department ofEntomological Sciences, University of California, Berkeley, California 94720,2 and Department
of Veterinary Microbiology and Immunology, School of Veterinary Medicine,University of California, Davis, California 956161
Received 12 February 1993/Returned for modification 1 April 1993/Accepted 8 June 1993
Twenty-nine Borrelia burgdorferi isolates, obtained from dusky-footed wood rats (Neotoma fuscipes) andCalifornia kangaroo rats (Dipodomys californicus) in California, were analyzed genetically. Chromosomal DNAwas examined by restriction endonuclease analysis (REA) and gene probe restriction fragment lengthpolymorphism. Pulsed-field gel electrophoresis was used to analyze the plasmid profiles of the isolates. REA,the method with the greatest discrimination, disclosed 24 distinct restriction patterns among the 29 isolates.These restriction patterns were sorted into four restriction fragment length polymorphism groups on the basisof their gene hybridization patterns. Results of the REA and plasmid profile analysis supported this grouping.The degree of genetic diversity among Californian isolates demonstrated by our findings is greater than thatpreviously reported among other groups of North American isolates and is similar or greater than the diversityreported among European isolates.
Borrelia burgdorferi was first described as a new speciesin 1984 (23) following its isolation from Ixodes dammini tickscollected on Shelter Island, N.Y. (15). Isolation of a novel,seemingly identical spirochete from the skin (42), blood (10,42), and cerebrospinal fluid (42) of human patients subse-quently confirmed suspicions that this organism was theetiologic agent of Lyme disease. In the past 10 years, Lymedisease has become the most commonly reported vector-borne disease in the United States and Europe, and B.burgdorferi has been identified throughout the temperateclimate zones of the world (38). In California, Lyme borre-liosis was declared a reportable disease in 1989, with a totalof 345 cases being registered the following year (3). In 1990,the majority (92%) of the cases contracted within the statewere diagnosed in northern California residents. The fourneighboring counties of Humboldt, Marin, Mendocino, andSonoma contributed 42% of these patients (3). The remain-ing 8% of reported cases in California may have beencontracted in southern California (3), where B. burgdorferiwas only recently isolated for the first time (13).The primary vertebrate reservoir for the Lyme disease
spirochete in the northeastern United States and the GreatLakes region is the white-footed mouse (Peromyscus leuco-pus) (18, 29, 31). Although Peromyscus spp. mice are alsocommon on the West Coast of the United States, they serveonly as a peripheral reservoir, and the enzootic cycle of B.burgdorferi maintenance in California differs from that in theNortheast and the Midwest (14). In California the primaryreservoir host for B. burgdorferi appears to be the dusky-footed wood rat (Neotoma fuscipes). The more geographi-cally and ecologically restricted California kangaroo rat(Dipodomys californicus) may be a locally significant reser-voir (14, 24).
Classification and identification of spirochetes as B. burg-dorferi are generally based on their reactivities with species-specific monoclonal antibodies (2, 8, 11, 22, 24, 25, 45).
* Corresponding author.
Other methods have also been used to characterize theseorganisms such as sodium dodecyl sulfate (SDS)-polyacryl-amide gel electrophoresis, immunoblotting, multilocus en-zyme electrophoresis, DNA homology, restriction enzymeanalysis (REA), rRNA gene restriction patterns (restrictionfragment length polymorphisms [RFLPs]), and plasmid pro-file (PP) analysis (1, 4, 6, 11, 12, 14, 25-27, 34). Theoutcomes of some of these assays may be affected byprolonged in vitro cultivation, resulting in changes in proteinexpression and reactivity to monoclonal antibodies, as wellas in the loss or rearrangement of plasmids (36, 37). Molec-ular characterization of the chromosomal genome is a pref-erable technique of assessing diversity among bacteria at thespecies level, since the bacterial chromosome is not typicallyaffected by the length of time that the organism has beenkept in culture (20, 26, 27). Additionally, chromosomal DNAanalysis represents a sensitive and discriminating method oftyping strains within a bacterial species, because often DNApolymorphisms are attributable to neutral mutations thatwould not be detected by protein or antigen analysis (32).For these reasons, we investigated B. burgdorferi isolates
from California at the genetic level using REA. We furtherexamined the DNA by gene RFLP, and we used pulsed-fieldgel electrophoresis (PFGE) for linear plasmid analysis. Pre-vious studies reporting on the genetic characteristics ofEuropean and North American B. burgdorferi isolates indi-cated a greater degree of diversity among European isolates(8, 12, 44, 45). The North American isolates analyzed inthose studies, however, originated primarily from the mid-western and northeastern United States. Reports regardingprotein profiles of B. burgdorferi isolates from Californiaindicated an increased level of diversity among these spiro-chetes from the western United States (14, 24, 25). Geneticanalysis of a small number of Californian isolates alsosuggested a disparate population of B. burgdorferi in thisgeographic locale (26). The objective of the present studywas to assess the genetic diversity of a significant number ofrodent-derived isolates of B. burgdorferi from California.
3109
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
cm o
n 14
Feb
ruar
y 20
22 b
y 61
.98.
150.
148.

3110 ZINGG ET AL.
MATERIALS AND METHODS
B. burgdorferi isolates. Each northern Californian isolatewas cultured from ear-punch biopsy specimens taken fromdifferent rodents (40). The rodents were trapped at theUniversity of California Hopland Field Station in MendocinoCounty (14a, 24). Seven B. burgdorfen isolates originatedfrom Californian kangaroo rats and 22 isolates originatedfrom dusky-footed wood rats. In addition, CA 100, the firstsouthern Californian isolate reported (13), was included inthis analysis. CA 100 had been cultured from centrifugedblood cells of a wood rat trapped in the San BernardinoMountains east of Los Angeles. The B31 type strain of B.burgdorfeni (tick isolate from Shelter Island, N.Y. [15]) wasused as a reference strain. All spirochetes were maintainedin modified Barbour-Stoenner-Kelly (BSK) medium at 33°C(5). The isolates analyzed for plasmid content were at a lowpassage number (.10), with the exception of the referencestrain B31.Chromosomal DNA enrichment and REA. Washed pellets
of the spirochetes (resulting from 30 ml of a stationary-phaseculture) were resuspended in 250 p,l of TES (50 mM Tris [pH8.0], 50 mM EDTA, 15% [wt/vol] sucrose). After the addi-tion of an equal volume of 5 M NaCl as well as 0.01%(vol/vol) sodium deoxycholate, the samples were incubatedon ice for 30 min. The partially lysed cell suspension was
then centrifuged at 16,000 x g for 10 min. The supernatantfraction was discarded, and the pellet was resuspended in250 p,l of TES; this was followed by the addition of 250 ,ul of10% SDS and 3 p.l of RNase Ti. After a 30-min incubation at37°C, 50 ,ul of proteinase K (20 mg/ml) was added and thetubes were incubated at 50°C for 30 min. This solution wassubsequently phenol extracted twice, chloroform extractedonce, and ethanol precipitated. The DNA pellets were
resuspended in 44 p,l of TE buffer (10 mM Tris [pH 7.6], 1mM EDTA). A fluorometer (Hoefer Scientific Instruments,San Francisco, Calif.) was used to determine the DNAconcentration. REA was performed by digesting 2.5 ,ug ofsample DNA for 4 h in an excess of HindIII, EcoRI, HhaI,or SmaI (GIBCO BRL Life Technologies, Inc., Gaithers-burg, Md.) or ClaI (Boehringer Mannheim, Indianapolis,Ind.). These enzymes were chosen on the basis of informa-tion presented in a previous study (26). Following digestionwith one of the restriction endonucleases, the DNA frag-ments were separated in a 0.7% agarose gel (20 by 25 cm) byusing Tris-borate-EDTA buffer (89 mM Tris-borate, 89 mMboric acid, 2 mM EDTA) at 60 V for 14.5 to 16.5 h,depending on the enzyme used. Subsequently, the gels werestained in a solution of 0.5 ,ug of ethidium bromide per ml,illuminated with UV light, and photographed. To enhancethe analysis of the DNA profiles, the negatives were scannedwith a computing densitometer (Molecular Dynamics,Sunnyvale, Calif.). The scans of all isolates were comparedby visual analysis and by superimposition. Isolates were
rerun and compared on a single gel when initial REA profiles
were similar.Southern blotting and DNA hybridization. Electrophoresed
DNA restriction fragments were transferred to a nylon
membrane (Hybond N+; Amersham, Arlington Heights, Ill.)
following the manufacturer's instructions for the Vacugene
2016 system (LKB Pharmacia, Piscataway, N.J.). The trans-
ferred DNA was covalently bound to the membrane by UV
cross-linking with a Stratalinker UV cross-linker (Strata-
gene, La Jolla, Calif.). Blots were wrapped in aluminum foil
and were stored at room temperature.The cloned 83-kDa antigen gene of B. burgdorfien B31 (28,
33) was excised from its vector with FokI (New EnglandBiolabs, Beverly, Mass.), purified (GeneClean; Bio 101, LaJolla, Calif.), and used for hybridizing to the Southern blotsof the EcoRI digests. An enhanced chemiluminescencedirect nucleic acid labeling and detection system was usedaccording to the manufacturer's specifications (Amersham).Hybridizations were performed under stringent conditionsovernight at 42°C in a hybridization incubator (RobbinsScientific Corporation, Sunnyvale, Calif.). The blots wereexposed to Kodak XAR film at room temperature for 1 to 5min.PFGE. Borrelial cells were embedded into plugs of 0.5%
low-melting-temperature agarose (InCert; FMC BioProd-ucts, Rockland, Maine) at a concentration of approximately8 x 108 cells per ml. These agarose inserts were incubated inlysis buffer (50 mM Tris [pH 8], 50 mM EDTA, 1% SDS)containing 1 mg of proteinase K per ml for 48 h in a 50°Cwater bath. The plugs were washed twice for 1 h at roomtemperature in TE buffer containing 40,ug phenylmethylsul-fonyl fluoride per ml. The plugs were rinsed briefly in TE andwere then stored at 4°C in 0.5 M EDTA for use in PFGE (35).Molecular size markers consisted of lambda phage concate-mers (multiples of 48.5 kbp) and HindIII-digested lambdaDNA (Promega, Madison, Wis.). Insert slices of about 50 ,ulin volume (containing the DNAs of approximately 4 x 107microorganisms) were added to each well of a 1% agarosegel. Electrophoresis was performed in 0.5 x TBE buffer at15°C in an LKB 2015 Pulsaphor system (Pharmacia LKBBiotechnology, Uppsala, Sweden). Two electrode systemswere used for the plasmid analysis: point electrodes for thedouble inhomogeneous mode and a contour-clamped homo-geneous electric field electrode. The electrodes in the doubleinhomogeneous mode were positioned identically to those ina previous report (26). Both arrays were run at constantvoltage, according to the manufacturer's recommendation,for 24 h with a switch time of 1 s. All gels were stained withethidium bromide and were visualized with UV light.
RESULTS
REA. Twenty-nine B. burgdorfen isolates from California(Table 1), 22 of these from dusky-footed wood rats and 7from California kangaroo rats, were evaluated by REA. Anexample of an REA gel illustrating the electrophoretic DNAfragment configurations of strain B31 and 17 of the 29isolates digested with HhaI is displayed in Fig. 1. Fourteenmarkedly different electrophoretic patterns are visible in Fig.1. Isolates CA 32, CA 33, and CA 123 appeared to be nearlyidentical, except for the lack of a band at approximately 25kbp in CA 123. Strains CA 49 and CA 104 as well as strainsCA 36 and CA 46 also exhibited homologous banding pat-terns. CA 28 was identical to CA 36 and CA 46 in thelower-molecular-size range (<4 kbp). All other isolatesshown on the gel in Fig. 1 demonstrated individual, specificpatterns of chromosomal DNA restriction fragments. Of theremaining 12 isolates digested with HhaI not shown in Fig. 1,CA 37 was identical to CA 36 and CA 46. Isolates CA 120and CA 121 shared similar banding patterns, differing mainlyin the upper molecular size range. Isolates CA 24, CA 25,CA 29, CA 30, CA 35, CA 84, CA 100, CA 112, and CA 125presented unique banding patterns. Chromosomal digestionwith other restriction enzymes (HindIII, EcoRI, ClaI, andSmaI) supported the similarities and differences exhibitedfor HhaI (Fig. 1).RFLP of the 83-kDa antigen gene. Southern blots of EcoRI
digests were probed with a chromosomally conserved gene.
J. CLIN. MICROBIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
cm o
n 14
Feb
ruar
y 20
22 b
y 61
.98.
150.
148.

VOL.31,1993~~~~GENETICDIVERSITY AMONG B. BURGDORFERI ISOLATES 3111
TABLE 1. Source and genotypic profiles of B. burgdorferi isolates from California
RFLP Islt ot80-kDa antigen gene; RFLP REA (identical or PP (identical orgroup Islt otfragmnent bands (kbp) very similar)a very similar)b
I B31 Ixodes dammini 14; 28I CA 21 Dipodomys califomnicus 14;28(0I CA 30 Neotoma fuscipes 14; 28I CA 31 Dipodomys californicus 14; 28I CA 57 Neotoma fuscipes 14; 28I CA 59 Neotoma fuscipes 14; 28I CA 79 Neotoma fuscipes 14; 28 (0)I CA lOOc Neotoma fuscipes 14; 28I CA 102 Neotoma fuscipes 14; 28I CA 120 Neotoma fiuscipes 14; 28 (0) 0I CA 121 Neotoma fuiscipes 14; 28 (0) 0
II CA 20 Dipodomys califomicus 4.2; 14II CA 22 Dipodomys califomnicus 4.2;14(AII CA 32 Dipodomys califomicus 4.2;14AII CA 33 Dipodomys californicus 4.2;14AII CA 123 Dipodomys califomnicus 4.2;14()(A
III CA 24 Neotoma fuscipes 6; 28 C>III CA 25 Neotoma fiuscipes 6; 28 (C>)III CA 29 Neotoma fuscipes 6;28()III CA 35 Neotoma fuiscipes 6;28()III CA 49 Neotoma fiuscipes 6; 28 K>III CA 84 Neotoma fuscipes 6; 28III CA 101 Neotomna fuscipes 6; 28III CA 104 Neotoma fuscipes 6; 28 C>III CA 112 Neotoma fuiscipes 6;28()III CA 125 Neotoma fuiscipes 6; 28
IV CA 28 Neotoma fuscipes 8; 28 (U)IV CA 36 Neotoma futscipes 8;28U0IV CA 37 Neotoma fuscipes 8;28UIV CA 46 Neotoma futscipes 8;28U0a Identical symbols, identical-REA profiles; identical symbols, but one or both in parentheses, very similar REA profiles.b"Identical symbols, identical PPs; identical symbols, but one or both in parentheses, very similar plasmid profiles.Only southern California isolate.
The gene cloned from the B31 type strain expresses an83-kDa protein, as determined by immunoblot analysis (28).DNA hybridization experiments revealed that this protein ishighly conserved and specific for B. butgdorferi (33). Thereis a single EcoRI restriction site within the 83-kDa antigengene in the B31 type strain, thus producing two hybridizationfragmnents sized at 14 and 28 kbp, respectively. An autorad-iograph of an EcoRI digest of 12 representative isolateshybridized with the labeled gene probe is shown in Fig. 2.The hybridization produced two bands of various sizes in theisolates, hence demonstrating RFLPs. On the basis of thisRFLP analysis, the Californian isolates were arranged intofour different groups. Table 1 lists these groups and enumer-ates the approximate sizes of the EcoRI DNA fragmentswhich hybridized to the 83-kDa gene probe.
Interestingly, five of the seven isolates from Californiakangaroo rats fell into the same specific RFLP group char-acterized by hybridizing fragments of approximately 4.2 and14 kbp (group II). The remaining two isolates displayed amatching RFLP with 10 of the wood rat isolates and with theB31 reference strain (group I).PP analysis. Extrachromosomal DNA in the B. burgdorferi
isolates was examined by PFGE. By using the electrophore-sis conditions described above, the separation of the whole-cell DNA revealed that the sizes of the plasmids from thesedifferent isolates ranged from approximately 16 to 65 kbp onthe basis of linear DNA sizing. The number of plasmids
C\ CY) v-'It
-0 -t O OCC '0cN QQ DWQD D00 J
.sN O LOooooooooooooooc--- MId'N 4-
Kbp
23.1 -
9.4 -
6.6 -
4.4 -
2.3 -2.0 -
FIG. 1. REA of chromosomal B. burgdorferi DNA digested withHhaI. The DNA was size fractionated on a 0.7% agarose gel, stainedwith ethidium bromide, and UV illuminated.
VOL. 31, 1993
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
cm o
n 14
Feb
ruar
y 20
22 b
y 61
.98.
150.
148.
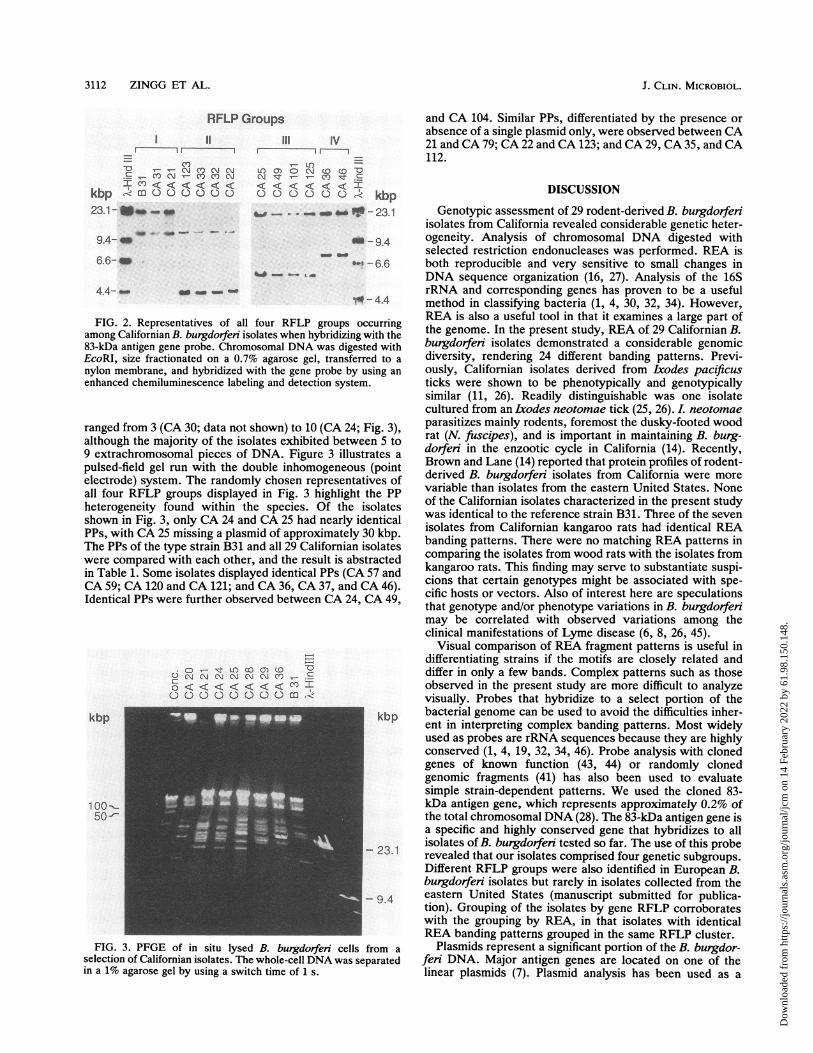
3112 ZINGG ET AL.
RFLP GroupsI~ ~~~~~~llII
-o -- Ir cv cm.9 -- co N- CO COI Cn < < <I < < <K
kbp -R m0 0 c.) D cJ C) c23.1 - Ifso _
III IV
O m coCM 0 -N Ol~ 0- N- CY) (C) g
< c< <t < 'I < ko) C) 0 0 cD C) kbp
and CA 104. Similar PPs, differentiated by the presence orabsence of a single plasmid only, were observed between CA21 and CA 79; CA 22 and CA 123; and CA 29, CA 35, and CA112.
DISCUSSION
9.4- _
6.6-4
4.4- _ _
FIG. 2. Representatives of all four RFLPamong Californian B. burgdorferi isolates when h)83-kDa antigen gene probe. Chromosomal DNA i
EcoRI, size fractionated on a 0.7% agarose gel.nylon membrane, and hybridized with the gene renhanced chemiluminescence labeling and detecti
ranged from 3 (CA 30; data not shown) to 10although the majority of the isolates exhibit(9 extrachromosomal pieces of DNA. Figuripulsed-field gel run with the double inhom(electrode) system. The randomly chosen relall four RFLP groups displayed in Fig. 3heterogeneity found within the species.shown in Fig. 3, only CA 24 and CA 25 hadPPs, with CA 25 missing a plasmid of approxThe PPs of the type strain B31 and all 29 Calwere compared with each other, and the resin Table 1. Some isolates displayed identicalCA 59; CA 120 and CA 121; and CA 36, CAIdentical PPs were further observed betweer
o < <
C) C) C)
t LOC)D O (D r
< < < < C< In0c) 0 mCo
kbp
100-0_5050-
FIG. 3. PFGE of in situ lysed B. burgdorfselection of Californian isolates. The whole-cell Drin a 1% agarose gel by using a switch time of 1 s.
- _ ! - 23.1 Genotypic assessment of 29 rodent-derived B. burgdorfeniisolates from California revealed considerable genetic heter-
-9-4 ogeneity. Analysis of chromosomal DNA digested with_ _ - 6.6 selected restriction endonucleases was performed. REA is
b-6.6 both reproducible and very sensitive to small changes inDNA sequence organization (16, 27). Analysis of the 16SrRNA and corresponding genes has proven to be a usefulmethod in classifying bacteria (1, 4, 30, 32, 34). However,
groups occurring REA is also a useful tool in that it examines a large part of,^bridizing With the the genome. In the present study, REA of 29 Californian B.was digested with burgdorfen isolates demonstrated a considerable genomictransferred to a diversity, rendering 24 different banding patterns. Previ-
probe by using an ously, Californian isolates derived from Lxodes pacificusion system. ticks were shown to be phenotypically and genotypically
similar (11, 26). Readily distinguishable was one isolatecultured from an Lxodes neotomae tick (25, 26). L neotomae
(CA 24; Fig. 3) parasitizes mainly rodents, foremost the dusky-footed wooded betw'eeng5 to rat (N. fuscipes), and is important in maintaining B. burg-e 3 illustrates a dorfen in the enzootic cycle in California (14). Recently,ogeneous (point Brown and Lane (14) reported that protein profiles of rodent-presentatives of derived B. burgdorfen isolates from California were more
presegtatives of variable than isolates from the eastern United States. Noneiighlight thePa of the Californian isolates characterized in the present studyOf the isolateswas identical to the reference strain B31. Three of the sevennearly identical isolates from Californian kangaroo rats had identical REA
dimately 30 kbp. banding patterns. There were no matching REA patterns inult is abstracted comparing the isolates from wood rats with the isolates fromPPs (CA 57 and kangaroo rats. This finding may serve to substantiate suspi-37, and CA 46) cions that certain genotypes might be associated with spe-CA 24, CA 49 cific hosts or vectors. Also of interest here are speculations' that genotype and/or phenotype variations in B. burgdorferi
may be correlated with observed variations among theclinical manifestations of Lyme disease (6, 8, 26, 45).
Visual comparison of REA fragment patterns is useful indifferentiating strains if the motifs are closely related anddiffer in only a few bands. Complex patterns such as thoseobserved in the present study are more difficult to analyzevisually. Probes that hybridize to a select portion of the
kbp bacterial genome can be used to avoid the difficulties inher-ent in interpreting complex banding patterns. Most widelyused as probes are rRNA sequences because they are highlyconserved (1, 4, 19, 32, 34, 46). Probe analysis with clonedgenes of known function (43, 44) or randomly clonedgenomic fragments (41) has also been used to evaluatesimple strain-dependent patterns. We used the cloned 83-kDa antigen gene, which represents approximately 0.2% ofthe total chromosomal DNA (28). The 83-kDa antigen gene isa specific and highly conserved gene that hybridizes to all
>- 23.1isolates of B. burgdorferi tested so far. The use of this probe
- 23.1 revealed that our isolates comprised four genetic subgroups.Different RFLP groups were also identified in European B.burgdorfeii isolates but rarely in isolates collected from the
- 9.4 eastern United States (manuscript submitted for publica-tion). Grouping of the isolates by gene RFLP corroborateswith the grouping by REA, in that isolates with identicalREA banding patterns grouped in the same RFLP cluster.
fen cells from a Plasmids represent a significant portion of the B. burgdor-NA was separated feri DNA. Major antigen genes are located on one of the
linear plasmids (7). Plasmid analysis has been used as a
J. CLIN. MICROBIOL.
tin* - - - 0-0
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
cm o
n 14
Feb
ruar
y 20
22 b
y 61
.98.
150.
148.

GENETIC DIVERSITY AMONG B. BURGDORFERI ISOLATES 3113
strain-typing tool in various bacteria (20, 39) and may beuseful for classifying B. burgdorferi (6). It is an efficientprocedure since DNA extraction is not necessary and prep-
aration of the bacterial cells for in situ lysis requires only a
small amount of cultured organisms. Plasmid typing andREA profiling are comparable in their efficiencies of discrim-inating B. burgdorferi isolates. Among the 29 Californianstrains tested, 23 different plasmid types were found. Al-though plasmid loss may occur in B. burgdorferi, this seemsto be a relatively rare event (37). The difficulty of comparingPPs between laboratories (22) could be overcome if electro-phoresis conditions were standardized. There was a strong
correlation between REA and plasmid typing. The corre-
spondence of the PPs to the REA and RFLP types, whichhad not been observed previously, was unexpected andmight be used as an additional argument in favor of mini-chromosomes (9, 17, 21).Three different methods were used in the present study to
analyze the genomic diversity of 29 Californian isolates of B.burgdorferi, all but 1 of which originated from a singlelocality in Mendocino County. The degree of genotypicvariation found among these isolates corresponds to thephenotypic diversity described within this group of isolates(14). It stands in striking contrast to previous findings fromthe eastern United States, where isolates of B. burgdorferiare much more homogeneous (6, 12, 45). The level ofdiversity observed in the Californian isolates is at leastsimilar in magnitude to that observed in European isolates.Our findings confirm that DNA REA is a useful tool forstudying the diversity among B. burgdorferi strains (26, 27).
ACKNOWLEDGMENT
We thank Tom Schwan, NIH-Rocky Mountain Laboratories, forhis collaboration and openness with us in performing the experi-ments and in preparing the manuscripts pertaining to Borreliaburgdorferi isolates of California published together in this journal.
REFERENCES1. Adam, T., G. S. Gassmann, C. Rasiah, and U. B. Gobel. 1991.
Phenotypic and genotypic analysis of Borrelia burgdorfen iso-lates from various sources. Infect. Immun. 59:2579-2585.
2. Anderson, J. F., L. A. Magnarelli, R. B. LeFebvre, T. G.Andreadis, J. B. McAninch, G. C. Perng, and R. C. Johnson.1989. Antigenically variable Borrelia burgdorferi isolated fromcottontail rabbits and Ixodes dentatus in rural and urban areas.
J. Clin. Microbiol. 27:13-20.3. Anonymous. 1991. The surveillance of Lyme disease in Califor-
nia. Calif. Morbid. 47/48.4. Baranton, G., D. Postic, I. Saint Girons, P. Boerlin, J. C.
Piffaretti, M. Assous, and P. A. Grimont. 1992. Delineation ofBorrelia burgdorferi sensu stricto, Borrelia garinii sp. nov., andgroup VS461 associated with Lyme borreliosis. Int. J. Syst.Bacteriol. 42:378-383.
5. Barbour, A. G. 1984. Isolation and cultivation of Lyme diseasespirochetes. Yale J. Biol. Med. 57:521-525.
6. Barbour, A. G. 1988. Plasmid analysis of Borrelia burgdorferi,the Lyme disease agent. J. Clin. Microbiol. 26:475-478.
7. Barbour, A. G., and C. F. Garon. 1988. The genes encodingmajor surface proteins of Borrelia burgdorferi are located on a
plasmid. Ann. N.Y. Acad. Sci. 539:144-153.8. Barbour, A. G., R. H. Heiland, and T. R. Howe. 1985. Hetero-
geneity of major proteins in Lyme disease borreliae: a molecularanalysis of North American and European isolates. J. Infect.Dis. 152:478-484.
9. Baril, C., C. Richaud, G. Baranton, and I. S. Saint Girons. 1989.Linear chromosome of Borrelia burgdorferi. Res. Microbiol.140:507-516.
10. Benach, J. L., E. M. Bosler, J. P. Hanrahan, J. L. Coleman,G. S. Habicht, L. F. Bast, D. J. Cameron, J. L. Ziegle, A. G.
Barbour, W. Burgdorfer, R. Edelman, and R. A. Kaslow. 1983.Spirochetes isolated from the blood of two patients with Lymedisease. N. Engl. J. Med. 308:740-742.
11. Bisset, M. L., and W. Hill. 1987. Characterization of Borreliaburgdorfen strains isolated from Icodes pacificus ticks in Cali-fornia. J. Clin. Microbiol. 25:2296-2301.
12. Boerlin, P., 0. Peter, A. G. Bretz, D. Postic, G. Baranton, andJ. C. Piffaretti. 1992. Population genetic analysis of Borreliaburgdorfen isolates by multilocus enzyme electrophoresis. In-fect. Immun. 60:1677-1683.
13. Boyce, W. M., R. N. Brown, B. C. Zingg, R. B. LeFebvre, andR. S. Lane. 1992. First isolation of Borrelia burgdorferi insouthern California. J. Med. Entomol. 29:496-500.
14. Brown, R. N., and R. S. Lane. 1992. Lyme disease in California:a novel enzootic transmission cycle of Borrelia burgdorferi.Science 256:1439-1442.
14a.Brown, R. N., and R. S. Lane. Unpublished data.15. Burgdorfer, W., A. G. Barbour, S. F. Hayes, J. L. Benach, E.
Grunwaldt, and J. P. Davis. 1982. Lyme disease-a tick-bornespirochetosis? Science 216:1317-1319.
16. Denny, T. P., M. N. Gilmour, and R. K. Selander. 1988. Geneticdiversity and relationships of two pathovars of Pseudomonassyringae. J. Gen. Microbiol. 134:1949-1960.
17. Ferdows, M. S., and A. G. Barbour. 1989. Megabase-sized linearDNA in the bacterium Borrelia burgdorferi, the Lyme diseaseagent. Proc. Natl. Acad. Sci. USA 6:5969-5973.
18. Godsey, M. S., T. E. Amundson, E. C. Burgess, W. Schell, J. P.Davis, R. Kaslow, and R. Edelman. 1987. Lyme disease ecologyin Wisconsin: distribution and host preferences of Ixodes dam-mini, and prevalence of antibody to Borrelia burgdorferi insmall mammals. Am. J. Trop. Med. Hyg. 37:180-187.
19. Grimont, F., and P. A. D. Grimont. 1986. Ribosomal ribonucleicacid gene restriction patterns as potential taxonomic tools. Ann.Inst. Pasteur Microbiol. 137B:165-175.
20. Hartmann, A., and N. Amarger. 1991. Genotypic diversity of anindigenous Rhizobium meliloti field population assessed byplasmid profiles, DNA fingerprinting, and insertion sequencetyping. Can. J. Microbiol. 37:600-608.
21. Hinnebusch, J., and A. G. Barbour. 1992. Linear- and circular-plasmid copy numbers in Borrelia burgdorferi. J. Bacteriol.174:5251-5257.
22. Hughes, C. A., C. B. Kodner, and R. C. Johnson. 1992. DNAanalysis of Borrelia burgdorferi NCH-1, the first northcentralU.S. human Lyme disease isolate. J. Clin. Microbiol. 30:698-703.
23. Johnson, R. C., G. P. Schmid, F. W. Hyde, A. G. Steigerwalt,and D. J. Brenner. 1984. Borrelia burgdorferi sp. nov.: etiologicagent of Lyme disease. Int. J. Syst. Bacteriol. 34:496-497.
24. Lane, R. S., and R. N. Brown. 1991. Wood rats and kangaroorats: potential reservoirs of the Lyme disease spirochete inCalifornia. J. Med. Entomol. 28:299-302.
25. Lane, R. S., and J. A. Pascocello. 1989. Antigenic characteristicsof Borrelia burgdorfen isolates from ixodid ticks in California.J. Clin. Microbiol. 27:2344-2349.
26. LeFebvre, R. B., R. S. Lane, G.-C. Perng, J. A. Brown, andR. C. Johnson. 1990. DNA and protein analyses of tick-derivedisolates of Borrelia burgdorfen from California. J. Clin. Micro-biol. 28:700-707.
27. LeFebvre, R. B., G.-C. Perng, and R. C. Johnson. 1989. Char-acterization of Borrelia burgdorfen isolates by restriction endo-nuclease analysis and NA hybridization. J. Clin. Microbiol.27:636-639.
28. LeFebvre, R. B., G.-C. Perng, and R. C. Johnson. 1990. The83-kilodalton antigen of Borrelia burgdorferi which stimulatesimmunoglobulin M (IgM) and IgG responses in infected hosts isexpressed by a chromosomal gene. J. Clin. Microbiol. 28:1673-1675.
29. Levine, J. F., M. L. Wilson, and A. Spielman. 1985. Mice asreservoir of the Lyme disease spirochete. Am. J. Trop. Med.Hyg. 34:355-360.
30. Marconi, R. T., and C. F. Garon. 1992. Identification of a thirdgenomic group of Borrelia burgdorferi through signature nucle-otide analysis and 16S rRNA sequence determination. J. Gen.
VOL. 31, 1993
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
cm o
n 14
Feb
ruar
y 20
22 b
y 61
.98.
150.
148.

3114 ZINGG ET AL.
Microbiol. 138:533-536.31. Mather, T. N., M. L. Wilson, S. I. Moore, J. M. C. Ribeiro, and
A. Spielman. 1989. Comparing the relative potential of rodentsas reservoirs of the Lyme disease spirochete (Borrelia burgdor-fen). Am. J. Epidemiol. 130:143-150.
32. Owen, R. J. 1989. Chromosomal DNA fingerprinting-a newmethod of species and strain identification applicable to micro-bial pathogens. J. Med. Microbiol. 30:89-99.
33. Perng, G.-C., R. B. LeFebvre, and R. C. Johnson. 1991. Furthercharacterization of a potent immunogen and the chromosomalgene encoding it in the Lyme disease agent, Borrelia burgdor-fen. Infect. Immun. 59:2070-2074.
34. Postic, D., C. Edlinger, C. Richaud, F. Grimont, Y. Dufresne, P.Perolat, G. Baranton, and P. A. D. Grimont. 1990. Two genomicspecies in Borrelia burgdorferi. Res. Microbiol. 141:465-475.
35. Sambrook, J., E. F. Fritsch, and T. Maniatis. 1989. Molecularcloning: a laboratory manual, 2nd ed. Cold Spring HarborLaboratory Press, Cold Spring Harbor, N.Y.
36. Schwan, T. G., and W. Burgdorfer. 1987. Antigenic changes ofBorrelia burgdorferi as a result of in vitro cultivation. J. Infect.Dis. 156:852-853.
37. Schwan, T. G., W. Burgdorfer, and C. F. Garon. 1988. Changesin infectivity and plasmid profile of the Lyme disease spirocheteBorrelia burgdorferi as a result of in vitro cultivation. Infect.Immun. 56:1831-1836.
38. Sigal, L. H., and A. S. Curran. 1991. Lyme disease: a multifocalworldwide epidemic. Annu. Rev. Public Health 12:85-109.
39. Simor, A. E., B. Shames, B. Drumm, P. Sherman, D. E. Low,
and J. L. Penner. 1990. Typing of Campylobacter pylon bybacterial DNA restriction endonuclease analysis and determi-nation of plasmid profile. J. Clin. Microbiol. 28:83-86.
40. Sinsky, R. J., and J. Piesman. 1989. Ear punch biopsy methodfor detection and isolation of Borrelia burgdorfen from rodents.J. Clin. Microbiol. 29:1723-1727.
41. Stalhammar-Carlemaim, M., E. Jenny, L. Gern, A. Aeschli-mann, and J. Meyer. 1990. Plasmid analysis and restrictionfragment length polymorphisms of chromosomal DNA allow adistinction between Borrelia burgdorferi strains. Zentralbl. Bak-teriol. Parasitenkd. Infektionskr. Hyg. Abt. 1 Orig. 274:28-39.
42. Steere, A. C., R. L. Grodzicki, A. N. Kornblatt, J. E. Craft,A. G. Barbour, W. Burgdorfer, G. P. Schmid, E. Johnson, andS. E. Malawista. 1983. The spirochetal etiology of Lyme dis-ease. N. Engl. J. Med. 308:733-740.
43. Timms, P., F. W. Eaves, A. A. Girjes, and M. F. Lavin. 1988.Comparison of Chlamydia psittaci isolates by restriction endo-nuclease and DNA probe analyses. Infect. Immun. 56:287-290.
44. Wallich, R., C. Helmes, U. E. Schaible, Y. Lobet, S. E. Moter,M. D. Kramer, and M. M. Simon. 1992. Evaluation of geneticdivergence among Borrelia burgdorfeni isolates by use of OspA,fla, HSP60, and HSP70 gene probes. Infect. Immun. 60:4856-4866.
45. Wilske, B., V. Preac-Mursic, G. Schierz, R. Kuhlbeck, A. G.Barbour, and M. Kramer. 1988. Antigenic variability of Borreliaburgdorfeni. Ann. N.Y. Acad. Sci. 539:126-143.
46. Woese, C. R. 1987. Bacterial evolution. Microbiol. Rev. 51:221-271.
J. CLIN. MICROBIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
cm o
n 14
Feb
ruar
y 20
22 b
y 61
.98.
150.
148.





























