Embed Size (px)
Citation preview

EMBRYONIC STEM CELLS/INDUCED PLURIPOTENT STEM CELLS
Generation of Parthenogenetic Induced Pluripotent Stem Cells
from Parthenogenetic Neural Stem Cells
JEONG TAE DO,aJIN YOUNG JOO,
bDONG WOOK HAN,
bMARCOS J. ARAUZO-BRAVO,
bMIN JUNG KIM,
a,c
BORIS GREBER,b HOLM ZAEHRES,b INGEBORG SOBEK-KLOCKE,b HYUNG MIN CHUNG,c HANS R. SCHOLERb
aLaboratory of Stem Cell and Developmental Biology, CHA Stem Cell Institute, CHA University, Seoul, Republic
of Korea; bDepartment of Cell and Developmental Biology, Max Planck Institute for Molecular Biomedicine,
Munster, Germany; cCHA Bio & Diostech Co., Ltd., Seoul, Republic of Korea
Key Words. Reprogramming • Parthenogenetic maternal iPS cells • Genomic imprinting
ABSTRACT
Somatic cells can achieve a pluripotent cell state in a pro-cess called pluripotential reprogramming. Multipotent stemcells can differentiate into cells of only one lineage, but plu-
ripotent stem cells can give rise to cells of all three germlayers of an organism. In this study, we generated inducedpluripotent stem (iPS) cells from bimaternal (uniparental)
parthenogenetic neural stem cells (pNSCs) by transductionwith either four (4F: Oct4, Klf4, Sox2, and c-Myc) or two
(2F: Oct4 and Klf4) transcription factors. The resultantmaternal iPS cells, which were reprogrammed directlyfrom pNSCs, were capable of generating germ line-compe-
tent chimeras. Interestingly, analysis of global gene expres-sion and imprinting status revealed that parthenogeneticiPS cells clustered closer to parthenogenetic ESCs than to
female ESCs, with patterns that were clearly distinct fromthose of pNSCs. STEM CELLS 2009;27:2962–2968
Disclosure of potential conflicts of interest is found at the end of this article.
INTRODUCTION
Cells inherit a stable genetic program partly through variousepigenetic marks, such as DNA methylation and histone mod-ifications. This cellular memory, however, gets erased as cellsundergo ‘‘genetic reprogramming’’, and the cellular programreverts to that of an earlier developmental stage. Somatic cellscan therefore re-establish a pluripotent cell state in a processcalled pluripotential reprogramming. Induced pluripotent stem(iPS) cells have recently been generated by reprogrammingsomatic cells by transduction with a transcription-factor cock-tail [1]. Such iPS cells have been derived from somatic cellsof all three germ layers: mesoderm (fibroblasts, B lympho-cytes, and stomach cells) [2, 3], endoderm (liver and pancre-atic beta cells) [2, 4], and ectoderm (neural cells) [5–7].These iPS cells all originate from biparental tissue. In thisstudy, we have generated iPS cells from bimaternal (uniparen-tal) parthenogenetic neural stem cells (pNSCs) by transduc-tion with either four (4F) or two (2F) transcription factors. Todate, parthenogenetic embryos and ES cells (pESCs) havebeen used to investigate the biological significance ofgenomic imprinting during embryonic development. Partialloss of imprinting has been observed in parthenogenetic blas-
tocysts and their derivative pESCs but not in control blasto-cysts and their derivative ESCs [8]. However, it is not clearwhy the imprinting status of parthenogenetic blastocysts andpESCs differs, as the constituent cells originate from totipo-tent fertilized embryos. Here we first generated parthenoge-netic somatic cell lines and then used them to directly deriveparthenogenetic maternal pluripotent stem (miPS) cells. Wethen compared these miPS cells with ESCs and biparental iPScells and determined the similarity of their parthenogeneticimprinting pattern to that of pESCs or control ESCs. Withthis unique system it is now possible to study the molecularevents that alter the genomic imprint during reprogrammingof parthenogenetic somatic cells into their pluripotentderivatives.
MATERIALS AND METHODS
Generation of Parthenogenetic Embryos
OG2þ/� mice were induced to superovulate by serial injections of5 IU of pregnant mare serum gonadotropin (Sigma-Aldrich, St.Louis, http://www.sigmaaldrich.com) and 5 IU of human
Author contributions: J.T.D.: conception and design, collection and/or assembly of data, data analysis and interpretation, manuscriptwriting, final approval of manuscript; J.Y.J.: conception and design, collection and/or assembly of data; D.W.H.: collection and/orassembly of data, data analysis and interpretation; M.J.A.-B.: data analysis and interpretation; M.J.K.: collection and/or assembly ofdata; B.G.: data analysis and interpretation; H.Z. and I.S.-K.: provision of study material or patients; H.M.C.: provision of study materialor patients, financial support; H.R.S.: conception and design, financial support, manuscript writing, final approval of manuscript.
Correspondence: Jeong Tae Do, Ph.D. CHA University, CHA Stem Cell Institute, Laboratory of Stem Cell and Developmental Biology,605-21 Yoeksam 1-dong, Gangnam-gu, Seoul 135-081, Republic of Korea. Telephone: 82-2-3468-2830; Fax: 82-2-3468-3373; e-mail:[email protected]; or Hans R. Scholer, Max Planck Institute for Molecular Biomedicine, Department of Cell and Developmental Biology,Rontgenstrasse 20, 48149 Munster, Germany. Telephone: 49-251-70365-300; Fax: 49-251-70365-399; e-mail: [email protected] Received August 13, 2009; accepted for publication September 18, 2009; first published online in STEM CELLS EXPRESS October8, 2009. VC AlphaMed Press 1066-5099/2009/$30.00/0 doi: 10.1002/stem.233
STEM CELLS 2009;27:2962–2968 www.StemCells.com

chorionic gonadotropin (hCG) (Sigma-Aldrich), administered 48hours apart. Fourteen hours after the last hCG injection, cumulus-oocyte complexes were recovered from the mouse oviducts, andcumulus cells were removed by treatment with 0.1% hyaluoni-dase (Sigma-Aldrich) in M2 medium. For parthenogenetic activa-tion, denuded oocytes were cultured in M16 medium supple-mented with 10 mmol/l SrCl2 and 5 lg/ml cytochalasin B.Activated oocytes were cultured first in G1.3 medium (Vitrolife,Goteborg, Sweden, http://www.vitrolife.com) for 2 days and laterin G2.3 medium (Vitrolife) under mineral oil at 37�C, 5% CO2.Blastocysts were transferred into one uterine horn of 2.5-dpc(days postcoitum) pseudopregnant mice; 10.5-dpc parthenogeneticembryos were recovered from the pregnant mice. OG2þ/� micewere also used for derivation of parthenogenetic ESCs and femaleESCs. All mouse strains were bred and housed at the mouse fa-cility of the Max Planck Institute for Molecular Biomedicine orwere bought from Harlan Winkelmann (Harlan GmbH, Borchen,Germany, http://www.harlan.com).
Generation of Parthenogenetic NSCs
Brain tissue was collected from 10.5-dpc parthenogeneticembryos (OG2þ/�). Neurospheres were cultured as thoroughlydescribed in our previous article [9]. Primary neurospheres werere-plated onto gelatin-coated dishes in NSC expansion medium.Outgrowing cells were trypsinized, re-plated, and cultured inNSC expansion medium. NSCs were established by dissociationand were re-plated onto gelatin-coated dishes in NSC expansionmedium, comprising NS-A media (Euroclone, Siziano, Italy,http://www.euroclonegroup.it) supplemented with N2 supplement,10 ng/ml of each epidermal growth factor and basic fibroblastgrowth factor (Invitrogen, Carlsbad, CA, http://www.invitrogen.com), 50 lg/ml of bovine serum albumin (Fraction V; Gibco-BRL, Gaithersburg, MD, http://www.gibcobrl.com), 1� penicil-lin/streptomycin/glutamine, and 1� nonessential amino acids(Gibco BRL). Passage 10-12 pNSCs were used as donor cells forreprogramming.
Generation and Culture of Induced PluripotentStem Cells
pMX-based retroviral vectors encoding the mouse cDNAs ofOct4, Sox2, KLf4, and c-Myc [1] were separately co-transfectedby packaging defective helper plasmids into 293T cells usingFugene 6 transfection reagent (Roche Diagnostics, Basel, Switzer-land, http://www.roche-applied-science.com). Forty-eight hoursafter infection, virus supernatants were collected, filtered, andconcentrated as previously described [10]. Female NSCs (fNSCs)or pNSCs (OG2þ/�) were seeded at a density of 1 � 105 cellsper 6-well plate and incubated with virus-containing supernatantsof the four factors (4F; 1:1:1:1) supplemented with 6 lg/ml ofprotamine sulfate (Sigma-Aldrich) for 24 hours. Cells were re-plated onto mitomycin C (MMC)-treated mouse embryonic fibro-blast (MEF) feeders in ESC medium. Oct4-GFPþ iPS cells weresorted by fluorescence-activated cell sorting and subcultured ontoMMC-treated MEF feeders. Passage 12-15 miPS cells were usedfor further analysis.
Aggregation of iPS Cells with Zona-Free Embryos
iPS cells were aggregated and cultured with denuded postcom-pacted 8-cell-stage mouse embryos. Morula embryos were flushedfrom 2.5-dpc B6C3F1 female mice. Clumps of iPS cells (10–20cells per clump) were selected after a brief period of trypsiniza-tion and transferred into microdrops of KSOM medium (10% fe-tal calf serum) under mineral oil containing zona-free morulaembryos. Aggregates were cultured overnight at 37�C, 5% CO2.After 24 hours of culture, the aggregated blastocysts were trans-ferred into one uterine horn of a 2.5-dpc pseudopregnantrecipient.
Whole Genome Expression Analysis
RNA samples for microarrays were prepared using QiagenRNeasy columns with on-column DNA digestion. Three hundrednanograms of total RNA per sample was used as the starting ma-terial for the linear amplification protocol (Ambion, Austin, TX,http://www.ambion.com), which involved synthesis of T7-linkeddouble-stranded cDNA and 12 hours of in vitro transcriptionincorporating biotin-labeled nucleotides. Purified and labeledcRNA was then hybridized for 18 hours onto MouseRef-8 v2gene expression BeadChips (Illumina, Inc., San Diego, http://www.illumina.com) following the manufacturer’s instructions. Af-ter being washed, the chips were stained with streptavidin-Cy3(GE Healthcare, Chalfont St. Giles, UK, http://www.gehealthcar-e.com) and scanned using the iScan reader and accompanyingsoftware. Samples were exclusively hybridized as biologicalreplicates.
Microarray Analysis
The intensities for each bead were mapped onto gene informationusing BeadStudio 3.2 (Illumina). Background correction was per-formed using the Affymetrix Robust Multi-array Analysis back-ground correction model, variance stabilization was performedusing the log2 scaling, and gene expression normalization wascalculated with the quantile method implemented in the lumipackage of R-Bioconductor. Data postprocessing and graphicswere performed with in-house developed functions in Matlab.Microarray data can be assessed at Gene Expression Omnibus(GEO) under the following link: http://www.ncbi.nlm.nih.gov/geo/query/acc.cgi.
RESULTS
Generation of Parthenogenetic MEFs and NeuralStem Cells
To derive parthenogenetic mouse embryonic fibroblasts(pMEFs) and pNSCs, parthenogenetic blastocysts (Fig. 1A)were obtained from parthenogenetically activated OG2þ/�
heterozygous embryos and transferred into pseudopregnantmice. The pregnant mice were then sacrificed at 10.5 dpc, asmouse parthenogenetic embryos die by day 10 of gestation[11, 12]. One fetus, which exhibited growth retardation, wasrecovered from a foster mother (Fig. 1A). The fetal brain wasdissected and cultured for neurosphere culture; the rest of thefetus was cultured for pMEFs. pMEFs exhibited slow prolifer-ation and appeared to undergo senescence after passage 2,indicating a pathenogenetic defect in proliferation (data notshown). However, parthenogenetic neurospheres wereobserved to form normally until day 6 of culture (Fig. 1B),when pNSCs were derived under adherent culture conditions.The pNSCs were morphologically similar to control fNSCsand stained positive for the two major NSC markers Sox2 andNestin (Fig. 1B). Moreover, pNSCs and fNSCs expressedsimilar levels of Sox2, Nestin, and Olig2 (Fig. 1C). Interest-ingly, however, pNSCs exhibited a much slower proliferationrate than fNSCs (Fig. 1D). Since maternally imprinted genes,such as Impact, Ndn, and Snrpn, are associated with the de-velopment of the mouse brain [13–15], it is possible that theslower proliferation rate of pNSCs may be due to the lowerexpression of maternally imprinted genes in pNSCs comparedwith those in control fNSCs (supplemental online Fig. 1). ButpNSCs derived from parthenogenetic embryos can still self-renew for more than 30 passages (3 months).
Do, Joo, Han et al. 2963
www.StemCells.com

Generation of miPS Cells
We first attempted to derive miPS cells from passage 2pMEFs, which had already undergone senescence. However,we failed to generate Oct4-GFPþ or ESC-like colonies frompMEFs by transduction with four transcription factors (Oct4,Sox2, c-Myc, and Klf4) in two independent experiments.
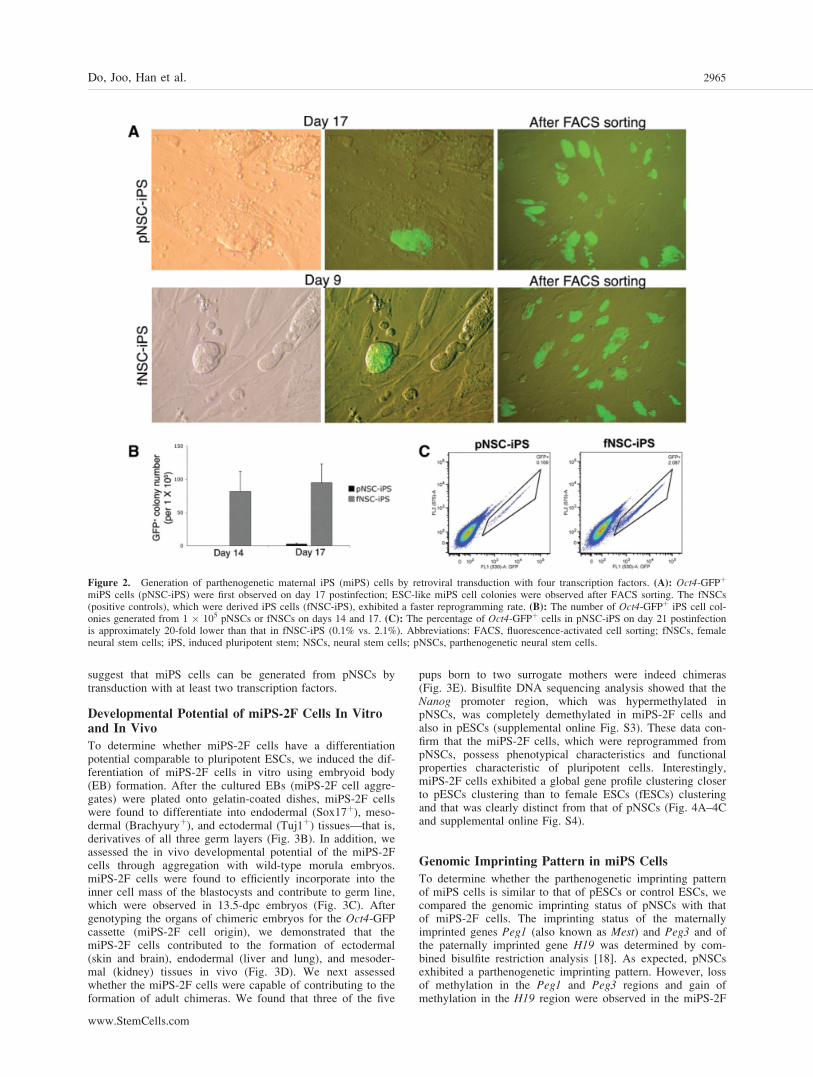
Next, we used pNSCs to generate miPS cells. NSCs havebeen successfully reprogrammed by transduction with fewerthan four transcription factors [6, 7, 16], and thus appear tobe a somatic cell type ideal for reprogramming. We attemptedto establish miPS cells from pNSCs using retroviral vectorstransducing either four or two transcription factors and, inparallel, transduced normal fNSCs as a control. During thereprogramming of control fNSCs, ESC-like colonies were firstobserved on day 4 postinfection (supplemental online Fig. S2)and Oct4-GFPþ colonies on day 9 postinfection (Fig. 2A).However, during the reprogramming of pNSCs, ESC-like col-onies were first observed on day 6 postinfection, 2 days laterthan in control fNSCs (supplemental online Fig. S2), andOct4-GFPþ colonies were observed on day 17 postinfection(Fig. 2A), 8 days later than in fNSCs. The number of Oct4-
GFPþ ESC-like colonies on day 17 was calculated to be 956 21.2 (mean 6 SD) and 3 6 1.0 for a starting cell popula-tion of 1 � 105 fNSCs and pNSCs, respectively (Fig. 2B).The reprogramming efficiency was estimated by flow cyto-metric analysis for Oct4-GFPþ cell count on day 21 postinfec-tion. The percentage of Oct4-GFPþ cells obtained after thereprogramming of pNSCs was approximately 20-fold lowerthan that obtained after the reprogramming of fNSCs (0.1%vs. 2.1%) (Fig. 2C). There is no obvious explanation for thelower reprogramming efficiency of pNSCs compared withthat of fNSCs, but the slow proliferation rate of pNSCs maypartially account for this, as viral infection-induced reprog-ramming may depend on the active proliferation of somaticcells [17]. Next, we generated miPS cells from pNSCs bytransduction with two transcription factors: Oct4 and Klf4.Oct4-GFPþ colonies were first observed on day 30 postinfec-tion in both pNSCs and control fNSCs (Fig. 3A). The effi-ciency of reprogramming of pNSCs and fNSCs using thesetwo factors was similarly reduced, indicating that the use offewer than four transcription factors reduces the reprogram-ming rate of both cell types in a similar fashion. These data
Figure 1. Derivation of pNSCs. (A): 3.5-dpc parthenogenetic blastocysts (Oct4-GFPþ) and 10.5-dpc parthenogenetic embryos developed fromOG2þ/� oocytes. 10.5-dpc parthenogenetic embryos are smaller than their normal embryo counterparts. (B): Parthenogenetic neurospheres formedin brain tissue culture; pNSCs stained positive for Sox2 and Nestin. (C): Real-time reverse transcription-polymerase chain reaction reveals thatpNSCs and control fNSCs have the same levels of the NSC markers Sox2, Nestin, and Olig2. (D): Cell proliferation rate of pNSCs comparedwith that of fNSCs. pNSCs and fNSCs seeded at 2 � 104 cells reach 5.3 6 0.7 (mean 6 SD) and 248.0 6 17.7 � 104 cells, respectively, after 6days of culture. Abbreviations: dpc, days postcoitum; fES cells, female embryonic stem cells; fNSCs, female neural stem cells; NSCs, neuralstem cells; pNSCs, parthenogenetic neural stem cells.
2964 Parthenogenetic iPS Cells
suggest that miPS cells can be generated from pNSCs bytransduction with at least two transcription factors.
Developmental Potential of miPS-2F Cells In Vitroand In Vivo
To determine whether miPS-2F cells have a differentiationpotential comparable to pluripotent ESCs, we induced the dif-ferentiation of miPS-2F cells in vitro using embryoid body(EB) formation. After the cultured EBs (miPS-2F cell aggre-gates) were plated onto gelatin-coated dishes, miPS-2F cellswere found to differentiate into endodermal (Sox17þ), meso-dermal (Brachyuryþ), and ectodermal (Tuj1þ) tissues—that is,derivatives of all three germ layers (Fig. 3B). In addition, weassessed the in vivo developmental potential of the miPS-2Fcells through aggregation with wild-type morula embryos.miPS-2F cells were found to efficiently incorporate into theinner cell mass of the blastocysts and contribute to germ line,which were observed in 13.5-dpc embryos (Fig. 3C). Aftergenotyping the organs of chimeric embryos for the Oct4-GFPcassette (miPS-2F cell origin), we demonstrated that themiPS-2F cells contributed to the formation of ectodermal(skin and brain), endodermal (liver and lung), and mesoder-mal (kidney) tissues in vivo (Fig. 3D). We next assessedwhether the miPS-2F cells were capable of contributing to theformation of adult chimeras. We found that three of the five
pups born to two surrogate mothers were indeed chimeras(Fig. 3E). Bisulfite DNA sequencing analysis showed that theNanog promoter region, which was hypermethylated inpNSCs, was completely demethylated in miPS-2F cells andalso in pESCs (supplemental online Fig. S3). These data con-firm that the miPS-2F cells, which were reprogrammed frompNSCs, possess phenotypical characteristics and functionalproperties characteristic of pluripotent cells. Interestingly,miPS-2F cells exhibited a global gene profile clustering closerto pESCs clustering than to female ESCs (fESCs) clusteringand that was clearly distinct from that of pNSCs (Fig. 4A–4Cand supplemental online Fig. S4).
Genomic Imprinting Pattern in miPS Cells
To determine whether the parthenogenetic imprinting patternof miPS cells is similar to that of pESCs or control ESCs, wecompared the genomic imprinting status of pNSCs with thatof miPS-2F cells. The imprinting status of the maternallyimprinted genes Peg1 (also known as Mest) and Peg3 and ofthe paternally imprinted gene H19 was determined by com-bined bisulfite restriction analysis [18]. As expected, pNSCsexhibited a parthenogenetic imprinting pattern. However, lossof methylation in the Peg1 and Peg3 regions and gain ofmethylation in the H19 region were observed in the miPS-2F
Figure 2. Generation of parthenogenetic maternal iPS (miPS) cells by retroviral transduction with four transcription factors. (A): Oct4-GFPþ
miPS cells (pNSC-iPS) were first observed on day 17 postinfection; ESC-like miPS cell colonies were observed after FACS sorting. The fNSCs(positive controls), which were derived iPS cells (fNSC-iPS), exhibited a faster reprogramming rate. (B): The number of Oct4-GFPþ iPS cell col-onies generated from 1 � 105 pNSCs or fNSCs on days 14 and 17. (C): The percentage of Oct4-GFPþ cells in pNSC-iPS on day 21 postinfectionis approximately 20-fold lower than that in fNSC-iPS (0.1% vs. 2.1%). Abbreviations: FACS, fluorescence-activated cell sorting; fNSCs, femaleneural stem cells; iPS, induced pluripotent stem; NSCs, neural stem cells; pNSCs, parthenogenetic neural stem cells.
Do, Joo, Han et al. 2965
www.StemCells.com

cells, indicating that pNSCs lose the parthenogenetic imprint-ing patterns during pluripotential reprogramming.
Two clonal lines of the four factor-induced miPS cellsand pESCs (passage 7) also exhibited the same imprintingpatterns as miPS-2F cells (Fig. 4D), which is consistent withobservations in pESCs [8]. Our results were confirmed byDNA bisulfite sequencing analysis. pNSCs exhibited hypome-thylation of the paternally imprinted genes H19 and Igf2 andhypermethylation of the maternally imprinted genes Peg1 andPeg3 (Fig. 4E). Interestingly, however, de novo DNA methyl-ation was observed in the H19 and Igf2 regions of pNSC-2Fcells (Fig. 4E), confirming that loss of parthenogeneticimprinting occurs during pluripotential reprogramming.Changes in the parthenogenetic imprinting patterns observed
during the reprogramming process are reminiscent of thoseobserved during the establishment of induced pluripotent stemcells from germ cells [19]. Therefore, loss of parthenogeneticimprinting appears to be a phenomenon specific to partheno-genetic pluripotent stem cells, such as pESCs and miPS cells.Collectively, miPS cells and pESCs display similar imprintingpatterns and global gene expression profiles.
DISCUSSION
Here, we first generated pNSCs and then used them todirectly derive parthenogenetic miPS cells by transduction
Figure 3. Generation of two factor-inducedmaternal iPS (miPS-2F) cells and their differ-entiation potential in vivo and in vitro. (A):
Oct4-GFPþ iPS cells were generated fromfNSCs and pNSCs transduced with Oct4 andKlf4 on day 30 postinfection, forming ESC-like colonies, which were observed after FACSsorting. (B): In vitro differentiation potential ofmiPS-2F cells. miPS-2F cells differentiate intocells of ectoderm (Tuj1þ), endoderm (Sox17þ),and mesoderm (Brachyuryþ). (C): miPS-2Fcells efficiently incorporate into the inner cellmass of a blastocyst after aggregation withwild-type morula embryos and contribute toformation of germ line cells (Oct4-GFPþ) (D):A 13.5-dpc (days postcoitum) chimeric embryocontains cells originating from miPS-2F cells(with Oct4-GFP transgene) in tissues of allthree germ layers: ectoderm (skin and brain),endoderm (liver and lung), and mesoderm (kid-ney). (E): Adult chimeric mice derived aftermorula aggregation of miPS-2F cells identifiedby Oct4-GFP genotyping of their tail tips.Abbreviations: DAPI, 40,6-diamidino-2-phenyl-indole; FACS, fluorescence-activated cell sort-ing; fNSCs, female neural stem cells; iPS,induced pluripotent stem; NSCs, neural stemcells; pNSCs, parthenogenetic neural stemcells.
2966 Parthenogenetic iPS Cells

with reprogramming factors. The direct reprogramming ofparthenogenetic somatic cells renders possible a direct com-parison between the original parthenogenetic somatic cellsand their pluripotent derivative miPS cells, which would notbe possible using pESCs.
Interestingly, pNSCs exhibited a slower proliferation ratethan control biparental fNSCs, whereas miPS cells and pESCsshowed comparable proliferation rates. This may be due tothe re-establishment of maternally imprinted genes, which areassociated with the development of the mouse neural system,during the pluripotential reprogramming of miPS cells. Alsoconceivable is the possibility that maternally imprinted genes
are not necessary for the self-renewal of pluripotent cells orthat maternally imprinted genes are reprogrammed during theinduction of pluripotency. pMEF cells had a very severe pro-liferation defect, however, exhibited by slow proliferation andeventual senescence after passage 2, which may have ulti-mately resulted in their failure to generate iPS cells by trans-duction with four transcription factors. Hanna et al. reportedthat terminally differentiated B cells could not be induced tobecome iPS cells unless the B-cell-specific gene Pax5 hadbeen inactivated by overexpressing CCAAT/enhancer-binding-protein-a (C/EBPa) [3]. Therefore, successful reprogrammingof pMEFs by a four transcription factor-transduction process
Figure 4. Global gene expression profile and loss of imprinting in miPS cells. (A): Heat map of the global gene expression of the tested sam-ples. (B): Pairwise scatter plots comparing the global gene expression patterns of miPS-2F cells with pNSCs and of miPS-2F cells with pESCs.Black lines indicate twofold differences in gene expression levels between the paired cell types. Genes upregulated in ordinate samples comparedwith abscissa samples are shown in blue; those downregulated are shown in red. The positions of the pluripotent markers (Oct4, Sox2, Nanog,Klf4, and Lin28) are shown as green dots. The gene expression levels are in log2 scale. (C): Hierarchical clustering of the global gene expressionprofile. The red branches cluster pluripotent parthenogenetic populations and the blue branches NSC populations. (D): Combined bisulfiterestriction analysis in fNSCs, pNSCs, iPS cells derived from fNSCs, miPS cells, miPS-2F cells, and pESCs. (E): Methylation pattern in the H19,Igf2, and Peg1 regions of pNSCs, miPS-2F cells, and pESCs. Each line represents a separate clone and the sequence position of each CpG site isshown. Black and white circles represent methylated and unmethylated CpGs, respectively. Abbreviations: DAPI, 40,6-diamidino-2-phenylindole;FACS, fluorescence-activated cell sorting; fiPS, female induced pluripotent stem; fNSCs, female neural stem cells; iPS, induced pluripotentstem; miPS, maternal induced pluripotent stem; NSCs, neural stem cells; pESCs, parthenogenetic embryonic stem cells; pNSCs, parthenogeneticneural stem cells.
Do, Joo, Han et al. 2967
www.StemCells.com

may actually require the presence of yet additional factors.This senescence barrier also might be overcome by hypoxiacondition [20] or manipulation of the p53 pathway [21–25]since these systems remarkably increase the reprogramming ef-ficiency. Park et al. recently reported the derivation of disease-specific iPS cells from individual patients with different geneticdiseases [26]. These immortalized cells directly induced frompatients’ somatic cells represent a potential alternative to thesparse cell lines used for research into the etiology and treat-ment of various diseases. Consistent with this approach, weprovide two alternative resources for investigations into theparthenogenetic and genomic imprinting status of cells: parthe-nogenetic somatic cells and pluripotent stem cells.
In this study, we found that miPS cells are capable ofgenerating germ line chimeras and exhibit a global geneexpression profile clustering closer to that of pESCs than tothat of control fESCs. Interestingly, miPS cells, which weredirectly reprogrammed from pNSCs, exhibited changes ingenomic imprinting patterns characteristic of the donor cells;pNSCs containing only maternal alleles regain the biparentalimprinting patterns after reprogramming. To date, ESCsderived from parthenogenetic embryos have been widely usedto study parthenogenetic development. pNSCs, which are alsocapable of self-renewal, maintain their imprinting pattern; yetonce the cells are reprogrammed to pluripotent miPS cells,their parthenogenetic imprinting pattern becomes similar tothat of pESCs. Thus, changes in the genomic imprinting ofparthenogenetic cells occur concomitantly with the inductionof pluripotency in multipotent stem cells. Here, we suggest
that pluripotential reprogramming induced the loss of parthe-nogenetic imprinting and postulate that changes in genomicimprinting are a marker for pluripotency but not for multipo-tency. This phenomenon may be explored in an effort to dis-cover treatments for a number of diseases, such as Prader-Williand Angelman syndrome, which are caused by an abnormalityin imprinted genes [27]. It could be envisioned that cells froma patient afflicted with an imprinting-related disease that ex-hibit abnormal imprinting patterns could be subjected to invitro reprogramming and differentiation and then transplantedback to the patient to replace cells of the impaired tissue.Therefore, pNSCs and miPS cells represent suitable ex vivomodel systems for exploring how genomic imprinting is regu-lated in both somatic and pluripotent cell types.
ACKNOWLEDGMENTS
We are grateful to Claudia Ortmeier for assisting with the real-time RT-PCR experiments. We thank Dr. Toshio Kitamura forproviding the MX retroviral vector.
DISCLOSURE OF POTENTIAL CONFLICTS
OF INTEREST
The authors indicate no potential conflicts of interest.
REFERENCES
1 Takahashi K, Yamanaka S. Induction of pluripotent stem cells frommouse embryonic and adult fibroblast cultures by defined factors. Cell2006;126:663–676.
2 Aoi T, Yae K, Nakagawa M et al. Generation of pluripotent stem cellsfrom adult mouse liver and stomach cells. Science 2008;321:699–702.
3 Hanna J, Markoulaki S, Schorderet P et al. Direct reprogramming ofterminally differentiated mature B lymphocytes to pluripotency. Cell2008;133:250–264.
4 Stadtfeld M, Brennand K, Hochedlinger K. Reprogramming of pancre-atic beta cells into induced pluripotent stem cells. Curr Biol 2008;18:890–894.
5 Eminli S, Utikal J, Arnold K et al. Reprogramming of neural progeni-tor cells into induced pluripotent stem cells in the absence of exoge-nous Sox2 expression. Stem Cells 2008;26:2467–2474.
6 Kim JB, Zaehres H, Wu G et al. Pluripotent stem cells induced fromadult neural stem cells by reprogramming with two factors. Nature2008;454:646–650.
7 Shi Y, Do JT, Desponts C et al. A combined chemical and geneticapproach for the generation of induced pluripotent stem cells. CellStem Cell 2008;2:525–528.
8 Horii T, Kimura M, Morita S et al. Loss of genomic imprinting inmouse parthenogenetic embryonic stem cells. Stem Cells 2008;26:79–88.
9 Do JT, Scholer HR. Nuclei of embryonic stem cells reprogram so-matic cells. Stem Cells 2004;22:941–949.
10 Zaehres H, Daley GQ. Transgene expression and RNA interference inembryonic stem cells. Methods Enzymol 2006;420:49–64.
11 McGrath J, Solter D. Completion of mouse embryogenesis requiresboth the maternal and paternal genomes. Cell 1984;37:179–183.
12 Surani MA, Barton SC. Development of gynogenetic eggs in themouse: Implications for parthenogenetic embryos. Science 1983;222:1034–1036.
13 Hasegawa K, Yoshikawa K. Necdin regulates p53 acetylation via Sir-tuin1 to modulate DNA damage response in cortical neurons. J Neuro-sci 2008;28:8772–8784.
14 Lee S, Walker CL, Wevrick R. Prader-Willi syndrome transcripts areexpressed in phenotypically significant regions of the developingmouse brain. Gene Expr Patterns 2003;3:599–609.
15 Pereira CM, Sattlegger E, Jiang HY et al. IMPACT, a protein prefer-entially expressed in the mouse brain, binds GCN1 and inhibits GCN2activation. J Biol Chem 2005;280:28316–28323.
16 Kim JB, Sebastiano V, Wu G et al. Oct4-induced pluripotency inadult neural stem cells. Cell 2009;136:411–419.
17 Jaenisch R, Young R. Stem cells, the molecular circuitry of pluripo-tency and nuclear reprogramming. Cell 2008;132:567–582.
18 Xiong Z, Laird PW. COBRA: A sensitive and quantitative DNAmethylation assay. Nucleic Acids Res 1997;25:2532–2534.
19 Kanatsu-Shinohara M, Inoue K, Lee J et al. Generation of pluripotentstem cells from neonatal mouse testis. Cell 2004;119:1001–1012.
20 Yoshida Y, Takahashi K, Okita K et al. Hypoxia enhances the genera-tion of induced pluripotent stem cells. Cell Stem Cell 2009;5:237–241.
21 Hong H, Takahashi K, Ichisaka T et al. Suppression of induced pluri-potent stem cell generation by the p53-p21 pathway. Nature 2009;460:1132–1135.
22 Kawamura T, Suzuki J, Wang YV et al. Linking the p53 tumour suppres-sor pathway to somatic cell reprogramming. Nature 2009;460:1140–1144.
23 Li H, Collado M, Villasante A et al. The Ink4/Arf locus is a barrierfor iPS cell reprogramming. Nature 2009;460:1136–1139.
24 Marion RM, Strati K, Li H et al. A p53-mediated DNA damageresponse limits reprogramming to ensure iPS cell genomic integrity.Nature 2009;460:1149–1153.
25 Utikal J, Polo JM, Stadtfeld M et al. Immortalization eliminates aroadblock during cellular reprogramming into iPS cells. Nature 2009;460:1145–1148.
26 Park IH, Arora N, Huo H et al. Disease-specific induced pluripotentstem cells. Cell 2008;134:877–886.
27 Nakao M, Sasaki H. Genomic imprinting: Significance in development anddiseases and the molecular mechanisms. J Biochem 1996;120:467–473.
See www.StemCells.com for supporting information available online.
2968 Parthenogenetic iPS Cells

![STEM CELLS EMBRYONIC STEM CELLS/INDUCED PLURIPOTENT STEM CELLS Stem Cells.pdf · germ cell production [2]. Human embryonic stem cells (hESCs) offer the means to further understand](https://img.dokumen.tips/doc/110x75/6014b11f8ab8967916363675/stem-cells-embryonic-stem-cellsinduced-pluripotent-stem-cells-stem-cellspdf.jpg)